Introduction
Periodontal ligament (PDL) is a connective tissue
that anchors the tooth root to the alveolar bone (1). PDL contains primary mechanosensitive
PDL (PM-PDL) neurons running into the trigeminal mesencephalic
nucleus (Vmes) (2). Following
their excitation by mechanical stimulation of the tooth, PM-PDL
neurons transmit information to the trigeminal ganglion, resulting
in the excitement of the ventral posteromedial nucleus of the
thalamus (3,4). PM-PDL neuron innervation comprises
Ruffini-like endings, which are partially covered by special
lamellar Schwann cells and complex basal lamina (5,6).
Thus, Ruffini-like endings function as a mechanosensor of the PDL
tissue, collecting information on the mechanical stress of PDL
influenced by the dental occlusal force and exciting the sense
perception of chewing and crunching in the brain. Uemura et
al (7) previously reported
that afferent neurons of jaw muscles running into Vmes also
function as a mechano-sensor of stress information during chewing
and crunching. In addition, immunohistological analysis revealed
that PDL neurons exhibit phosphorylated extracellular
signal-regulated kinases 1 and 2 (ERK1/2), which are found in
particular around blood vessels, suggesting that PDL neurons
regulate peripheral blood supply in PDL tissue in an
ERK1/2-dependent manner (8).
During occlusal trauma of PDL tissues,
mechanosensitive (MS) neurons are injured, resulting in atrophic
neurites and eventual degeneration of MS neurons (9). Nerve growth factor (NGF) is a
neurotrophic factor that is known to serve important roles in
neurite extension and regeneration in injured sensory neurons
(10,11). NGF serves as a potential guidance
cue for the axon outgrowth of dorsal root ganglion neurons through
tropomyosin receptor kinase A (TrkA). The primary subunit of NGF is
composed of 118 amino acids, as first identified in the mouse
submaxillary gland, and the native protein comprises two subunits
(12). Two types of receptors
with high and low affinities for NGF have been identified,
including TrkA, which is a receptor tyrosine kinase on the cell
membrane and has a high affinity for NGF, and p75 neurotrophin
receptor that has a low affinity for NGF (13).
Transforming growth factor β (TGF-β) is known to
play important roles in immunosuppression (14). In particular, TGF-β1 is involved
in the inhibition of renal inflammation progression in vitr
and in viv in a Smad7-dependent manner (15). TGF-β is mainly synthesized by
macrophages and secreted by these cells homing into inflammatory
tissues (16), and directly binds
to its type II receptors on the cell membrane. The type II receptor
kinase activates the type I receptor kinase following the formation
of a tetrameric complex composed of two type I and two type II
receptors. The activated type I receptor then induces intracellular
signal transduction through the phosphorylation of
receptor-regulated Smads (R-Smads) (17-19). Smads are major signaling molecules
of the TGF-β superfamily and comprise three groups as follows: i)
R-Smads, including Smad1, Smad5 and Smad8 that are primarily
activated by the bone morphogenetic protein-specific type I
receptors, as well as Smad2 and Smad3 that are activated by
TGF-β-specific type I receptors; ii) common mediator Smad
(Co-Smads), such as Smad4; and iii) inhibitory Smads (I-Smads),
such as Smad6 and Smad7. The activated R-Smads form complexes with
Co-Smad, which enter the nucleus and regulate the transcription of
specific target genes. Furthermore, I-Smads suppress the activation
of R-Smads by competing with R-Smads for type I receptor
interaction and recruiting specific ubiquitin ligases, resulting in
their proteasomal degradation.
TGF-β is also known to induce NGF expression in
various types of cells (20,21). In particular, TGF-β1 promotes NGF
expression in chondrocytes in a Smad2/3-dependent manner (22). By contrast, TGF-β relays its
intracellular signaling through non-Smad signaling pathways, such
as c-Jun N-terminal kinase (JNK) and p38 mitogen-activated protein
kinase (MAPK) signaling (23).
TGF-β1 promotes NGF expression in dental pulp cells through JNK and
p38 MAPK signal transduction (24). However, whether TGF-β induces the
expression of NGF in PDL fibroblasts through Smad and/or non-Smad
signaling pathways is questionable.
Inflammatory cytokines, including interleukin 1β
(IL-1β) and tumor necrosis factor α (TNF-α), are known to modulate
TGF-β-induced NGF expression. TGF-β1 and IL-1β cooperatively and
additively promote NGF production/secretion in the astroglial cell
line RC7 (25). A previous study
reported that IL-1β or TNF-α alone promoted NGF expression in the
mouse fibroblast cell line Swiss 3T3, and notably, these cytokines
synergistically promoted NGF expression in the cells (26). Intracellular signaling induced by
IL-1β and TNF-α activates nuclear factor-κB (NF-κB), JNK and p38
MAPK signaling pathways (27,28). Hengerer et al (29) reported that the c-Fos/c-Jun
heterodimer activator protein 1 activated the promoter of NGF gene,
while Lee et al (30)
demonstrated that cyclosporine A induced NGF expression in the
human corneal epithelial HCECL cell line in a p38 MAPK-dependent
manner. A study by Heese et al (31) further reported that NF-κB
activation was a key positive regulatory event for NGF expression
in B cells. These results suggest that IL-1β- and TNF-α-induced
activation of JNK, p38 MAPK and NF-κB positively regulates NGF
expression. However, the effect of IL-1β or TNF-α on NGF expression
in PDL fibroblasts remains to be elucidated.
In the present study, the effects of the
anti-inflammatory cytokine TGF-β1, the inflammatory cytokines IL-1β
or TNF-α, and the combination of inflammatory and anti-inflammatory
cytokines on the expression of NGF in PDL fibroblasts were
investigated. In addition, the study evaluated the effect of NGF
secreted by PDL fibroblasts on the status of neurite extension and
the expression of tyrosine hydroxylase (TH), which is a
rate-limiting enzyme in dopamine synthesis (32), in rat pheochromocytoma PC12 cells.
The current study clarified the molecular mechanisms underlying the
TGF-β-mediated regenerative effect of PDL fibroblasts on injured
and inflamed PDL neurons, and the findings may aid in identifying
drug targets for the treatment of PDL nerve injury at a molecular
level.
Materials and methods
Reagents
Recombinant human TGF-β1 was obtained from
PeproTech, Inc. (Rocky Hill, NJ, USA), recombinant rat NGF was
obtained from Alomone Labs (Jerusalem, Israel), and recombinant rat
IL-1β and TNF-α were purchased from Miltenyi Biotec GmbH (Bergisch
Gladbach, Germany). SB-431542, a TGF-β type I receptor inhibitor,
was supplied by Cell Signaling Technology, Inc. (Beverly, MA, USA).
The p38 MAPK inhibitor SB203580, JNK inhibitor SP600125, and Smad3
inhibitor SIS3 (33) were
obtained from Merck KGaA (Calbiochem; Darmstadt, Germany).
Adenosine 5′-O-(3-thio) triphosphate (ATPγS; ab138911) was
purchased from Abcam (Cambridge, UK). The TrkA inhibitor GW441756
was purchased from Selleck Chemicals (Houston, TX, USA), while the
NF-κB inhibitor BAY 11-7085 was from Cayman Chemical (Ann Arbor,
MI, USA).
Cell culture
The isolation of rat PDL-derived fibroblasts and the
establishment of single cell-derived cultures (SCDCs) have been
previously described (34). SCDC2
cells (1×105 cells) were cultured on type I
collagen-coated plastic dishes (Sumilon Celltight Dishes with 90 mm
diameters; Sumitomo Bakelite Co., Ltd., Tokyo, Japan) in Ham's F-12
(Sigma-Aldrich; Merck KGaA) medium supplemented with 2 mM glutamine
(100X solution; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), 10% fetal bovine serum (FBS), penicillin (50
units/ml)-streptomycin (50 µg/ml) (Invitrogen; Thermo Fisher
Scientific, Inc.), 10 ng/ml fibroblast growth factor (FGF)-1
(R&D Systems Inc., Minneapolis, MN, USA), and 15 µg/ml
heparin (Sigma-Aldrich; Merck KGaA) in a humidified atmosphere of
5% CO2 at 37°C. Rat pheochromocytoma PC12 cells obtained
from RIKEN BioResource Research Center Cell Bank (Ibaragi, Japan)
were cultured on type I collagen tissue culture plastic dishes in
Dulbecco's modified Eagle's medium (Sigma-Aldrich; Merck KGaA)
supplemented with 2 mM glutamine (100X solution), 5% FBS, 10% horse
serum and penicillin (50 units/ml)-streptomycin (50 µg/ml)
(Invitrogen; Thermo Fisher Scientific, Inc.) in a humidified
atmosphere of 5% CO2 at 37°C.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was isolated from SCDC2 cells with ISOGEN
I reagent (Nippon Gene Co., Ltd., Toyama, Japan) according to the
manufacturer's protocol. Concentration/quality of total RNA was
measured by using spectrophotometer Gene Spec III (Hitachi
High-Tech Science Corp., Tokyo, Japan). First-strand complementary
DNA (cDNA) was then synthesized from 1 µg of total RNA using
the PrimeScript RT reagent kit (Takara Bio, Inc., Shiga, Japan).
qPCR was subsequently performed on a Thermal Cycler Dice Real Time
System (two-step cycle procedure for 40 cycles: Denaturation at
95°C for 5 sec and annealing and extension at 60°C for 30 sec;
Takara Bio, Inc.) using SYBR Premix Ex Taq II (Takara Bio, Inc.)
with specific oligonucleotide primers, as follows: Rat TH,
5′-GGAGCTGAAGGCTTATGGTG-3′ (forward) and 5′-CATTGAAGCTCTCGGACACA-3′
(reverse); rat NGF, 5′-TGCCAAGGACGCAGCTTTC-3′ (forward) and
5′-TGAAGTTTAGTCCAGTGGGCTTCAG-3′ (reverse); and rat glyceraldehyde
3-phosphate dehydrogenase (GAPDH), 5′-GGCACAGTCAAGGCTGAGAATG-3′
(forward) and 5′-ATGGTGGTGAAGACGCCAGTA-3′ (reverse). The mRNA
levels of NGF and TH were normalized to the level of GAPDH, and the
relative expression levels of genes are expressed as the fold
increase or decrease relative to the control (35).
