Introduction
Obesity is a serious public health epidemic, with
~2,000,000,000 adults being overweight or obese worldwide, and is
considered a major contributor to hyperlipidemia, cardiovascular
disease, type 2 diabetes, and cancer (1). As a result, obesity is now among the
leading contributors to global morbidity and mortality rates
(2). Although the development of
obesity is influenced by a number of factors, including excessive
food intake, lack of physical activity, genetic susceptibility,
medications, and endocrine abnormalities (2,3),
increasing evidence indicates that it also arises from excessive
preadipocyte differentiation in adipose tissue (4,5).
Therefore, any compound that inhibits the disproportionate
differentiation of these cells constitutes a potential therapeutic
option for obesity.
Preadipocyte differentiation, also known as
adipogenesis, is the process during which fibroblast-like
preadipocytes develop into mature adipocytes (5). This process is influenced by the
adipogenic program, which is controlled by adipogenic transcription
factors, including CCAAT/enhancer-binding proteins (C/EBPs),
peroxisome proliferator-activated receptors (PPARs), and signal
transducers and activators of transcription (STATs) (6-10),
lipogenic enzymes, including fatty acid synthase (FAS) and acetyl
CoA carboxylase (ACC) (11,12), and lipid droplet (LD)-associated
proteins, including perilipin A (13,14). In addition, there is evidence that
adenosine 3',5'-cyclic mono-phosphate (cAMP), cAMP-activated
protein kinase (AMPK), protein kinase A (PKA), protein kinase C
family members, mitogen-activated protein kinase kinase-1/2, and
extracellular signal-regulated protein kinase-1/2, are known to
mediate preadipocyte differentiation (15-18). Furthermore, several studies have
shown that members of the Raf kinases, including c-Raf, A-Raf, and
B-Raf, are expressed and phosphorylated during preadipocyte
differentiation, and their expression and activities are crucial to
this process (19-21).
In our previous study, 85 known protein kinase
inhibitors with anticancer activities were screened using the
AdipoRed assay to assess the ability of each to lower lipid
(triglyceride, TG) content during the differentiation of 3T3-L1
preadipocytes into adipociytes. Several, including the pan-Raf
kinase inhibitor LY3009120 (22),
were found to have an anti-adipogenic effect. LY3009120 is known to
have anticancer activities (23,24); however, its anti-obesity effect
and its mode of action in adipocytes remain to be fully elucidated.
The present study investigated the effect of LY3009120 on
adipogenesis in 3T3-L1 preadipocytes. The results revealed for the
first time, to be the best of our knowledge, that LY3009120 had an
anti-adipogenic effect on differentiating 3T3-L1 cells, which
appeared to be mediated through modulation of the expression and/or
phosphorylation of C/EBP-α, PPAR-γ, STAT-3, FAS, ACC, perilipin A,
and AMPK.
Materials and methods
Materials
LY3009120 was purchased from Apexbio Technology
(Houston, TX, USA). Polyclonal C/EBP-α (cat. no. sc-61), monoclonal
PPAR-γ (cat. no. sc-7273), monoclonal STAT-3 (cat. no. sc-8019),
and monoclonal phosphorylated (p)-STAT-3 (cat. no. sc-8059)
antibodies were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). Monoclonal FAS (cat. no. 610962), monoclonal
β-actin (cat. no. A5441), and polyclonal perilipin A (cat. no.
3948-200) antibodies were purchased from BD Biosciences (San Jose,
CA, USA), Sigma, EMD Millipore (Billerica, MA, USA), and BioVision,
Inc. (Milpitas, CA, USA), respectively. Polyclonal p-AMPK (T172,
cat. no. 2535), monoclonal AMPK (cat. no. 2793), polyclonal p-ACC
(S79, cat. no. 3661), polyclonal ACC (cat. no. 3662), polyclonal
liver kinase B1 (LKB1; cat. no. 3047), polyclonal p-LKB1 (S428,
cat. no. 3482), polyclonal p-A-Raf (S299, cat. no. 4431),
polyclonal A-Raf (cat. no. 4432), polyclonal p-B-Raf (S445, cat.
no. 2696), polyclonal B-Raf (cat. no. 9433), polyclonal p-C-Raf
(S259, cat. no. 9421), and monoclonal C-Raf (cat. no. 12552)
antibodies were acquired from Cell Signaling Technology, Inc.
(Danvers, MA, USA). Monoclonal procas-pase-9 (cat. no.
ADI-AAM-139), polyclonal poly (ADP-ribose) polymerase (PARP; cat.
no. 11 835 238 001), and polyclonal death receptor 5 (DR5; cat. no.
NBP1-45951) antibodies were purchased from Enzo Life Sciences, Inc.
(Farmingdale, NY, USA), Roche Diagnostics (Basel, Switzerland), and
Novus Biologicals, LLC (Littleton, CO, USA), respectively.
Culture and differentiation of 3T3-L1
cells
Murine white preadipocytes of the 3T3-L1 line (ATCC,
Manassas, VA, USA) were cultured to the contact-inhibition stage
and maintained at 37°C and 5% CO2 in the post-confluent
stage for 2 days in DMEM supplemented with 10% fetal bovine serum
(FBS; Gibco, Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
penicillin-streptomycin (WelGENE, Inc., Daegu, Korea).
Differentiation was then induced in the presence or absence of
LY3009120 at the indicated concentrations (0.1, 0.2, 0.3, and 0.4
µM) by replacing the medium with DMEM supplemented with 10%
FBS (WelGENE, Inc.) and a hormone cocktail (MDI; Sigma, EMD
Millipore) comprising 0.5 mM IBMX (M), 0.5 µM dexamethasone
(D), and 5 µg/ml insulin (I). Following 48 h of exposure to
MDI, the differentiation medium was replaced with DMEM supplemented
with 10% FBS and 5 µg/ml insulin, with or without LY3009120
at the indicated concentrations. The cells were then provided every
other day with DMEM containing 10% FBS with or without LY3009120 at
the indicated concentrations until day 8, by which point, the
preadipocytes had become mature adipocytes with a rounded
morphology and filled with LDs.
Oil Red O staining
On day 8 of differentiation, the control and
LY3009120-treated 3T3-L1 cells were washed twice with PBS, fixed
with 10% formaldehyde for 2 h at room temperature (RT), washed with
60% isopropanol, and dried completely. The fixed cells were then
stained with Oil Red O working solution for 1 h at RT, and then
washed twice with distilled water. The LDs were observed by light
microscopy (Nikon Corporation, Tokyo, Japan).
