Introduction
Posterior capsule opacification (PCO) is a secondary
cataract, which is the most important complication of extracapsular
cataract extraction and the main reason for postoperative visual
decline (1). The main molecular
biological mechanism underlying the development of PCO involves the
transformation of residual lens epithelial cells (LECs) into
mesenchymal cells and the migration of these cells to the posterior
capsule, leading to abnormal extracellular matrix (ECM) deposition
on and fibrosis of the posterior capsule of the lens (1-4).
The expression of transforming growth factor (TGF)
is elevated in the anterior chamber following cataract surgery, and
TGF-β is currently considered one of the most important promoters
of LEC transdifferentiation, which leads to pathological fibrosis
(5). During the
epithelial-mesenchymal transition (EMT), LECs transform into
myofibroblasts and undergo cytoskeletal rearrangements, with events
associated with the deposition of several ECM proteins, including
lumican, collagen, fibronectin, hyaluronic acid (HA) (6) and osteopontin (OPN) (7). It has been reported that TGF-β
regulates numerous cell biological behaviors by activating mothers
against decapentaplegic homolog (Smad)-mediated signal transduction
pathways (8). Smad2 and Smad3 are
phosphorylated receptor-activated Smads, which are essential for
the transmission of TGF-β signals. However, Smad2 and Smad3 have
different biological functions. Smad2 knockout (KO) causes mice to
die before they are born, whereas Smad3-KO mice survive until birth
but exhibit mucosal immunodeficiency (9). Granulation tissue lacking Smad3
produces limited quantities of ECM (10). Additionally, the loss of Smad3
disturbs the TGF-β-mediated induction of the EMT process and renal
fibrosis (11,12).
Smad3, an integral part of the TGF-β/Smad3 signaling
pathway, is closely associated with fibrotic diseases and the
TGF-β1/Smad3 pathway is involved in renal fibrosis in patients with
kidney disease (13). Smad3 KO
(Smad3−/−) was found to reduce esophageal fibrosis in a
model of egg-induced eosinophilic esophagitis (14). Skin cells derived from Smad3-KO
mice show less severe inflammation and enhanced wound healing
(15). According to the results
of our previous in vitro experiments, silencing of Smad3
interfered with TGF-β2-induced cell proliferation and downstream
ECM protein (10). In addition,
the overexpression of Smad3 was found to activate the TGF-β2
signaling pathway and increased the secretion of ECM components in
the human lens epithelial (HLE)-B3 human lens epithelial cell line
(16).
The present study investigated whether Smad3 KO
prevented trauma-induced EMT of the lens epithelium in vivo.
In addition, the study investigated whether inhibition of the
phosphorylation of Smad3 (p-Smad) by SIS3 affected ECM production
in vitro. The potential association between apoptosis and
Smad3 was also determined. Wild-type (Smad3+/+; WT) and
Smad3−/− mice were used to reveal the role of Smad3 in
the lens epithelium following injury. The mRNA or protein
expression levels of multiple EMT markers, including α-smooth
muscle actin (α-SMA), Snail1, Slug (Snail 2), Twist1,
SMAD-interacting protein 1 (SIP1), E-cadherin, lumican,
fibronectin, OPN, HA and collagen, were measured. Inhibition of the
TGF-β/Smad3 pathway may be clinically desirable for the prevention
of capsular opacification, a potential complication of cataract
surgery.
Materials and methods
Animals and traumatic cataract model
generation
All experimental procedures were performed according
to the ARVO Declaration on the Use of Animals in Ophthalmic and
Vision Research (17) and were
approved by the Tianjin Medical University Animal Ethics Committee
(Tianjin, China; approval no. TMUaMEC 2016009; June 6, 2016).
Adult C57BL/6J mice and Smad3-KO mice (18) were employed in the present study
(age, 6-8 weeks; weight, 20±2 g; sex ratio, 76 male and 76 female;
76 KO and 76 WT mice). C57BL/6J mice were purchased from the
Institute of Genetics and Developmental Biology at the Chinese
Academy of Sciences (Beijing, China). The Smad3-KO mice were
provided by Dr Xiao Yang from the Genetic Laboratory of Development
and Disease (Institute of Biotechnology, Academy of Military
Medical Sciences, Beijing, China). All mice were maintained under a
12 h light/dark cycle at 22-24°C with 45-50% humidity. Food and
water was available ad libitum. The mice were anesthetized
by intraperitoneal injections of pentobarbital sodium (70 mg/kg)
(19). A 26-gauge hypodermic
needle was applied to the right eye to puncture the corresponding
central anterior capsule through the corneal limbus, following
mydriasis and the administration of topical anesthesia. The depth
of the puncture wound was ~25% of the length of the blade portion
of the needle, as previously reported (17,19). The left eye served as an uninjured
control. The mice were treated with erythromycin eye ointment and
ofloxacin eye drops from 0 h to 4 weeks post-surgery. Finally,
healing mice in the different groups were sacrificed by sevoflurane
anesthesia and cervical dislocation to obtain the eyeballs for the
subsequent analyses.
Cell culture and in vitro
experiments
The HLE-B3 cell line was purchased from American
Type Culture Collection (Manassas, VA, USA). The cells were
cultured in Dulbecco's modified Eagle's medium/F-12 (HyClone; GE
Healthcare Life Sciences, Logan, UT, USA) containing 20% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) at 37°C with 5% CO2. The cells were serum-starved
overnight and then treated with TGF-β2 (10 ng/ml; R&D Systems,
Inc., Minneapolis, MN, USA) for 24 h to measure the ECM component
expression levels. In a separate group, the cells were pretreated
with 3 µM SIS3 (p-Smad3 inhibitor; Sigma-Aldrich; EMD
Millipore, Billerica, MA, USA) for 4 h prior to being treated with
TGF-β2 (10 ng/ml). SIS3 was dissolved in 100% dimethyl sulfoxide
(DMSO). DMSO was maintained at a final concentration of 0.1% in all
experiments.
Immunofluorescence staining
Following extraction from the mice, the eyeballs
were fixed in 40 g/l paraformaldehyde solution for 24 h at 4°C.
Subsequently, the fixed eyeballs were rinsed with PBS three times
for 5 min each, and the eyeballs were dehydrated in 15% sucrose
solution for 24 h, prior to being placed in 30% sucrose solution.
Each eye was then embedded in optimum cutting temperature compound
and sliced into 6-µm sagittal sections with a cryostat
(Leica CM 1850; Leica Microsystems GmbH, Wetzlar, Germany). The
sections were stored at −80°C until staining.
For the α-SMA, E-cadherin, lumican, OPN,
fibronectin, HA and collagen type I immunofluorescence localization
experiments, the frozen sections were removed from the −80°C
freezer, warmed for 30 min at room temperature and rinsed with PBS
three times for 5 min. The sections were then permeabilized with
0.1% Triton X-100 for 15 min; 5% bovine serum albumin (BSA; Roche
Diagnostics, Basel, Switzerland) was added to the slides for
non-specific antigen blocking, and then the slides were incubated
for 60 min at room temperature. The sections were subsequently
incubated with primary antibodies specific for α-SMA (1:500
dilution; cat. no. ab5694), OPN (1:200 dilution; cat. no. ab8448),
lumican (1:200 dilution; cat. no. ab168348), fibronectin (1:500
dilution; cat. no. ab2413) E-cadherin (1:500 dilution; cat. no.
ab11512), HA (1:100; cat. no. Sc-7392) and collagen type I (1:500
dilution; cat. no. ab34710) overnight at 4°C. With the exception of
the HA antibody, which was purchased from Santa Cruz Biotechnology,
Inc. (Dallas, TX, USA), all of the primary antibodies were
purchased from Abcam (Cambridge, MA, USA). The following day, the
slides were washed with PBS three times, and they were then
incubated with an Alexa Fluor 488-conjugated goat anti-rabbit
(A11034), goat anti-rat (A21210), and goat anti-mouse (A11029)
secondary antibodies (1:200 dilution; Thermo Fisher Scientific,
Inc.) for 60 min at room temperature. The HLE-B3 cells were
cultured in 24-well plates (2×104 cells per well) and
then treated with TGF-β2 (10 ng/ml) with or without SIS3 (3
µM) for 24 h. The cells were rinsed with PBS three times for
5 min each and then fixed with 4% paraformaldehyde for 30 min.
