Introduction
Ovarian cancer (OC) is the most common malignant
tumor threatening the health of women in China. In 2015, an
estimated 52,100 incident cases were diagnosed, with >22,500
mortalities (1). Although
patients with OC are generally treated with a series of standard
treatments, including surgery, chemotherapy and radiotherapy, the
prognosis remains unsatisfactory for the majority of patients, with
an overall 5-year survival rate of ~30% (2). Additionally, advanced OC may only be
observed at the point of the initial diagnosis, which may be too
late for patients to receive optimal treatment. Therefore, a screen
of effective diagnostic biomarkers and the development of novel
therapeutic strategies are urgently required.
Based on previous evidence, microRNAs (miRNAs/miRs)
serve important roles in the development and progression of OC by
specifically binding to the 3′-untranslated region (3′-UTR) of
target mRNAs to inhibit translation or induce degradation (3). For example, Qin et al
(4) observed a significant
decrease in the expression of miR-152 in OC specimens and three OC
cell lines. The transfection of miR-152 mimic into OC cells
inhibited proliferation and migration by targeting the 3′-UTR of
the forkhead box protein 1 (FOXP1) and resulted in improved overall
survival. Liu et al (5)
confirmed the over-expression of miR-26a in human OC specimens and
revealed correlations between lymph node metastasis, an advanced
International Federation of Gynecology and Obstetrics (FIGO) stage
and poor survival. miR-216a promoted the metastatic behaviors and
epithelial-mesenchymal transition (EMT) of OC cells by inhibiting
its direct downstream target phosphatase and tensin homolog and
subsequently regulating the protein kinase B pathway. Therefore,
miRNAs may serve as promising biomarkers for the diagnosis of OC
and attractive therapeutic targets.
In addition to their intracellular functions, miRNAs
are also packaged into membrane-bound vesicles, including exosomes,
and released to the extracellular environment to facilitate
tumorigenesis and progression (6). For example, in the study by Ying
et al (7), OC-derived
exosomes released miR-222-3p into macrophages and induced the
formation of a tumor-associated macrophage-like phenotype by
decreasing the expression of suppressor of cytokine signaling 3
(SOCS3) and activating SOCS3/signal transducer and activator of
transcription (STAT)-3 signaling pathway, thereby promoting the
growth and metastasis of OC. According to the study by Kanlikilicer
et al (8), miR-6126 is
released from OC cells via exosomes. miR-6126 significantly reduces
tumor growth, invasion and migration in vitro and in
vivo by suppressing the expression of integrin-β1. De et
al (9) observed the
anti-neoplastic effects of an Emblica officinalis extract on
OC that were mediated by the upregulation of cellular and exosomal
miR-375 expression. Meng et al (10) observed correlations between high
levels of exosomal miR-373, miR-200a, miR-200b and miR-200c
expression with a higher FIGO stage (III-IV) and a shorter overall
survival. Therefore, exosomal miRNAs may also be crucial targets
for cancer diagnosis and treatment. Strategies targeting
co-expressed miRNAs in cancer cells and their exosomes may
represent more effective approaches for inducing cancer remission
(11), however, this information
has been rarely reported in OC.
The present study aimed to investigate the shared
miRNAs in OC cells and their exosomes using microarray data
downloaded from Gene Expression Omnibus (GEO) database and validate
their clinical significance with The Cancer Genome Atlas (TCGA)
datasets. The present study preliminarily identified that
downregulated hsa-miR-145 was common in OC and OC-derived exosomes
and thus may be a crucial biomarker for the diagnosis and treatment
of OC.
Materials and methods
Data collection
The miRNA microarray data (accession number
GSE103708) were downloaded from the GEO database (http://www.ncbi.nlm.nih.gov/geo/) on January 23,
2018; the dataset contained exosomal samples isolated from 13 human
epithelial OC cell lines (A2780, ES-2, CAOV3, SKOV3, OV-90, OAW42,
MCAS, COV362, RMG-1, RMUG-S, KURAMOCHI, NIH-OVCAR3 and A2780cis), 3
normal ovarian surface epithelial cell lines (HOSE1, HOSE2 and
HOSE3) and their original cell samples. Each cell type had one
biological repeat, resulting in 32 samples.
Data preprocessing and identification of
differentially expressed miRNAs (DE-miRNAs)
The GSE Series Matrix and annotations files were
retrieved from the Agilent-046064
Unrestricted_Human_miRNA_V19.0_Microarray Array platform GPL18402.
The probe names were transferred to gene symbols based on the
platform annotation information. When multiple probes were mapped
to a given gene, the mean expression value was used. Data were then
log2-transformed and quantile normalized using the Robust
Multiarray Average function available in the Bioconductor R package
(v3.32.5; http://www.biocon-ductor.org/packages/release/bioc/html/limma.html).
The DE-miRNAs between OC and normal ovarian surface epithelial
cells and exosomes and original cells were identified using the
Linear Models for Microarray data method in the Bioconductor R
package (v3.32.5; http://www.bioconductor.org/packages/release/bioc/html/limma.html)
(12). The significance of
differences of DE-miRNAs were assessed using the empirical Bayes
moderated t-test. Subsequently, the P-value was corrected for
multiple comparisons using the Benjamini-Hochberg (BH) procedure
(13). miRNAs with a false
discovery rate <0.05 and |logFC(fold change)|>1 were
considered differentially expressed. The ability of DE-miRNAs to
differentiate the samples was tested using the Euclidean
distance-based bidirectional hierarchical clustering analysis in
the Pheatmap package (v1.0.8; http://cran.r-project.org/web/packages/pheatmap/index.html),
following which a heat map was produced. A Venn diagram (http://bioinfor-matics.psb.ugent.be/webtools/Venn/)
was used to visualize the shared DE-miRNAs between different
exosomes and original cells.
Target gene prediction
The mRNA targets of DE-miRNAs were predicted using
the miRWalk database (v2.0; http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2/)
(14). Then, the miRNA-target
gene interaction network was constructed and visualized using
Cytoscape software (v3.4; www.cytoscape.org/) (15).
Screening prognosis-associated miRNAs and
their target genes
The miRNA and mRNA Seq data from OC samples (Level
3) were obtained from the TCGA database (https://tcga-data.nci.nih.gov/). The expression levels
of targeted miRNAs and mRNAs in OC were analyzed using the fragment
per kilobase per million mapped reads value from the TCGA data.
A univariate Cox regression analysis was performed
to screen prognosis-associated miRNAs/mRNAs using the survival
package in R (v2.4; https://cran.r-project.org/web/packages/survival/index.html)
(16), with the log-rank
P<0.05 set as the threshold. Univariate and multivariate Cox
regression analyses were used to screen prognosis-associated
clinical characteristics, including age, radiation therapy, stage
(17), histological grade
(18), neoplasm subdivision,
lymphatic invasion, recurrence and survival, and then the potential
associations between crucial miRNAs and clinical characteristics
were also calculated using the survival package. The Kaplan-Meier
curve with the log-rank test was plotted using GraphPad Prism
software (v5; GraphPad Software, Inc., La Jolla, CA, USA) to
determine significant associations between the expression of
miRNAs/mRNAs and patient survival outcomes. Pearson’s correlation
coefficients were calculated to assess the correlations between
miRNAs and mRNAs. P<0.05 was considered to indicate a
statistically significant difference.
