Introduction
Breast cancer (BC) is the most commonly diagnosed
cancer, which holds the highest mortality rate in women around the
world (1-4). It has been documented that radiation
therapy and chemotherapy have been employed in treating BC, which
can lead to the improvement of prognosis (5). However, chemotherapy resistance and
metastasis may lead to failure of BC therapy (6). BC is a complex disease, which is
influenced by several environmental factors and genetic
alterations. The environmental factors include unhealthy diet and
psychology, environmental contamination and radiation. Gene
dysregulation may lead to the spreading of tumor cells from the
primary neoplasm to distant sites (7). Therefore, it is urgent to gain
further insight into the gene expression alterations and molecular
mechanism in BC progression that may aid better therapeutic
developments for BC patients.
MicroRNAs (miRNAs/miRs) are small noncoding
single-stranded RNA molecule of 20-25 nucleotides in length that
regulate gene expression (8). It
has been demonstrated that miRNAs function as oncogenes or tumor
suppressors and modulate a variety biological processes, including
cell proliferation, migration, invasion, metastasis, apoptosis and
differentiation (9-13). Dysregulation of several miRNAs has
been demonstrated in BC, including miR-122 (14), miR-34a (15) and miR-155 (16), which associated with the
development and progression of malignancy. miR-135 has been
reported to serve important roles in the development of various
types of cancer, including colorectal cancer, non-small cell lung
cancer and renal cancer (17-19). It was reported that miR-135 was
highly expressed in metastatic breast tumors and promotes cell
migration and invasion by targeting HOXA10 (20). Furthermore, a considerable
upregulation of miR-135 in colorectal adenomas and carcinomas was
observed, which is significantly correlated with low APC mRNA
levels and miR-135 serves as a tumor suppressor in breast cancer
metastasis (17). In addition,
miR-135 inhibits prostate cancer cell growth and migration by
targeting the epidermal growth factor receptor (EGFR) (21). However, it remains unclear whether
miR-135 is involved in BC progression. Therefore, the development
of novel effective therapeutic strategies is essential and
urgent.
It is known that epithelial-mesenchymal transition
(EMT) is a complex biological process. Specifically, cells
undergoing EMT have increased expression of mesenchymal molecules
including snail, slug and vimentin and decreased expression of the
epithelial adhesion marker epithelial (E)-cadherin, leading to
enhanced motility and metastasis (22). Increasing evidence has
demonstrated that aberrant activation of Wnt/β-catenin signaling
promoted cell proliferation and is associated with poor prognosis
of BC patients (23-25). Aberrant activation of the
Wnt/β-catenin signaling pathway is identified and promoted tumor
progression in a number of types of human cancer (26-30).
Therefore, miR-135 expression was measured in BC
cells and biological functions of miR-135 were investigated in BC.
In the present study, it was demonstrated that expression of
miR-135 was significantly reduced in BC cells and overexpression of
miR-135 inhibited cell proliferation, migration, invasion and
metastasis. Finally, it was demonstrated that miR-135 suppression
was followed by Wnt/β-catenin signaling pathway activation.
Collectively, the results of the present study demonstrated that
miR-135 suppresses the progression of BC by regulating the
Wnt/β-catenin signaling pathway.
Materials and methods
Cell lines and transfection
Human breast cancer cell lines MDA-MB-468,
MDA-MB-231 and normal epithelial cell line MCF-10A were purchased
from the Cell Type Culture Collection of the Chinese Academy of
Sciences (Shanghai, China) and routinely maintained in RPMI-1640
medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific,
Inc.) and human breast cancer cell line MCF-7 was cultured in RPMI
1640 medium. All cells were incubated in a humidified 5%
CO2 atmosphere at 37°C. The miR-135 mimic, miR-135
inhibitor or scrambled miRNA control which expressed green
fluorescent protein (GFP) were transfected to the MDA-MB-468 and
MCF-7 cells with Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer’s protocol. The
sequences were as follows: miR-135 mimic forward, 5′-UAU GGC UUU
UCA UUC CUA UGU GA-3′ and reverse, 5′-ACA UAG GAA UGA AAA GCC AUA
UU-3′; miR-135 inhibitor forward, 5′-UCA CAU AGG AAU GAA AAG CCA
UA-3′ and reverse, 5′-CAG UAC UUU UGU GUA GUA CAA-3′; and scrambled
miRNA control forward, 5′-UUC UCC GAA CGU GUC ACG UTT-3′ and
reverse, 5′-ACG UGA CAC GUU CGG AGA ATT-3′. A total of 2 days
later, cells were collected and stored at −80°C.
RNA extraction and quantitative
polymerase chain reaction (qPCR)
Total RNA was extracted from cell lines using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer’s protocol. TaqMan MicroRNA Reverse Transcription
kit (Takara Bio, Inc., Otsu, Japan) was used to synthesize cDNA
from total RNA at 37°C for 30 min. qPCR was performed on an Applied
Biosystems StepOne Plus Real-Time PCR System (Takara Bio, Inc.,
Otsu, Japan). The PCR conditions consisted of 95°C for 30 sec,
followed by 40 cycles of amplification (95°C for 3 sec and 60°C for
30 sec). The primer sequences were as follows: miR-135 forward,
5′-TCT GCT GTG GCC TAT GGC TT-3′, reverse, 5′-CTG TAG CCC ATG GCT
TTT AGC-3′ and U6 forward, 5′-CTC GCT TCG GCA GCA CA-3′, reverse,
5′-AAC GCT TCA CGA ATT TGC GT-3′. The small nuclear RNA U6 was used
as control. Fold-changes for mRNA expressions were calculated using
the 2−ΔΔCq method (31) and all experiments were performed
in at least triplicate.
Western blot analysis
For western blot analysis, total protein was
extracted from transfected cells using radioimmunoprecipitation
assay (RIPA) buffer (Beijing Solarbio Science & Technology Co.,
Ltd., Beijing, China) and the supernatants were collected with
centrifugation at 12,000 × g at 4°C for 20 min. Protein
concentration was measured with a Bicinchoninic acid Protein Assay
kit (Beyotime Institute of Biotechnology, Haimen, China).
Subsequently, equal amounts of protein (30 µg) from each
sample were separated by 10% SDS-PAGE and transferred onto
polyvinylidene difluoride membranes. The membranes were blocked
with 5% non-fat milk for 1 h at room temperature and then incubated
with primary antibodies overnight at 4°C. The specific primary
antibodies were as follows: Anti-wnt (ab15251; 1:1,000; Abcam,
Cambridge, UK), p-GSK3 (ab75745; 1:1,000; Abcam), GSK3 (ab32391;
1:1,000; Abcam), β-catenin (ab32572; 1:1,000; Abcam). Following the
membranes were washed with TBST containing 0.2% Tween-20 three
times and incubated with the horseradish peroxidase-conjugated
anti-rabbit secondary antibody (cat. no. ab6721; 1:2,000; Abcam) at
room temperature for 1 h. The blot was visualized by an odyssey
infrared imaging system with ECL western blotting substrate kit
(Amersham; GE Healthcare, Chicago, IL, USA). Expression of GAPDH
was used as a loading control.
Cell proliferation colony formation
assay
Cell proliferation was measured by the MTT assay
(Nanjing KeyGEN Biotech, Co., Ltd., Nanjing, China). MDA-MB-468 and
MCF-7 cells (1.0×104/well) were plated in a 6-well plate
and transfected with miR-135 mimics, miR-135 mimics negative
control (NC), miR-135 inhibitors and miR-135 inhibitors NC. At 24,
48 and 72 h of incubation, cells were washed with PBS, and dimethyl
sulfoxide used to dissolve the purple formazan. Then, 20 µl
MTT were added into each well for 4 h. The absorbance of the
resulting solution was assessed at 490 nm wavelength with a
microplate reader (BioTek Instruments, Inc., Winooski, VT, USA).
For colony formation assay, cells were subcultured in a 6-well
plate for 2 weeks. The cells were washed twice with PBS, fixed in
4% paraformaldehyde at room temperature for 30 min and stained with
5% crystal violet for 4 h at room temperature, and counted under a
light microscope.
