Introduction
Hydrogen sulfide (H2S), an endogenous
gaseous signaling molecule, performs important regulatory
functions, such as vascular toning, leukocyte adhesion and smooth
muscle cell proliferation, in the cardiovascular system. Recently,
Li et al demonstrated that H2S reduces
atherosclerosis by regulating lipid metabolism (1). Exogenous H2S can mitigate
fatty liver and reduce triglyceride and total cholesterol levels in
obese mice (2). Cytathiohine
β-synthase (a key enzyme for the biosynthesis of H2S)
knockout mice have been reported to exhibit higher low-density
lipoprotein cholesterol (LDL-C) and lower plasma levels of
high-density lipoprotein cholesterol than normal mice (3). Additionally, patients with
atherosclerosis exhibit reduced serum H2S concentrations
(4). However, the precise
mechanisms through which H2S regulates lipid metabolism
remain unclear.
It is well established that proprotein convertase
subtilisin/kexin type 9 (PCSK9) plays an important role in lipid
metabolism and atherosclerosis (5). PCSK9 alters LDL-C concentrations by
promoting hepatic low-density lipoprotein receptor (LDLR)
degradation and decreasing the hepatic clearance of plasma LDL-C
levels (6). Numerous factors
regulate PCSK9 expression. For example, statin treatment leads to
an increased transcription of PCSK9 (7), in contrast to berberine treatment,
which decreases PCSK9 expression (8). However, to date, whether
H2S is involved in the regulation of hepatic PCSK9
expression remains unknown.
In the present study, we thus aimed to examine the
effects of H2S on the expression of PCSK9 in HepG2 cells
and to elucidate the mechanisms through which H2S
regulates lipid metabolism.
Materials and methods
Reagents
Sodium hydrosulfide (NaHS) and Oil Red O were
purchased from Sigma-Aldrich (St. Louis, MO, USA). Specific
monoclonal anti-PCSK9 (cat. no. 55206), anti-LDLR (cat. no. 66414),
anti-SREBP-2 (cat. no. 557037) and anti-SREBP-1-c (cat. no. 66875)
antibodies were purchased from Proteintech (Rosemont, IL, USA).
Specific monoclonal anti-protein kinase B (Akt) antibody (cat. no.
BM4400) was purchased from Wuhan Boshide Bioengineering Co. (Wuhan,
China). Specific monoclonal anti-phosphorylated Akt antibody (cat.
no. YM3621) was purchased from ImmunoWay Biotechnology (Plano, TX,
USA). Anti-β-actin antibody (cat. no. LCA01) was purchased from
Aijia Biological Technology Co., Ltd. (Changsha, China).
Cy3-conjugated goat anti-rabbit IgG secondary antibody (cat. no.
E031620) was purchased from EarthOx Life Sciences (Millbrae, CA,
USA). Dulbecco's modified Eagle's medium (DMEM) was purchased from
HyClone (Logan, UT, USA). Fetal bovine serum (FBS) was purchased
from Hangzhou Sijiqing Biocompany (Hangzhou, China). The Hoechst
Staining kit was purchased from Beyotime Institute of Biotechnology
(Jiangsu, China).
1,1′-Dioctadecyl-3,3,3′,3′-tetramethyl-indocarbocyanine
perchlorate-labeled LDL (DiI-LDL) was purchased from Haoyuan
Biological Technology Co., Ltd. (Guangzhou, China). siRNAs were
synthesized by Ruibo Biotechnology Co., Ltd. (Guangzhou,
China).
Cells and cell culture
The HepG2 human liver cancer cell line was obtained
from the Cell Bank of the Chinese Academy of Sciences (Shanghai,
China) and maintained in tissue culture bottles with DMEM
supplemented with 10% heat-inactivated FBS at 37°C under an
atmosphere of 5% CO2 and 95% air. The cell culture
medium was replaced with fresh medium 3 times a week.
Oil Red O staining protocol
Oil Red O staining was used to quantify
intracytoplasmic lipid droplets. The HepG2 cells were seeded in a
6-well plate covered with coverslips. After the cells had become
adherent, they were incubated for 12 h at 37°C in serum-free
medium, treated with various concentrations (0, 50, 100 and 200
µmol/l) of NaHS (a H2S donor) for 0.5 h, and
subsequently incubated with 30% FBS for 24 h. The HepG2 cells were
first fixed with 4% paraformaldehyde for 30 min at room
temperature, and washed 3 times with distilled water. Subsequently,
the cells were stained with Oil Red O at room temperature for 15
min. The cells were then washed 3 times with distilled water, and
nuclear counterstaining was performed with hematoxylin and eosin
(Beyotime Institute of Biotechnology, Jiangsu, China) staining for
30 sec at room temperature. Lipid droplets were observed by light
microscopy (Olympus, Tokyo, Japan) and then imaged using Image
Pro-Plus 6.0 software (Media Cybernetics, Rockville, MD, USA).
DiI-LDL uptake assay
The HepG2 cells were seeded in 24-well plates.
Following treatment with various concentrations of NaHS (0, 50, 100
and 200 µmol/l) for 24 h, the cells were rinsed twice with
phosphate-buffered saline (PBS). Subsequently, the cells were
incubated with 25 µg/ml DiI-LDL for 4 h at 37°C, followed by
fixation in 3% paraformaldehyde for 20 min. The nuclei were
counterstained with Hoechst 33258 (250 nM; Beyotime Institute of
Biotechnology) for 5 min at room temperature. Immunofluorescence
was visualized using a Nikon TE2000-U fluorescence microscope
(Nikon, Tokyo, Japan).
Immunofluorescence assay
The HepG2 cells were plated in 24-well plates.
Following treatment with various concentrations of NaHS (0, 50, 100
and 200 µmol/l) for 24 h, the cells were rinsed 3 times with
PBS, fixed with 4% paraformaldehyde for 30 min, permeabilized with
0.5% Triton X-100 in PBS for 30 min, and incubated with anti-LDLR
antibody (1:1,000) overnight at 4°C. The cells were rinsed 3 times
for 5 min with PBS, followed by incubation with the appropriate
fluorophore-conjugated secondary antibody (SA012; 1:1,000; Aijia
Biological Technology Co., Ltd.) for 2 h. The nuclei were
counterstained with Hoechst 33258 (250 nM; Beyotime Institute of
Biotechnology). Immunofluorescence was visualized using a Nikon
TE2000-U fluorescence microscope (Nikon).
Western blot analysis
The cells were washed twice with PBS and incubated
on ice with cell lysis buffer and phenylmethanesulfonyl fluoride
for 30 min, followed by cell lysate collection and centrifugation
at 10,000 x g for 10 min at 4°C. The BCA protein assay kit was used
for the quantification of total protein in cell lysates (Beyotime
Institute of Biotechnology) according to the manufacturer's
instructions. Proteins were separated by 12% SDS-PAGE and
electrotransferred onto a polyvinylidene fluoride membrane. The
membrane was incubated in a blocking buffer (0.1% Tween-20, 150 mM
NaCl, 20 mM Tris base, pH 7.6 and 5% non-fat milk) for 2 h at 37°C
and subsequently with the appropriate primary antibodies (PCSK9,
1:400; LDLR, 1:1,000; SREBP-2, 1:5,000; SREBP-1c, 1:2,000; Akt,
1:400; phosphorylated Akt, 1:2,000; and β-actin, 1:1,000) in
blocking buffer at 4°C overnight. The membranes were then incubated
with anti-rabbit or anti-mouse horseradish peroxidase-conjugated
secondary antibodies (SA009 and SA001; 1:1,000; Aijia Biological
Technology Co., Ltd.) for 2 h. Proteins were visualized by
chemiluminescence using AuraECL (Aijia Biological Technology Co.,
Ltd.) following the manufacturer's instructions. Image Pro-Plus 6.0
software (Media Cybernetics) was used to analyze relative protein
band density.
siRNA transfection
The PCSK9 mRNA sequence was retrieved from GenBank
(no. NM 199253). The target sequences of PCSK9 were as follows:
siPCSK9 sense strand, 5′-GGC AGA GAC UGA UCC ACU UdT dT-3′ and
siPCSK9 antisense strand, 5′-AAG UGG AUC AGU CUCU GCC TdT d-3′. The
sequences of the negative control siRNA were as follows: Sense
strand, 5′-UAU AGC UGU CUC GAG CAA GdT dT-3′ and antisense strand,
5′-CUU GCU CGA GAC AGC UAU ATd Td-3′. The HepG2 cells were seeded
in 6-well plates. siRNA transfection was performed using riboFECT
™CP (Ruibo Biotechnology Co., Ltd.) following the manufacturer's
instructions. Cells were harvested for the subsequent experiments
at 48 h following transfection.