Cell count analysis
The preadipocytes (3T3-L1 cells) were seeded in
24-well plates and cultured under the above-mentioned
differentiation conditions. On day 8 of differentiation, the
control and LY3009120-treated 3T3-L1 cells, which cannot be stained
with trypan blue dye, were counted under an Olympus phase contrast
microscope equipped with a digital camera (Nikon Corporation). The
assay was performed in triplicate, and the data are presented as
the mean ± standard error (SE) of three independent
experiments.
Quantification of intracellular TG
content by AdipoRed assay
On day 8 of differentiation, the lipid content of
control and LY3009120-treated 3T3-L1 was measured using the
commercially available AdipoRed Assay Reagent kit according to the
manufacturer's protocol (Lonza Group AG, Basel, Switzerland).
Following incubation for 10 min, fluorescence was measured on a
Victor3 plate reader (PerkinElmer, Inc., Waltham, MA,
USA) with excitation and emission wavelengths of 485 and 572 nm,
respectively.
Preparation of whole cell lysates
At the designated time point, the 3T3-L1 cells were
washed twice with PBS and exposed to a modified RIPA buffer [50
µM Tris-Cl (pH 7.4), 150 µM NaCl, 0.1% SDS, 0.25%
sodium deoxycholate, 1% Triton X-100, 1% Nonidet P-40, 1 mM EDTA, 1
mM EGTA, and 1X proteinase inhibitor cocktail]. The cell lysates
were then collected and centrifuged at 12,074 x g for 20 min at
4°C. The supernatants were retained and their protein
concentrations were determined with Bradford reagent (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Western blot analysis
The proteins (50 µg) were separated by
SDS-PAGE on 10% gels and transferred onto nitrocellulose membranes
(EMD Millipore). The membranes were washed with TBST [10 mM Tris,
and 150 mM NaCl supplemented with 0.05% (vol/vol) Tween 20] and
subsequently blocked with TBST containing 5% (wt/vol) non-fat dried
milk. The membranes were then incubated overnight with antibodies
specific to C/EBP-α (1:1,000), PPAR-γ (1:1,000), STAT-3 (1:1,000),
p-STAT-3 (1:1,000), FAS (1:1,000), perilipin A (1:2,000), p-AMPK
(1:1,000), AMPK (1:1,000), p-ACC (1:2,000), ACC (1:2,000),
p-LKB1(1:1,000), LKB1 (1:1,000), p-A-Raf (1:1,000), A-Raf
(1:1,000), p-B-Raf (1:1,000), B-Raf (1:1,000), p-C-Raf (1:1,000),
C-Raf (1:1,000), procaspase-9 (1:1,000), PARP (1:5,000), DR5
(1:1,000) or β-actin (1:10,000) at 4°C. The membranes were then
exposed to horseradish peroxidase-conjugated secondary antibodies
(an goat anti-rabbit IgG (H+L) (1:2,000), catalog no. 111-035-045
or goat anti-mouse IgG (H+L) (1:2,000), catalog no. 115-035-062,
Jackson ImmunoResearch, West Grove, PA, USA) for 2 h at RT, and
were then washed three times with TBST at RT. The immunoreactivity
was detected using enhanced chemiluminescence reagents, and
variations in protein loading were assessed by observing actin
protein levels using ImageJ software (ImageJ version 1.8.0;
National Institutes of Health, Bethesda, Maryland, USA).
Statistical analysis
The cell count analysis was performed in triplicate
and repeated three times. Data are expressed as the mean ± SE.
Differences were analyzed by one-way analysis of variance (SPSS
software, version 11.5; SPSS, Inc., Chicago, IL, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
LY3009120 inhibits 3T3-L1 cell
adipogenesis
The present study first investigated the effect of
LY3009120 at different concentrations (0.1, 0.2, 0.3, and 0.4
µM) on lipid accumulation in differentiating 3T3-L1 cells
using the Oil Red O staining. The protocol followed to induce
3T3-L1 preadipocyte differentiation is shown in Fig. 1A. Treatment of these preadipocytes
with LY3009120 for 8 days decreased the number of LDs in the
corresponding differentiated adipocytes in a
concentration-dependent manner (Fig.
1B, upper panels). This LY3009120-mediated inhibition of LD
accumulation was also observed in differentiating 3T3-L1 cells by
light microscopy on days 2, 5 and 8 (Fig. 1B, lower panels). Subsequently,
whether exposure of the differentiating 3T3-L1 cells to LY3009120
for 8 days affects their TG content was determined using the
AdipoRed assay. Although cellular TG content was marginally
enhanced by treatment with 0.1 µM LY3009120, it was
effectively reduced by administration of this drug at 0.2, 0.3, or
0.4 µM in a concentration-dependent manner (Fig. 1C). A cell counting assay was used
to determine whether treatment with LY3009120 for 8 days at the
concentrations assessed was cytotoxic to the differentiating 3T3-L1
cells. As shown in Fig. 1D,
3T3-L1 cell survival was marginally reduced by 0.1, 0.2, or 0.3
µM, and markedly decreased following exposure to this drug
at 0.4 µM (Fig. 1D),
indicating that the latter concentration was cytotoxic to
differentiating 3T3-L1 cells. In addition, whether 8 days of
LY3009120 administration at the concentrations assessed induces the
apoptosis of differentiating 3T3-L1 cells was analyzed by measuring
the levels of procaspase-9, PARP, and DR5, which are known
apoptotic proteins. The results of the western blotting analysis
revealed that, at the concentrations used, LY3009120 did not
markedly affect the expression of procaspase-9, PARP, or DR5. As
0.3 µM of LY3009120 markedly reduced lipid accumulation and
TG content with minimal cytotoxicity, this concentration was
selected for further experiments.