Functional enrichment analysis
The enriched Kyoto Encyclopedia of Genes and Genomes
(KEGG) (19) pathways of target
genes were predicted by searching the Database for Annotation,
Visualization and Integrated Discovery online tool (v6.8;
http://david.abcc.ncifcrf.gov) (20). P<0.05 was set as the cut-off
criterion. In addition, all known OC-associated pathways were also
downloaded from the Comparative Toxicogenomics Database (CTD;
http://ctd.mdibl.org/) (21). The pathways obtained from these
two sources were overlapped to obtain crucial miRNAs involved in
OC-associated pathways.
Results
Identification of DE-miRNAs between
cancer and normal cells
In the exosomal samples, 77 miRNAs were
differentially expressed between 13 OC cancer and 3 normal cells,
including 37 upregulated and 40 downregulated miRNAs. For the
original cell samples, 259 DE-miRNAs were identified between 13 OC
cancer and 3 normal cells, including 87 upregulated and 172
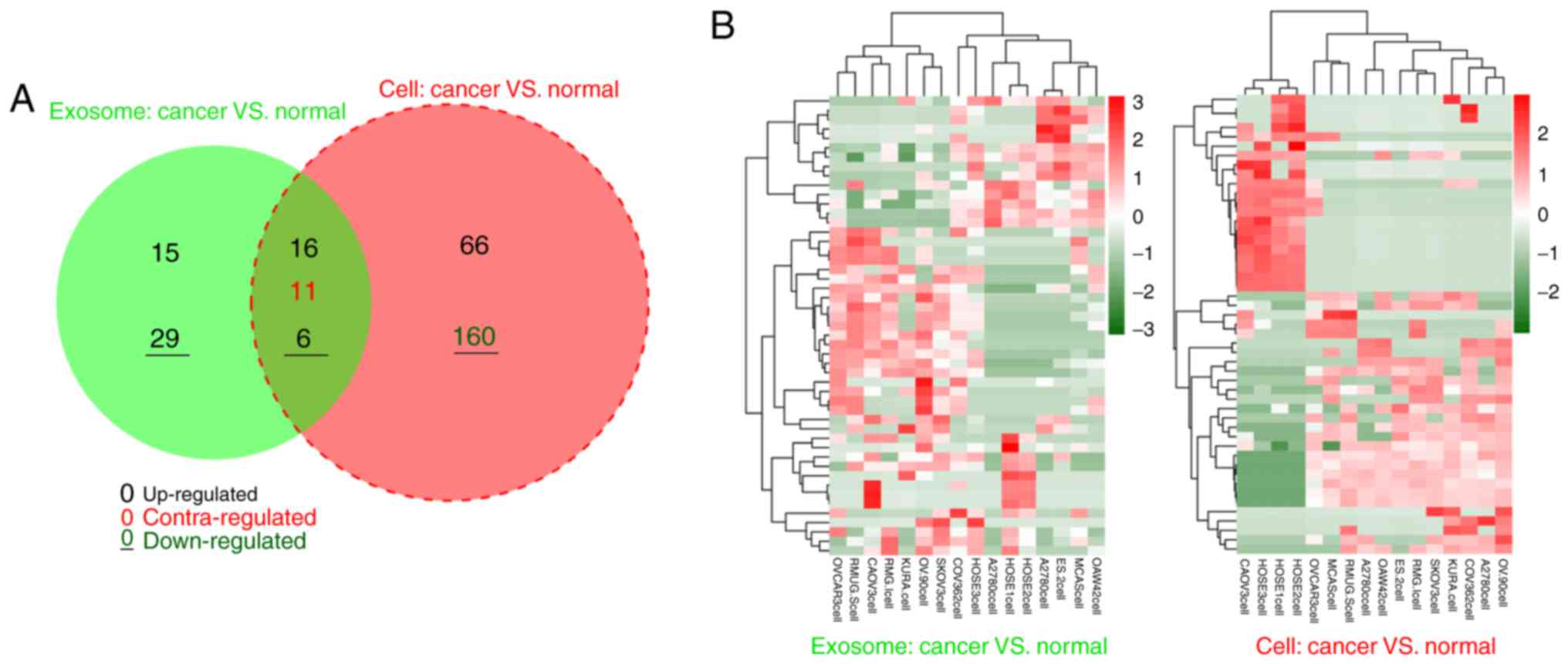
downregulated miRNAs. According to the Venn diagram, 22 DE-miRNAs
with similar expression trends (16 upregulated and 6 downregulated)
were shared between exosomal and original cell samples (Fig. 1; Table I), suggesting that these 22
DE-miRNAs may be particularly crucial for OC development in an
exosomal or non-exosomal manner. Therefore, these DE-miRNAs were
examined in subsequent analyses.
 | Table IShared differentially expressed
miRNAs in exosomes and original cells. |
Table I
Shared differentially expressed
miRNAs in exosomes and original cells.
| miRNA | Exosome
| Original cells
|
|---|
| logFC | P-value | FDR | logFC | P-value | FDR |
|---|
| hsa-miR-202-3p
- | 1.14 |
7.29×10−5 |
1.48×10−2 | −2.14 |
1.13×10−2 |
3.92×10−2 |
| hsa-miR-5684 | −1.73 |
5.76×10−5 |
1.17×10−2 | −1.58 |
1.94×10−3 |
8.36×10−3 |
| hsa-miR-376a-3p
- | 1.74 |
9.67×10−6 |
1.97×10−3 | −1.80 |
3.36×10−3 |
1.38×10−2 |
| hsa-miR-141-3p | 2.13 |
8.71×10−6 |
1.77×10−3 | 2.01 |
6.05×10−3 |
2.29×10−2 |
| hsa-miR-376c-3p
- | 2.07 |
4.41×10−6 |
8.95×10−4 | −2.08 |
6.82×10−4 |
3.32×10−3 |
| hsa-miR-381-3p
- | 2.11 |
6.51×10−6 |
1.33×10−3 | −2.35 |
4.88×10−4 |
2.47×10−3 |
| hsa-miR-145-5p
- | 2.67 |
1.15×10−8 |
2.35×10−6 | −2.16 |
1.84×10−4 |
1.03×10−3 |
| hsa-miR-378i | 2.79 |
2.54×10−6 |
5.15×10−4 | 2.13 |
2.05×10−3 |
8.70×10−3 |
| hsa-miR-98-5p | 1.286 |
2.42×10−4 |
4.91×10−2 | 5.30 |
4.07×10−6 |
4.21×10−5 |
| hsa-miR-7-5p | 1.48 |
9.31×10−5 |
1.90×10−2 | 4.92 |
6.08×10−5 |
3.89×10−4 |
|
hsa-miR-374b-5p | 1.34 |
1.57×10−4 |
3.19×10−2 | 5.59 |
5.44×10−7 |
1.45×10−5 |
|
hsa-miR-374a-5p | 1.42 |
1.08×10−4 |
2.20×10−2 | 5.74 |
1.54×10−7 |
8.57×10−6 |
|
hsa-miR-301a-3p | 1.56 |
8.62×10−5 |
1.76×10−2 | 5.60 |
5.44×10−7 |
1.45×10−5 |
| hsa-miR-17-3p | 2.06 |
1.84×10−4 |
3.74×10−2 | 5.42 |
1.64×10−6 |
2.15×10−5 |
| hsa-miR-335-5p | 3.09 |
2.31×10−4 |
4.70×10−2 | 4.22 |
1.65×10−3 |
7.19×10−3 |
| hsa-miR-186-5p | 3.34 |
7.16×10−6 |
1.46×10−3 | 3.93 |
6.75×10−3 |
2.53×10−2 |
|
hsa-miR-148a-3p | 2.50 |
1.50×10−5 |
3.05×10−3 | 5.27 |
5.01×10−6 |
4.90×10−5 |
| hsa-miR-532-5p | 3.35 |
1.78×10−4 |
3.63×10−2 | 4.30 |
1.21×10−3 |
5.51×10−3 |
| hsa-miR-660-5p | 3.45 |
1.29×10−4 |
2.62×10−2 | 4.43 |
7.61×10−4 |
3.64×10−3 |
| hsa-miR-205-5p | 4.31 |
6.74×10−6 |
1.37×10−4 | 4.48 |
7.61×10−4 |
3.64×10−3 |
| hsa-miR-126-3p | 4.15 |
1.17×10−5 |
2.37×10−3 | 4.85 |
6.98×10−5 |
4.44×10−4 |
|
hsa-miR-200c-3p | 4.00 |
7.13×10−7 |
1.45×10−4 | 5.05 |
2.99×10−5 |
2.06×10−4 |
Comparison of target genes of DE-miRNAs
between OC cells and normal controls
Subsequent to searching the miRWalk 2.0 database,
only the target genes of 11 shared DE-miRNAs [upregulated: homo
sapiens (has)-miR-126-3p, hsa-miR-141-3p, hsa-miR-17-3p,
hsa-miR-186-5p, hsa-miR-301a-3p, hsa-miR-335-5p, hsa-miR-532-5p,
hsa-miR-7-5p, hsa-miR-374b-5p, and hsa-miR-98-5p; down-regulated:
hsa-miR-145-5p] were obtained. These common miRNAs and their target
genes were then used to construct a miRNA-mRNA regulatory network,
as demonstrated in Fig. 2. This
network included 4,198 nodes (11 miRNAs and 4187 target genes) and
462 edges (interaction associations).