Following washing with PBS, the cells were incubated with 5% BSA
for 30 min and permeabilized with 0.1% Triton X-100 for 15 min. The
cells were then incubated with primary antibodies specific for
fibronectin (dilution 1:200) and E-cadherin (dilution 1:200)
overnight at 4°C. The cells were washed with PBS three times, and
then incubated with an Alexa Fluor 488-conjugated goat anti-rabbit
and goat anti-mouse secondary antibody (1:200 dilution; Thermo
Fisher Scientific, Inc.) for 60 min at room temperature. Following
washing with PBS, the sections and cells were sealed with mounting
medium with DAPI (Beijing Solarbio Science & Technology Co.,
Ltd., cat. no. S2110). Finally, LEC protein expression was observed
with a laser scanning confocal microscope (FV1000; Olympus
Corporation, Tokyo, Japan), and images were captured.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from the lenses (n=4 samples
per group) with the RNeasy Mini kit in accordance with the
manufacturer's protocol (Qiagen GmbH, Hilden, Germany). When the
HLE-B3 cells had been subjected to the above treatments, they were
lysed in RLT buffer. The total RNA concentration was measured using
a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Inc.).
A total of 2 µg of total RNA was used as a template to
synthesize cDNA with M-MLV Reverse Transcriptase kit (Promega
Corporation, Madison, WI, USA) according to the manufacturer's
protocols. The cDNA was then stored at −20°C for later use. qPCR
was performed with SYBR Premix Ex Taq™ (Takara Biotechnology Co.,
Ltd., Dalian, China), and the data was collected using an
StepOnePlus™ Real-Time PCR system (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The qPCR thermocycling conditions were as
follows: 95°C for 3 min, followed by 40 cycles at 95°C for 12 sec
and 62°C for 40 sec, a final extension step of 95°C for 15 sec,
60°C for 1 min, 95°C for 15 sec. The relative expression levels of
genes were detected and GAPDH served as an internal control. The
primer sequences are presented in Table I. The data were calculated by the
2−ΔΔCq method (20).
 | Table IPrimers used for reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I
Primers used for reverse
transcription-quantitative polymerase chain reaction analysis.
| Gene | Forward primer | Reverse primer |
|---|
| α-SMA |
5′-TCACCATTGGAAACGAACGC-3′ |
5′-GCTGTTATAGGTGGTTTCGTGG-3′ |
| Snail1 |
5′-CACACGCTGCCTTGTGTCT-3′ |
5′-GGTCAGCAAAAGCACGGTT-3′ |
| CDH1 |
5′-CAGTTCCGAGGTCTACACCTT-3′ |
5′-TGAATCGGGAGTCTTCCGAAAA-3′ |
| Slug |
5′-CAGCGAACTGGACACACACA-3′ |
5′-ATAGGGCTGTATGCTCCCGAG-3′ |
| Twist1 |
5′-GGACAAGCTGAGCAAGATTCA-3′ |
5′-CGGAGAAGGCGTAGCTGAG-3′ |
| SIP1 |
5′-TCTGTGTAGCCCTGGCTGTC-3′ |
5′-GGTGGTCTCGCACTCCTTTA-3′ |
| HAS1 |
5′-GCGAGCACTCACGATCATCTT-3′ |
5′-GTCCATAGCGATCTGAAGCCA-3′ |
| HAS2 |
5′-GTACGGTGCCTTTTTAGCCTC-3′ |
5′-TAATCGGGGTTTCAAGGGACT-3′ |
| HAS3 |
5′-CCTGGAGCACCGTCGAATG-3′ |
5′-CCTTGAGGTTTGGAAAGGCAA-3′ |
| Lumican |
5′-TGGGACCACTGTCTTACT-3′ |
5′-CTGCTCCAGAACATACTT-3′ |
| OPN |
5′-TCCTAGACCCTAAGAGTAA-3′ |
5′-TAAGCTAAGAGCCCAAAA-3′ |
| Fibronectin |
5′-GAAGACAGATGAGCTTCCCCA-3′ |
5′-GGTTGGTGATGAAGGGGGTC-3′ |
| Collagen type
I |
5′-TTCTGCAGGGTTCCAACGAT-3′ |
5′-GCAGGCGAGATGGCTTATTTG-3′ |
| GAPDH |
5′-GCAACTCCCACTCTTCCA-3′ |
5′-GTCCAGGGTTTCTTACTCC-3′ |
| hGAPDH |
5′-GGAGCGAGATCCCTCCAAAAT-3′ |
5′-GGCTGTTGTCATACTTCTCATGG-3′ |
| hα-SMA |
5′-GTGTTGCCCCTGAAGAGCAT-3′ |
5′-GCTGGGACATTGAAAGTCTCA-3′ |
| hSnail |
5′-TCGGAAGCCTAACTACAGCGA-3′ |
5′-AGATGAGCATTGGCAGCGAG-3′ |
| hCDH1 |
5′-CGAGAGCTACACGTTCACGG-3′ |
5′-GGGTGTCGAGGGAAAAATAGG-3′ |
| hFN1 |
5′-CGGTGGCTGTCAGTCAAAG-3′ |
5′-AAACCTCGGCTTCCTCCATAA-3′ |
| hCOL1A1 |
5′-GAGGGCCAAGACGAAGACATC-3′ |
5′-CAGATCACGTCATCGCACAAC-3′ |
Western blot analysis
The lenses (n=4 per group) from each group were
collected and homogenized in lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China) containing a protease and phosphatase
inhibitor cocktail (Biotool, Shanghai, China). The precipitate was
removed through centrifugation (12,000 × g for 5 min at 4°C), and
the protein concentration of the supernatant was measured using a
bicinchoninic acid assay kit (Thermo Fisher Scientific, Inc.). The
protein sample from the supernatant was then mixed with 5X SDS-PAGE
loading buffer and heated for 5 min at 98°C. Equal quantities of
proteins (30 µg) were separated by 8-10% SDS-PAGE and then
transferred onto a PVDF membrane for 2 h at 300 mA. The membrane
was then incubated with 5% skim milk for 2 h at room temperature,
and then incubated with primary antibodies specific for α-SMA
(1:2,000 dilution), lumican (1:2,000 dilution), fibronectin
(1:10,000 dilution), OPN (1:5,000 dilution) and GAPDH (1:10,000
dilution) in Blotto overnight at 4°C. Following three washes with
PBST, the membrane was incubated with the appropriate secondary
antibody (1:20,000; cat. no. ab6721; Abcam) for 2 h at room
temperature. The membrane was then subjected to three washes with
PBST, and protein expression was evaluated by chemiluminescence
reagent (EMD Millipore) and exposure to chemiluminescent film. The
grayscale images of the proteins were analyzed using ImageJ
software (v.1.42q; National Institutes of Health, Bethesda, MD,
USA).
TUNEL assay
Apoptosis was measured using a TUNEL detection kit
(Roche Diagnostics). Briefly, the frozen sections were removed from
storage and air-dried for 30 min at room temperature. Following
washing with PBS, the sections were permeabilized with citrate
buffer containing 0.1% Triton X-100 for 2 min on ice. DNase I was
prepared to treat the positive control samples prior to TUNEL
staining. The TUNEL reaction mixture was then added to the
sections, which were incubated for 1 h at 37°C. Finally, the
sections were mounted in medium with DAPI, and the resultant images
were observed under a fluorescence microscope (DM4000 B LED; Leica
Microsystems GmbH). The TUNEL-positive cell nuclei contained green
fluorescent particles. For quantitative analysis, five fields in
each of the sections from each group were randomly selected and
observed at ×400 magnification, and the numbers of TUNEL-positive
nuclei among 120 LECs were counted.