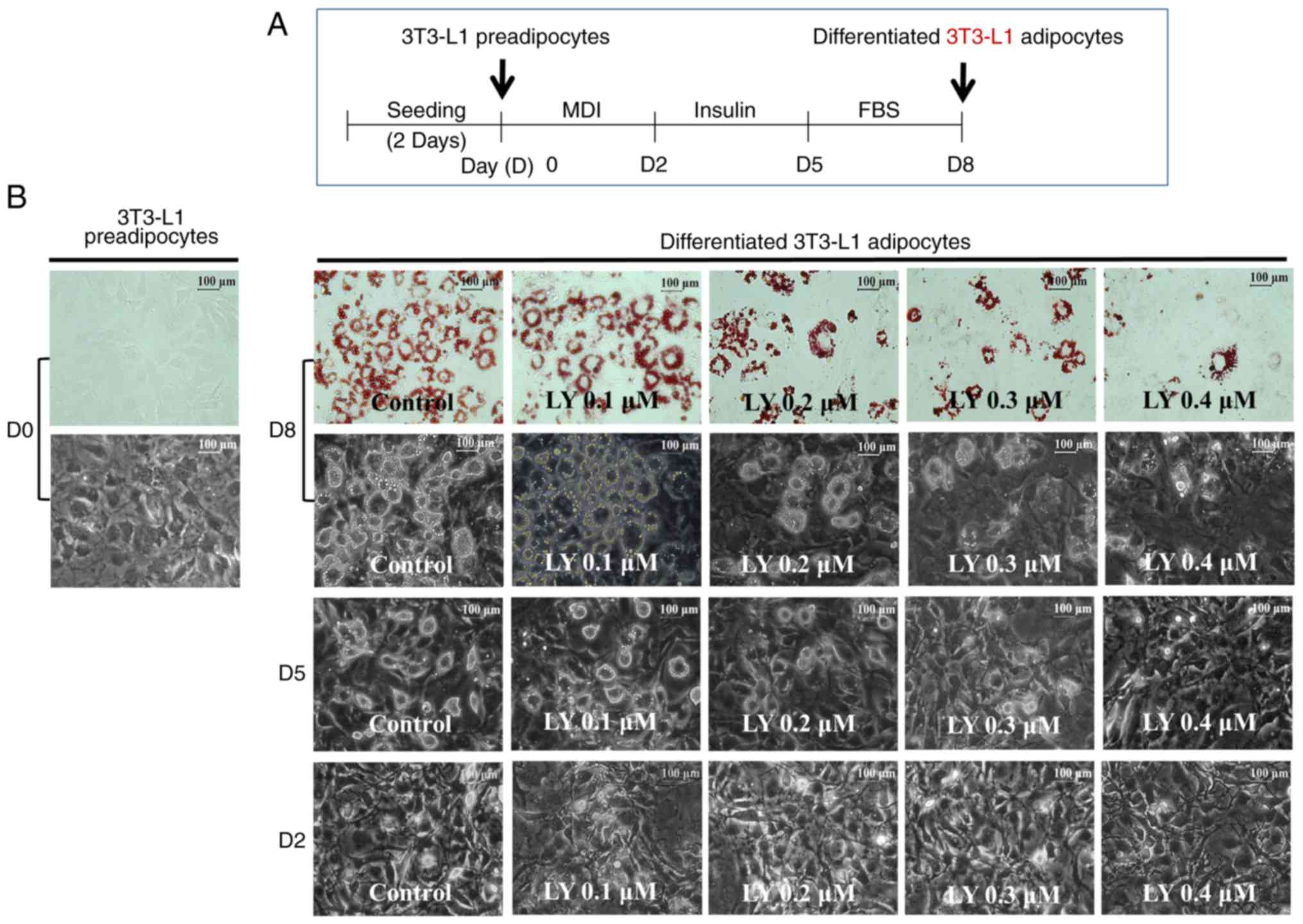 | Figure 1Effects of LY3009120 on the
adipogenesis and growth of differentiating 3T3-L1 cells. (A)
Protocol used to induce 3T3-L1 preadipocyte differentiation. 3T3-L1
preadipocytes were induced to differentiate with induction medium
in the presence or absence of LY3009120 at the indicated
concentrations and for the indicated times. (B) On D8, cellular
lipid content was assessed by Oil Red O staining (upper panels).
Phase-contrast images of the cells were also recorded following
treatment (lower panels). (C) On D8, cellular TG content was
quantified using the AdipoRed assay. Values are presented as the
mean ± standard error of the mean of data from three independent
experiments with three replicates. *P<0.05, vs.
control. (D) On D8, LY3009120-treated 3T3-L1 cells, which are not
stained by trypan blue dye, were counted under a microscope. The
cell count assay was performed in triplicate. Data are presented as
the mean ± standard error of the mean of data from three
independent experiments. *P<0.05, vs. control. (E)
3T3-L1 preadipocytes were induced to differentiate with induction
medium in the presence or absence of LY3009120 at the indicated
concentrations for 8 days. Cellular proteins were extracted and
analyzed by western blot analysis. LY, LY3009120; MDI, IBMX,
dexamethasone, and insulin; FBS, fetal bovine serum; D, day; TG,
triglyceride; PARP, polyclonal poly (ADP-ribose) polymerase; DR5,
death receptor 5. |
LY3009120 does not substantially alter
the expression and phosphorylation of Raf kinases in
differentiating 3T3-L1 cells
As LY3009120 targets Raf kinases, western blotting
analysis was performed to assess whether C-Raf, A-Raf, and B-Raf,
members of this protein kinase family, are expressed and
phosphorylated in differentiating 3T3-L1 cells and whether this is
affected by LY3009120. Substantial expression and phosphorylation
of C-Raf, A-Raf, and B-Raf was observed in these cells (Fig. 2A); however, LY3009120 treatment
had no notable effect on the expression and phosphorylation of
these Raf kinases. Densitometry data obtained from Fig. 2A are shown in Fig. 2B.