Screening prognosis-associated miRNAs and
their target genes
The miRNA/mRNA expression and clinical data
associated with OC were also extracted from the TCGA database to
validate the clinical importance of the miRNAs identified in the
present study. Subsequently, the miRNA and mRNA expression data
were matched with the clinical data for 414 patients. The
expression of 11 shared DE-miRNAs was extracted from the TCGA data
and then combined with the clinical data to screen the
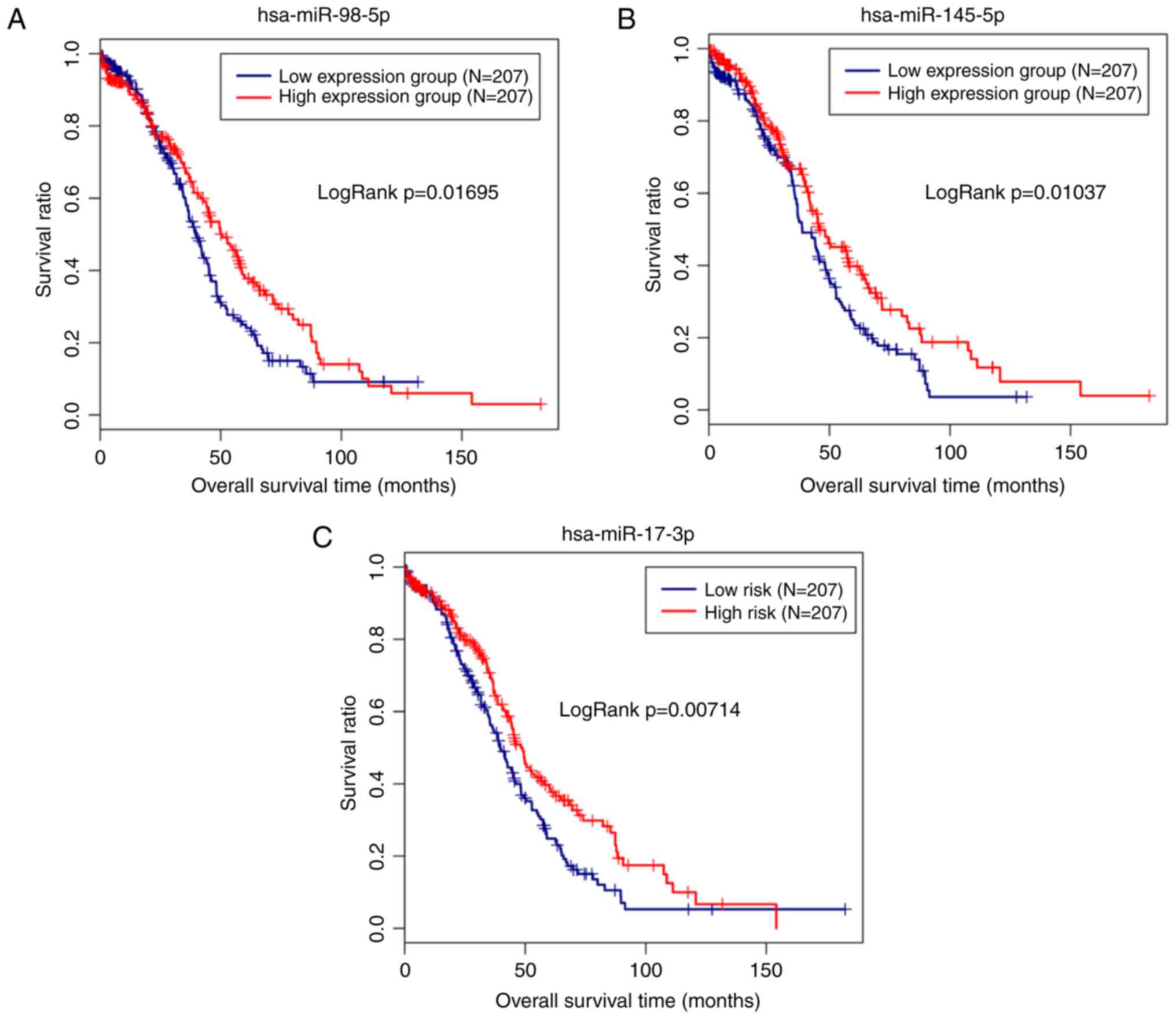
prognosis-associated miRNAs. Notably, hsa-miR-98-5p (upregulated;
P=0.01046), hsa-miR-145-5p (downregulated; P=0.0178) and
hsa-miR-17-3p (upregulated; P=0.0294) were significantly correlated
with the survival outcomes of patients with OC. The Kaplan-Meier
analysis additionally revealed a poor prognosis for patients with
low expression levels of hsa-miR-145-5p, as expected (Fig. 3). Similarly, the subsequent
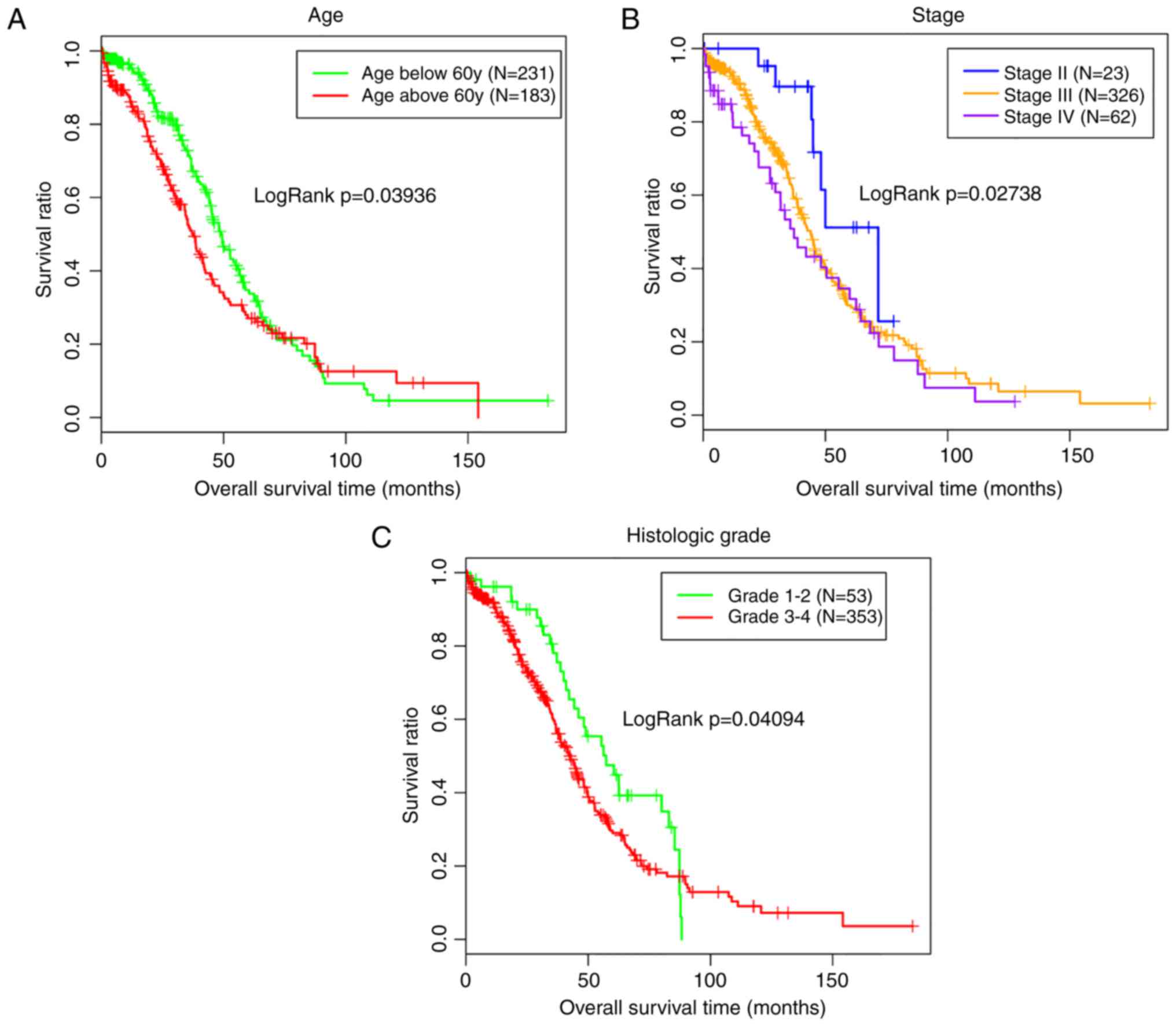
univariate analyses revealed an association between hsa-miR-145-5p
expression and the tumor stage (Table II), which was also an independent
prognosis factor with survival (Table III; Fig. 4).
| Table IIAssociations between miRNAs and
clinical characteristics using data from The Cancer Genome Atlas
data. |
Table II
Associations between miRNAs and
clinical characteristics using data from The Cancer Genome Atlas
data.
| hsa-miR-17-3p
| hsa-miR-145-5p
| hsa-miR-98-5p
|
|---|
| Clinical
characteristics | P-value | P-value | P-value |
|---|
| Age
(59.44±11.42) | 0.10 | 0.30 | 0.26 |
| Radiation therapy
(yes/no) | 0.90 | 0.61 |
6.62×10−3 |
| Neoplasm
subdivision (bilateral/left/right) | 0.95 | 0.09 | 0.30 |
| Stage
(II/III/IV) | 0.01 | 0.01 | 0.39 |
| Lymphatic invasion
(yes/no) | 0.53 | 0.65 | 0.12 |
| Histologic grade
(G1-G2/G3-G4) | 0.26 | 0.85 | 0.88 |
| Recurrence
(yes/no) | 0.14 | 0.74 | 0.09 |
| Table IIIPrognosis-associated clinical
characteristics using data from The Cancer Genome Atlas data. |
Table III
Prognosis-associated clinical
characteristics using data from The Cancer Genome Atlas data.
| Variables | Univariate analysis
| Multivariate
analysis
|
|---|
| HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Radiation therapy
(yes/no) | 0.84 | 0.53-1.34 | 0.47 | - | - | - |
| Neoplasm