Statistical analysis
All experimental results are presented as the mean ±
standard deviation of at least three independent experiments. All
statistical analyses were performed using SPSS (v.19.0; IBM SPSS,
Armonk, NY, USA). Student's t-test was used to assess the
differences between two groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
Absence of Smad3 suppresses
trauma-induced EMT
To determine whether the absence of Smad3 inhibited
the EMT of the lens epithelium following injury, the present study
first examined the expression levels of α-SMA, a commonly used EMT
marker, using immunofluorescence staining. α-SMA was expressed in
Smad3−/− mice, however, the intensity of
immunofluorescence staining intensity in the Smad3−/−
mice was reduced, compared with that in the WT mice on days 7, 14
and 28 post-injury (Fig. 1A).
Additionally, the expression of α-SMA was detected by western blot
analysis, the results of which confirmed those of the
immunofluorescence experiments. Specially, western blot analysis
revealed that the expression levels of α-SMA in the
Smad3−/− mice were reduced by 87.5, 92.7 and 90.6%,
compared with those in the Smad3+/+ mice on days 7, 14
and 28 post-injury, respectively (P<0.05; Fig. 1B). The expression of the
epithelial marker, E-cadherin, was also detected, the
downregulation of which is the most important molecular event in
EMT development (21). The
Smad3−/− mice expressed higher levels of E-cadherin,
compared with the WT mice at all time points during EMT.
Immunofluorescence staining confirmed that the expression levels of
E-cadherin were increased in Smad3−/− mice, compared
with those in WT mice (Fig. 1C).
Additionally, to further confirm the EMT process, the EMT-related
transcription factors Snail1, Slug, SIP1 and Twist1 were examined
by RT-qPCR analysis. The results showed that the expression of
these four transcription factors increased in trauma-induced EMT of
LECs and decreased significantly in Smad3-KO mice, compared with WT
mice (Fig. 1D-G). As shown in
Fig. 1H, the RT-qPCR results
revealed that the expression of α-SMA in the Smad3-KO mice had
decreased to levels that were 35.5, 25.3 and 20% of those in the WT
mice on days 7, 14 and 28, respectively (P<0.001). The mRNA
expression levels of E-cadherin in the Smad3-KO mice were 4-, 6-,
and 2.8-fold higher than those in the WT mice on days 7, 14 and 28
post-surgery, respectively (Fig.
1I).
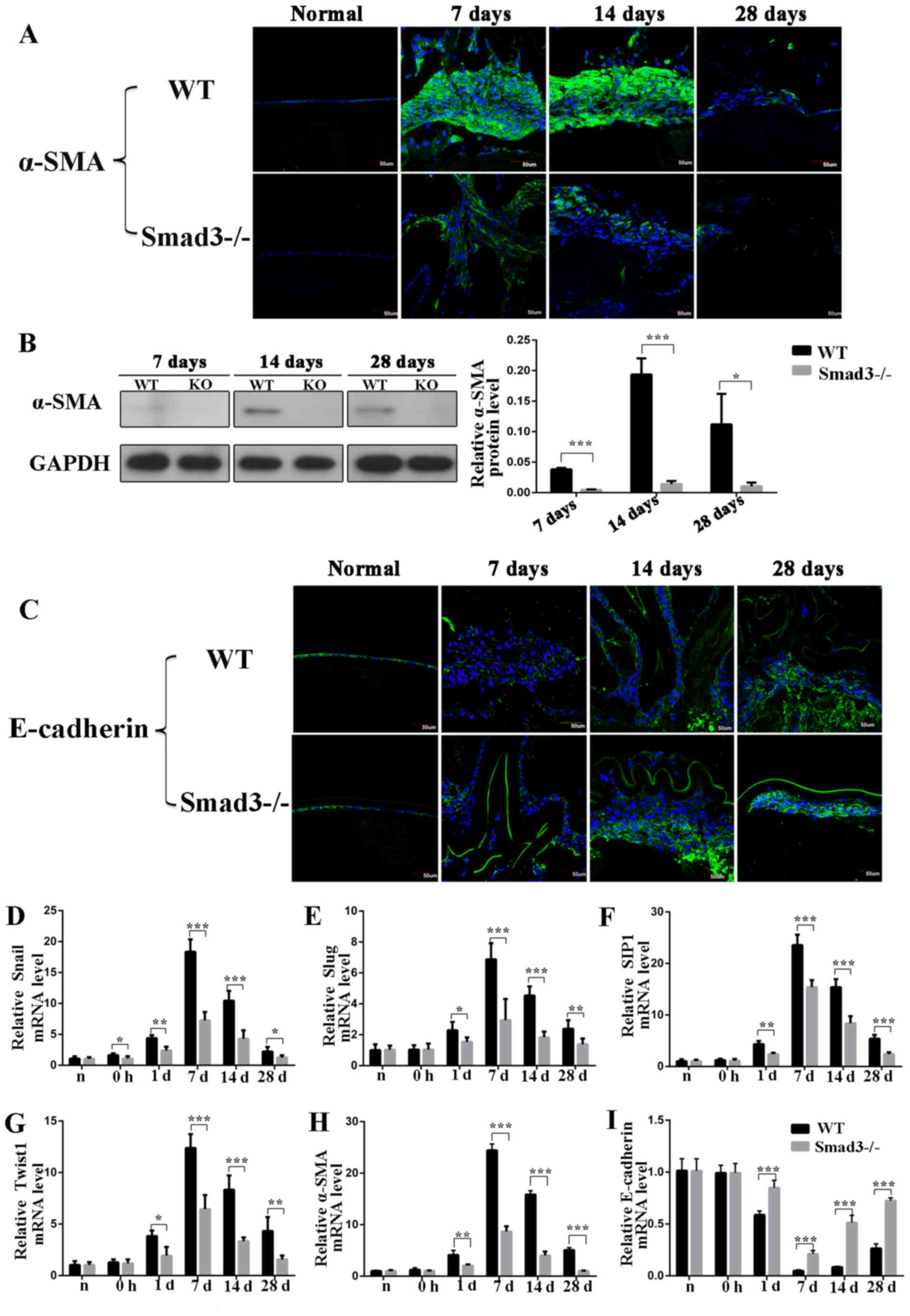 | Figure 1Expression of epithelial-mesenchymal
transition biomarkers at different time points following anterior
capsular injury. (A) Immunohistochemical analysis of α-SMA. (B)
Western blot analysis revealed that expression of α-SMA was reduced
to levels that were 87.5, 92.7 and 90.6% in Smad3-KO mice of those
in WT mice on days 7, 14 and 28, respectively. (C) In situ
protein expression of E-cadherin in lens epithelial cells following
injury. Transcription factor expression of (D) Snail, (E) Slug, (F)
SIP1 and (G) Twist1 in the lens epithelium following injury. (H)
Reverse transcription-quantitative polymerase chain reaction
analysis revealed that mRNA expression of α-SMA in Smad3-KO mice
decreased to levels that were 35.5, 25.3 and 20% of those in WT
mice on days 7, 14 and 28 post-injury, respectively. (I) mRNA
expression level of E-cadherin at different time points following
anterior capsule injury. Scale bar=50 µm.
