Introduction
Brain ageing is a progressive and gradual
accumulation of harmful changes in the brain tissue with age,
including oxidative stress injury and neuronal inflammation, which
increase the risk of age-related learning and memory impairment
(1,2). At present, the oxidative stress
theory is recognised as the most feasible hypothesis to explain the
ageing process, particularly age-related neuronal degeneration
(3-5). Increasing evidence has suggested
that reactive oxygen species (ROS) may cause neuronal oxidative
stress, inflammation, and neurodegeneration with age (3,6-7).
There are several enzymes that can generate intracellular ROS.
NADPH oxidase (NOX) is a major source of ROS generation (4,5).
It has been found that NOX2 (gp91phox) is constitutively expressed
in neurons and significantly increased in the brain of ageing mice
(4). The expression of NOX2
subunits p47phox and p67phox are also increased in the cortex of
patients with mild cognitive disorders (5). Inflammasomes are NOD-like receptor
(NLR) family multiprotein complexes that are responsible for the
formation of pro-inflammatory molecules. Neuroinflammation mediated
by inflammasomes is important in several age-related
neurodegenerative diseases (8,9).
NOD-like receptor protein 1 (NLRP1) was the first reported
inflammasome (10). The NLRP1
inflammasome is widely expressed in the body and is the main
inflammasome in neurons. An increasing number of studies have shown
that the NLRP1 inflammasome is associated with age-related neuronal
damage and cognitive impairment (8,11).
NADPH oxidase-derived ROS accumulation is one of the main pathways
for NLRP3 inflammasome activation (12,13). However, whether NOX2-derived ROS
can activate the NLRP1 inflammasome, and whether it is involved in
neuronal senescence and damage, remain to be elucidated.
Ginsenoside Rg1 (Rg1) is one of the active
ingredients of ginseng, which has been used to improve health
conditions and delay senescence in China (14,15). It has been reported that Rg1 has
significant antioxidant, anti-ageing and neuroprotective effects
(16), and Rg1 can improve
learning and memory impairment in ageing mice (17,18). Our previous studies showed that
Rg1 significantly decreased ROS generation and attenuated the
neuronal oxidative stress damage induced by chronic restrain stress
in mice (19), and Rg1 protected
against neuroinflammation and neuronal injury induced by chronic
glucocorticoid exposure (20).
These data suggest that Rg1 has a protective effect on neuronal
damage due to decreasing ROS generation. Therefore, it was
hypothesised that Rg1 may downregulate NOX2 and reduce the
production of ROS, thereby inhibiting activation of the NLRP1
inflammasome in hippocampal neurons and protecting against
age-related neuronal damage.
In the present study, primary cultured hippocampal
neurons were treated with H2O2 (200
µM) for 24 h to mimic age-related neuronal damage. The study
evaluated the protective effects of ROS scavenger tempol and Rg1 on
H2O2-induced neuronal damage and assessed
whether Rg1 regulated the activation of the NLRP1 inflammasome via
the inhibition of NOX2 in hippocampal neurons.
Materials and methods
Culture and treatment of hippocampal
neurons
Primary hippocampal neurons were prepared from
postnatal Sprague-Dawley rats (within 24 h; 6-8
neonates/experiment) obtained from the Center of Laboratory Animals
of Anhui Medical University (Hefei, China) and placed in plates
coated with poly-L-lysine (10 µg/ml). Neurobasal medium with
B-27 supplement (Thermo Fisher Scientific, Inc., Waltham, MA, USA)
was used to culture the neurons at 37°C with 5% CO2 as
described previously (21). The
neurons were cultured for 7 days and divided into six groups:
Control group, H2O2 (200 µM) group,
H2O2 (200 µM) + tempol (100 µM)
group, and H2O2 (200 µM) + Rg1 (1, 5
and 10 µM) groups. With the exception of the control group,
the neurons in the groups were treated with
H2O2 (200 µM) and tempol (100
µM) or Rg1 (1, 5 or 10 µM) for 24 h. Tempol (Santa
Cruz Technology, Inc., Dallas, TX, USA) and Rg1 (content of Rg1
>98%; Chengdu Desite Biotechnology Co., Chengdu, China) were
dissolved in distilled water and stored at −80°C. All experimental
procedures were performed in accordance with the approved protocol
of the Ethics Committee of Anhui Medical University (Anhui,
China).
Neuronal apoptosis assay (Hoechst 33258
staining)
Hoechst 33258 can stain the nucleus and is often
used to assess the apoptotic rate of neurons (22). First, the neurons were fixed with
4% paraformaldehyde and incubated with Hoechst 33258 (5
µg/ml, Zhongshan Golden Bridge Biotechnology Co., Beijing,
China) for 15 min. Following washing with PBS, the neurons were
sealed onto slides with anti-fade mounting medium. Fluorescence
microscopy was then used to examine the apoptosis of neurons
(Olympus IX71; Olympus Corporation, Tokyo, Japan). Morphologically,
the apoptotic neurons appeared smaller and bright blue, the nucleus
was condensed and deeply stained (23). The apoptotic neurons were counted
and the relative neuronal apoptotic rate was examined in each
culture and compared with the control.
Examination of ROS production with
dihydroethidium (DHE) staining
DHE is the most commonly used ROS fluorescent probe
and is often used for labelling living cells to detect the
production of ROS (24). For the
ROS examination, DHE (10 µM) was added to the medium and
incubated for 30 min at 37°C. The production of ROS was then
detected using a fluorescence microscope (Olympus IX71, Olympus
Corporation). The red mean optical density was performed using
Image-Pro Plus 6.0 software (Media Cybernetics, Inc., Rockville,
MD, USA), to indicate the ROS production in the hippocampal
neurons.
Immunofluorescence
Caspase-3 is the common pathway and the executor of
apoptosis (25). To confirm the
effect of Rg1 on neuronal apoptosis in
H2O2-damaged hippocampal neurons, the
expression of caspase-3 was detected by immunofluorescence. First,
the hippocampal neurons were fixed with 4% paraformaldehyde for 30
min and washed thoroughly with PBS. Following permeabilisation with
Triton X-100 (0.25%) for 30 min, and blocking with 1% BSA
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) in PBS for 1 h, the
neurons were incubated with caspase-3 primary antibody (cat. no.
BS7003; lot no. CJ32131; 1:200; Bioworld Technology, Inc., St.
Louis Park, MN, USA) overnight at 4°C. Subsequently, the neurons
were incubated in secondary antibody conjugated to FITC (cat. no.
ZF-0311; lot no. 127805; 1:200; ZSGB-BIO; OriGene Technologies,
Inc., Beijing, China) for 1 h at room temperature. The slides were
then mounted and examined with a fluorescence microscope (Olympus
IX71; Olympus Corporation). The relative fluorescent intensity was
quantified from three random fields per slide using the Image-Pro
Plus 6.0 analysis system to indicate the expression of
caspase-3.
Immunoblotting assay
Radioimmunoprecipitation assay lysis buffer
(Beyotime Institute of Biotechnology, Haimen, China) were used to
extract the total protein. The protein concentration was determined
by BCA Protein Assay kit (Beyotime Institute of Biotechnology).
Equal samples of protein (30 µg) were separated by 12%
SDS-PAGE and transferred onto a PVDF membrane. The membrane was
blocked with 5% skim milk for 1 h at room temperature. The membrane
was then incubated with primary antibodies targeting
β-galactosidase (β-Gal), NOX2, p22phox, p47phox, NLRP1,
apoptosis-associated speck-like protein containing a
carboxy-terminal CARD (ASC), caspase-1 and β-actin (1:1,000)
overnight at 4°C. The primary antibodies for β-Gal (cat. no.
ab9361; lot no. GR27325-12), NOX2 (cat. no. ab31092; lot no.