subdivision (bilateral/left/right) | 0.99 | 0.80-1.21 | 0.89 | - | - | - |
| Lymphatic invasion
(yes/no) | 1.14 | 0.70-1.86 | 0.59 | - | - | - |
| Recurrence
(yes/no) | 1.73 | 1.45-2.20 | 0.21 | - | - | - |
| Age
(59.44±11.42) | 1.02 | 1.01-1.03 |
3.78×10−3 | 1.02 | 1.01-1.03 |
2.49×10−3 |
| Stage
(II/III/IV) | 1.38 | 1.04-1.82 | 0.03 | 1.36 | 1.02-1.83 | 0.04 |
| Histologic grade
(G1-G2/G3-G4) | 1.39 | 0.96-2.00 | 0.04 | 1.35 | 0.93-1.95 | 0.04 |
The 91 target genes of hsa-miR-145-5p were included
in the screen of prognosis-associated genes. As a result, 11 genes,
including ADP ribosylation factor-like GTPase 6 interacting protein
5 (P=0.017), connective tissue growth factor (CTGF; P=0.047),
matrix metallopeptidase 12 (P=0.0068), myotubularin-related protein
14 (MTMR14; P=0.03), p21 (RAC1)-activated kinase 4 (P=0.026),
protein phosphatase 3 catalytic subunit alpha (PPP3CA; P=0.018),
suppressor of cytokine signaling 7 (SOCS7; P=0.0084), STAT1
(P=0.015), tropomyosin 3 (P=0.0015), tetraspanin 6 (P=0.036),
vascular endothelial growth factor A (P=0.0049) and AT-rich
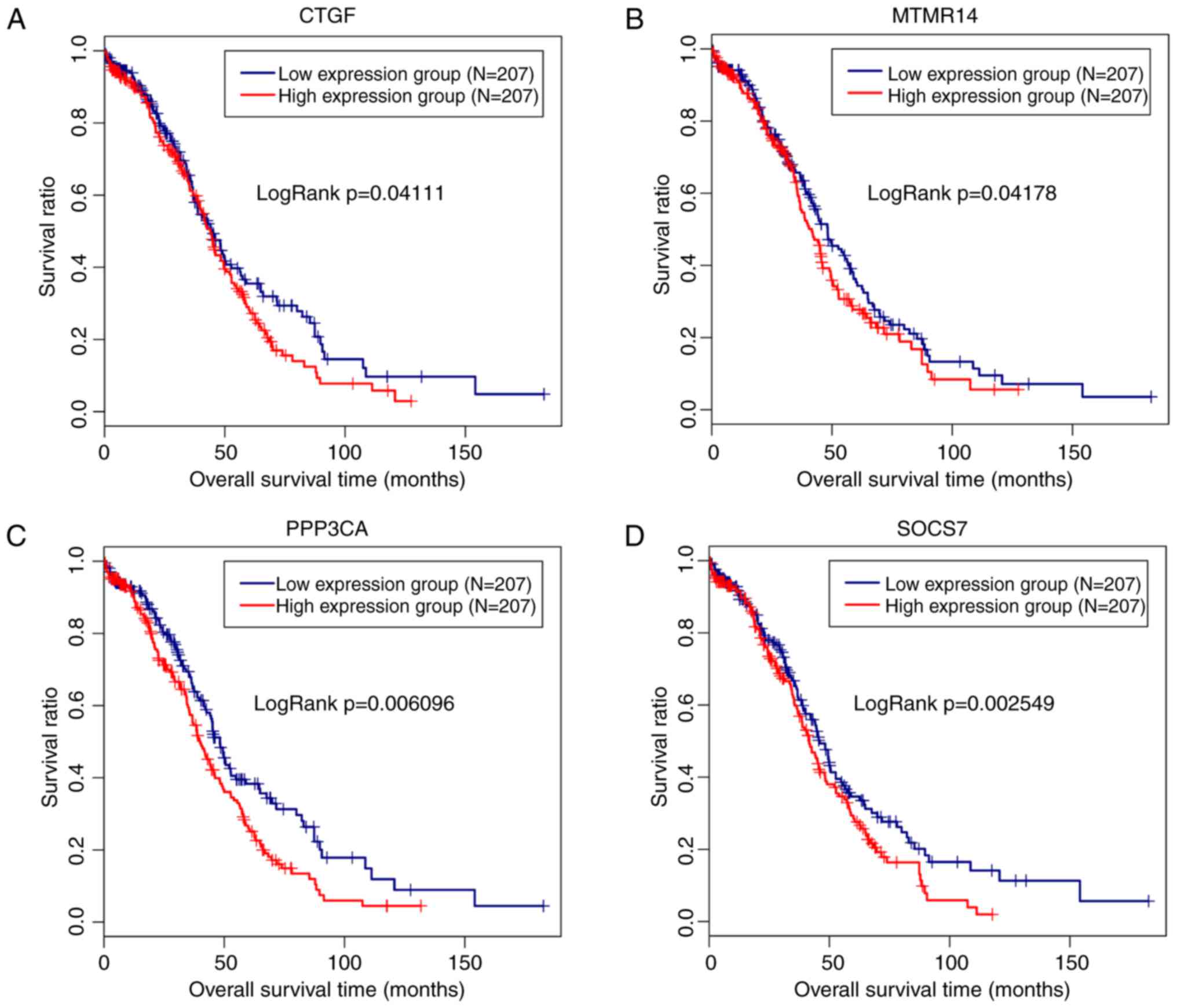
interaction domain 4B (P=0.015), were obtained, among which 4 genes
(CTGF, exp(coef)=1.1; MTMR14, exp(coef)=1.42; PPP3CA,
exp(coef)=1.31; SOCS7, exp(coef)=1.43) were suggested to be risk
factors for prognosis. Therefore, a Kaplan-Meier survival curve was
drawn for these 4 genes to reveal their associations with survival.
As anticipated, a poor prognosis was observed for patients
expressing high levels of these 4 genes, and the difference was
significant (Fig. 5). However,
only a significant negative correlation was observed between
hsa-miR-145-5p and CTGF (r=−0.1126, P=0.02188) using Pearson’s
correlation analysis (Fig.
6).
According to the functional analysis, the target
genes of hsa-miR-145-5p participated in OC development by affecting
26 KEGG pathways, 20 of which overlapped with 56 known
OC-associated pathways downloaded from the CTD (Table IV). CTGF was identified to be
involved in the Hippo signaling pathway (hsa04390).
| Table IVFunctional enrichment for the target
genes of microR-145. |
Table IV
Functional enrichment for the target
genes of microR-145.
| Term | P-value | Genes |
|---|
| hsa05219:Bladder
cancer |
3.91×10−7 | NRAS, CDKN1A,
VEGFA, MDM2, CDK4, MYC, MMP1 |
| hsa04550:Signaling
