Introduction
The aging population is increasing at a rapid rate
worldwide, giving rise to a number of age-related diseases that
have a significant social and economic burden on the community.
With normal aging, the brain undergoes synaptic dysfunction,
extensive neuronal death and declined neurogenesis. Learning and
memory impairment and cognitive deficits are well-known
characteristics of the aging process (1-3).
In addition, aging is associated with various debilitating
neurodegenerative conditions, including Alzheimer's disease (AD).
Thus, the prevention or delay of the onset of age-related diseases
and age-related cognitive decline may improve the quality of
life.
The hippocampus, located in the medial temporal lobe
of the brain, is crucial for normal learning and memory
consolidation. This region is particularly vulnerable to the aging
process (2,4). The hippocampus has been shown to
undergo several structural and functional changes with age
(2). Significant aged-related
neuronal atrophy and volume decreases of the hippocampus, as well
as hippocampal-dependent learning and memory decline have been
demonstrated (5). An upregulation
in the levels of pro-inflammatory genes and inflammatory parameters
has also been observed in the hippocampus during aging (6,7).
Additionally, changes in synaptic plasticity have been detected in
the hippocampi of aged humans and rodents (8,9).
Although the mechanisms underlying age-related synaptic plasticity
impairment are still under investigation, dysregulations and
alterations in the expression levels of several proteins, that play
key roles in synaptogenesis and synaptic stabilization, in the
hippocampus have been reported (2,10).
Piperlongumine (PL,
5,6-dihydro-1-[(2E)-1-oxo-3-(3,4,5-trimethoxyphenyl)-2-propenyl]-2(1H)-pyridinone)
is a natural alkaloid that can be isolated from the long pepper
(Piper longum L.). PL is found in the fruits and roots of
the plant (11). Cumulative
evidence has indicated that PL has a number of pharmacological
activities, including antidepressant, anxiolytic, anti-fungal,
antidiabetic, antinociceptive and antitumour properties (11-16). Moreover, in our previous study, it
was demonstrated that administration of PL improves cognitive
function in a transgenic mouse model of AD (17). Thus, we hypothesized that PL would
enhance cognitive function in aged mice. In the present study, we
demonstrate that PL treatment modulates age-related cognitive
decline and hippocampal dysfunction in aged mice.
Materials and methods
Preparation of PL
PL was isolated from Piper longum.
Preparation was performed as described in previous studies
(17-19). Dried fruits (500 g) of Piper
longum were extracted with ethyl acetate (EtOH; 1 liter x 3
times) at room temperature for 1 week. The combined EtOH extracts
were concentrated to yield a dry residue (32.5 g), which was
subsequently suspended in water (H2O; 500 ml) and
partitioned with EtOAc (3×500 ml). The partial EtOAc extract (6.0
g), which was subjected to a silica gel column chromatography (CC;
5×40 cm), was eluted with a gradient n-hexane/acetone system
(20:1 to 1:1) to yield 5 fractions (F1-F5). Fractions F3 and F4
were combined and further applied to a reversed
phase-C18 CC (3×30 cm) with methanol
(MeOH)/H2O (1:1 to 9:1). Subfraction F34.3 (60.8 mg) was
purified by high-performance liquid chromatography [mobile phase:
MeOH in H2O containing (0-40 min: 65% MeOH); flow rate:
2 ml/min; UV detection at 205 and 254 nm] to yield a compound
(tR=17.2 min, 14 mg). The chemical structure of
the isolated compound was confirmed by comparison with the reported
chemical structure of PL using 1D and 2D nuclear magnetic resonance
spectroscopy.
Animals
Female C57BL/6J mice, at 3 months (n=7, weighing
19-22 g) and 23 months of age (n=28, 28-34 g), were obtained from
the Korea Research Institute of Bioscience and Biotechnology
(KRIBB, Daejeon, Korea) and housed in regular polycarbonate plastic
cages in an environment with a controlled temperature (21-22°C) and
humidity (50-60%) and a 12-h light/dark cycle (lights on at 7
a.m.). The mice were maintained on an ad libitum diet of lab
chow (Teklad 2018S, Harlan, WI, USA) with free access to water. The
cages were filled to an approximate depth of 1.5 cm with bedding
made of chopped wood particles (JSBio, Daejeon, Korea). All
materials used were autoclaved and gamma-irradiated. The animal
room was maintained in specific-pathogen-free conditions. The
C57BL/6J mice at 23 months of age were randomized into the vehicle
[0.5% carboxymethyl cellulose (CMC), Aged vehicle, n=14)] and PL
(Aged PL, n=14) groups. The PL extract was suspended in 0.5% CMC at
a concentration of 5 mg/ml as a stock solution. The 23-month-old
female mice were orally administrated 10 μl/g/day of PL
stock solution or 0.5% CMC for 8 weeks. The 3-month-old female mice
were used as young controls (n=7). Multiple behaviour tests were
performed on a single cohort of mice and the following order was
obeyed: Open field test → novel object recognition test →
nest-building behaviour test (17,20). All the animal experiments were
approved by the Institutional Animal Use and Care Committee of the
KRIBB (KRIBB-AEC-14074).
Open filed locomotor activity
The mice were individually placed in an open field
box (45×45×45 cm3) for 30 min. The horizontal locomotion
of the mouse was measured using a computerized video tracking
system, SMART (Panlab, Barcelona, Spain).
Novel object recognition test
The novel object recognition test was performed as
described in previous studies (21,22). The mice were individually
habituated to a testing chamber (40×20×20 cm3) with no
objects for 5 min and then placed in a testing chamber for 10 min
with two identical objects (familiar, acquisition session). The
mice were then returned to the home cages. One day later, the mice
were placed back into the testing chamber in the presence of one of
the original objects and one novel object (novel, recognition
session) for 10 min. The original objects were cylindrical wooden
blocks 10 cm high x 2 cm in diameter. The novel object was a
10×2.5×2 cm rectangular wooden block. The acquisition and
recognition sessions were video-recorded and an observer, who was
blinded to the drug treatment, scored the time spent exploring the
objects. The chambers and objects were cleaned with ethanol between
trials. Exploration was defined as sniffing and touching the object
with the nose and/or forepaws. Sitting on the object was not
considered exploratory behaviour. A discrimination index was
calculated for each animal and expressed using the following
formula: [time (number) of contacts with the novel object-time
(number) of contacts with the familiar object]/[time (number) of
contacts with the novel object + time (number) of contacts with the
familiar object] on day 2.
Nest-building behaviour test
The nest building behaviour test was performed as
described in a previous study (23). The mice were housed in single
cages containing chopped wood particles for 5 days. On the first
day of testing, one piece of cotton (5×5 cm; Nestlets, Ancare,
Bellmore, NY, USA) was introduced into the home cage to permit
nesting. The presence and quality of nesting was rated 1 day later
on a 5-point scale ranging from 1 to 5 as follows: 1, nestlet not
noticeably touched (>90% intact); 2, nestlet partially torn up
(50-90% remaining intact); 3, mostly shredded, but often no
identifiable nest site; 4, an identifiable but flat nest; and 5, a
(near) perfect nest. Immediately afterward, the mice were
group-housed as before.
