Introduction
Malignant gliomas are a life-threatening form of
primary brain cancer characterized by uncontrollable and
infiltrative growth that destroys surrounding normal brain tissues
and causes neurological deficits (1). Following maximal surgical tumor
excision, the standard treatment is chemo-radiotherapy. Despite
multidisciplinary treatment approaches, gliomas have a high rate of
recurrence, with few patients surviving >1 year (2). The failure of current therapeutics
is partly attributed to drug resistance. Therefore, it is critical
to focus on identifying novel target genes and the molecular
mechanisms involved in the restoration of drug sensitivity
(3).
Forkhead box O1 (FOXO1) belongs to the FOXO family
of transcription factors, which are characterized by a conserved
winged-helix DNA binding domain (4). The gene encoding FOXO1 is located at
chromosoe 13q14, where methylation, mutation and allelic losses are
common occurrences in the presence of cancer. These characteristics
suggest that there are potential tumor-associated genes involved in
the origination and progression of human malignancies harbored in
this region (5,6). Increasing evidence has demonstrated
that FOXO1 is downregulated in several types of cancer with adverse
outcomes (7). In addition, FOXO1
proteins are usually accumulated in the nucleus and act as a
transcriptional regulator in non-tumor tissues. Once FOXO1 is
phosphorylated in tumors, its proteins can migrate to the cytoplasm
and become inactive, eliminating the expression of certain
anticancer genes and leading to tumorigenesis (8). FOXO1 is able to inhibit tumor growth
and has been identified as a tumor suppressor gene (TSG). Previous
studies have shown that FOXO1 inhibited proliferation, prevented
invasion and induced apoptosis in gliomas (9). Other data have also provided
evidence that FOXO1 reversed chemotherapy resistance in certain
types of cancer (10), however,
this function in glioma remains to be fully elucidated.
Multidrug resistance (MDR) is a major barrier to
radiotherapy and chemotherapy in cancer cells. A variety of
mechanisms induce MDR phenotypes, however, the upregulation of
ATP-binding cassette transporters represents the most common factor
involved in MDR development (11). Trifluoperazine (TFP) is a
phenothiazine derivative and is widely used as an antipsychotic
drug. TFP may be clinically potent as a calmodulin antagonist and
an inhibitor of the dopamine receptor (12). It has been reported that TFP
reverses drug resistance in tumors through the inhibition of MDR
genes, including P-glycoprotein (P-gp) (13). Of note, the anticancer effects of
doxorubicin (DOX), bleomycin and gefitinib were found to be
reinforced when TFP was combined with them (13-15). This suggests that TFP may be a
valuable tool in overcoming MDR in cancer.
In the present study, the effects of FOXO1 on MDR
phenotypes in glioma cells were examined. The data indicated that
FOXO1 is a TSG, and that the nuclear translocation of FOXO1 was
conducive for exerting anticancer functions. It was also confirmed
that TFP may overcome drug resistance by limiting the nuclear
excretion of FOXO1 in gliomas. This identification may accelerate
TFP as a molecular therapy for gliomas.
Materials and methods
Reagents
The glioma SHG44 cell line was from Shanghai Life
Academy of Sciences Cell Library (Shanghai, China). RPMI-1640
medium and fetal bovine serum (FBS) were obtained from Gibco;
Thermo Fisher Scientific, Inc. (Waltham, MA, USA). Penicillin and
streptomycin were purchased from HyClone; GE Healthcare Life
Sciences (Logan, UT, USA). Primary antibodies targeting
P-glycoprotein (P-gp, cat. no. sc-13131), multidrug
resistance-associated protein 1 (MRP1, cat. no. sc-18835), lung
resistance protein (LRP, cat. no. sc-23916), Ki67 nuclear antigen
(Ki67, cat. no. sc-15402), and proliferating cell nuclear antigen
(PCNA, cat. no. sc-25280) were purchased from Santa Cruz
Biotechnology, Inc. (San Francisco, CA, USA). Antibodies targeting
FOXO1 (cat. no. ab52857), α-tubulin (cat. no. ab18251) and Lamin B1
(cat. no. ab133741) were purchased from Abcam (Cambridge, MA, USA).
Horseradish peroxidase- (cat. no. ZB2301) or TRITC-conjugated (cat.
no. ZF0313) secondary antibodies were purchased from Zhongshan
Golden Bridge Biotechnology (Beijing, China). Epidermal growth
factor (EGF), DAPI, RNase, propidium iodide (PI), the Annexin
V-PE/7-AAD apoptosis reagent kit and TFP were purchased from Sigma;
EMD Millipore (Billerica, MA, USA). The caspase-3 activity assay
kit (cat. no. 12012952001) was purchased from Roche Diagnostics
GmbH (Mannheim, Germany). DOX (cat. no. D1515; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany). The RNAiso Plus, primescript RT reagent
kit and DNA polymerase were purchased from Takara Biotechnology
Co., Ltd. (Dalian, China). Cell Counting Kit-8 (CCK8), BSA,
bicinchoninic acid (BCA) kit and the protein extraction kits
(total, nuclear and cytosolic) were purchased from Beyotime
Institute of Biotechnology (Beijing, China).
Cell culture
The human glioma SHG44 cell line and the
DOX-resistant SHG44 cell line (SHG44/DOX) have been described
previously (16). In brief, the
SHG44 cells were maintained in a 5% CO2 atmosphere at
37°C in RPMI-1640 medium supplemented with 100 U/ml penicillin, 100
mg/ml streptomycin and 10% FBS. The concentrations of DOX were
gradually increased between 0.01 and 1 μg/ml, resulting in
the SHG44/DOX cells being able to grow in 0.1 μg/ml DOX.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from the cells using RNAiso
Plus. The RNA sample concentrations were measured using a
spectrophotometer and then reverse transcribed into cDNA using a
primescript RT reagent kit. The primer sequences were as follows:
MDR1 forward, 5′-CCCATCATTGCAATAGCAGG-3′ and reverse,
5′-GTTCAAACTTCTCTGCTGCTCCTGA-3′; MRP1 forward,
5′-GGCATCTCAGCAACTCGTCTT-3′ and reverse,
5′-ATTAGCTTCCACGTCTCCTCCTT-3′; LRP forward,
5′-ACAACTACTGCGTGATTCTC-3′ and reverse, 5′-CTAGCATGTAGGTGCTTCCA-3′;
and GAPDH forward, 5′-CTTTGGTATCGTGGAAGGACTC-3′ and reverse,
5′-GTAGAGGCAGGGATGATGTTCT-3′. The reaction systems were as follows:
The DNA polymerase 10 μl, forward primer sequences (10
μM) 0.8 μl, reverse primer sequences (10 μM)
0.8 μl, cDNA 2.0 μl and DEPC 6.4 μl. The
amplification conditions were as follows: 95°C for 30 sec, followed
by 40 cycles at 95°C for 15 sec, and 60°C for 45 sec. The relative
fold-changes in mRNA levels were calculated with the
2-ΔΔCq method (17).