GR91755-40), NLRP1 (cat. no. ab3683; lot no. GR281560-21) and
caspase-1 (cat. no. ab1872; lot no. GR615230-38) were from Abcam
(Cambridge, UK). The p22phox (cat. no. BS60290; lot no. CN21141)
and p47phox (cat. no. BS4852; lot no. CN33161) antibodies were from
Bioworld Technologies, Inc. ASC antibody (cat. no. SC-514414; lot
no. G1916) was from Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA) The dilutions of β-Gal, NOX2, NLRP1 and p47phox were 1:1,000.
The dilutions of ASC, caspase-1 and p22phox were 1:500. The
membranes were then washed with TBS-Tween 20 and incubated with
anti-rabbit IgG antibody conjugated to HRP (cat. no. ZF-2301; lot
no. 128615; 1:10,000; ZSGB-BIO; OriGene Technologies, Inc.) at room
temperature for 1 h. The protein bands were visualised by using a
Chemi Q4800 mini imaging system (Shanghai Bioshine Technology,
Shanghai, China). The protein bands were measured with ImageJ 1.44
software (National Institutes of Health, Bethesda, MD, USA) and
normalised to the corresponding β-actin bands. The relative density
of each target protein over the control was used to represent the
changes in expression of target proteins.
Enzyme-linked immunosorbent assay
(ELISA)
Following incubation with H2O2
and tempol or Rg1, the supernatants were collected from each group.
ELISA kits were used to detect the levels of interleukin (IL)-1β
and IL-18 (Cloud-Clone Corp., Houston, TX, USA). Briefly, the
samples and the standards of IL-1β and IL-18 were placed into the
assay plate and incubated for 1 h at 37°C. Following incubation for
1 h, the HRP-conjugated reagent (100 µl) was added to the
wells and incubated for 1 h at 37°C. Subsequently, the chromogen
solution (100 µl) was added to the wells. The plate was
mixed and incubated for 15 min at 37°C. Stop solution (50
µl) was then added to the wells and the absorbance at 450 nm
was detected with a microplate reader (Thermo Fisher Scientific,
Inc.) within 15 min.
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical differences were analysed by one-way analysis of
variance and the between-group comparisons were subjected to the
Bonferroni post hoc test using SPSS 17.0 software (SPSS, Inc.,
Chicago, IL, USA. P<0.05 was considered to indicate a
statistically significant difference.
Results
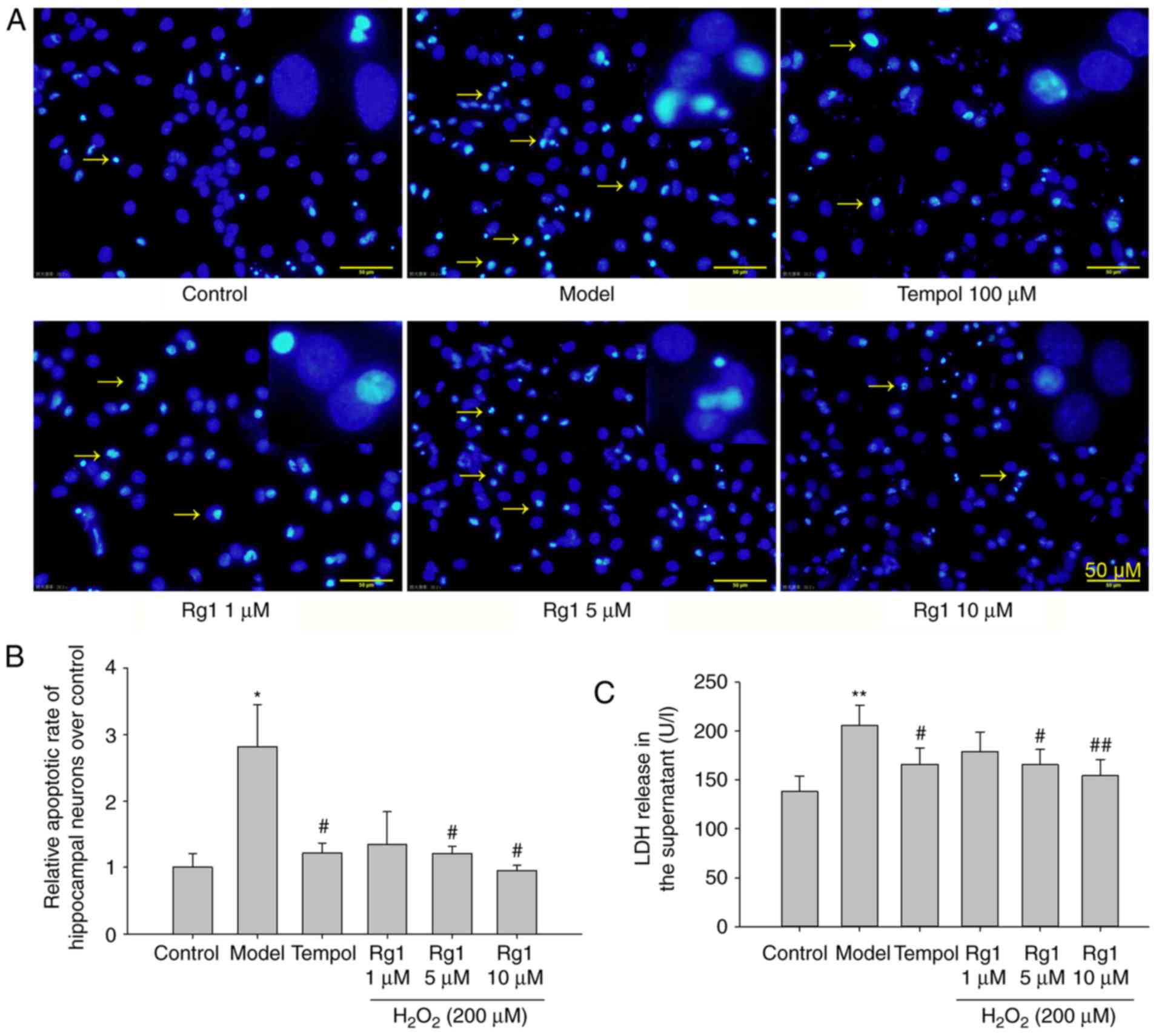
Rg1 inhibits
H2O2-induced neuronal apoptosis and the
expression of caspase-3
The present study first examined the effects of Rg1
treatment on H2O2-induced neuronal apoptosis
by staining with Hoechst 33258, which can bind to chromatin
allowing the visualisation of normal and condensed chromosomes with
a fluorescence microscope (22).
The results showed that there were few apoptotic neurons in the
control group. Compared with the control group,
H2O2 exposure significantly increased
neuronal apoptosis (Fig. 1A and
B; P<0.05). Compared with the
H2O2-treated group, Rg1 (5 and 10 µM)
treatment significantly reduced neuronal apoptosis (Fig. 1A and B; P<0.05). Treatment with
the ROS scavenger tempol (100 µM) significantly decreased
the apoptosis of hippocampal neurons induced by
H2O2 treatment (Fig. 1A and B; P<0.05).