Wound healing assay
For the wound healing assay, MDA-MB-468 and MCF-7
were seeded into 6-well plates at density of 1×105 cells
per well following transfection and cultured in fresh culture the
confluence reached 90%. Furthermore, sterile pipette tips was used
to scratch the wound uniformly and incubated at 37°C for 0, 24 and
48 h. The cell migration ability was assessed using a light
microscope and images were captured at 0, 24 and 48 h following
scraping. Each experiment was conducted in triplicate.
Transwell assay
The migration and invasion of transfected MDA-MB-468
and MCF-7 cells were evaluated by using Boyden chambers consisting
of transwell with 8-µm pore size polycarbonate membrane
filters. For migration assay, cells were plated at a density of
2×105 cells/well in the upper chamber of a transwell
filter. For invasion assay, matrigel was added to the inner
surface. Cells were placed in RPMI-1640 medium on the top chamber
and the lower chambers were filled with RPMI-1640 medium
supplemented with 10% FBS. After incubation for 48 h, nonmigrated
or noninvasive cells on the upper surface were removed carefully
with a cotton swab. The cells bound to the lower side of the filter
were washed twice with PBS and fixed with 5% gluteraldehyde at room
temperature for 10 min. Fixed cells were washed twice with PBS and
fixed 4% paraformaldehyde for 30 min at room temperature. After
that, cells were stained with 0.1% crystal violet for another 30
min at room temperature. The migration and invasion cells were
counted and imaged in six random fields under a light microscope at
×200 magnification.
Immunofluorescence assay
Cells (1×105) were seeded on 6-well
plates and incubated for 48 h. After that, MDA-MB-468 and MCF-7
cells were fixed with 4% paraformaldehyde for 30 min at room
temperature and incubated with the primary antibodies at room
temperature for 2 h after blocking cell with 3% bovine serum
albumin (Gibco; Thermo Fisher Scientific, Inc.). The specific
primary antibodies are as follows: Anti-wnt (ab15251; 1:1,000;
Abcam), p-GSK3 (ab75745; 1:1,000; Abcam), GSK3 (ab32391; 1:1,000;
Abcam), β-catenin (ab32572; 1:1,000; Abcam). Thereafter, an Alexa
Fluor 594-Conjugated Anti-Rabbit IgG (H+L) (SA00006-4; 1:1,000;
ProteinTech Group, Inc., Chicago, IL, USA) second antibody was
incubated for 1 h at room temperature. The nuclei were
counterstained with DAPI for 5 min at room temperature. The slides
were examined under a confocal microscopy.
Xenografted tumor model
A total of 30 female nude mice (6 weeks old, 18-22
g) were provided by the Nanjing Medical University and housed under
germ free conditions. Animal care and use were carried out
according to the ethical guidelines by Nanjing Medical University
Animal Care and Use Committee and approved by the Nanjing Medial
University Experimental Animal Ethics Committee and the permit
number is SYXK(su)2016-0016. Mice were maintained in laminar
airflow chambers under specific pathogen-free conditions in a
temperature controlled room (24°C) under a 12 h light/dark cycle.
They were provided free access to water and standard laboratory
rodent chow. MCF-7 cells (5×106) transfected with an
miR-135 mimic, miR-135 inhibitor or the control vector were
inoculated subcutaneously into the right flanks of nude mice,
respectively. The volume of tumor was measured every week and
calculated using the equation volume (mm3)=length x
(width)2/2. After five weeks, the mice were sacrificed
by cervical dislocation and stored in −80°C for further use. In
addition, xenografted tumor tissue samples were obtained and were
fixed in formalin for 24 h at room temperature and embedded in
paraffin. Hematoxylin and eosin (H&E) staining was performed to
obtain histological samples of the tumor.
H&E staining
The tumors tissues were fixed in 4%
para-formaldehyde for 30 min at room temperature and then embedded
in paraffin. The 4-µm sections were stained with H&E
staining for 20 min at room temperature. At least three different
sections of tumor tissues were examined for each group using a
light microscope to assess the histopathological changes.
Terminal
deoxynucleotidyl-transferase-mediated dUTP nick end labelling
(TUNEL) staining
The apoptosis of paraffin-embedded tumor 4-µm
sections was detected using a TUNEL assay kit. Briefly, fixed and
paraffin-embedded sections were dewaxed, then proteinase K was
added into for 20 min at room temperature. The sections had 3%
H2O2 added following washing with PBS and
then were incubated with the equilibration buffer and terminal
deoxynucleotidyl transferase (TdT) enzyme for 1 h at room
temperature in the dark. Sections were treated with 3%
H2O2 for 5 min and horseradish peroxidase for
30 min following washing with PBS. Finally, sections were incubated
with DAB for 5 min at room temperature to evaluate the tissue
peroxidase activity and five fields per view were analyzed with a
light microscope.
Statistical analysis
All data were expressed as the mean ± standard
deviation and statistical analyses were performed with 19.0 SPSS
software (IBM, Corp., Armonk, NY, USA); three independent
experiments were performed. Statistical analyses of pairs of
samples were performed using the Student’s t-test. Multiple group
comparisons were carried out by one-way analysis of variance with
Bonferroni’s post-hoc test. P<0.01 was considered to indicate a
statistically significant difference.
Results
miR-135 is downregulated in BC cell
lines
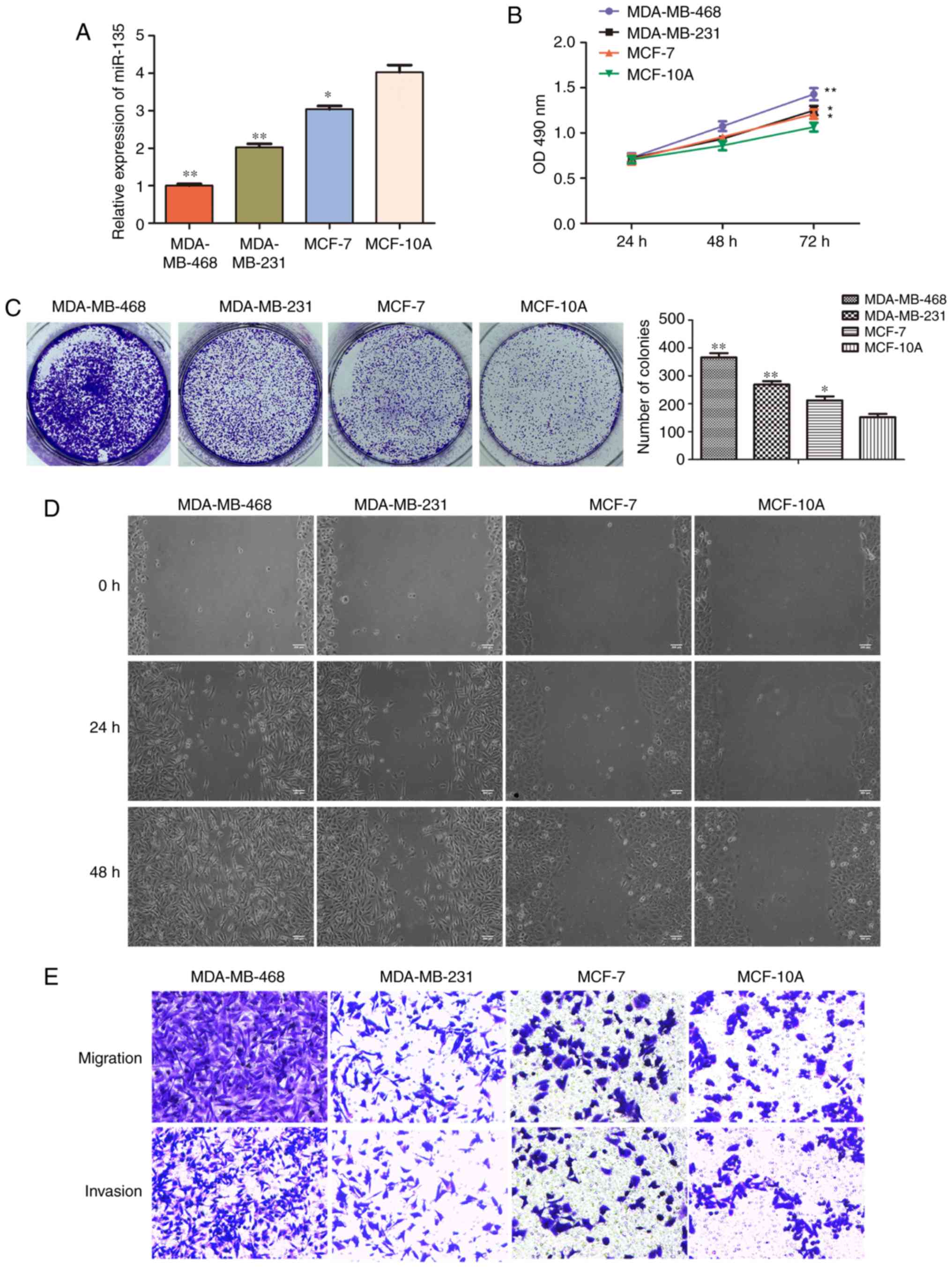
To investigate the role of miR-135 in BC
pathogenesis, reverse transcription (RT)-qPCR was used to quantify
its expression in different cell lines, including BC cell lines
MDA-MB-468, MDA-MB-231, MCF-7 and normal immortalized MCF-10A
breast cell line. RT-qPCR analysis demonstrated that expression was
significantly downregulated in BC cell lines (MDA-MB-468,
MDA-MB-231 and MCF-7) compared with breast epithelial cell line
MCF-10A (P<0.05; Fig. 1A).