Western blot analysis
The nuclear, cytoplasmic and total proteins were
extracted using their respective extraction kits according to the
manufacturer’s protocols. The protein concentrations were detected
using the BCA method. An equal quantity (35 μg) of each
sample was separated by 6-12% SDS-PAGE and transferred onto PVDF
membranes. The membranes were blocked with 5% goat serum for 1 h at
4°C and incubated with FOXO1 (dilution 1:200), P-gp (dilution
1:300), MRP1 (dilution 1:300), LRP (dilution 1:300), GAPDH
(dilution 1:1,000), α-tubulin (dilution 1:500) and Lamin B1
(dilution 1:500) primary antibodies overnight at 4°C, and then
washed with 5% TBST for three times and incubated with secondary
antibody (dilution 1:5,000) for 1 h at 37°C. The membranes were
then washed three times, and the protein in each band was
quantified using Quantity One 4.6 computer software (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Immunofluorescence
The SHG44 and SHG44/DOX cells were collected to
mount on the coverslip and fixed in 4% paraformaldehyde for 20 min.
The cells were blocked with 5% BSA for 45 min then incubated with
FOXO1 primary antibody (dilution 1:100) overnight at 4°C. The cells
were then stained with a TRITC-labeled secondary antibody (1 h,
dilution 1:500) and DAPI (4 min). The sections were coverslipped
with 50% glycerol, and the location of the FOXO1 proteins was
detected using a laser scanning confocal microscope (TCS SP2, Leica
Microsystems GmbH, Wetzlar, Germany). For the negative control, 5%
BSA was used in place of the primary antibodies.
Cell viability
The SHG44/DOX cells were divided into a blank group,
a DOX group and a DOX + TFP group. The cells were grown in 96-well
plates at a density of 2,000 cells/well. The cells were washed with
PBS following various treatments (complete medium for blank group,
0.1 μg/ml DOX for DOX group, 10 μM TFP + 0.1
μg/ml DOX for DOX + TFP group) for 24, 48 and 72 h. A total
of 100 μl medium and 10 μl CCK-8 were added to each
well for an additional 2 h. The absorbance values (OD values) were
read at 450 nm using an enzyme-labeled instrument.
Flow cytometry
For cell cycle distribution, 5×105 cells
were harvested from each group and fixed in 70% ice-cold ethanol
overnight. The cells were then treated with 10 mg/ml RNase and 50
mg/ml PI at 37°C for 30 min in the dark. The cell cycle
distribution was analyzed by flow cytometry (BD Biosciences,
Franklin Lakes, NJ, USA). Apoptosis was measured by flow cytometry
using the Annexin V-PE/7-AAD apoptosis reagent kit according to the
manufacturer’s protocol. The SHG44/DOX cells from the blank group,
the DOX group and the DOX + TFP group were harvested, re-suspended,
and stained with phycoerythrin (PE)-labeled Annexin V and
7-aminoactinomycin D (7-AAD) for measuring early apoptosis.
Caspase-3 activity assay
To determine the intra-cellular caspase-3 activity
in the SHG44/DOX control group, the DOX group and the DOX + TFP
group, a commercial caspase-3 activity assay kit was utilized
according to the manufacturer’s protocol. The caspase-3 activities
in implanted tumors were also analyzed following injection with DOX
or DOX + TFP. The values are presented as the percentage of the
blank control.
DOX uptake
The SHG44/DOX cells were exposed to 0.1 μg/ml
DOX, 0.1 μg/ml DOX + 10 μM TFP or 0.1 μg/ml
DOX + 10 μM TFP + EGF for 2 h. Following cell lysis and
supernatant collection, the intracellular concentrations of DOX
were measured using a spectrophotometer (absorbance: 490 nm), and
the protein concentrations were measured for standardizing the
uptake of DOX.
Xenograft tumor model
All animal experiments were approved by the Ethics
Committee of Chongqing Medical University (Chonqing, China). A
total of 24 Male nude mice (4 weeks old, 14.4±3.1 g) were provided
by the Experimental Animal Center of Chongqing Medical University.
The nude mice were housed in light for 10 h/day at 26°C. Food and
water were sterilized by high pressure steam and multivitamins were
added into distilled water. The subcutaneous tumor model was
produced as previously described (18). For treatment, 5 mg/kg DOX alone or
5 mg/kg DOX + 5 mg/kg/day TFP was injected the tail vein (DOX every
7 days and TFP every day). Tumor volumes were recorded at 7, 14, 21
and 28 days post-implantation in accordance with the previously
described formula (18).
Immunohistochemistry (IHC)
The mice were injected with 10% chloral hydrate (300
mg/kg) for anesthesia and sacrificed depending on cervical vertebra
dislocation at 28 days. No mice exhibited signs of peritonitis. The
tumor samples were dissected and embedded in paraffin.
Paraffin-embedded sections (4 mm) were prepared and IHC procedures
were performed based on prior methods (19). The proliferation indices of Ki-67
(dilution 1:100) and PCNA (dilution 1:100) were defined as the
percentage of positive cells from five randomly selected fields at
×400 magnification using a laser scanning confocal microscope (TCS
SP2; Leica Microsystems GmbH, Wetzlar, Germany).
Statistical analysis
Statistical analyses were performed using SPSS 20.0
software (IBM SPSS, Armonk, NY, USA). Statistical differences among
groups were analyzed by one-way analysis of variance and the
Student-Neuman-Keuls post hoc test, two-sample t-test or
χ2 test. P<0.05 was considered to indicate a
statistically significant difference. All data are presented as the
mean ± standard deviation.
Results
Downregulation of nuclear FOXO1 in
SHG44/DOX cells
The SHG44/DOX cells were established according to
previous methods. The RT-qPCR analysis showed that three MDR genes
(MDR1, MRP1 and LRP) were upregulated in the SHG44/DOX cells
compared with the SHG44 cells (Fig.
1A). The protein levels of P-gp, MRP1 and LRP were also
increased in the SHG44/DOX cells, as determined by western blot
analysis (Fig. 1B). The levels of
nuclear FOXO1 (dephospho-FOXO1) were lower in the SHG44/DOX cells
than in the SHG44 cells, whereas a higher expression of cytoplasmic
FOXO1 (p-FOXO1) was detected in the SHG44/DOX cells (Fig. 1C). The immunofluorescence also
verified that the FOXO1 proteins were excreted into the cytoplasm
in SHG44/DOX cells (Fig. 1D).