Effects of SB-431542, SB203580, SP600125,
and SIS3 on the TGF-β1-promoted expression of NGF mRNA in SCDC2
cells
The cells were seeded onto type I collagen-coated
12-well tissue culture plates at a density of 7×104
cells/well in SCDC2 growth medium and maintained for 24 h. The
growth medium was replaced with Ham's F-12 without FBS, FGF-1, and
heparin for 24 h for cell starvation. The cells were starved and
treated with or without TGF-β1 (10 ng/ml) for 24 h. Some cells were
pretreated with SB-431542 (10 µM), SB203580 (10 µM),
Smad3 inhibitor SIS3 (10 µM), or SP600125 (10 µM) for
30 min before TGF-β1 administration. The cells without pretreatment
were incubated with DMSO as a vehicle control. The relative
expression level of NGF was evaluated using RT-qPCR analysis, as
described above.
Effect of IL-1β and TNF-α on
TGF-β1-promoted expression of NGF mRNA in SCDC2 cells
The cells were seeded onto type I collagen-coated
12-well tissue culture plates at a density of 7×104
cells/well in SCDC2 growth medium and maintained for 24 h. The
growth medium was replaced with Ham's F-12 without FBS, FGF-1, and
heparin for 24 h for cell starvation. The cells were starved and
treated with or without TGF-β1 (10 ng/ml) alone, TGF-β1 (10 ng/ml)
plus IL-1β (10 ng/ml), or TGF-β1 (10 ng/ml) plus TNF-α (10 ng/ml)
for 24 h. Some cells were pretreated with BAY 11-7085 (10
µM) for 30 min before IL-1β or TNF-α administration. The
cells without pretreatment were incubated with dimethyl sulfoxide
as a vehicle control. The relative expression level of NGF was
evaluated using RT-qPCR analysis, as described above.
Western blot analysis
Cells were lysed in radioimmunoprecipitation assay
buffer [containing 50 mM Tris-HCl (pH 7.2), 150 mM NaCl, 1% NP-40,
0.5% sodium deoxycholate and 0.1% sodium dodecyl sulfate (SDS);
Sigma-Aldrich; Merck KGaA] or lysis buffer [including 20 mM HEPES
(pH 7.5), 150 mM NaCl, 1 mM EDTA and 1% Triton X-100] containing
protease and phosphatase inhibitor cocktails (Sigma-Aldrich; Merck
KGaA). The protein content of the samples was measured using a
bicinchoninic acid reagent (Pierce; Thermo Fisher Scientific,
Inc.). Samples containing equal amounts of protein were separated
on a 10% SDS-polyacrylamide gel by electrophoresis and then
transferred onto polyvinylidene difluoride membranes (EMD
Millipore, Bedford, MA, USA). Subsequent to blocking with 1% bovine
serum albumin or 1% skim milk in TBS/Tween-20 [containing 50 mM
Tris-HCl (pH 7.2), 150 mM NaCl and 0.05% Tween-20], the membranes
were incubated with the appropriate primary antibody for 24 h in
4°C. The following antibodies were used: Anti-Smad2/3 purified
mouse monoclonal antibody (1:1,000; cat. no. 610842; BD
Transduction Laboratories™; BD Biosciences, Franklin Lakes, NJ,
USA), as well as anti-phospho-Smad2/3, anti-p38 MAPK,
anti-phospho-p38 MAPK, anti-MAPK-activated protein kinase 2
(MAPKAPK-2), anti-phospho-MAPKAPK-2 (Thr222), anti-JNK and
anti-phospho-JNK rabbit antibodies (1:1,000; cat. nos. 8828, 9212,
9211, 12155, 3316, 9252S and 9251S, respectively; Cell Signaling
Technology, Inc.). Next, the blots were incubated with alkaline
phosphatase (AP)-conjugated secondary antibodies [anti-mouse IgG-AP
donkey antibody or anti-rabbit IgG-AP goat antibody (1:2,000; cat.
nos. sc-2320 and sc-2057, respectively; Santa-Cruz Biotechnology,
Inc.)] for 1 h at room temperature. Signals were detected using an
alkaline phosphatase substrate kit (BCIP/NBT Substrate kit; Vector
Laboratories, Inc., Burlingame, CA, USA), and then images of the
detected bands were captured. The expression level of the
phosphorylated target protein was compared with that of the total
target protein for evaluation of the phosphorylation status of the
target protein.
Enzyme-linked immunosorbent assay
(ELISA)
SCDC2 cells were seeded onto a type I
collagen-coated 6-well plate at a cell density of
1.4×105 cells/well with the aforementioned growth medium
for SCDC2 cells and cultured for 24 h. The growth medium was then
replaced with Ham's F-12 without any supplementation, and SCDC2
cells were stimulated with or without 10 ng/ml of TGF-β1, IL-1β or
TNF-α for 5 days. The amount of NGF secreted into the medium was
measured using Rat Beta-NGF ELISA kit (ab193736; Abcam) according
to the manufacturer's protocol.
Immunofluorescence analysis of cultured
cells
For immunofluorescence analysis of cultured cells,
SCDC2 cells (2×104 cells) and PC12 cells
(1×104 cells) were subcultured on non-coated 4-well
glass culture slides and maintained in Ham's F-12 supplemented with
2 mM glutamine, penicillin-streptomycin and 10% FBS. Cells were
cultured with or without TrkA inhibitor (10 µM), SB-431542
(10 µM), TGF-β1 (10 ng/ml), TNF-α (10 ng/ml) and IL-1β (10
ng/ml) for 4 days. Following incubation, cells were fixed in 4%
paraformaldehyde (Nacalai Tesque, Inc., Kyoto, Japan) for 15 min
and permeabilized with Triton X-100 (Sigma-Aldrich; Merck KGaA).
Following background reduction with normal goat serum, the cells
were incubated with the anti-Smad2/3 purified mouse monoclonal
antibody (1:200), as well as the anti-NF-κB p65 (D14E12) rabbit and
anti-neurofilament H (RMdO 20) mouse monoclonal antibodies (1:200
and 1:1,000, respectively; cat. nos. 8242 and 2836S, respectively;
Cell Signaling Technology, Inc.) at room temperature for 1 h. After
washing with phosphate-buffered saline (PBS) to remove any excess
primary antibody, the cells were incubated with Alexa Fluor
568-conjugated goat anti-rabbit IgG or Alexa Fluor 568-conjugated
goat anti-mouse IgG, as appropriate (1:400; A-11011 and A-11031,
respectively; Molecular Probes; Thermo Fisher Scientific, Inc.).
Subsequent to washing further with PBS to remove any excess
secondary antibody, certain cells were stained with Alexa Fluor
488-conjugated phalloidin (1:1,000; A12379; Thermo Fisher
Scientific, Inc.), which specifically detects F-actin. Next, nuclei
were stained with the 4′,6-diamidino-2-phenylindole
(DAPI)-containing mounting medium, DAPI Fluoromount-G
(SouthernBiotech, Birmingham, AL, USA). The fluorescent signal was
detected using an Olympus IX70 fluorescence microscope with the
LCPIanFI 20 objective lens (Olympus Corp., Tokyo, Japan).
Evaluation of neurite extension from PC12
cells co-cultured with SCDC2 cells and TH expression in the PC12
cells
SCDC2 cells (2×104) and PC12 cells
(1×104) were mixed and seeded into each well of a
non-coated 4-well glass culture slide (Thermo Fisher Scientific,
Inc.) with the aforementioned PC12 cell growth medium. The cells
were subsequently incubated with or without TGF-β1 (10 ng/ml),
IL-1β (10 ng/ml), TNF-α (10 ng/ml), SB-431542 (10 µM) and
TrkA inhibitor GW441756 (2 nM) for 5 days. In the ATPγS-treated
PC12 cell group, ATPγS (100 µM) was added to all cultures
from the beginning of the co-culture. PC12 cells in the co-culture
were specifically and immunofluorescently labeled with
anti-neurofilament H antibody. The status of neurite extensions
emerging from PC12 cells was observed using an Olympus IX70
fluorescence microscope with the LCPIanFI 20 objective lens
(Olympus Corp.). In addition, statistical assessment of neurite
extension was performed as follows: Cells bearing neurites longer
than the length of the cell body were counted as cells with neurite
extension. Neurite-extended and non-extended PC12 cells were
counted in eight different fluorescent microscopic fields. The
ratio of the number of neurite-extended cells to the number of
total PC12 cells in the fluorescent microscopic field was
statistically analyzed (n=8).