MTT, colony formation, wound healing and transwell assay were used
to evaluate cell proliferation, migration and invasion ability
(Fig. 1B-E). The results
indicated that the BC cell proliferation, migration and invasion
ability were stronger than MCF-10A cells.
miR-135 inhibited BC cell proliferation
in vitro
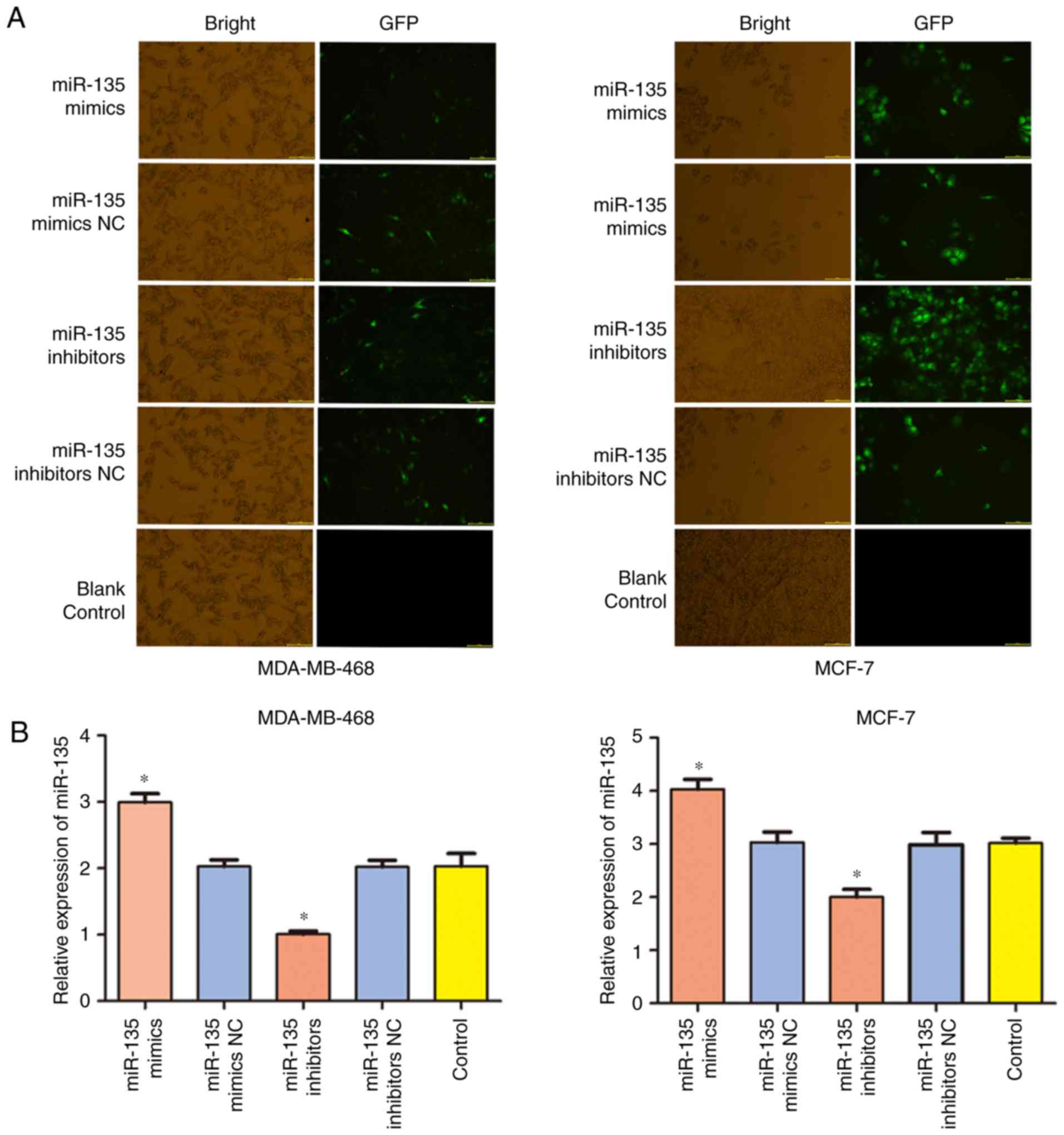
To determine the role of miR-135 in regulating cell
proliferation, miR-135 mimics, mimics NC, miR-135 inhibitors and
inhibitors NC were separately transfected into MDA-MB-468 and MCF-7
cells, respectively. The transfection efficiency of miR-135 was
examined by green fluorescent protein (GFP) and RT-qPCR assay. As
presented in Fig. 2A and B, cells
transfection of miR-135 mimic significantly increased the level of
miR-135 (P<0.05), while miR-135 inhibitors significantly
inhibited the expression of miR-135 in MDA-MB-468 and MCF-7 cells
(P<0.05). MTT and colony formation assays were performed to
investigate the effect of miR-135 on cell proliferation following
transfection for 48 h. As presented in Fig. 2C, the results revealed that the
miR-135 inhibitor caused a decrease in optical density, which
indicated that the proliferation ability of cells was significantly
increased in the miR-135 inhibitor group compared with the control
group, while upregulation of the expression of miR-135 attenuated
the proliferation of MDA-MB-468 and MCF-7 cells. Furthermore,
MDA-MB-468 and MCF-7 cells which transfected miR-135 mimics
exhibited fewer and smaller colonies compared with the control
cells (Fig. 2D). Taken together,
these results revealed that miR-135 overexpression inhibited cell
proliferation of BC in vitro.