These data suggested that FOXO1 proteins were expressed in the
cytoplasm, resulting in a loss of transcriptional activity and
tumor inhibitory effects in the SHG44/DOX drug-resistant glioma
cells.
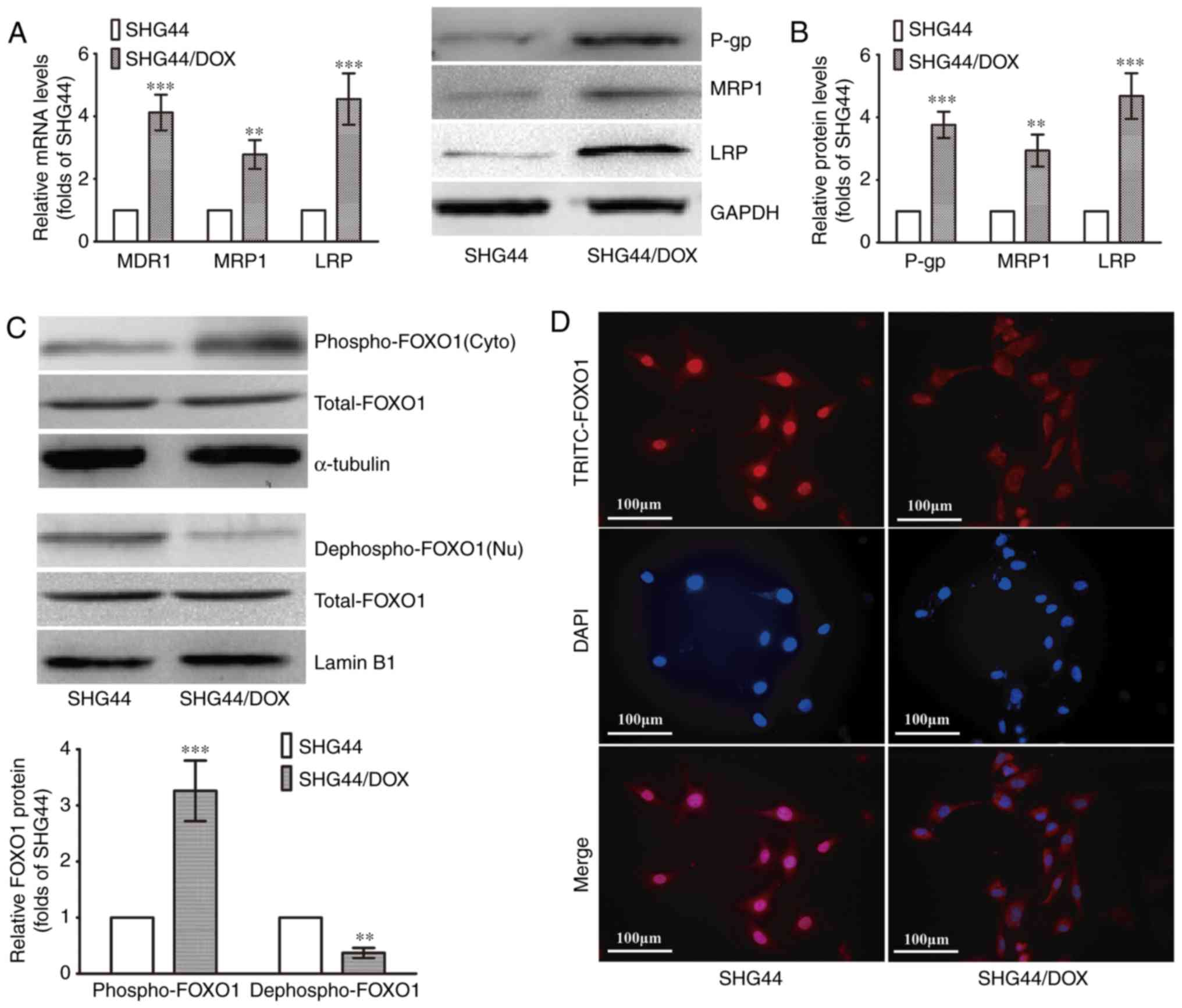 | Figure 1Expression of FOXO1 and
MDR-associated molecules analyzed in SHG44 and SHG44/DOX glioma
cells. (A) mRNA levels of MDR1, MRP1, LRP in SHG44 and SHG44/DOX
cells. (B) Protein expression of P-gp, MRP1 and LRP in SHG44 and
SHG44/DOX cells. (C) Western blot analysis was used to observe
nuclear and cytoplasmic FOXO1 protein in the two cell lines.
α-tubulin was the cytoplasmic protein loading control, and Lamin B1
was the nuclear protein loading control. (n=5,
**P<0.01 and ***P<0.001, compared with
SHG44). (D) Immunofluorescence for examining the location of FOXO1
proteins in the two cell lines. FOXO1, Forkhead box O1; DOX,
doxorubicin; MDR, multidrug resistance; LRP, lung resistance
protein; P-gp, P-glycoprotein; Nu, nuclear; Cyto, cytoplasmic. |
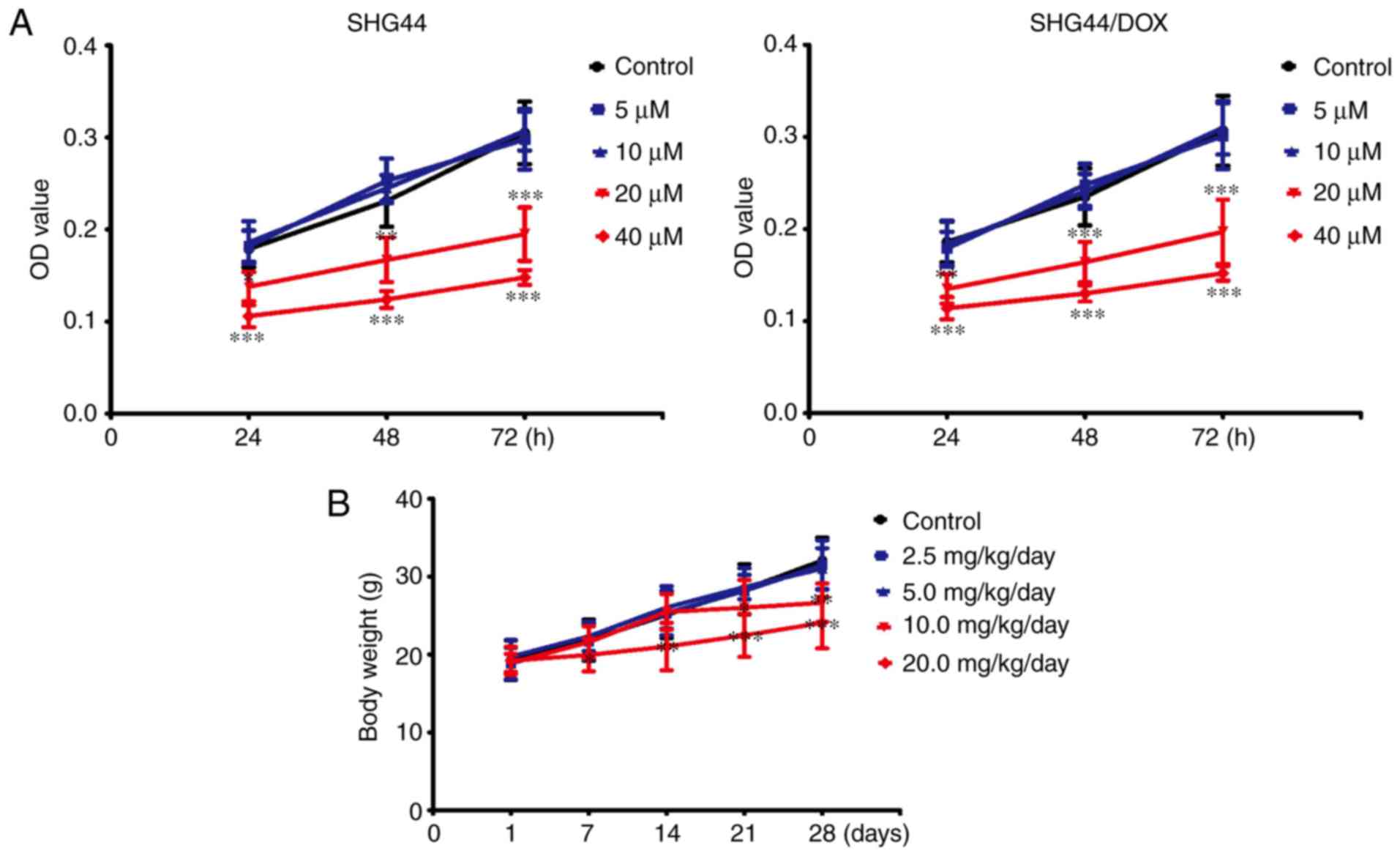
Non-toxic concentrations of TFP in vitro
and in vivo
To avoid toxicity from TFP in the SHG44 and
SHG44/DOX cells, the appropriate dose of TFP was selected.
Following treatment with concentrations of ≤10 μM for 24,
48, or 72 h, TFP did not inhibit growth of the two selected cell
lines. However, TFP may reduce cell viability at a concentration of
20 μM in SHG44 and SHG44/DOX cells (Fig. 2A). TFP toxicity surveys were
performed in vivo. The mice were administered with 0, 2.5,
5, 10, or 20 mg/kg/day of TFP through tail vein injections. No
