Introduction
Astrocytes represent housekeeper cells in the
central nervous system (CNS), and participate in CNS development
and homeostatic regulation by secreting a wide array of signaling
molecules, including classical neurotransmitters, hormone peptides,
neurotrophins and inflammatory factors (1,2).
The neuroprotective and/or harmful roles of astrocytes depend
largely on the molecules they release into, and take up from, the
extracellular microenvironment (3). In addition, reactive astrogliosis is
a multistage defensive response that has been observed in acute
injuries, such as trauma and ischemic damage, and progressive
pathological conditions, including neuroinflammation and
neurodegeneration in the CNS. This process is characterized by the
alteration of intermediate filament proteins, such as glial
fibrillary acidic protein (GFAP) and vimentin (4,5).
In addition, it has previously been reported that various
categories of functional genes and molecules are modulated by
activated astrocytes in vivo and in vitro, including
inflammatory cell regulators, extracellular transmitters and
neurotrophic factors (6).
The granin family, which comprises markers of the
regulated secretory pathway, contains numerous acidic proteins,
including chromogranin A (CgA), chromogranin B and secretogranin
II-VII. The granins are located in the secretory granules of
various endocrine and neuroendocrine cells, and are involved in
numerous neurological diseases (7-9).
Secretogranin III (SCG3) participates in the early processing of
peptide hormones within the trans-Golgi network and the subsequent
secretion of these peptide hormones via the regulated secretory
pathway (10). By binding with
CgA and carboxypeptidase E (CPE) at specific domains, SCG3 acts as
a molecular bridge between the core aggregate and the
cholesterol-rich secretory granule membrane, and it is regulated
for efficient secretion and processing of proopiomelanocortin
(POMC)-derived peptides in endocrine cells (11,12). Furthermore, SCG3 has been reported
to be associated with the regulated secretory pathway in numerous
types of non-endocrine cells, such as mast cells and platelets
(13,14). Previous studies have confirmed
abundant expression of SCG3 in astrocytes and neuronal populations
(15,16). In addition, SCG3 has been reported
to be involved in neuropathology, such as in major depressive
disorder, multiple sclerosis and Alzheimer's disease (AD); however,
its potential role in the pathogenesis and progression of nervous
system dysfunction has yet to be determined (17,18).
The common pesticide paraquat (PQ), together with
6-hydroxydopamine, 1-methyl-4-phenyl-1,2,3,6-tetrahydro-pyridine
(MPTP) and rotenone, has been extensively used to induce the
pathology of Parkinson's disease in previous experiments (19-21). In our previous studies, a
PQ-activated U118MG astrocytoma cell model was established, and it
was observed that secretogranin II (SCG2), another member of the
granin family, was partially involved in the expression and sorting
of inflammatory factors and neurotransmitters in astroglial
activation. The secretory pathway component SCG3 has previously
been demonstrated to act as a caspase substrate in PQ-induced
dopaminergic cell apoptosis, and its transport is disrupted
following PQ-induced toxicity (16,22,23). However, its role as an anchor
protein of dense-core vesicles (DCVs) remains to be elucidated.
Therefore, in the present study, the PQ-activated astroglial
activation model was used to investigate the role of SCG3 in this
process.
Materials and methods
Materials
Dulbecco's modified Eagle's medium (DMEM),
penicillin, streptomycin and fetal bovine serum (FBS) were obtained
from Gibco; Thermo Fisher Scientific, Inc. (Waltham, MA, USA). PQ
dichloride and Hoechst 33258 were purchased from Sigma-Aldrich;
Merck KGaA (Darmstadt, Germany). RNAiso Plus, PrimeScript RT
Reagent kit and SYBR Prime EX Taq were obtained from Takara
Biotechnology Co., Ltd. (Dalian, China). The Bicinchoninic Acid
Protein Assay kit, SDS-PAGE gel preparing kit, bovine serum albumin
(BSA), radioimmunoprecipitation assay (RIPA) lysis buffer and
protease inhibitor were purchased from Beyotime Institute of
Biotechnology (Shanghai, China), and the Easysee chemiluminescence
reagent was obtained from TransGen Biotech, Co., Ltd. (Beijing,
China). Polyvinylidene difluoride (PVDF) membranes were purchased
from Merck KGaA. Antibodies against β-actin (cat. no. sc-47778) and
SCG3 (cat. no. sc-50289) were obtained from Santa Cruz
Biotechnology, Inc. (Dallas, TX, USA), and anti-GFAP (cat. no.
WL0836) and anti-S100-calcium binding protein β (S100β; cat. no.
WL00789) antibodies were purchased from Wanleibio Co., Ltd.
(Shenyang, China). Anti-interleukin (IL)-6 (cat. no. ab9324) and
anti-brain-derived neurotrophic factor (BDNF; cat. no.
H00000627-M02) were purchased from Abcam (Cambridge, MA, USA) and
Abnova (Taipei, Taiwan), respectively. In addition, Alexa
Fluor® 594-conjugated donkey anti-rabbit immunoglobulin
G (IgG) secondary antibody (cat. no. A-21207), Alexa
Fluor® 488-conjugated goat anti-mouse IgG secondary
antibody (cat. no. A-11029) and Lipofectamine® 3000 were
purchased from Invitrogen; Thermo Fisher Scientific, Inc. SCG3
small interfering RNAs (siRNA_001-003) and the negative control
(NC) siRNA (cat. no. siN05815122147) were obtained from Guangzhou
RiboBio Co., Ltd. (Guangzhou, China).
Cell line
Human astrocytoma U118MG cells (cat. no. TCHu216)
were obtained from Shanghai Institutes for Biological Science
(Shanghai, China), and grown in DMEM High Glucose, supplemented
with 10% FBS and 1% penicillin-streptomycin (10,000 U/ml and 10,000
µg/ml v/v, respectively). U118MG astroglia (5x106) were
seeded in 10 cm2 tissue culture dishes, grown at 37°C in
a humidified incubator containing 5% CO2, and were
exposed to 0.25 mM PQ for 48 h, in order to establish an activated
astroglia model, as described previously (23). PQ treatment was performed in
accordance with ISO 15190:2003 Medical laboratories-Requirements
for safety (24).
Western blotting
Cultured cells treated with 0.25 mM PQ at 37°C for
various durations (0-96 h) were homogenized in ice-cold RIPA with
protease inhibitor. Subsequently, protein concentration was
detected using the bicinchoninic acid method. In order to collect
the conditioned media, cells were grown in serum-starved medium and
were treated with 0.25 mM PQ at 37°C for 0-96 h; the medium
containing PQ was replaced every 24 h. Subsequently, the
supernatant was separated after centrifugation at 2,000 x g for 5
min to remove dislodged cells. The post-nuclear lysate (30
µg/lane) and secretory proteins in conditioned media (30
µl/lane) were separated by 10% SDS-PAGE and were transferred
to PVDF membranes. The membranes were then blocked in a solution
containing 5% non-fat milk powder in Tris-buffered saline
containing 0.05% Tween-20 (TBST) for 2 h at room temperature.