Statistical analysis
Quantitative data are presented as the means ±
standard error of the mean, with the significance determined by a
Student's t-test or one-way analysis of variance with Tukey's
multiple comparisons post hoc test. The results were plotted using
GraphPad Prism 5.0 software. Differences between datasets were
considered statistically significant at P<0.05.
Results
H2S inhibits PCSK9 expression
in HepG2 cells
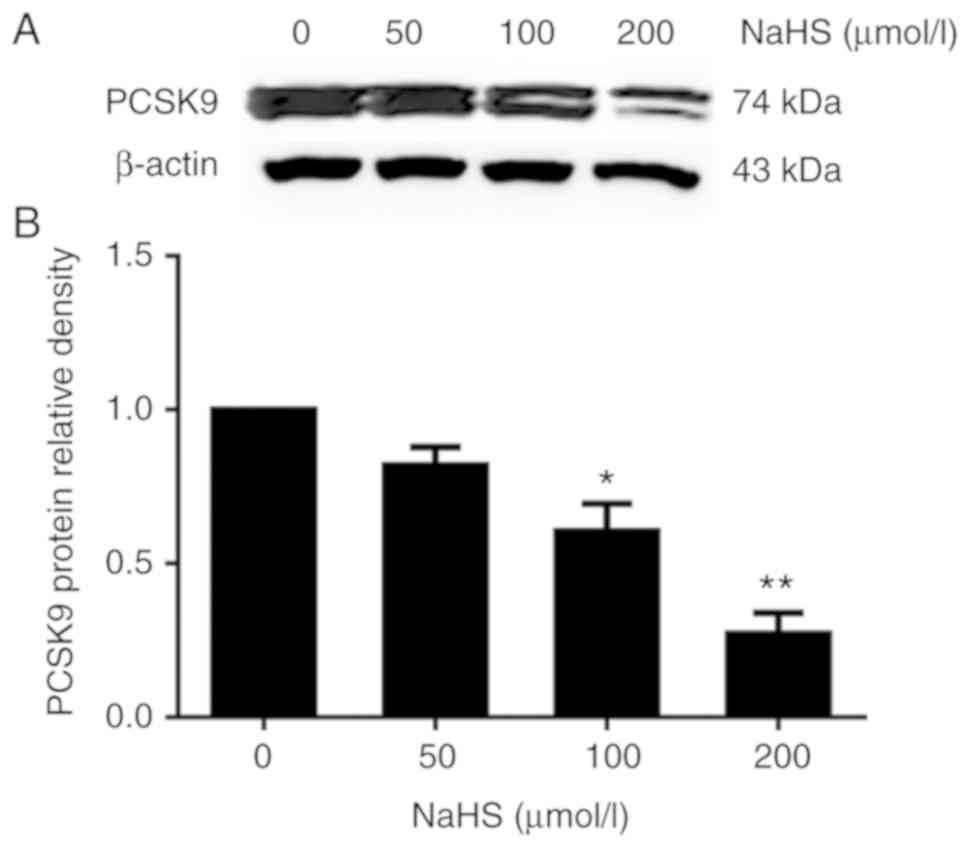
To elucidate the role of H2S in PCSK9
expression, we first incubated the HepG2 cells with various
concentrations of NaHS (0, 50, 100 and 200 µmol/l) for 24 h.
PCSK9 protein expression was detected by western blot analysis. As
shown in Fig. 1, H2S
inhibited PCSK9 expression in the HepG2 cells in a
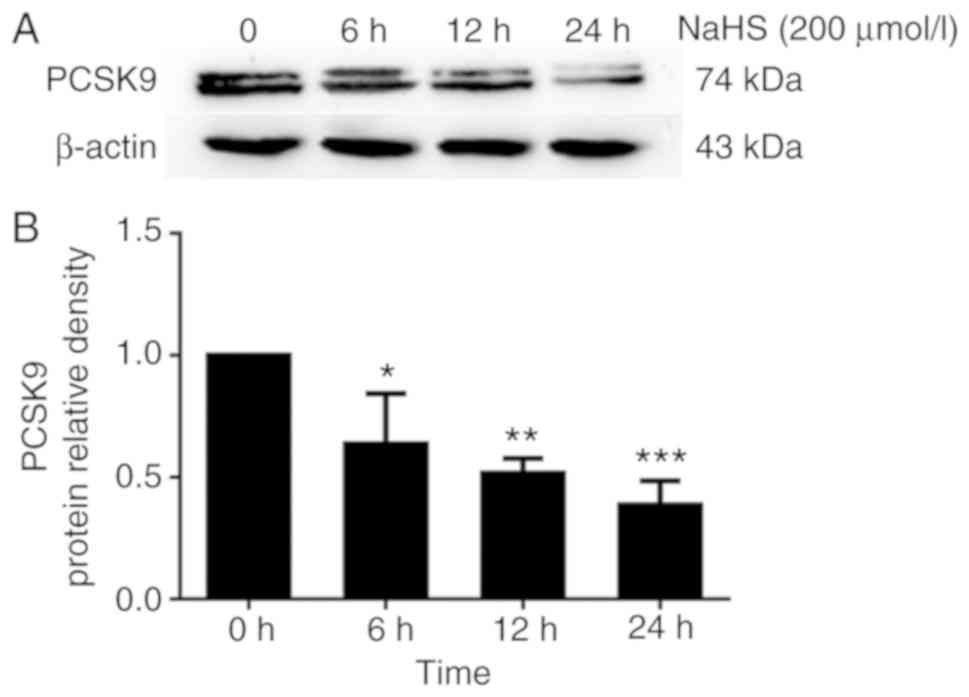
concentration-dependent manner. Subsequently, we incubated the
HepG2 cells with 200 µmol/l NaHS for the indicated periods
of time (0, 6, 12 and 24 h). As shown in Fig. 2, H2S inhibited PCSK9
expression in the HepG2 cells in a time-dependent manner.
H2S inhibits the expression of
PCSK9, upregulating the expression of LDLR and increase lipid
uptake in HepG2 cells
The above-mentioned results indicated that
H2S inhibited PCSK9 expression. Thus, the LDLR levels