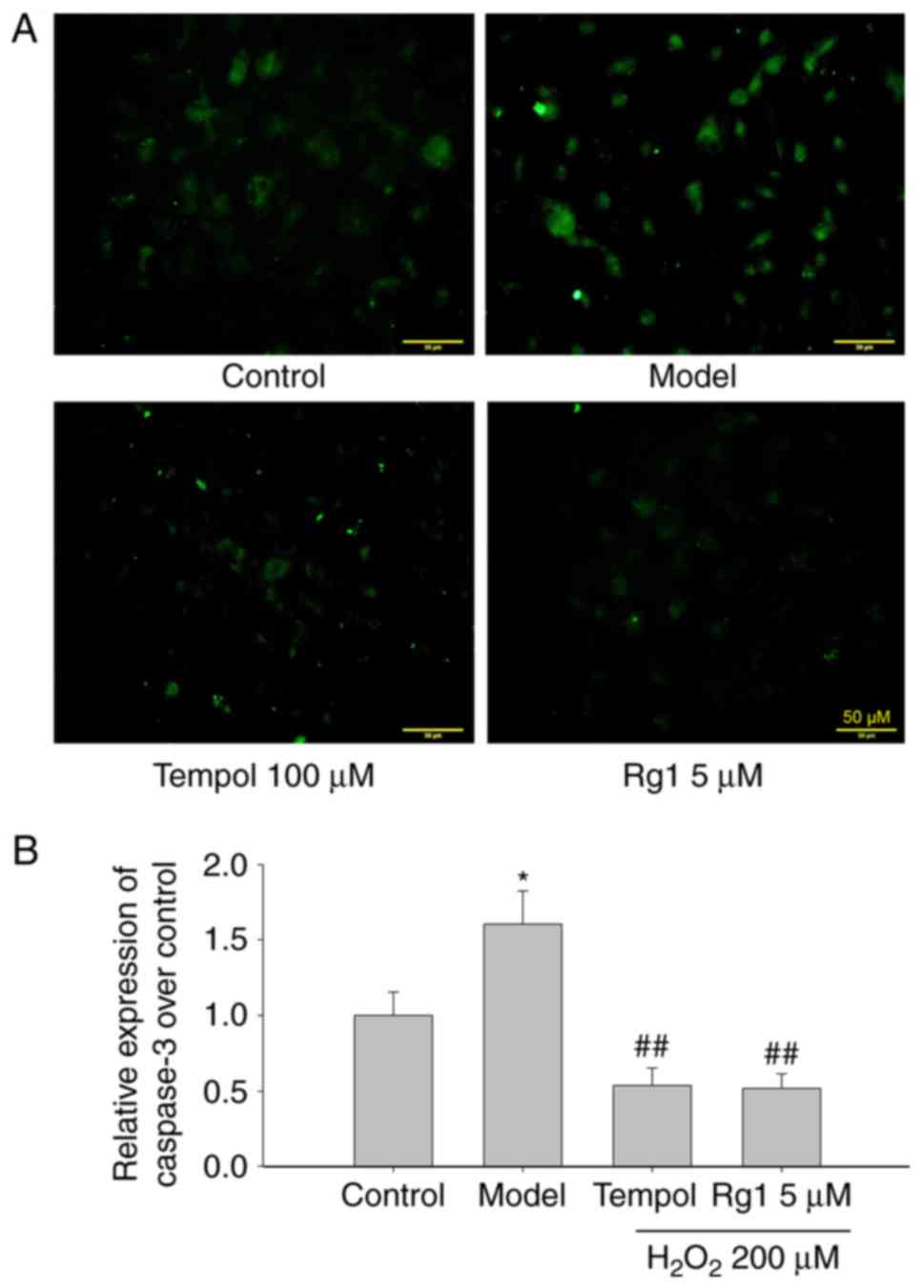
To confirm the anti-apoptotic effect of Rg1, the
effects of Rg1 (5 µM) on the expression of caspase-3 in
H2O2-damaged hippocampal neurons was further
examined by immunofluorescence. The results showed that, compared
with the control group, H2O2 treatment
significantly increased the expression of caspase-3. Compared with
the H2O2-treated model group, tempol and Rg1
treatments significantly decreased the expression of caspase-3 in
H2O2-treated hippocampal neurons (Fig. 2A and B; P<0.05).
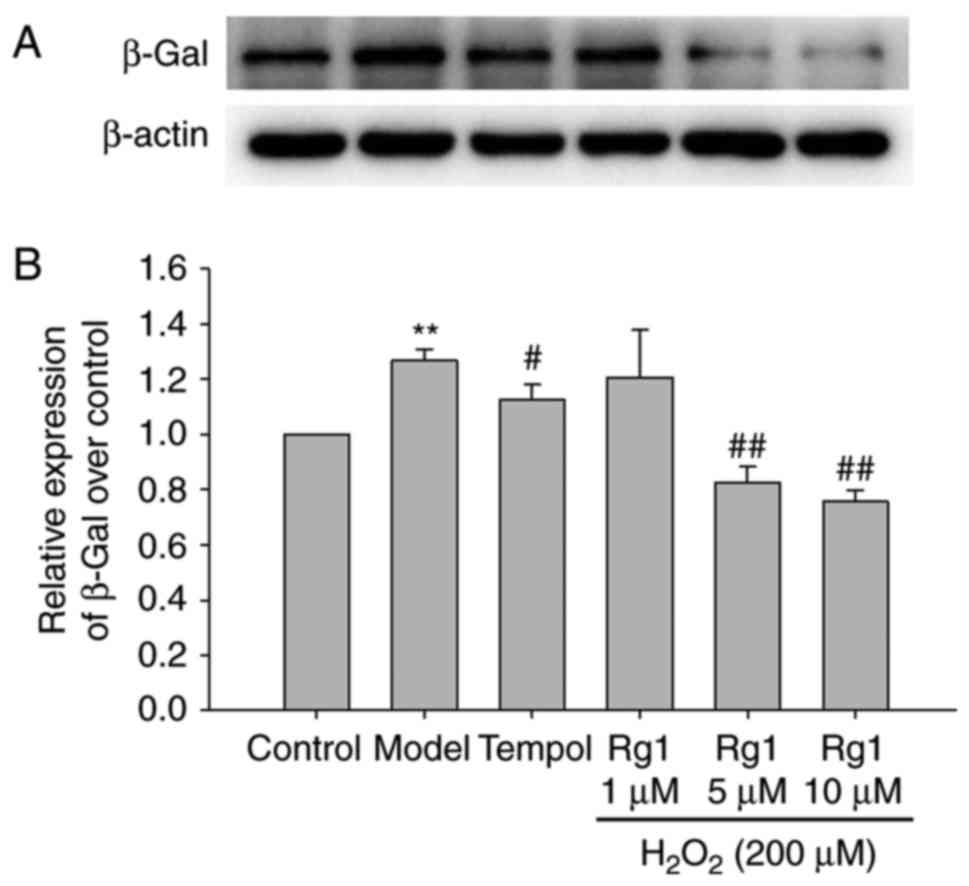
Rg1 decreases the expression of
senescence-associated β-gal in H2O2-treated
hippocampal neurons
Senescent cells exhibit increased
senescence-associated β-Gal activity (26). In order to observe the effect of
Rg1 on H2O2-induced senescence in hippocampal
neurons, the expression of β-Gal was detected by western blot
analysis. The results showed that H2O2 (200
µM) treatment significantly increased the expression of
β-Gal in the hippocampal neurons compared with the control group
(Fig. 3A and B; P<0.01).
Compared with the H2O2-treated group, tempol
(100 µM) and Rg1 (5 and 10 µM) treatments
significantly decreased the expression of β-Gal in the hippocampal
neurons (Fig. 3A and B; P<0.05
and P<0.01, respectively).