*P<0.05, **P<0.01,
***P<0.001. Smad3, mothers against decapentaplegic 3;
KO, knockout; WT, wild-type; α-SMA, α-smooth muscle actin; SIP1,
SMAD-interacting protein 1. |
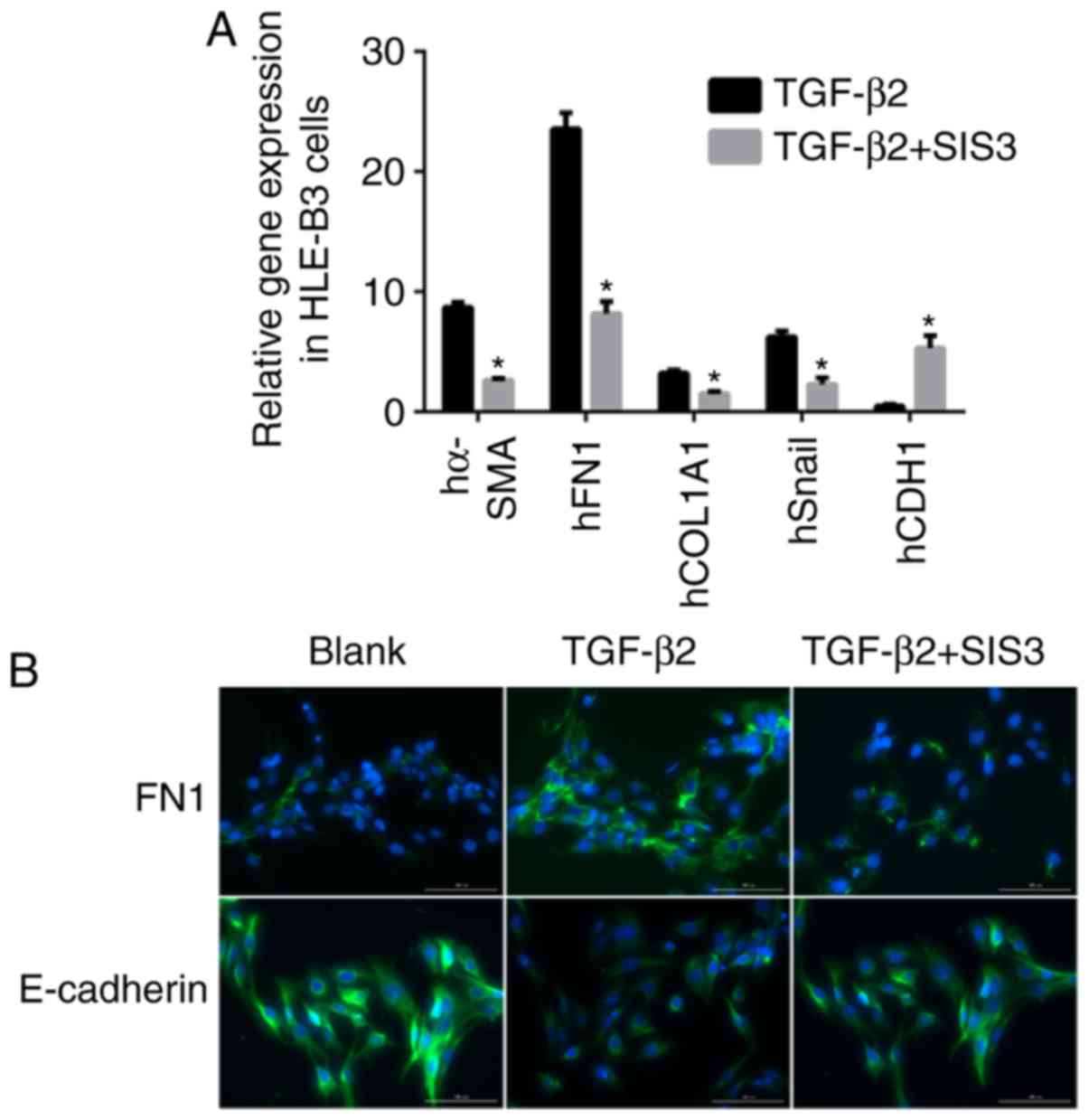
For the in vitro experiments SIS3 was added
to the culture to inhibit the phosphorylation of Smad3. As shown in
Fig. 2A, the mRNA expression
levels of α-SMA and Snail were decreased by 3.6- and 3.7-fold,
respectively, and that of E-cadherin was increased by 5.2-fold in
the TGF-β2+SIS3 group, compared with the TGF-β2 group. The
immunofluorescence staining confirmed these results (Fig. 2B).
Loss of Smad3 decreases ECM accumulation
and suppresses fibrosis during trauma-induced EMT
To examine ECM accumulation during lens epithelium
wound healing, the present study detected the expression levels of
lumican, OPN, fibronectin, collagen type I and HA (Fig. 3A-a-g). Lumican is an ECM protein
belonging to the small leucine-rich proteoglycan family, which
binds to cytokines, regulates ECM secretion by mesenchymal cells,
and is involved in epithelial cell migration and tissue repair
(22). The expression of lumican
was examined using RT-qPCR and western blot analyses. As shown in
Fig. 3A-a, the mRNA expression of
lumican in the Smad3−/− mice was decreased to levels
that were 21.3, 11.5 and 13.2% of those in the WT mice on days 7,
14 and 28 post-surgery, respectively (P<0.001). The western blot
analysis revealed that the expression of lumican in the Smad3-KO
mice decreased to levels that were 52.6, 7.9 and 8.8% of those in
the WT mice on days 7, 14 and 28 post-injury, respectively
(P<0.05), which was consistent with the RT-qPCR results
(Fig. 3B). Immunofluorescence
staining confirmed that the expression of lumican was decreased in
the absence of Smad3 (Fig.
3C).
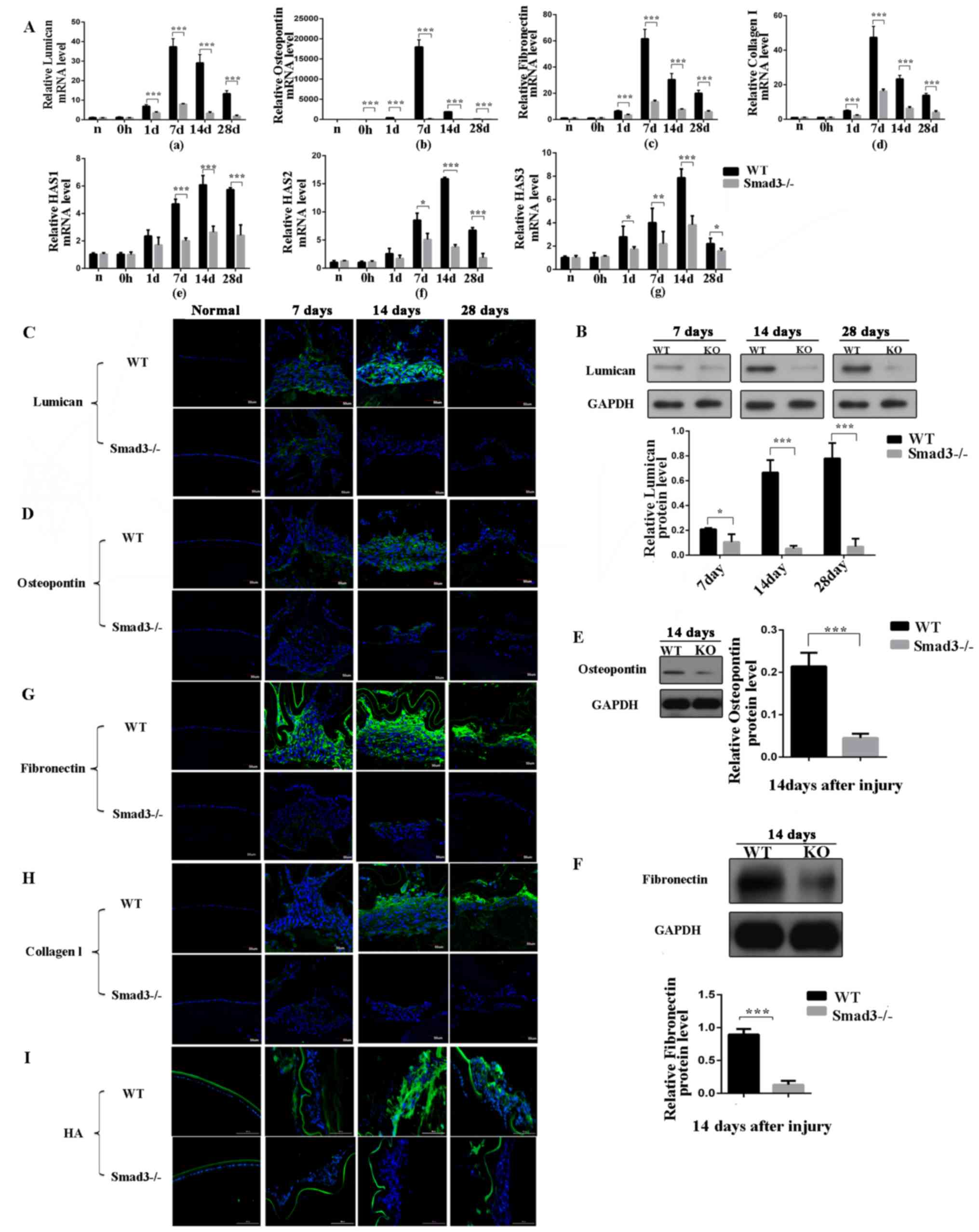 | Figure 3Expression levels of extracellular
matrix components at different time points following anterior
capsule injury. (A) mRNA expression levels of (a) lumican, (b)
oesteopontin, (c) fibronectin, (d) collagen type I and (e-g) HAS in
lens epithelial cells following anterior capsule injury. In
situ protein expression levels were determined. (B) Western
blot and (C) immunofluorescence of lumican; (D) immunofluorescence
and (E) western blot of osteopontin; (F) western blot and (G)
immunofluorescence of fibronectin. Immunofluorescence of (H)
collagen type I and (I) HA in lens epithelial cells following
injury. Scale bar=50 or 100 µm. *P<0.05,
**P<0.01, ***P<0.001. KO, knockout; WT,
wild-type; Smad3, mothers against decapentaplegic 3; HAS,
hyaluronan synthase; HA, hyaluronan. |
OPN is a glycoprotein, which is secreted into the
ECM by several cells and is expressed in PCO and anterior
subcapsular cataract (ASC) tissues (23). To obtain additional insight into
the role of OPN in the EMT of the lens epithelium and to identify
the steps of the EMT that are dependent on Smad3, the present study
examined OPN expression patterns in WT and KO lenses following
injury. The WT LECs exhibited strong immunoreactivity to OPN
protein, compared with the Smad3-KO LECs at all time points
following injury (Fig. 3D). As
shown in Fig. 3A-b, the mRNA
expression of OPN in the Smad3-KO mice was decreased to levels that
were 4.4, 2.2 and 5.6% of those in the WT mice on days 7, 14 and 28
post-surgery, respectively (P<0.001). The protein expression
levels of OPN were measured by western blot analysis on day 14, and
the protein expression of OPN in the Smad3-KO mice was 79.2% lower
than that in the WT mice (P<0.001; Fig. 3E).