may be increased in the HepG2 cells. To determine whether
H2S inhibits the expression of PCSK9 to influence the
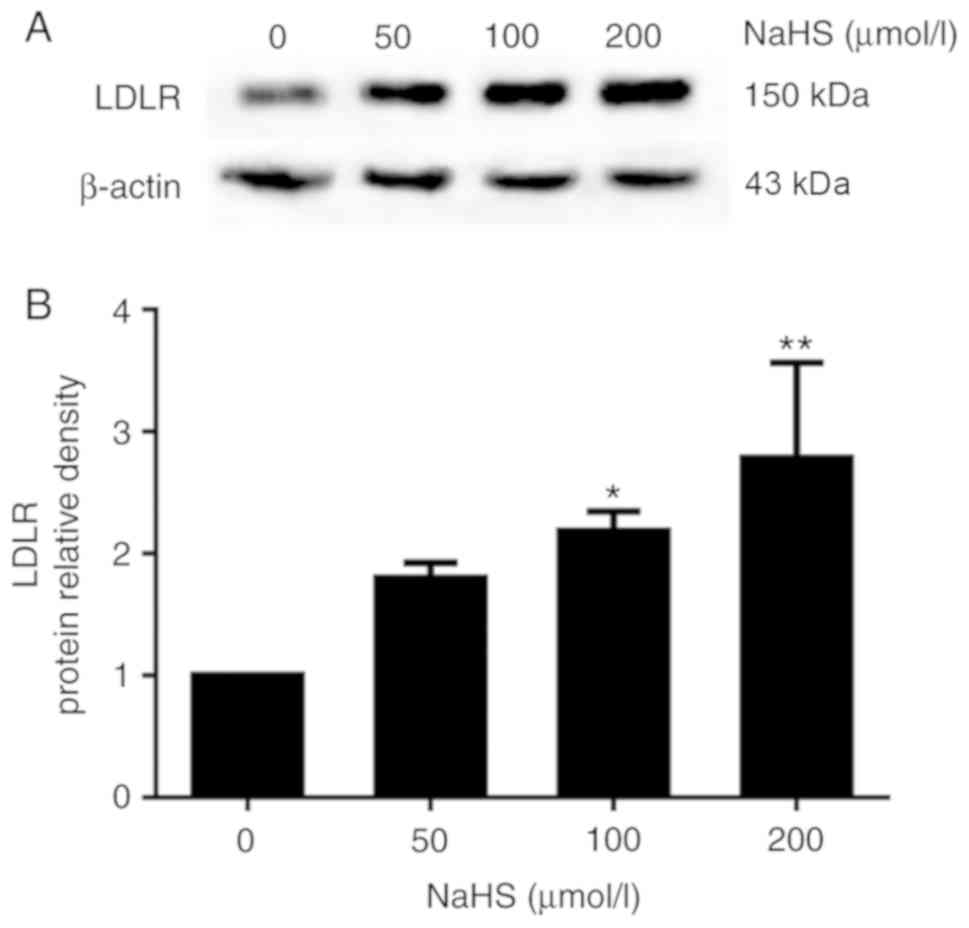
expression of LDLR and lipid uptake, we first examined the effects
of H2S on LDLR expression in the HepG2 cells. The HepG2
cells were incubated with various concentrations of NaHS (0, 50,
100 and 200 µmol/l) for 24 h. LDLR expression was detected
by western blot analysis and immunofluorescence assay. As shown in
Fig. 3, H2S
upregulated LDLR expression in the HepG2 cells in a
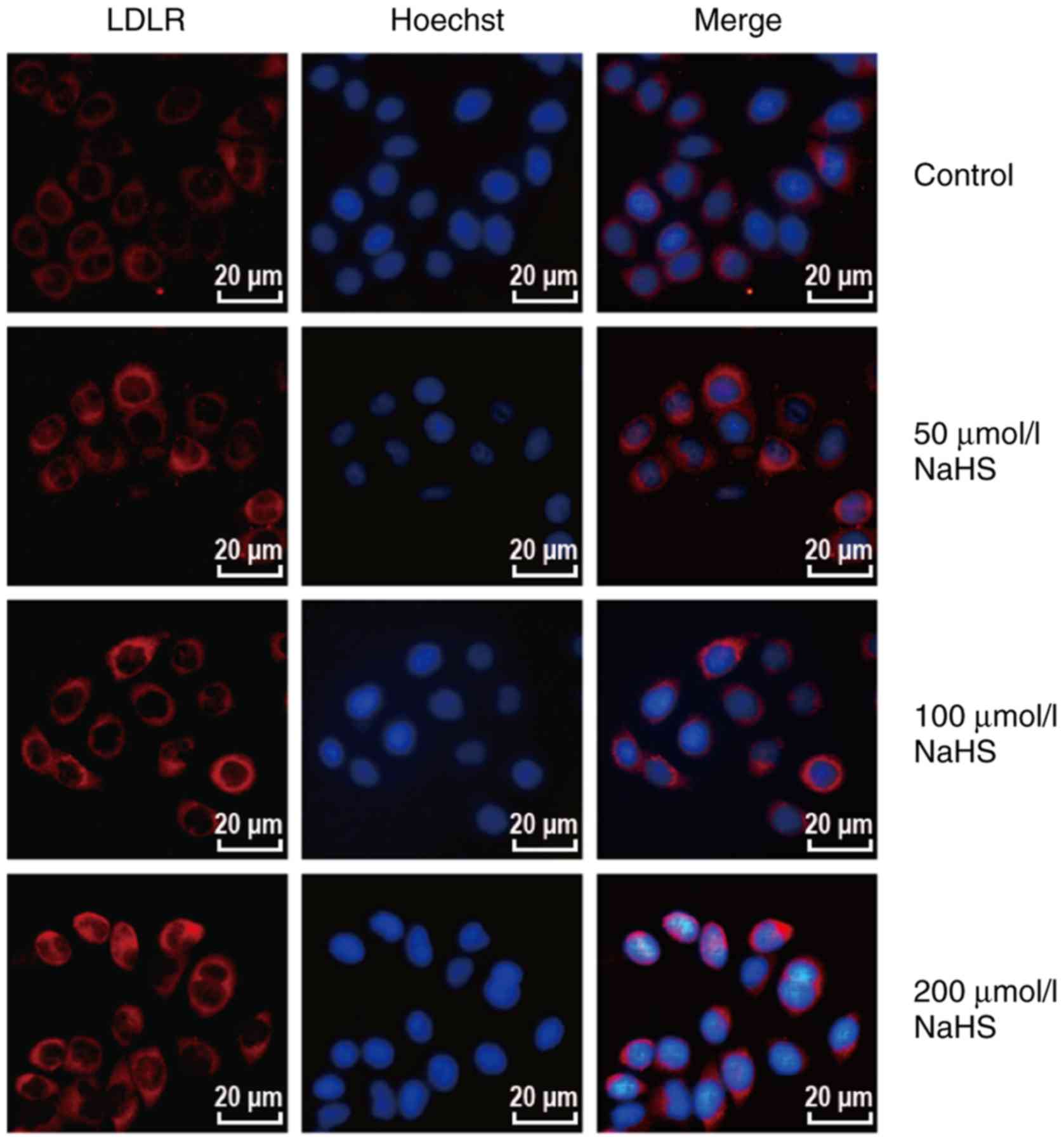
concentration-dependent manner. In addition, as shown in Fig. 4, red fluorescence intensity
increased with the increasing NaHS concentrations. This finding
indicates that H2S upregulates the LDLR protein
levels.
It is well known that PCSK9 can promote LDLR
degradation (9). Thus, to
determine whether H2S upregulates LDLR expression by
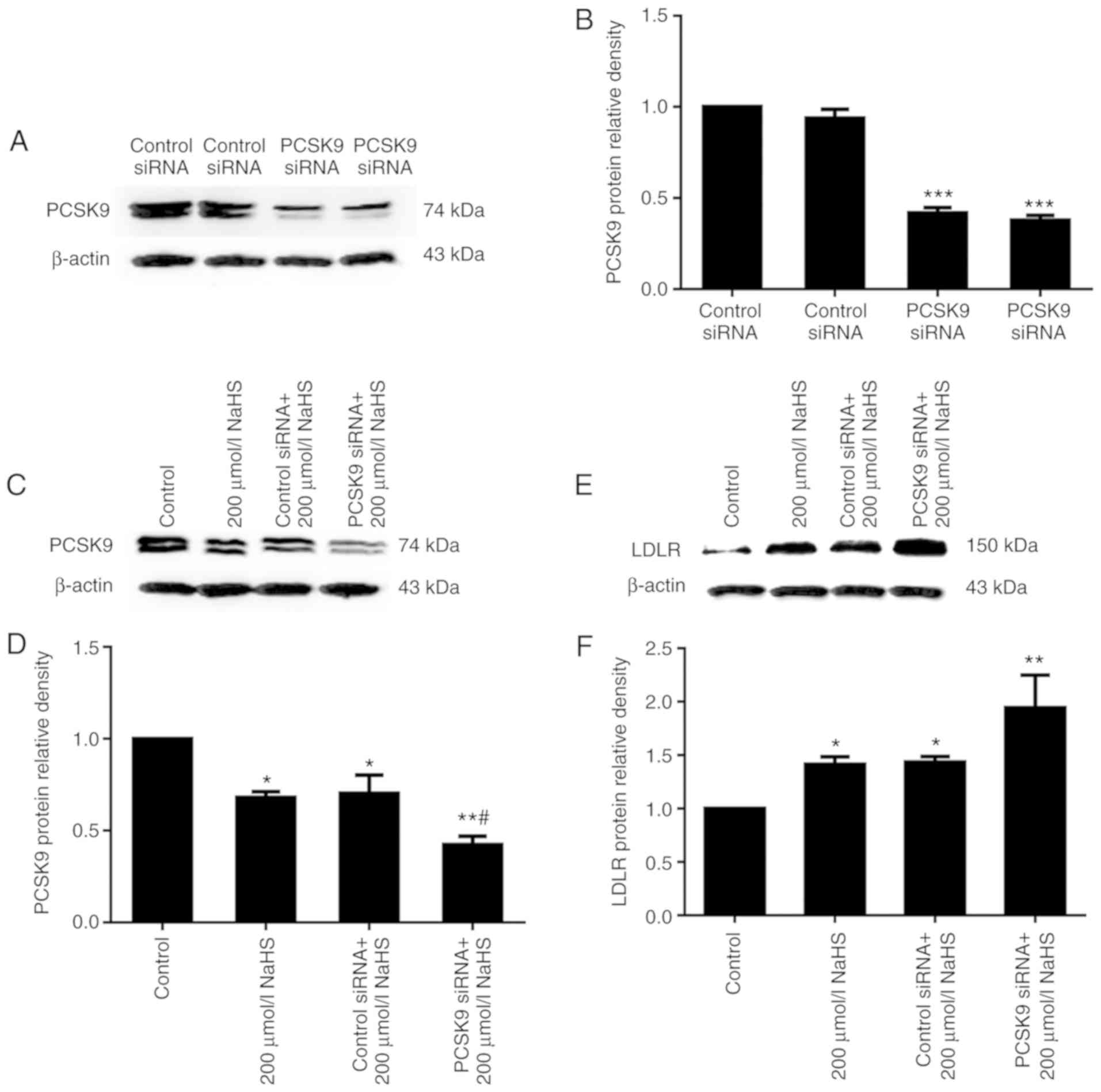
inhibiting PCSK9 expression, we first used RNA interference to
suppress PCSK9 expression, and LDLR expression was detected. The
results of western blot analysis revealed that PCSK9 inhibition was
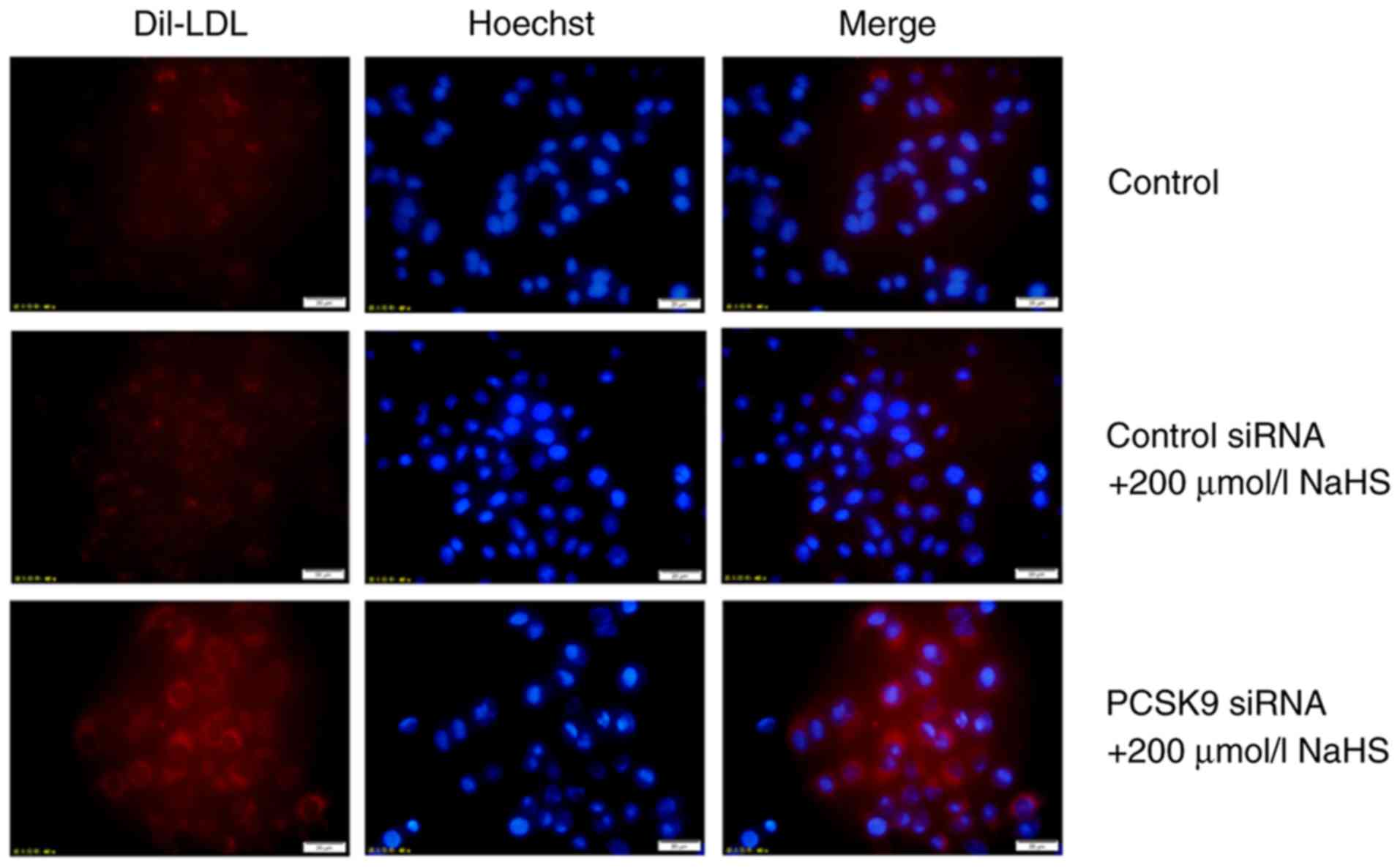