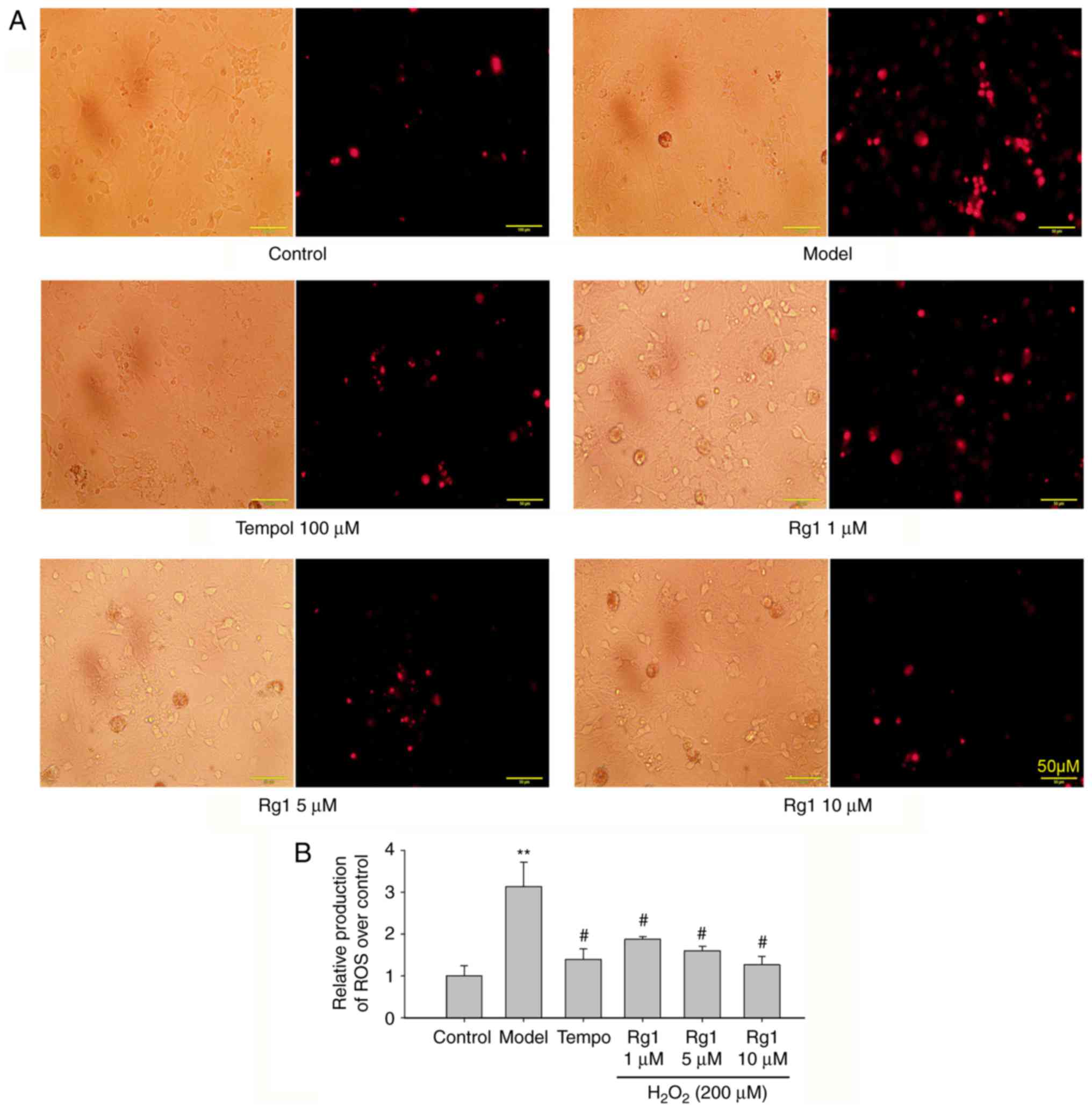
Rg1 reduces ROS generation in
H2O2-treated hippocampal neurons
H2O2 exposure induces cell
redox balance disorder, which may cause neuronal senescence and
damage. Therefore, ROS production was detected using DHE staining.
Compared with the control group, H2O2
treatment significantly increased the production of ROS in the
hippocampal neurons (Fig. 4A and
B; P<0.01). Compared with the
H2O2-treated group, tempol (100 µM)
and ginsenoside Rg1 (1, 5 and 10 µM) treatments
significantly reduced the production of ROS in hippocampal neurons
(Fig. 4A and B; P<0.05).
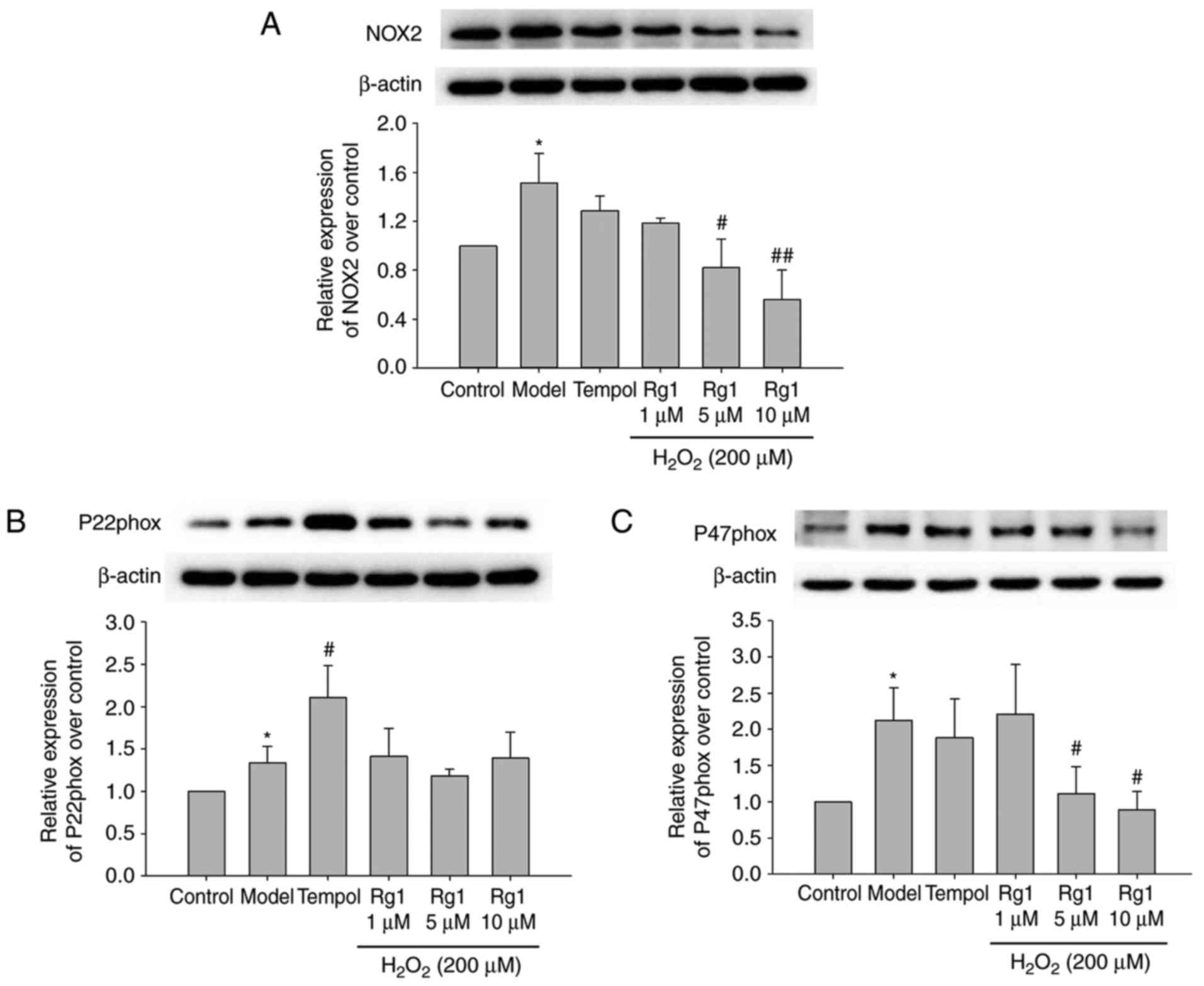
Effects of Rg1 on the expression of NOX2,
p22phox and p47phox in H2O2-treated
hippocampal neurons
To confirm whether NADPH oxidase is involved in
H2O2-induced neuronal senescence and damage
in primary hippocampal neurons, the expression levels of NOX2,
p22phox and p47phox in hippocampal neurons were measured by
immunoblotting. The results showed that, compared with the control
group, the expression levels of NOX2, p22phox and p47phox were
significantly increased in the H2O2-treated
group (Fig. 5A-C; P<0.05).