In order to evaluate the TH expression status at the
mRNA level in the PC12 cells co-cultured with SCDC2 cells as
described earlier, mRNA was extracted from the co-culture without
separating PC12 and SCDC2 cells. TH is known as a specific marker
for the identification of dopaminergic neuron (32), therefore it was hypothesized that
PDL-derived fibroblasts SCDC2 cells are unlikely to express TH.
Statistical analysis
The data are presented as the mean ± standard
deviation (n=6 or 8) and were statistically analyzed by Tukey's
multiple comparison test. A value of P<0.05 was considered to
indicate a difference that was statistically significant. The
results presented are representative of at least three separate
experiments.
Results
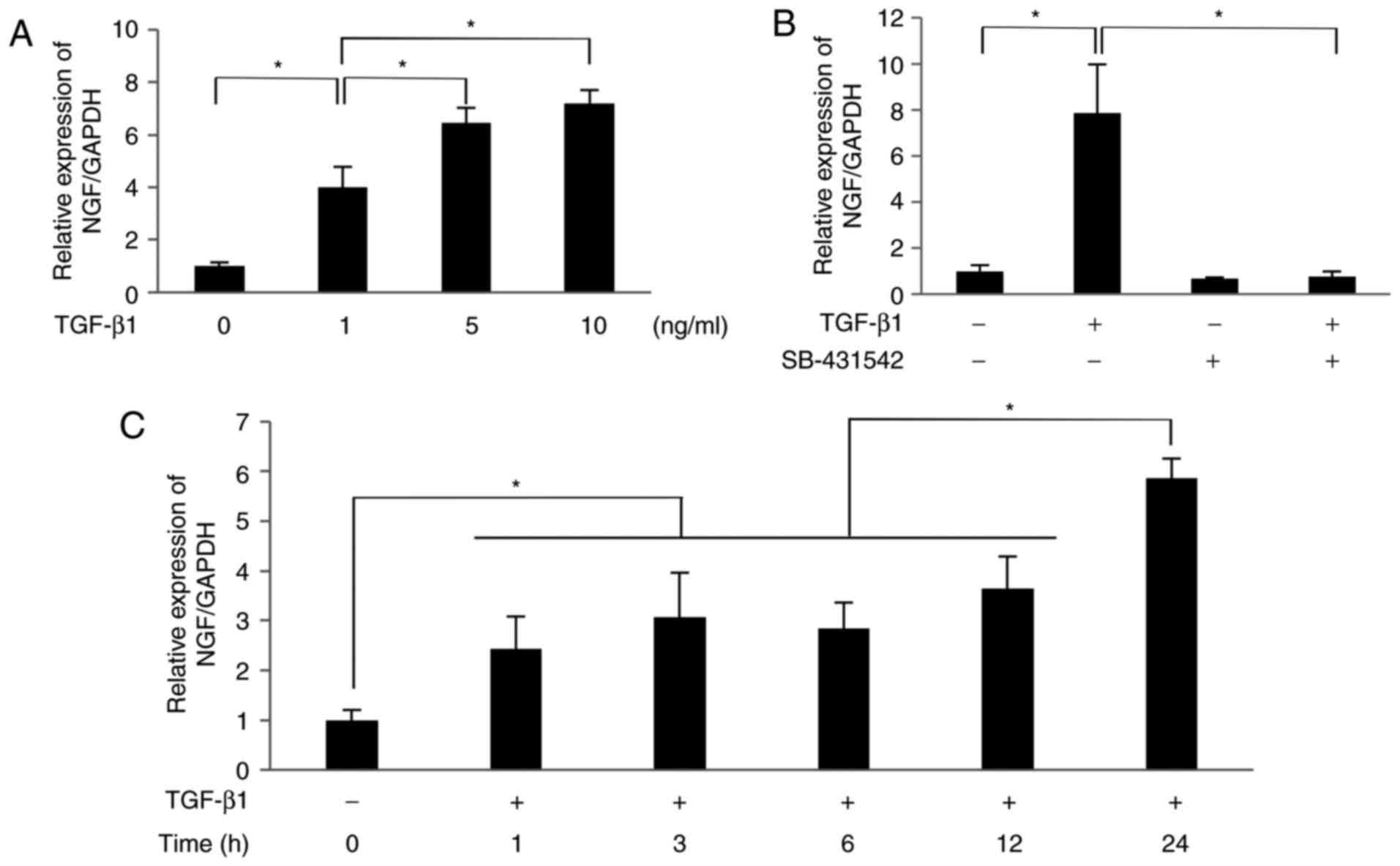
TGF-β1 promotes the expression of NGF
mRNA in SCDC2 cells through its type I receptor kinase activity in
a dose-dependent manner
The results of RT-qPCR analysis revealed that TGF-β1
(1-10 ng/ml) significantly increased the mRNA expression level of
NGF in SCDC2 cells in a dose-dependent manner (1 ng/ml, 4.0-fold
increase; 5 ng/ml, 6.5-fold increase; 10 ng/ml, 7.2-fold increase;
Fig. 1A). The selective inhibitor
of TGF-β type I receptor SB-431542 (10 µM) completely
suppressed NGF mRNA expression induced by 10 ng/ml TGF-β1 (Fig. 1B). These results indicate that
TGF-β1 upregulated the mRNA expression of NGF in PDL-derived
fibroblasts through its type I receptor kinase activity. As shown
in Fig. 1C, TGF-β1 (10 ng/ml)
significantly increased the mRNA level of NGF in SCDC2 cells in a
time-dependent manner, with a 2.4-fold, 3.1-fold, 2.8-fold,
3.7-fold and 5.9-fold increase observed at 1, 3, 6, 12 and 24
h.
TGF-β1 promotes the expression of NGF
mRNA in SCDC2 cells in Smad2/3-dependent and p38 MAPK-dependent
manners
Treatment with TGF-β1 (10 ng/ml) significantly
increased the mRNA expression level of NGF in SCDC2 cells by
10-fold (Fig. 2A). The
TGF-β1-mediated increase in NGF mRNA was completely suppressed by
the Smad3 inhibitor SIS3 (10 µM; Fig. 2A). In addition, TGF-β1-mediated
increase in NGF mRNA expression was markedly suppressed by the p38
MAPK inhibitor SB203580 (10 µM; Fig. 2B). It was also confirmed that
TGF-β1 upregulated the phosphorylation of Smad2/3 and p38 MAPK in
SCDC2 cells within 15 and 30 min of administration, respectively
(Fig. 2C). Furthermore, the
inhibitor SIS3 completely abrogated TGF-β1-induced nuclear
translocation of Smad2/3, as evident from the immuno-fluorescence
analysis (Fig. 2D). The results
further confirmed the effect of SB203580 on the phosphorylation
status of MAPKAPK-2, which is known as the direct target of p38
MAPK (Fig. 2E). The
phosphorylation level of MAPKAPK-2 was upregulated at 30 min
following 10 ng/ml TGF-β1 administration. SB203580 inhibited the
TGF-β1-induced MAPKAPK-2 phosphorylation, indicating that this
inhibitor attenuated p38 MAPK-mediated signaling activated by
TGF-β1. Although the phosphorylation status of JNK was not at a
detectable level following TGF-β1 stimulation (data not shown), the
JNK inhibitor SP600125 (10 µM) partially and significantly
suppressed the TGF-β1-mediated increase in NGF mRNA expression
(Fig. 2F).