maintained following transfection for 48 h (Fig. 5A and B). Subsequently, DiI-LDL was
incubated with the HepG2 cells for 4 h, and DiI-LDL uptake by the
HepG2 cells was detected. The uptake of DiI-LDL at 37°C represented
the cell surface-bound DiI-LDL. As shown in Figs. 5 and 6, compared with the cells treated with
200 µmol/l NaHS, those that were transfected with PCSK9
siRNA exhibited an increased LDLR expression and increased lipid
uptake. This result indicated that H2S reduced the
effects of PCSK9 on LDLR degradation by inhibiting PCSK9
expression, thereby increasing lipid uptake by HepG2 cells.
H2S regulates the expression
of PCSK9 by affecting the PI3K/Akt-SREBP-2 signaling pathway in
HepG2 cells
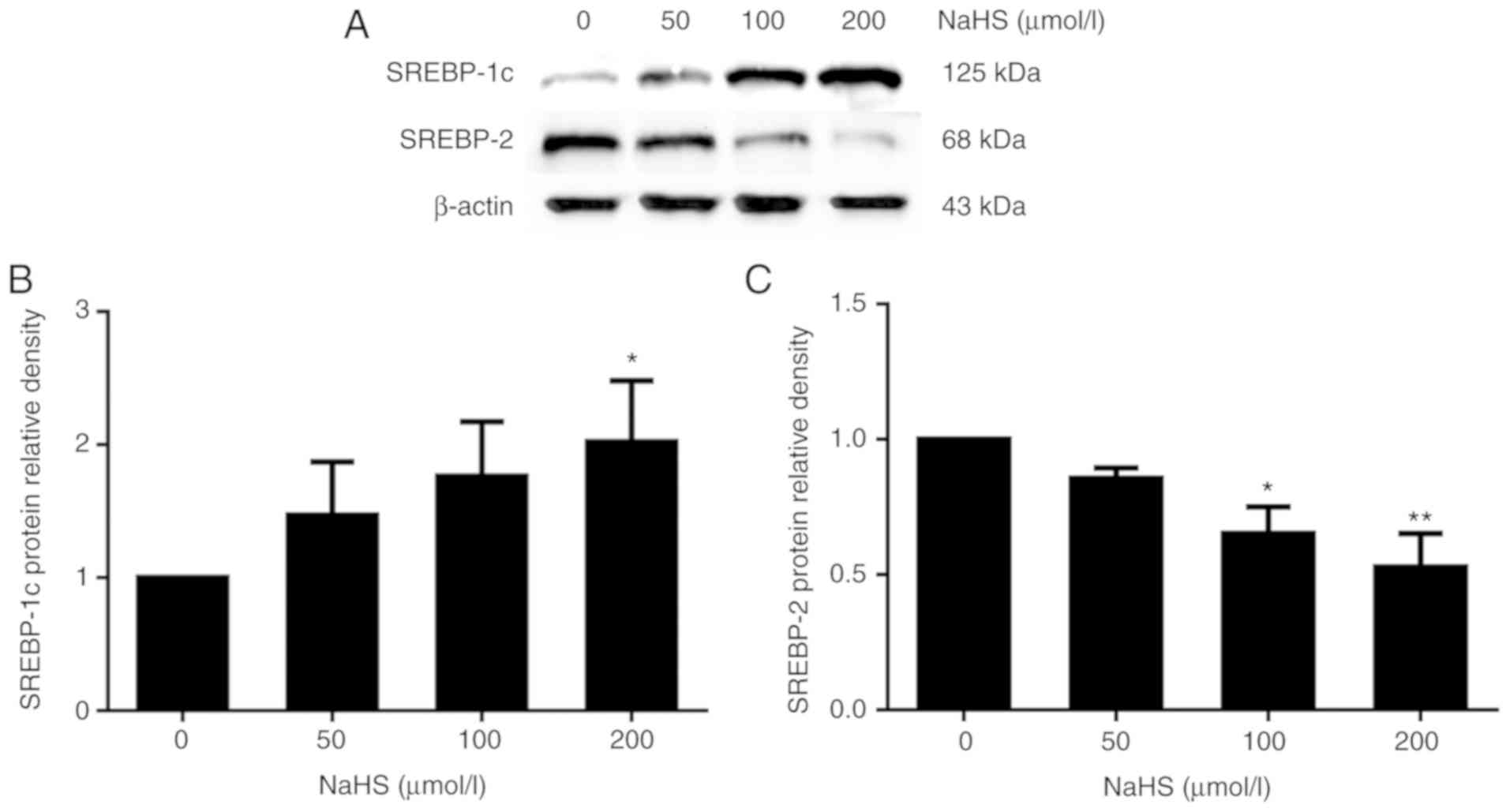
It has been reported that SREBP-2 may upregulate
PCSK9 expression (10). Thus, to
explore the potential mechanisms underlying the H2S
regulation if PCSK9 expression, we investigated whether
H2S regulates PCSK9 expression by regulating SREBP-2
expression. Following treatment with various concentrations of NaHS
(0, 50, 100 and 200 µmol/l) for 24 h, SREBP-1c and SREBP-2
expression was detected by western blot analysis. As shown in
Fig. 7, SREBP-1c expression
increased and that of SREBP-2 decreased with the increasing NaHS
concentration.
