Introduction
Acute-on-chronic liver failure (ACLF) has been
increasingly recognized as a distinct disease entity. ACLF has been
newly-defined as a serious syndrome characterized by the acute
decompensation (AD) of hepatic cirrhosis, which encompasses the
acute deterioration of hepatic function and involves the
dysfunction of multiple other organs in patients with chronic liver
disease (1). Based on an analytic
study of 1,343 patients, the European Association for the Study of
the Liver and Chronic Liver Failure (EASL-CLIF) consortium
suggested the following diagnostic criteria for ACLF: i) Liver
cirrhosis and AD as defined by the development of ascites,
encephalopathy, gastrointestinal hemorrhage and bacterial
infection; ii) organ failure assessed according to the Chronic
Liver Failure-Sequential Organ Failure Assessment (CLIF-SOFA)
score; iii) a 28-day mortality rate >15%. In this CANONIC study,
the prevalence of ACLF reached as high as 30.9%, and this
considerably relevant syndrome appeared to be different from AD,
which was not only based on liver failure, multiple organ failure
and high mortality rates, but also on age, precipitating events and
systematic inflammation (2). It
has been reported that alcoholic liver cirrhosis represents 50-70%
of the underlying liver diseases of ACLF in Western countries,
whereas hepatitis-associated liver cirrhosis constitutes 10-30% of
all ACLF cases. In the majority of Asian countries with high a
prevalence of hepatitis B virus (HBV) infection, chronic hepatitis
B (CHB) is considered to be the major causative factor for ACLF,
accounting for ~70% of ACLF cases, and accounts for as little as
15% of alcohol-related ACLF (2-4).
To date, the mechanism underlying the development
and progression of ACLF remain to be fully elucidated. It has been
noted that that ACLF is correlated to immune dysregulation.
However, studies have mainly focused on T-cell response, despite
the fact that human lymphocytes are well known to comprise T- and
B-cells, which account for almost 20% of peripheral lymphocytes.
These fulfil an important role in the immune response. An effective
B-cell immune response is largely dependent on B-cells with diverse
B-cell receptors (BCRs), which are capable of specifically
recognizing and binding antigens. Each BCR is structurally
connected by two heavy chains with variable (V), constant (C),
transmembrane and cytoplasmic regions, and two light (L) chains
which have V and C regions.
It has been documented that the V region comprises
two domains, namely, VH and VL, and possesses three
complementarity-determining regions(CDRs): CDR1, CDR2 and CDR3.
CDRs are variable in amino acid composition and sequence, and the
range can reach as high as 69-612 in the
library, allowing the identification of a variety of antigens and
the production of specific antibodies. These three CDRs act to
identify and jointly determine BCR antigen specificity (5). Similar to the T-cell receptor
repertoire, the diversity of the BCR repertoire is possibly formed
through the recombination of multiple sets of similar genes to
generate functional genes. Immunoglobulin heavy (IGH) chains are
encoded by recombination of variable (V), diversity (D) and joining
(J) genes, which are formed from sets of V, D and J genes (IGHV and
IGHJ), whereas VJ rearrangements of the κ and λ chain V genes
(IGKV, IGLV) and J genes (IGKJ, IGLJ) encode the immunoglobulin
light chains (6,7). It has long been observed that there
is a broad deficit of B-cells in patients who suffer from acute
liver diseases, and that there is an association between exposure
to circulating antigens and the abnormality of shunting or Kupffer
cells, which is presumably the consequence (8).
High-throughput sequencing (HTS) is an advanced
genetic tool that enables the investigation of more complex
sequencing targets. In the present study, HTS was coupled with
multiplex polymerase chain reaction (PCR), Illumina sequencing and
the international ImMunoGeneTics database to assess the composition
and alteration of BCR heavy chain repertoires associated with ACLF.
It is anticipated that the results of the present study may provide
direct evidence of the role of BCR heavy chain repertoires in the
development and progression of ACLF.
Materials and methods
Patients
In the present prospective study, six patients with
HBV-related ACLF and six healthy controls (control subjects) were
recruited between January and September 2013 at the No. 181
Hospital of Guilin (Guilin, China). The diagnosis of ACLF was
pathologically and clinically confirmed on the basis of the
diagnostic criteria developed by the Asia-Pacific Association for
the Study of the Liver (3,9),
which has been accepted for defining this novel clinical entity as
follows: ‘An acute hepatic insult manifesting as jaundice and
coagulopathy, which is complicated within four weeks by ascites
and/or encephalopathy in a patient with previously diagnosed or
undiagnosed chronic liver disease’. The patients with HBV-related
ACLF were confirmed to have CHB, without any evidence of
sepsis/bacterial infection, combined hepatitis C virus infection,
or active alcoholism. Therefore, the patients with ACLF in the
present study shared a similar etiology, which was HBV infection.
However, there were no differences among BCRs associated with the
etiologies of ACLF. It is noteworthy that jaundice with serum
bilirubin ≥5 mg/dl or 85 µmol/l, and coagulopathy with INR
>1.5 or prothrombin activity <40% are required for the final
diagnosis of ACLF (3). The
patients with HBV-related ACLF had a mean age of 34.12 years old,
with an age range of 20-54 years old. The six healthy controls
(control subjects) were matched for age, gender and ethnicity, and
had no clinical signs of liver disease.
Prior to the present study, written informed consent
was obtained from each participant or from their guardians. The
present study complied with the Declaration of Helsinki for
investigations involving humans, and the study protocol was
carefully reviewed and approved by the Ethics Committee of the No.
181 Hospital of Guilin.
Multiplex PCR
Amplification of the BCR CDR3 region by
multiplex-PCR. In order to prepare the template library for
subsequent analysis using a Genome Analyzer, multiplex-PCR was
performed. DNA was extracted from PBMCs using GenFIND DNA
(Agencourt; Beckman Coulter, Inc., Brea, CA, USA) extraction kits
according to the manufacturer’s protocol. The following were added
to a 200-µl PCR tube: 500 ng DNA, Mul-PCR Primer (V primer
and J primer) using a QIAGEN multiplex PCR kit. The tubes were
placed into a thermal cycler and the PCR cycling program was run
with 30 cycles. Agarose gel electrophoresis: Using 2% agarose gel,
electrophoresis conditions: 100 V for 2 h. Size selection: 100-190
bp. The amplicons were purified using the QIAquick Gel Extraction
kit (Qiagen GmbH, Hilden, Germany). Samples were quantified using
Quant-iT PicoGreen (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The rearranged BCR CDR3 regions were amplified
using genomic DNA as the template, with 12 forward primers, each
specific to the functional BCR-V segment, and four reverse primers,
each unique to the BCR-J segment. All the forward and reverse
primers harbored the universal forward and reverse primer sequences
(forward, 5′-AAT GAT ACG GCG ACC ACC GA-3′ reverse, 5′-CAA GCA GAA
GAC GGC ATA CGA-3′) at their 5′ ends, respectively, which are
compatible with the GA2 cluster station solid-phase PCR. Following
the multiplex-PCR amplification, the targeted PCR products were
purified using agarose gel electrophoresis and a QIA quick PCR
purification kit, according to the manufacturer’s protocol. The
generated library was then quantified using the following two
approaches: Determination of the average length on the Agilent 260
bioanalyzer instrument with Agilent DNA 600 reagents, and reverse
transcription-quantitative PCR analysis using the TaqMan probe. For
multiplex PCR amplification of DNA, 30 ng DNA was mixed with the JH
reverse primer and the FR1 forward primer set (0.25 µM
each), using 0.5 µl Phusion® High-Fidelity DNA
Polymerase (Finnzymes; Thermo Fisher Scientific, Inc.), 1 µl
dNTPs (0.25 µM) and 1 µl DTT (0.25 µM), per 50
µl reaction. The following PCR program was used: 3 min at
94°C, 35 cycles of 30 sec at 94°C, 30 sec at 60°C and 1 min at
72°C, with a final extension cycle of 7 min at 72°C on an MJ
Thermocycler (10).