Subsequently, the membranes were incubated with anti-β-actin
(1:1,000) and anti-SCG3 (1:100) primary antibodies in blocking
buffer overnight at 4°C. After three washes in TBST, the membranes
were incubated for a further 2 h at room temperature with
peroxidase-conjugated goat anti-mouse antibody (1:5,000; cat. no.
ZB-2305) and peroxidase-conjugated goat anti-rabbit antibody
(1:2,000; cat. no. ZB-2301) (both ZSGB-BIO Technologies, Inc.,
Beijing, China). Bound antibodies were visualized via Easysee
chemiluminescence reagent using a 5500 Luminescent imaging
workstation (Tanon Science & Technology Co., Ltd., Shanghai,
China). Densitometric analyses of the bands were semi-quantified
using ImageJ software V1.8.0 (National Institutes of Health,
Bethesda, MD, USA), with β-actin as an internal reference.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Cultured cells were treated with 0.25 mM PQ at 37°C
for 0-72 h. To perform RT-qPCR experiments, total RNA was isolated
from cultured astroglia using RNAiso Plus, in accordance with the
manufacturer's protocol. The obtained RNA was then reverse
transcribed to cDNA using a PrimeScript™ RT Reagent kit in a
20-µl reaction mixture containing 4 µl 5X PrimeScript
buffer, 1 µl PrimeScript RT Enzyme Mix I, 1 µl Oligo
dT Primer, 1 µl Random 6 primers and 1 µg total RNA,
under the following thermocycling conditions: 37°C for 15 min
followed by 85°C for 5 sec, in accordance with the manufacturer's
protocol. Subsequently, cDNA was amplified using SYBR Prime EX Taq
via an ABI 7500 Real-Time PCR system (Applied Biosystems; Thermo
Fisher Scientific, Inc.) with the following primers: SCG3 forward,
5′-TGT TAG TGC TCC CGA TTC AAG-3′ and reverse, 5′-TCA AAG GTC TTT
CTG CAC TTA ATT C-3′; and β-actin forward, 5′-AGA TGA CCC AGA TCA
TGT TTG-3′ and reverse, 5′-ATC ACG ATG CCA GTG GTA-3′. The
thermocycling conditions used were as follows: Initial denaturation
at 95°C for 30 sec, followed by 40 cycles at 95°C for 5 sec and
60°C for 34 sec. Each sample was tested in triplicate wells and the
experiment was repeated at least three times. β-actin was used as
an internal reference, and a positive PCR control was performed.
The results were quantified using in the 2-ΔΔCq method
(25).
Transfection with siRNA
U118MG cells (cell density ~70%) were transfected
with SCG3 siRNA or NC siRNA (~100 nM) using
Lipofectamine® 3000 and were treated with PQ immediately
after transfection; RT-qPCR was conducted 48 h later to confirm
successful knockdown of SCG3 mRNA in vitro. A total of 48 h
post-transfection, total RNA was isolated, and the mRNA expression
levels of SCG3 and other astrocytic factors were determined using
RT-qPCR. The primers used for the amplification of astrocytic
factors were described in a previous study (23). The sequences of the most efficient
SCG3 siRNA (siRNA_002) were: Sense, 5′-GAA CAA TAT CTC CAG AAG AdT
dT-3′, and antisense, 5′-UCU UCU GGA GAU AUU GUU CdT dT-3′. In
addition, cell morphology was observed under phase contrast
microscopy (Leica Microsystems, Inc., Buffalo Grove, IL, USA)
post-transfection with SCG3 siRNA.
Fluorescence microscopy
Cultured cells were grown on sterile glass cover
slips at 37°C overnight. After incubation under experimental
conditions (control or 0.25 mM PQ-treated for 48 h), cells were
washed twice with PBS containing 1 mM MgCl2 and 1 mM
CaCl2 (CM), and were fixed with 4% parafor-maldehyde
solution for 15 min. For permeabilization, the fixed cells were
washed in PBS/CM and were incubated for 10 min at room temperature
with 0.2% Triton X-100 in PBS/CM. After three washes with PBS/CM,
the cells were blocked with PBS/CM containing 0.2% BSA for 30 min
at room temperature. The cells were subsequently incubated with
anti-S100β, anti-GFAP, anti-SCG3 and/or anti-IL-6 and anti-BDNF
antibodies (1:50 in PBS/CM containing 0.2% BSA) overnight at 4°C.
After three further washes with PBS/CM, the cells were incubated
with Alexa Fluor® 488-conjugated secondary antibody
(1:2,000 in PBS/CM) and/or Alexa Fluor® 594-conjugated
secondary antibody (1:2,000 in PBS/CM) for 1 h at room temperature.
Finally, the cells were washed three times with PBS/CM and
incubated with Hoechst 33258, in accordance with the manufacturer's
protocol. The cells were observed under a fluorescence microscope
or confocal microscope (Leica Microsystems, Inc.).
Semi-quantitative data were obtained from at least five randomly
selected visual fields from the three different assays. Identical
acquisition parameters were applied when cells were labeled with
the same antibody. The process by which the number of cells were
counted was double-blind and the labeled cell body areas were
measured using ImageJ software V1.8.0 (National Institutes of
Health), according to the methods described in a previous study
(26). Colocalization was
measured using the Co-Localization function of Image Pro Plus 6.0
software (Media Cybernetics, Inc., Rockville, MD, USA). Finally,
the density of each image was normalized against the mean value of
the controls and presented as arbitrary units of the control.
Pearson's correlation coefficient and the overlay percentage of
SCG3 expression from five images per condition were used to
determine colocalization.
Statistical analysis
For statistical analysis, all quantitative data were
collected from at least three independent experiments and the final
data were expressed as the means ± standard deviation.