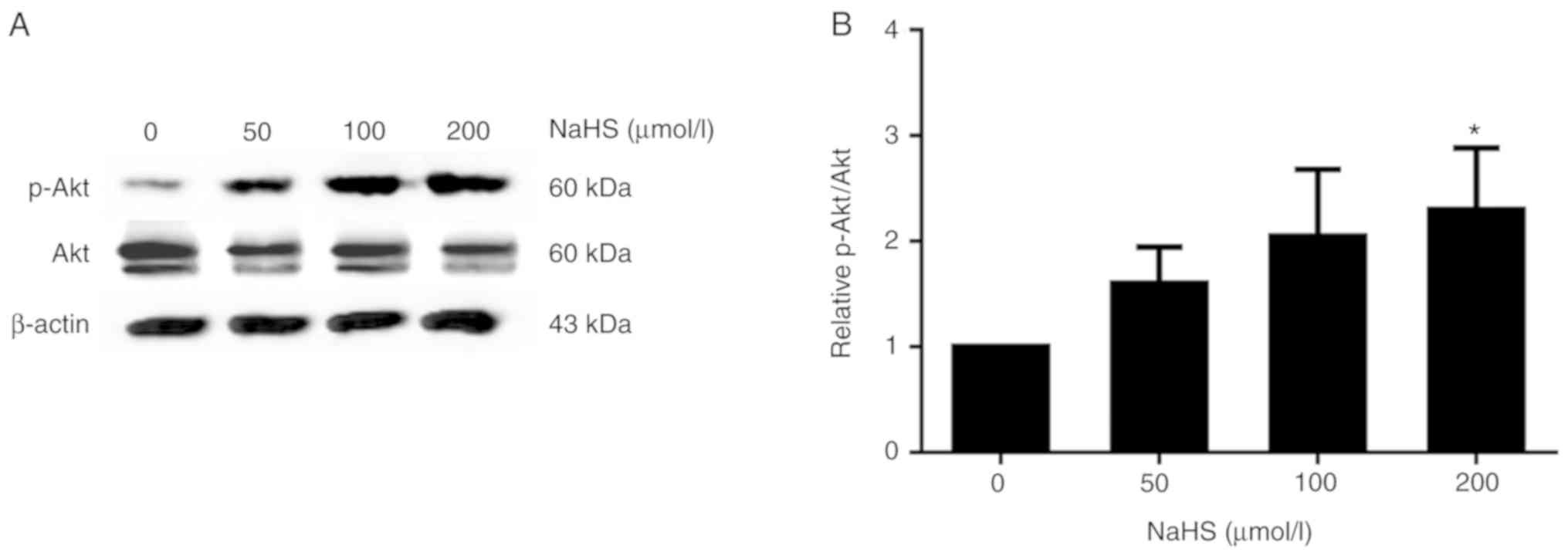
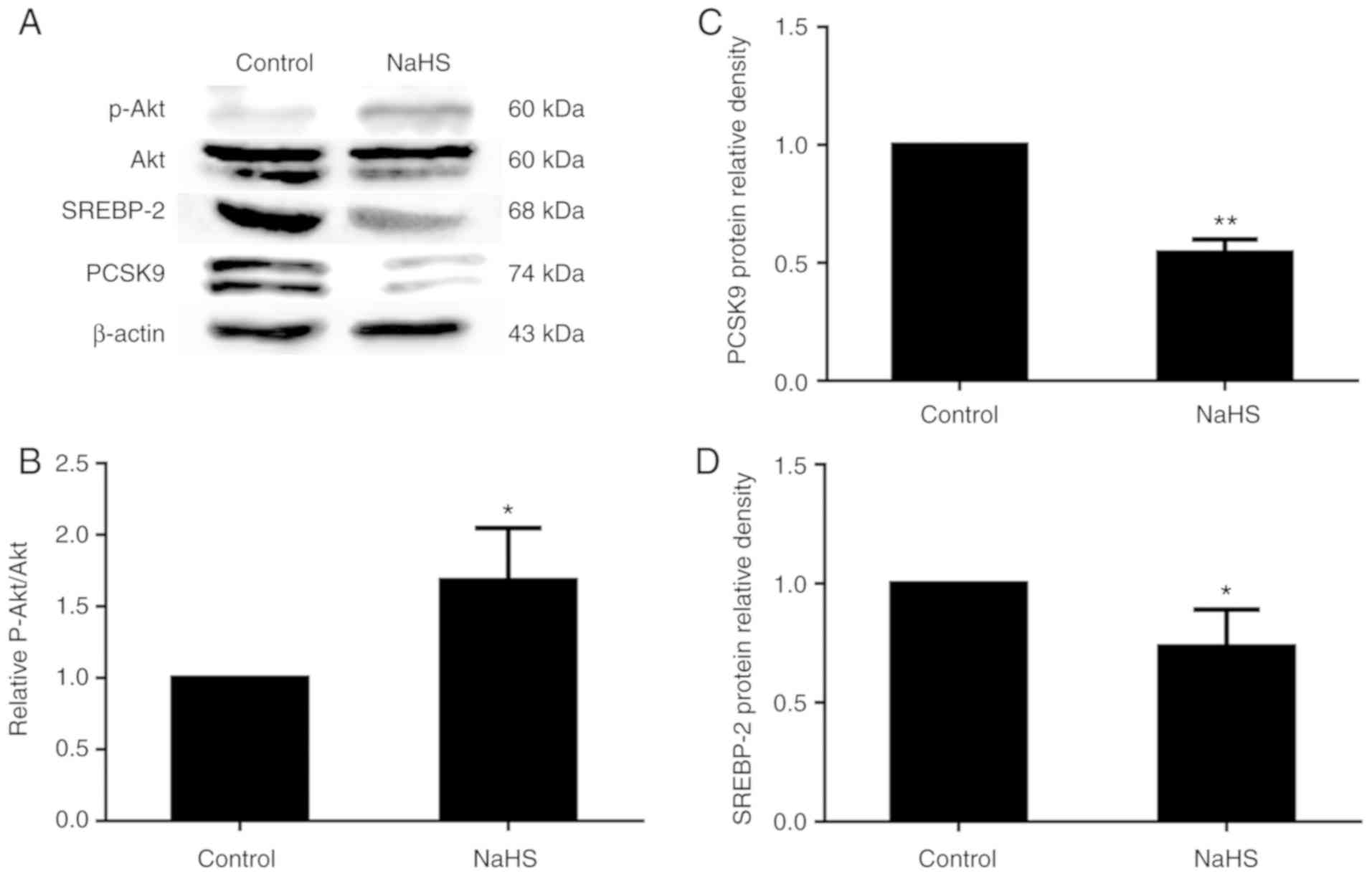
PI3K/Akt is known to regulate the expression of
SREBPs (11). Thus, in this
study, we treated the HepG2 cells with various concentrations of
NaHS (0, 50, 100 and 200 µmol/l) for 24 h, and
phosphorylated Akt protein expression was detected by western blot
analysis. As shown in Fig. 8,
phosphorylated Akt protein expression increased with the increasing
NaHS concentration. Following treatment with 200 µmol/l
NaHS, the level of Akt phosphorylation increased, and SREBP-2 and
PCSK9 expression decreased accordingly (Fig. 9).