The libraries were subsequently amplified with the
c-Bot system to create a cluster on the flow cell, followed by
paired-end sequencing of the amplified flow cell using an Illumina
MiSeq instrument, in which the generally accepted read length of 60
bp was used. In order to ensure analysis quality, any raw reads
that contained the low quality sequence and adaptor sequence were
filtered, following which clean reads suitable for the subsequent
data analysis were obtained. The data processing procedures were
summarized as follows: i) Removal of reads with the adapter; ii)
elimination of N reads, as N is unable to determine the information
for a base with a proportion >6%; iii) removal of reads of
relatively low quality (sequence quality value ≤5 bases accounting
for >50% of the entire read).
Data analysis
During the development of B-lymphocytes, the V, D
and J gene rearrangement of BCR, the V-J fragments, or that between
the V-D-J fragments may be randomly inserted into various numbers
of nucleotides. By contrast, different numbers of nucleotides may
be missed in the V-J fragments or those between the V-D-J
fragments. Therefore, a highly diverse CDR3 variable area, which
has marked differences in length and amino acid sequence, was
formed. This allowed the diversity of B-lymphocytes to be assessed
through the length of CDR3. The length of CDR3 in normal
individuals is similarly normally distributed. In the present
study, systematic analysis of the frequent use of V, D J and V-J
fragments was performed. The upregulated and downregulated genes
were identified using Student’s t-test to compare the BCR heavy
chain V, D, J and V-J fragments. Furthermore, the diversity of the
BCR repertoire was assessed using the Simpson index of diversity
(Ds) (10) and Shannon-Wiener
index (H′) (11) in the present
study. The statistical analyses were conducted using GraphPad Prism
software (version 4.03; GraphPad Software, San Diego, CA,
USA). The Mann-Whitney test was used to analyze the
differences between the ACLF and control groups, given the
relatively small sample size in the present study. P<0.05 was
considered to indicate a statistically significant difference.
Results
BCR heavychainCDR3 sequences from
patients with HBV-related ACLF and control subjects
The HTS technique was used to capture the BCR
heavychainCDR3 sequences of B-cells prepared from peripheral blood
obtained from the recruited patients with ACLF and control
subjects. As a result, an average number of 12,243,860.30 in the
control group and an average number of 12,299,65.30 in the ACLF
group were initially obtained. As the Raw Reads or Sequenced Reads
contain low-quality sequences and adaptors sequences, these Raw
Reads were filtered to obtain high-quality clean reads with an
average number of 6,674,277.80 in the control group and an average
number of 6,114,722.70 in the ACLF group, ensuring the quality of
the information analysis. These were used for the subsequent data
analysis. The total reads/sequences, BCR sequences, in-frame
sequences, total IGH CDR3 sequences, unique CDR3 nt sequences,
unique CDR3 aa sequences, highly expanded clone (HEC) numbers, and
HEC ratios are shown in Table
I.
 | Table IComparison of B-cell receptor
statistical data. |
Table I
Comparison of B-cell receptor
statistical data.
| Individual | Total reads | In-frame
sequences | Total CDR3
sequences | Unique cdr3 nt
sequences | Unique cdr3 aa
sequences | HEC number, all |
|---|
| NC-1 | 15,410,305 | 13,317,137 | 12,584,061 | 414,914 | 292,378 | 0 |
| NC-2 | 15,517,826 | 13,016,806 | 12,249,709 | 391,751 | 276,673 | 1 |
| NC-3 | 14,022,700 | 11,415,501 | 6,600,111 | 260,471 | 181,219 | 4 |
| NC-4 | 537,114 | 465,901 | 439,959 | 43,655 | 30,075 | 2 |
| NC-5 | 14,696,122 | 12,483,863 | 11,778,081 | 392,836 | 279,047 | 2 |
| NC-6 | 13,279,095 | 11,231,688 | 6,575,248 | 432,519 | 316,507 | 0 |
| ACLF-1 | 13,494,348 | 11,594,496 | 6,925,522 | 388,512 | 281,302 | 2 |
| ACLF-2 | 11,264,437 | 9,942,837 | 9,479,612 | 452,729 | 329,633 | 0 |
| ACLF-3 | 9,958,559 | 8,099,585 | 7,733,592 | 190,996 | 140,723 | 201 |
| ACLF-4 | 12,978,762 | 9,887,906 | 8,771,599 | 168,652 | 118,150 | 446 |
| ACLF-5 | 12,819,431 | 6,862,593 | 6,206,099 | 316,046 | 228,715 | 6 |
| ACLF-6 | 12,539,789 | 6,300,919 | 9,651,145 | 446,872 | 326,426 | 0 |
Comparison of immune diversity in
patients with ACLF and control subjects
In order to compare the immune diversity between the
ACLF and control groups, the normalized Shannon entropy index was
applied, which has been well-accepted for quantitatively measuring
diversity. In the analysis of immune diversity in these two groups,
the index was calculated and rated between 0 and 1, with 1 as the
highest diversity and 0 as an indication for no immune diversity.
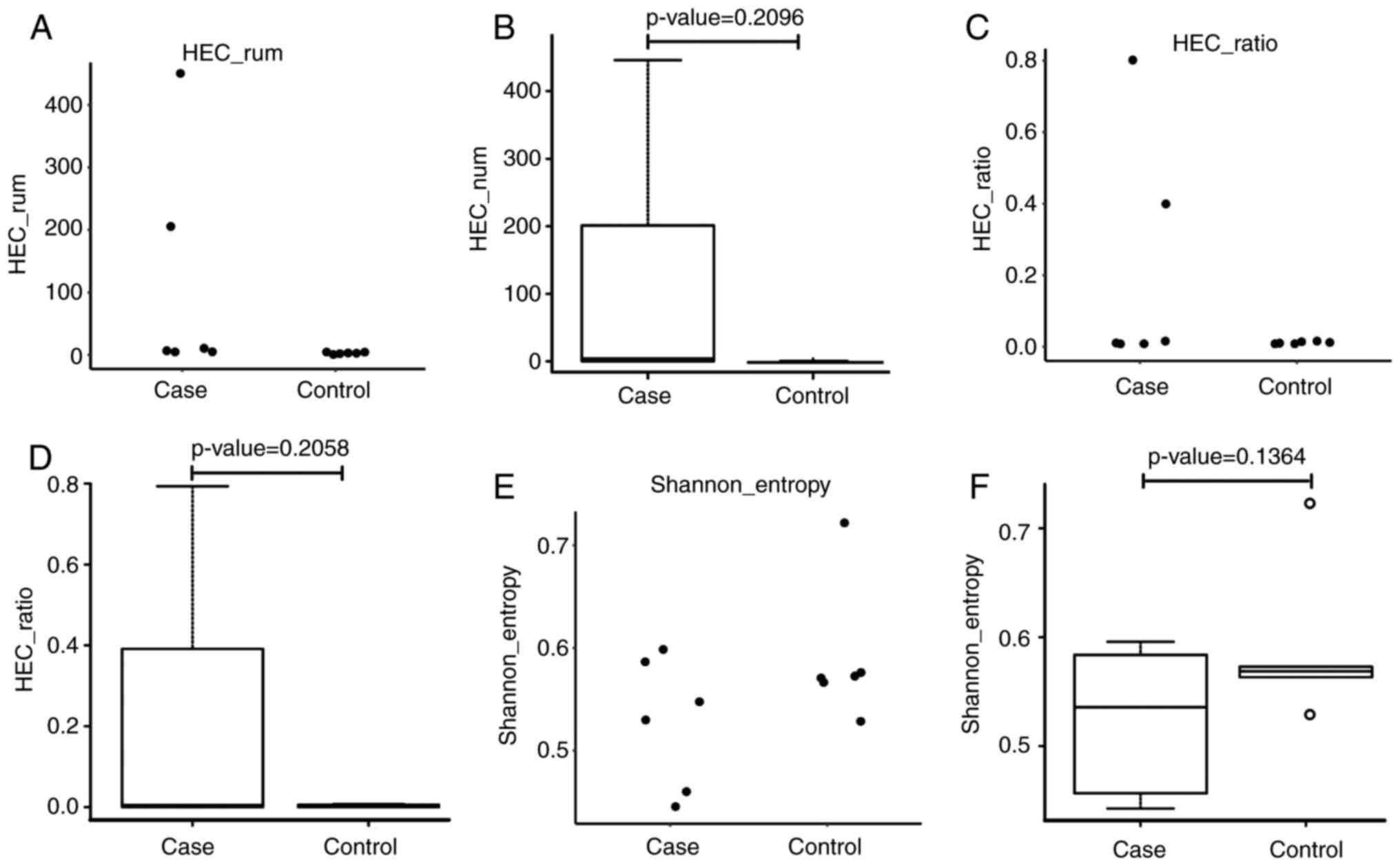
As shown in Fig. 1A-F, immune
diversity was marginally higher in the ACLF group, when compared
with that in the control subjects, although this difference was not
statistically significant (P=0.1364). Subsequently, the HEC in
these two groups were examined, and it was found that the HEC was
higher in the ACLF group than in the control group, indicating the
amplification of abnormal CDR3 sequences. Although differences
between these two groups were observed, there was no statistical
significance, which was possibly due to the relatively small sample
size used in the present study. In comparing the Shannon entropy of
the ACLF group with the control group, it was found that the
Shannon entropy value distribution of the patients with ACLF was
scattered, whereas the value distribution of control subjects was
relatively concentrated. As shown in Fig. 1A and B, the Shannon entropy
distribution of patients with ACLF was substantially skewed,
whereas the Shannon entropy of the control subjects presented with
a normal distribution. Although different Shannon entropy
distributions were identified in these two groups, the difference
was not statistically significant (P=0.2096). The CDR3 HEC was
higher in the ACLF patient group, compared with that in the control
subjects, however, the difference between these two groups was not
statistically significant.