Statistically significant differences between three or more groups
were determined via one-way analysis of variance followed by
Dunnett's post hoc test using GraphPad Prism 6 (GraphPad Software,
Inc., La Jolla, CA, USA). The differences in immunofluorescence
between the PQ-treated group and the control group were determined
using the Student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
mRNA and protein expression levels of
SCG3 are upregulated in PQ-activated U118MG astroglia
In our previous study, a PQ-activated U118MG
astroglia model was established in vitro; the results
revealed that the cell cycle and expression levels of astrocytic
factors are markedly altered following treatment with PQ (23). To further investigate the activity
of U118MG astroglia, the expression levels of GFAP and S100β, which
are commonly used as markers of astroglial activation, were
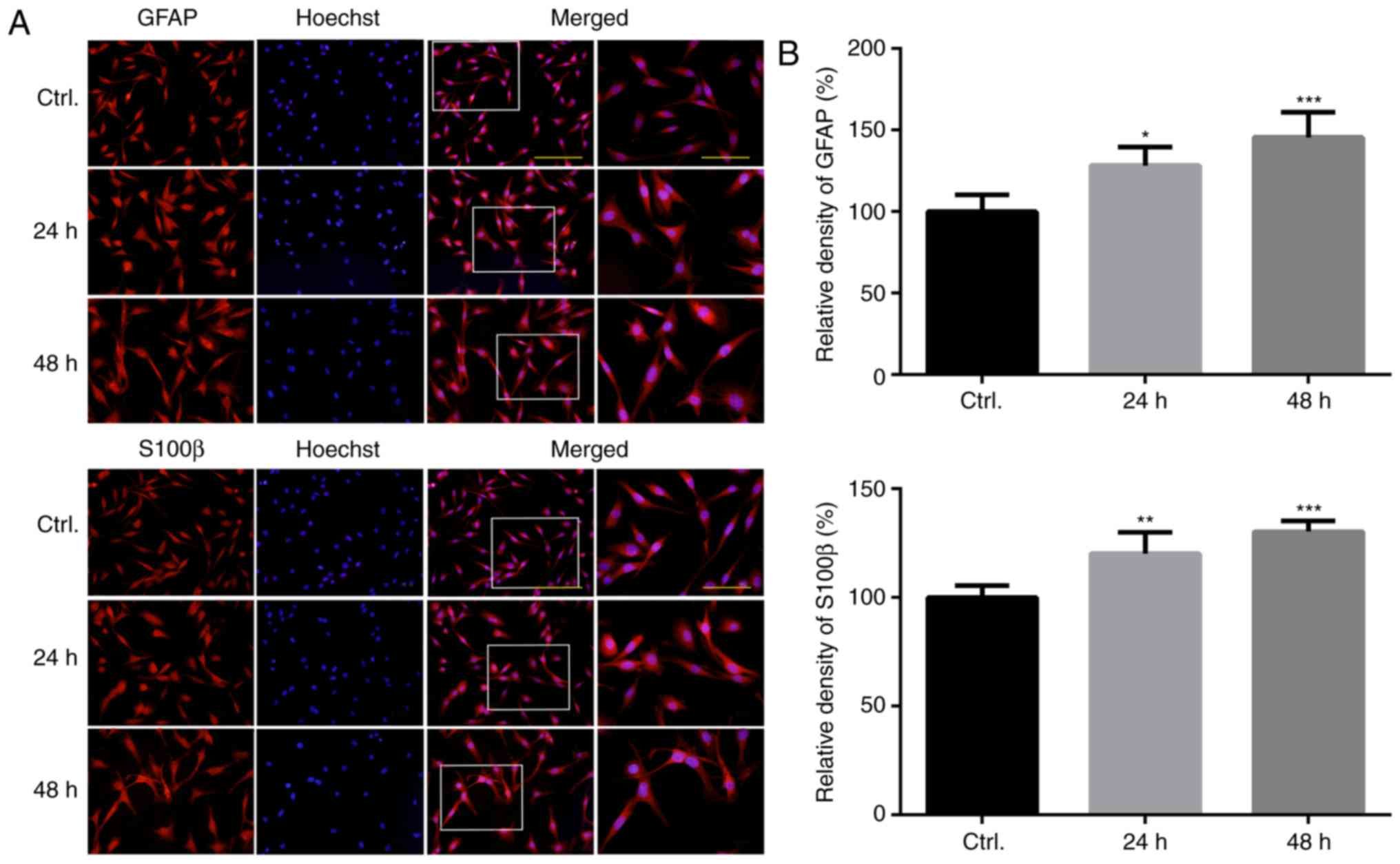
determined in the present study. As presented in Fig. 1, the results of a cell
immunofluorescence analysis revealed that GFAP and S100β proteins
were overexpressed in U118MG astroglia following 48 h of PQ
treatment, thus suggesting that the cells were in a state of
activation (Fig. 1A and B).
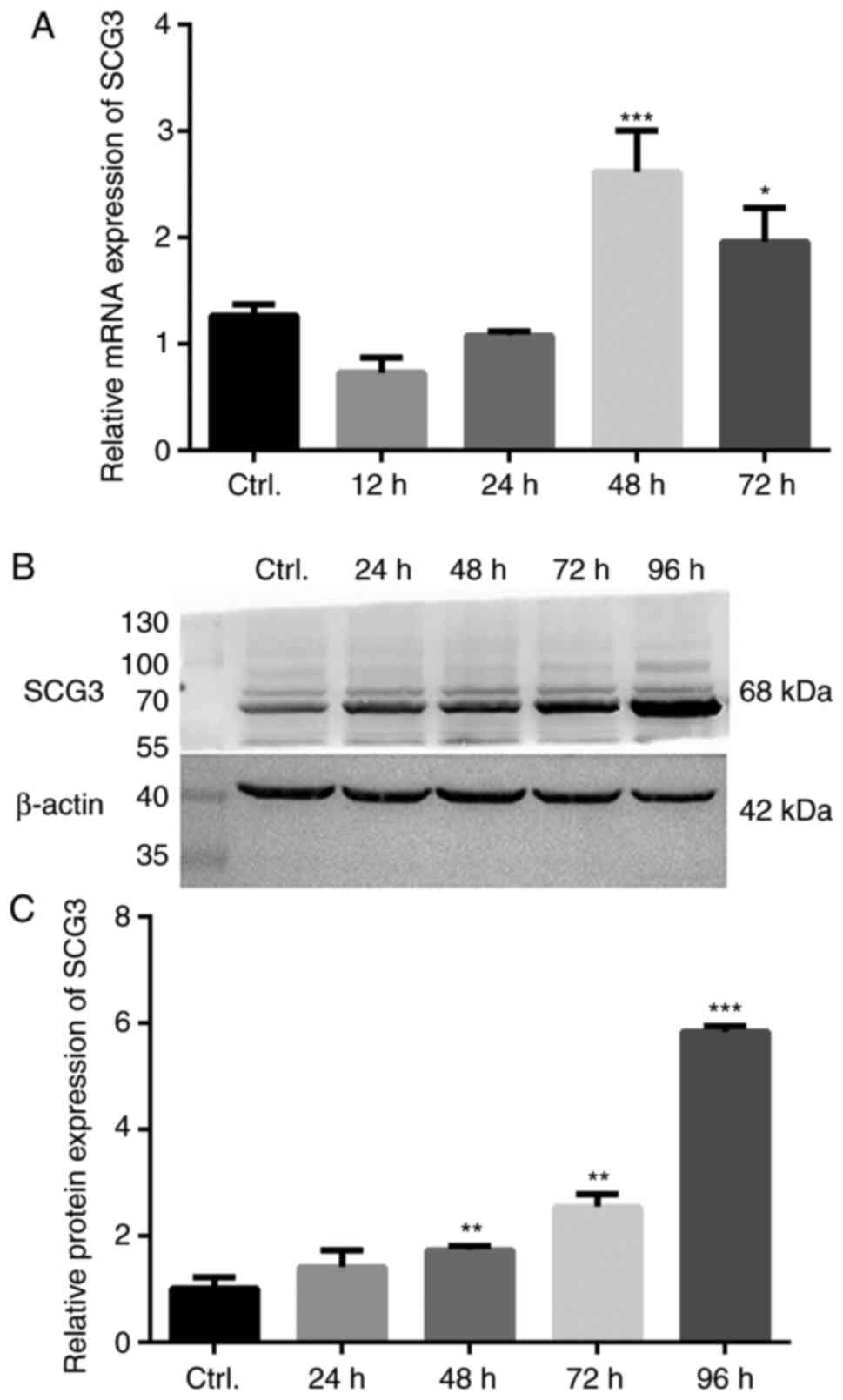
The expression levels of the regulated secretory
pathway component SCG3 were subsequently investigated. The results
of RT-qPCR revealed that the mRNA expression levels of SCG3 were
significantly increased (2.1-fold) at 48 h, and were slightly
decreased at 72 h compared with at 48 h (Fig. 2A). Furthermore, the protein
expression levels of SCG3 were significantly increased at the 48 h
time point and continued to gradually increase in a time-dependent
manner, which was mostly consistent with the mRNA expression
results (Fig. 2B and C).