pathways regulating |
5.82×10−5 | NRAS, IGF1R, NANOG,
POU5F1, SOX2, MYC, FZD7, KLF4 |
| pluripotency of
stem cells | | |
| hsa05200:Pathways
in cancer |
8.73×10−5 | NRAS, IGF1R,
CDKN1A, HDAC2, VEGFA, MDM2, STAT1, |
| | CDK4, MYC, FZD7,
MMP1, TPM3 |
| hsa04151:PI3K-Akt
signaling pathway |
1.45×10−4 | NRAS, IGF1R,
CDKN1A, EIF4E, ITGB8, IFNB1, VEGFA, |
| | MDM2, CDK4, IRS1,
MYC |
| hsa05220:Chronic
myeloid leukemia |
1.57×10−4 | NRAS, CDKN1A,
HDAC2, MDM2, CDK4, MYC |
|
hsa05205:Proteoglycans in cancer |
5.32×10−4 | NRAS, IGF1R,
CDKN1A, NANOG, VEGFA, MDM2, MYC, |
| | FZD7 |
| hsa05161:Hepatitis
B |
5.72×10−4 | NRAS, CDKN1A,
IFNB1, TIRAP, STAT1, CDK4, MYC |
|
hsa05214:Glioma |
1.19×10−3 | NRAS, IGF1R,
CDKN1A, MDM2, CDK4 |
|
hsa05202:Transcriptional misregulation in
cancer |
1.24×10−3 | IGF1R, ERG, CDKN1A,
FLI1, HDAC2, MDM2, MYC |
| hsa04115:p53
signaling pathway |
1.33×10−3 | PPM1D, CDKN1A,
SERPINE1, MDM2, CDK4 |
|
hsa05218:Melanoma |
1.65×10−3 | NRAS, IGF1R,
CDKN1A, MDM2, CDK4 |
| hsa05206:MicroRNAs
in cancer |
4.11×10−3 | NRAS, CDKN1A, IRS2,
PAK4, VEGFA, MDM2, IRS1, MYC |
| hsa04919:Thyroid
hormone signaling pathway |
9.03×10−3 | NRAS, HDAC2, MDM2,
STAT1, MYC |
| hsa04110:Cell
cycle |
1.20×10−2 | CDKN1A, HDAC2,
MDM2, CDK4, MYC |
| hsa04360:Axon
guidance |
1.31×10−2 | NRAS, PAK4, ROBO2,
PPP3CA, SRGAP1 |
| hsa05216:Thyroid
cancer |
1.84×10−2 | NRAS, MYC,
TPM3 |
| Shsa04390:Hippo
signaling pathway |
2.32×10−2 | CTGF, SOX2,
SERPINE1, MYC, FZD7 |
| hsa05215:Prostate
cancer |
2.54×10−2 | NRAS, IGF1R,
CDKN1A, MDM2 |
| hsa05166:HTLV-I
infection |
3.66×10−2 | NRAS, CDKN1A,
PPP3CA, CDK4, MYC, FZD7 |
|
hsa05169:Epstein-Barr virus infection |
4.78×10−2 | CDKN1A, HDAC2,
MDM2, MYC, MAP2K6 |
Discussion
In the present study, the miRNA expression profiles
of OC cells and their exosomes compared with normal ovarian surface
epithelial cell lines were examined using a microarray. A total of
22 miRNAs were co-expressed in exosomes and the OC cells from which
they were derived. Among these miRNAs, the downregulation of
hsa-miR-145-5p and its negatively regulated target gene CTGF were
additionally demonstrated to be associated with the prognosis of
patients with OC by affecting the Hippo signaling pathway, and
therefore they may be potentially important diagnostic biomarkers
and therapeutic targets for OC.