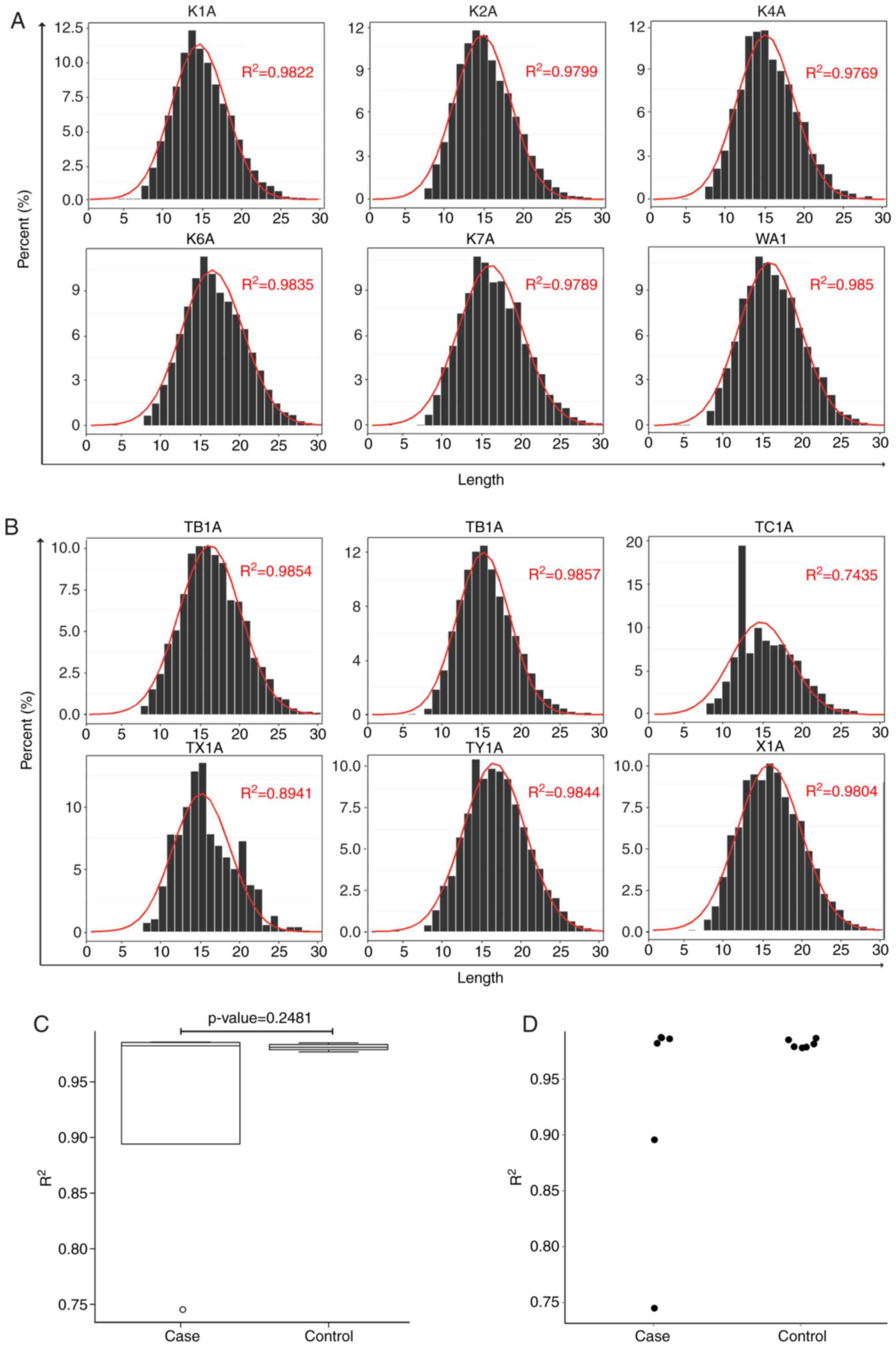
Analysis of CDR3 length distribution
between patients with ACLF and control subjects
It is well-recognized that CDR3 length distributions
in normal subjects are approximately normally distributed. In the
present study, CDR3 length distributions were analyzed in patients
with ACLF (X1A, TX-1A,TY-1A,TA-1A,TB-1A and TC-1A) and control
subjects (K1A, K2A, K4A, K6A, K7A and W1A). The results are shown
in Fig. 2A and B. The CDR3 normal
length distribution of patients with ACLF was compared with that of
control subjects. Through Gaussian fitting, the normal CDR3 length
distribution of each sample was obtained. Furthermore, the normal
distribution between the ACLF and control groups was compared, and
it was found that the CDR3 length distribution of control subjects
tended to be more normally distributed, when compared with that of
patients with ACLF (Fig. 2C and
D).