Subcellular localization and secretion of
SCG3 are altered in PQ-activated U118MG astroglia
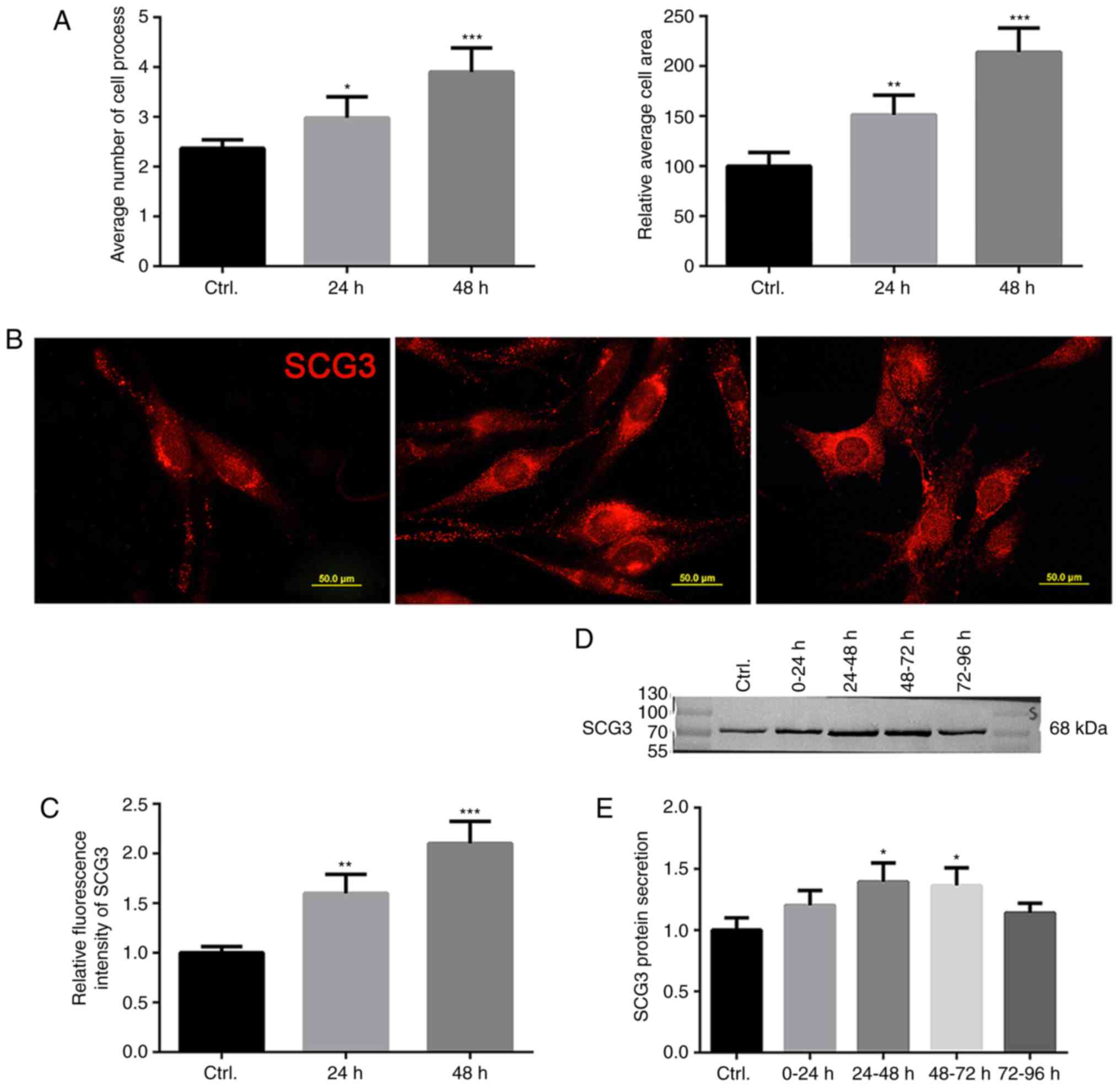
In the present study, the effects of PQ on the
morphology of U118MG cells were investigated; the results
demonstrated that the mean number of cell processes and area of
cell bodies were markedly increased following treatment with PQ for
48 h, thus suggesting that the astroglia developed a hypertrophic
and bushy phenotype following activation (Fig. 3A). Subsequently, the expression
and distribution of SCG3 at the subcellular level were determined
via immunofluorescence. The results revealed that SCG3 was
abundantly expressed and distributed evenly throughout the
cytoplasm and processes of U118MG cells. However, when astroglia
were activated by PQ, the relative fluorescence intensity of SCG3
was markedly increased. Morphologically, punctate-patterned SCG3
markedly accumulated in the perinuclear region in hypertrophic
U118MG astroglia (Fig. 3B and C),
thus indicating that a large number of SCG3-positive vesicles was
rapidly synthesized in the cells following treatment with PQ. In
addition, to determine the levels of SCG3 protein secreted from
PQ-activated astroglia, conditioned culture media were collected at
intervals of 24 h and were further analyzed. When equal volumes of
media were loaded, it was revealed that levels of externally
secreted SCG3 were increased in activated astroglia between 24 and
72 h (Fig. 3D and E).
SCG3 knockdown partially attenuates
PQ-induced alterations to astrocytic morphology, but does not
affect the expression levels of astrocytic factors
To further confirm the involvement of SCG3 in
PQ-stimulated U118MG astroglial activation, the effects of PQ on
SCG3 and astrocytic factor expression were determined after
knockdown of SCG3 via siRNA transfection. Firstly, the efficiency
of three candidate SCG3 siRNAs (siRNA_001-003) was investigated,
and the siRNA that exerted the most marked effect (siRNA_002) was
selected for use in subsequent experiments (Fig. 4A). The results, as observed by
phase contrast microscopy revealed that swelling of U118MG cells
was partially attenuated by transfection with SCG3 siRNA (Fig. 4B). The RT-qPCR results revealed
that SCG3 expression was markedly decreased (0.22-fold) following
transfection with SCG3 siRNA compared with the NC group
(P<0.05). In subsequent experiments, U118MG cells were treated
with 0.25 mM PQ immediately after SCG3 siRNA transfection; RT-qPCR
was conducted after 48 h. The results demonstrated that SCG3
expression was significantly decreased in the siRNA_002 + PQ group
compared with in the PQ group (P<0.05; Fig. 4C). Furthermore, the expression
levels of various astrocyte-derived factors were investigated
following SCG3 knockdown in the present study, including GFAP,
S100β, glutamine synthetase (GS) and glutamate transporter (GLT-1)
astrocyte-specific proteins; BDNF and glial-derived neurotrophic
factor (GDNF) neurotrophic factors; and tumor necrosis factor
(TNF-α), IL-1β and IL-6 inflammatory cytokines (Fig. 4D). However, no significant
alterations in the mRNA expression levels of these factors were
detected between the SCG3 knockdown group and the NC group.