miR-135 suppresses the migration and
invasion in BC cells
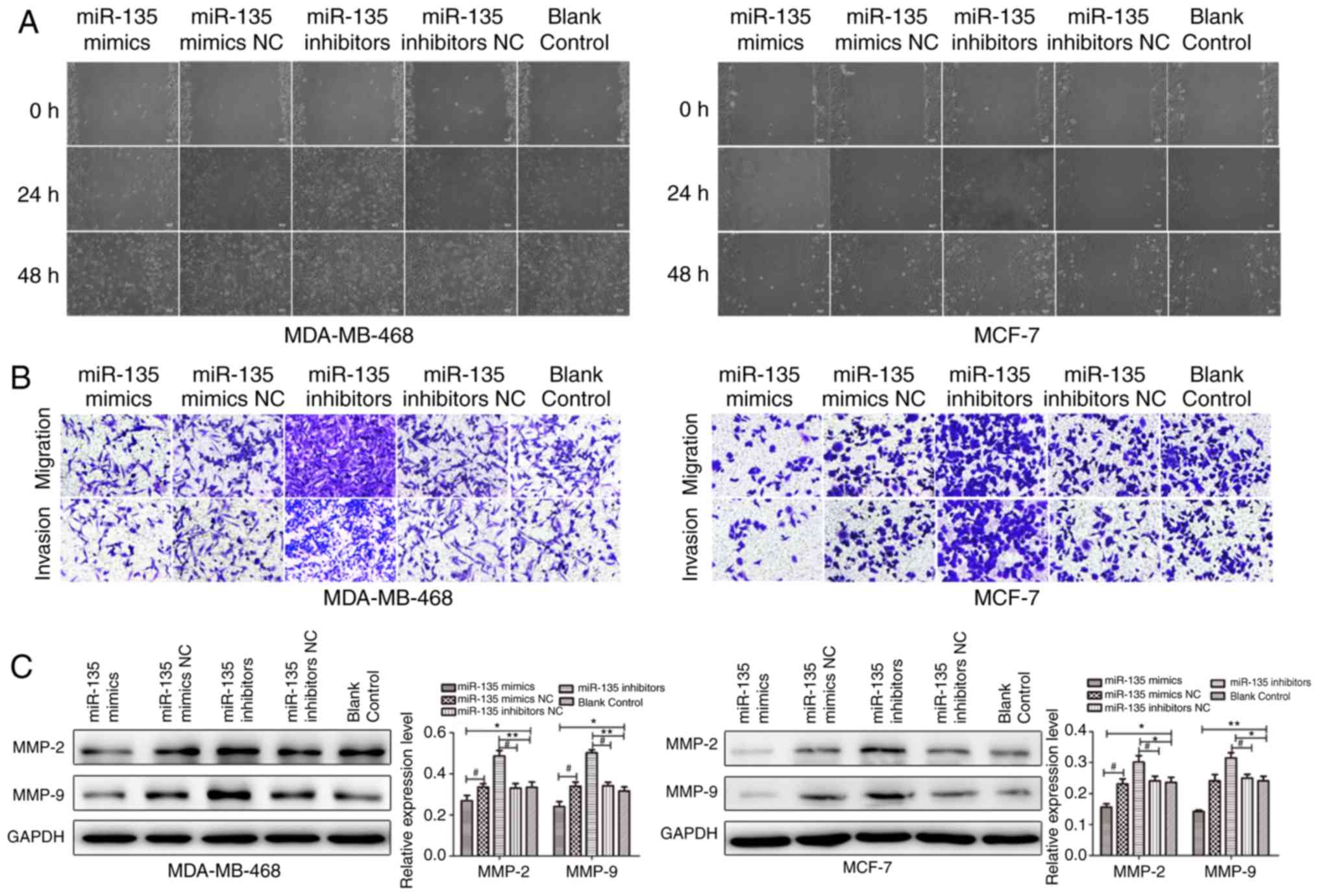
Given that cellular migration and invasion are key
processes underlying metastasis, whether miR-135 could affect BC
was investigated. To further validate the role of miR-135 in cell
migration and invasion in vitro, cell migration and invasion
in MDA-MB-468 and MCF-7 cells following transfection with miR-135
mimic NC, miR-135 mimics, miR-135 inhibitor NC and miR-135
inhibitor was determined by wound healing assay and transwell
assays. As presented in Fig. 3A,
ectopic restoration of miR-135 delayed the migration rate of cells
from the wound area, compared with the control group, while miR-135
inhibitor promoted the migration potential of MDA-MB-468 and MCF-7
cells. Transwell assay demonstrated that the migration and invasion
capacity of cells transfected with miR-135 mimics was markedly
suppressed compared with the control group (Fig. 3B). Moreover, the protein levels of
matrix metalloproteinase (MMP)-2 and MMP-9 were examined in
MDA-MB-468 and MCF-7 cells. It was demonstrated that miR-135 mimic
suppressed the levels of MMP-2 and MMP-9, while the expression of
MMP-2 and MMP-9 were remarkably promoted in cells which were
transfected with miR-135 inhibitors (Fig. 3C). The results suggested that
miR-135 may have the ability to inhibit migration and invasion in
BC cells.
Effects of miR-135 on xenograft tumour
growth of breast cancer cells in vivo
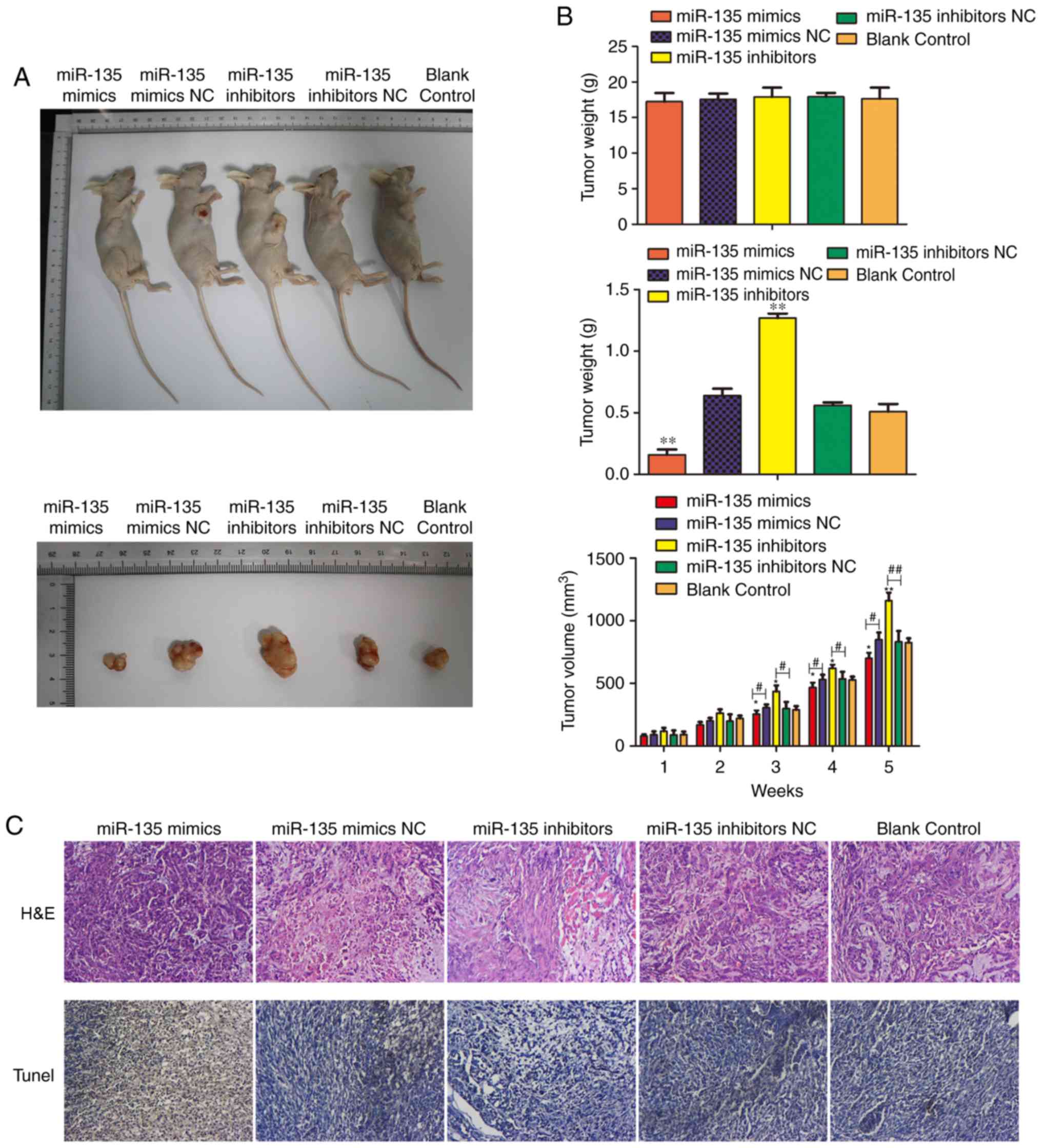
To further investigate the role of miR-135 in
tumorigenicity of BC cells in vivo, cells stably transfected
with miR-135 mimic, miR-135 inhibitors or controls were injected
subcutaneously into nude mice. As presented in Fig. 4A, the tumors formed by the cells
which were transfected with miR-135 inhibitors were larger and
heavier compared with the control tumors. Consistently, miR-135
over-expression significantly inhibited tumor growth of cells of
xenografted mice compared with the control (P<0.01). The tumor
sizes were measured every week. Data demonstrates that the tumors
in the group injected with cells stably overexpressing miR-135 grew
at a slower rate and exhibited smaller volumes compared with the
controls. Consistent with the tumor volume, the average tumor
weight was also significantly reduced (P<0.01; Fig. 4B). Furthermore, the subcutaneous
tumor tissues were examined by histochemical staining and the
effect of miR-135 on the cell apoptosis was assessed by TUNEL assay
(Fig. 4C). The results indicated
that the degree of apoptosis was notably decreased in cells
transfected with miR-135 inhibitor. In addition, overexpression of
miR-135 markedly increased the number of apoptotic cells when
compared with the control group. Based on these observations, it
was demonstrated that miR-135 overexpression inhibited tumor growth
in vivo.