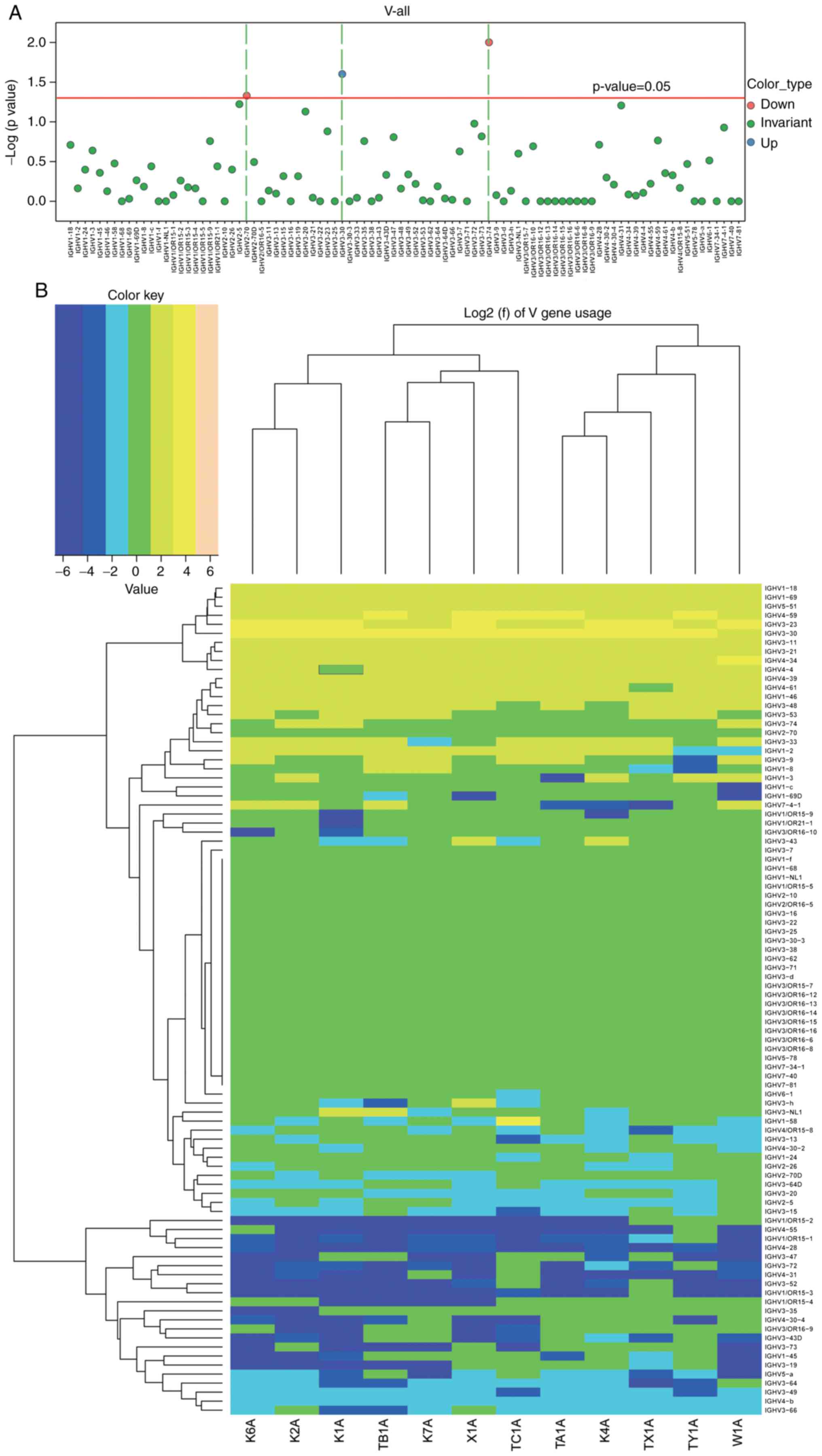
Comparison of usage frequency of the V,
D, and J gene segments in the BCR heavy chain
In order to compare the usage frequency of the V, D
and J gene segments in the BCR heavy chain, a usage frequency
distribution histogram and hierarchical clustering heat map were
generated according to each usage frequency in the ACLF and control
groups. The differential expression, including the upregulation and
downregulation of the individual gene fragments, were then
identified in the ACLF group, and compared with the control group.
A t-test was used to analyze the usage frequency and distribution
ratio of the V, D, J and V-J genes in the six patients with ACLF
(X1A, TX-1A, TY-1A, TA-1A, TB-1A and TC-1A) and six control
subjects (K1A, K2A, K4A, K6A, K7A and W1A). It was found that the
usage of the IGHV gene, which is a well-accepted assessment for BCR
repertoire structure, exhibited lower frequencies in the majority
or all IGHV genes in the control subjects, when compared with that
in the patients with ACLF. Furthermore, these clonal populations
had significantly higher frequencies of a single IGHV gene or group
of IGHV genes (12). Based on the
usage frequency distribution histogram of the V gene of the BCR
heavy chain and clustering heat map (Fig. 3A and B), and the results obtained
from the t-test for the distribution ratio of the V gene of
patients with ACLF and control subjects (Fig. 3; Table II), it was found that the
IGHV3-30 gene was significantly upregulated (P<0.05), whereas
the IGHV3-74 and IGHV2-70 genes were significantly downregulated
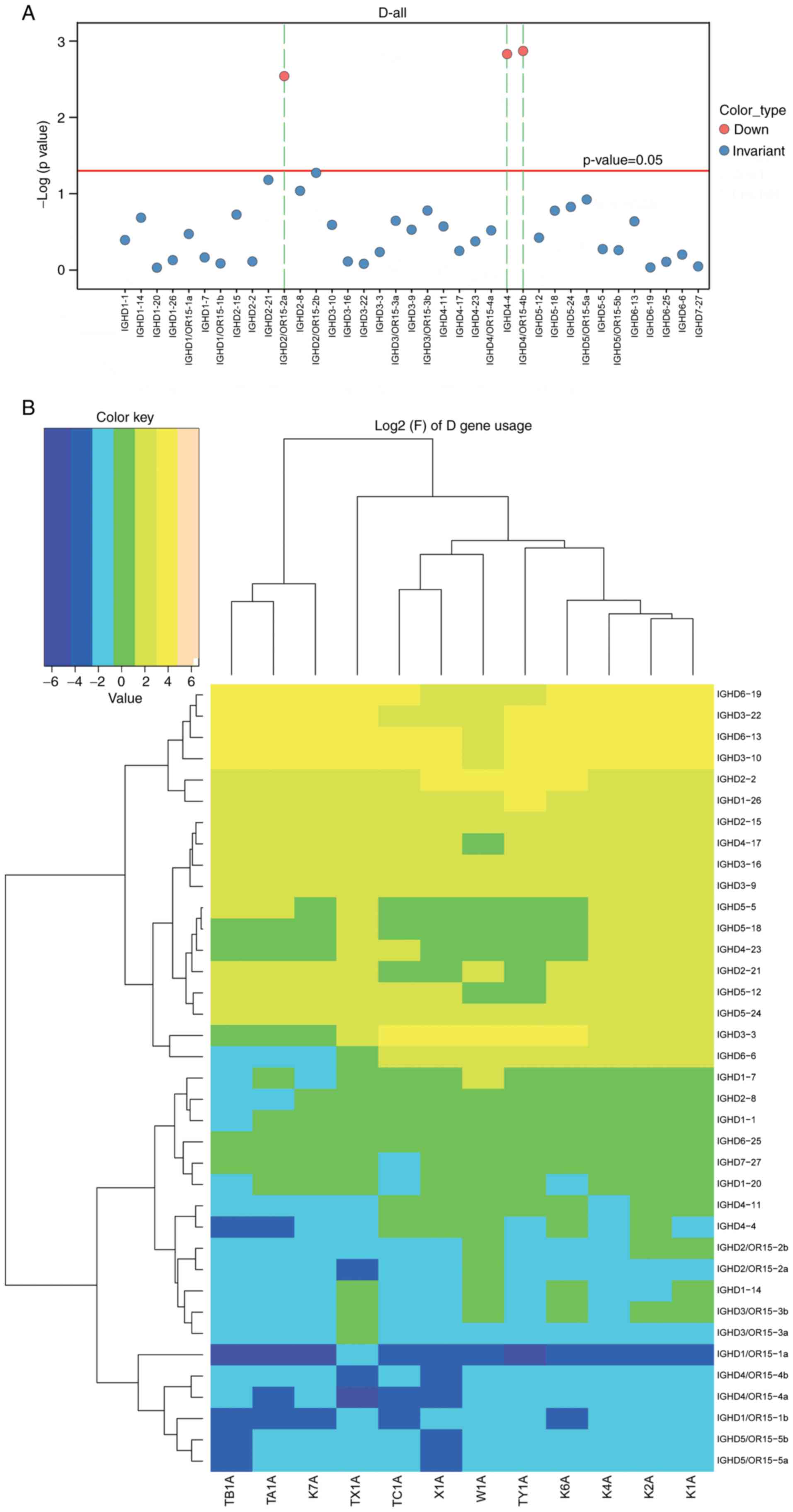
(P<0.05). Furthermore, a distribution histogram of the D region
usage frequency of the BCR heavy chain and the D sub-genotype of
each frequency clustering heat map were generated (Fig. 4A and B). The t-test for the