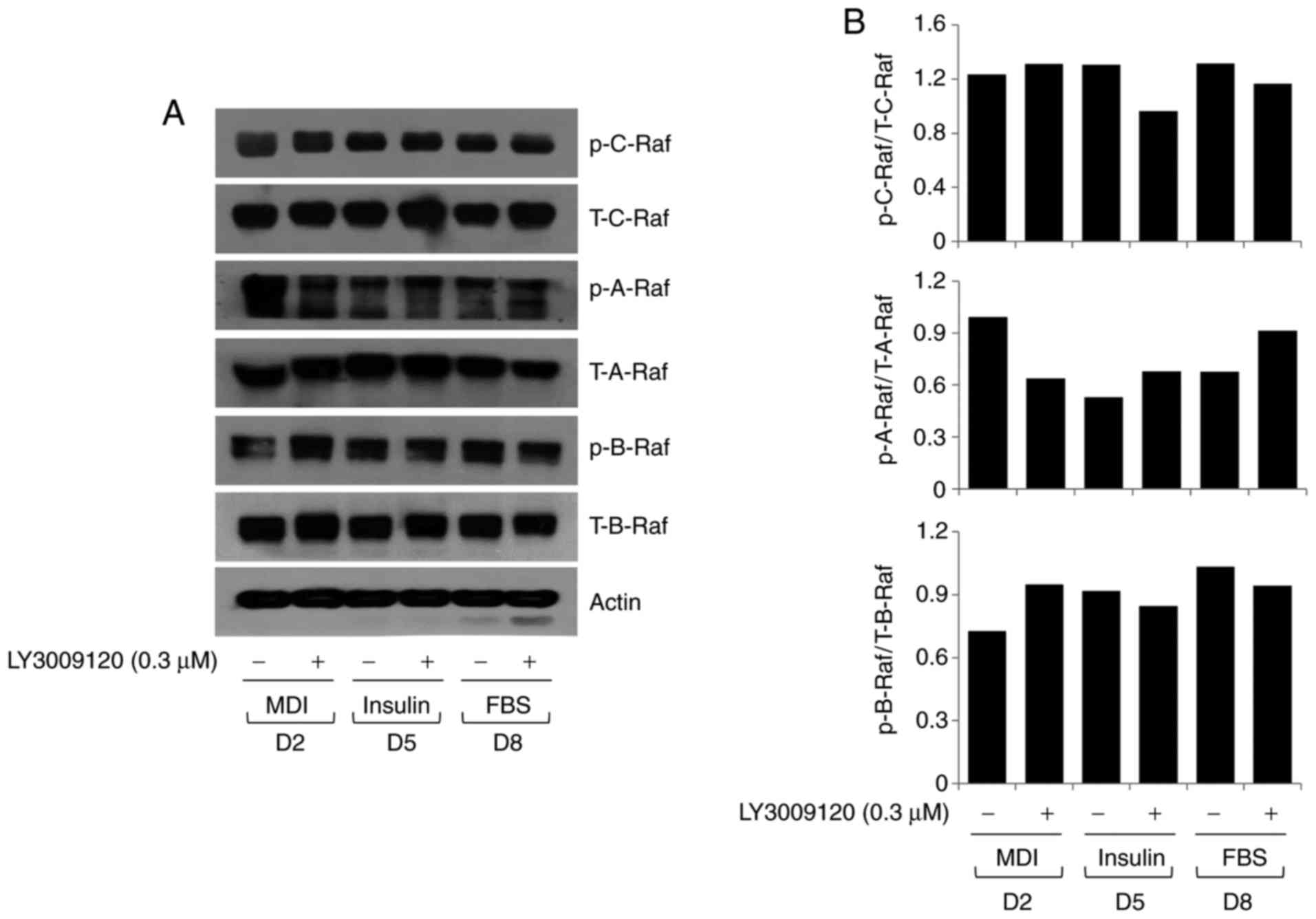 | Figure 2Effect of LY3009120 on the expression
and/or phosphorylation of three Raf kinases in differentiating
3T3-L1 cells. (A) 3T3-L1 preadipocytes were induced to
differentiate with induction medium in the presence or absence of
LY3009120, and harvested on D2, 5, and 8. Proteins were extracted
from cells at the indicated time points and analyzed by western
blot analysis. (B) Densitometry data from triplicate experiments
showing levels of p-C-Raf, p-A-Raf, and p-B-Raf relative to those
of T-C-Raf, T-A-Raf, and T-B-Raf, respectively, on D2, 5 and 8.
MDI, IBMX, dexamethasone, and insulin; FBS, fetal bovine serum;
p-C-Raf, phosphorylated C-Raf; T-C-Raf, total C-Raf; p-A-Raf,
phosphorylated A-Raf; T-A-Raf, total A-Raf; p-B-Raf, phosphorylated
B-Raf; T-B-Raf, total B-Raf; D, day. |
LY3009120 reduces the expression and/or
phosphorylation of C/EBP-α, PPAR-γ, and STAT-3 in differentiating
3T3-L1 cells
To obtain a better understanding of the mechanisms
associated with the anti-adipogenic effect of LY3009120, the
present study aimed to determine whether this drug modulates the
expression and/or phosphorylation (activation) of C/EBP-α, PPAR-γ,
and STAT-3 in differentiating 3T3-L1 cells. As shown in Fig. 3A, LY3009120 treatment considerably
lowered the levels of C/EBP-α and PPAR-γ in the cells. Furthermore,
it reduced the phosphorylation of STAT-3 without affecting the
expression of total STAT-3 (Fig.
3B). Densitometry data from the experiments performed in
triplicate confirmed that LY3009120 reduced the expression of
C/EBP-α and PPAR-γ (Fig. 3C) and
phosphorylation of STAT-3 (Fig.
3D) during 3T3-L1 preadipocyte differentiation.
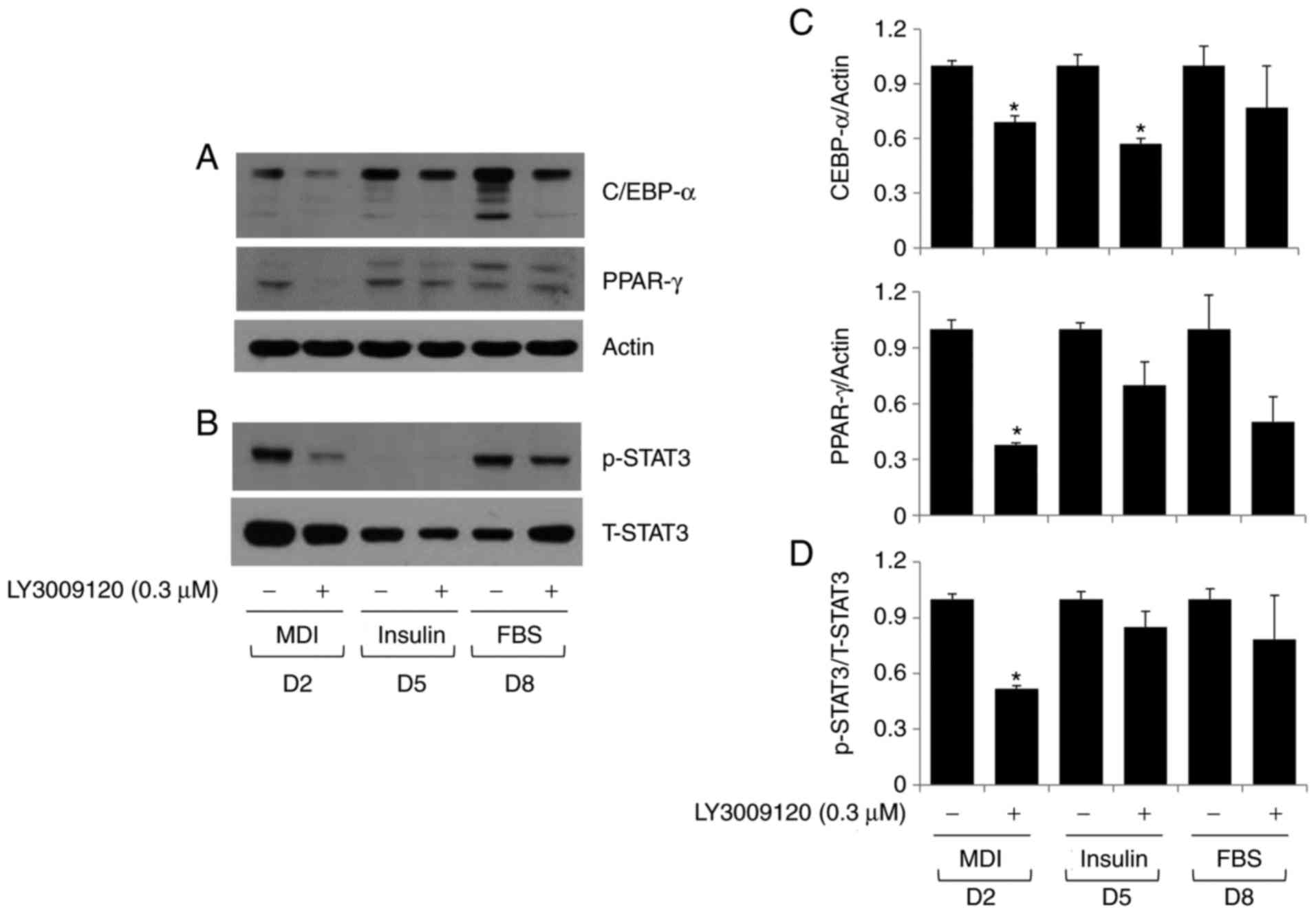 | Figure 3Effect of LY3009120 on the expression
and/or phosphorylation of C/EBP-α, PPAR-γ, and STAT-3 in
differentiating 3T3-L1 cells. 3T3-L1 preadi-pocytes were induced to
differentiate with induction medium in the presence or absence of
LY3009120, and harvested on D2, 5, and 8. Proteins were extracted
from cells collected at the indicated time points and analyzed by
western blot analysis for (A) C/EBP-α and PPAR-γ or (B) p-STAT-3
and T-STAT-3. Densitometry data from the western blot experiments
for (C) C/EBP-α and PPAR-γ and (D) p-STAT-3/T-STAT3, which were
performed in triplicate on cells collected on D2, 5, and 8. Data
are expressed as the mean ± standard error. *P<0.05
vs. control at the indicated day. MDI, IBMX, dexamethasone, and
insulin; FBS, fetal bovine serum; C/EBP-α, CCAAT/enhancer-binding
protein-α; PPAR-γ, peroxisome proliferator-activated receptor-γ;
p-STAT-3, phosphorylated STAT-3; T-STAT-3, total STAT3; D, day. |
LY3009120 alters the expression and/or
phosphorylation of FAS, perilipin A, AMPK, ACC, and LKB1 in
differentiating 3T3-L1 cells
The effects of LY3009120 on the expression of FAS
and perilipin A in differentiating 3T3-L1 cells were also examined.
As shown in Fig. 4A, LY3009120
treatment led to a decrease in the levels of these proteins in the
cells. Subsequently, the effect of LY3009120 on the expression
and/or phosphorylation of AMPK and ACC proteins in 3T3-L1 cells
during differentiation was investigated. LY3009120 markedly
increased the phosphorylation of AMPK but decreased that of ACC.
LY3009120 did not affect the levels of total AMPK protein, but
marginally reduced the levels of total ACC protein. Whether
LY3009120 alters the expression and/or phosphorylation of LKB1, the
main kinase responsible for AMPK phosphorylation (24), was also determined in the
differentiating 3T3-L1 cells. The levels of total LKB1 and the