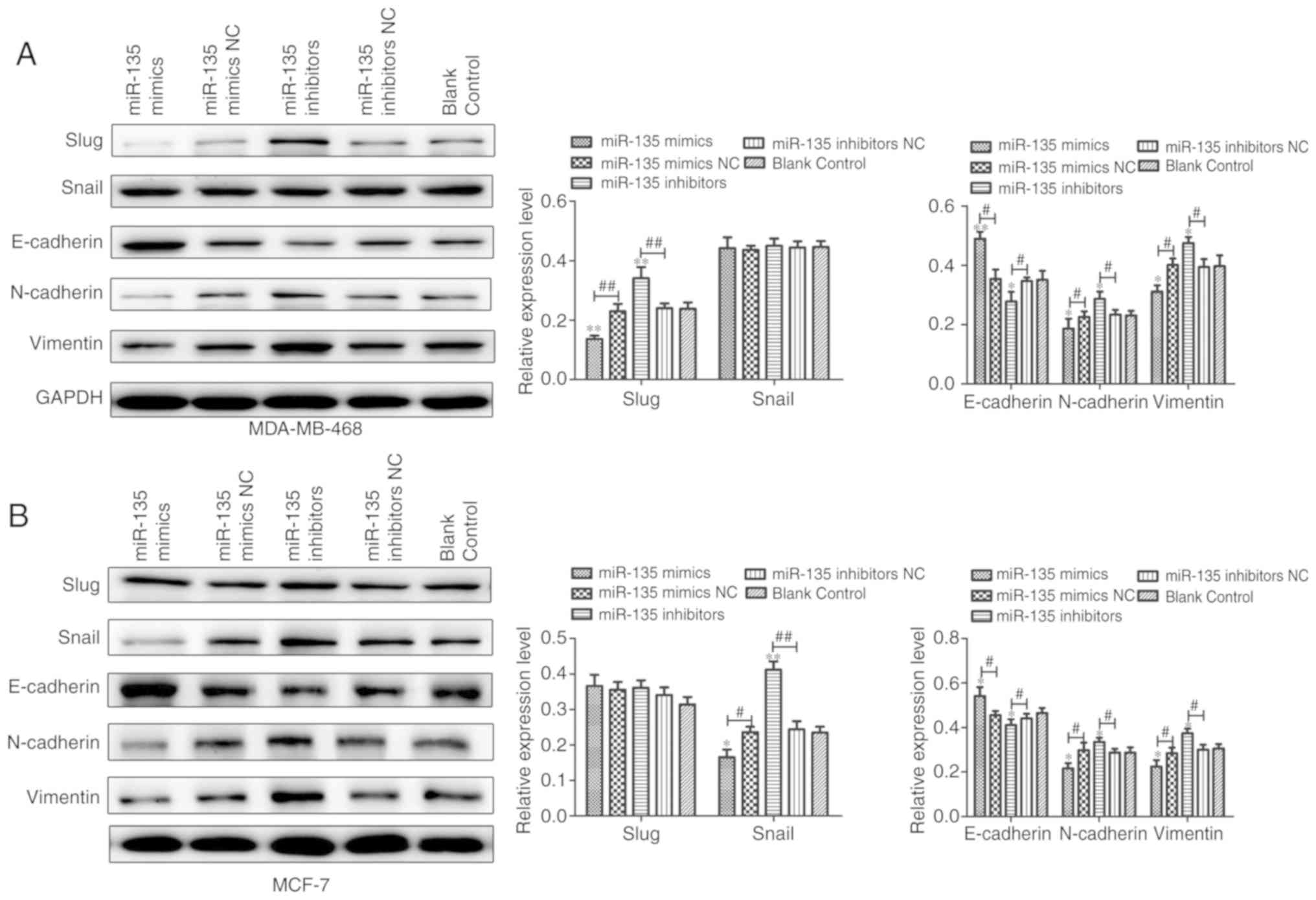
miR-135 suppresses EMT and is associated
with Wnt/β-catenin signaling activation in BC
To identify whether miR-135 can affect cell EMT, the
expression of EMT markers in MDA-MB-468 and MCF-7 cells was
measured by western blot analysis. It was demonstrated that miR-135
mimic transfection enhanced the expression of E-cadherin and
displayed the lower expression of mesenchymal markers including
Snail, Slug, neural (N)-cadherin and Vimentin at mRNA and protein
levels (Fig. 5) in cells, while
the results of cells transfected with miR-135 inhibitors
demonstrated the opposite activities. The results of the present
study revealed that miR-135 inhibited cells metastasis may be
through regulation of EMT in BC cells.