distribution ratio of the D gene in the six patients with ACLF and
six control subjects allowed for the identification of the
differential gene expression between these two groups (Fig. 4; Table III). As a result, three
significantly downregulated genes, IGHD4/OR15-4b, IGHD4/OR15-4a and
IGHD2-8, were identified in the ACLF group, compared with the
control group (P<0.05). Similarly, a distribution histogram of
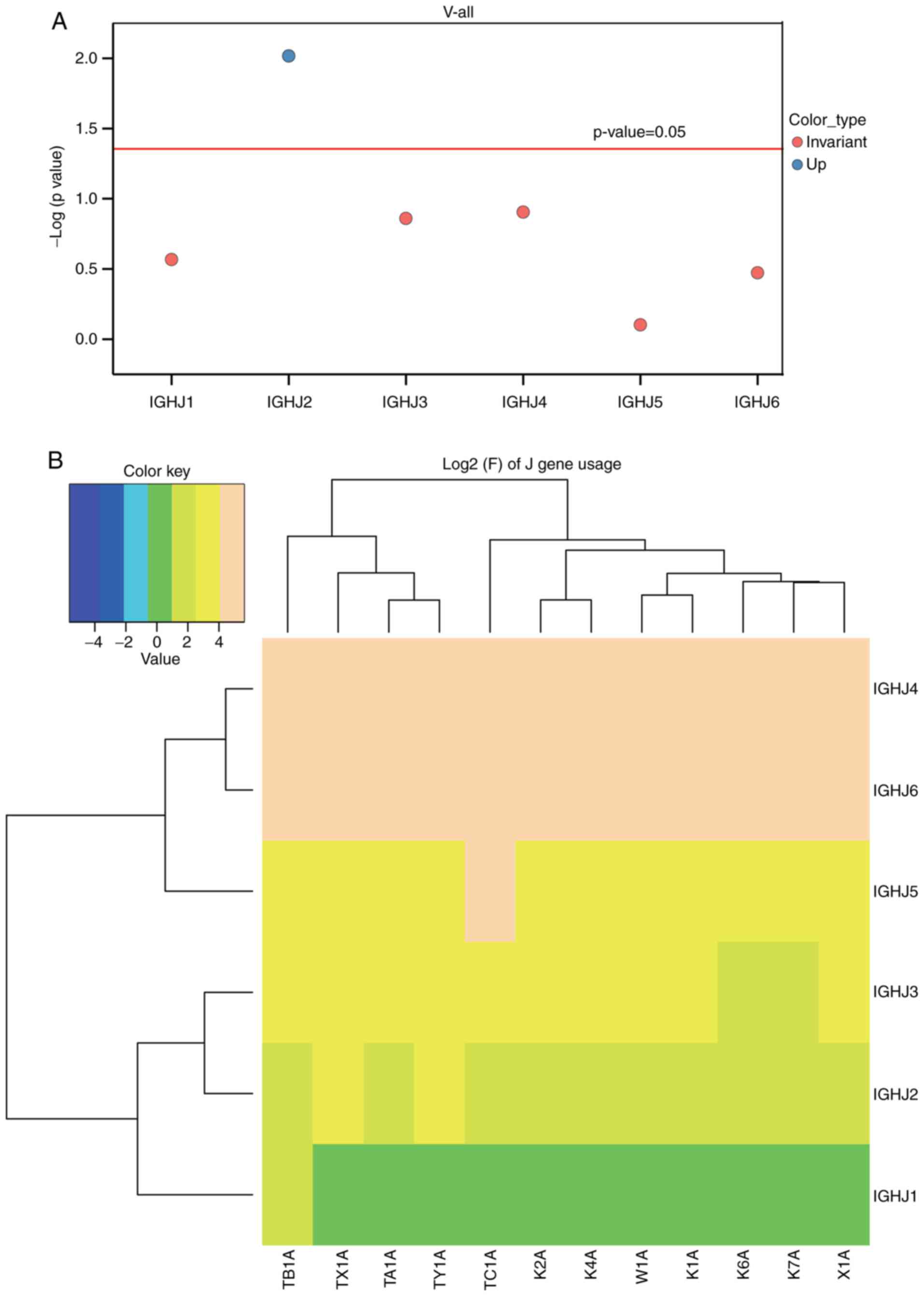
the J region usage frequency of BCR heavy chain and the J
sub-genotype of each frequency clustering heat-map was established
(Fig. 5A and B). At-test was then
performed for the distribution ratio of the J gene of the six
patients with ACLF and six control subjects (Fig. 5, Table IV). The IGHJ2 gene was identified
to be significantly upregulated in the ACLF group, when compared
with the control group (P<0.05), whereas the IGHJ1, IGHJ5,
IGHJ3, IGHJ4 and IGHJ6 genes remained statistically unchanged
between these two groups.
| Table IIResults of the t-test for the
distribution ratio of the V gene. |
Table II
Results of the t-test for the
distribution ratio of the V gene.
| Type | Control | Case | P-value | T-value |
|---|
| IGHV3-74 | 2.191677 | 1.526667 | 0.009973 | −0.26749 |
| IGHV2-70 | 1.486667 | 1.266667 | 0.046805 | −2.26805 |
| IGHV3-30 | 8.946667 | 11.93333 | 0.024998 | 2.645089 |
| Table IIIResults of the t-test for the
distribution ratio of the D gene. |
Table III
Results of the t-test for the
distribution ratio of the D gene.
| Type | Control | Case | P-value | T-value |
|---|
| IGHD4/OR15-4b | 0.35 | 0.158333 | 0.001349 | −5.11743 |
| IGHD4/OR15-4a | 0.293333 | 0.118333 | 0.001483 | −4.87766 |
| IGHD2-8 | 1.393333 | 0.731667 | 0.002883 | −3.929 |
| Table IVResults of the t-test for the
distribution ratio of the J gene. |
Table IV
Results of the t-test for the
distribution ratio of the J gene.