Based on extensive evidence, miR-145-5p functions as
a tumor suppressor gene in various cancers, including OC. For
example, Zhang et al (22)
observed the downregulation of miR-145-5p in gastric cancer tissues
compared with the adjacent normal tissues. Low expression of
miR-145-5p was significantly associated with lymph node metastasis,
metastasis stage and distant metastasis, ultimately leading to
poorer overall survival. In the study by Ozen et al
(23), overexpression of
miR-145-5p inhibited proliferation and decreased the migration of
prostate cancer cells. Similarly, miR-145-5p is also an important
target for non-small cell lung cancer (NSCLC) as miR-145-5p
overexpression suppressed the EMT in NSCLC cells, which is an
important biological process associated with cancer migration and
metastasis (24). Upon
transfection of miR-145-5p, the angiogenesis ability of SW480 colon
carcinoma cells was significantly inhibited (25). Concomitantly, the overexpression
of miR-145 significantly suppressed the proliferation, migration
and invasion of OC cells and inhibited tumor growth and metastasis
in vivo (26,27). Furthermore, tumor protein 53
(TP53) upregulated the expression of miR-145 to enhance its tumor
suppressor roles (28-30). However, TP53 mutations occur in
~95% of high-grade serous OC (31). Therefore, miR-145 may be
specifically down regulated in advanced OC. Consistent with these
studies, miR-145 was expressed at lower levels in all 13 OC cell
lines and its downregulation was significantly associated with a
shorter survival and higher tumor stage in the present study.
Although miR-145 serves crucial roles in the
development of cancer, the majority of studies have focused on the
original cells, and the expression of miR-145 in exosomes and its
effect on OC remains largely unknown. However, one previous study
revealed that the exposure of pancreatic ductal adenocarcinoma to
miR-145-5p-enriched tumor-associated stroma exosomes cells
decreased cell viability in a dose-dependent manner (32). To the best of our knowledge, the
present study is the first to demonstrate the significant
downregulation of miR-145-5p expression in OC exosomes, indicating
that the enhanced secretion of miR-145 via exosomes may be an
underlying approach to treating OC (33,34). In addition, exosomes secreted from
primary tumors are transferred and released to the peripheral
circulation (35-37). Therefore, the expression of
exosomal miRNAs detected in cancer cells is generally similar to
their levels in blood. This hypothesis has been verified in the
study by Hannafon et al (38), in which miR-1246 and miR-21 were
first detected at significantly higher levels in exosomes from
breast cancer cells and then validated in patient-derived
orthotopic xenograft models, and mouse and human plasma exosomal
samples. According to their analysis of the receiver operating
characteristic curves, the diagnostic accuracy (area under the
curve) of the combination of plasma exosomal miR-1246 and miR-21
levels was 72.66% for breast cancer. Consistent with these data, we
hypothesize that the exosomal miR-145-5p identified in OC cells may
also be a potential biomarker for the detection of malignant
OC.
miRNAs are considered to participate in disease
development by regulating target genes through interactions with
complementary sequences in the 3′UTR. Several studies have explored
the mechanisms of miR-145-5p in cancer. For example, Matsushita
et al (39) predicted
putative binding sites for miR-145-5p in 1,735 genes following a
search of the microRNA.org database, and additionally
confirmed that ubiquitin-like with PHD and ring finger domains 1
may be an indirect target of miR-145-5p to regulate bladder cancer
cell aggressiveness in subsequent dual luciferase reporter assays.
Wu et al (27) revealed a
negative regulatory effect of miR-145 on levels of the ribosomal
protein S6 kinase, polypeptide 1 (P70S6K1) and mucin 1, cell
surface associated (MUC1) proteins in OC cells. Overexpression of
p70S6K1 and MUC1 restored the colony formation and invasion
abilities that were inhibited by miR-145. As indicated in the study
by Chen et al (40),
transfection of an miR-145 agomiR (activator) into OC cells
downregulated the expression of its direct target tripartite motif
containing 2 and subsequently induced apoptosis. However, the
mechanisms of miR-145 in OC are not well understood. In the present
study, 91 target genes of miR-145 were screened using the
miRwalk2.0 database, and the subsequent clinical correlation
analysis using TCGA data suggested that the Hippo signaling
pathway-associated gene CTGF may be an important target of miR-145,
due to its significant association with a poor prognosis and
negative correlation with miR-145 expression. The data from the
present study appear to be consistent with previous studies, as
high CTGF expression promotes the migration and peritoneal adhesion
of OC cells, resulting in a poor prognosis; these changes were
abrogated by a human monoclonal antibody against CTGF FG-3019
(41). CTGF is a key downstream
intermediate in the Hippo-YAP1/TEAD cascade that controls cell
growth and initiates OC formation (42-44). CTGF was indicated to be a direct
target of miR-145 using a dual luciferase reporter gene assay, and
miR-145 negatively regulated CTGF to affect the proliferation,
migration and invasion of esophageal squamous cell carcinoma
(45) and malignant glioma cells
transfected with miR-145 mimics and CTGF small interfering RNA
(46). Nevertheless, to the best
of our knowledge, studies of this nature in OC have not been
described, requiring additional investigation. Notably, we
hypothesized that the upregulation of CTGF in OC cells due to the
downregulation of miR-145 may be one potential contributor to the
decreased sorting of miR-145 into exosomes and subsequent transfer
to acceptor cells, including cancer associated fibroblasts,
endothelial cells and tumor-associated macrophages, to inhibit the
pro-tumor environment (47).
Furthermore, CTGF may also be a crucial target of exosomal miR-145
in the tumor environment. Low expression of miR-145 in exosomes was
insufficient to block the transcription of CTGF, leading to its
overexpression, acceleration of tumor cell invasion and the
carcinogenesis of adjacent normal ovarian epithelial cells
(48,49).
The present study had certain limitations: Firstly,
13 OC cell lines were used to screen crucial miRNAs in the present
study. Due to the presence of underlying differences in phenotype,
the expression of miRNAs varied among these cell lines, which lead
to unexpected and paradoxical results for specific miRNAs,
including miR-98 and miR-17. According to the prognosis analysis,
miR-98 and miR-17 should be expressed at lower levels in OC, but in
fact they were upregulated in OC and exosomes. Secondly, the TCGA
data did not include a normal control, and potential deviations in
the identified correlations between the identified miRNAs and
clinical characteristics may have occurred. Thirdly, although the
analysis preliminarily revealed a negative correlation between
miR-145 and its target gene CTGF in clinical samples, in
vitro and in vivo experiments using OC models are
required. Fourthly, the loss of exosomal miR-145 as a potentially
important factor contributing to OC pathogenesis was identified,
but the mechanism underlying its formation and whether modification
of exosomes by transfection with miR-145 decrease oncogenesis by
affecting the tumor environment remain unknown (33,34,50). Fifthly, although previous studies
have suggested that the levels of exosomal miRNAs in cancer cells
are consistent with their levels in the peripheral circulation
(35-38), the expression of exosomal miR-145
in plasma samples from patients with OC requires additional
verification to confirm its underlying value as a biomarker.