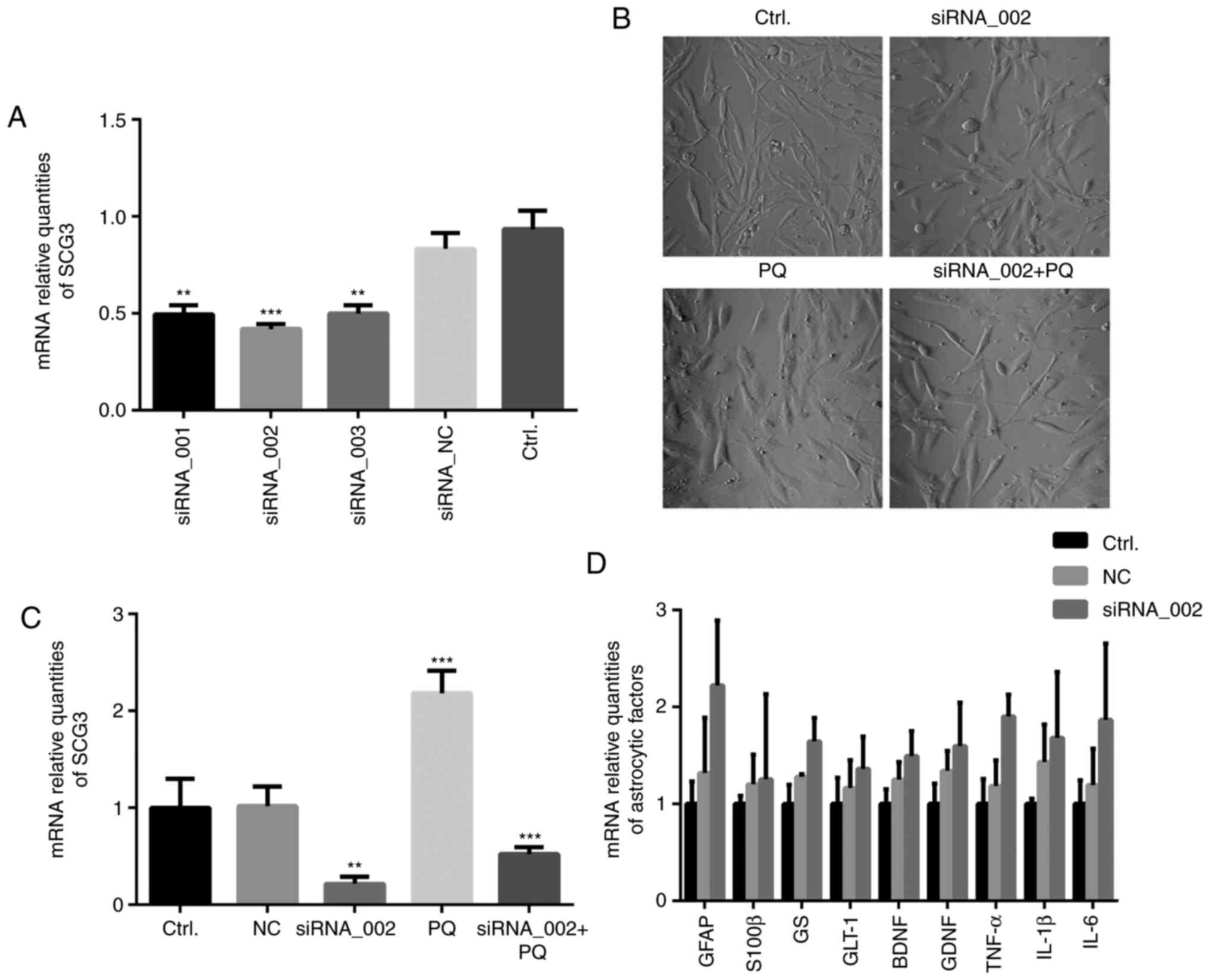 | Figure 4Cell morphology and mRNA expression
levels of astrocytic factors in U118MG astroglia following
knockdown of SCG3. (A) SCG3 mRNA expression levels were analyzed by
RT-qPCR post-transfection with three candidate siRNAs (n=3). SCG3
expression levels were presented relative to β-actin. (B)
Astroglial swelling was partially attenuated by SCG3 siRNA under
phase contrast microscopy (magnification, x20). (C) RT-qPCR was
used to detect SCG3 expression levels following SCG3 knockdown. (D)
mRNA expression levels of astrocyte-specific functional proteins in
U118MG cells were analyzed by RT-qPCR post-transfection with
siRNA_002 (n=3). Data are presented as the means ± standard
deviation. **P<0.01 and ***P<0.001
compared with the control group. BDNF, brain-derived neurotrophic
factor; GDNF, glial-derived neurotrophic factor; GFAP, glial
fibrillary acidic protein; GLT-1, glutamate transporter-1; GS,
glutamine synthetase; IL, interleukin; NC, non-specific control;
PQ, paraquat; RT-qPCR, reverse transcription-quantitative
polymerase chain reaction; S100β, S100-calcium-binding protein β;
SCG3, secretogranin III; siRNA, small interfering RNA; TNF-α, tumor
necrosis factor-α. |
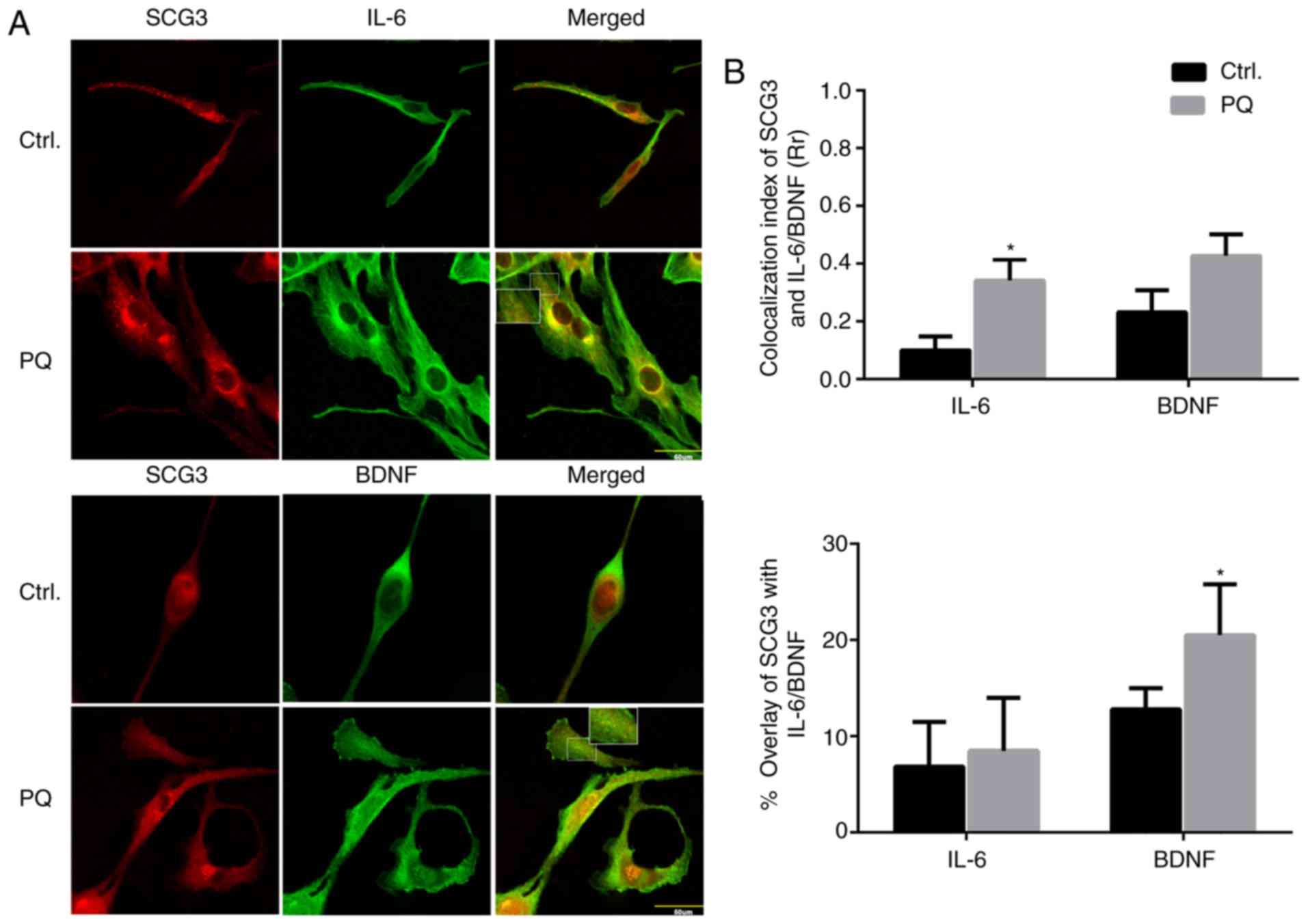
Intracellular SCG3-positive DCVs may
participate in the trafficking of BDNF and IL-6 in PQ-activated
astroglia
It has previously been reported that there are
numerous DCV variants present in astrocytes (27); however, whether SCG3-positive DCVs
are present in astroglia remains unknown. Despite the present
results revealing that there were no marked associations between
SCG3 and gliotransmitters (data not shown), other potential
associations between neurotrophic BDNF and neuropoietic cytokine
IL-6 with SCG3-positive DCVs were investigated via colocalization
analysis in PQ-treated U118MG astroglia. In PQ-activated astroglia,
co-distribution of BDNF immunoreactivity with SCG3-positive
vesicles was observed, and a vesicular pattern of both BDNF and
SCG3 staining was visualized in the perinuclear space and
cytoplasm. However, overlap of IL-6 and SCG3 expression was only
observed in the perinuclear region, which may be due to early
synthesis of the corresponding proteins in the endoplasmic
reticulum (ER; Fig. 5A). In
Pearson's correlation coefficient analysis, the overlapping was
determined by colocalization index: 0.0-0.2 is considered to
indicate an extremely weak or no correlation, whereas 0.2-0.4 and
0.4-0.6 are considered weak and moderate correlations,
respectively. In the control groups, slight colocalization was
demonstrated between BDNF and SCG3, IL-6 and SCG3 (0.23 and 0.1,
respectively). However, the colocalizations were increased
following PQ treatment (0.43 and 0.34, respectively). Furthermore,
when the percentage of overlay in the cytoplasm and processes was
measured, the overlay of SCG3 with BDNF following PQ-induced
activation was increased from 13 to 20.5% (P<0.05), whereas the
overlay of SCG3 with IL-6 was increased from 7.4 to 8.8% (Fig. 5B). These results indicated that
SCG3 DCVs possibly affect the recruitment of BDNF, but not IL-6, in