| Type | Control | Case | P-value | T-value |
|---|
| IGHJ2 | 2.776667 | 4.56 | 0.009614 | 3.624858 |
| IGHJ4 | 45.35 | 39.975 | 0.124539 | −1.72416 |
| IGHJ3 | 6.173333 | 7.073333 | 0.137946 | 1.661918 |
| IGHJ1 | 1.206667 | 1.495 | 0.270726 | 1.197279 |
| IGHJ6 | 31.35833 | 34.225 | 0.337043 | 1.008973 |
| IGHJ5 | 13.13167 | 12.66667 | 0.79099 | −0.27929 |
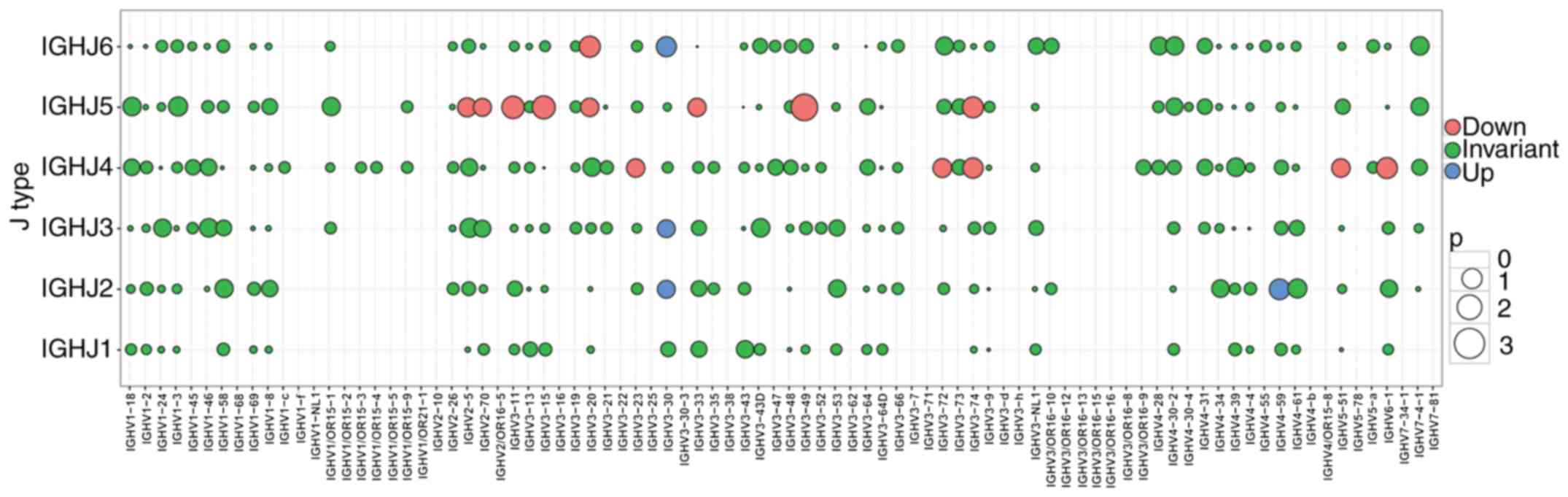
Subsequently, a bubble chart for the expression of
the combinations of each V and J gene segment was established
(Fig. 6), and different colors
were used for different genes. The differential composition of the
V-J combinations in the patients with ACLF (X1A, TX-1A, TY-1A,
TA-1A, TB-1A and TC-1A) were then compared with those in the
control subjects (K1A, K2A, K4A, K6A, K7A and W1A). Subsequently, a
t-test for the distribution ratio of the combinations of the six
patients with ACLF and six control subjects was performed, and the
negative P-values were plotted (Fig.
6; Table V). As observed in
the alterations in gene expression, the upregulated genes are
denoted in blue, downregulated genes are denoted in red, and
non-significant change combinations are denoted in green (Fig. 6). Furthermore, four significantly
upregulated combination genes were found, including (IGHV3-30,
IGHJ2),(IGHV3-30, IGHJ3), (IGHV3-30,IGHJ6) and (IGHV4-59,IGHJ2)
(P<0.05), whereas 14 combination genes were identified to be
significantly downregulated, including (IGHV3-49,IGHJ5),
(IGHV3-15,IGHJ5), (IGHV3-11,IGHJ5), (IGHV3-20,IGHJ6),
(IGHV3-74,IGHJ5), (IGHV6-1,IGHJ4), (IGHV3-74,IGHJ4),
(IGHV2-5,IGHJ5), (IGHV3-72,IGHJ4), (IGHV5-51,IGHJ4),
(IGHV3-23,IGHJ4), (IGHV3-20,IGHJ5), (IGHV3-33,IGHJ5) and
(IGHV2-70,IGHJ5) (P<0.05).
| Table VResults of the t-test for the
distribution ratio of the V-J gene combinations. |
Table V
Results of the t-test for the
distribution ratio of the V-J gene combinations.
| V-type | J-type | Control | Case | P-value | T-value |
|---|
| IGHV3-49 | IGHJ5 | 0.026666 | 0.006666 | 0.000815 | −5.070925 |
| IGHV3-15 | IGHJ5 | 0.0716666 | 0.025 | 0.005040 | −3.838859 |
| IGHV3-11 | IGHJ5 | 1.3766666 | 0.995 | 0.00639811 | −3.6823573 |
| IGHV6-1 | IGHJ4 | 0.446666 | 0.278333 | 0.012146 | −3.235252 |
| IGHV3-20 | IGHJ6 | 0.1933333 | 0.63333 | 0.0118824 | −3.0769355 |
| IGHV3-74 | IGHJ5 | 0.3833333 | 0.193333 | 0.0119918 | −3.062569 |
| IGHV3-74 | IGHJ4 | 1.0833333 | 0.6483333 | 0.0158769 | −2.9282990 |
| IGHV3-33 | IGHJ5 | 0.1466666 | 0.045 | 0.035139 | −2.747301 |
| IGHV3-20 | IGHJ5 | 0.0833333 | 0.03 | 0.034568 | −2.651974 |
| IGHV2-5 | IGHJ5 | 0.1183333 | 0.058333 | 0.026997 | −2.617230 |
| IGHV3-72 | IGHJ4 | 0.05 | 0.013333 | 0.028606 | −2.607250 |
| IGHV3-23 | IGHJ4 | 4.5783333 | 3.253333 | 0.033451 | −2.52706 |
| IGHV5-51 | IGHJ4 | 2.0316666 | 1.346666 | 0.032790 | −2.515948 |
| IGHV2-70 | IGHJ5 | 0.3716666 | 0.21 | 0.039841 | −2.362483 |
| IGHV3-30 | IGHJ3 | 0.5366666 | 0.743333 | 0.041738 | 2.340706 |
| IGHV3-30 | IGHJ2 | 0.1733333 | 0.483333 | 0.041722 | 2.637409 |
| IGHV3-30 | IGHJ6 | 2.78 | 4.225 | 0.017769 | 2.879356 |
| IGHV4-59 | IGHJ2 | 0.2283 | 0.52 | 0.014176 | 3.497063 |
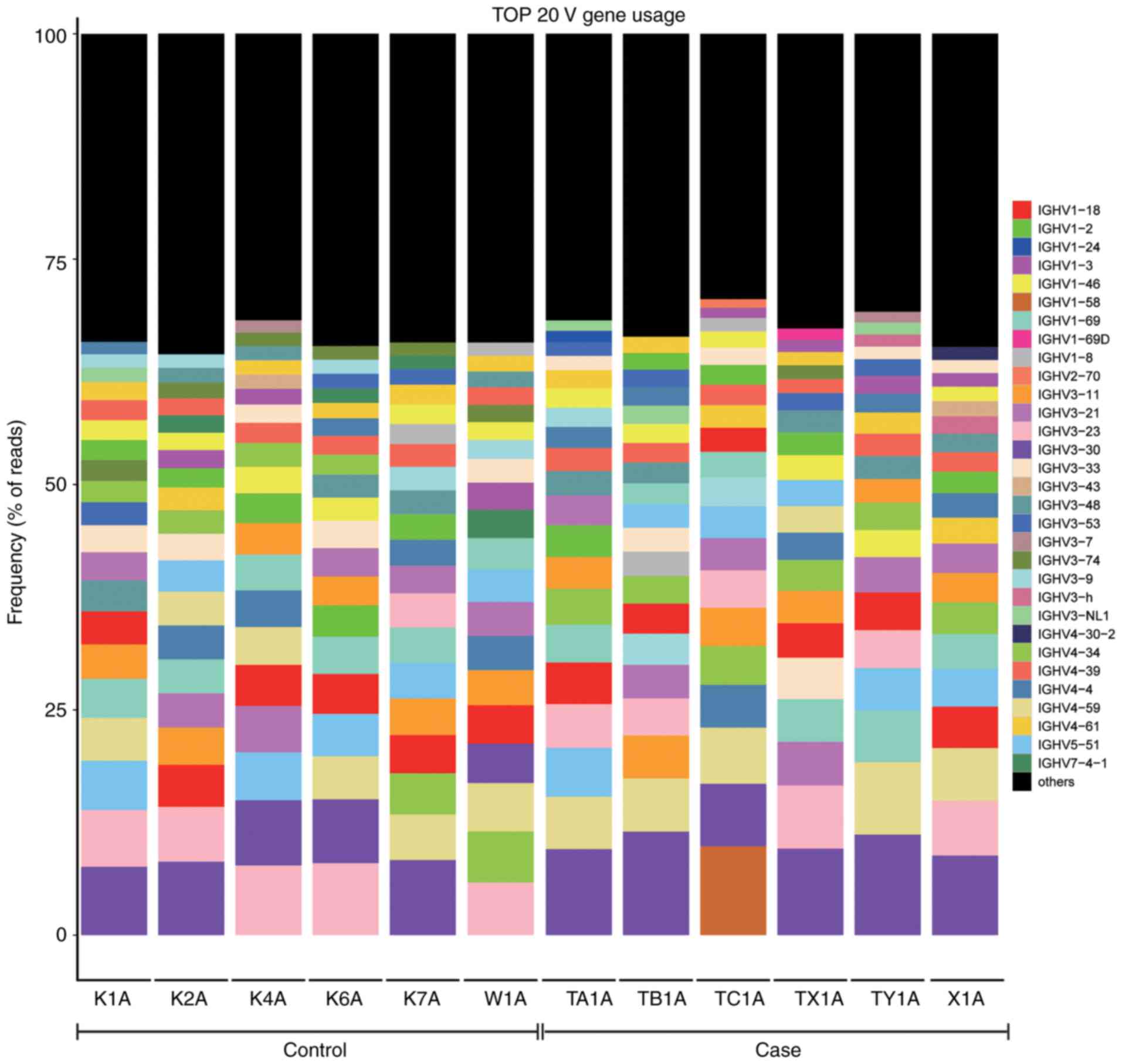
Comparison of the top 20 BCR heavy chain
V region genes used between the ACLF and control groups
The top 20 frequently used genes of the BCR heavy
chain V subtypes are listed in Fig.