Compared with the model group, Rg1 (5 and 10 µM) treatment
significantly reduced the expression levels of NOX2 and p47phox in
the H2O2-treated hippocampal neurons, whereas
tempol (100 µM) treatment had no significant influence on
their expression. The results showed that Rg1 had no significant
influence on the expression of p22phox, whereas tempol treatment
increased the expression of p22phox in the
H2O2-treated hippocampal neurons (Fig. 5B; P<0.05).
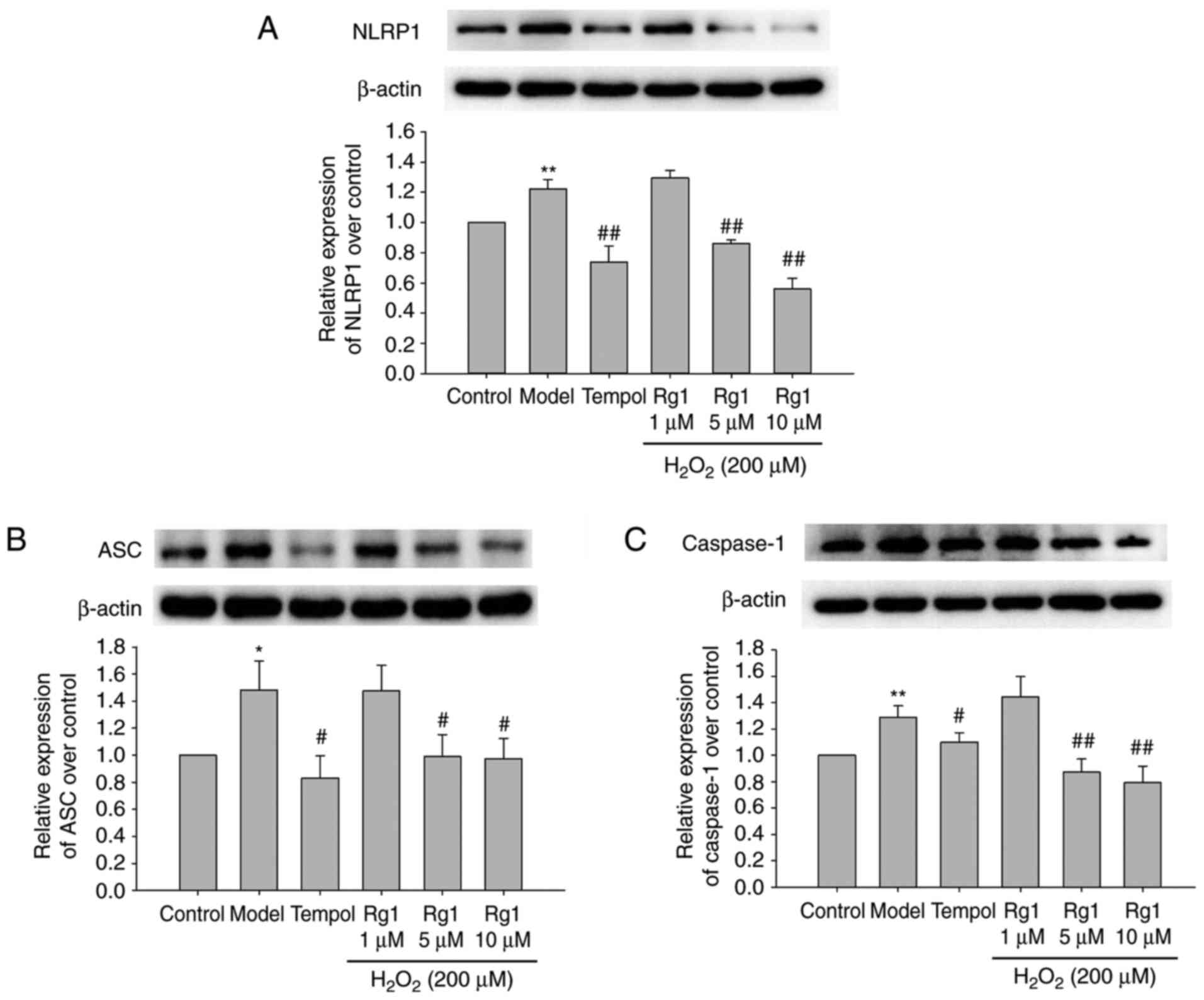
Effect of Rg1 on the expression levels of
NLRP-1, ASC and caspase-1 in H2O2-treated
hippocampal neurons
To confirm whether Rg1 regulated NLRP1 inflammasome
activation, the expression levels of NLRP1, ASC and caspase-1 in
primary hippocampal neurons were detected by immunoblotting. The
results showed that, compared with the control group, the
expression levels of NLRP-1, ASC and caspase-1 were significantly
increased in the H2O2-treated group. Compared
with the model group, tempol (100 µM) and Rg1 (5 and 10
µM) treatments significantly decreased the expression levels
of NLRP1, ASC and caspase-1 in the
H2O2-treated hippocampal neurons (Fig. 6A-C; P<0.05).