U118MG cells during PQ-induced astrocyte activation.
Discussion
At present, GFAP is the most commonly used hallmark
for astrocyte activation; however, GFAP overexpression is not
always observed during the activation of all astro-cytes. For
example, astrocytes activated by ATP, IL-1 and MPTP may express
unmodified and even decreased levels of GFAP (28-30). In addition, the role of GFAP in
neuronal regeneration, scar formation and amyloid load assimilation
remains unclear (31,32). Therefore, the present study aimed
to identify an indicator that reflects functional modifications
occurring during the activation of astrocytes. Previous studies
have demonstrated that numerous transporters and carrier proteins,
such as vesicular transporters of γ-aminobutyric acid and glutamate
(excitatory amino acid transporter 1 and 2), are affected following
the activation of astrocytes (33,34). In addition, two individual
microarray analyses identified enhanced expression of the
secretogranin family members secretogranin V and secretogranin VII
in reactive astrocytes isolated from various neuropathologies
(6,20). In the present study, a
PQ-activated astrocyte model established in our previous study was
used, and the results revealed that PQ exposure led to
morphological alterations, and increased expression levels of GFAP
and S100β astrocyte activation markers in U118MG astroglia
(23). In addition, a regulated
secretory pathway component, SCG3, was further investigated in
PQ-activated astrocytes. Our previous study revealed that PQ
exposure induced the apoptosis of dopaminergic SH-SY5Y cells and
decreased SCG3 expression (16).
These findings indicated that alterations in SCG3 expression may
vary in different cell types. In the present study, it was
demonstrated that the expression levels of SCG3 were gradually
increased, in accordance with the degree of U118MG astroglial
activation induced by PQ treatment, thus suggesting that altered
SCG3 may reflect astrocyte reactivity to a certain extent. This
effect has previously been reported in brain perforation and AD
plaque-activated astrocytes (15,35). Conversely, depletion of SCG3 via
RNA interference attenuated the hypertrophic phenotype of U118MG
astroglia and reversed PQ-induced increases in SCG3 expression,
which further suggested that SCG3 may have an active role in
PQ-induced astrocyte activation. Therefore, it was hypothesized
that SCG3 secretion in astrocytes may represent an indicator of
cellular activation and provide further information regarding
transmitter signaling.
SCG3-positive vesicles have been identified in
primary astrocyte cultures and human brain samples (35). Furthermore, aberrant accumulation
of SCG3 and CPE in senile plaque-surrounding astrocytes and
dystrophic neurites obtained from patients with AD suggests that
SCG3 may affect neuropeptide trafficking and processing (15,35). However, the mechanism by which
neurotransmitters and peptides are transported by SCG3 secretory
vesicles in astrocytes remains largely unknown. In the present
study, both extracellular and intracellular SCG3 expression levels
were upregulated in U118MG astroglia following treatment with PQ.