Fibronectin is a fibrotic marker and is a major ECM
molecule, which is absent in the clear adult lens; however,
fibronectin accumulates around LECs in PCO (24). Therefore, the present examined the
expression of fibronectin in Smad3-KO and WT mice following
puncturing the central anterior lens capsule. The absence of Smad3
decreased the mRNA expression of fibronectin in the Smad3-KO mice
to levels that were 22.2, 25 and 29.4% of those in the WT mice on
days 7,14 and 28 post-injury, respectively, as shown in Fig. 3A-c. Additionally, the western blot
analysis revealed that the expression of fibronectin in the
Smad3-KO was decreased by 85.5%, compared with that in the WT mice
on day 14 post-injury (Fig. 3F).
Immunoreactivity to fibronectin was compared between the two
groups. Fibronectin immunofluorescence intensity in the Smad3-KO
group was lower than that in the WT group (Fig. 3G). In the in vitro
experiments described above, the mRNA expression levels of
fibronectin were 3-fold lower in the TGF-β2+SIS3 group, compared
those in the TGF-β2 group (Fig. 2A
and B), and the immunofluorescence staining confirmed these
results.
In the transparent lens, collagen expression is
limited to the capsule of the lens; however, abnormal collagen
deposition has been identified in ASC plaques and PCO (25). Analysis of the expression of
collagen type I was performed using RT-qPCR and immunofluorescence.
The mRNA expression of collagen type I in the Smad3-KO mice was
decreased to levels that were 65.5, 73 and 71.4% of those in the WT
mice on days 7, 14 and 28 following anterior capsular injury,
respectively (Fig. 3A-d).
Immunofluorescence staining confirmed that the expression of
collagen type I was decreased in the absence of Smad3 (Fig. 3H). In the in vitro
experiments, the mRNA expression levels of collagen type I were
2.2-fold lower in the TGF-β2+SIS3 group than in the TGF-β2 group
(Fig. 2A).
HA is found throughout the body, regulates cell
proliferation, migration and differentiation, and is key in
inflammation and tumorigenesis (26). Hyaluronan synthase (HAS) is an
enzyme, which is vital in HA synthesis and exists as one of the
following three subtypes: HAS1, HAS2 and HAS3. In the in
vivo mouse model, the expression of HA was increased following
injury; however, Smad3-KO decreased the expression of HA (Fig. 3I). The expression levels of HAS1,
HAS2 and HAS3 were further examined (Fig. 3A-e-g). As shown in Fig. 3A-f, the expression of HAS2 in the
Smad3-KO mice was decreased to levels that were 58, 23.4 and 27.6%
of those in the WT mice on days 7, 14 and 28 post-injury,
respectively (P<0.05), and the expression of HAS3 in the
Smad3-KO mice was decreased to levels that were 20, 48.9 and 63% of
those in the WT mice on days 7, 14 and 28 post-injury, respectively
(P<0.05) (Fig. 3A-g). However,
the expression of HAS1 in the Smad3-KO mice was decreased to levels
that were 42.6, 43.3 and 42.1% of those in the WT mice on days 7,
14 and 28, respectively (Fig.
3A-e).
Taken together, the above data indicated that Smad3
signaling is important in trauma-induced EMT in LECs in vivo
and in vitro. The deletion of the Smad3 gene reduced cell
transdifferentiation and ECM accumulation, and suppressed fibrosis
in trauma-induced EMT in LECs.
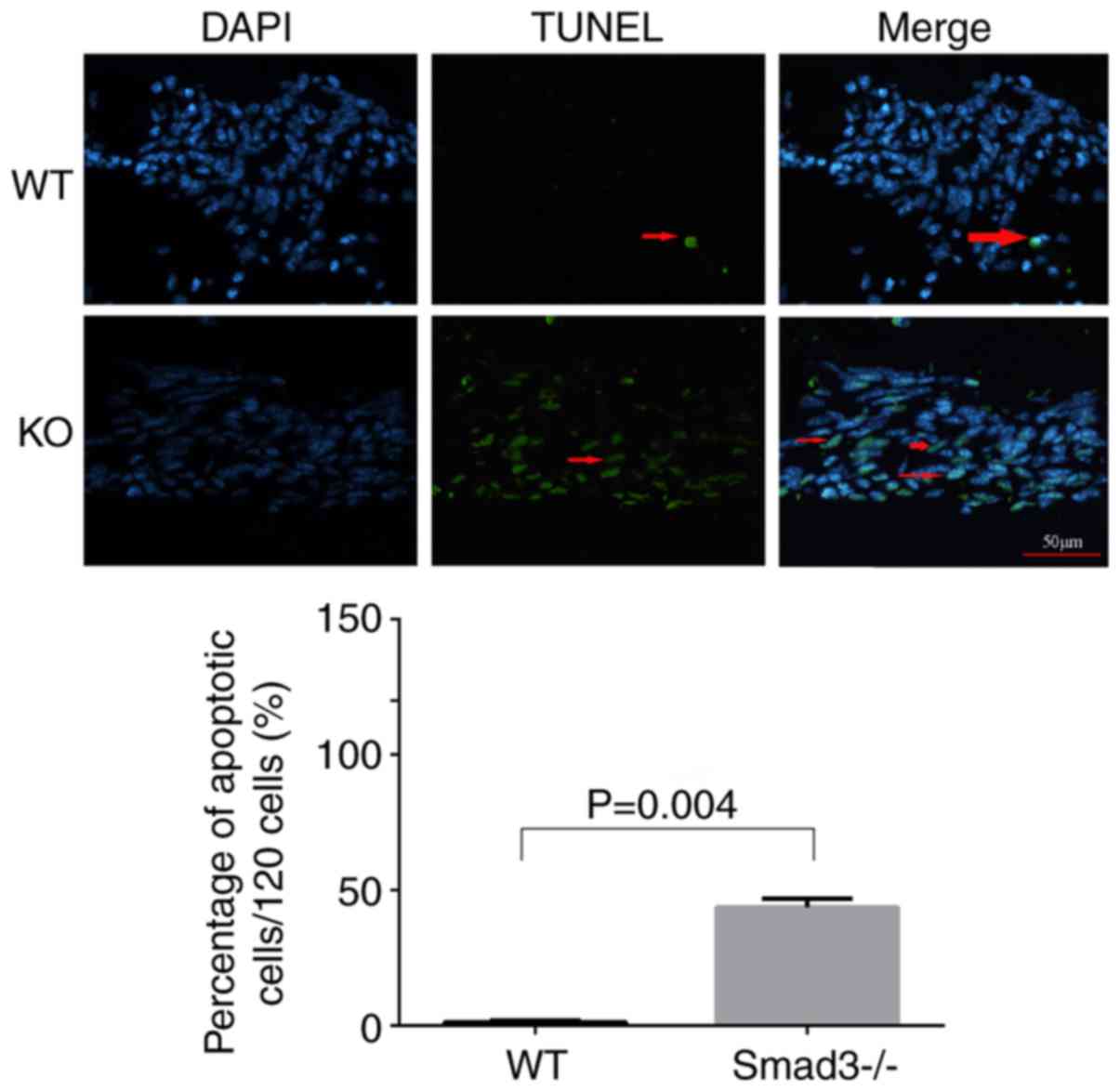
Smad3 deficiency induces LEC
apoptosis
TUNEL labeling was applied to compare apoptosis
levels between the two experimental groups in the present study.