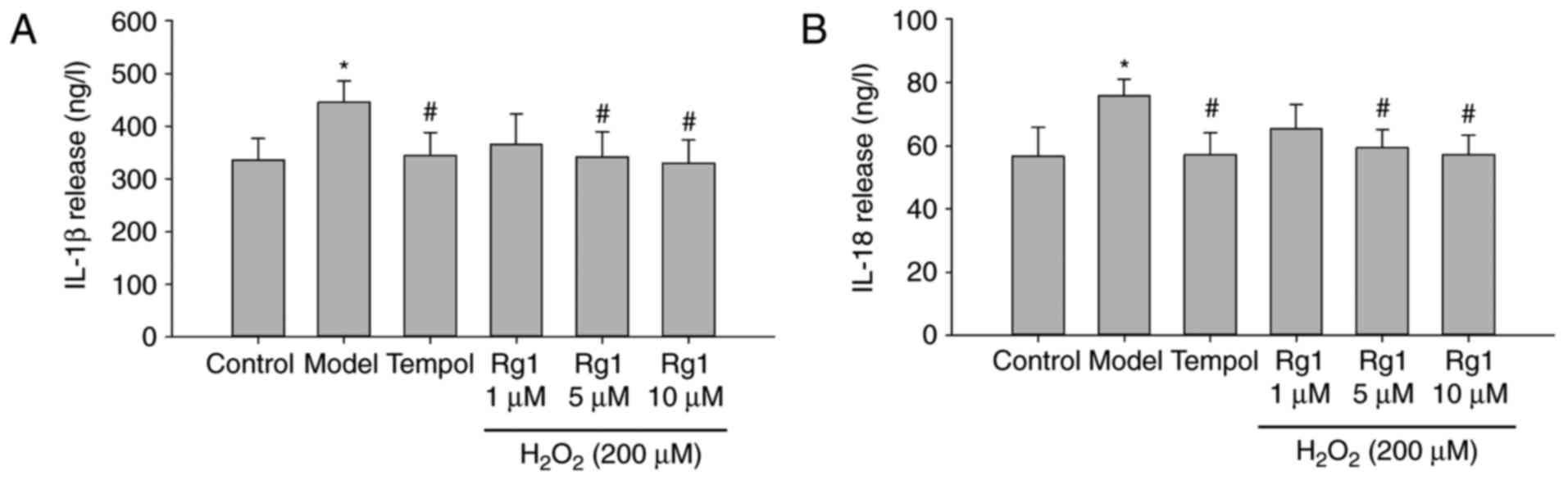
Rg1 decreases the release of IL-1β and
IL-18 in H2O2-treated hippocampal
neurons
To confirm the effect of Rg1 on downregulation of
the NLRP1 inflammasome, the levels of IL-1β and IL-18 released in
supernatants were examined by ELISA. The results showed that,
compared with the control group, the release of IL-1β and IL-18 was
significantly increased in the H2O2-treated
group (Fig. 7A and B; P<0.05).
Compared with the model group, tempol (100 µM) and Rg1 (5
and 10 µM) treatments significantly reduced the release of
IL-1β and IL-18 in supernatants from the
H2O2-treated hippocampal neurons (Fig. 7A and B; P<0.05).
Discussion
It has been reported that oxidative stress and
neuroinflammation are involved in neuronal senescence and
neurodegenerative diseases, including Alzheimer’s disease (27). The effect of Rg1 is largely
associated with its antioxidant and other scavenging properties,
for example, inhibiting the expression of inducible nitric oxide
synthase and the overgeneration of nitric oxide. However, whether
Rg1 can alleviate H2O2-induced senescence and
neuronal damage, and its mechanism, warrants further investigation.
In the present study, it was demonstrated that
H2O2 treatment significantly induced
oxidative stress damage and apoptosis in primary hippocampal
neurons, and the NOX2-NLRP1 inflammasome pathway was important in
H2O2-induced neuronal senescence and damage.
In addition, Rg1 significantly reduced the production of ROS and
the expression of NOX2 and the NLRP1 inflammasome, and inhibited
neuronal senescence and damage in the
H2O2-treated hippocampal neurons.
Oxidative stress is caused by the imbalance of redox
in the body, resulting in excessive ROS generation. The
accumulation of ROS can induce neuronal oxidative stress damage and
is involved in the development of ageing-related neurodegenerative
diseases (28). In the process of
ageing, excessive ROS can lead to destruction of neuronal structure
and impairment of learning and memory (29,30). H2O2
treatment is often used to simulate ROS accumulation-induced
neuronal damage (31). In the
present study, the results showed that H2O2
treatment significantly increased the generation of ROS, promoted
neuronal apoptosis and increased the expression of
senescence-associated β-Gal in hippocampal neurons.
Ginseng has been used as an anti-ageing drug for
thousands of years in China. Numerous studies have shown that Rg1,
a main active ingredient in ginseng, has antioxidant and
anti-ageing effects, in addition to promoting cognitive function
and improving immunity (32,33). Rg1 protects against neuronal
damage induced by various mechanisms, including oxidative stress
and inflammatory responses (20,34). Tempol is an effective antioxidant
and can attenuate the damage caused by excessive accumulation of
ROS (35). In the present study,
the results showed that both Rg1 and tempol significantly decreased
ROS generation and inhibited the expression of β-Gal in
H2O2-treated hippocampal neurons. In
addition, the Hoechst 33258 staining showed that Rg1 (5 and 10
µM) had a significant effect on
H2O2-induced hippocampal neuron apoptosis,
whereas Rg1 (1 µM) had no effect on the neuronal apoptosis.
Caspase-3 is the most important terminal shear enzyme in the
process of apoptosis (33). To
confirm the anti-apoptotic effect of Rg1, the effects of Rg1 (5
µM) on the expression of caspase-3 were examined by
immunofluorescence. The results showed that Rg1 (5 µM)
significantly decreased the expression of caspase-3 in
H2O2-induced hippocampal neurons. These
results indicate that Rg1 may decrease ROS generation and inhibit
neuronal senescence and apoptosis. However, the mechanism
underlying the effect of Rg1 on reducing the production of ROS
remains to be fully elucidated.
NOX is the most important enzyme system involved in
the generation of ROS and has been shown to be important in various
neurological diseases (28). NOX2
is a major source of ROS involved in the development of age-related
cognitive dysfunction (28). The
expression of NOX2 has been found to be increased in the brains of
ageing mice (36). NOX2 consists
of a membrane-bound catalytic core, including NOX2 (gp91phox) and
p22phox), and several cytosolic subunits, including p40phox,
p47phox, p67phox and rac1 (37).
NOX2 is activated when the cytosolic subunits translocate to the
membrane and combine with gp91phox (37,38). There is a linear relationship
between the activity of NOX2 and cognitive decline in aged mice
(39). In addition, NOX
contributes to oxidative stress and neuronal apoptosis.
NOX-deficient mice exhibit reduced injury following stroke
(40). It is not entirely clear
whether Rg1 can reduce NOX2 and, thus, reduce the production of ROS
in H2O2-treated hippocampal neurons. In the
present study, the results showed that the expression levels of
NOX2, p22phox and p47phox were significantly increased in
H2O2-treated hippocampal neurons. Rg1
treatment significantly reduced the expression levels of NOX2 and
p47phox in H2O2-treated hippocampal neurons,
whereas tempol (100 µM) treatment had no significant
influence on their expression. The results also showed that Rg1 had
no significant influence on the expression of p22phox, whereas
tempol treatment increased the expression of p22phox in
H2O2-treated hippocampal neurons. Therefore,
Rg1 may protect against neuronal ageing and damage by scavenging
ROS and inhibiting the formation of ROS derived from NOX2.
Neuroinflammation can accelerate brain ageing,
causing neuronal damage and cognitive deficits (41). The chronic, progressive
pro-inflammatory response is an important feature of the ageing
process and the leading cause of neuronal apoptosis (42). Inflammasomes are multimeric
proteins in the cytoplasm of a wide variety of cells (43). Among them, the NLRP1 inflammasome
was the first characterised and is expressed widely in the body,
particularly in neurons (44,45). The NLRP1 inflammasome consists
mainly of NLRP1, caspase-1 and ASC proteins. NLRP1 can activate the
key regulatory agent caspase-1 through its own oligomerization.
Activated caspase-1 can further cleave pro-IL-18 and pro-IL-1β into
active IL-18 and IL-1β, which in turn promote inflammatory
responses and apoptosis (46). It
has been reported that the NLRP3 inflammasome can be activated by
excessive ROS production (47).