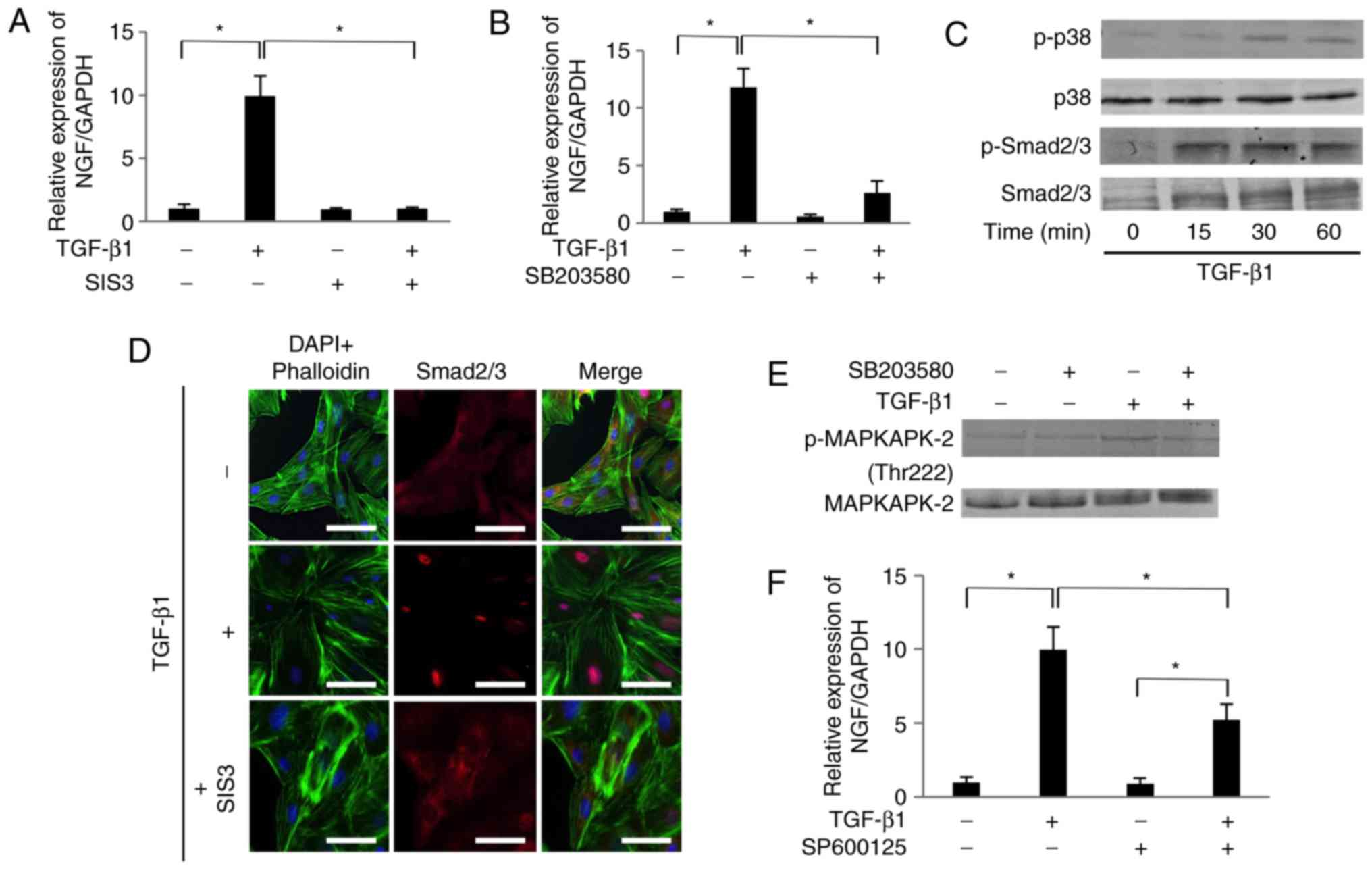 | Figure 2TGF-β1 promoted the mRNA expression
of NGF in SCDC2 cells in Smad2/3-dependent and p38 MAPK-dependent
manners. Effects of (A) SIS3 (10 µM), and (B) SB203580 (10
µM) on expression of NGF mRNA were evaluated as described in
Materials and methods. Data represent the mean ± standard deviation
(n=6). *P<0.05. (C) Phosphorylation status of Smad2/3
and p38 MAPK in cells stimulated with TGF-β1 (10 ng/ml) for the
indicated times, evaluated using western blot analysis. (D) After
24-h starvation, cells were pretreated with Smad3 inhibitor SIS3
(10 µM) for 30 min and then treated with or without TGF-β1
(10 ng/ml) for 30 min, and the status of nuclear translocation of
Smad2/3 following TGF-β1 stimulation was examined using
immunofluorescence analysis (×200 magnification; scale bar, 50
µm). (E) Phosphorylation status of MAPKAPK-2 evaluated using
western blot analysis in cells stimulated with TGF-β1 (10 ng/ml)
and/or with the inhibitor SB203580. (F) Effect of SP600125 (10
µM) on expression of NGF mRNA was evaluated as described in
Materials and methods. Data represent the mean ± standard deviation
(n=6). *P<0.05. TGF, transforming growth factor; NGF,
nerve growth factor; SCDC, single cell-derived culture; RT-qPCR,
reverse transcription-quantitative polymerase chain reaction; MAPK,
mitogen-activated protein kinase; MAPKAPK-2, MAPK-activated protein
kinase 2. |
IL-1β and TNF-α suppress the
TGF-β1-induced secretion of NGF in SCDC2 cells by abrogating the
activities of Smad2/3 and p38 MAPK
As shown in Fig. 3A
and B, IL-1β (1-10 ng/ml) or TNF-α (1-10 ng/ml) failed to
significantly affect the mRNA expression level of NGF in SCDC2
cells in the absence of TGF-β1. However, both IL-1β (10 ng/ml) and
TNF-α (10 ng/ml) were able to significantly suppress the
TGF-β1-mediated NGF mRNA expression (Fig. 3C and D, respectively).
Additionally, NF-κB inhibitor BAY 11-7085 (10 µM) did not
reverse the IL-1β-induced and TNF-α-induced suppression of the
TGF-β1-promoted NGF mRNA expression (data not shown). Intriguingly,
IL-1β (10 ng/ml) and TNF-α (10 ng/ml) significantly suppressed the
TGF-β1-induced phosphorylation of Smad2/3 and p38 MAPK,
particularly at 60 min after stimulation (Fig. 3E and F, respectively). The study
also evaluated the protein level of NGF in the conditioned medium
of SCDC2 cells using ELISA. TGF-β1 (10 ng/ml) treatment increased
the concentration of NGF from 0.5 to 5 ng/ml in SCDC2 conditioned
medium. However, IL-1β (10 ng/ml) and TNF-α (10 ng/ml)
significantly decreased the TGF-β1-mediated increase in the NGF
protein level from 5 ng/ml to 2.5 and 3.8 ng/ml, respectively, in
the conditioned medium (Fig. 3G).
Immunofluorescence analysis further revealed that IL-1β (10 ng/ml)
and TNF-α (10 ng/ml) induced the nuclear trans-location of NF-κB
from the cytoplasm (Fig. 3H),
indicating the induction of signal transduction in SCDC2 cells
following stimulations with IL-1β and TNF-α.
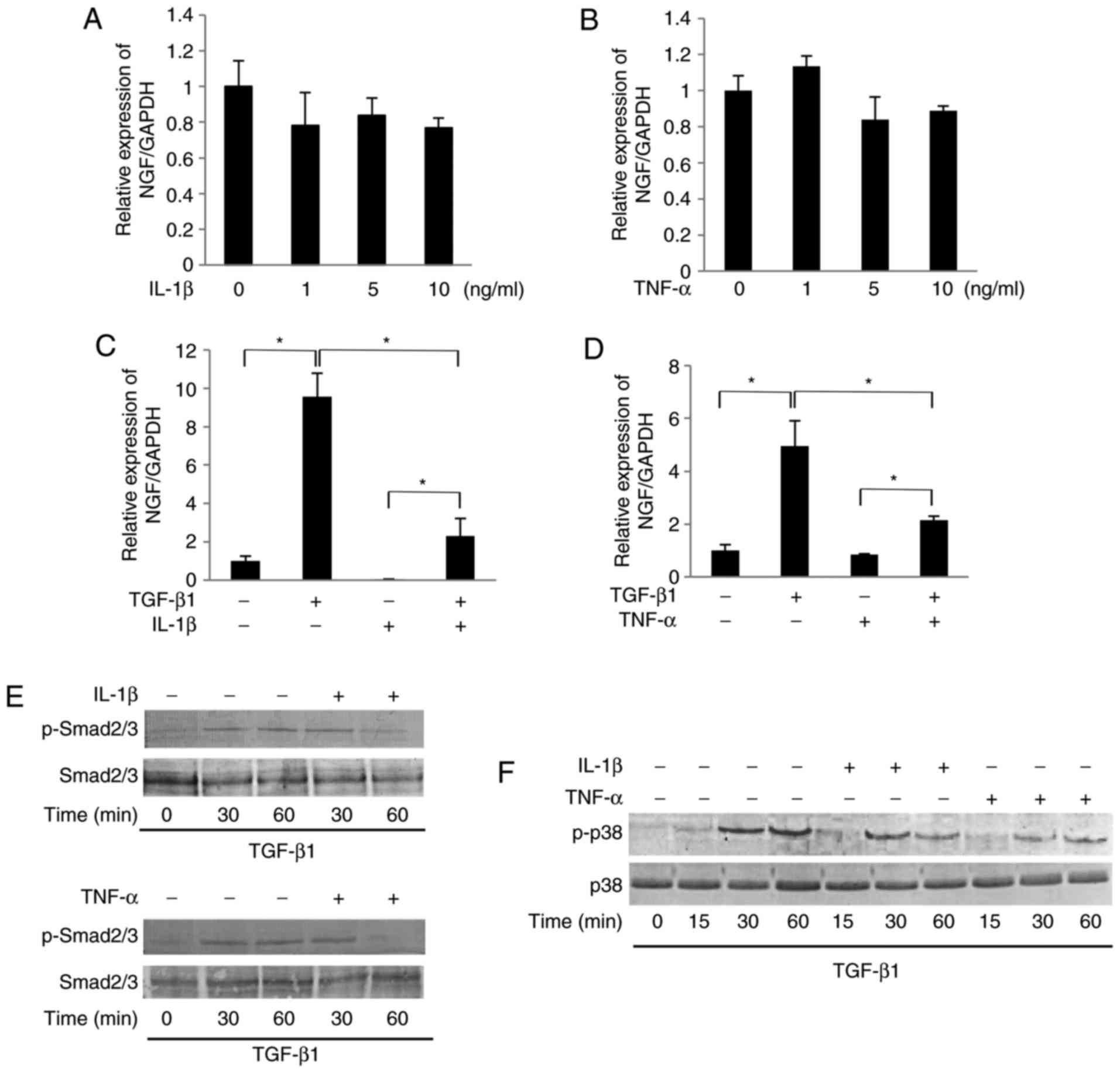 | Figure 3IL-1β and TNF-α suppressed the
TGF-β1-induced mRNA expression of NGF in SCDC2 cells by abrogating
Smad2/3 and p38 MAPK activities. The effects of IL-1β and TNF-α on
TGF-β1-induced mRNA expression of NGF in SCDC2 cells were evaluated
using RT-qPCR. The cells were treated with or without (A) IL-1β
alone or (B) TNF-α alone at indicated concentrations, (C) TGF-β1
(10 ng/ml) and/or IL-1β (10 ng/ml), and (D) TGF-β1 (10 ng/ml)
and/or TNF-α (10 ng/ml). Data represent the mean ± standard
deviation (n=6). *P<0.05. Phosphorylation status of
(E) Smad2/3 and (F) p38 MAPK was evaluated using western blot
analysis in cells treated with or without TGF-β1 (10 ng/ml) alone,
TGF-β1 (10 ng/ml) + IL-1β (10 ng/ml), or TGF-β1 (10 ng/ml) + TNF-α
(10 ng/ml) for the indicated times. (G) NGF protein concentration
secreted into the culture medium was determined using ELISA in
cells cultured with or without TGF-β1 (10 ng/ml) alone, TGF-β1 (10
ng/ml) + IL-1β (10 ng/ml), or TGF-β1 (10 ng/ml) + TNF-α (10 ng/ml)
for 5 days. (H) Nuclear translocation status of NF-κB p65 (red) was