According to the data from the present study, the
decreased expression of hsa-miR-145 in OC and OC-derived exosomes
may be required for the development of OC via the targeted
modulation of a downstream intermediate in the Hippo signaling
pathway, CTGF. Therefore, upregulation of hsa-miR-145 in exosomes
and cells to inhibit CTGF expression may be a potential therapeutic
approach for OC.
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
The microarray data GSE103708 were downloaded from
the GEO database in NCBI (http://www.ncbi.nlm.nih.gov/geo/).
Authors’ contributions
WH, QH and XX conceived the design of the original
study. WH and YF conducted the statistical analysis and drafted the
manuscript. ZS, YY and YZ were involved with the interpretation of
the data. QH and XX participated in critical revisions of the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cliby WA, Powell MA, Alhammadi N, Chen L,
Miller JP, Roland PY, Mutch DG and Bristow RE: Ovarian cancer in
the United States: Contemporary patterns of care associated with
improved survival. Gynecol Oncol. 136:11–17. 2015. View Article : Google Scholar
|
|
3
|
Hausser J and Zavolan M: Identification
and consequences of miRNA-target interactions-beyond repression of
gene expression. Nat Rev Genet. 15:599–612. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Qin W, Xie W, He Q, Sun T, Meng C, Yang K,
Luo Y and Yang D: MicroRNA-152 inhibits ovarian cancer cell
proliferation and migration and may infer improved outcomes in
ovarian cancer through targeting FOXP1. Exp Ther Med. 15:1672–1679.
2018.PubMed/NCBI
|
|
5
|
Liu H, Pan Y, Han X, Liu J and Li R:
MicroRNA-216a promotes the metastasis and epithelial-mesenchymal
transition of ovarian cancer by suppressing the PTEN/AKT pathway.
Onco Targets Ther. 10:2701–2709. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Neviani P and Fabbri M: Exosomic microRNAs
in the tumor microenvironment. Front Med. 2:472015. View Article : Google Scholar
|
|
7
|
Ying X, Wu Q, Wu X, Zhu Q and Wang X,
Jiang L, Chen X and Wang X: Epithelial ovarian cancer-secreted
exosomal miR-222-3p induces polarization of tumor-associated
macrophages. Oncotarget. 7:43076–43087. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kanlikilicer P, Rashed MH, Bayraktar R,
Mitra R, Ivan C, Aslan B, Zhang X, Filant J, Silva AM,
Rodriguez-Aguayo C, et al: Ubiquitous release of exosomal tumor
suppressor miR-6126 from ovarian cancer cells. Cancer Res.
76:7194–7207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
De A, Powers B, De A, Zhou J, Sharma S,
Van VP, Bansal A, Sharma R and Sharma M: Emblica officinalis
extract downregulates pro-angiogenic molecules via upregulation of
cellular and exosomal miR-375 in human ovarian cancer cells.
Oncotarget. 7:31484–31500. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Meng X, Müller V, Mildelangosch K,
Trillsch F, Pantel K and Schwarzenbach H: Diagnostic and prognostic
relevance of circulating exosomal miR-373, miR-200a, miR-200b and
miR-200c in patients with epithelial ovarian cancer. Oncotarget.
7:16923–16935. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qin X, Yu S, Xu X, Bo S and Feng J:
Comparative analysis of microRNA expression profiles between A549,
A549/DDP and their respective exosomes. Oncotarget. 8:42125–42135.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Smyth GK: Limma: Linear models for
microarray data. Bioinformatics and Computational Biology Solutions
using R and Bioconductor. Gentleman R, Carey V, Du S, Irizarry R
and Huber W: Springer; New York: pp. 397–420. 2005, View Article : Google Scholar
|
|
13
|
Benjamini Y and Hochberg Y: Controlling
the false discovery rate: A practical and powerful approach to
multiple testing. J R Stat Soc B. 57:289–300. 1995.
|
|
14
|
Dweep H and Gretz N: miRWalk2.0: A
comprehensive atlas of microRNA-target interactions. Nat Methods.
12:6972015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kohl M, Wiese S and Warscheid B:
Cytoscape: Software for visualization and analysis of biological
networks. Methods Mol Biol. 696:291–303. 2011. View Article : Google Scholar
|
|
16
|
Therneau TM: A package for survival
analysis in S. R package version 2. 37:72014.
|
|
17
|
Shimizu Y, Kamoi S, Amada S, Akiyama F and
Silverberg SG: Toward the development of a universal grading system
for ovarian epithelial carcinoma: Testing of a proposed system in a
series of 461 patients with uniform treatment and follow-up.
Cancer. 82:893–901. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kosary CL: FIGO stage, histology,
histologic grade, age and race as prognostic factors in determining
survival for cancers of the female gynecological system: An
analysis of 1973-87 SEER cases of cancers of the endometrium,
cervix, ovary, vulva, and vagina. Semin Surg Oncol. 10:31–46. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ogata H, Goto S, Sato K, Fujibuchi W, Bono
H and Kanehisa M: KEGG: Kyoto encyclopedia of genes and genomes.
Nucleic Acids Res. 27:29–34. 1999. View Article : Google Scholar
|
|
20
|
Huang Da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Davis AP, Grondin CJ, Johnson RJ, Sciaky
D, King BL, McMorran R, Wiegers J, Wiegers TC and Mattingly CJ: The
comparative toxicogenomics database: Update 2017. Nucleic Acids
Res. 45:D972–D978. 2017. View Article : Google Scholar :
|
|
22
|
Zhang Y, Wen X, Hu XL, Cheng LZ, Yu JY and
Wei ZB: Downregulation of miR-145-5p correlates with poor prognosis
in gastric cancer. Eur Rev Med Pharmacol Sci. 20:3026–3030.
2016.PubMed/NCBI
|
|
23
|
Ozen M, Karatas OF, Gulluoglu S, Bayrak
OF, Sevli S, Guzel E, Ekici ID, Caskurlu T, Solak M, Creighton CJ
and Ittmann M: Overexpression of miR-145-5p inhibits proliferation
of prostate cancer cells and reduces SOX2 expression. Cancer
Invest. 33:251–258. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chang Y, Yan W, Sun C, Liu Q, Wang J and
Wang M: miR-145-5p inhibits epithelial-mesenchymal transition via
the JNK signaling pathway by targeting MAP3K1 in non-small cell
lung cancer cells. Oncol Lett. 14:6923–6928. 2017.
|
|
25
|
Thuringer D, Jego G, Berthenet K, Hammann
A, Solary E and Garrido C: Gap junction-mediated transfer of
miR-145-5p from microvascular endothelial cells to colon cancer
cells inhibits angiogenesis. Oncotarget. 7:28160–28168. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dong R, Liu X, Zhang Q, Jiang Z, Li Y, Wei
Y, Li Y, Yang Q, Liu J, Wei JJ, et al: miR-145 inhibits tumor
growth and metastasis by targeting metadherin in high-grade serous
ovarian carcinoma. Oncotarget. 5:10816–10829. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu H, Xiao ZH, Wang K, Liu W and Hao Q:
MiR-145 is down-regulated in human ovarian cancer and modulates
cell growth and invasion by targeting p70S6K1 and MUC1. Biochem
Biophys Res Commun. 441:693–700. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Suzuki HI, Yamagata K, Sugimoto K, Iwamoto
T, Kato S and Miyazono K: Modulation of microRNA processing by p53.