phosphorylation of this protein on days 2 and 5 of 3T3-L1
preadipocyte differentiation were not affected by LY3009120, but
were considerably reduced on day 8 due to administration of this
drug. Densitometry data from experiments performed in triplicate
confirmed the ability of LY3009120 to increase the phosphorylation
of AMPK and decrease the phosphorylation and/or total levels of
FAS, perilipin A, ACC, and LKB1 during 3T3-L1 preadipocyte
differentiation (Fig. 4B).
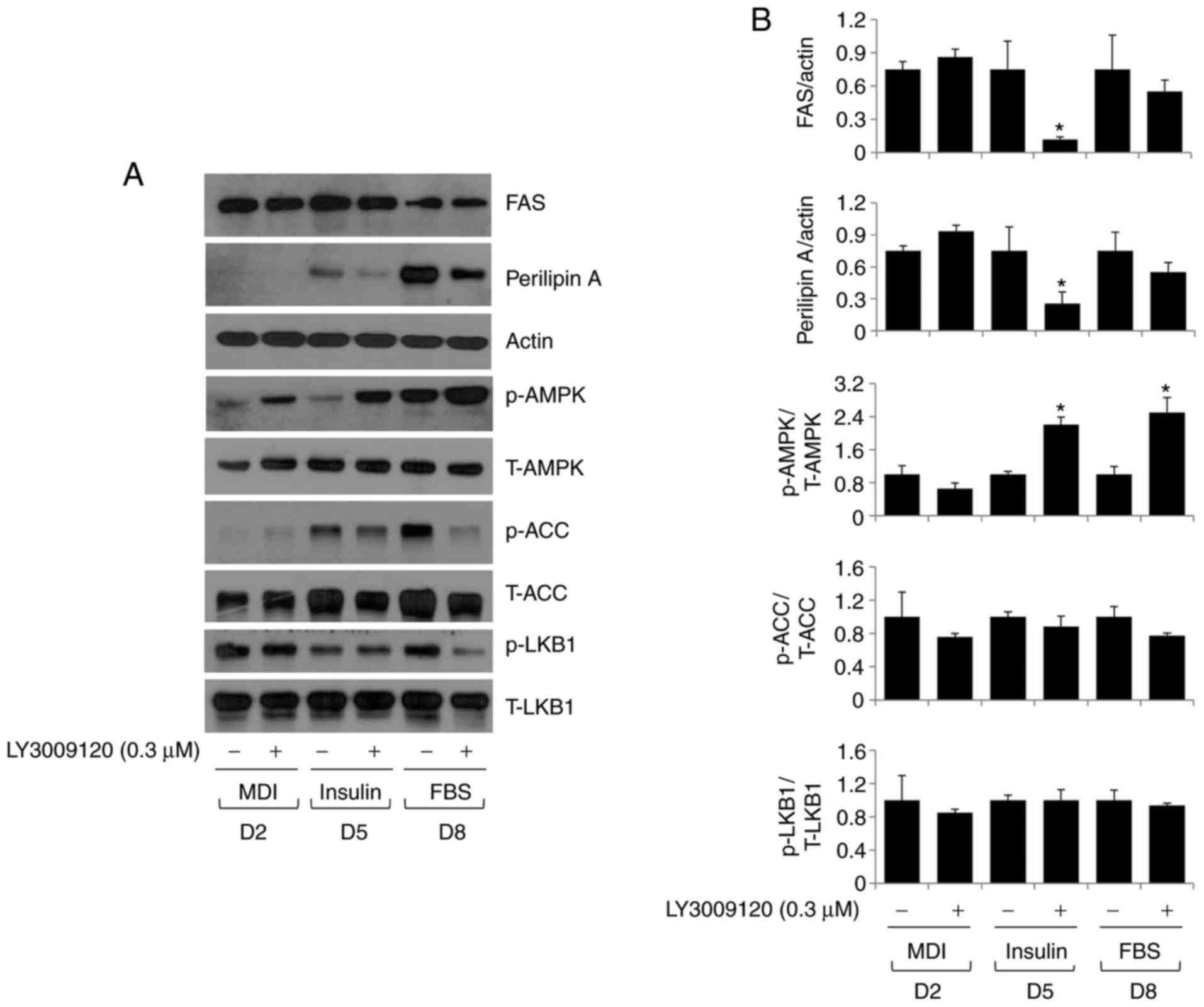 | Figure 4Effect of LY3009120 on the expression
and/or phosphorylation of FAS, perilipin A, AMPK, ACC, and LKB1 in
differentiating 3T3-L1 cells. (A) 3T3-L1 preadipocytes were induced
to differentiate with induction medium in the presence or absence
of LY3009120, and harvested on D2, 5, and 8. Proteins were
extracted from cells collected at the indicated time points and
analyzed by western blot analysis. (B) Densitometry data from
western blot experiments performed in triplicate, showing
expression levels of FAS, perilipin A, p-/T-AMPK, p-/T-ACC, and
p-/T-LKB1 on D2, 5, and 8. Data are expressed as the mean ±
standard error. *P<0.05 vs. control at the indicated
day. MDI, IBMX, dexamethasone, and insulin; FAS, fatty acid
synthase; AMPK, adenosine 3',5'-cyclic monophosphate-activated
protein kinase; ACC, acetyl CoA carboxylase; LKB1; liver kinase B1;
p-, phosphorylated; T-, total; D, day. |
Discussion
Preadipocyte differentiation/adipogenesis, excessive
levels of which are correlated with the development of obesity, is
known to be mediated by Raf kinases. LY3009120 is a pan-Raf kinase
inhibitor and demonstrates anticancer activity. In order to
establish whether LY3009120 may also serve as an alternative to
currently available anti-obesity medications, the present study
investigated its effect on adipogenesis in differentiating 3T3-L1
cells. It was demonstrated that LY3009120 had an anti-adipogenic
effect on these cells by controlling the expression and
phosphorylation of C/EBP-α, PPAR-γ, STAT-3, FAS, ACC, perilipin A,
and AMPK.
It has previously been shown that C-Raf kinase is
phosphorylated in insulin-treated 3T3-L1 cells, and its
hyper-phosphorylation is crucial for hormone-induced 3T3-L1 cell
differentiation (19). The
present study revealed that the three Raf kinase isoforms, C-Raf,
A-Raf, and B-Raf, were expressed and phosphorylated in
differentiating 3T3-L1 cells. Notably, at the concentrations
assessed, LY3009120 did not appreciably influence the expression
and phosphorylation of these Raf kinases, but markedly reduced
lipid accumulation and TG content in the differentiating 3T3-L1
cells, particularly when administered at a concentration of 0.3
µM, with no cytotoxic or apoptotic effects. These results
suggested that LY3009120 exerts a potent anti-adipogenic effect
through a Raf-independent mechanism. At present, the mechanism
underlying the restriction of adipo-genesis in differentiating
3T3-L1 cells by LY3009120 at 0.3 µM in the absence of
pan-Raf kinase inhibition remains to be fully elucidated. It has
been shown that LY03009120 inhibits A-Raf, B-Raf, and C-Raf kinase
in A375 melanoma cells, with IC50 values of 44, 31-47,
and 42 nM, respectively (22). It
is suggested that the inhibitory effect of LY3009120 on Raf kinases
(and its inhibitory concentration) differs between cell types;
therefore, a concentration >0.3 µM may be required to
inhibit all Raf kinases in differentiating 3T3-L1 cells. However,
the administration of LY3009120 at a concentration >0.3
µM (0.4 µM in the present study) was cytotoxic to
these cells. Taken together, 0.3 µM LY3009120 may be used to