Our previous study showed that Rg1 protected against chronic
dexamethasone-induced neuronal degeneration by inhibiting NLRP-1
inflammasomes in mice (19). It
has also been reported that Rg1 can protect against
H2O2-induced neuronal apoptosis (33). These studies suggest that Rg1 has
an anti-oxidative effect and can inhibit the NLRP1 inflammasome in
hippocampal neurons. However, whether Rg1 can down-regulate the
NOX2-mediated generation of ROS and thereby inhibit the NLRP1
inflammasome in H2O2-treated hippocampal
neurons remained to be elucidated. The present study found that
H2O2 treatment significantly increased the
expression levels of NLRP-1, ASC and caspase-1 in hippocampal
neurons and the levels of IL-1β and IL-18 released into the
supernatant. These results suggest that the accumulation of ROS can
activate the NLRP1 inflammasome, which may be involved in
senescence and damage in hippocampal neurons. The results also
showed that tempol and Rg1 treatments significantly decreased the
expression levels of NLRP1, ASC and caspase-1, and reduced the
levels of IL-1β and IL-18 released into the supernatant. A previous
study showed that Rg1 attenuated H2O2-induced
neuronal oxidative stress and apoptosis via the downregulation of
caspase-3, Rho-associated kinase1 activation and myosin light chain
(Ser-19) phosphorylation (33),
whereas the present study suggested that Rg1 alleviated
H2O2-induced neuronal oxidative stress and
apoptosis via the inhibition of NOX2-derived ROS generation and
downregulation of the NLRP1 inflammasome.
Overall, the present study demonstrated that Rg1 has
antioxidant effects and can reduce the expression of NOX2 and the
production of ROS, which in turn inhibits the activation of the
NLRP1 inflammasome in H2O2-treated
hippocampal neurons. These findings provide support for the
hypothesis that Rg1 reduces NOX2-mediated ROS generation and
inhibits NLRP1 inflammasome activation. However, whether the
overexpression of NOX2 can activate the NLRP1 inflammasome and
accelerate neuronal senescence remains to be fully elucidated.
Other associated mechanisms of Rg1 on neuronal senescence
associated with the NOX2-NLRP1 inflammasome signalling pathway
warrant further investigation.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81671384 and 81371329) and
the Natural Science Foundation of Anhui Province Education
Department (grant no. KJ2016A357).
Availability of data and materials
The datasets used and analysed during the present
study are available from the corresponding author on reasonable
request.
Authors’ contributions
TZX analysed data, performed the experiments, and
was a major contributor in writing the manuscript. XYS, LLS and YLC
collated the data, BQZ contributed to the immunoblot analysis and
interpretation of the results. DKH was mainly responsible for the
immunofluorescence detection. WZL designed the study, critically
revised the manuscript for intellectually important content,
supervised the study and wrote the manuscript. All authors read and
approved the final submitted manuscript.
Ethics approval and consent to
participate
All experiments involving animals were approved by
the Ethics Committee of Laboratory Animals of Anhui Medical
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank Mr. Bao Li
(Synthetic Laboratory of Basic Medicine College, Anhui Medical
University) for their technical assistance.
References
|
1
|
Floyd RA and Hensley K: Oxidative stress
in brain aging. Implications for therapeutics of neurodegenerative
diseases. Neurobiol Aging. 23:795–807. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Joseph JA, Shukitt-Hale B, Casadesus G and
Fisher D: Oxidative stress and inflammation in brain aging:
Nutritional considerations. Neurochem Res. 30:927–935. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Calabrese V, Cornelius C, Mancuso C,
Lentile R, Stella AM and Butterfield DA: Redox homeostasis and
cellular stress response in aging and neurodegeneration. Methods
Mol Biol. 610:285–308. 2010. View Article : Google Scholar
|
|
4
|
Fan LM, Cahill-Smith S, Geng L, Du J,
Brooks G and Li JM: Aging-associated metabolic disorder induces
Nox2 activation and oxidative damage of endothelial function. Free
Radic Biol Med. 108:940–951. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ansari MA and Scheff SW: NADPH-oxidase
activation and cognition in Alzheimer disease progression. Free
Radic Biol Med. 51:171–178. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rehman SU, Shah SA, Ali T, Chung JI and
Kim MO: Anthocyanins reversed d-galactose-induced oxidative stress
and neuroinflammation mediated cognitive impairment in adult rats.
Mol Neurobiol. 54:255–271. 2017. View Article : Google Scholar
|
|
7
|
Zotova E, Bharambe V, Cheaveau M, Morgan
W, Holmes C, Harris S, Neal JW, Love S, Nicoll JA and Boche D:
Inflammatory components in human Alzheimer’s disease and after
active amyloid-β42 immunization. Brain. 136:2677–2696. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kaushal V, Dye R, Pakavathkumar P, Foveau
B, Flores J, Hyman B, Ghetti B, Koller BH and LeBlanc AC: Neuronal
NLRP1 inflammasome activation of Caspase-1 coordinately regulates
inflammatory interleukin-1-beta production and axonal
degeneration-associated Caspase-6 activation. Cell Death Differ.
22:1676–1686. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang B, Zhang Y, Xu T, Yin Y, Huang R,
Wang Y, Zhang J, Huang D and Li W: Chronic dexamethasone treatment
results in hippocampal neurons injury due to activate NLRP1
inflam-masome in vitro. Int Immunopharmacol. 49:222–230. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Faustin B, Lartigue L, Bruey JM, Luciano
F, Sergienko E, Bailly-Maitre B, Volkmann N, Hanein D, Rouiller I
and Reed JC: Reconstituted NALP1 inflammasome reveals two-step
mechanism of caspase-1 activation. Mol Cell. 25:713–724. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mawhinney LJ, de Rivero Vaccari JP, Dale
GA, Keane RW and Bramlett HM: Heightened inflammasome activation is
linked to age-related cognitive impairment in fischer 344 rats. BMC
Neurosci. 12:1232011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Choi AJ and Ryter SW: Inflammasomes:
Molecular regulation and implications for metabolic and cognitive
diseases. Mol Cells. 37:441–448. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Moon JS, Nakahira K, Chung KP, DeNicola
GM, Koo MJ, Pabón MA, Rooney KT, Yoon JH, Ryter SW, Stout-Delgado H
and Choi AM: NOX4-dependent fatty acid oxidation promotes NLRP3
inflammasome activation in macrophages. Nat Med. 22:1002–1012.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang L, Zhang J, Zheng K, Shen H and Chen
X: Long-term ginsenoside Rg1 supplementation improves age-related
cognitive decline by promoting synaptic plasticity associated
protein expression in C57BL/6J mice. J Gerontol A Biol Sci Med Sci.
69:282–294. 2014. View Article : Google Scholar
|
|
15
|
Sun XC, Ren XF, Chen L, Gao XQ, Xie JX and
Chen WF: Glucocorticoid receptor is involved in the neuroprotective
effect of ginsenoside Rg1 against inflammation-induced dopaminergic
neuronal degeneration in substantia nigra. J Steroid Biochem Mol
Biol. 155:94–103. 2016. View Article : Google Scholar
|
|
16
|
Attele AS, Wu JA and Yuan CS: Ginseng
pharmacology: Multiple constituents and multiple actions. Biochem
Pharmacol. 58:1685–1693. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu J, Mu X, Zeng J, Xu C, Liu J, Zhang M,
Li C, Chen J, Li T and Wang Y: Ginsenoside Rg1 prevents cognitive
impairment and hippocampus senescence in a rat model of
D-galactose-induced aging. PLoS One. 9:e1012912014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhu G, Wang Y, Li J and Wang J: Chronic
treatment with ginsenoside Rg1 promotes memory and hippocampal
long-term potentiation in middle-aged mice. Neuroscience.