Nature. 460:529–533. 2009. View Article : Google Scholar
|
|
29
|
Spizzo R, Nicoloso MS, Lupini L, Lu Y,
Fogarty J, Rossi S, Zagatti B, Fabbri M, Veronese A, Liu X, et al:
miR-145 participates with TP53 in a death-promoting regulatory loop
and targets estrogen receptor-alpha in human breast cancer cells.
Cell Death Differ. 17:246–254. 2010. View Article : Google Scholar
|
|
30
|
Boominathan L: The guardians of the genome
(p53, TA-p73, and TA-p63) are regulators of tumor suppressor miRNAs
network. Cancer Metastasis Rev. 29:613–639. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Brachova P, Thiel KW and Leslie KK: The
consequence of oncomorphic TP53 mutations in ovarian cancer. Int J
Mol Sci. 14:19257–19275. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Han S, Belsare S, Zhang DY, Beveridge M,
Rinaldi C, Trevino JG, Schmittgen TD and Hughes SJ: Stroma-derived
extracellular vesicles deliver tumor-suppressive miRNAs to
pancreatic cancer cells. Oncotarget. 9:5764–5777. 2017.
|
|
33
|
Shi M, Jiang Y, Yang L, Yan S, Wang YG and
Lu XJ: Decreased levels of serum exosomal miR-638 predict poor
prognosis in hepatocellular carcinoma. J Cell Biochem.
119:4711–4716. 2018. View Article : Google Scholar
|
|
34
|
Zhang Z, Li X, Sun W, Yue S, Yang J, Li J,
Ma B, Wang J, Yang X, Pu M, et al: Loss of exosomal miR-320a from
cancer-associated fibroblasts contributes to HCC proliferation and
metastasis. Cancer Lett. 397:33–42. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Taylor DD 1 and Gercel-Taylor C: MicroRNA
signatures of tumor-derived exosomes as diagnostic biomarkers of
ovarian cancer. Gynecol Oncol. 110:13–21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Suetsugu A, Honma K, Saji S, Moriwaki H,
Ochiya T and Hoffman RM: Imaging exosome transfer from breast
cancer cells to stroma at metastatic sites in orthotopic nude-mouse
models. Adv Drug Deliv Rev. 5:383–390. 2013. View Article : Google Scholar
|
|
37
|
Ostenfeld MS, Jeppesen D, Morth JP, Khanh
HB, Dan T, Borre M, Dyrskjøt L and Ørntoft T: Abstract 3387:
Secreted exosomes from cultured bladder cells are enriched for
distinct miRNAs detected in circulation of metastatic bladder
cancer patients. Cancer Res. 72(Suppl): S33872012. View Article : Google Scholar
|
|
38
|
Hannafon BN, Trigoso YD, Calloway CL, Zhao
YD, Lum DH, Welm AL, Zhao ZJ, Blick KE, Dooley WC and Ding WQ:
Plasma exosome microRNAs are indicative of breast cancer. Breast
Cancer Res. 18:902016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Matsushita R, Yoshino H, Enokida H, Goto
Y, Miyamoto K, Yonemori M, Inoguchi S, Nakagawa M and Seki N:
Regulation of UHRF1 by dual-strand tumor-suppressor microRNA-145
(miR-145-5p and miR-145-3p): Inhibition of bladder cancer cell
aggressiveness. Oncotarget. 7:28460–28487. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen X, Dong C, Law PT, Chan MT, Su Z,
Wang S, Wu WK and Xu H: MicroRNA-145 targets TRIM2 and exerts
tumor-suppressing functions in epithelial ovarian cancer. Gynecol
Oncol. 139:513–519. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim MJ, Gloss BS, Rajmohan M, Chang DK,
Colvin EK, Jones MD, Samuel Y, Howell VM, Brown LM, Wong CW, et al:
Connective tissue growth factor as a novel therapeutic target in
high grade serous ovarian cancer. Oncotarget. 6:44551–44562.
2015.
|
|
42
|
Kang W, Huang T, Zhou Y, Zhang J, Lung
RWM, Tong JHM, Chan AWH, Zhang B, Wong CC, Wu F, et al: miR-375 is
involved in Hippo pathway by targeting YAP1/TEAD4-CTGF axis in
gastric carcinogenesis. Cell Death Dis. 9:n922018. View Article : Google Scholar
|
|
43
|
Pobbati AV and Hong W: Emerging roles of
TEAD transcription factors and its coactivators in cancers. Cancer
Biol Ther. 14:390–398. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tan G, Cao X, Dai Q, Zhang B, Huang J,
Xiong S, Zhang Y, Chen W, Yang J and Li H: A novel role for
microRNA-129-5p in inhibiting ovarian cancer cell proliferation and
survival via direct suppression of transcriptional co-activators
YAP and TAZ. Oncotarget. 6:8676–8686. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Han Q, Zhang HY, Zhong BL, Wang XJ, Zhang
B and Chen H: MicroRNA-145 inhibits cell migration and invasion and
regulates epithelial-mesenchymal transition (EMT) by targeting
connective tissue growth factor (CTGF) in esophageal squamous cell
carcinoma. Med Sci Monit. 22:3925–3934. 2016. View Article : Google Scholar :
|
|
46
|
Lee HK, Bier A, Cazacu S, Finniss S, Xiang
C, Twito H, Poisson LM, Mikkelsen T, Slavin S, Jacoby E, et al:
MicroRNA-145 is downregulated in glial tumors and regulates glioma
cell migration by targeting connective tissue growth factor. PLoS
One. 8:e546522013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Squadrito ML, Baer C, Burdet F, Maderna C,
Gilfillan G, Lyle R, Ibberson M and De Palma M: Endogenous RNAs
modulate microRNA sorting to exosomes and transfer to acceptor
cells. Cell Rep. 8:1432–1446. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li L, Li C, Wang S, Wang Z, Jiang J, Wang
W, Li X, Chen J, Liu K, Li C and Zhu G: Exosomes derived from
hypoxic oral squamous cell carcinoma cells deliver miR-21 to
normoxic cells to elicit a prometastatic phenotype. Cancer Res.
76:1770–1780. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Fang T, Lv H, Lv G, Li T, Wang C, Han Q,
Yu L, Su B, Guo L, Huang S, et al: Tumor-derived exosomal
miR-1247-3p induces cancer-associated fibroblast activation to
foster lung metastasis of liver cancer. Nat Commun. 9:1912018.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Trivedi M, Talekar M, Shah P, Ouyang Q and
Amiji M: Modification of tumor cell exosome content by transfection
with wt-p53 and microRNA-125b expressing plasmid DNA and its effect
on macrophage polarization. Oncogenesis. 5:e2502016. View Article : Google Scholar : PubMed/NCBI
|