changes in bodyweight were observed with administration of 5
mg/kg/day within 4 weeks (Fig.
2B). Therefore, these non-toxic concentrations of TFP (10
μM in vitro and 5 mg/kg/day in vivo) were used
for the experiments.
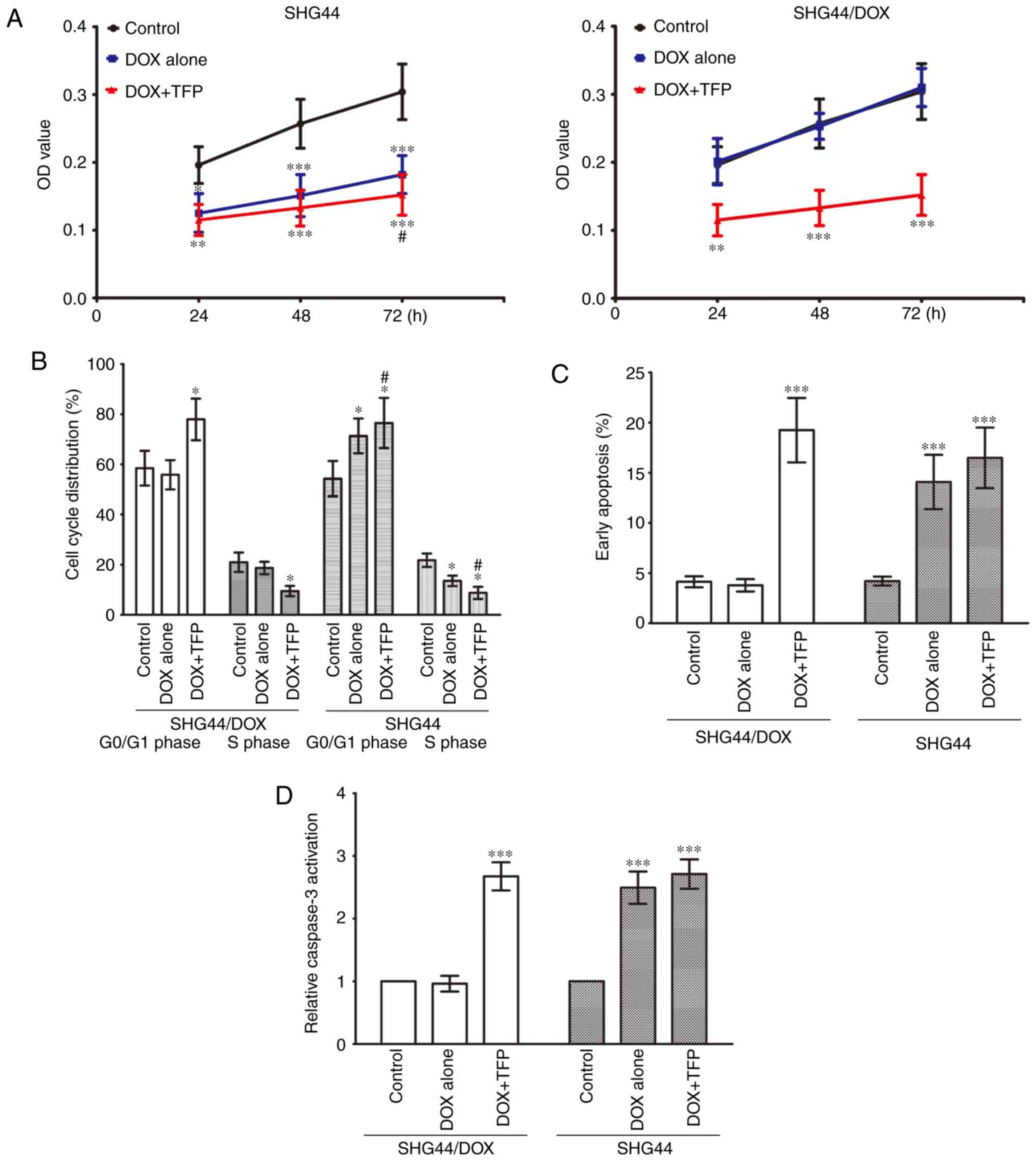
TFP overcomes DOX-resistance in SHG44/DOX
cells in vitro
The CCK-8 assay showed that the cell viability of
the SHG44 cells was suppressed by 0.1 μg/ml DOX, and 10
μM TFP marginally promoted the growth inhibition effect of
DOX at 72 h. The growth of SHG44/DOX cells was not inhibited by 0.1
μg/ml DOX, however, 10 μM TFP + 0.1 μg/ml DOX
prevented the growth of SHG44/DOX cells at 24, 48 and 72 h
(Fig. 3A). Flow cytometry
revealed that the percentage of G0/G1 phase
cells was increased and the percentage of S phase cells was
decreased in the SHG44 group following treatment with 0.1
μg/ml DOX. However, these effects were more pronounced when
10 μM TFP was added. Treatment with 0.1 μg/ml DOX did
not alter cell cycle distribution in SHG44/DOX cells; however, the
percentage of G0/G1 phase cells was higher in
the 10 μM TFP + 0.1 μg/ml DOX group than in the
control group and the 0.1 μg/ml DOX group, whereas the
percentage of S phase cells was lower in the 10 μM TFP + 0.1
μg/ml DOX group than in the other two groups (Fig. 3B). Furthermore, 0.1 μg/ml
DOX induced early apoptosis in SHG44 cells, but this did not occur
in the SHG44/DOX cells. DOX did accelerate early apoptosis and
caspase-3 activities in SHG44/DOX cells at 72 h when it was
combined with 10 μM TFP (Fig.