LECs around the capsular puncture undergo apoptosis relatively
early in the healing process (27). Therefore, lens epithelial cells
apoptosis was detected on day 7 using TUNEL staining. The
Smad3−/− lenses exhibited a significantly higher
percentage of TUNEL-positive cells (39.82±4.4%), compared with the
Smad3+/+ lenses (1.1±0.44%) (Fig. 4).
Discussion
To determine whether Smad3-KO affects trauma-induced
EMT in the lens epithelium, differences between the WT and KO
groups were investigated using RT-qPCR analysis, western blot
analysis and immunofluorescence staining. It was found that Smad3
deficiency suppressed the EMT. The loss of Smad3 partially
inhibited the TGF-β signaling pathway in trauma-induced EMT in the
lens epithelium. The expression of the EMT marker α-SMA, the
hallmark of myofibroblasts, was significantly decreased in the
Smad3-KO mice, compared with that in the WT mice. The most
important change in protein expression that occurs during the EMT
is the decrease in the expression of E-cadherin (28). The expression of E-cadherin is
downregulated by Snail in the EMT (29). However, the expression of
E-cadherin, an epithelial marker, was increased in the Smad3-KO
mice, indicating that Smad3-KO effectively inhibited TGF-β/Smad
signaling pathway activity and thus inhibited the EMT. Furthermore,
transcription factors Snail1, Slug, SIP1 and Twist1 are involved in
the regulation of EMT (29-32), and the data showed that the
expression of these transcription factors was increased in
trauma-induced EMT and decreased in Smad3-KO mice. The Snail family
was mainly involved in this process and consistent with the
findings of a previous study (17). The levels of ECM components,
including lumican, OPN, fibronectin, HA and collagen type I, were
decreased in the Smad3-KO mice, compared with those in the WT mice
and were also decreased in the in vitro experiment. These
results showed that Smad3 regulated the production of ECM and were
consistent with those of a previous study (10,16). Smad3 may be used as a drug target
for the treatment of PCO, as previous studies showed that tranilast
and pirfenidone were effective in treating fibrosis by targeting
the TGF-β/Smad pathway (33-36). Further in vivo experiments
in the mouse model used are required to confirm the antifibrotic
drug effects. However, in the present study the absence of Smad3
did not completely inhibit trauma-induced EMT in mice in
vivo. Capsular injury resulted in the expression of α-SMA in
the local proliferating cell multilayer, and aberrant ECM
deposition, a change reflected by alterations in mRNA and protein
expression levels of the marker in Smad3-KO mice; however, the KO
lenses exhibited a lower intensity of α-SMA staining, compared with
that in the WT lenses. This result suggested that Smad3-KO
decreased trauma-induced EMT in the lens epithelium and activated
non-Smad signaling pathways (25,37,38).
Non-Smad pathways are also important in the EMT.
TGF-β modulates the EMT by activating protein kinases, including
mitogen-activated protein kinase, Ras homolog family member A and
phosphoinositide 3-kinase, among others (39-42).
The role of Smad3 in the EMT of epithelial cells
remains disputed (8). TGF-β
activates the Smad3 signaling pathway to promote the occurrence of
cataracts (25). Robertson et
al utilized recombinant adenoviruses that bind TGF-β in the
anterior chamber of WT mice and found that these mice developed
subcapsular cataracts, however, Smad3-KO mice exhibited delayed
cataract development (37).
Shirai et al (43) induced
a novel ASC model in which the EMT was inhibited in the Smad3-KO
mouse lens epithelium by exposing the ocular surface to topical
alkali agents. Unexpectedly, the EMT of the LECs was not completely
eliminated but was significantly attenuated in Smad3-KO mice. This
observation may be associated with the occurrence of a severe
inflammatory reaction in the anterior chamber, as inflammatory
reactions can activate TGF-β and other proinflammatory cytokines,
which may result in the activation of Smad3-dependent pathways or
non-Smad signaling pathways to regulate EMT (43).
In the present study, ECM deposition was observed in
Smad3-KO and WT lenses following injury. The pathologic bases of
PCO are residual LEC proliferation, migration and
transdifferentiation, collagen deposition, and fibrosis formation
following cataract surgery. Abnormal ECM accumulation is common in
ASC plaque formation and PCO (1,44,45). However, lower expression levels of
lumican, OPN, collagen type I, HA and fibronectin were observed in
the Smad3−/− lenses, compared with those in the WT
lenses, and fewer myofibroblasts were observed in the
Smad3−/− mice than in the WT mice. These results
corresponded with those pertaining to the ECM deposition patterns
of the two groups. TGF-β is vital in regulating the expression of
ECM, and increasing evidence suggest that this process depends on
Smad3. Fibrosis is significantly decreased in Smad3-KO wound beds,
compared with HT and WT wounds beds (46). In addition, Smad3-KO mice exhibit
less TGF-β2-induced ocular hypertension and fibronectin deposition
in trabecular meshwork cells, compared with WT mice (47). These results indicate that Smad3
is a mediator of the fibrotic response.
Of note, the expression levels of EMT markers and
ECM in the KO group peaked at day 7 post-injury and began to
decline at 2 weeks post-injury, but remained higher than the
corresponding levels in the normal control group. The EMT markers
and ECM components were expressed at low levels at 4 weeks
post-surgery. A previous study showed that expression levels of
α-SMA were highest on day 7 post-surgery (17), which may have been caused by
increases in the levels of various cytokines, leading to the
development of a high-cytokine 'microenvironment', prompting LEC
proliferation and differentiation by 2 weeks post-surgery.
Apoptosis is another characteristic of TGF-β-induced
ASC and PCO (48,49). TGF-β1/Smad3−/− mice
have been reported to exhibit a higher percentage of dead cells
than WT mice (25,37). In the present study, the number of
apoptotic cells was significantly increased in the
Smad3−/− mice, compared with that in the WT mice. This
result was consistent with those of the experiments in which mRNA
and protein expression levels were measured, as Smad3−/−
mice exhibited lower mRNA and protein expression levels than in WT
mice. Smad3-KO may increase apoptosis in trauma-induced EMT in the
lens epithelium. The activity of TGF-β is generally considered to
induce apoptosis. However, the present study found that Smad3-KO
increased cell apoptosis, a finding for which the following
explanations were suggested: i) a non-canonical TGF-β signaling
pathway was activated; ii) when arterial injury occurs, levels of
Smad3 are high, and TGF-β appears to stimulate smooth muscle cell
proliferation (50); iii) Smad3
may be required for the survival of intermediate progenitor cells
(51), as Smad3 gene-KO disrupts
normal cell proliferation, leading to increased apoptosis.