induce a Raf-independent anti-adipogenic effect with no
cytotoxicity.
It is well established that the expression and
activities of C/EBP-α, PPAR-γ, and STAT-3/5 are critical for 3T3-L1
cell adipogenesis (6-10,25,26). In the present study, LY3009120
decreased not only the levels of C/EBP-α and PPAR-γ, but also the
phosphorylation of STAT-3 in differentiating 3T3-L1 cells.
Therefore, it is likely that the anti-adipogenic effect of
LY3009120 may be linked to the reduced expression/phosphorylation
of these adipogenic transcription factors. Adipocyte-specific
proteins, including FAS and perilipin A, are also involved in
adipocyte differentiation. It has been reported that FAS is a
lipogenic enzyme involved in fatty acid synthesis (11), and perilipin A is an LD-associated
protein that binds and stabilizes newly formed LDs during the
differentiation of adipocytes (13,14,27). As LY3009120 was shown to lower
levels of FAS and perilipin A in differentiating 3T3-L1 cells, it
is possible that the attenuation of lipid accumulation and TG
content induced by this drug is attributable to the downregulation
of these proteins.
Another observation in the present study was the
regulation of AMPK phosphorylation at T172, indicating activation,
by LY3009120 in differentiating 3T3-L1 cells. AMPK is a key
regulator of fat and energy metabolism (28,29), and notably, there have been
several reports indicating that its activation inhibits
adipogenesis (30-32). There is also evidence that the
activation of AMPK induces the phosphorylation (at S79) of its
downstream effector ACC, which lacks the enzymatic activity
required to synthesize fatty acids (12). ACC is a multi-subunit lipogenic
enzyme that catalyzes the irreversible carboxylation of acetyl-CoA
to produce malonyl-CoA for the biosynthesis of fatty acids
(33). In addition, there is
accumulating evidence that the activation of AMPK inhibits
ATP-consuming anabolic processes but activates ATP-producing
catabolic processes (34), in
part via the phosphorylation of ACC (12). In the present study, LY3009120
demonstrated the ability to increase the phosphorylation of AMPK
and decrease the phosphorylation and expression of ACC in
differentiating 3T3-L1 cells. These results suggested that the
anti-adipogenic and lipid-lowering effects of this drug are further
mediated through the activation of AMPK and downregulation of ACC,
which may lead to the inhibition of ATP-consuming anabolic
processes, including fatty acid synthesis. In addition, its
suppressive effect on the phosphorylation and expression of ACC is
unlikely to be AMPK-dependent. LKB1 is the kinase principally
responsible for AMPK phosphorylation (24,35,36), and a previous study demonstrated
the regulation of AMPK by LKB1 in adipocytes (37). However, the present study showed
that the phosphorylation and expression of LKB1 in differentiating
3T3-L1 cells were not markedly altered by LY3009120, being only
marginally decreased on days 2 and 8 of differentiation. This
indicated that promotion of the phosphorylation of AMPK by
LY3009120 occurs independently of LKB1. The phosphorylation of AMPK
is also controlled by other kinases, including CAMKK2, also known
as CAMKKb (38,39), and other mechanisms entirely,
including changes in the intracellular AMP/ATP ratio (40). Therefore, future investigations
are required to examine whether LY3009120 alters intracellular ATP
levels (and the AMP/ATP ratio) and/or the expression and activity
of CAMKK2 in differentiating 3T3-L1 cells, which is likely to
improve current understanding of the activation of AMPK by this
pan-Raf kinase inhibitor.