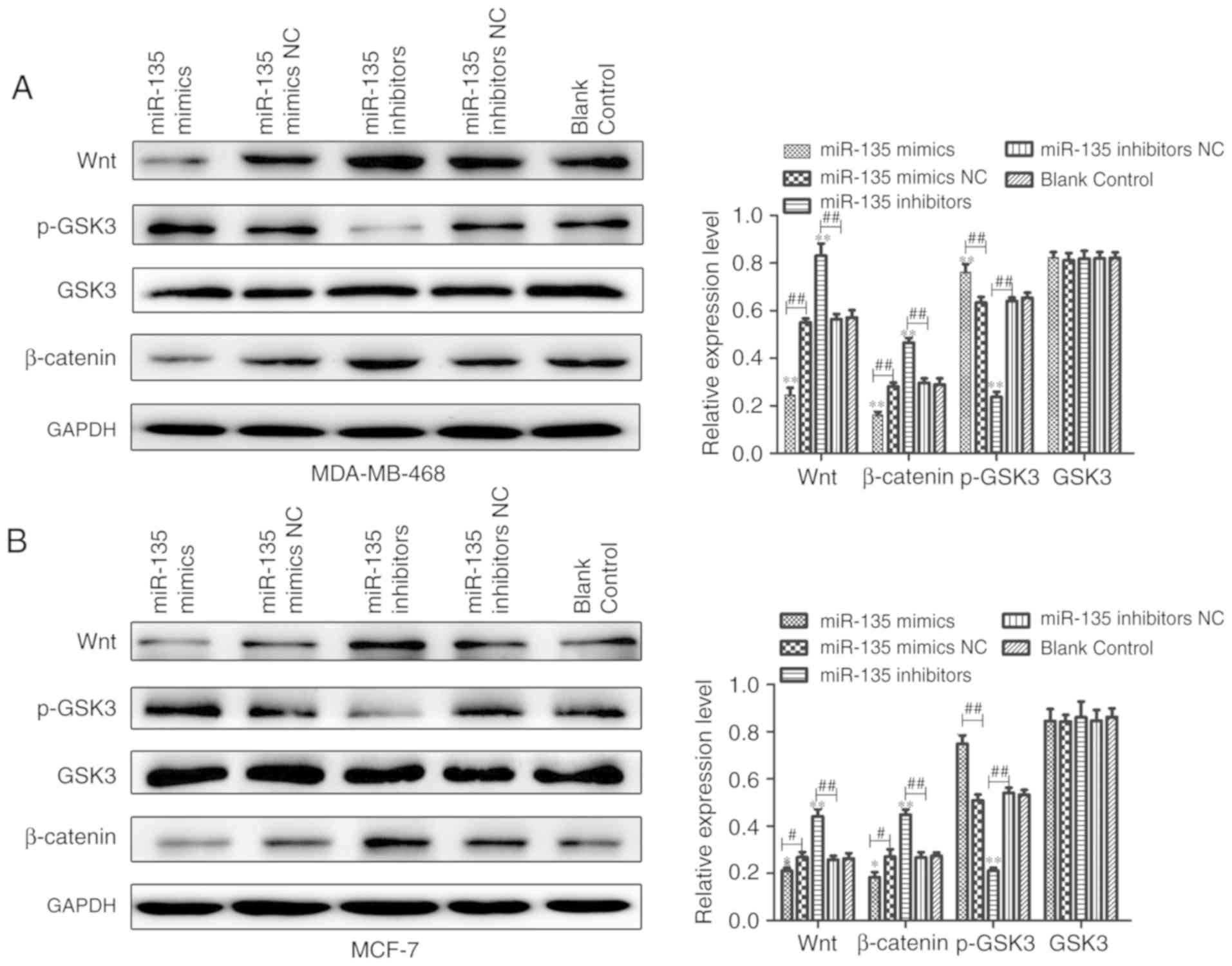
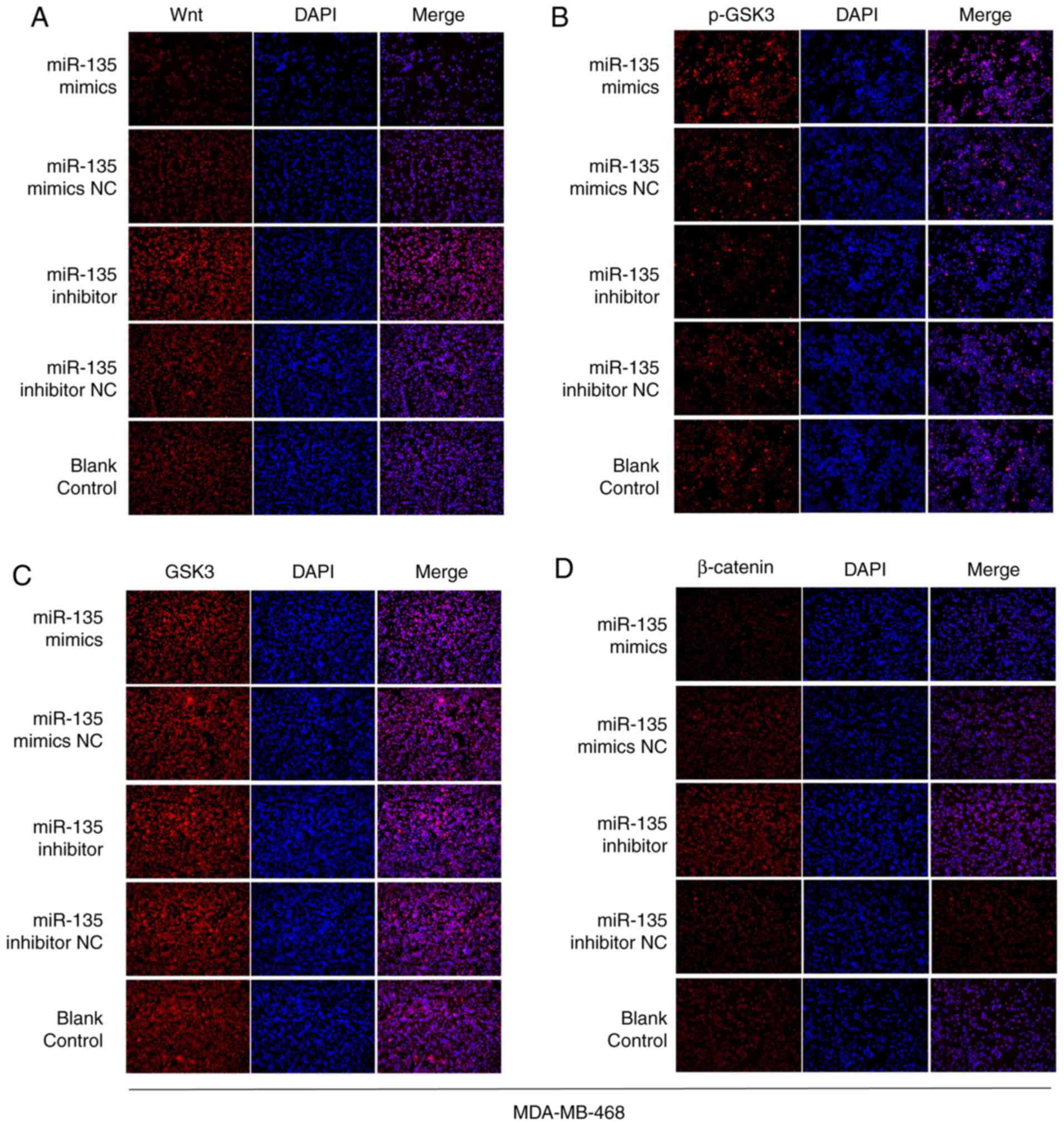
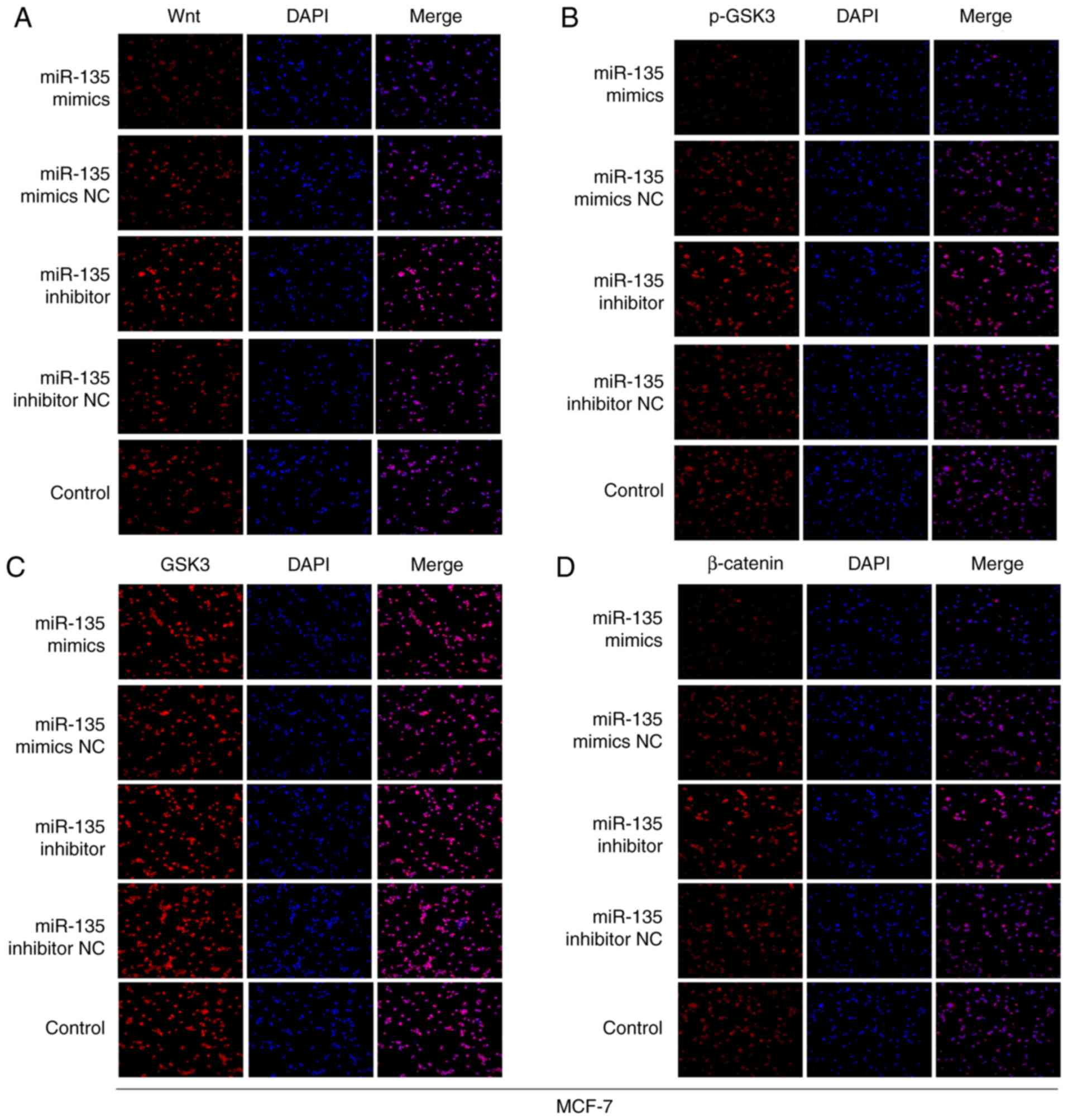
To further elucidate whether miR-135 levels were
associated with activation of Wnt/β-catenin signaling to
regulate-miR-135 inhibited cell EMT, western blotting and
immunofluorescence staining assays were performed to detect the
proteins levels that were associated with the Wnt/β-catenin
signaling pathway. As presented in Fig. 6, there was a significant positive
association between miR-135 and p-GSK3 expression, but a
significant inverse association between miR-135 and Wnt and
β-catenin expression. Furthermore, immunofluorescence staining
observed similar results (Figs. 7
and 8). The levels of Wnt and
β-catenin were remarkably upregulated in miR-135 inhibitors group
compared with the control group. In contrast, overexpression of
miR-135 increased the levels of phosphorylated (p)-glycogen
synthase kinase (GSK)3. Therefore, the aforementioned findings
indicate that downregulation of miR-135 participates in the
regulation of cell biological functions, at least in part through
activating Wnt/β-catenin signaling in BC.