Western blot analysis
Western blot analysis was performed as described in
a previous study (21). Following
8 weeks of PL treatment, the mice were sacrificed and the
hippocampal tissues were rapidly removed and homogenized in a
homogenization buffer (50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1%
Nonidet P-40, 0.1% sodium dodecyl sulfate and 0.1% sodium
deoxycholate) containing a cocktail of protease inhibitors (Roche
Diagnostics GmbH, Mannheim, Germany). Protein samples were resolved
by performing sodium dodecyl sulfate-polyacrylamide gel
electrophoresis. The samples were then transferred onto
polyvinylidene fluoride membranes (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The blots were incubated with primary
antibodies followed by secondary antibodies, and specific signals
were visualized using an Enhanced Chemi Luminescence kit (Intron
Biotechnology, Gyeonggi-do, Korea). Western blot images were
quantified using Quantity One 1-D analysis software version 4.6.1
(Bio-Rad Laboratories, Inc.). The primary antibodies used were
vesicular glutamate transporter 1 (VGLUT1; 1:1,000, #135 302, SYSY,
Göttingen, Germany), vesicular glutamate transporter 2 (VGLUT2;
1:1,000, #75-067 UC Davis/NIH NeuroMab Facility, Davis, CA, USA),
glutamate receptor 1 (GluR1; a gift from Dr J.R. Lee, KRIBB,
Daejeon, Korea, 1:1,000), N-methyl-D-aspartate receptor
subtype 2B [(NR2B, 1:1,000, #4212, Cell Signaling Technology (CST),
Danvers, MA, USA)], phosphorylated (p-)NR2B (p-Tyr-1472-NR2B,
1:1,000, #4208, CST), synaptophysin (1:1,000, #S5768, Sigma-Aldrich
Co. LLC; Merck KGaA, Darmstadt, Germany), post-synaptic density
protein 95 (PSD-95, 1:1,000, #124 014, SYSY), glutamate
decarboxylase 65/67 (GAD65/67, 1:1,000, #AB1511, Merck KGaA),
gephyrin (1:1,000, #147 011, SYSY), vesicular GABA transporter
(VGAT, 1:1,000, #131 002, SYSY), cAMP response element binding
protein (CREB, 1:1,000, #06-863, Merck KGaA), p-CREB
(p-Ser133-CREB, 1:1,000, #06-519, Merck KGaA),
calcium/calmodulin-dependent protein kinase type II α (CaMKIIα,
1:1,000, #sc-13141, Santa Cruz Biotechnology, Inc., Dallas, TX,
USA), p-CaMKIIα (p-Thr-286-CaMKIIα, 1:1,000, #sc-12886, Santa Cruz
Biotechnology, Inc.), extra cellular signal-regulated kinases 1/2
(ERK1/2, 1:1,000, #9102, CST), p-ERK1/2 (p-Thr202/Tyr204-ERK1/2,
1:1,000, #9101, CST) and β-actin (1:1,000, #MAB1501, Merck KGaA).
The secondary antibodies used were horseradish
peroxidase-conjugated goat anti-rabbit IgG (1:2,000, #NCI1460KR,
Thermo Fisher Scientific, Inc., Waltham, MA, USA) or goat
anti-mouse (1:2,000, #sc-2005, Santa Cruz Biotechnology).
Histological analysis
Immunohistochemistry and immunofluorescence staining
were performed as previously described (21,24-26). Following 8 weeks of PL treatment,
the mice were deeply anesthetized (250 mg/kg Avertin,
intraperitoneally) and transcardially perfused with saline followed
by 4% paraformaldehyde in phosphate-buffered saline (PBS). The
brains were removed, post-fixed overnight, and then cut into
40-μm-thick coronal sections using a vibratome (Vibratome
VT1000A, Leica Microsystems GmbH, Wetzlar, Germany). The
free-floating sections were then incubated in PBS containing 3%
H2O2 (v/v), rinsed 3 times in PBS, and
blocked with serum for 1 h at room temperature. The sections were
then incubated with the phospho-CREB (Ser133, 1:1,000, #06-519,
Merck KGaA), doublecortin (DCX, 1:1,000, #sc-8666, Santa Cruz
Biotechnology), 4-hydroxy-2-nonenal (4-HNE, 1:1,000, #HNE11-S,
Alpha Diagnostic, San Antonio, TX, USA), ionized calcium-binding
adapter molecule 1 (Iba1, 1:1,000, #019-19741, Wako Chemicals USA,
Inc., Richmond, VA, USA) and glial fibrillary acidic protein (GFAP,
1:1,000, #Z-0334, Dako, Glostrup, Denmark) primary antibodies
overnight at 4°C. The sections were then washed and incubated with
biotinylated secondary anti-rabbit IgG (1:200, #BA-1000, Vector
Laboratories, Inc., Burlingame, CA, USA), followed by the
avidin-biotinylated peroxidase complex (Vector Laboratories, Inc.)
and 3,3′-diaminobenzidine (Sigma-Aldrich Co. LLC; Merck KGaA).
Immunofluorescence staining was then performed with an Alexa Fluor
594 goat anti-rabbit IgG antibody (secondary antibody, 1:200,
#A11012, Thermo Fisher Scientific, Inc.). Sections containing the
hippocampus were selected and the number of doublecortin-positive
cells in the dentate gyrus (DG) were counted under a microscope
(Olympus Corp., Tokyo, Japan). The intensity of 4-HNE- and
p-CREB-stained cells and the percentage area occupied by GFAP- and
Iba-1-positive cells in hippocampal CA1, CA3 and DG were assessed
using the MetaMorph image analyser (Molecular Devices, LLC,
Sunnyvale, CA, USA).
Statistical analysis
GraphPad PRISM (GraphPad Software, Inc., La Jolla,
CA, USA) software was used to perform the statistical analyses.
Two-sample comparisons were performed using a Student's t-test,
while multiple comparisons were made using a one-way ANOVA followed
by the Tukey-Kramer's post hoc test. Associations between distance
and discrimination index were examined by Pearson's correlation
coefficient. All data are presented as the means ± SEM and
statistical differences are accepted at the 5% level (P<0.05),
unless otherwise indicated.
Results
PL improves the performance of aged mice
in novel object recognition and nest building tasks
The aged female C57BL/6J mice (23 months old) were
randomly separated into the vehicle- and PL-treated groups. PL was
administered at a dose of 50 mg/kg/day for 8 weeks, from the ages
of 23 to 25 months. The experimental design is presented in
Fig. 1A. The aged mice (24 months
of age) exhibited a significantly lower locomotor activity in the
open field test than the young control mice (Fig. 1B, P<0.05). PL treatment did not
markedly affect the exploratory behaviour of the aged mice compared
to the aged vehicle group (Fig.
1B, P>0.05). To determine whether PL can improve cognitive
function in aged mice, we performed the novel object recognition
test. In the recognition session, with two different objects (one
novel and the other familiar), the young control mice explored the
novel object for a relatively long time period and a made contact
with it a relatively high number of times, yielding a
discrimination index (DI) of approximately 0.24±0.08 and 0.15±0.07,
indicating that they had a memory of the familiar object (Fig. 1D and E). By contrast, the aged
mice treated with the vehicle exhibited a DI that was significantly
lower than that of the younger controls (−0.05±0.05 and −0.06±0.04,
Fig. 1D and E), which is
consistent with impaired cognition. PL treatment markedly increased
the DI in aged mice to approximately 0.24±0.06 and 0.17±0.05
(Fig. 1D and E), reflecting a
therapeutic effect of PL on age-related cognitive impairment. PL
treatment did not alter the total exploration time (aged vehicle,
10.14±1.11 sec; aged PL, 8.97±0.48 sec, P=0.466) and total number
of contacts (aged vehicle, 16.92±2.22; aged PL, 19.71±1.25,
P=0.680) to both objects (familiar + novel) on day 2, indicating no
influence on the total exploration activity of PL in the novel
object recognition test. Additionally, we could not find any
association between the distance in the open field test and the DI
in the novel object recognition test in the aged mice (Fig. 1F and G, P>0.05).