In conclusion, the present study is the first, to
the best of our knowledge, to show that LY3009120 has an
anti-adipogenic effect on 3T3-L1 cells, which may be mediated via
control of the expression and/or phosphorylation of C/EBP-α,
PPAR-γ, STAT-3, FAS, ACC, perilipin A, and AMPK. Although important
issues remain to be elucidated, for example, whether this effect
can be reproduced in animal models of obesity, the present findings
reveal that LY3009120 demonstrates potential as a treatment for
obesity.
Acknowledgements
Not applicable.
Funding
This study was supported by the Yeungnam University
Research Grant of 2016 (grant no. 215A480003).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SMY and YKP performed experiments. TYL, BCJ, JK, and
YHL designed the work and analysed the data. TYL and BCJ wrote the
paper. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
ACC
|
acetyl CoA carboxylase
|
|
AMP
|
adenosine monophosphate
|
|
AMPK
|
cAMP-activated protein kinase
|
|
ATP
|
adenosine triphosphate
|
|
CAMKK2
|
calcium-/calmodulin-dependent kinase
kinase 2
|
|
cAMP
|
adenosine 3',5'-cyclic
monophosphate
|
|
C/EBP-α
|
CCAAT/enhancer-binding protein-α
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
DR5
|
death receptor 5
|
|
FAS
|
fatty acid synthase
|
|
FBS
|
fetal bovine serum
|
|
LD
|
lipid droplet
|
|
LKB1
|
liver kinase B1
|
|
MDI
|
IBMX, dexamethasone, and insulin
|
|
MEK-1/2
|
mitogen-activated protein kinase
kinase-1/2
|
|
PKA
|
protein kinase A
|
|
PPAR-γ
|
peroxisome proliferator-activated
receptor-γ
|
|
RT
|
room temperature
|
|
SDS
|
sodium dodecyl sulfate
|
|
SE
|
standard error
|
|
STAT-3/5
|
signal transducer and activator of
transcription-3/5
|
|
TG
|
triglyceride
|
References
|
1
|
Schwartz MW, Seeley RJ, Zeltser LM,
Drewnowski A, Ravussin E, Redman LM and Leibel RL: Obesity
pathogenesis: An endocrine society scientific statement. Endocr
Rev. 38:267–296. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Karageorgi S, Alsmadi O and Behbehani K: A
review of adult obesity prevalence, trends, risk factors, and
epidemiologic methods in Kuwait. J Obes. 2013:3786502013.
View Article : Google Scholar
|
|
3
|
Rubinstein M and Low MJ: Molecular and
functional genetics of the proopiomelanocortin gene, food intake
regulation and obesity. FEBS Lett. 591:2593–2606. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cristancho AG and Lazar MA: Forming
functional fat: A growing understanding of adipocyte
differentiation. Nat Rev Mol Cell Biol. 28:722–734. 2011.
View Article : Google Scholar
|
|
5
|
Farmer SR: Transcriptional control of
adipocyte formation. Cell Metab. 4:263–273. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mota de Sá P, Richard AJ, Hang H and
Stephens JM: Transcriptional regulation of adipogenesis. Compr
Physiol. 7:635–674. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guo L, Li X and Tang QQ: Transcriptional
regulation of adipo-cyte differentiation: A central role for
CCAAT/enhancer-binding protein (C/EBP) β. J Biol Chem. 290:755–761.
2015. View Article : Google Scholar
|
|
8
|
Ali AT, Hochfeld WE, Myburgh R and Pepper
MS: Adipocyte and adipogenesis. Eur J Cell Biol. 92:229–236. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Richard AJ and Stephens JM: The role of
JAK-STAT signaling in adipose tissue function. Biochim Biophys
Acta. 1842:431–439. 2014. View Article : Google Scholar :
|
|
10
|
Zhang K, Guo W, Yang Y and Wu J:
JAK2/STAT3 pathway is involved in the early stage of adipogenesis
through regulating C/EBPβ transcription. J Cell Biochem.
112:488–497. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lenhard JM: Lipogenic enzymes as
therapeutic targets for obesity and diabetes. Curr Pharm Des.
17:325–331. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Peng IC, Chen Z, Sun W, Li YS, Marin TL,
Hsu PH, Su MI, Cui X, Pan S, Lytle CY, et al: Glucagon regulates
ACC activity in adipocytes through the CAMKKβ/AMPK pathway. Am J
Physiol Endocrinol Metab. 302:E1560–E1568. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Beller M, Bulankina AV, Hsiao HH, Urlaub
H, Jäckle H and Kühnlein RP: PERILIPIN-dependent control of lipid
droplet structure and fat storage in Drosophila. Cell Metab.
12:521–532. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kern PA, Di Gregorio G, Lu T, Rassouli N
and Ranganathan G: Perilipin expression in human adipose tissue is
elevated with obesity. J Clin Endocrinol Metab. 89:1352–1358. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Martini CN, Plaza MV and Maria del Vila C:
PKA-dependent and independent cAMP signaling in 3T3-L1 fibroblasts
differentiation. Mol Cell Endocrinol. 298:42–47. 2009. View Article : Google Scholar
|
|
16
|
Yavari A, Stocker CJ, Ghaffari S, Wargent
ET, Steeples V, Czibik G, Pinter K, Bellahcene M, Woods A, Martínez
de Morentin PB, et al: Chronic activation of γ2 AMPK induces
obesity and reduces β cell function. Cell Metab. 23:821–836. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Prusty D, Park BH, Davis KE and Farmer SR:
Activation of MEK/ERK signaling promotes adipogenesis by enhancing
peroxi-some proliferator-activated receptor gamma (PPARgamma) and
C/EBPalpha gene expression during the differentiation of 3T3-L1
preadipocytes. J Biol Chem. 277:46226–46232. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou Y, Wang D, Li F, Shi J and Song J:
Different roles of protein kinase C-betaI and -delta in the
regulation of adipocyte differentiation. Int J Biochem Cell Biol.