Discussion
BC is the most frequent cancer of women due to a
complicated etiology involving environmental and genetic factors.
miRNAs, as the class of endogenous non-coding small RNA and can
modulate a wide variety of biological processes including tumor
cell proliferation, differentiation, migration, invasion, apoptosis
and metastasis (32-35). Numerous miRNAs have been reported
to be abnormally expressed in BC cells and tissues (36). However, the role of miR-135 in BC
remains elusive.
The present study identified the decreased
expression of miR-135 in MDA-MB-468, MDA-MB-231 and MCF-7 compared
with breast epithelial cell line MCF-10A. Therefore it was inferred
that miR-135 may act a tumor suppressor involved in the progression
and development in BC. Xu et al reported (21) that miR-135 inhibits prostate
cancer cell growth and migration by targeting EGFR. Wu et al
(37) demonstrated that miR-135a
targets JAK2 and inhibits gastric cancer cell proliferation. The
hypothesis was further confirmed by in vitro and in
vivo analysis in MDA-MB-468 and MCF-7 cells with MTT, colony
formation, wound healing, transwell and xenograft tumor growth
assays. The results demonstrated that overexpression of miR-135
inhibited the proliferation, migration, invasion and
tumorigenicity.
A number of studies have demonstrated that tumor
invasion and metastasis are a series of complex processes and MMPs
are associated with tumor invasion and metastasis (38-40). In the present study, the data
demonstrated that MMP-2 and MMP-9 expression was promoted following
transfection with miR-135 inhibitors, while miR-135 mimics could
suppress the expression of MMP-2 and MMP-9. The role of EMT has
been paid more attention in tumor metastasis as EMT is a complex
process in which epithelial cells lose their epithelial morphology
and acquire a mesenchymal phenotype (41,42). Therefore, it was demonstrated that
miR-135 could inhibit the EMT process by inducing the levels of
E-cadherin and reducing Slug, Snail, N-cadherin and Vimentin
expression. The Wnt/β-catenin signaling pathway has a significant
impact on the maintenance of stem cell properties and cancer
metastasis (43,44). Additionally, the nuclear
accumulation of β-catenin is a crucial step in the activation of
the Wnt signaling pathway (45).
In the present study, evidence was provided that miR-135 could
inhibit the activation of the Wnt/β-catenin pathway.
From these data, it was demonstrated that miR-135
could inhibit cell proliferation, migration and invasion, at least
in part through Wnt/β-catenin signaling pathway in BC. It was
concluded that miR-135 was a potential tumor suppressor and could
serve a critical role in the progression of BC. In conclusion, the
results of the present study could represent a potential
therapeutic target for the diagnosis and treatment of breast
cancer. Furthermore, the present study provides novel insights into
the pathogenesis and therapeutics of breast cancer. The current
study is a preliminary study on the anti-tumor effect of miR-135 in
BC. Previous studies have reported that signal transducer and
activator of transcription (STAT)3 is connected with the nuclear
factor (NF)-κB signaling pathway (46-49). The association between miR-135 and
the STAT3/NF-κB signaling pathway on BC based on current findings
will be investigated in future studies.
Funding
No funding was received.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors’ contributions
DJ and HC conceived and designed the study. BZ and
YX performed the experiments. DJ wrote the manuscript. All authors
have read and approved the manuscript and agree to be accountable
for all aspects of the research in ensuring that the accuracy and
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
Animal care and use were carried out according to
the ethical guidelines by Nanjing Medical University Animal Care
and Use Committee and approved by the Nanjing Medial University
Experimental Animal Ethics Committee and the permit number is
SYXK(su)2016-0016.
Patient consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ismail NI, Kaur G, Hashim H and Hassan MS:
S100A4 overexpression proves to be independent marker for breast
cancer progression. Cancer Cell Int. 8:122008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fedewa SA, Sauer AG, Siegel RL and Jemal
A: Prevalence of major risk factors and use of screening tests for
cancer in the United States. Cancer Epidemiol Biomarkers Prev.
26:1192–1208. 2017. View Article : Google Scholar
|
|
4
|
Zou DH, He XM, Chen B, Xu XH, Zhang XP, Ni
JF and Mao WM: Expression and association of reproductive hormones
and receptors in postmenopausal patients with breast cancer. Eur J
Gynaecol Oncol. 38:727–732. 2017.
|
|
5
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rivera E and Gomez H: Chemotherapy
resistance in metastatic breast cancer: The evolving role of
ixabepilone. Breast Cancer Res. 12(Suppl 2): S22010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lorusso G and Rüegg C: The tumor
microenvironment and its contribution to tumor evolution toward
metastasis. Histochem Cell Biol. 130:1091–1103. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gromak N: Intronic microRNAs: A crossroad
in gene regulation. Biochem Soc Trans. 40:759–761. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fernandez S, Risolino M, Mandia N, Talotta
F, Soini Y, Incoronato M, Condorelli G, Banfi S and Verde P:
miR-340 inhibits tumor cell proliferation and induces apoptosis by
targeting multiple negative regulators of p27 in non-small cell
lung cancer. Oncogene. 34:3240–3250. 2015. View Article : Google Scholar
|
|
10
|
Sandhu R, Rein J, D’Arcy M, Herschkowitz
JI, Hoadley KA and Troester MA: Overexpression of miR-146a in
basal-like breast cancer cells confers enhanced tumorigenic
potential in association with altered p53 status. Carcinogenesis.