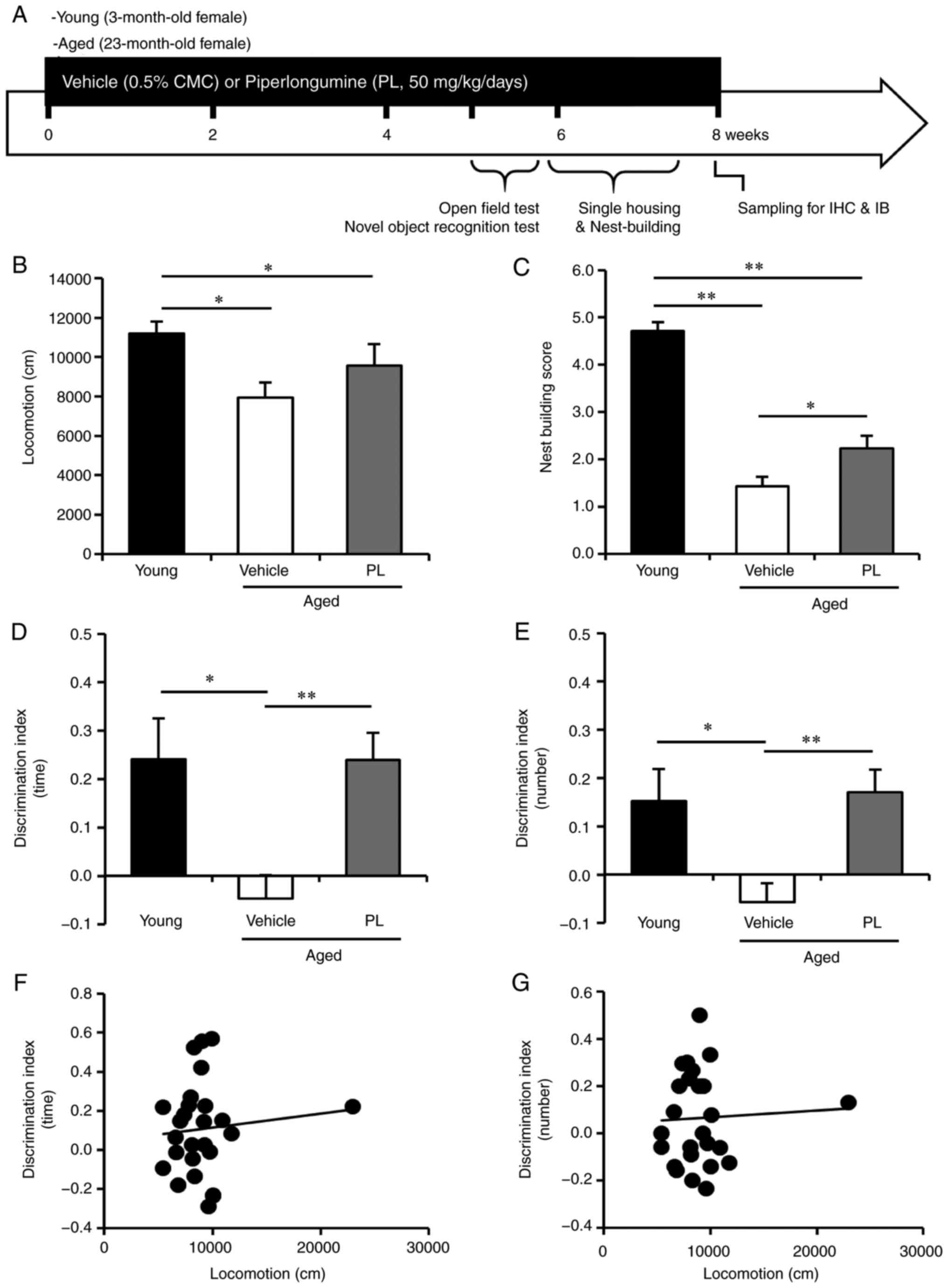 | Figure 1Effect of piperlongumine (PL) on
novel object recognition and nest building in aged female mice. (A)
Experimental design for PL treatment, behaviour testing and
sampling. Open field test, novel object recognition test and nest
building behaviour test were performed at 39, 42 and 54 days of PL
treatment. (B) Total locomotor activity for a 30-min period in
young control mice and aged mice following treatment with the
vehicle or PL (young control; n=7, aged vehicle; n=14, aged PL;
n=14). (C) The presence and quality of nesting over a 24 h period,
rated on a 5-point scale, in young control mice and aged mice
following treatment with the vehicle, or PL (young control; n=7,
aged vehicle; n=8, aged PL; n=9). (D and E) The discrimination
index [(D) the time spent exploring and (E) the number of contacts]
of the young mice, and aged mice following treatment with the
vehicle or PL in the novel object recognition test (young control;
n=6, aged vehicle; n=14, aged PL; n=14). (F and G) The correlation
between locomotor activity in open field test and discrimination
index in novel object recognition test in aged mice was absent [F,
between locomotion (B) and DI (time, D), r=0.09955, P=0.6285; G,
between locomotion (B) and DI (number, E), r=0.04872, P=0.8132,
n=26]. *P<0.05 and **P<0.01,
significant differences from an indicated group, determined by
one-way ANOVA, followed by Tukey-Kramer's post-hoc test. |
Previous studies have reported that nest building,
which is an indicator of well-being and social context in mice, is
decreased in aging in rodent models of AD (27,28). Reduced nesting has also been
observed in mice with hippocampal lesions (29). In this study, the nesting score in
the nest building test was significantly lower in the aged mice
than in the young control mice (Fig.
1C, P<0.05). PL significantly increased the nesting score in
the aged mice (Fig. 1C,
P<0.05). These results indicate that treatment with PL may
improve cognitive and social decline without affecting locomotion
in aged mice.
PL did not alter the glia activation and
lipid peroxidation in the hippocampus of aged mice
An upregulation of inflammatory responses and
oxidative stress have been observed in the hippocampus in aging
(30-33). An increase in inflammation in
aging implicates the activation of microglia and astrocytes in the
brain over this period (34). In
aged brains, there is an increase in the number, size and
activation of microglia (34). In
this study, to investigate the effects of PL on microglia and
astrocytes in aging, we measured the percentage area occupied by
astrocytes (Fig. 2A and D) and
microglia (Fig. 2B and E) in
hippocampus through immunohistochemical assay. Additionally,
immunofluorescence analysis for oxidative stress (4-HNE, an
indicator of lipid peroxidation) in the hippocampus was performed
(Fig. 2C and F). PL
administration at a dose of 50 mg/kg/day for 8 weeks had no
significant effect on glial activation and oxidative stress in the
hippocampus at this point in aging.
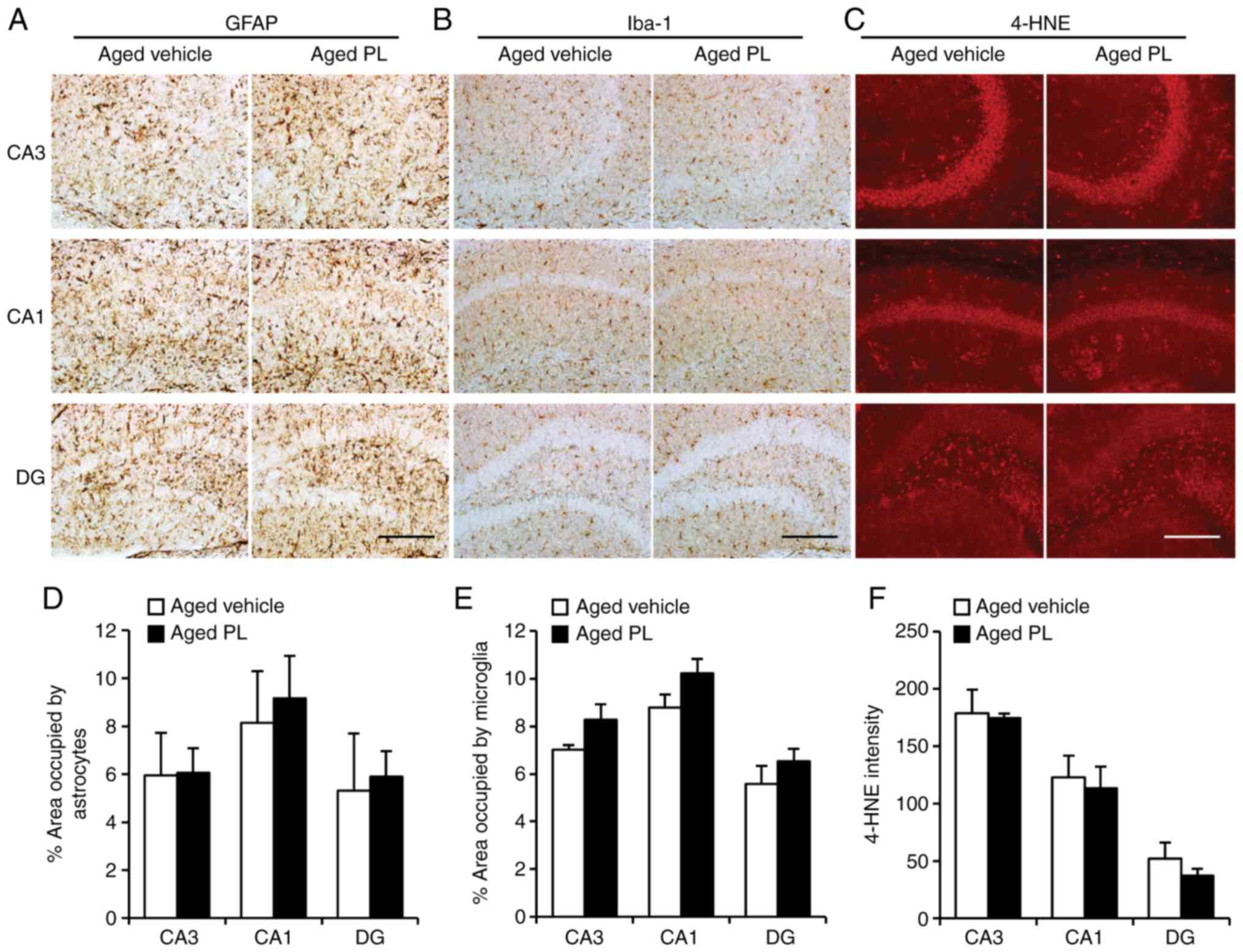 | Figure 2Effect of piperlongumine (PL) on
neuroinflammation and oxidative stress in the hippocampus.