The results of the present study revealed that the
deletion of Smad3 inhibited trauma-induced EMT in vivo in
the Smad3-KO mouse model and in vitro. As Smad3-KO mice
exhibited lower levels of lumican, OPN, collagen type I, HA and
fibronectin deposition, compared with those in WT mice, it is
possible that signaling pathways independent of Smad3 are activated
and that fibrosis is an Smad3-dependent process. Smad3-KO may
increase cell death in trauma-induced EMT in the lens epithelium,
and Smad3 may be a target for the treatment of PCO and other
fibrosis-related diseases.
In conclusion, Smad3-KO decreased trauma-induced EMT
in the lens epithelium, but did not completely inhibit EMT.
Inhibition of the TGF-β/Smad3 pathway may be clinically desirable
for preventing capsular opacification, a potential complication of
cataract surgery.
Acknowledgments
The authors would like to thank Dr Jinyong Lin and
Dr Yuchuan Wang (Department of Pathology of Tianjin Eye Hospital,
Tianjin, China), Mr Ming Ying, Mr Peng Hao and Ms Ruifang Han
(Tianjin Key Laboratory of Ophthalmology and Visual Science,
Tianjin, China) for their technical assistance.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (grant no. 81670837), the
Tianjin Research Program of Application Foundation and Advanced
Technology (grant no. 13JCYBJC21500), the Tianjin Application
Foundation and Advanced Technology Program (15JCYBJC27600), and the
Youth Project of Tianjin Applied Basic Research and Cutting-edge
Technology Research Programs (grant no. 15JCQNJC45000).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FM performed the experiments, analyzed the data and
wrote the paper. XT, XY and JL made substantial contributions to
the concept and design of the present study. XY provided the Smad3
knockout mice. JL made substantial contributions to the analysis
and interpretation of data. XT and XY revised the paper critically
revised the manuscript for important intellectual content. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
All experimental procedures were performed according
to the ARVO Declaration on the Use of Animals in Ophthalmic and
Vision Research and were approved by the Tianjin Medical University
Animal Ethics Committee (approval no. TMUaMEC 2016009; June 6,
2016).
Consent for publication
Not applicable.
Competing interests
The authors confirm that they have no competing
interests.
References
|
1
|
Awasthi N, Guo S and Wagner BJ: Posterior
capsular opacification: A problem reduced but not yet eradicated.
Arch Ophthalmol. 127:555–562. 2009. View Article : Google Scholar
|
|
2
|
Nibourg LM, Gelens E, Kuijer R, Hooymans
JM, van Kooten TG and Koopmans SA: Prevention of posterior capsular
opacification. Exp Eye Res. 136:100–115. 2015. View Article : Google Scholar
|
|
3
|
Wallentin N, Wickström K and Lundberg C:
Effect of cataract surgery on aqueous TGF-beta and lens epithelial
cell proliferation. Invest Ophthalmol Vis Sci. 39:1410–1418.
1998.
|
|
4
|
Meacock WR, Spalton DJ and Stanford MR:
Role of cytokines in the pathogenesis of posterior capsule
opacification. Br J Ophthalmol. 84:332–336. 2000. View Article : Google Scholar
|
|
5
|
Lee EH and Joo CK: Role of transforming
growth factor-beta in transdifferentiation and fibrosis of lens
epithelial cells. Invest Ophthalmol Vis Sci. 40:2025–2032.
1999.
|
|
6
|
Chandler HL, Haeussler DJ Jr,
Gemensky-Metzler AJ, Wilkie DA and Lutz EA: Induction of posterior
capsule opacification by hyaluronic acid in an ex vivo model.
Invest Ophthalmol Vis Sci. 53:1835–1845. 2012. View Article : Google Scholar
|
|
7
|
Saika S, Shirai K, Yamanaka O, Miyazaki K,
Okada Y, Kitano A, Flanders KC, Kon S, Uede T, Kao WW, et al: Loss
of osteopontin perturbs the epithelial-mesenchymal transition in an
injured mouse lens epithelium. Lab Invest. 87:130–138. 2007.
View Article : Google Scholar
|
|
8
|
Derynck R and Zhang YE: Smad-dependent and
smad-independent pathways in TGF-beta family signalling. Nature.
425:577–584. 2003. View Article : Google Scholar
|
|
9
|
Uemura M, Swenson ES, Gaca MD, Giordano
FJ, Reiss M and Wells RG: Smad2 and Smad3 play different roles in
rat hepatic stellate cell function and alpha-smooth muscle actin
organization. Mol Biol Cell. 16:4214–4224. 2005. View Article : Google Scholar :
|
|
10
|
Li J, Tang X and Chen X: Comparative
effects of TGF-β2/Smad2 and TGF-β2/Smad3 signaling pathways on
proliferation, migration, and extracellular matrix production in a
human lens cell line. Exp Eye Res. 92:173–179. 2011. View Article : Google Scholar
|
|
11
|
Flanders KC: Smad3 as a mediator of the
fibrotic response. Int J Exp Pathol. 85:47–64. 2004. View Article : Google Scholar
|
|
12
|
Brodeur AC, Roberts-Pilgrim AM, Thompson
KL, Franklin CL and Phillips CL: Transforming growth
factor-β1/Smad3-independent epithelial-mesenchymal transition in
type I collagen glomerulopathy. Int J Nephrol Renovasc Dis.
10:251–259. 2017. View Article : Google Scholar
|
|
13
|
Zhao K, He J, Zhang Y, Xu Z, Xiong H, Gong
R, Li S, Chen S and He F: Activation of FXR protects against renal
fibrosis via suppressing Smad3 expression. Sci Rep. 6:372342016.
View Article : Google Scholar
|
|
14
|
Cho JY, Doshi A, Rosenthal P, Beppu A,
Miller M, Aceves S and Broide D: Smad3-deficient mice have reduced
esophageal fibrosis and angiogenesis in a model of egg-induced
eosinophilic esophagitis. J Pediatr Gastroenterol Nutr. 59:10–16.
2014. View Article : Google Scholar :
|
|
15
|
Wormstone IM, Tamiya S, Eldred JA,
Lazaridis K, Chantry A, Reddan JR, Anderson I and Duncan G:
Characterisation of TGF-beta2 signalling and function in a human
lens cell line. Exp Eye Res. 78:705–714. 2004. View Article : Google Scholar
|
|
16
|
Li H, Yuan X, Li J and Tang X: Implication
of Smad2 and Smad3 in transforming growth factor-β-induced
posterior capsular opacification of human lens epithelial cells.
Curr Eye Res. 40:386–397. 2015. View Article : Google Scholar
|
|
17
|
Saika S, Kono-Saika S, Ohnishi Y, Sato M,
Muragaki Y, Ooshima A, Flanders KC, Yoo J, Anzano M, Liu CY, et al:
Smad3 signaling is required for epithelial-mesenchymal transition
of lens epithelium after injury. Am J Pathol. 164:651–663. 2004.
View Article : Google Scholar
|
|
18
|
Yang X, Letterio JJ, Lechleider RJ, Chen
L, Hayman R, Gu H, Roberts AB and Deng C: Targeted disruption of
SMAD3 results in impaired mucosal immunity and diminished T cell
responsiveness to TGF-beta. EMBO J. 18:1280–1291. 1999. View Article : Google Scholar
|
|
19
|
Xiao W, Chen X, Li W, Ye S, Wang W, Luo L
and Liu Y: Quantitative analysis of injury-induced anterior
subcapsular cataract in the mouse: A model of lens epithelial cells
proliferation and epithelial-mesenchymal transition. Sci Rep.
5:83622015. View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-delta delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
21
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar
|
|
22
|
Yamanaka O, Yuan Y, Coulson-Thomas VJ,
Gesteira TF, Call MK, Zhang Y, Zhang J, Chang SH, Xie C, Liu CY, et
al: Lumican binds ALK5 to promote epithelium wound healing. PloS
One. 8:e827302013. View Article : Google Scholar :
|
|
23
|
Saika S, Miyamoto T, Ishida I, Ohnishi Y
and Ooshima A: Osteopontin: A component of matrix in capsular
opacification and subcapsular cataract. Invest Ophthalmol Vis Sci.