7, in which the percentage of each sample V subtype in the top
20 frequently used genesis shown.
CDR3amino acid sequences commonly used in
the ACLF and control groups
In the present study, three amino acid sequences
commonly used in all patients with ACLF (n=6) and control subjects
(n=6) were found, which were AR, FDY and MDV. The nine cloned
numbers for patients with ACLF in the CDR3amino acid sequences are
listed in Table VI, and the
total number of CDR3 clones are shown in the last row. In addition,
the three cloned numbers for control subjects in the CDR3amino acid
sequences are shown in Table
VII, and the total number of all CDR3 clones are shown in the
last row.
| Table VINumber of CDR3 amino acid sequences
for the clones of patients with ACLF. |
Table VI
Number of CDR3 amino acid sequences
for the clones of patients with ACLF.
| CDR3 | TA1A | TB1A | TC1A | TX1A | TY1A | X1A | |
|---|
| AR |
2.08×10−5 |
6.54×10−6 |
5.17×10−7 |
1.04×10−4 |
9.01×10−6 |
1.02×10−5 |
| DY |
3.97×10−6 |
6.96×10−6 |
2.72×10−6 |
2.28×10−7 |
4.12×10−6 |
6.84×10−6 |
| FDY |
3.34×10−6 |
3.69×10−6 |
7.76×10−7 |
1.14×10−7 |
5.88×10−7 |
1.76×10−6 |
| GMDV |
6.91×10−6 |
3.38×10−6 |
9.05×10−7 |
2.39×10−6 |
4.21×10−6 |
1.09×10−5 |
| LDV |
7.95×10−8 |
3.16×10−7 |
9.05×10−7 |
1.03×10−6 |
4.90×10−7 |
3.11×10−7 |
| MDV |
5.48×10−6 |
1.48×10−6 |
3.49×10−6 |
8.89×10−6 |
3.23×10−6 |
6.94×10−6 |
| YGMDV |
1.11×10−6 |
3.06×10−6 |
1.81×10−6 |
1.25×10−6 |
2.06×10−6 |
2.69×10−6 |
| YYGMDV |
1.19×10−6 |
7.38×10−7 |
7.76×10−7 |
1.03×10−6 |
1.27×10−6 |
3.11×10−7 |
| YYYGMDV |
1.59×10−7 |
8.44×10−7 |
1.29×10−7 |
5.70×10−7 |
2.74×10−6 |
2.07×10−7 |
| Total | 12,584,061 | 9,479,612 | 7,733,592 | 8,771,599 | 10,206,099 | 9,651,145 |
| Table VIINumber of CDR3 amino acid sequences
for the clones of control subjects. |
Table VII
Number of CDR3 amino acid sequences
for the clones of control subjects.
| CDR3 | K1A | K2A | K4A | K6A | K7A | W1A |
|---|
| AR |
2.08×10−5 |
1.12×10−5 |
9.15×10−6 |
4.55×10−6 |
6.79×10−6 |
2.84×10−7 |
| FDY |
3.34×10−6 |
3.02×10−6 |
6.51×10−6 |
6.82×10−6 |
1.44×10−6 |
1.13×10−6 |
| MDV |
5.48×10−6 |
4.90×10−6 |
1.23×10−6 |
2.27×10−6 |
1.02×10−6 |
1.38×10−5 |
| Total | 12,584,061 | 12,249,709 | 10,600,111 | 439,959 | 11,778,081 | 10,575,248 |
Discussion
ACLF has been recognized as a distinct syndrome from
AD, and the development of ACLF has been found to occur under the
setting of systematic inflammation (2). However, to date, the mechanism
underlying the development and progression of ACLF remains to be
fully elucidated (13). In
particular, the role of the immune system remains unclear. In the
present study, NGS was used to characterize the polymorphisms of
the BCR CDR3 gene segments in PBMCs prepared from patients with
HBV-ACLF (n=6) and control subjects (n=6). The main novel findings
were follows: i) Patients with ACLF exhibited a significantly
higher extent of clonal expansion for B-cells, when compared with
control subjects; ii) patients with ACLF presented with a skewed
usage of BCR CDR3 V, D, J and V-J gene segments, whereas control
subjects had a close-to-normal distribution; iii) the BCR heavy
chain CDR3 of the V, D, J and V-J combinations of subfamily genes
were abnormally expressed in response to ACLF; and iv) there was a
significant association between the BCR CDR3 repertoire and
ACLF.
It has been reported that a large proportion of
B-cell clones in the BCR sequence are at lower frequencies, and
these clones indicate its inability for clonal expansion (14). In the present study, certain HECs
were identified in each individual BCR heavy chain CDR3, and these
clones were able to form in a complex environment with antigen or
pathogen stimulation. Furthermore, clonal expansion was observed in
patients with ACLF and control subjects, and a higher extent of
clonal expansion was present in the ACLF group, when compared with
the control group.