Activation of microglia and astrocytes was analysed by
immunohistochemical staining against glial fibrillary acidic
protein (GFAP), ionized calcium binding adaptor molecule 1 (Iba1)
and 4-hydroxynonenal (4-HNE), respectively. Images showing (A)
GFAP, (B) Iba1, and (C) 4-HNE labelling in the hippocampus of aged
mice treated with the vehicle or PL. (D) Percentage area of
hippocampus [CA3, CA1 and dentate gyrus (DG)] occupied by
astrocytes (thus GFAP labelled; aged vehicle; n=6, aged PL; n=9) in
aged mice treated with the vehicle or PL. (E) Percentage area of
hippocampus (CA3, CA1, and DG) occupied by microglia (thus Iba-1
labelled; aged vehicle; n=6, aged PL; n=9) in aged mice treated
with the vehicle or PL. (F) 4-HNE-intensity in the hippocampus
(CA3, CA1, DG) in aged mice treated with the vehicle or PL aged
vehicle; n=4, aged PL; n=4). Scale bar, 200 μm. Data are
presented as the means ± SEM. |
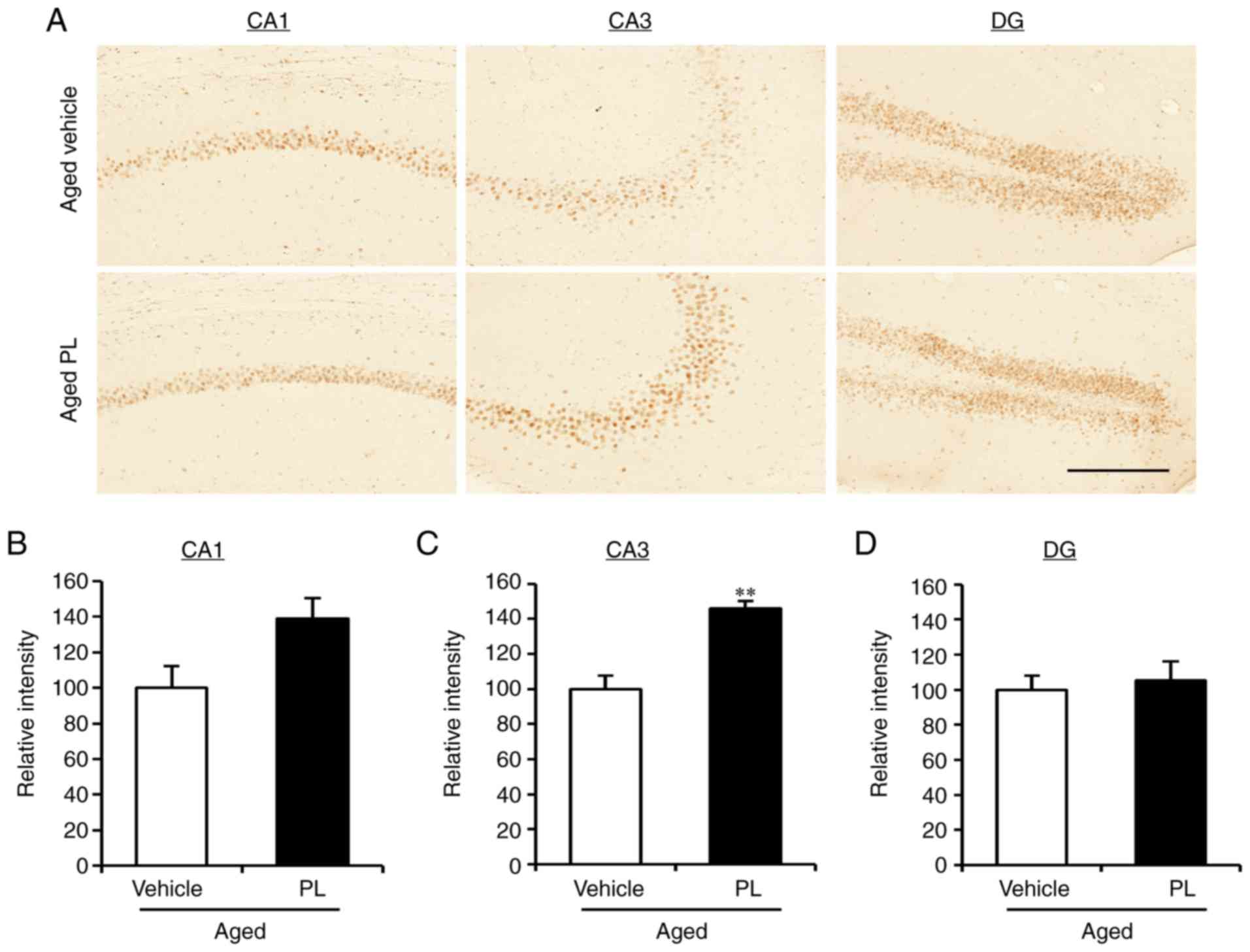
PL increases the phosphorylation of NR2B,
ERK1/2 and CaMKIIα in the hippocampus of aged mice
As the results from the behavioural tests pointed to
a reduction in age-related cognitive impairment with PL treatment,
we examined the level of synaptic markers in the hippocampus of the
aged mice treated with the vehicle or PL. As indicated by the
results of western blot analysis, the expression levels of
gephyrin, VGAT, GAD65/67, PSD95, VGLUT1, VGLUT2 and synaptophysin
were similar between the aged vehicle and aged PL groups (Fig. 3A and B). Additionally, PL had no
effect on the protein expression of the AMPA (GluR1) or NMDA (NR2B)
receptors (Fig. 3C and D). Of
note, the levels of phosphorylation of NR2B (Tyr1472), ERK1/2
(Thr202/Tyr204) and (Thr286) were significantly higher in the aged
mice treated with PL than in the aged mice treated with the vehicle
(Fig. 3C and D). There was a
tendency for the phosphorylation of CREB (Ser133) to be slightly
higher in the aged PL group than the aged vehicle group, although
this difference was not significant. To further investigate the
level of p-CREB in the areas of the hippocampus, we measured the
integrated optical density (IOD) of p-CREB by immunohistochemical
assay in the CA1, CA3, and DG of the aged vehicle- and aged
PL-treated mice (Fig. 4). The IOD
in the CA3 was markedly higher in the aged mice treated with PL
than in the aged mice treated with the vehicle (Fig. 4A and C, P<0.01); however, the
level of p-CREB in the CA1 and DG did not differ significantly
between the groups (Fig. 4A, B and
D). Taken together, these results suggest that the molecular
signalling pathways involving NR2B, CaMKIIα, ERK1/2 and CREB are
regulated by PL treatment in the hippocampus of the aged mice.