3C and D).
TFP decreases MDR genes and facilitates
DOX uptake through restoration of the nuclear localization of FOXO1
in SHG44/DOX cells
Western blot analysis was used to elucidate the
molecular mechanisms involved in the effects of TFP inhibition of
DOX drug-resistance. As shown in Fig.
4A, there was an increase in nuclear FOXO1 and a decrease in
cytoplasmic FOXO1 in the 10 μM TFP group, coupled with a
downregulation of P-gp, MRP1 and LRP. EGF stimulates FOXO1 nuclear
exclusion (20). When EGF was
added to the TFP group, the TFP-induced FOXO1 nuclear accumulation
was counteracted, and the levels of the three MDR proteins were
increased (Fig. 4A). The changes
in the mRNA expression of MDR1, MRP1 and LRP were consistent with
their protein expression (Fig.
4B). Following treatment with 0.1 μg/ml DOX, 0.1
μg/ml DOX + 10 μM TFP, or 0.1 μg/ml DOX + 10
μM TFP + EGF for 2 h, the intracellular concentrations of
DOX were assessed. Spectrophotometer analysis revealed that 10
μM TFP was able to enhance the intracellular concentrations
of DOX in SHG44/DOX cells. However, EGF restored the expression of
MDR proteins and contributed to DOX excretion from the glioma cells
(Fig. 4C).
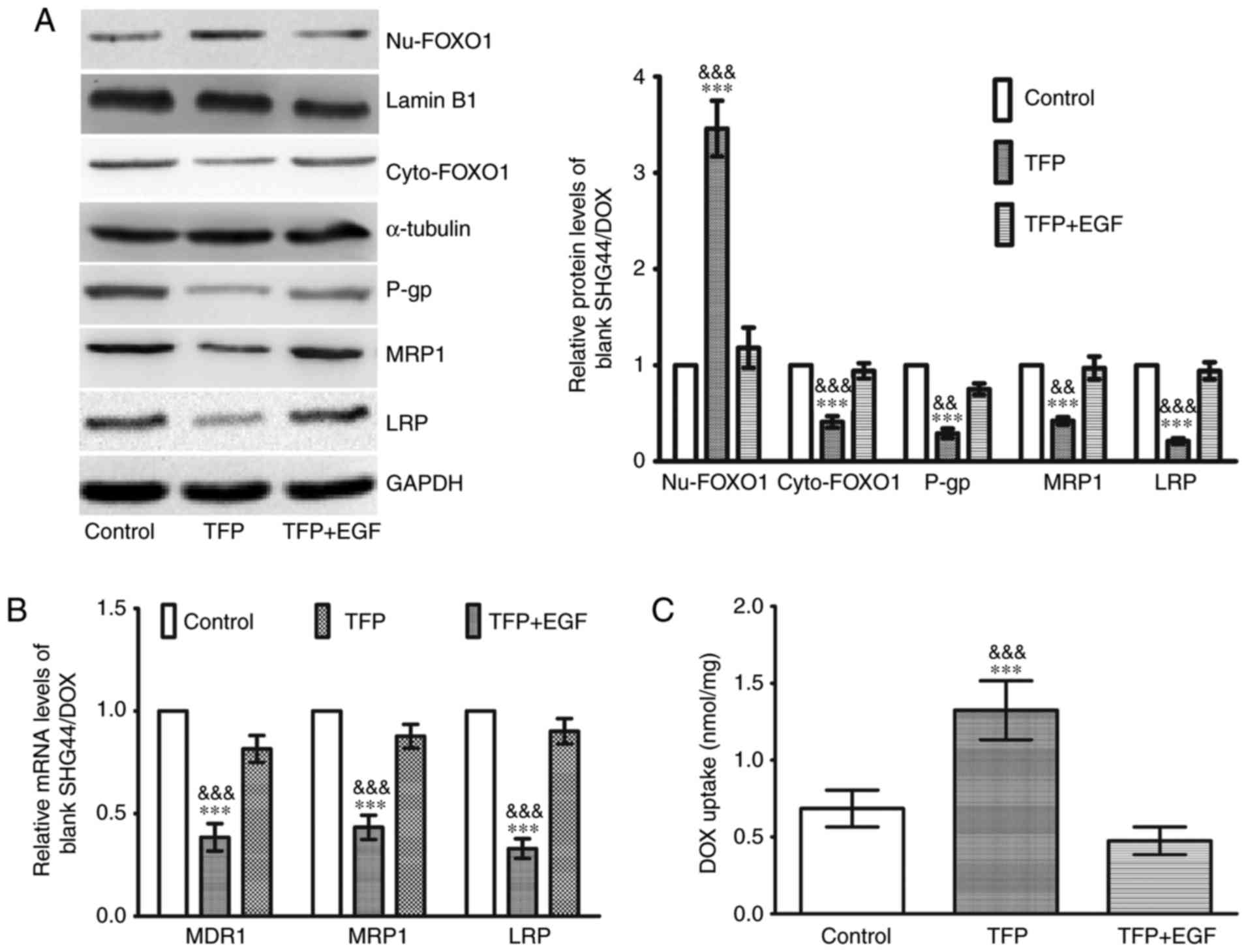 | Figure 4TFP induces DOX uptake in SHG44/DOX
cells through the inhibition of FOXO1 nuclear exclusion and MDR
genes. (A) Following treatment with TFP or TFP + EGF for 24 h,
western blot analysis showed that TFP reduced the protein levels of
P-gp, MRP1, and LRP by upregulating nuclear FOXO1 in SHG44/DOX
cells (n=5). (B) Reverse transcription-quantitative polymerase
chain reaction analysis was used to observe the effects of TFP on
mRNA levels of MDR1, MRP1 and LRP in SHG44/DOX cells (n=5). (C)
Following incubation with DOX (0.1 μg/ml) for 2 h,
spectrophotometry was used to measure intracellular concentrations
of DOX in the control, TFP and TFP + EGF groups (n=5).