H2S reduces lipid accumulation
in HepG2 cells
The above-mentioned results indicated that
H2S increased lipid uptake. Therefore, we hypothesized
that lipid accumulation may be increased in the HepG2 cells. To
explore whether H2S influences lipid accumulation in the
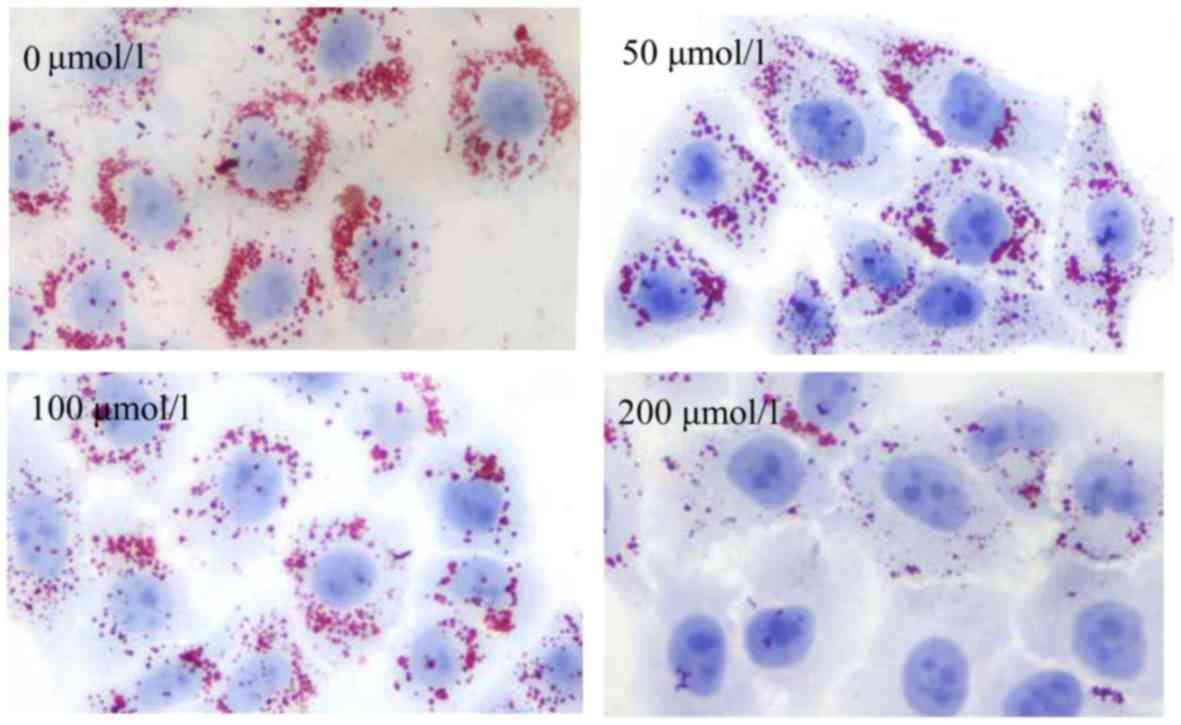
HepG2 cells, the HepG2 cells were incubated with culture medium
supplemented with 30% FBS for 24 h, and intracellular lipids were
detected by Oil Red O staining. Surprisingly, as shown in Fig. 10, lipid accumulation in the HepG2
cells exhibited no increase, but exhibited a decrease with the
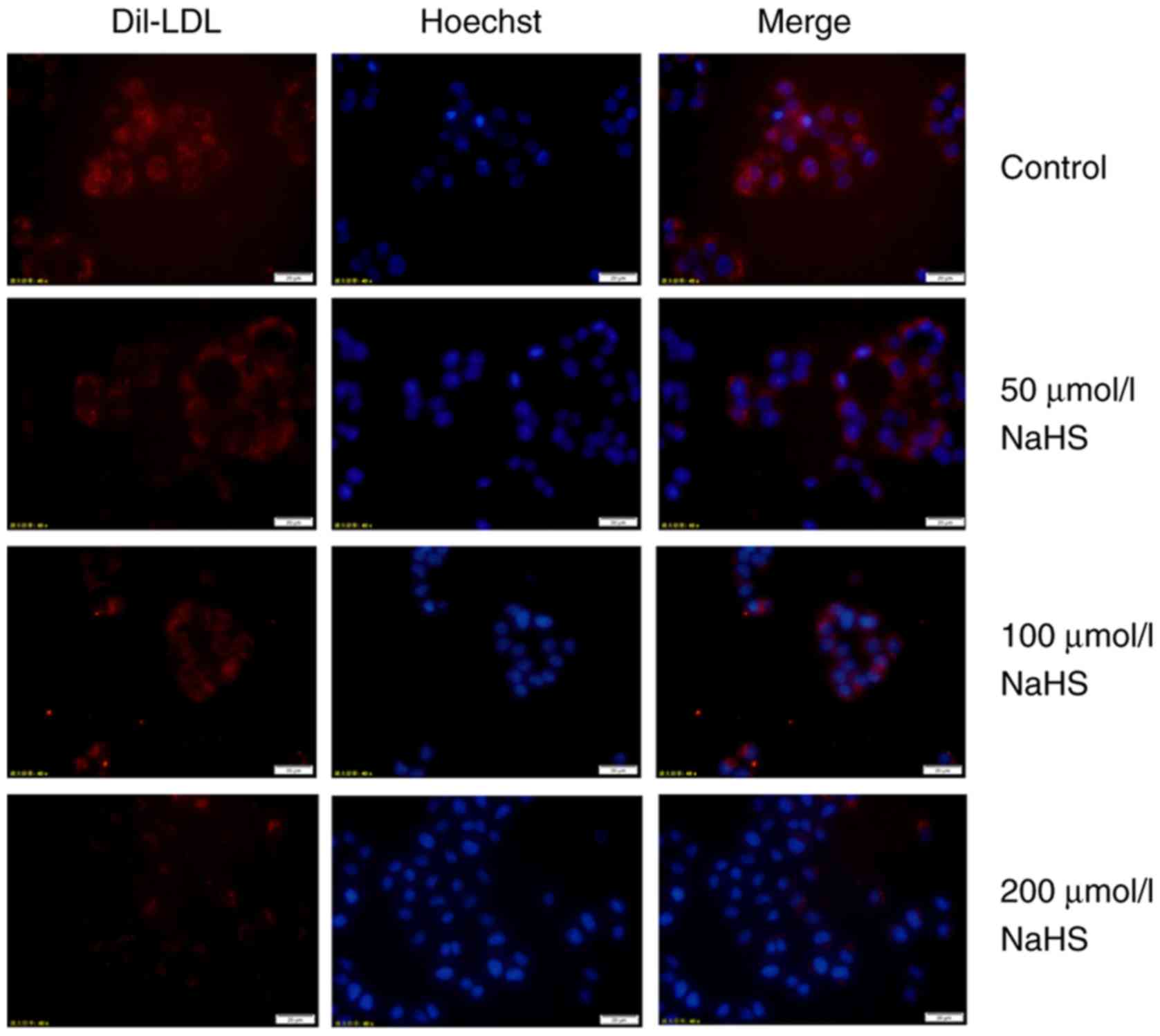
increasing H2S concentration. Subsequently, DiI-LDL was
incubated with the HepG2 cells for 24 h, and intracellular red
fluorescence was also reduced, suggesting the reduced lipid
accumulation in HepG2 cells (Fig.
11). The above-mentioned results indicate that H2S
can inhibit lipid accumulation in HepG2 cells induced by high serum
concentrations.
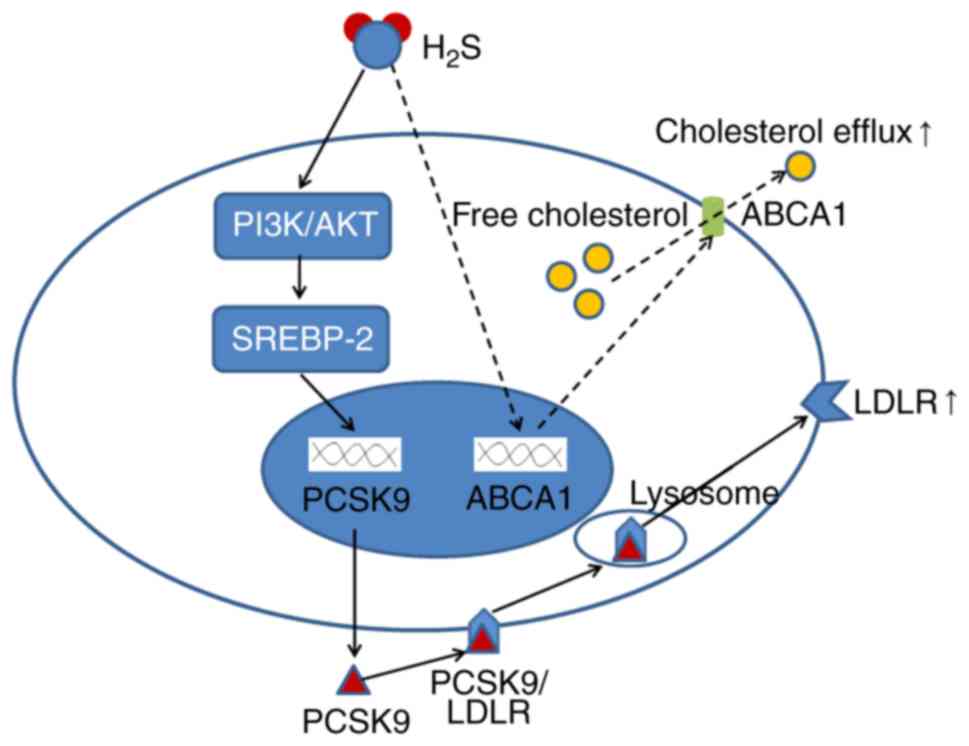
Taken together, these results indicated that
H2S downregulated the expression of PCSK9 in a time- and
concentration-dependent manner, thereby regulating HepG2 lipid
metabolism (Fig. 12).