evaluated using immunofluorescence analysis (blue, nuclei; green,
filamentous actin) in SCDC2 cells treated with or without IL-1β (10
ng/ml) or TNF-α (10 ng/ml) for 24 h (×200 magnification; scale bar,
50 µm). IL, interleukin; TNF, tumor necrosis factor; TGF,
transforming growth factor; NGF, nerve growth factor; SCDC, single
cell-derived culture; MAPK, mitogen-activated protein kinase. |
NGF secreted by SCDC2 cells upon
stimulation with TGF-β1 promotes neurite extension from the surface
of PC12 cells treated with ATPγS
The present study then examined whether NGF secreted
by the TGF-β1-treated SCDC2 cells retained the neurotrophic
activity to induce neurite extension from the surface of PC12
cells. Arthur et al (36)
reported that ATPγS enhanced the sensitivity of PC12 cells to NGF
stimulation through P2Y2 receptor-mediated activation of
ERK1/2 and p38 MAPK. As a consequence, NGF-induced neurite
extension from the surface of PC12 cells was accelerated (36). In the current study, the
neurogenerative effect of NGF secreted by SCDC2 cells upon TGF-β1
stimulation was evaluated, and whether the secreted NGF promoted
neurite extension from the surface of PC12 cells was investigated
using a co-culture system. ATPγS-treated PC12 cells co-cultured
with non-treated SCDC2 cells failed to exhibit neurite extension
(Fig. 4A). However, ATPγS-treated
PC12 cells co-cultured with TGF-β1-treated SCDC2 cells clearly
exhibited neurite outgrowth (Fig.
4A). In addition, SB-431542 and GW441756 suppressed both the
morphological change of PC12 cells from round shape to elongated
shape and the neurite extension from the surface of PC12 cells
induced by the SCDC2-secreted NGF (Fig. 4A). By contrast, IL-1β and TNF-α
did not completely suppress the morphological changes of PC12 cells
from round shape to elongated shape, whereas they clearly
suppressed neurite extension from the surface of ATPγS-treated PC12
cells induced by the SCDC2-secreted NGF (Fig. 4A). In addition, the results
confirmed that ATPγS itself did not affect the expression level of
NGF mRNA in the SCDC2 cells using RT-qPCR analysis (data not
shown). Statistical assessment of neurite extension in PC12 cells
co-cultured with SCDC2 cells was also performed. Approximately 50%
of ATPγS-treated PC12 cells co-cultured with TGF-β1-stimulated
SCDC2 cells exhibited neurite extension, whereas none of the
ATPγS-treated PC12 cells co-cultured with non-stimulated SCDC2
cells exhibited neurite growth (Fig.
4B). Furthermore, SB-431542 (10 µM) and TrkA inhibitor
GW441756 (2 nM) significantly reduced the ratio of neurite-extended
PC12 cells to total PC12 cells, which were activated by treatment
with ATPγS (100 µM) and co-cultured with TGF-β1-treated
SCDC2 cells (from 50.7% to 7.8 and 7.5., respectively).
Furthermore, IL-1β and TNF-α significantly reduced the ratio of
neurite-extended PC12 cells to total PC12 cells, which were
activated by treatment with ATPγS (100 µM) and co-cultured
with TGF-β1-stimulated SCDC2 cells (from 47.9% to 9.4 and 8.9%,
respectively).
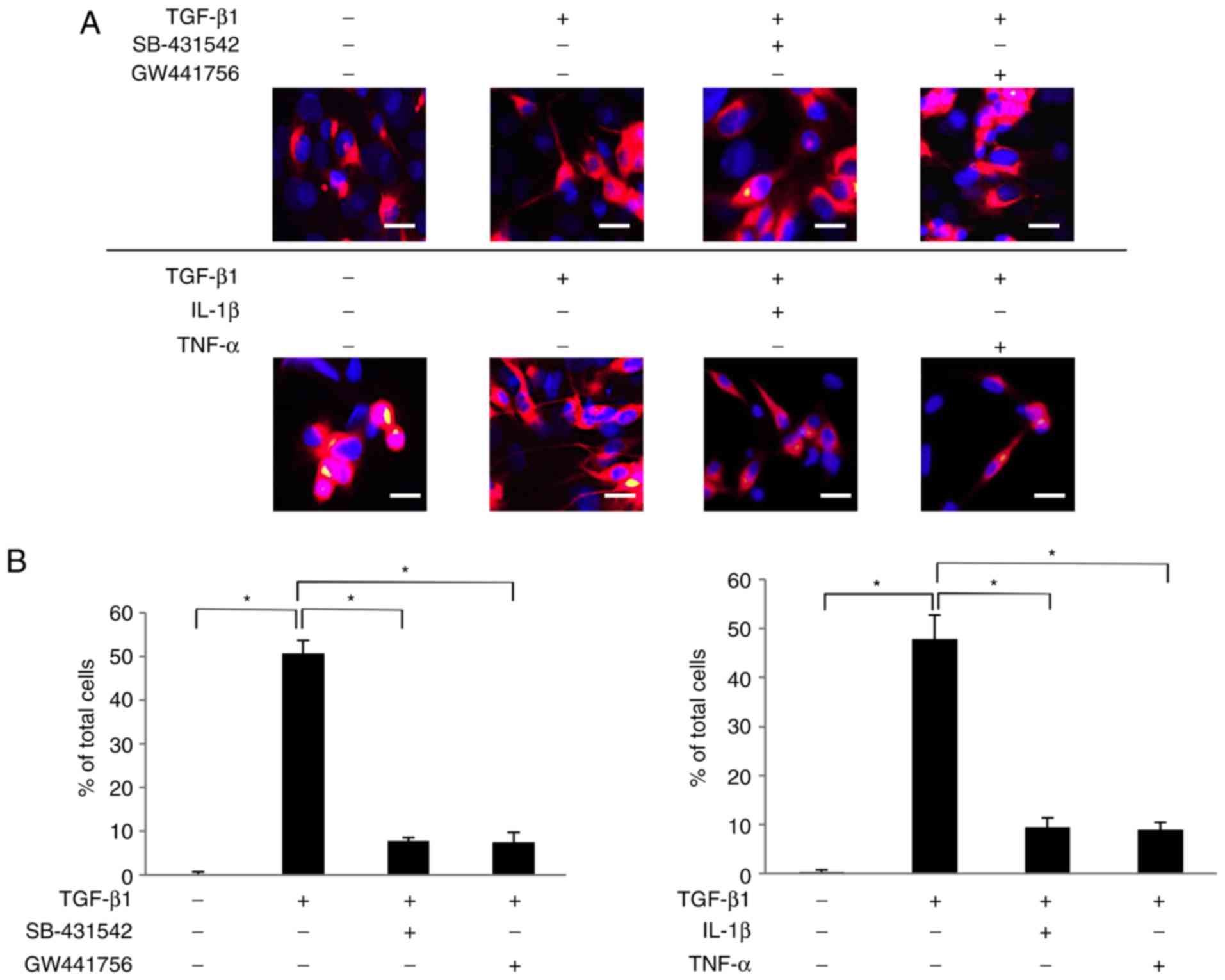 | Figure 4Nerve growth factor secreted by SCDC2
cells following TGF-β1 stimulation promoted neurite extension from
the surface of ATPγS-treated PC12 cells. (A) Neurite extension of
PC12 cells was visualized by immunostaining (×200 magnification;
scale bar, 50 µm) with anti-neurofilament H antibody (red)
and nuclei were stained with DAPI (blue). SCDC2 cells
(2×104 cells) and rat pheochromocytoma cells PC12
(1×104 cells) were co-cultured and treated with or
without TGF-β1 (10 ng/ml) for 4 days. Cells were also treated with
TGF-β type I receptor inhibitor SB-431542 (10 µM), TrkA
inhibitor GW441756 (2 nM), IL-1β (10 ng/ml), or TNF-α (10 ng/ml)
from the beginning of the co-culture. In addition, ATPγS (100
µM) was added to all cultures during cell seeding. Dimethyl
sulfoxide was added to cell cultures as a vehicle control for
SB-431542 and GW441756, respectively. (B) Statistical assessment of
neurite extension in PC12 cells co-cultured with SCDC2 cells. Data
represent the mean ± standard deviation (n=8).