***P<0.001, vs. control;
&&P<0.01 and
&&&P<0.001, vs. TFP + EGF group). FOXO1,
Forkhead box O1; MDR, multidrug resistance; DOX, doxorubicin; TFP,
trifluoperazine; P-gp, P-glycoprotein; MRP1, multidrug
resistance-associated protein 1; LRP, lung resistance protein; EGF,
epidermal growth factor; Nu, nuclear; Cyto, cytoplasmic. |
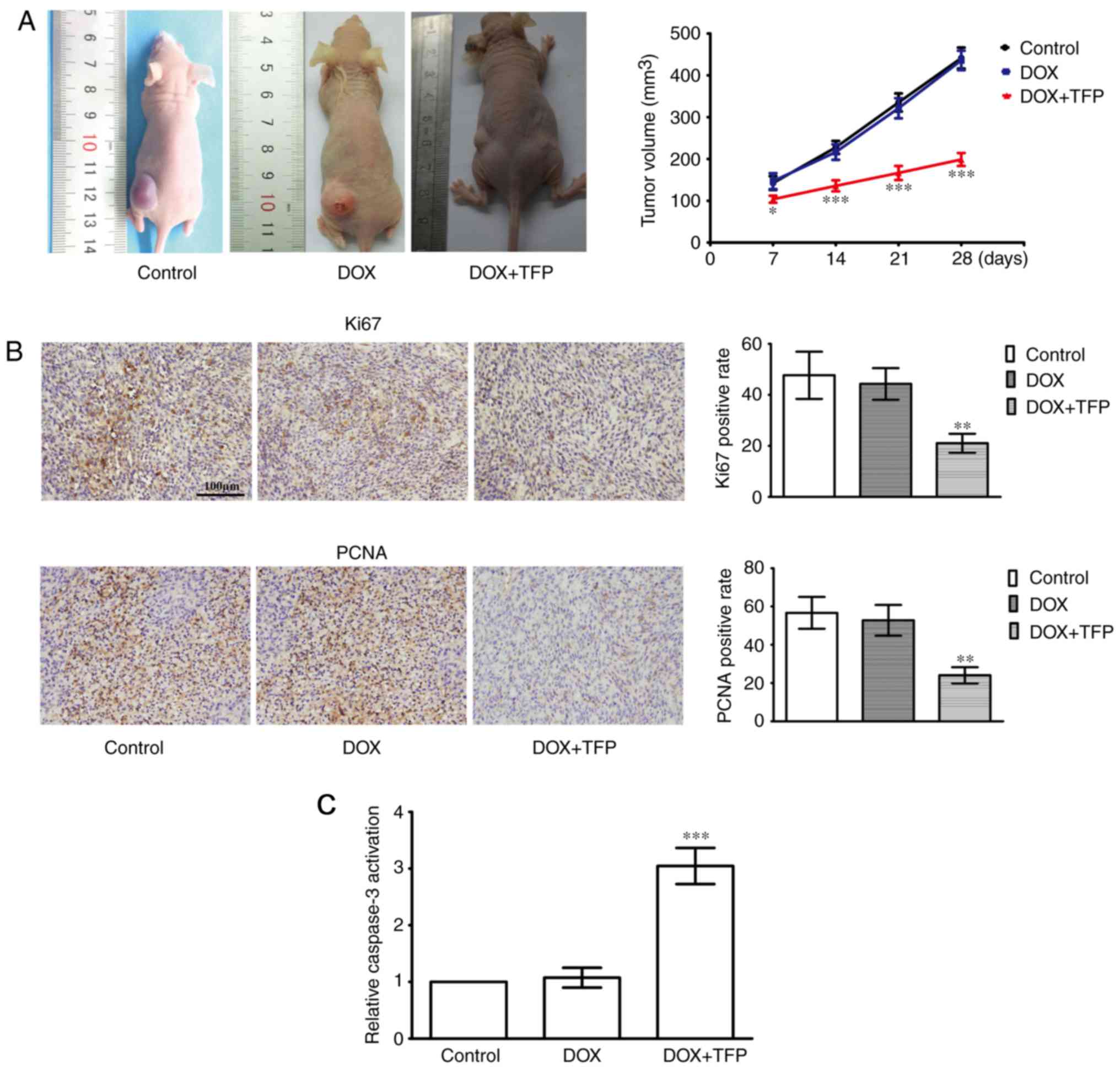
DOX-resistance of SHG44/DOX cells is
attenuated by TFP in vivo
The efficacy of TFP on the enhanced cell toxicity of
DOX was examined in subcutaneous xenotransplanted tumors. Although
5 mg/kg DOX did not suppress the xenograft tumors, 5 mg/kg DOX + 5
mg/kg/day TFP reduced SHG44/DOX growth in vivo compared with
that in the control group and DOX alone group (Fig. 5A). IHC showed that the Ki-67 and
PCNA proliferation indices were lower in the 5 mg/kg DOX + 5
mg/kg/day TFP group than that in the control group and the 5 mg/kg
DOX alone group (Fig. 5B). In
addition, the effect of DOX-induced apoptosis was also restored
when TFP (5 mg/kg/day) was added (Fig. 5C).
Discussion
Numerous studies have reported that FOXO1 functions
as a tumor suppressor and inhibits the development of different
types of cancer, with FOXO1 inactivation being accompanied with a
poor prognosis in patients (6,7).