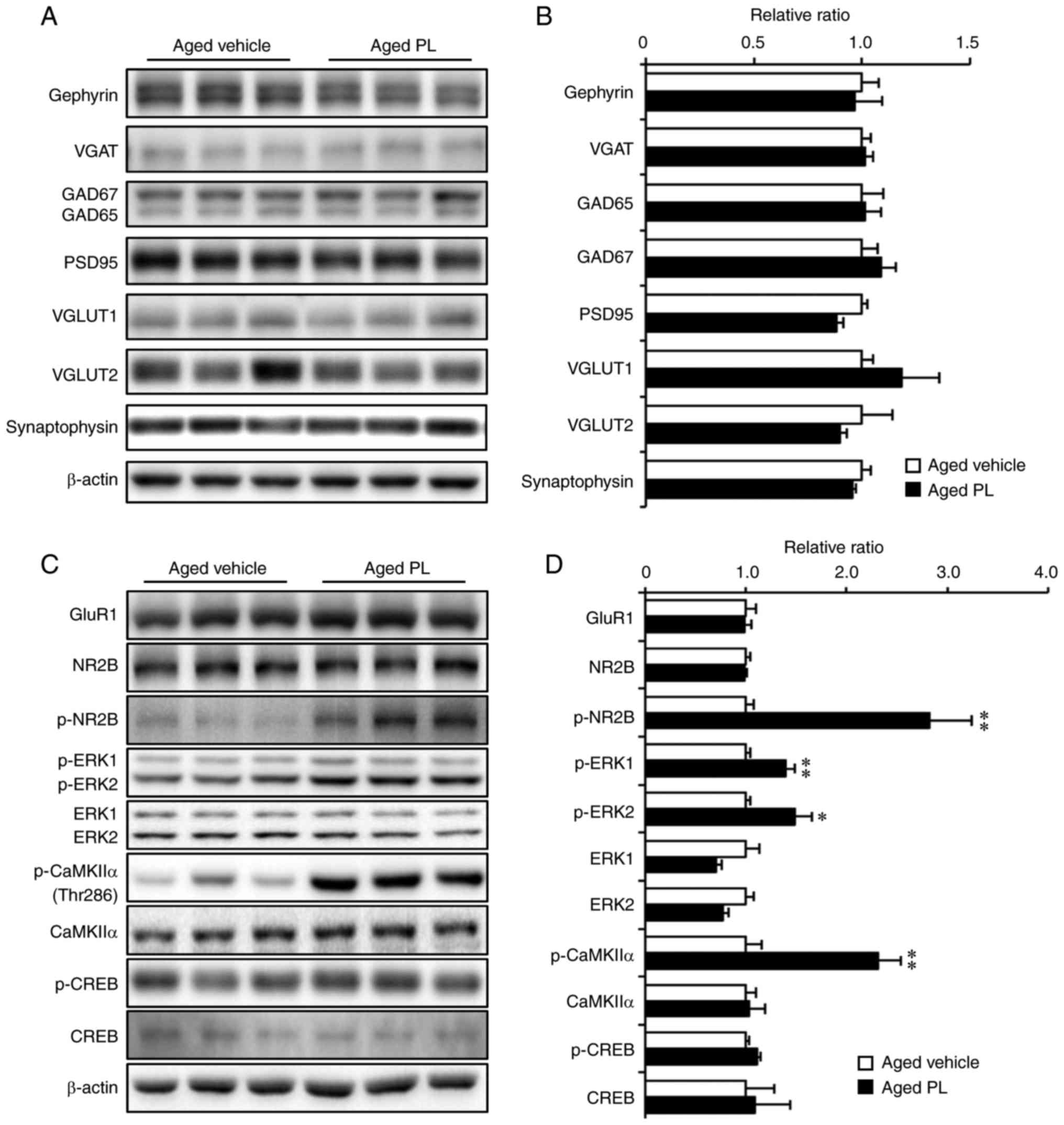 | Figure 3Effect of piperlongumine (PL) on the
expression of synaptic proteins and NMDAR signalling proteins. (A
and B) Western blot analysis and quantitative analysis of the
expression of synaptic proteins [gephyrin, vesicular GABA
transporter (VGAT), glutamate decarboxylase 65/67 (GAD65/67),
postsynaptic density protein 95 (PSD95), vesicular glutamate
transporter 1 (VGLUT1), vesicular glutamate transporter 2 (VGLUT2)
and synaptophysin, aged vehicle; n=5, aged PL; n=7] in hippocampal
homogenates of aged mice treated with the vehicle or PL. (C and D)
Western blot analysis and quantitative analysis of the expression
of NMDAR signalling proteins [glutamate receptor 1 (GluR1),
N-methyl-D-aspartate receptor subtype 2B (NR2B), p-NR2B,
extracellular signal-regulated kinase (ERK)1/2, p-ERK1/2,
calcium/calmodulin-dependent protein kinase type II α (CaMKIIα),
p-CaMKIIα, cAMP response element binding protein (CREB) and p-CREB,
aged vehicle; n=5, aged PL; n=7] in hippocampal homogenates of aged
mice treated with the vehicle or PL. *P<0.05 and
**P<0.01, significant differences from the aged
vehicle, as shown by the Student's t-test. Data are presented as
the means ± SEM. |
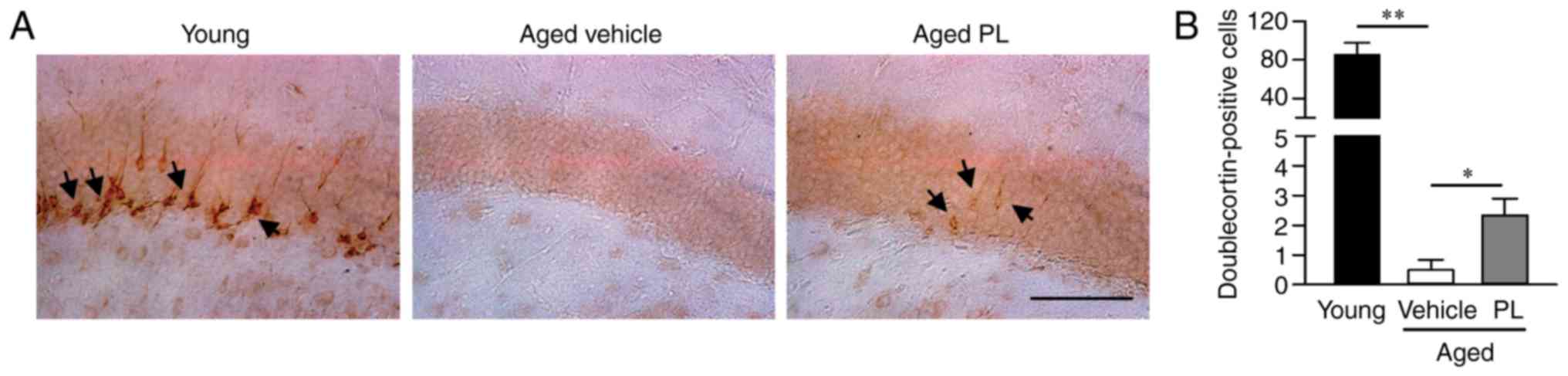
PL increases neurogenesis in the DG of
aged mice
Neurogenesis markedly declines with aging and, thus,
the maintenance of an adequate level of hippocampal neurogenesis is
another important factor to consider in maintaining cognitive
function (35). In this study, to
investigate whether PL treatment affects hippocampal neurogenesis,
we examined neuronal proliferation by immunohistochemistry using
the neuroblast marker, DCX, in the DG of aged mice treated with the
vehicle or PL (Fig. 5). The
number of DCX-positive cells was markedly lower in the aged mice
treated with the vehicle than in the young control group (Fig. 5). However, the number of
DCX-positive cells was significantly higher in the PL treated aged
mice than in the vehicle treated aged mice (Fig. 5, P<0.05). These results suggest
that PL increases adult neurogenesis in the DG of aged mice.