*P<0.05. TGF, transforming growth factor; SCDC,
single cell-derived culture; IL, interleukin; TNF, tumor necrosis
factor; ATPγS, adenosine 5′-O-(3-thio)triphosphate; TrkA,
tropomyosin receptor kinase A. |
NGF secreted by SCDC2 cells upon TGF-β1
stimulation promotes mRNA expression of TH in PC12 cells
The study further evaluated how NGF secreted by
TGF-β1-treated SCDC2 cells affected the expression status of TH,
which is a rate-limiting enzyme in dopamine synthesis (32), in PC12 cells. As shown in Fig. 5A, the mRNA expression level of TH
in PC12 cells co-cultured with TGF-β1 (10 ng/ml)-treated SCDC2
cells was 3.0 times higher than that in PC12 cells co-cultured with
non-treated SCDC2 cells at 24 h after initiation of the co-culture.
In addition, ATPγS (100 µM) significantly enhanced the TH
expression in PC12 cells co-cultured with TGF-β1-treated SCDC2
cells compared with cells cultured with TGF-β1 or ATPγS alone
(Fig. 5A). Furthermore, treatment
with the TGF-β type I receptor inhibitor SB-431542, TrkA inhibitor
GW441756, IL-1 or TNF-α significantly suppressed the
TGF-β1-promoted expression of TH (Fig. 5B). Thus, NGF secreted from SCDC2
cells stimulated with TGF-β1 exerted a neurofunctional activity in
PC12 cells.
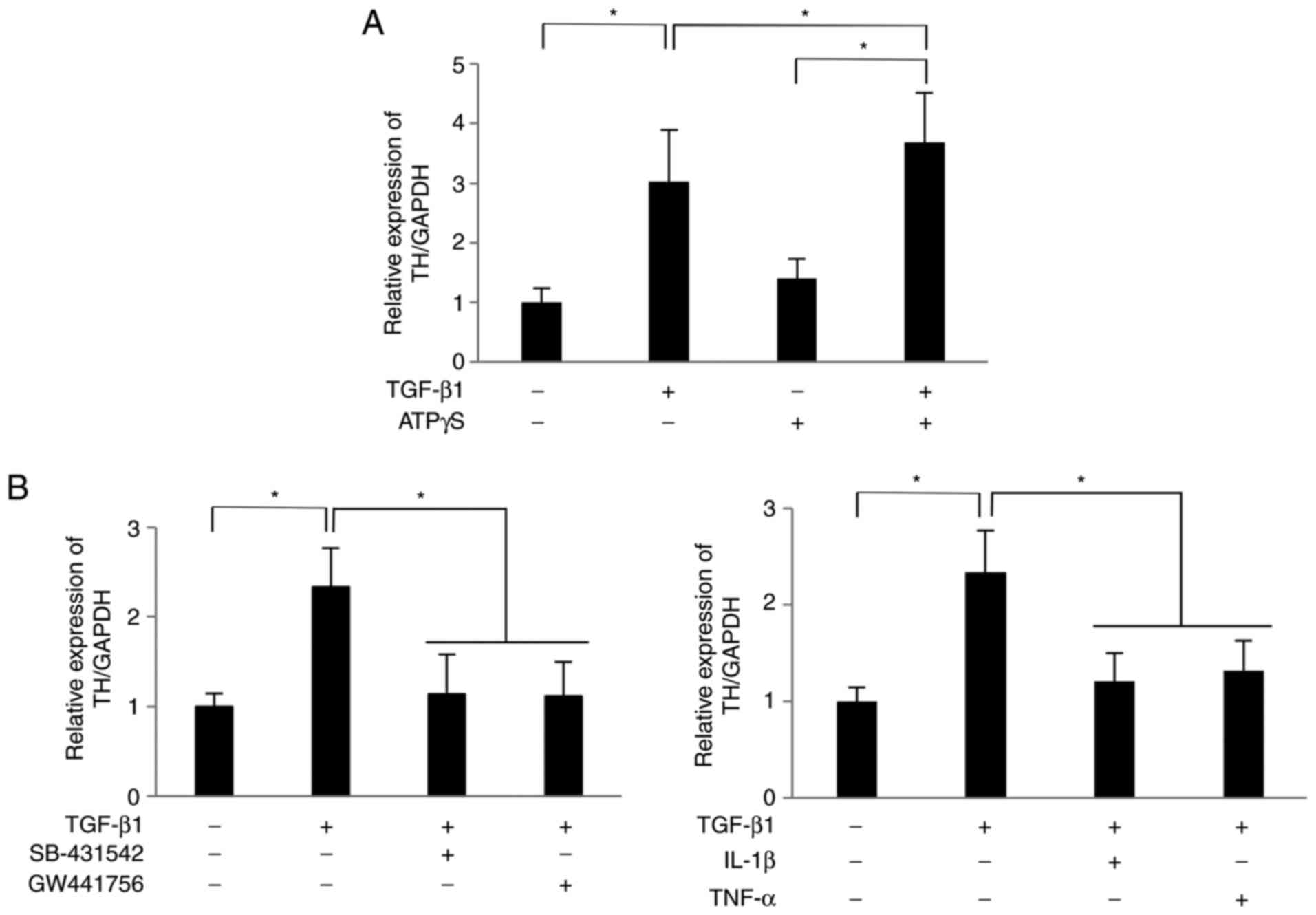 | Figure 5Nerve growth factor secreted by SCDC2
cells subsequent to TGF-β1 stimulation promoted the expression of
TH mRNA in PC12 cells. SCDC2 cells (7×104 cells) and rat
pheochromocytoma cells PC12 cells (3.5×104 cells) were
co-cultured and stimulated with or without TGF-β1 (10 ng/ml) for 24
h. The relative expression level of TH was evaluated using reverse
transcription-quantitative polymerase chain reaction in cells also
treated with (A) ATPγS (100 µM), and with (B) TGF-β type I
receptor inhibitor SB-431542 (10 µM), TrkA inhibitor
GW441756 (2 nM), IL-1β (10 ng/ml) or TNF-α (10 ng/ml) during the
co-culture. Dimethyl sulfoxide was added to cell cultures as a
vehicle control for SB-431542 and GW441756, respectively. Data
represent the mean ± standard deviation (n=6).
*P<0.05. TGF, transforming growth factor; SCDC,
single cell-derived culture; IL, interleukin; TNF, tumor necrosis
factor; ATPγS, adenosine 5′-O-(3-thio)triphosphate; TrkA,
tropomyosin receptor kinase A; TH, tyrosine hydroxylase. |
Discussion
The present study demonstrated that TGF-β1
significantly enhanced the expression levels of NGF mRNA and
protein in rat PDL fibroblasts SCDC2 (Figs. 1 and 3G, respectively). In addition, TGF-β1
upregulated the phosphorylation levels of Smad2/3 and p38 MAPK
(Fig. 2C). It was also reported
that the Smad3 inhibitor SIS3 and the p38 MAPK inhibitor SB203580
(10 µM) significantly abrogated TGF-β1-induced upregulation
of NGF expression in SCDC2 cells (Fig. 2A and B). These results strongly
suggest that TGF-β1 upregulated NGF expression in PDL fibroblasts
in Smad2/3-dependent and p38 MAPK-dependent manners. Next, the
effect of the inflammatory cytokine on TGF-β-mediated expression of
NGF was evaluated. Hattori et al (26) previously reported that IL-1β and
TNF-α promoted NGF expression in mouse fibroblasts, whereas IL-1β
or TNF-α alone failed to exhibit any effect on NGF expression in
SCDC2 cells in the current study (Fig. 3A and B). In addition, Hahn et
al (25) demonstrated that
TGF-β1 and IL-1β cooperatively and additively promoted NGF
production/secretion in astroglial cells. The present study
revealed that the pro-inflammatory cytokines IL-1β and TNF-α
presented a partial but significant inhibition of TGF-β1-mediated
NGF expression (Fig. 3C and D,
respectively). These results indicate that the differential effects
of pro-inflammatory cytokines on NGF expression may vary according
to the cell type. In addition, IL-1β and TNF-α downregulated the
phosphorylation levels of Smad2/3 and p38 MAPK (Fig. 3E and F, respectively). These
results strongly suggest that IL-1β and TNF-α suppressed the
TGF-β1-induced expression of NGF mRNA in SCDC2 cells by abrogating
Smad2/3 and p38 MAPK activities. It was also confirmed that
TGF-β1-mediated secretion of NGF by SCDC2 cells was partially but
significantly inhibited by IL-1β and TNF-α (Fig. 3G).