FOXO1 contains a conserved DNA-binding domain and links to the
consensus DNA-binding sequence (TTGTTTAC) of target genes (9). For example, FOXO1 makes contact with
the p27 promoter, initiates transcription and suppresses cell cycle
progression (21). FOXO1 also
decreases tumor invasion by inhibiting matrix metalloproteinase
(MMP)7, MMP9, vascular endothelial growth factor and
hypoxia-inducible factor-1α (22,23). However, the inactivation of FOXO1
has been noted to occur in several tumor types (24-26). FOXO1 has a direct effect in the
Akt signaling pathway, which is active when malignancies are
present. Akt phosphorylates FOXO1, resulting in nuclear exclusion
and loss of binding to target regulatory elements (27,28). In the present study, it was
demonstrated that FOXO1 was conducive to enhancing chemotherapy
sensitivity and indicated that it may be involved in glioma
pathogenesis. It has been suggested that c-Jun N-terminal kinase is
an upstream regulatory molecule of FOXO1 and may inhibit its
activation, contributing to a tolerance for 5-fluorouracil and
chemotherapy failure (29). Serum
and glucocorticoid-regulated kinase isoform 1, extracellular
signal-regulated kinase, inhibitor of nuclear factor-κB kinase, and
AMP-activated protein kinase have also been described as negative
regulators of FOXO1 (30,31). Together, this suggests that FOXO1
is a convergence target for several signaling pathways and has an
opposing role in tumorigenesis (32).
TFP is used as a neuroleptic for controlling
psychotic disturbances, but it has also been reported to exert
anticancer effects on several cancer cells. Studies have revealed
that calmodulin antagonists can interfere with
Ca2+-calmodulin interactions, prevent
Ca2+-dependent cellular events and thereby limiting
tumor growth (33). As a
calmodulin antagonist, TFP also induces apoptosis and inhibits
tumorigenesis, proliferation and metastasis in several tumor types
(12,15). Furthermore, TFP is responsible for
inactivating the dopamine receptor D2, and inhibiting angiogenesis
and invasion (34). It has also
been reported that TFP is able to prevent the phosphorylation and
activation of Akt, which is an important molecular mechanism for
oncotherapy. The inactive Akt enhances the nuclear translocation of
FOXO1 and prevents tumor progression (34). Therefore, it was hypothesized that
TFP is capable of inhibiting gliomas by modulating FOXO1. Data have
suggested that TFP promotes the tumor inhibitory activity of FOXO1
(20). In the present study, it
was also shown that TFP increased nuclear FOXO1 protein, decreased
the levels of MDR genes, suppressed the efflux of DOX from glioma
cells and restricted glioma growth. These results are consistent
with those of other studies, which have confirmed that TFP may be
useful as a chemotherapy adjuvant for gliomas and for reversing ATP
binding cassette transporter-relevant MDR (13). The present study also found that
the levels of breast cancer resistance protein (BCRP) in SHG44/DOX
cells were not increased as much as the P-gp, MRP1 and LRP proteins
(data not shown). The reason for this may involve the upregulation
of P-gp in SHG44/DOX to result in the inhibition of BCRP. However,
the levels of BCRP may have been increased when P-gp was inhibited.
Therefore, it is important to emphasize that chemotherapy failure
may occur if only P-gp levels are reduced, leading to BCRP being
restored and becoming a candidate MDR mechanism.
In conclusion, FOXO1 appears to be a novel
therapeutic target for MDR. TFP is conducive in regulating FOXO1
and downstream drug resistance genes, and restoring DOX-induced
cytotoxicity. The present study provides the experimental basis for
a clinical application of TFP to reverse glioma chemotherapy
resistance and improve the efficacy of chemotherapeutic drugs for
this disease. These results may assist in the development of a
novel strategy for molecular cancer therapy for glioma.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors’ contributions
YC designed the study. XL cultured the SHG44/DOX
cell line and completed the xenograft tumor model. XC performed the
in vitro and in vivo experiments. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Ethics
Committee of Chongqing Medical University (Chonqing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Sontheimer H: Brain cancer: Tumour cells
on neighbourhood watch. Nature. 528:49–50. 2015.PubMed/NCBI
|
|
2
|
Geng F, Cheng X, Wu X, Yoo JY, Cheng C,
Guo JY, Mo X, Ru P, Hurwitz B, Kim SH, et al: Inhibition of SOAT1
suppresses glioblastoma growth via blocking SREBP-1-mediated
lipogenesis. Clin Cancer Res. 22:5337–5348. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stupp R, Brada M, van den Bent MJ, Tonn JC
and Pentheroudakis G; ESMO Guidelines Working Group: High-grade
glioma: ESMO clinical practice guidelines for diagnosis, treatment
and follow-up. Ann Oncol. 25:iii93–iii101. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xu Y, Zhao S, Cui M and Wang Q:
Down-regulation of microRNA-135b inhibited growth of cervical
cancer cells by targeting FOXO1. Int J Clin Exp Pathol.
8:10294–10304. 2015.PubMed/NCBI
|
|
5
|
Yamagata K, Daitoku H, Takahashi Y, Namiki
K, Hisatake K, Kako K, Mukai H, Kasuya Y and Fukamizu A: Arginine
methylation of FOXO transcription factors inhibits their
phosphorylation by Akt. Mol Cell. 32:221–231. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Coomans de Brachène A and Demoulin JB:
FOXO transcription factors in cancer development and therapy. Cell
Mol Life Sci. 73:1159–1172. 2016. View Article : Google Scholar
|
|
7
|
Song HM, Song JL, Li DF, Hua KY, Zhao BK
and Fang L: Inhibition of FOXO1 by small interfering RNA enhances
proliferation and inhibits apoptosis of papillary thyroid carcinoma
cells via Akt/FOXO1/Bim pathway. Onco Targets Ther. 8:3565–3573.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li CF, Zhang WG, Liu M, Qiu LW, Chen XF,
Lv L and Mei ZC: Aquaporin 9 inhibits hepatocellular carcinoma
through up-regulating FOXO1 expression. Oncotarget. 7:44161–44170.
2016.PubMed/NCBI
|
|
9
|
Cheng C, Jiao JT, Qian Y, Guo XY, Huang J,
Dai MC, Zhang L, Ding XP, Zong D and Shao JF: Curcumin induces G2/M
arrest and triggers apoptosis via FoxO1 signaling in U87 human
glioma cells. Mol Med Rep. 13:3763–3770. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shen H, Wang D, Li L, Yang S, Chen X, Zhou
S, Zhong S, Zhao J and Tang J: MiR-222 promotes drug-resistance of
breast cancer cells to adriamycin via modulation of PTEN/Akt/FOXO1
pathway. Gene. 596:110–118. 2017. View Article : Google Scholar
|
|
11
|
Arrigoni E, Galimberti S, Petrini M,
Danesi R and Di Paolo A: ATP-binding cassette transmembrane
transporters and their epigenetic control in cancer: An overview.