Discussion
Aging is a natural biological process that is
associated with physical and cognitive decline. Notably, in both
normal aging and under pathological conditions, cognitive decline
can diminish the quality of life. In the present study, we found
that treatment with piperlongumine (PL), isolated from the long
pepper, significantly improved cognitive function in novel object
recognition and performance in nest building in 25-month-old female
mice. These effects appear to be partly due to the modulation of
neuronal activity and neurogenesis in the hippocampus. We found
that treatment with PL increased the phosphorylation levels of the
NR2B subunit of the NMDA receptor in the hippocampus of aged mice.
Furthermore, we observed that PL significantly increased the
phosphorylation of ERK1/2 at Thr202/Tyr204, CaMKIIα at Thr286, and
CREB at Ser133, and increased the number of doublecortin-positive
cells.
PL is a primary constituent of Piper longum,
which has been reported to kill multiple types of cancer cells
through the targeting of the stress response to reactive oxygen
species (ROS) (14,36). Diagnosis with certain tumours,
such as age-related degenerative diseases, increases with age and
the molecular alterations that occur in aging can favour
carcinogenesis (37). Senescent
cells can drive hyperplastic pathology and promote age-related
neurodegeneration (38,39). Recently, PL has been reported to
be a potential novel lead for the development of senolytic agents
(40) and the selective depletion
of senescence cells as an anti-aging strategy may prevent cancer
and aging-related degenerative diseases. Although in this study, we
did not investigate the anti-tumour activities of PL in aged mice,
PL treatment may be beneficial through the apoptosis of age-related
senescence cells. Cellular senescence is associated with oxidative
stress and inflammation (39). An
increase in the expression of GFAP has been the most common change
to be observed in astrocytes with aging (41). The results of this study
demonstrated that PL did not affect the size of area occupied by
glia, such as microglia and astrocytes, in the hippocampus of the
aged mice (Fig. 2). We also
observed that lipid peroxidation in the hippocampus was not altered
in the aged mice (Fig. 2).
However, previously, we have demonstrated that PL effectively
decreases astrogliosis and microglia activation in the parietal
cortex in animal models of AD (17). The results indicated that the
inflammation and microglia activation that was triggered by
pathological conditions were effectively suppressed by PL
treatment.
The precise mechanism of action through which PL
improves cognitive function remains unclear. The results of this
study demonstrated that PL modulates the NR2B subunit of the NMDA
receptor and CaMKII in the hippocampus (Fig. 3). The phosphorylation of NR2B at
Tyr-1472 in hippocampus was increased by treatment with PL
(Fig. 3C and D). The level of
Tyr-1472 phosphorylation is increased after the induction of
long-term potentiation (LTP) in the hippocampus, indicating that
the phosphorylation of Tyr-1472 is involved in synaptic plasticity
(42). Additionally, CaMKII is
the main protein of post-synaptic density and is an essential
protein for the induction of NMDAR-dependent LTP (43). CaMKIIα promotes synaptic
formation, strengthening, and integration into existing neural
circuits (44).
Autophosphorylation at Thr286 of CaMKIIα is also required for
NMDAR-dependent LTP and hippocampus-dependent learning (45). However, CaMKIIα activation is
impaired in an age-dependent manner in the hippocampus and amygdala
(46). The loss of CaMKIIα
activity results in severe electrophysiological abnormalities that
are associated with impaired synaptic plasticity and memory
formation, while the overexpression of CaMKIIα improves cognitive
performance, as assessed by Morris water maze testing (45,47). NR2B-containing NMDARs is coupled
to ERK activation (48). The
present study demonstrates that the oral administration of PL also
significantly increased ERK1/2 and CREB phosphorylation in the
hippocampus (Figs. 3 and 4). One of the key signalling proteins
activated downstream of CaMKII and ERK is CREB (49,50). It has been well-documented that
CREB plays a role in LTP and memory formation (51). A reduction and deficit in CREB
signalling has been observed in aged animals (52). The phosphorylation of Ser133 seems
to be a critical step in CREB activation (51,53). Total CREB levels do not appear to
change; however, the level of p-CREB is decreased in aged rats
(53,54). Additionally, the level of p-CREB
expression has been found to be associated with performance in
emotional memory tests, where a higher level of p-CREB is
indicative of a better emotional memory performance (56,57). In the current study, PL
significantly increased the phosphorylation of CREB in the CA3
region of the hippocampus (Fig.
4). Therefore, considering the functional role of these
molecules in the regulation of cognitive function, the modulation
of CaMKII/ERK/CREB signalling transduction could account for the
therapeutic effect of PL.
The age-related decline in adult neurogenesis is a
well-documented process (58). In
mice, aging is associated with a decreased number of neural stem
cells in the hippocampus (59).
New-born neurons in aged mice are highly associated with
neurogenesis-dependent cognition (60). Moreover, hippocampal neurogenesis
in response to exercise and enriched environment contributes to
hippocampal plasticity (58,61). Previously, we reported that PL
markedly increases sirtuin 1 deacetylase activity in in
vitro assays (17). Sirtuin 1
is one of seven mammalian sirtuins and has been shown to modulate
aging and memory (62,63). Although the regulation of
neurogenesis by sirtuin 1 has not been investigated in this study,
it has been reported that the activation of sirtuin 1 restores
cognitive performance and neurogenesis in mice exhibiting reduced
adult neurogenesis and lowered hippocampal cognitive abilities
(64). In the present study,
there were few DCX-positive neuroblasts in the DG of 25-month-old
female mice (Fig. 5). Moreover,
the aged mice treated with PL exhibited significantly higher number
of DCX-positive cells in the DG than in the aged mice treated with
the vehicle (Fig. 5). These
results suggest that PL may have an effect on neurogenesis by
preventing or reversing age-related decline. However, the precise
mechanisms responsible for the effect of PL on neurogenesis in aged
mice are not yet clear. Further studies, therefore, are warranted
to investigate the effects of PL on neurogenesis, including in
in vitro models. Additionally, studies on target mediators
of signalling pathways involved in the formation of new neurons can
be utilized to determine the effect of PL on neurogenesis in the
adult brain.
In conclusion, our in vivo analysis of aged
female mice demonstrates that PL improves some properties of aging,
such as age-associated cognitive impairments, synaptic dysfunction
and the decline in neurogenesis. Although additional studies are
required to elucidate the underlying molecular mechanisms and
validate the anti-aging effects of PL in male mice, the results of
the present study suggest that the activation of NR2B, CaMKIIα,
ERK1/2 and CREB, and the increase in neurogenesis following PL
treatment may contribute to hippocampal neuronal activity in the
aged brain.
Acknowledgments
The authors would like to thank Dr Jae-Ran Lee
(KRIBB, Republic of Korea) for the gift of GluR1 antiserum and Mr.
In-Bok Lee, Ms. Jung-Hyun Choi, Mr. Young-Keun Choi and Ms.
Yun-Jeong Seo for their technical assistance.
Abbreviations:
|
PL
|
piperlongumine
|
|
AD
|
Alzheimer's disease
|
|
EtOH
|
ethanol
|
|
EtOAc
|
ethyl acetate
|
|
MeOH
|
methanol
|
|
CMC
|
carboxymethyl cellulose
|
|
VGLUT1
|
vesicular glutamate transporter 1
|
|
VGLUT2
|
vesicular glutamate transporter 2
|
|
NR2B
|
N-methyl-D-aspartate receptor subtype
2B
|
|
PSD-95
|
postsynaptic density protein 95
|
|
GAD65/67
|
glutamate decarboxylase 65/67
|
|
VGAT
|
vesicular GABA transporter
|
|
CREB
|
cAMP response element binding
protein
|
|
CaMKIIα
|
calcium/calmodulin-dependent protein
kinase type II α
|
|
ERK1/2
|
extracellular signal-regulated kinases
1/2
|
|
PBS
|
phosphate-buffered saline
|
|
Iba1
|
ionized calcium-binding adapter
molecule 1
|
|
GFAP
|
glial fibrillary acidic protein
|
|
4-HNE
|
4-hydroxy-2-nonenal
|
|
LTP
|
long-term potentiation
|
Funding
This study was supported by the KRIBB Research
Initiative Program of the Republic of Korea, and the Development of
Platform Technology for Innovative Medical Measurements funded by
Korea Research Institute of Standards and Science
(KRISS-2017-GP2017-0020).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JG, TSP, HYP, GHH, and YKR carried out the
experiment and analysed the data. CHL, SK, WKO, and KSK conceived
and planned the experiments. JG, CHL, and KSK wrote the manuscript.