NF-κB signaling is known to induce Smad7, a negative
regulator of TGF-β signaling (37). Therefore, the current study
examined the effect of the NF-κB inhibitor BAY 11-7085 on
IL-1β-induced and TNF-α-induced suppression of the TGF-β1-promoted
expression of NGF. BAY 11-7085 (10 µM) did not reverse the
IL-1β-induced and TNF-α-induced suppression of the TGF-β1-promoted
expression of NGF (data not shown). These results suggested that
IL-1β and TNF-α suppressed the TGF-β1-promoted expression of NGF
through the activation of signal transduction molecules other than
NF-κB.
The present study also investigated how JNK affected
the TGF-β1-mediated NGF expression. Although JNK was not
phosphorylated following TGF-β1 stimulation (data not shown), JNK
inhibitor SP600125 significantly and partially suppressed the
TGF-β1-mediated expression of NGF mRNA (Fig. 2F), suggesting that the basal level
of JNK activity may be important for the TGF-β1-mediated NGF
expression in SCDC2 cells.
The rat pheochromocytoma PC12 cell line has been
established and used as a model for the growth and differentiation
of neural crest cells (38).
Furthermore, the neural crest cells are known to differentiate into
various types of cells, which construct the neural network in the
orofacial region during the mouse embryonic development, such as
sensory and autonomic nerves, ganglia, Schwann cells and cells of
the dental pulp involving odontoblasts (39). Therefore, the neurotrophic effect
of the secreted NGF from PDL fibroblasts on the status of neurite
extension in PC12 cells may reflect the NGF effect on PM-PDL
neurons running into Vmes. The present study reported that ~50% of
ATPγS-treated PC12 cells exhibited neurite extension when
co-cultured with TGF-β1-stimulated SCDC2 cells, whereas this was
not detected in the ATPγS-treated PC12 cells that were co-cultured
with non-stimulated SCDC2 cells (Fig.
4A and B). In addition, TGF-β type I receptor inhibitor
SB-431542 clearly abrogated the neurite extension from the
ATPγS-treated PC12 cells co-cultured with TGF-β1-stimulated SCDC2
cells. However, TGF-β1 alone failed to promote neurite extension
from the ATPγS-treated PC12 cells in the absence of co-culture
(data not shown). The inhibitor of TrkA (GW441756) clearly
abrogated the neurite extension from the ATPγS-treated PC12 cells
co-cultured with TGF-β1-stimulated SCDC2 cells. Thus, NGF secreted
from TGF-β1-treated SCDC2 cells appeared to retain a physiological
activity as a neurotrophic factor.
By contrast, the mRNA expression level of TH in PC12
cells co-cultured with TGF-β1 (10 ng/ml)-treated SCDC2 cells was
3.0-fold higher in comparison with that in PC12 cells co-cultured
with non-treated SCDC2 cells (Fig.
5A). Furthermore, treatment with TGF-β type I receptor
inhibitor SB-431542, TrkA inhibitor GW441756, IL-1 or TNF-α
significantly suppressed the TGF-β1-promoted expression of TH
(Fig. 5B). These results
suggested that NGF secreted from the TGF-β1-treated SCDC2 cells
upregulated the mRNA expression of TH in PC12 cells, which was not
inconsistent with the findings of a previous study reporting that
NGF induced TH expression at the transcriptional level in PC12
cells (40). Thus, NGF secreted
from SCDC2 cells stimulated with TGF-β1 exerted a neurofunctional
activity in PC12 cells.
During occlusal trauma of PDL tissues, MS neurons
are injured, and their neurites undergo atrophy and eventually
degenerate. As an inflammatory response, the injured PDL tissues
recruit inflammatory blood cells involving pro-inflammatory M1
macrophages and anti-inflammatory M2 macrophages that advance and
complete inflammation, respectively (41). M1 macrophages synthesize and
secrete pro-inflammatory cytokines, including IL-1β and TNF-α,
whereas M2 macrophages synthesize and secrete anti-inflammatory
cytokines, such as TGF-β1 and IL-10 (42). TGF-β1 derived from M2 macrophages
stimulates PDL fibroblasts in the inflammatory PDL tissue,
resulting in the upregulation of NGF synthesis and secretion by PDL
fibroblasts. NGF secreted from TGF-β1-stimulated PDL fibroblasts
subsequently and neurotrophically regenerates the injured MS
neurons in the damaged PDL tissue. IL-1β and TNF-α derived from M1
macrophages suppress TGF-β1-mediated secretion of NGF from PDL
fibroblasts. Therefore, the inhibition of IL-1β and TNF-α signaling
in the fibroblasts from the inflamed PDL tissue may be an efficient
therapeutic strategy for NGF-mediated regeneration of injured PDL
neurons. Neutralizing agents against IL-1β (such as canakinumab)
(43) and TNF-α (including
infliximab, adalimumab, golimumab and etanercept) (44) are clinically used for the
treatment of chronic inflammatory diseases and may be available for
the regenerative therapy of injured PDL neurons. Furthermore, Li
et al (45) reported that
the transplantation of PDL-derived mesenchymal cells exhibited a
potential value in the repair of the crush-injured left mental
nerve in rats. Hence, the possibility of the establishment of
regenerative medicine for the damaged neurons by transplantation of
PDL-derived fibroblasts treated with TGF-β1 following the
administration of neutralizing agents against IL-1β and TNF-α is
proposed.
In conclusion, the present study findings partly
clarified the molecular mechanisms underlying the regeneration of
neuronal tissues by PDL-derived fibroblasts and may benefit future
research on neuronal regenerative medicine by transplantation of
PDL-derived fibroblasts.
Acknowledgments
Not applicable.
Abbreviations:
|
MS
|
mechanosensitive
|
|
PDL
|
periodontal ligament
|
|
NGF
|
nerve growth factor
|
|
IL-1β
|
interleukin 1β
|
|
TNF-α
|
tumor necrosis factor α
|
|
TGF-β
|
transforming growth factor β
|
|
MAPK
|
mitogen-activated protein kinase
|
|
Vmes
|
trigeminal mesencephalic nucleus
|
|
ERK1/2
|
extracellular signal-regulated kinases
1/2
|
|
TrkA
|
tropomyosin receptor kinase A
|
|
R-Smad
|
receptor-regulated Smad
|
|
co-Smad
|
common mediator Smad
|
|
I-Smad
|
inhibitory Smad
|
|
JNK
|
c-Jun N-terminal kinase
|
|
NF-κB
|
nuclear factor-κB
|
|
TH
|
tyrosine hydroxylase
|
|
ATPγS
|
adenosine
5′-O-(3-thio)triphosphate
|
|
SCDC
|
single cell-derived culture
|
|
FBS
|
fetal bovine serum
|
|
FGF
|
fibroblast growth factor
|
|
MAPKAPK-2
|
MAPK-activated protein kinase 2
|
|
PBS
|
phosphate-buffered saline
|
Funding
This study was supported in part by the JSPS KAKENHI
(grant nos. JP22592076 and JP17K11851 awarded to MK, JP25463053 and
JP16K11654 awarded to NC, JP26462823 awarded to SK, and JP26670852
and JP16H05534 awarded to AI) and the Grant-in-Aid for Strategic
Medical Science Research Centre (awarded to AI), both from the
Ministry of Education, Culture, Sports, Science, and Technology of
Japan.
Availability of data and materials
The data sets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
MO performed and interpreted data from RT-qPCR,
western blot analysis and immunofluorescence analysis, and was a
contributor in writing the manuscript. NC analyzed and interpreted
ELISA, and was a contributor in writing the manuscript. SK
performed RNA isolation and protein extraction from cells, and was
a contributor in writing the manuscript. SY, NO, AN, and MK were
contributors in asisting MO in performing ELISA, western blot
analysis, and immunofluorescence analysis. SJ and KS were
contributors in designing the experimental plan and writing
manuscript. AI made substantial conributions to the conception and
design of the study, and was a major contributor in writing the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jiang N, Guo W, Chen M, Zheng Y, Zhou J,
Kim SG, Embree MC, Songhee Song K, Marao HF and Mao JJ: Periodontal
ligament and alveolar bone in health and adaptation: Tooth
movement. Front Oral Biol. 18:1–8. 2016.
|
|
2
|
Shienaga Y, Doe K, Suemune S, Mitsuhiro Y,
Tsuru K, Otani K, Shirana Y, Hoshi M, Yoshida A and Kagawa K:
Physiological and morphological characteristics of periodontal
mesencephalic trigeminal neurons in the cat-intra-axonal staining
with HRP. Brain Res. 505:91–110. 1989. View Article : Google Scholar
|
|
3
|
Karita K and Tabata T: Response properties
of periodontal mechanosensitive units in the cat's thalamus. Exp
Brain Res. 86:341–346. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tabata T, Takahashi Y and Hayashi H:
Response properties of mechanosensitive neurons in the rat
trigeminal sensory complex projecting to the posteromedial ventral
nucleus of the thalamus. Arch Oral Biol. 46:881–889. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Byers MR: Sensory innervation of
periodontal ligament of rat molars consists of unencapsulated
Ruffini-like mechanoreceptors and free nerve endings. J Comp
Neurol. 231:500–518. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Maeda T, Sato O, Kobayashi S, Iwanaga T
and Fujita T: The ultrastructure of Ruffini endings in the
periodontal ligament of rat incisors with special reference to the
terminal Schwann cells (K-cells). Anat Rec. 223:95–103. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Uemura T, Yasuda K, Ishihara K, Yasuda H,
Okayama M, Hasumi-Nakayama Y and Furusawa K: A comparison of the
postnatal development of muscle-spindle and periodontal-ligament
neurons in the mesencephalic trigeminal nucleus of the rat.