Extensive studies have shown that the V, D and J
gene segments of BCRs are rearranged in the development of
B-lymphocytes, during which different numbers of nucleotides can be
randomly inserted into or deleted from the V-J gene segments or
V-D-J gene segments. Therefore, a variable CDR3 region is formed
with diversity in its length and sequence, in which the CDR3
sequence is able to determine a unique BCR clone type, allowing for
the assessment of the clonal diversity of B-lymphocytes through the
length of CDR3 (15,16). In the present study, the normal
distribution of CDR3 lengths was compared between patients with
ACLF and control subjects. The head of the distribution tended to
be skewed in patients with ACLF, whereas the distribution was close
to normal in control subjects; the difference between these two
groups was not statistically significant. In the comparative
analysis of Shannon entropy and HEC between patients with ACLF and
control subjects, there were differences between these two groups
with respect to number and proportion. However, the difference was
not statistically significant, which was possibly due to the small
sample size. A Student’s t-test was used to analyze the frequency
usage of the BCR heavy chain V, D, J and V-J segments, and the
following six upregulated genes were identified: IGHV3-30, IGHJ2,
(IGHV3-30, IGHJ2), (IGHV3-30, IGHJ3), (IGHV3-30, IGHJ6) and
(IGHV4-59, IGHJ2). Furthermore, 19 downregulated genes were
identified: IGHV3-74, IGHV2-70, IGHD4/OR15-4b, IGHD4/OR15-4a,
IGHD2-8, (IGHV3-49, IGHJ5), (IGHV3-15, IGHJ5), (IGHV3-11, IGHJ5),
(IGHV3-20, IGHJ6), (IGHV3-74, IGHJ5), (IGHV6-1, IGHJ4), (IGHV3-74,
IGHJ4), (IGHV2-5, IGHJ5), (IGHV3-72, IGHJ4), (IGHV5-51, IGHJ4),
(IGHV3-23, IGHJ4), (IGHV3-20, IGHJ5), (IGHV3-33, IGHJ5) and
(IGHV2-70, IGHJ5). The identified dysregulation of the BCR heavy
chain CDR3 V, D, J and V-J combinations of the sub-family gene may
be involved in the development and progression of ACLF. It was
hypothesized that the upregulated genes IGHV3-30,IGHJ2, (IGHV3-30,
IGHJ2), (IGHV3-30, IGHJ3), (IGHV3-30, IGHJ6) and (IGHV4-59, IGHJ2)
are involved in certain BCR-specific clonal proliferations of
B-lymphocytes and undermine BCR diversity. This has been shown to
be essential in maintaining the normal immune system in control
subjects. The more subtypes of immune proteins present, the more
effective the immune system against various pathogens. By contrast,
the lower the number of subtypes an immune protein has, the more
the individual becomes susceptible to diseases. In the present
study, the downregulated genes IGHV3-74, IGHV2-70, IGHD4/OR15-4b,
IGHD4/OR15-4a, IGHD2-8, (IGHV3-49, IGHJ5), (IGHV3-15, IGHJ5),
(IGHV3-11, IGHJ5), (IGHV3-20, IGHJ6), (IGHV3-74, IGHJ5), (IGHV6-1,
IGHJ4), (IGHV3-74, IGHJ4), (IGHV2-5, IGHJ5), (IGHV3-72, IGHJ4),
(IGHV5-51, IGHJ4), (IGHV3-23, IGHJ4), (IGHV3-20, IGHJ5), (IGHV3-33,
IGHJ5) and (IGHV2-70, IGHJ5) may be associated with inhibition of
the clonal proliferation of certain specific B lymphocytes of BCRs.
It is noteworthy that a number of the above subtypes were
significantly reduced in the patients with ACLF, compared with the
control subjects, indicating that B-lymphocytes were significantly
reduced inpatients with ACLF.
In conclusion, an assay to determine the entire
diversity of the B-cell repertoire at a sequence-level resolution
in patients with HBV-ACLF was successfully developed. The findings
suggested that the development of HBV-ACLF is linked with the BCR
heavy chain CDR3 repertoire. Future insightful investigations are
required to better understand the role of the BCR repertoire in
immune responses, autoimmunity and alloreactivity, and in driving
the development and progression of HBV-ACLF.
Acknowledgments
My deepest gratitude goes first and foremost to
Professor Weiguo Sui, my supervisor, for his constant encouragement
and guidance. He has walked me through all the stages of the
writing of this thesis. Without his consistent and illuminating
instruction, this thesis could not have reached its present form.
Second, I would like to express my heartfelt gratitude to Professor
Yong Dai, who instructed and helped me a lot in the past four
years. Last my thanks would go to my beloved family for their
loving considerations and great confidence in me all through these
years. I also owe my sincere gratitude to my friends who gave me
their help and time in listening to me and helping me work out my
problems during the difficult course of the thesis.
Funding
This study was supported by the Technology Plan of
Shenzhen, Guangdong (grant no. JCYJ20160422150329190), the China
Postdoctoral Science Foundation (grant no. 2017M610575) and the
Guilin Scientific Research and Technology Development Program
(grant no. 20170117-1).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors’ contributions
WS and YD conceived and designed the study. QY
conducted the experiments. LL, SL and HC participated in the
completion of the experiments. QY and LW analyzed the data. LW
wrote the manuscript. WS and JZ revised the manuscript. All the
authors read and approved the final paper.
Ethics approval and consent to
participate
Written informed consent was obtained from each
participant or from their guardians. The present study complies
with the Declaration of Helsinki for investigations involving
humans, and the study protocol was carefully reviewed and approved
by the Ethics Committee of the No. 181 Hospital of Guilin (Guilin,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jalan R, Gines P, Olson JC, Mookerjee RP,
Moreau R, Garcia-Tsao G, Arroyo V and Kamath PS: Acute-on chronic
liver failure. J Hepatol. 57:1336–1348. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Moreau R, Jalan R, Gines P, Pavesi M,
Angeli P, Cordoba J, Durand F, Gustot T, Saliba F, Domenicali M, et
al: Acute-on-chronic liver failure is a distinct syndrome that
develops in patients with acute decompensation of cirrhosis.
Gastroenterology. 144:1426–1437. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sarin SK, Kumar A, Almeida JA, Chawla YK,
Fan ST, Garg H, de Silva HJ, Hamid SS, Jalan R, Komolmit P, et al:
Acute-on-chronic liver failure: Consensus recommendations of the
Asian Pacific Association for the study of the liver (APASL).
Hepatol Int. 3:269–282. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Olson JC and Kamath PS: Acute-on-chronic
liver failure: Concept, natural history, and prognosis. Curr Opin
Crit Care. 17:165–169. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Arstila TP, Casrouge A, Baron V, Even J,
Kanellopoulos J and Kourilsky P: A direct estimate of the human
alphabeta T cell receptor diversity. Science. 286:958–961. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schroeder HW Jr and Cavacini L: Structure
and function of immunoglobulins. The J Allergy Clin Immunol.
125:S41–S52. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tonegawa S: Somatic generation of antibody
diversity. Nature. 302:575–581. 1983. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nouri-Aria KT, Alexander GJ, Portmann BC,
Hegarty JE, Eddleston AL and Williams R: T and B cell function in
alcoholic liver disease. J Hepatol. 2:195–207. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sarin SK, Kedarisetty CK, Abbas Z,
Amarapurkar D, Bihari C, Chan AC, Chawla YK, Dokmeci AK, Garg H,
Ghazinyan H, et al: Acute-on-chronic liver failure: consensus
recommendations of the Asian Pacific Association for the Study of
the Liver (APASL) 2014. Hepatol Int. 8:453–471. 2014. View Article : Google Scholar
|
|
10
|
Londoño JC and Philipp SE: A reliable
method for quantification of splice variants using RT-qPCR. BMC Mol
Biol. 17:82016. View Article : Google Scholar
|
|
11
|
Venturi V, Kedzierska K, Turner SJ,
Doherty PC and Davenport MP: Methods for comparing the diversity of
samples of the T cell receptor repertoire. J Immunol Methods.
321:182–195. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Keylok CJ: Simpson diversity and the
Shannon-Wiener index as special cases of a generalized entropy.
Oikos. 109:203–207. 2005. View Article : Google Scholar
|
|
13
|
Wang FS and Zhang Z: Host immunity
influences disease progression and antiviral efficacy in humans
infected with hepatitis B virus. Expert Rev Gastroenterol Hepatol.
3:499–512. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu YC, Kipling D, Leong HS, Martin V,
Ademokun AA and Dunn-Walters DK: High-throughput immunoglobulin
repertoire analysis distinguishes between human IgM memory and
switched memory B-cell populations. Blood. 116:1070–1078. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Honjo T: Immunoglobulin Genes. Annu Rev
Immunol. 1:499–528. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Foreman AL, Van de Water J, Gougeon ML and
Gershwin ME: B cells in autoimmune diseases: Insights from analyses
of immunoglobulin variable (Ig V) gene usage. Autoimmun Rev.
6:387–401. 2007. View Article : Google Scholar : PubMed/NCBI
|