YHK, JHH, DHC, DYH and JRN contributed to sample preparation and
analysed the data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the
Institutional Animal Use and Care Committee of the KRIBB
(KRIBB-AEC-14074).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Morrison JH and Hof PR: Life and death of
neurons in the aging brain. Science. 278:412–419. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bettio LEB, Rajendran L and Gil-Mohapel J:
The effects of aging in the hippocampus and cognitive decline.
Neurosci Biobehav Rev. 79:66–86. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Aaboe K, Knop FK, Vilsboll T, Vølund A,
Simonsen U, Deacon CF, Madsbad S, Holst JJ and Krarup T: KATP
channel closure ameliorates the impaired insulinotropic effect of
glucose-dependent insulinotropic polypeptide in patients with type
2 diabetes. J Clin Endocrinol Metab. 94:603–608. 2009. View Article : Google Scholar
|
|
4
|
Geinisman Y, Detoledo-Morrell L, Morrell F
and Heller RE: Hippocampal markers of age-related memory
dysfunction: Behavioral, electrophysiological and morphological
perspectives. Prog Neurobiol. 45:223–252. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Driscoll I, Howard SR, Stone JC, Monfils
MH, Tomanek B, Brooks WM and Sutherland RJ: The aging hippocampus:
A multi-level analysis in the rat. Neuroscience. 139:1173–1185.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Griffin R, Nally R, Nolan Y, McCartney Y,
Linden J and Lynch MA: The age-related attenuation in long-term
potentiation is associated with microglial activation. J Neurochem.
99:1263–1272. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ojo JO, Rezaie P, Gabbott PL and Stewart
MG: Impact of age-related neuroglial cell responses on hippocampal
deterioration. Front Aging Neurosci. 7:572015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gureviciene I, Gurevicius K and Tanila H:
Aging and alpha-synuclein affect synaptic plasticity in the dentate
gyrus. J Neural Transm (Vienna). 116:13–22. 2009. View Article : Google Scholar
|
|
9
|
Lister JP and Barnes CA: Neurobiological
changes in the hippocampus during normative aging. Arch Neurol.
66:829–833. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nyffeler M, Zhang WN, Feldon J and Knuesel
I: Differential expression of PSD proteins in age-related spatial
learning impairments. Neurobiol Aging. 28:143–155. 2007. View Article : Google Scholar
|
|
11
|
Bezerra DP, Pessoa C, de Moraes MO,
Saker-Neto N, Silveira ER and Costa-Lotufo LV: Overview of the
therapeutic potential of piplartine (piperlongumine). Eur J Pharm
Sci. 48:453–463. 2013. View Article : Google Scholar
|
|
12
|
Cícero Bezerra Felipe F, Trajano Sousa
Filho J, de Oliveira Souza LE, Alexandre Silveira J, Esdras de
Andrade Uchoa D, Rocha Silveira E, Deusdênia Loiola Pessoa O and de
Barros Viana GS: Piplartine, an amide alkaloid from Piper
tuberculatum, presents anxiolytic and antidepressant effects in
mice. Phytomedicine. 14:605–612. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rodrigues RV, Lanznaster D, Longhi
Balbinot DT, de Gadotti VM, Facundo VA and Santos AR:
Antinociceptive effect of crude extract, fractions and three
alkaloids obtained from fruits of Piper tuberculatum. Biol Pharm
Bull. 32:1809–1812. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Raj L, Ide T, Gurkar AU, Foley M, Schenone
M, Li X, Tolliday NJ, Golub TR, Carr SA, Shamji AF, et al:
Selective killing of cancer cells by a small molecule targeting the
stress response to ROS. Nature. 475:231–234. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rao VR, Muthenna P, Shankaraiah G,
Akileshwari C, Babu KH, Suresh G, Babu KS, Chandra Kumar RS, Prasad
KR, Yadav PA, et al: Synthesis and biological evaluation of new
piplartine analogues as potent aldose reductase inhibitors (ARIs).
Eur J Med Chem. 57:344–361. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Navickiene HM, Alécio AC, Kato MJ, Bolzani
VD, Young MC, Cavalheiro AJ and Furlan M: Antifungal amides from
Piper hispidum and Piper tuberculatum. Phytochemistry. 55:621–626.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Go J, Ha TKQ, Seo JY, Park TS, Ryu YK,
Park HY, Noh JR, Kim YH, Hwang JH, Choi DH, et al: Piperlongumine
activates Sirtuin1 and improves cognitive function in a murine
model of Alzheimer's disease. J Funct Foods. 43:103–111. 2018.
View Article : Google Scholar
|
|
18
|
Peng S, Zhang B, Meng X, Yao J and Fang J:
Synthesis of piper-longumine analogues and discovery of nuclear
factor erythroid 2-related factor 2 (Nrf2) activators as potential
neuroprotective agents. J Med Chem. 58:5242–5255. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tabuneng W, Bando H and Amiya T: Studies
on the constituents of the crude drug 'piperis longi fructus'. On
the alkaloids of fruits of piper longum L. Chem Pharm Bull.
31:3562–3565. 1983. View Article : Google Scholar
|
|
20
|
Jang S, Dilger RN and Johnson RW: Luteolin
inhibits microglia and alters hippocampal-dependent spatial working
memory in aged mice. J Nutr. 140:1892–1898. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park HY, Ryu YK, Kim YH, Park TS, Go J,
Hwang JH, Choi DH, Rhee M, Lee CH and Kim KS: Gadd45β ameliorates
L-DOPA-induced dyskinesia in a Parkinson's disease mouse model.
Neurobiol Dis. 89:169–179. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Park TS, Ryu YK, Park HY, Kim JY, Go J,
Noh JR, Kim YH, Hwang JH, Choi DH, Oh WK, et al: Humulus japonicus
inhibits the progression of Alzheimer's disease in a APP/PS1
transgenic mouse model. Int J Mol Med. 39:21–30. 2017. View Article : Google Scholar
|
|
23
|
Deacon RM, Cholerton LL, Talbot K,
Nair-Roberts RG, Sanderson DJ, Romberg C, Koros E, Bornemann KD and
Rawlins JN: Age-dependent and -independent behavioral deficits in
Tg2576 mice. Behav Brain Res. 189:126–138. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim YJ, Kang Y, Park HY, Lee JR, Yu DY,
Murata T, Gondo Y, Hwang JH, Kim YH, Lee CH, et al: STEP signaling
pathway mediates psychomotor stimulation and morphine withdrawal
symptoms, but not for reward, analgesia and tolerance. Exp Mol Med.
48:e2122016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ryu YK, Kang Y, Go J, Park HY, Noh JR, Kim
YH, Hwang JH, Choi DH, Han SS, Oh WK, et al: Humulus japonicus
prevents dopaminergic neuron death in 6-hydroxydopamine-induced
models of Parkinson's disease. J Med Food. 20:116–123. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ryu YK, Park HY, Go J, Choi DH, Kim YH,
Hwang JH, Noh JR, Lee TG, Lee CH and Kim KS: Metformin inhibits the
development of L-DOPA-induced dyskinesia in a murine model of
Parkinson's disease. Mol Neurobiol. 55:5715–5726. 2018. View Article : Google Scholar
|
|
27
|
Wesson DW and Wilson DA: Age and gene
overexpression interact to abolish nesting behavior in Tg2576
amyloid precursor protein (APP) mice. Behav Brain Res. 216:408–413.