Neurosci Lett. 473:155–157. 2010. View Article : Google Scholar
|
|
8
|
Korkmaz Y, Bloch W, Klinz FJ, Kübler AC,
Schneider K, Zimmer S, Addicks K and Raab WH: The constructive
activation of extracellular signal-regulated kinase 1 and 2 in
periodontal ligament nerve fibers. J Periodontol. 80:850–859. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Atsumi Y, Imai T, Matsumoto K, Sakuda M,
Maeda T, Kurisu K and Wakisaka S: Effects of different types of
injury to the inferior alveolar nerve on the behavior of Schwann
cells during the regeneration of periodontal nerve fibers of rat
incisor. Arch Histol Cytol. 63:43–54. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Webber CA, Xu Y, Vanneste KJ, Martinez JA,
Verge VM and Zochodne DW: Guiding adult mammalian sensory axons
during regeneration. J Neuropathol Exp Neurol. 67:212–222. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Donnerer J: Regeneration of primary
sensory neurons. Pharmacology. 67:169–181. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Angeletti RH and Bradshaw RA: Nerve growth
factor from mouse submaxillary gland: Amino acid sequence. Proc
Natl Acad Sci USA. 68:2417–2420. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hirose M, Kuroda Y and Murata E: NGF/TrkA
signaling as a therapeutic target for pain. Pain Pract. 16:175–182.
2016. View Article : Google Scholar
|
|
14
|
Li MO, Wan YY, Sanjabi S, Robertson AK and
Flavell RA: Transforming growth factor-beta regulation of immune
responses. Annu Rev Immunol. 24:99–146. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang W, Huang XR, Li AG, Liu F, Li JH,
Truong LD, Wang XL and Lan HY: Signaling mechanism of TGF-beta1 in
prevention of renal inflammation: Role of Smad7. J Am Soc Nephrol.
16:1371–1383. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Korns D, Frasch SC, Fernandes-Boyanapalli
R, Henson PM and Bratton DL: Modulation of macrophage efferocytosis
in inflammation. Front Immunol. 2:572011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ahmed S, Bradshaw AD, Gera S, Dewan MZ and
Xu R: The TGF-β/Smad4 signaling pathway in pancreatic
carcinogenesis and clinical significance. J Clin Med. 6:E52017.
View Article : Google Scholar
|
|
18
|
Heldin CH and Moustakas A: Signaling
receptors for TGF-β family members. Cold Spring Harb Perspect Biol.
8:a0220532016. View Article : Google Scholar
|
|
19
|
Macias MJ, Martin-Malpartida P and
Massagué J: Structural determinations of Smad function in TGF-β
signaling. Trends Biochem Sci. 40:296–308. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu G and Fahnestock M: Differential
expression of nerve growth factor transcripts in glia and neurons
and their regulation by transforming growth factor-beta1. Mol Brain
Res. 105:115–125. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lindholm D, Hengerer B, Zafra F and
Thoenen H: Transforming growth factor-beta 1 stimulates expression
of nerve growth factor in the rat CNS. Neuroreport. 1:9–12. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Blaney Davidson EN, van Carm APM, Vitters
EL, Bennink MB, Thijssen E, van der Berg WB, Koenders MI, van Lent
PLEM, van deLoo FAJ and van der Kraan PM: TGF-β is a potent inducer
of nerve growth factor in articular cartilage via the ALK5-Smad2/3
pathway. Potential role in OA related pain? Osteoarth Cartil.
23:478–486. 2015. View Article : Google Scholar
|
|
23
|
Mu Y, Gudey SK and Landström M: Non-Smad
signaling pathways. Cell Tissue Res. 347:11–20. 2012. View Article : Google Scholar
|
|
24
|
Yongchaitracl T and Pavasant P:
Transforming growth factor-beta1 up-regulates the expression of
nerve growth factor through mitogen-activated protein kinase
signaling pathways in dental pulp cells. Eur J Oral Sci. 115:57–63.
2007. View Article : Google Scholar
|
|
25
|
Hahn M, Lorez H and Fischer G: The
immortalized astrogrial cell line RC7 is a new model system for the
study of nerve growth factor (NGF) regulation: Stimulation by
interleukin-1 beta and transforming growth factor-beta 1 is
additive and affected differently by dibutyryl cyclic AMP. Glia.
10:286–295. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hattori A, Iwasaki S, Murase K, Tsujimoto
M, Sato M, Hayashi K and Kohno M: Tumor necrosis factor is markedly
synergistic with interleukin 1 and interferon-gamma in stimulating
the production of nerve growth factor n fibroblasts. FEBS Lett.
340:177–180. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Weber A, Wasiliew P and Kracht M:
Interleukin-1 (IL-1) pathway. Sci Signal. 3:2010.
|
|
28
|
Kalliolias GD and Ivashkiv LB: TNF
biology, pathogenic mechanisms and emerging therapeutic strategies.
Nat Rev Rheumatol. 12:49–62. 2016. View Article : Google Scholar :
|
|
29
|
Hengerer B, Lindholm D, Heumann R, Rüther
U, Wagner EF and Thoenen H: Lesion-induced increase in nerve growth
factor mRNA is mediated by c-fos. Proc Natl Acad Sci USA.
87:3899–3903. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee JH, Kim JW, Im YS, Seong GJ and Lee
HK: Cyclosporine A induces nerve growth factor expression via
activation of p38 and NFAT5. Cornea Suppl. 1:S19–S24. 2011.
View Article : Google Scholar
|
|
31
|
Heese K, Inoue N and Sawada T: NF-κB
regulates B-cell-derived nerve growth factor expression. Cell Mol
Immunol. 3:63–66. 2006.PubMed/NCBI
|
|
32
|
White RB and Thomas MG: Moving beyond
tyrosine hydroxylase to defined dopaminergic neurons for use in
cell replacement therapies for Parkinson's disease. CNS Neurol
Disord Drug Targets. 11:340–349. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jinnin M, Ihn H and Tamaki K:
Characterization of SIS3, a novel specific inhibitor of Smad3, and
its effect on transforming growth factor-beta1-induced
extracellular matrix expression. Mol Pharmacol. 69:597–607. 2006.
View Article : Google Scholar
|
|
34
|
Okubo N, Ishisaki A, Iizuka T, Tamura M
and Kitagawa Y: Vascular cell-like potential of undifferentiated
ligament fibroblasts to construct vascular cell-specific
marker-positive blood vessel structures in a
PI3K-activation-dependent manner. J Vasc Res. 47:369–383. 2010.
View Article : Google Scholar
|
|
35
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−delta delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
36
|
Arthur DB, Akassoglou K and Insel PA: P2Y2
receptor activates nerve growth factor/TrkA signaling to enhance
neuronal differentiation. Proc Natl Acad Sci USA. 102:19138–19143.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bitzer M, von Gersdorff G, Liang D,
Dominguez-Rosales A, Beg AA, Rojkind M and Böttinger EP: A
mechanism of suppression of TGF-beta/SMAD signaling by NF-kappa
B/RelA. Genes Dev. 15:187–197. 2000.
|
|
38
|
Greene LA and Tischler AS: Establishment
of a noradrenergic clonal line of rat adrenal pheochromocytoma
cells which respond to nerve growth factor. Proc Natl Acad Sci USA.
73:2424–2428. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chigero DJ Jr: The early distribution and
possible role of nerves during odontogenesis. Int J Dev Biol.
39:191–194. 1995.
|
|
40
|
Gizang-Ginsberg E and Ziff EB: Nerve
growth factor regulates tyrosine hydroxylase gene transcription
through a nucleoprotein complex that contains c-Fos. Genes Dev.
4:477–491. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu YC, Zou XB, Chai YF and Yao YM:
Macrophage polarization in inflammatory diseases. Int J Biol Sci.
10:520–529. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ferrante CJ and Leibovich SJ: Regulation
of macrophage polarization and wound healing. Adv Wound Care (New
Rochelle). 1:10–16. 2012. View Article : Google Scholar
|
|
43
|
Gram H: Preclinical characterization and
clinical development of ILARIS(®) (canakinumab) for the treatment
of autoinflammatory diseases. Curr Opin Chem Biol. 32:1–9. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Billmeier U, Dieterich W, Neurath MF and
Atreya R: Molecular mechanism of action of anti-tumor necrosis
factor antibodies in inflammatory bowel diseases. World J
Gastroenterol. 22:9300–9313. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li B, Jung HJ, Kim SM, Kim MJ, Jahng JW
and Lee JH: Human periodontal ligament stem cells repair mental
nerve injury. Neural Regen Res. 8:2827–2837. 2013.
|