Immunofluorescence analysis demonstrated that SCG3-positive
vesicles accumulated in the perinuclear ER, where they were
generated following exposure to PQ. These results indicated that PQ
stimulated the biosynthesis of SCG3 secretory vesicles, thus
suggesting that SCG3 may be involved in the recruitment of cellular
factors during PQ-induced astrocyte activation. A previous study
indicated that DCVs isolated from astrocytes contain a range of
gliotransmitters, such as atrial natriuretic peptide, neuropeptide
Y and ATP, which serve important roles in astrocyte metabolism
(36). In our previous study, it
was demonstrated that three categories of astrocytic proteins were
upregulated to different extents in PQ-activated U118MG astroglia,
including astrocytic marker proteins (GFAP, S100β, GS and GLT-1),
neurotrophins (BDNF and GDNF), and cytokines (TNF-α, IL-1β, IL-6)
(23). However, the mRNA
expression levels of the aforementioned proteins did not exhibit
any significant alterations following the silencing of SCG3.
Therefore, it was hypothesized that SCG3 secretory vesicles may be
associated with the regulation of secretory peptide secretion,
rather than the regulation of gene expression, in PQ-activated
astrocytes.
Previous studies have demonstrated that BDNF targets
to SCG2-positive DCVs in neuronal dendrites, and BDNF and SCG2
genes are co-activated in response to intracellular Ca2+
signaling (33,37,38). In addition, the sorting signal of
BDNF can specifically bind to the sorting receptor CPE, which is
localized closely with SCG3 at the periphery region of endocrine
cells (12,39,40). In the present study, it was
observed that double-labeled vesicles were distributed in the
peri-nuclear ER and cytoplasm of U118MG astroglia, and their
numbers were increased during PQ stimulation. Therefore, it was
hypothesized that SCG3 may act as an analogous sorting receptor to
CPE during the processing and trafficking of BDNF. In addition,
SCG3 is involved in inflammatory mediator biogenesis and release in
mast cells, which is partially analogous to astrocytes during
immune regulation (14). It has
also been reported that IL-6 is stored in SCG2-positive DCVs and is
released via the regulated secretory pathway following ATP
stimulation in PC12 cells (41).
Therefore, the role of SCG3 in IL-6 trafficking was analyzed. The
results of the present study revealed that the overlap of IL-6 and
SCG3 vesicles was predominantly observed in the perinuclear space
of PQ-activated U118MG astroglia, which may be due to their common
biosynthesis in the ER. However, when the percentage of overlay in
the cytoplasm and processes was measured, there was no significant
increase in the overlay between SCG3 and IL-6 following PQ-induced
activation. Therefore, it may be hypothesized that SCG3-positive
vesicles exert a significant effect on BDNF trafficking but do not
participate in the subsequent processing and trafficking of IL-6 in
astrocytes.
In conclusion, the results of the present study
revealed that the regulated secretory pathway component SCG3 was
expressed in U118MG astroglia, and SCG3 expression and astrocyte
activation were both gradually increased in response to PQ
stimulation. Despite the fact that SCG3 knockdown did not affect
the expression of other astrocytic factors, de novo SCG3
vesicles contributed to the trafficking of BDNF, but not IL-6, in
PQ-activated astrocytes. These results suggested that SCG3 may act
as an indicator of astrocyte activation. In addition, SCG3 may have
a role in astrocyte self-sustenance as well as neuroglia peptide
communication; however, the present data were derived from an in
vitro astrocytoma cell line model, and only one PQ activator
was used. Therefore, some limitations may exist with regards to the
alteration of SCG3 in response to astrocyte activation. For these
reasons, future experiments should be performed using primary
astrocytes or animal models, and the role of SCG3 in astrocyte
activation should be reported in response to other activators
reported in previous studies; for example, ATP, IL-1 and MPTP
(28-30). Furthermore, considering the small
number of DCVs in astrocytes, further experiments to
pre-concentrate and separate SCG3-positive vesicles would be
beneficial to confirm the target aggregates of SCG3 and to
determine its role in the pathogenesis of neurodegenerative
disorders.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81172713
and 81471826).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XZ wrote the manuscript. XZ and QC conducted the
experiments. XZ and FL analyzed the results and modified the
manuscript. HP made substantial contributions to conception and
design, he also revised the manuscript critically for important
intellectual content. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Parpura V, Heneka MT, Montana V, Oliet SH,
Schousboe A, Haydon PG, Stout RF Jr, Spray DC, Reichenbach A,
Pannicke T, et al: Glial cells in (patho)physiology. J Neurochem.
121:4–27. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schousboe A and Waagepetersen HS: Role of
astrocytes in glutamate homeostasis: Implications for
excitotoxicity. Neurotox Res. 8:221–225. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yu B, Changsheng Y, Wenjun Z, Ben L, Hai
Q, Jing M, Guangwei X, Shuhua W, Fang L, Aschner M and Rongzhu L:
Differential protection of pre-versus post-treatment with curcumin,
Trolox, and N-acetylcysteine against acrylonitrile-induced
cytotoxicity in primary rat astrocytes. Neurotoxicology. 51:58–66.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ben Haim L, Carrillo-de Sauvage MA,
Ceyzeriat K and Escartin C: Elusive roles for reactive astrocytes
in neurodegen-erative diseases. Front Cell Neurosci. 9:2782015.
View Article : Google Scholar
|
|
5
|
Eng LF, Ghirnikar RS and Lee YL: Glial
fibrillary acidic protein: GFAP-thirty-one years (1969–2000).
Neurochem Res. 25:1439–1451. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zamanian JL, Xu L, Foo LC, Nouri N, Zhou
L, Giffard RG and Barres BA: Genomic analysis of reactive
astrogliosis. J Neurosci. 32:6391–6410. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Taupenot L, Harper KL and O'Connor DT: The
chromo-granin-secretogranin family. N Engl J Med. 348:1134–1149.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bartolomucci A, Possenti R, Mahata SK,
Fischer-Colbrie R, Loh YP and Salton SR: The extended granin
family: Structure, function, and biomedical implications. Endocr
Rev. 32:755–797. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
van Luijn MM, van Meurs M, Stoop MP,
Verbraak E, Wierenga-Wolf AF, Melief MJ, Kreft KL, Verdijk RM, 't
Hart BA, Luider TM, et al: Elevated expression of the cerebrospinal
fluid disease markers chromogranin a and clusterin in astrocytes of
multiple sclerosis white matter lesions. J Neuropathol Exp Neurol.
75:86–98. 2016. View Article : Google Scholar
|
|
10
|
Hosaka M and Watanabe T: Secretogranin
III: A bridge between core hormone aggregates and the secretory
granule membrane. Endocr J. 57:275–286. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hosaka M, Suda M, Sakai Y, Izumi T,
Watanabe T and Takeuchi T: Secretogranin III binds to cholesterol
in the secretory granule membrane as an adapter for chromogranin A.
J Biol Chem. 279:3627–3634. 2004. View Article : Google Scholar
|
|
12
|
Hosaka M, Watanabe T, Sakai Y, Kato T and
Takeuchi T: Interaction between secretogranin III and
carboxypeptidase E facilitates prohormone sorting within secretory
granules. J Cell Sci. 118:4785–4795. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Coppinger JA, Cagney G, Toomey S,
Kislinger T, Belton O, McRedmond JP, Cahill DJ, Emili A, Fitzgerald
DJ and Maguire PB: Characterization of the proteins released from
activated platelets leads to localization of novel platelet
proteins in human atherosclerotic lesions. Blood. 103:2096–2104.