2011. View Article : Google Scholar
|
|
28
|
Filali M, Lalonde R and Rivest S:
Subchronic memantine administration on spatial learning,
exploratory activity, and nest-building in an APP/PS1 mouse model
of Alzheimer's disease. Neuropharmacology. 60:930–936. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Deacon RM, Croucher A and Rawlins JN:
Hippocampal cytotoxic lesion effects on species-typical behaviours
in mice. Behav Brain Res. 132:203–213. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cribbs DH, Berchtold NC, Perreau V,
Coleman PD, Rogers J, Tenner AJ and Cotman CW: Extensive innate
immune gene activation accompanies brain aging, increasing
vulnerability to cognitive decline and neurodegeneration: A
microarray study. J Neuroinflamm. 9:1792012. View Article : Google Scholar
|
|
31
|
Harman D: Aging: A theory based on free
radical and radiation chemistry. J Gerontol. 11:298–300. 1956.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stebbings KA, Choi HW, Ravindra A and
Llano DA: The impact of aging, hearing loss, and body weight on
mouse hippocampal redox state, measured in brain slices using
fluorescence imaging. Neurobiol Aging. 42:101–109. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cini M and Moretti A: Studies on lipid
peroxidation and protein oxidation in the aging brain. Neurobiol
Aging. 16:53–57. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
von Bernhardi R, Eugenín-von Bernhardi L
and Eugenín J: Microglial cell dysregulation in brain aging and
neurodegeneration. Front Aging Neurosci. 7:1242015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee SW, Clemenson GD and Gage FH: New
neurons in an aged brain. Behav Brain Res. 227:497–507. 2012.
View Article : Google Scholar :
|
|
36
|
Bharadwaj U, Eckols TK, Kolosov M,
Kasembeli MM, Adam A, Torres D, Zhang X, Dobrolecki LE, Wei W,
Lewis MT, et al: Drug-repositioning screening identified
piperlongumine as a direct STAT3 inhibitor with potent activity
against breast cancer. Oncogene. 34:1341–1353. 2015. View Article : Google Scholar
|
|
37
|
Balducci L and Ershler WB: Cancer and
ageing: A nexus at several levels. Nat Rev Cancer. 5:655–662. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Salminen A, Ojala J, Kaarniranta K,
Haapasalo A, Hiltunen M and Soininen H: Astrocytes in the aging
brain express characteristics of senescence-associated secretory
phenotype. Eur J Neurosci. 34:3–11. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Campisi J: Aging, cellular senescence, and
cancer. Ann Rev Physiol. 75:685–705. 2013. View Article : Google Scholar
|
|
40
|
Wang Y, Chang J, Liu X, Zhang X, Zhang S,
Zhang X, Zhou D and Zheng G: Discovery of piperlongumine as a
potential novel lead for the development of senolytic agents. Aging
(Albany NY). 8:2915–2926. 2016. View Article : Google Scholar
|
|
41
|
Nichols NR, Day JR, Laping NJ, Johnson SA
and Finch CE: GFAP mRNA increases with age in rat and human brain.
Neurobiol Aging. 14:421–429. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nakazawa T, Komai S, Tezuka T, Hisatsune
C, Umemori H, Semba K, Mishina M, Manabe T and Yamamoto T:
Characterization of Fyn-mediated tyrosine phosphorylation sites on
GluR epsilon 2 (NR2B) subunit of the N-methyl-D-aspartate receptor.
J Biol Chem. 276:693–699. 2001. View Article : Google Scholar
|
|
43
|
Lisman J, Schulman H and Cline H: The
molecular basis of CaMKII function in synaptic and behavioural
memory. Nat Rev Neurosci. 3:175–190. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Asrican B, Lisman J and Otmakhov N:
Synaptic strength of individual spines correlates with bound
Ca2+-calmodulin-dependent kinase II. J Neurosci.
27:14007–14011. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Giese KP, Fedorov NB, Filipkowski RK and
Silva AJ: Autophosphorylation at Thr286 of the alpha
calcium-calmodulin kinase II in LTP and learning. Science.
279:870–873. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fang T, Kasbi K, Rothe S, Aziz W and Giese
KP: Age-dependent changes in autophosphorylation of alpha
calcium/calmodulin dependent kinase II in hippocampus and amygdala
after contextual fear conditioning. Brain Res Bull. 134:18–23.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Elgersma Y, Sweatt JD and Giese KP: Mouse
genetic approaches to investigating calcium/calmodulin-dependent
protein kinase II function in plasticity and cognition. J Neurosci.
24:8410–8415. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Krapivinsky G, Krapivinsky L, Manasian Y,
Ivanov A, Tyzio R, Pellegrino C, Ben-Ari Y, Clapham DE and Medina
I: The NMDA receptor is coupled to the ERK pathway by a direct
interaction between NR2B and RasGRF1. Neuron. 40:775–784. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kreusser MM and Backs J: Integrated
mechanisms of CaMKII-dependent ventricular remodeling. Front
Pharmacol. 5:362014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Tully T, Bourtchouladze R, Scott R and
Tallman J: Targeting the CREB pathway for memory enhancers. Nat Rev
Drug Discov. 2:267–277. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Silva AJ, Kogan JH, Frankland PW and Kida
S: CREB and memory. Ann Rev Neurosci. 21:127–148. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Bach ME, Barad M, Son H, Zhuo M, Lu YF,
Shih R, Mansuy I, Hawkins RD and Kandel ER: Age-related defects in
spatial memory are correlated with defects in the late phase of
hippo-campal long-term potentiation in vitro and are attenuated by
drugs that enhance the cAMP signaling pathway. Proc Natl Acad Sci
USA. 96:5280–5285. 1999. View Article : Google Scholar
|
|
53
|
Gonzalez GA and Montminy MR: Cyclic AMP
stimulates somatostatin gene transcription by phosphorylation of
CREB at serine 133. Cell. 59:675–680. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Monti B, Berteotti C and Contestabile A:
Dysregulation of memory-related proteins in the hippocampus of aged
rats and their relation with cognitive impairment. Hippocampus.
15:1041–1049. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hattiangady B, Rao MS, Shetty GA and
Shetty AK: Brain-derived neurotrophic factor, phosphorylated cyclic
AMP response element binding protein and neuropeptide Y decline as
early as middle age in the dentate gyrus and CA1 and CA3 subfields
of the hippocampus. Exp Neurol. 195:353–371. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Cowansage KK, Bush DE, Josselyn SA, Klann
E and Ledoux JE: Basal variability in CREB phosphorylation predicts
trait-like differences in amygdala-dependent memory. Proc Natl Acad
Sci USA. 110:16645–16650. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Yu XW, Oh MM and Disterhoft JF: CREB,
cellular excitability, and cognition: Implications for aging. Behav
Brain Res. 322:206–211. 2017. View Article : Google Scholar :
|
|
58
|
Fan X, Wheatley EG and Villeda SA:
Mechanisms of Hippocampal Aging and the Potential for Rejuvenation.
Ann Rev Neurosci. 40:251–272. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Encinas JM, Michurina TV, Peunova N, Park
JH, Tordo J, Peterson DA, Fishell G, Koulakov A and Enikolopov G:
Division-coupled astrocytic differentiation and age-related
depletion of neural stem cells in the adult hippocampus. Cell Stem
Cell. 8:566–579. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Morgenstern NA, Lombardi G and Schinder
AF: Newborn granule cells in the ageing dentate gyrus. J Physiol.
586:3751–3757. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kempermann G, Gast D and Gage FH:
Neuroplasticity in old age: Sustained fivefold induction of
hippocampal neurogenesis by long-term environmental enrichment. Ann
Neurol. 52:135–143. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Herskovits AZ and Guarente L: Sirtuin
deacetylases in neurodegenerative diseases of aging. Cell Res.
23:746–758. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Michan S, Li Y, Chou MM, Parrella E, Ge H,
Long JM, Allard JS, Lewis K, Miller M, Xu W, et al: SIRT1 is
essential for normal cognitive function and synaptic plasticity. J
Neurosci. 30:9695–9707. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Sellner S, Paricio-Montesinos R, Spieß A,
Masuch A, Erny D, Harsan LA, Elverfeldt DV, Schwabenland M, Biber
K, Staszewski O, et al: Microglial CX3CR1 promotes adult
neurogenesis by inhibiting Sirt 1/p65 signaling independent of
CX3CL1. Acta Neuropathol Commun. 4:1022016. View Article : Google Scholar : PubMed/NCBI
|