44:1622–1628. 2003. View Article : Google Scholar
|
|
24
|
de Jong-Hesse Y, Kampmeier J, Lang GK and
Lang GE: Effect of extracellular matrix on proliferation and
differentiation of porcine lens epithelial cells. Graefes Arch Clin
Exp Ophthalmol. 243:695–700. 2005. View Article : Google Scholar
|
|
25
|
Banh A, Deschamps PA, Gauldie J, Overbeek
PA, Sivak JG and West-Mays JA: Lens-specific expression of TGF-beta
induces anterior subcapsular cataract formation in the absence of
Smad3. Invest Ophthalmol Vis Sci. 47:3450–3460. 2006. View Article : Google Scholar
|
|
26
|
Toole BP, Wight TN and Tammi MI:
Hyaluronan-cell interactions in cancer and vascular disease. J Biol
Chem. 277:4593–4596. 2002. View Article : Google Scholar
|
|
27
|
Saika S, Miyamoto T, Ishida I, Ohnishi Y
and Ooshima A: Lens epithelial cell death after cataract surgery. J
Cataract Refract Surg. 28:1452–1456. 2002. View Article : Google Scholar
|
|
28
|
van Roy F and Berx G: The cell-cell
adhesion molecule E-cadherin. Cell Mol Life Sci. 65:3756–3788.
2008. View Article : Google Scholar
|
|
29
|
Vincent T, Neve EP, Johnson JR, Kukalev A,
Rojo F, Albanell J, Pietras K, Virtanen I, Philipson L, Leopold PL,
et al: A SNAIL1 SMAD3/4 transcriptional repressor complex promotes
TGF-beta mediated epithelial-mesenchymal transition. Nat Cell Biol.
11:943–950. 2009. View
Article : Google Scholar
|
|
30
|
Postigo AA, Depp JL, Taylor JJ and Kroll
KL: Regulation of Smad signaling through a differential recruitment
of coactivators and corepressors by ZEB proteins. EMBO J.
22:2453–2462. 2003. View Article : Google Scholar :
|
|
31
|
Postigo AA: Opposing functions of ZEB
proteins in the regulation of the TGFbeta/BMP signaling pathway.
EMBO J. 22:2443–2452. 2003. View Article : Google Scholar
|
|
32
|
Chen ZF and Behringer RR: Twist is
required in head mesenchyme for cranial neural tube morphogenesis.
Genes Dev. 9:686–699. 1995. View Article : Google Scholar
|
|
33
|
Choi K, Lee K, Ryu SW, Im M, Kook KH and
Choi C: Pirfenidone inhibits transforming growth factor-β1-induced
fibrogenesis by blocking nuclear translocation of Smads in human
retinal pigment epithelial cell line ARPE-19. Mol Vis.
18:1010–1020. 2012.
|
|
34
|
Polydorou C, Mpekris F, Papageorgis P,
Voutouri C and Stylianopoulos T: Pirfenidone normalizes the tumor
microenvironment to improve chemotherapy. Oncotarget.
8:24506–24517. 2017. View Article : Google Scholar :
|
|
35
|
Papageorgis P, Polydorou C, Mpekris F,
Voutouri C, Agathokleous E, Kapnissi-Christodoulou CP and
Stylianopoulos T: Tranilast-induced stress alleviation in solid
tumors improves the efficacy of chemo- and nanotherapeutics in a
size-independent manner. Sci Rep. 7:461402017. View Article : Google Scholar :
|
|
36
|
Kaneyama T, Kobayashi S, Aoyagi D and
Ehara T: Tranilast modulates fibrosis, epithelial-mesenchymal
transition and peritubular capillary injury in unilateral ureteral
obstruction rats. Pathology. 42:564–573. 2010. View Article : Google Scholar
|
|
37
|
Robertson JV, Nathu Z, Najjar A, Dwivedi
D, Gauldie J and West-Mays JA: Adenoviral gene transfer of
bioactive TGFbeta1 to the rodent eye as a novel model for anterior
subcapsular cataract. Mol Vis. 13:457–469. 2007.
|
|
38
|
Pervan CL: Smad-independent TGF-β2
signaling pathways in human trabecular meshwork cells. Exp Eye Res.
158:137–145. 2017. View Article : Google Scholar
|
|
39
|
Secker GA, Shortt AJ, Sampson E, Schwarz
QP, Schultz GS and Daniels JT: TGFbeta stimulated
re-epithelialisation is regulated by CTGF and Ras/MEK/ERK
signalling. Exp Cell Res. 314:131–142. 2008. View Article : Google Scholar
|
|
40
|
Bakin AV, Tomlinson AK, Bhowmick NA, Moses
HL and Arteaga CL: Phosphatidylinositol 3-kinase function is
required for transforming growth factor beta-mediated epithelial to
mesenchymal transition and cell migration. J Biol Chem.
275:36803–36810. 2000. View Article : Google Scholar
|
|
41
|
Bhowmick NA, Ghiassi M, Bakin A, Aakre M,
Lundquist CA, Engel ME, Arteaga CL and Moses HL: Transforming
growth factor-beta1 mediates epithelial to mesenchymal
transdifferentiation through a RhoA-dependent mechanism. Mol Biol
Cell. 12:27–36. 2001. View Article : Google Scholar :
|
|
42
|
Bhowmick NA, Zent R, Ghiassi M, McDonnell
M and Moses HL: Integrin beta 1 signaling is necessary for
transforming growth factor-beta activation of 38MAPK and epithelial
plasticity. J Biol Chem. 276:46707–46713. 2001. View Article : Google Scholar
|
|
43
|
Shirai K, Saika S, Tanaka T, Okada Y,
Flanders KC, Ooshima A and Ohnishi Y: A new model of anterior
subcapsular cataract: Involvement of TGFbeta/Smad signaling. Mol
Vis. 12:681–691. 2006.
|
|
44
|
de Iongh RU, Wederell E, Lovicu FJ and
McAvoy JW: Transforming growth factor-beta-induced
epithelial-mesenchymal transition in the lens: A model for cataract
formation. Cells Tissues Organs. 179:43–55. 2005. View Article : Google Scholar
|
|
45
|
Chen X, Xiao W, Chen W, Luo L, Ye S and
Liu Y: The epigenetic modifier trichostatin A, a histone
deacetylase inhibitor, suppresses proliferation and
epithelial-mesenchymal transition of lens epithelial cells. Cell
Death Dis. 4:e8842013. View Article : Google Scholar
|
|
46
|
Liu K, Gao Z, Wu X, Zhou G, Zhang WJ, Yang
X and Liu W: Knocking out Smad3 favors allogeneic mouse fetal skin
development in adult wounds. Wound Repair Regen. 22:265–271. 2014.
View Article : Google Scholar
|
|
47
|
McDowell CM, Tebow HE, Wordinger RJ and
Clark AF: Smad3 is necessary for transforming growth factor-beta2
induced ocular hypertension in mice. Exp Eye Res. 116:419–423.
2013. View Article : Google Scholar
|
|
48
|
Maruno KA, Lovicu FJ, Chamberlain CG and
McAvoy JW: Apoptosis is a feature of TGF beta-induced cataract.
Clin Exp Optom. 85:76–82. 2002. View Article : Google Scholar
|
|
49
|
Lee JH, Wan XH, Song J, Kang JJ, Chung WS,
Lee EH and Kim EK: TGF-beta-induced apoptosis and reduction of
Bcl-2 in human lens epithelial cells in vitro. Curr Eye Res.
25:147–153. 2002. View Article : Google Scholar
|
|
50
|
Edlin RS, Tsai S, Yamanouchi D, Wang C,
Liu B and Kent KC: Characterization of primary and restenotic
atherosclerotic plaque from the superficial femoral artery:
Potential role of Smad3 in regulation of SMC proliferation. J Vasc
Surg. 49:1289–1295. 2009. View Article : Google Scholar
|
|
51
|
Tapia-González S, Muñoz MD, Cuartero MI
and Sanchez-Capelo A: Smad3 is required for the survival of
proliferative intermediate progenitor cells in the dentate gyrus of
adult mice. Cell Commun Signal. 11:932013. View Article : Google Scholar
|