Discussion
H2S inhibits PCSK9 expression
to regulate lipid metabolism in HepG2 cells
H2S has emerged as an important
cardiovascular signaling molecule similar to nitric oxide and
carbon monoxide (12).
H2S carries out a wide range of physiological and
pathological functions in the cardiovascular system, including
maintaining endothelial homeostasis (13), inhibiting the proliferation of
vascular smooth muscle cells (14), and regulating lipid metabolism
(15). However, the precise
mechanisms through which H2S carries out its functions
remain unknown. In this study, we found that H2S
inhibited the expression of PCSK9 and increased lipid uptake by
HepG2 cells.
The PCSK9 gene was discovered in 2003 (16); it plays an important role in lipid
metabolism (17). PCSK9
accelerates the degradation of hepatic LDLR and prevents its
recycling to the cell surface, which reduces lipid removal from the
liver and increases serum lipid levels, thus promoting
atherogenesis (18). PCSK9
mutations cause autosomal dominant hypercholesterolemia (19). Numerous factors, such as statins
(20) and berberine (21), regulate the expression of PCSK9.
This study demonstrated that H2S downregulated the
expression of PCSK9 in HepG2 cells in a concentration- and
time-dependent manner. To the very best of our knowledge, this
study is the first to report that H2S can regulate the
expression of PCSK9.
Previous studies have confirmed that PCSK9 regulates
cellular lipid metabolism by degrading LDLR on the cell surface
(22,23). The overexpression of PCSK9 has
been shown to significantly increase LDLR degradation in HepG2
cells, thus reducing LDLR expression (24). In this study, H2S
treatment upregulated the expression of LDLR by decreasing the
expression of PCSK9 in HepG2 cells. The expression of LDLR further
increased after the expression of PCSK9 was interfered with by
siRNA. Consistent with this result, following incubation with
DiI-LDL for 4 h, DiI-LDL uptake by HepG2 cells increased in the 200
µmol/l NaHS and PCSK9 siRNA groups compared with that in the
200 µmol/l NaHS group. This result indicates that
H2S reduces the expression of PCSK9, thereby inhibiting
the degradation of LDLR by PCSK9, and increasing the expression of
LDLR on the cell surface to enhance lipid uptake. In addition, the
highest concentration of NaHS (200 µmol/l) used in the
treatment of HepG2 cells was confirmed by previous experiments, and
this concentration does not cause toxic effects on cells (25).
PI3K/Akt-SREBP-2 is involved in the
regulation of PCSK9 and LDLR expression by H2S
SREBPs play an important role in lipid metabolism by
regulating the expression of lipid metabolism-related genes
(26). SREBP-1c primarily
enhances the synthesis of fatty acids and the transcription of
metabolic enzymes, while SREBP-2 primarily regulates the
transcription of cholesterol-metabolizing enzymes (27). SREBP-1c and SREBP-2 are the
predominant isomers in the liver (27). The promoter region of PCSK9
contains a sterol-regulatory element, which implies that PCSK9
transcription depends on sterols (28). Dubuc et al demonstrated
that PCSK9 is a target gene of SREBP-2 (29), whereas SREBP-2 may participate in
lipid metabolism by regulating PCSK9 expression. In this study,
following treatment with various concentrations of H2S,
the expression of SREBP-1c and SREBP-2 was detected in HepG2 cells.
The results revealed that the expression of SREBP-1c increased,
while that of SREBP-2 decreased with the increasing NaHS
concentration. This opposite result may be due to the fact that
SREBP-1c primarily enhances the synthesis of fatty acids and
triglycerides, while SREBP-2 preferentially synthesizes
cholesterol. Chae et al demonstrated that LDLR was increased
by downregulating the expression of hepatic PCSK9 via SREBP-2
(30). In this study,
H2S inhibited the expression of SREBP-2 to inhibit the
expression of PCSK9 and reduced the degradation of LDLR by PCSK9 to
increase the LDLR level on the cell surface, consistent with the
findings in the study by Chae et al (30).
PI3K/Akt can upregulate SREBP-2 expression and
participate in lipid metabolism regulation (31). In this study, following treatment
with various concentrations of NaHS, the levels of phosphorylated
Akt were detected in the HepG2 cells. The results revealed that
H2S increased the content of phosphorylated Akt protein
to activate the PI3K/Akt signaling pathway, consistent with the
findings of the study by Zheng et al (32). The increased Akt phosphorylation
resulted in the decreased expression of SREBP-2. As a positive
transcriptional regulator of PCSK9, the inhibition of SREBP-2 by
phosphorylated Akt also downregulated the expression of PCSK9. The
above-mentioned results indicate that PI3K/Akt-SREBP-2 is involved
in the regulation of PCSK9 and LDLR expression by
H2S.
Notably, this study demonstrated that H2S
inhibited PCSK9 expression to reduce LDLR degradation at the HepG2
cell membrane to increase lipid uptake in the HepG2 cells. However,
following incubation with 30% FBS or DiI-LDL, lipid accumulation in
the HepG2 cells exhibited no increase, but instead decreased. This
result suggests that H2S may also promote lipid efflux
in addition to promoting lipid uptake through the SREBP-2
PCSK9/LDLR pathways in HepG2 cells. Gong et al (33) reported that H2S
upregulated ATP-binding cassette transporter A1 (ABCA1) expression,
whereas ABCA1 played an important role in promoting intracellular
cholesterol efflux, which may explain why H2S increases
lipid uptake, but causes no increase in lipid accumulation in HepG2
cells.
Notably, H2S can affect pH and
Na+, but whether pH or Na+ affects lipid
metabolism in HepG2 cells remains unclear. Therefore, further
studies are required to determine the effects of pH or
Na+ on lipid metabolism in HepG2 cells and whether other
liver cancer cells have the same effect as HepG2.
In conclusion, the findings of this study
demonstrate that H2S downregulates the expression of
PCSK9 in a time- and concentration-dependent manner, thereby
regulating HepG2 lipid metabolism (Fig. 12). To the very best of our
knowledge, this study is the first to report that H2S
can regulate the expression of PCSK9. Our findings provide new
insight into the understanding of H2S-regulated lipid
metabolism and anti-atherosclerosis. More importantly, the findings
of this study suggest that H2S may be a novel candidate
as PCSK9 inhibitor in addition to PCSK9 monoclonal antibodies and
may provide a novel method with which to discover novel PCSK9
inhibitors (34).
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (no. 81370376), the
Natural Science Foundation of Hunan province (nos. 2018JJ2343 and
2015JJ4097), the Key Project of the Educational Department of
Hu-nan Province (no. 15A137), the Visiting Scholar Foundation of
Key Laboratory of Biorheological Science and Technology (Chongqing
University), Ministry of Education (nos. CQKLBST-2014-002 and
CQKLBST-2015-004), and the first-class discipline in Hunan
Province, and Undergraduate Innovative Experiment Project (no.
2018XJXZ369).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LSL and MMW contributed to the conception and design
of the experiment. JX, XQB, LL, MZ, JP and QX performed the
experiments and collected the data. ZR, ZSJ, ZHT, HYW and MMW
analyzed the data and helped interpret the results. JX, XQB and LL
wrote the manuscript. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Li D, Xiong Q, Peng J, Hu B, Li W, Zhu Y
and Shen X: Hydrogen Sulfide upregulates the expression of
ATP-binding cassette transporter A1 via promoting nuclear
translocation of PPARα. Int J Mol Sci. 17:E6352016. View Article : Google Scholar
|
|
2
|
Wu D, Zheng N, Qi K, Cheng H, Sun Z, Gao
B, Zhang Y, Pang W, Huangfu C, Ji S, et al: Exogenous hydrogen
sulfide mitigates the fatty liver in obese mice through improving
lipid metabolism and antioxidant potential. Med Gas Res. 5:12015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li H, Mani S, Wu L, Fu M, Shuang T, Xu C
and Wang R: The interaction of estrogen and CSE/H2S
pathway in the development of atherosclerosis. Am J Physiol Heart
Circ Physiol. 312:H406–H414. 2017. View Article : Google Scholar
|
|
4
|
Gao L, Xu Z, Yin Z, Chen K, Wang C and
Zhang H: Association of hydrogen sulfide with alterations of
monocyte chemokine receptors, CCR2 and CX3CR1 in patients with
coronary artery disease. Inflamm Res. 64:627–635. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lin XL, Xiao LL, Tang ZH, Jiang ZS and Liu
MH: Role of PCSK9 in lipid metabolism and atherosclerosis. Biomed
Pharmacother. 104:36–44. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shapiro MD and Fazio S: PCSK9 and
atherosclerosis-lipids and beyond. J Atheroscler Thromb.