Expert Opin Drug Metab Toxicol. 12:1419–1432. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen QY, Wu LJ, Wu YQ, Lu GH, Jiang ZY,
Zhan JW, Jie Y and Zhou JY: Molecular mechanism of trifluoperazine
induces apoptosis in human A549 lung adenocarcinoma cell lines. Mol
Med Rep. 2:811–817. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shin SY, Choi BH, Kim JR, Kim JH and Lee
YH: Suppression of P-glycoprotein expression by antipsychotics
trifluoperazine in adriamycin-resistant L1210 mouse leukemia cells.
Eur J Pharm Sci. 28:300–306. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Polischouk AG, Holgersson A, Zong D,
Stenerlöw B, Karlsson HL, Möller L, Viktorsson K and Lewensohn R:
The antipsychotic drug trifluoperazine inhibits DNA repair and
sensitizes non small cell lung carcinoma cells to DNA double-strand
break induced cell death. Mol Cancer Ther. 6:2303–2309. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yeh CT, Wu AT, Chang PM, Chen KY, Yang CN,
Yang SC, Ho CC, Chen CC, Kuo YL, Lee PY, et al: Trifluoperazine, an
antipsychotic agent, inhibits cancer stem cell growth and overcomes
drug resistance of lung cancer. Am J Respir Crit Care Med.
186:1180–1188. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen J, Xu ZY and Wang F: Association
between DNA methylation and multidrug resistance in human glioma
SHG-44 cells. Mol Med Rep. 11:43–52. 2015. View Article : Google Scholar
|
|
17
|
Wen K, Fu Z, Wu X, Feng J, Chen W and Qian
J: Oct-4 is required for an antiapoptotic behavior of
chemoresistant colorectal cancer cells enriched for cancer stem
cells: effects associated with STAT3/Survivin. Cancer Lett.
333:56–65. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang N, Cheng S, Mi X, Tian Q, Huang Q,
Wang F, Xu Z, Xie Z, Chen J and Cheng Y: Downregulation of nitrogen
permease regulator like-2 activates PDK1-AKT1 and contributes to
the malignant growth of glioma cells. Mol Carcinog. 55:1613–1626.
2016. View
Article : Google Scholar
|
|
19
|
Chen S, Zhao H, Deng J, Liao P, Xu Z and
Cheng Y: Comparative proteomics of glioma stem cells and
differentiated tumor cells identifies S100A9 as a potential
therapeutic target. J Cell Biochem. 114:2795–2808. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sangodkar J, Dhawan NS, Melville H, Singh
VJ, Yuan E, Rana H, Izadmehr S, Farrington C, Mazhar S, Katz S, et
al: Targeting the FOXO1/KLF6 axis regulates EGFR signaling and
treatment response. J Clin Invest. 122:2637–2651. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao Z, Qin L and Li S: miR-411
contributes the cell proliferation of lung cancer by targeting
FOXO1. Tumour Biol. 37:5551–5560. 2016. View Article : Google Scholar
|
|
22
|
Ding H, Zhu Y, Chu T and Wang S: Epidermal
growth factor induces FoxO1 nuclear exclusion to activate
MMP7-mediated metastasis of larynx carcinoma. Tumour Biol.
35:9987–9992. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pei J, Lou Y, Zhong R and Han B: MMP9
activation triggered by epidermal growth factor induced FoxO1
nuclear exclusion in non-small cell lung cancer. Tumour Bio.
35:6673–6678. 2014. View Article : Google Scholar
|
|
24
|
Myatt SS, Wang J, Monteiro LJ, Christian
M, Ho KK, Fusi L, Dina RE, Brosens JJ, Ghaem-Maghami S and Lam EW:
Definition of microRNAs that repress expression of the tumor
suppressor gene FOXO1 in endometrial cancer. Cancer Res.
70:367–377. 2010. View Article : Google Scholar
|
|
25
|
Hou T, Ou J, Zhao X, Huang X, Huang Y and
Zhang Y: MicroRNA-196a promotes cervical cancer proliferation
through the regulation of FOXO1 and p27Kip1. Br J Cancer.
110:1260–1268. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Magro G, Righi A, Casorzo L, Antonietta T,
Salvatorelli L, Kacerovská D, Kazakov D and Michal M: Mammary and
vaginal myofibroblastomas are genetically related lesions:
Fluorescence in situ hybridization analysis shows deletion of 13q14
region. Hum Pathol. 43:1187–1193. 2012. View Article : Google Scholar
|
|
27
|
Zhang X, Tang N, Hadden TJ and Rishi AK:
Akt, FoxO and regulation of apoptosis. Biochim Biophys Acta.
1813.1978–1986. 2011.
|
|
28
|
Lian R, Lu B, Jiao L, Li S, Wang H, Miao W
and Yu W: MiR-132 plays an oncogenic role in laryngeal squamous
cell carcinoma by targeting FOXO1 and activating the PI3K/AKT
pathway. Eur J Pharmacol. 792:1–6. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Altan B, Yokobori T, Ide M, Mochiki E,
Toyomasu Y, Kogure N, Kimura A, Hara K, Bai T, Bao P, et al:
Nuclear PRMT1 expression is associated with poor prognosis and
chemosensitivity in gastric cancer patients. Gastric Cancer.
19:789–797. 2016. View Article : Google Scholar
|
|
30
|
Van Der Heide LP, Hoekman MF and Smidt MP:
The ins and outs of FoxO shuttling: Mechanisms of FoxO
translocation and transcriptional regulation. Biochem J.
380:297–309. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zou J, Hong L, Luo C, Li Z, Zhu Y, Huang
T, Zhang Y, Yuan H, Hu Y, Wen T, et al: Metformin inhibits
estrogen-dependent endometrial cancer cell growth by activating the
AMPK-FOXO1 signal pathway. Cancer Sci. 107:1806–1817. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sun F, Han DF, Cao BQ, Wang B, Dong N and
Jiang DH: Caffeine-induced nuclear translocation of FoxO1 triggers
Bim-mediated apoptosis in human glioblastoma cells. Tumour Biol.
37:3417–3423. 2016. View Article : Google Scholar
|
|
33
|
Yuan K, Yong S, Xu F, Zhou T, McDonald JM
and Chen Y: Calmodulin antagonists promote TRA-8 therapy of
resistant pancreatic cancer. Oncotarget. 6:25308–25319. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pulkoski-Gross A, Li J, Zheng C, Li Y,
Ouyang N, Rigas B, Zucker S and Cao J: Repurposing the
antipsychotic trifluoperazine as an antimetastasis agent. Mol
Pharmacol. 87:501–512. 2015. View Article : Google Scholar : PubMed/NCBI
|