35:2567–2575. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Waning DL, Mohammad KS and Guise TA:
Cancer-associated osteoclast differentiation takes a good look in
the miR(NA)ror. Cancer Cell. 24:407–409. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Valeri N, Braconi C, Gasparini P, Murgia
C, Lampis A, Paulus-Hock V, Hart JR, Ueno L, Grivennikov SI, Lovat
F, et al: MicroRNA-135b promotes cancer progression by acting as a
downstream effector of oncogenic pathways in colon cancer. Cancer
Cell. 25:469–483. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ma Y, Zhang P, Wang F, Zhang H, Yang Y,
Shi C, Xia Y, Peng J, Liu W, Yang Z and Qin H: Elevated oncofoetal
miR-17-5p expression regulates colorectal cancer progression by
repressing its target gene P130. Nat Commun. 3:12912012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang B, Wang H and Yang Z: MiR-122
inhibits cell proliferation and tumorigenesis of breast cancer by
targeting IGF1R. PLoS One. 7:e470532012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li L, Yuan L, Luo J, Gao J, Guo J and Xie
X: MiR-34a inhibits proliferation and migration of breast cancer
through down-regulation of Bcl-2 and SIRT1. Clin Exp Med.
13:109–117. 2013. View Article : Google Scholar
|
|
16
|
Sun Y, Wang M, Lin G, Sun S, Li X, Qi J
and Li J: Serum microRNA-155 as a potential biomarker to track
disease in breast cancer. PLoS One. 7:e470032012. View Article : Google Scholar :
|
|
17
|
Nagel R, le Sage C, Diosdado B, van der
Waal M, Oude Vrielink JA, Bolijn A, Meijer GA and Agami R:
Regulation of the adenomatous polyposis coli gene by the miR-135
family in colorectal cancer. Cancer Res. 68:5795–5802. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Holleman A, Chung I, Olsen RR, Kwak B,
Mizokami A, Saijo N, Parissenti A, Duan Z, Voest EE and Zetter BR:
miR-135a contributes to paclitaxel resistance in tumor cells both
in vitro and in vivo. Oncogene. 30:4386–4398. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yamada Y, Hidaka H, Seki N, Yoshino H,
Yamasaki T, Itesako T, Nakagawa M and Enokida H: Tumor-suppressive
microRNA-135a inhibits cancer cell proliferation by targeting the
c-MYC oncogene in renal cell carcinoma. Cancer Sci. 104:304–312.
2013. View Article : Google Scholar
|
|
20
|
Chen Y, Zhang J, Wang H, Zhao J, Xu C, Du
Y, Luo X, Zheng F, Liu R, Zhang H and Ma D: miRNA-135a promotes
breast cancer cell migration and invasion by targeting HOXA10. BMC
Cancer. 12:1112012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu B, Tao T, Wang Y, Fang F, Huang Y, Chen
S, Zhu W and Chen M: hsa-miR-135a-1 inhibits prostate cancer cell
growth and migration by targeting EGFR. Tumour Biol.
37:14141–14151. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–190. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu WH, Liu ZB, Yang C, Qin W and Shao ZM:
Expression of dickkopf-1 and beta-catenin related to the prognosis
of breast cancer patients with triple negative phenotype. PLoS One.
7:e376242012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lópezknowles E, Zardawi SJ, McNeil CM,
Millar EK, Crea P, Musgrove EA, Sutherland RL and O’Toole SA:
Cytoplasmic localization of beta-catenin is a marker of poor
outcome in breast cancer patients. Cancer Epidemiol Biomarkers
Prev. 19:301–309. 2010. View Article : Google Scholar
|
|
25
|
Zardawi SJ, O’Toole SA, Sutherland RL and
Musgrove EA: Dysregulation of Hedgehog, Wnt and Notch signalling
pathways in breast cancer. Histol Histopathol. 24:385–398.
2009.PubMed/NCBI
|
|
26
|
He X: Understanding Wnt/b-catenin
signaling in development and cancer. J Nanjing Med Univ.
22:802008.
|
|
27
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen Q, Cao HZ and Zheng PS: LGR5 promotes
the proliferation and tumor formation of cervical cancer cells
through the Wnt/β-catenin signaling pathway. Oncotarget.
5:9092–9105. 2014.PubMed/NCBI
|
|
29
|
Hua HW, Jiang F, Huang Q, Liao Z and Ding
G: MicroRNA-153 promotes Wnt/β-catenin activation in hepatocellular
carcinoma through suppression of WWOX. Oncotarget. 6:3840–3847.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou DS, Wang HB, Zhou ZG, Zhang YJ, Zhong
Q, Xu L, Huang YH, Yeung SC, Chen MS and Zeng MS: TACC3 promotes
stemness and is a potential therapeutic target in hepatocellular
carcinoma. Oncotarget. 6:24163–24177. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
32
|
Zhang B, Wang Q and Pan X: MicroRNAs and
their regulatory roles in animals and plants. J Cell Physiol.
210:279–289. 2007. View Article : Google Scholar
|
|
33
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in cancer. Ann Rev Med. 60:167–179. 2015. View Article : Google Scholar
|
|
34
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-C ordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Miranda KC, Huynh T, Tay Y, Ang YS, Tam
WL, Thomson AM, Lim B and Rigoutsos I: A pattern-based method for
the identification of MicroRNA binding sites and their
corresponding heteroduplexes. Cell. 126:1203–1217. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shi W, Gerster K, Alajez NM, Tsang J,
Waldron L, Pintilie M, Hui AB, Sykes J, P’ng C, Miller N, et al:
MicroRNA-301 mediates proliferation and invasion in human breast
cancer. Cancer Res. 71:2926–2937. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu H, Huang M, Cao P, Wang T, Shu Y and
Liu P: MiR-135a targets JAK2 and inhibits gastric cancer cell
proliferation. Cancer Biol Ther. 13:281–288. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bar-Or A, Nuttall RK, Duddy M, Alter A,
Kim HJ, Ifergan I, Pennington CJ, Bourgoin P, Edwards DR and Yong
VW: Analyses of all matrix metalloproteinase members in leukocytes
emphasize monocytes as major inflammatory mediators in multiple
sclerosis. Brain. 126:2738–2749. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Westermarck J and Kähäri VM: Regulation of
matrix metalloproteinase expression in tumor invasion. FASEB J.
13:781–792. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kraiem Z and Korem S: Matrix
metalloproteinases and the thyroid. Thyroid. 10:1061–1069. 2000.
View Article : Google Scholar
|
|
41
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:14202009.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kalluri R and Neilson EG:
Epithelial-mesenchymal transition and its implications for
fibrosis. J Clin Invest. 112:1776–1784. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Leung CO, Mak WN, Kai AK, Chan KS, Lee TK,
Ng IO and Lo RC: Sox9 confers stemness properties in hepatocellular
carcinoma through Frizzled-7 mediated Wnt/β-catenin signaling.
Oncotarget. 7:29371–29386. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yuan X, Sun X, Shi X, Wang H, Wu G, Jiang
C, Yu D, Zhang W, Xue B and Ding Y: USP39 promotes colorectal
cancer growth and metastasis through the Wnt/β-catenin pathway.
Oncol Rep. 37:2398–2404. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mao Y, Xu J, Li Z, Zhang N, Yin H and Liu
Z: The role of nuclear β-catenin accumulation in the Twist2-induced
ovarian cancer EMT. PLoS One. 8:e782002013. View Article : Google Scholar
|
|
46
|
Yamamoto Y and Gaynor RB: Therapeutic
potential of inhibition of the NF-κB pathway in the treatment of
inflammation and cancer. J Clin Invest. 107:135–142. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Takaesu G, Surabhi RM, Park KJ,
Ninomiya-Tsuji J, Matsumoto K and Gaynor RB: TAK1 is critical for
IkappaB kinase-mediated activation of the NF-kappaB pathway. J Mol
Biol. 326:105–115. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sasaki CY, Barberi TJ, Ghosh P and Longo
DL: Phosphorylation of RelA/p65 on serine 536 defines an
I{kappa}B{alpha}-independent NF-{kappa}B pathway. J Biol Chem.
280:34538–34547. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wajant H and Scheurich P: TNFR1-induced
activation of the classical NF-κB pathway. FEBS J. 278:862–876.
2015. View Article : Google Scholar
|