38:2151–2163. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Porras A, Muszynski K, Rapp UR and Santos
E: Dissociation between activation of Raf-1 kinase and the 42-kDa
mitogen-activated protein kinase/90-kDa S6 kinase (MAPK/RSK)
cascade in the insulin/Ras pathway of adipocytic differentiation of
3T3 L1 cells. J Biol Chem. 269:12741–12748. 1994.PubMed/NCBI
|
|
20
|
Kwak DH, Lee JH, Kim DG, Kim T, Lee KJ and
Ma JY: Inhibitory effects of hwangryunhaedok-tang in 3T3-L1
adipo-genesis by regulation of Raf/MEK1/ERK1/2 pathway and PDK1/Akt
phosphorylation. Evid Based Complement Alternat Med.
2013:4139062013. View Article : Google Scholar
|
|
21
|
Choi JS, Kim JH, Ali MY, Jung HJ, Min BS,
Choi RJ, Kim GD and Jung HA: Anti-adipogenic effect of epiberberine
is mediated by regulation of the Raf/MEK1/2/ERK1/2 and AMPKα/Akt
pathways. Arch Pharm Res. 38:2153–2162. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Henry JR, Kaufman MD, Peng SB, Ahn YM,
Caldwell TM, Vogeti L, Telikepalli H, Lu WP, Hood MM, Rutkoski TJ,
et al: Discovery of
1-(3,3-dimethylbutyl)-3-(2-fluoro-4-methyl-5-(7-m
ethyl-2-(methylamino)pyrido[2,3-d]pyrimidin-6-yl)phenyl)urea
(LY3009120) as a pan-RAF inhibitor with minimal paradoxical
activation and activity against BRAF or RAS mutant tumor cells. J
Med Chem. 58:4165–4179. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Peng SB, Henry JR, Kaufman MD, Lu WP,
Smith BD, Vogeti S, Rutkoski TJ, Wise S, Chun L, Zhang Y, et al:
Inhibition of RAF isoforms and active dimers by LY3009120 leads to
anti-tumor activities in RAS or BRAF mutant cancers. Cancer Cell.
28:384–398. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Woods A, Johnstone SR, Dickerson K, Leiper
FC, Fryer LG, Neumann D, Schlattner U, Wallimann T, Carlson M and
Carling D: LKB1 is the upstream kinase in the AMP-activated protein
kinase cascade. Cur Biol. 13:2004–2008. 2003. View Article : Google Scholar
|
|
25
|
Wang D, Zhou Y, Lei W, Zhang K, Shi J, Hu
Y, Shu G and Song J: Signal transducer and activator of
transcription 3 (STAT3) regulates adipocyte differentiation via
peroxisome-proliferator-activated receptor gamma (PPARgamma). Biol
Cell. 102:1–12. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shang CA and Waters MJ: Constitutively
active signal transducer and activator of transcription 5 can
replace the requirement for growth hormone in adipogenesis of
3T3-F442A preadipocytes. Mol Endocrinol. 17:2494–2508. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wolins NE, Brasaemle DL and Bickel PE: A
proposed model of fat packaging by exchangeable lipid droplet
proteins. FEBS Lett. 580:5484–5491. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Steinberg GR, Macaulay SL, Febbraio MA and
Kemp BE: AMP-activated protein kinase-the fat controller of the
energy railroad. Can J Physiol Pharmacol. 84:655–665. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lage R, Diéguez C, Vidal-Puig A and López
M: AMPK: A metabolic gauge regulating whole-body energy
homeostasis. Trends Mol Med. 14:539–549. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Han YH, Kee JY, Park J, Kim HL, Jeong MY,
Kim DS, Jeon YD, Jung Y, Youn DH, Kang J, et al: Arctigenin
inhibits adipogenesis by inducing AMPK activation and reduces
weight gain in high-fat diet-induced obese mice. J Cell Biochem.
117:2067–2077. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vingtdeux V, Chandakkar P, Zhao H, Davies
P and Marambaud P: Small-molecule activators of AMP-activated
protein kinase (AMPK), RSVA314 and RSVA405, inhibit adipogenesis.
Mol Med. 17:1022–1030. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Baek JH, Kim NJ, Song JK and Chun KH:
Kahweol inhibits lipid accumulation and induces glucose-uptake
through activation of AMP-activated protein kinase (AMPK). BMB Rep.
50:566–571. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zordoky BN, Nagendran J, Pulinilkunnil T,
Kienesberger PC, Masson G, Waller TJ, Kemp BE, Steinberg GR and
Dyck JR: AMPK-dependent inhibitory phosphorylation of ACC is not
essential for maintaining myocardial fatty acid oxidation. Circ
Res. 115:518–524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Saha AK and Ruderman NB: Malonyl-CoA and
AMP-activated protein kinase: An expanding partnership. Mol Cell
Biochem. 253:65–70. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Richter EA and Ruderman NB: AMPK and the
biochemistry of exercise: Implications for human health and
disease. Biochem J. 418:261–275. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shaw RJ, Kosmatka M, Bardeesy N, Hurley
RL, Witters LA, DePinho RA and Cantley LC: The tumor suppressor
LKB1 kinase directly activates AMP-activated kinase and regulates
apoptosis in response to energy stress. Proc Natl Acad Sci USA.
101:3329–3335. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu Z, Liu J and Shan T: New roles of Lkb1
in regulating adipose tissue development and thermogenesis. J Cell
Physiol. 232:2296–2298. 2017. View Article : Google Scholar
|
|
38
|
Gormand A, Henriksson E, Ström K, Jensen
TE, Sakamoto K and Göransson O: Regulation of AMP-activated protein
kinase by LKB1 and CaMKK in adipocytes. J Cell Biochem.
112:1364–1375. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hawley SA, Pan DA, Mustard KJ, Ross L,
Bain J, Edelmn AM, Frenguelli BG and Hardie DG: Calmodulin
dependent protein kinase kinase-beta is an alternative upstream
kinase for AMP-activated protein kinase. Cell Metab. 2:9–19. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gowans GJ and Hardie DG: AMPK: A cellular
energy sensor primarily regulated by AMP. Biochem Soc Trans.
42:71–75. 2014. View Article : Google Scholar : PubMed/NCBI
|