2004. View Article : Google Scholar
|
|
14
|
Prasad P, Yanagihara AA, Small-Howard AL,
Turner H and Stokes AJ: Secretogranin III directs secretory vesicle
biogenesis in mast cells in a manner dependent upon interaction
with chro-mogranin A. J Immunol. 181:5024–5034. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Paco S, Pozas E and Aguado F:
Secretogranin III is an astrocyte granin that is overexpressed in
reactive glia. Cereb Cortex. 20:1386–1397. 2010. View Article : Google Scholar
|
|
16
|
Li F, Tian X, Zhou Y, Zhu L, Wang B, Ding
M and Pang H: Dysregulated expression of secretogranin III is
involved in neurotoxin-induced dopaminergic neuron apoptosis. J
Neurosci Res. 90:2237–2246. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Teunissen CE, Koel-Simmelink MJ, Pham TV,
Knol JC, Khalil M, Trentini A, Killestein J, Nielsen J, Vrenken H,
Popescu V, et al: Identification of biomarkers for diagnosis and
progression of MS by MALDI-TOF mass spectrometry. Mult Scler.
17:838–850. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Teyssier JR, Ragot S, Chauvet-Gélinier JC,
Trojak B and Bonin B: Activation of a ΔFOSB dependent gene
expression pattern in the dorsolateral prefrontal cortex of
patients with major depressive disorder. J Affect Disord.
133:174–178. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Berry C, La Vecchia C and Nicotera P:
Paraquat and Parkinson's disease. Cell Death Differ. 17:1115–1125.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nakagawa T and Schwartz JP: Gene
expression profiles of reactive astrocytes in dopamine-depleted
striatum. Brain Pathol. 14:275–280. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sandström J, Broyer A, Zoia D, Schilt C,
Greggio C, Fournier M, Do KQ and Monnet-Tschudi F: Potential
mechanisms of development-dependent adverse effects of the
herbicide paraquat in 3D rat brain cell cultures. Neurotoxicology.
60:116–124. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li F, Tian X, Zhan X, Wang B, Ding M and
Pang H: Clathrin-dependent uptake of paraquat into SH-SY5Y cells
and its internalization into different subcellular compartments.
Neurotox Res. 32:204–217. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhan X, Li F, Chu Q and Pang H: Effects of
PQ's cytotoxicity on secretory vesicles in astroglia: Expression
alternation of secretogranin II and its potential interaction with
intracellular factors. Biochem Biophys Res Commun. 497:675–682.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Noble MA: ISO 15190:2003 medical
laboratories-requirements for safety. EJIFCC. 15:141–143.
2004.PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
26
|
Sazonova OV, Lee KL, Isenberg BC, Rich CB,
Nugent MA and Wong JY: Cell-cell interactions mediate the response
of vascular smooth muscle cells to substrate stiffness. Biophys J.
101:622–630. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Verkhratsky A, Matteoli M, Parpura V,
Mothet JP and Zorec R: Astrocytes as secretory cells of the central
nervous system: Idiosyncrasies of vesicular secretion. EMBO J.
35:239–257. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
John GR, Lee SC, Song X, Rivieccio M and
Brosnan CF: IL-1-regulated responses in astrocytes: Relevance to
injury and recovery. Glia. 49:161–176. 2005. View Article : Google Scholar
|
|
29
|
Niranjan R, Nath C and Shukla R: The
mechanism of action of MPTP-induced neuroinflammation and its
modulation by melatonin in rat astrocytoma cells, C6. Free Radic
Res. 44:1304–1316. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Adzic M, Stevanovic I, Josipovic N, Laketa
D, Lavrnja I, Bjelobaba IM, Bozic I, Jovanovic M, Milosevic M and
Nedeljkovic N: Extracellular ATP induces graded reactive response
of astrocytes and strengthens their antioxidative defense in vitro.
J Neurosci Res. 95:1053–1066. 2017. View Article : Google Scholar
|
|
31
|
Pekny M, Levéen P, Pekna M, Eliasson C,
Berthold CH, Westermark B and Betsholtz C: Mice lacking glial
fibrillary acidic protein display astrocytes devoid of intermediate
filaments but develop and reproduce normally. EMBO J. 14:1590–1598.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kraft AW, Hu X, Yoon H, Yan P, Xiao Q,
Wang Y, Gil SC, Brown J, Wilhelmsson U, Restivo JL, et al:
Attenuating astrocyte activation accelerates plaque pathogenesis in
APP/PS1 mice. FASEB J. 27:187–198. 2013. View Article : Google Scholar :
|
|
33
|
Calabrese F, Rossetti AC, Racagni G, Gass
P, Riva MA and Molteni R: Brain-derived neurotrophic factor: A
bridge between inflammation and neuroplasticity. Front Cell
Neurosci. 8:4302014. View Article : Google Scholar
|
|
34
|
Huang S, Tong H, Lei M, Zhou M, Guo W, Li
G, Tang X, Li Z, Mo M, Zhang X, et al: Astrocytic glutamatergic
transporters are involved in Aβ-induced synaptic dysfunction. Brain
Res. 1678:129–137. 2018. View Article : Google Scholar
|
|
35
|
Plá V, Paco S, Ghezali G, Ciria V, Pozas
E, Ferrer I and Aguado F: Secretory sorting receptors
carboxypeptidase E and secretogranin III in amyloid β-associated
neural degeneration in Alzheimer's disease. Brain Pathol.
23:274–284. 2013. View Article : Google Scholar
|
|
36
|
Potokar M, Stenovec M, Kreft M, Kreft ME
and Zorec R: Stimulation inhibits the mobility of recycling
peptidergic vesicles in astrocytes. Glia. 56:135–144. 2008.
View Article : Google Scholar
|
|
37
|
Fujita Y, Katagi J, Tabuchi A, Tsuchiya T
and Tsuda M: Coactivation of secretogranin-II and BDNF genes
mediated by calcium signals in mouse cerebellar granule cells.
Brain Res Mol Brain Res. 63:316–324. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kuczewski N, Porcher C, Ferrand N,
Fiorentino H, Pellegrino C, Kolarow R, Lessmann V, Medina I and
Gaiarsa JL: Backpropagating action potentials trigger dendritic
release of BDNF during spontaneous network activity. J Neurosci.
28:7013–7023. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lou H, Kim SK, Zaitsev E, Snell CR, Lu B
and Loh YP: Sorting and activity-dependent secretion of BDNF
require interaction of a specific motif with the sorting receptor
carboxypeptidase E. Neuron. 45:245–255. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cawley NX, Rathod T, Young S, Lou H, Birch
N and Loh YP: Carboxypeptidase E and Secretogranin III coordinately
facilitate efficient sorting of proopiomelanocortin to the
regulated secretory pathway in AtT20 cells. Mol Endocrinol.
30:37–47. 2016. View Article : Google Scholar :
|
|
41
|
Moller JC, Kruttgen A, Burmester R, Weis
J, Oertel WH and Shooter EM: Release of interleukin-6 via the
regulated secretory pathway in PC12 cells. Neurosci Lett.
400:75–79. 2006. View Article : Google Scholar : PubMed/NCBI
|