292:81–89. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Y, Kan H, Yin Y, Wu W, Hu W, Wang M
and Li W and Li W: Protective effects of ginsenoside Rg1 on chronic
restraint stress induced learning and memory impairments in male
mice. Pharmacol Biochem Behav. 120:73–81. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y, Hu W, Zhang B, Yin Y, Zhang J,
Huang D, Huang R and Li W and Li W: Ginsenoside Rg1 protects
against neuronal degeneration induced by chronic dexamethasone
treatment by inhibiting NLRP-1 inflammasomes in mice. Int J Mol
Med. 40:1134–1142. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang B, Zhang Y, Wu W, Xu T, Yin Y, Zhang
J, Huang D and Li W: Chronic glucocorticoid exposure activates
BK-NLRP1 signal involving in hippocampal neuron damage. J
Neuroinflammation. 14:1392017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cao G, Xiao M, Sun F, Xiao X, Pei W, Li J,
Graham SH, Simon RP and Chen J: Cloning of a novel
Apaf-1-interacting protein: A potent suppressor of apoptosis and
ischemic neuronal cell death. J Neurosci. 24:6189–6201. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hu W, Zhang Y, Wu W, Yin Y, Huang D, Wang
Y and Li W and Li W: Chronic glucocorticoids exposure enhances
neurodegeneration in the frontal cortex and hippocampus via NLRP-1
inflammasome activation in male mice. Brain Behav Immun. 52:58–70.
2016. View Article : Google Scholar
|
|
24
|
Kazama K, Anrather J, Zhou P, Girouard H,
Frys K, Milner TA and Iadecola C: Angiotensin II impairs
neurovascular coupling in neocortex through NADPH oxidase-derived
radicals. Circ Res. 95:1019–1026. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cheng XL and Zhang JJ: Effect of edaravone
on apoptosis of hippocampus neuron in seizures rats kindled by
pentylenetetrazole. Eur Rev Med Pharmacol Sci. 18:769–774.
2014.PubMed/NCBI
|
|
26
|
Dong W, Cheng S, Huang F, Fan W, Chen Y,
Shi H and He H: Mitochondrial dysfunction in long-term neuronal
cultures mimics changes with aging. Med Sci Monit. 17:BR91–BR96.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Quintanilla RA, Orellana JA and von
Bernhardi R: Understanding risk factors for Alzheimer’s disease:
Interplay of neuroinflammation, connexin-based communication and
oxidative stress. Arch Med Res. 43:632–644. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cahill-Smith S and Li JM: Oxidative
stress, redox signalling and endothelial dysfunction in
ageing-related neurodegenerative diseases: A role of NADPH oxidase
2. Br J Clin Pharmacol. 78:441–453. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee KW, Woo JM, Im JY, Park ES, He L,
Ichijo H, Junn E and Mouradian MM: Apoptosis signal-regulating
kinase 1 modulates the phenotype of α-synuclein transgenic mice.
Neurobiol Aging. 36:519–526. 2015. View Article : Google Scholar
|
|
30
|
Verri M, Pastoris O, Dossena M, Aquilani
R, Guerriero F, Cuzzoni G, Venturini L, Ricevuti G and Bongiorno
AI: Mitochondrial alterations, oxidative stress and
neuroinflammation in Alzheimer’s disease. Int J Immunopathol
Pharmacol. 25:345–353. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou X, Su CF, Zhang Z, Wang CY, Luo JQ,
Zhou XW, Cai L, Yan L, Zhang W and Luo HM: Neuroprotective effects
of methyl 3,4-dihydroxybenzoate against H(2)O(2)-induced apoptosis
in RGC-5 cells. J Pharmacol Sci. 125:51–58. 2014. View Article : Google Scholar
|
|
32
|
Ong WY, Farooqui T, Koh HL, Farooqui AA
and Ling EA: Protective effects of ginseng on neurological
disorders. Front Aging Neurosci. 7:1292015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang Y, Liu Q, Xu Y, Zhang Y, Lv Y, Tan Y,
Jiang N, Cao G, Ma X, Wang J, et al: Ginsenoside rg1 protects
against oxidative stress-induced neuronal apoptosis through myosin
IIA-actin related cytoskeletal reorganization. Int J Biol Sci.
12:1341–1356. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu Q, Kou JP and Yu BY: Ginsenoside Rg1
protects against hydrogen peroxide-induced cell death in PC12 cells
via inhibiting NF-kB activation. Neurochem Int. 58:119–125. 2011.
View Article : Google Scholar
|
|
35
|
Aksu U, Yanar K, Terzioglu D, Erkol T, Ece
E, Aydin S, Uslu E and Çakatay U: Effect of tempol on redox
homeostasis and stress tolerance in mimetically aged drosophila.
Arch Insect Biochem Physiol. 87:13–25. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dugan LL, Ali SS, Shekhtman G, Roberts AJ,
Lucero J, Quick KL and Behrens MM: IL-6 mediated degeneration of
forebrain GABAergic interneurons and cognitive impairment in aged
mice through activation of neuronal NADPH oxidase. PLoS One.
4:e55182009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tammariello SP, Quinn MT and Estus S:
NADPH oxidase contributes directly to oxidative stress and
apoptosis in nerve growth factor-deprived sympathetic neurons. J
Neurosci. 20:RC532000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
DeLeo FR and Quinn MT: Assembly of the
phagocyte NADPH oxidase: Molecular interaction of oxidase proteins.
J Leukoc Biol. 60:677–691. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ali SS, Young JW, Wallace CK, Gresack J,
Jeste DV, Geyer MA, Dugan LL and Risbrough VB: Initial evidence
linking synaptic superoxide production with poor short-term memory
in aged mice. Brain Res. 1368:65–70. 2011. View Article : Google Scholar :
|
|
40
|
Walder CE, Green SP, Darbonne WC, Mathias
J, Rae J, Dinauer MC, Curnutte JT and Thomas GR: Ischemic stroke
injury is reduced in mice lacking a functional NADPH oxidase.
Stroke. 28:2252–2258. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu YZ, Nygard M, Kristensson K and
Bentivoglio M: Regulation of cytokine signaling and T-cell
recruitment in the aging mouse brain in response to central
inflammatory challenge. Brain Behav Immun. 24:138–152. 2010.
View Article : Google Scholar
|
|
42
|
Glass CK, Saijo K, Winner B, Marchetto MC
and Gage FH: Mechanisms underlying inflammation in
neurodegeneration. Cell. 140:918–934. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zou P, Liu X, Li G and Wang Y: Resveratrol
pretreatment attenuates traumatic brain injury in rats by
suppressing NLRP3 inflammasome activation via SIRT1. Mol Med Rep.
17:3212–3217. 2018.
|
|
44
|
Yin Y, Yan Y, Jiang X, Mai J, Chen NC,
Wang H and Yang XF: Inflammasomes are differentially expressed in
cardiovascular and other tissues. Int J Immunopathol Pharmacol.
22:311–322. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tan CC, Zhang JG, Tan MS, Chen H, Meng DW,
Jiang T, Meng XF, Li Y, Sun Z, Li MM, et al: NLRP1 inflammasome is
activated in patients with medial temporal lobe epilepsy and
contributes to neuronal pyroptosis in amygdala kindling-induced rat
model. J Neuroinflammation. 12:182015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fann DY, Lee SY, Manzanero S, Tang SC,
Gelderblom M, Chunduri P, Bernreuther C, Glatzel M, Cheng YL,
Thundyil J, et al: Intravenous immunoglobulin suppresses NLRP1 and
NLRP3 inflammasome-mediated neuronal death in ischemic stroke. Cell
Death Dis. 4:e7902013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Aminzadeh M, Roghani M, Sarfallah A and
Riazi GH: TRPM2 dependence of ROS-induced NLRP3 activation in
Alzheimer’s disease. Int Immunopharmacol. 54:78–85. 2018.
View Article : Google Scholar
|