24:462–472. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shahreyar M, Salem SA, Nayyar M, George
LK, Garg N and Koshy SKG: Hyperlipidemia: Management with
proprotein convertase subtilisin/kexin type 9 (PCSK9) inhibitors. J
Am Board Fam Med. 31:628–634. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ochin CC and Garelnabi M: Berberine
encapsulated PLGA-PEG nanoparticles modulate PCSK-9 in HepG2 Cells.
Cardiovasc Hematol Disord Drug Targets. 18:61–70. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tavori H, Rashid S and Fazio S: On the
function and homeostasis of PCSK9: Reciprocal interaction with LDLR
and additional lipid effects. Atherosclerosis. 238:264–270. 2015.
View Article : Google Scholar :
|
|
10
|
Jeong HJ, Lee HS, Kim KS, Kim YK, Yoon D
and Park SW: Sterol-dependent regulation of proprotein convertase
subtilisin/kexin type 9 expression by sterol-regulatory element
binding protein-2. J Lipid Res. 49:399–409. 2008. View Article : Google Scholar
|
|
11
|
Krycer JR, Sharpe LJ, Luu W and Brown AJ:
The Akt-SREBP nexus: Cell signaling meets lipid metabolism. Trends
Endocrinol Metab. 21:268–276. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cao X, Ding L, Xie ZZ, Yang Y, Whiteman M,
Moore PK and Bian JS: A review of hydrogen sulfide synthesis,
metabolism, and measurement: Is modulation of hydrogen sulfide a
novel therapeutic for cancer? Antioxid Redox Signal. 2018.
View Article : Google Scholar
|
|
13
|
Altaany Z, Moccia F, Munaron L, Mancardi D
and Wang R: Hydrogen sulfide and endothelial dysfunction:
Relationship with nitric oxide. Curr Med Chem. 21:3646–3661. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang Y, Wang X, Liang X, Wu J, Dong S, Li
H, Jin M, Sun D, Zhang W and Zhong X: Inhibition of hydrogen
sulfide on the proliferation of vascular smooth muscle cells
involved in the modulation of calcium sensing receptor in high
homocysteine. Exp Cell Res. 347:184–191. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cheung SH and Lau JYW: Hydrogen sulfide
mediates atheroprotection against oxidative stress via
S-sulfhydration. PLoS One. 13:e01941762018. View Article : Google Scholar
|
|
16
|
Abifadel M, Varret M, Rabès JP, Allard D,
Ouguerram K, Devillers M, Cruaud C, Benjannet S, Wickham L, Erlich
D, et al: Mutations in PCSK9 cause autosomal dominant
hypercholesterolemia. Nat Genet. 34:154–156. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Singh A and Davidson M: Update on PCSK9
therapies for the treatment of dyslipidemia. Expert Rev Endocrinol
Metab. 11:87–95. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shapiro MD, Tavori H and Fazio S: PCSK9:
From basic science discoveries to clinical trials. Circ Res.
122:1420–1438. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Elbitar S, Susan-Resiga D, Ghaleb Y, El
Khoury P, Peloso G, Stitziel N, Rabès JP, Carreau V, Hamelin J,
Ben-Djoudi-Ouadda A, et al: New sequencing technologies help
revealing unexpected mutations in autosomal dominant
hypercholesterolemia. Sci Rep. 8:19432018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Reiss AB, Shah N, Muhieddine D, Zhen J,
Yudkevich J, Kasselman LJ and DeLeon J: PCSK9 in cholesterol
metabolism: From bench to bedside. Clin Sci (Lond). 132:1135–1153.
2018. View Article : Google Scholar
|
|
21
|
Spigoni V, Aldigeri R, Antonini M, Micheli
MM, Fantuzzi F, Fratter A, Pellizzato M, Derlindati E, Zavaroni I,
Bonadonna RC and Dei Cas A: Effects of a new nutraceutical
formulation (berberine, red yeast rice and chitosan) on non-HDL
cholesterol levels in individuals with dyslipidemia: Results from a
randomized, double blind, placebo-controlled study. Int J Mol Sci.
18:E14982017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Poirier S, Hamouda HA, Villeneuve L,
Demers A and Mayer G: Trafficking dynamics of PCSK9-induced LDLR
degradation: Focus on human PCSK9 mutations and C-terminal domain.
PLoS One. 11:e01572302016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tavori H, Giunzioni I, Predazzi IM,
Plubell D, Shivinsky A, Miles J, Devay RM, Liang H, Rashid S,
Linton MF and Fazio S: Human PCSK9 promotes hepatic lipogenesis and
atherosclerosis development via apoE- and LDLR-mediated mechanisms.
Cardiovasc Res. 110:268–278. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lagace TA, Curtis DE, Garuti R, McNutt MC,
Park SW, Prather HB, Anderson NN, Ho YK, Hammer RE and Horton JD:
Secreted PCSK9 decreases the number of LDL receptors in hepatocytes
and in livers of parabiotic mice. J Clin Invest. 116:2995–3005.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qu K, Liu YM, He XL, Zhang H, Zhang K,
Peng J, Tang YL, Yu XH, Zeng JF, Lei JJ, et al: H2 S inhibits
apo(a) expression and secretion through PKCα/FXR and Akt/HNF4α
pathways in HepG2 cells. Cell Biol Int. 40:906–916. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Park SW, Moon YA and Horton JD:
Post-transcriptional regulation of low density lipoprotein receptor
protein by proprotein convertase subtilisin/kexin type 9a in mouse
liver. J Biol Chem. 279:50630–50638. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Horton JD, Goldstein JL and Brown MS:
SREBPs: Activators of the complete program of cholesterol and fatty
acid synthesis in the liver. J Clin Invest. 109:1125–1131. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lebeau P, Al-Hashimi A, Sood S, Lhoták Š,
Yu P, Gyulay G, Paré G, Chen SR, Trigatti B, Prat A, et al:
Endoplasmic reticulum stress and Ca2+ depletion
differentially modulate the sterol regulatory protein PCSK9 to
control lipid metabolism. J Biol Chem. 292:1510–1523. 2017.
View Article : Google Scholar
|
|
29
|
Dubuc G, Chamberland A, Wassef H, Davignon
J, Seidah NG, Bernier L and Prat A: Statins upregulate PCSK9, the
gene encoding the proprotein convertase neural apoptosis-regulated
convertase-1 implicated in familial hypercholesterolemia.
Arterioscler Thromb Vasc Biol. 24:1454–1459. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chae HS, You BH, Kim DY, Lee H, Ko HW, Ko
HJ, Choi YH, Choi SS and Chin YW: Sauchinone controls hepatic
cholesterol homeostasis by the negative regulation of PCSK9
transcriptional network. Sci Rep. 8:67372018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang J, Chen S, Cai D, Bian D and Wang F:
Long noncoding RNA lncARSR promotes hepatic cholesterol
biosynthesis via modulating Akt/SREBP-2/HMGCR pathway. Life Sci.
203:48–53. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zheng D, Chen Z, Chen J, Zhuang X, Feng J
and Li J: Exogenous hydrogen sulfide exerts proliferation,
anti-apoptosis, migration effects and accelerates cell cycle
progression in multiple myeloma cells via activating the Akt
pathway. Oncol Rep. 36:1909–1916. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gong D, Cheng HP, Xie W, Zhang M, Liu D,
Lan G, Huang C, Zhao ZW, Chen LY, Yao F, et al: Cystathionine
γ-lyase(CSE)/hydrogen sulfide system is regulated by miR-216a and
influences cholesterol efflux in macrophages via the PI3K/AKT/ABCA1
pathway. Biochem Biophys Res Commun. 470:107–116. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
He NY, Li Q, Wu CY, Ren Z, Gao Y, Pan LH,
Wang MM, Wen HY, Jiang ZS, Tang ZH and Liu LS: Lowering serum
lipids via PCSK9-targeting drugs: Current advances and future
perspectives. Acta Pharmacol Sin. 38:301–311. 2017. View Article : Google Scholar : PubMed/NCBI
|