Introduction
Small bowel transplantation is the most effective
treatment of intestinal failure (1); however, the high incidence of
intestinal tissue injury, and the risk of acute and chronic
rejection following transplantation are difficulties associated
with small bowel transplantation (2). Although the development of novel
immunosuppressive agents has reduced the incidence of rejection and
significantly prolonged the survival of grafts, damage to function
and the poor postoperative recovery of intestinal epithelial cells
remain problems that require urgent resolution (3). Previous studies have found that bone
marrow mesenchymal stem cells (BMMSCs) are able to repair damaged
cells or tissues (4,5), with obvious biological and ethical
advantages (6,7), and is a potential cellular source
for cell therapy strategies (8).
In addition, animal experiments have demonstrated that the
transfusion of BMMSCs following small bowel transplantation can
significantly improve the function and repair the damage of the
transplanted intestinal tract (9). However, the viability of BMMSCs and
their capacity to home to lesions following transfusion into the
recipient are limited (10), and
the majority of the cells die within a few hours (11) even if the BMMSCs were transfused
into the local lesion site. Heme oxygenase-1 (HO-1) and its
metabolites have been found to exhibit anti-oxidative,
anti-inflammatory, antiproliferative and immunomodulatory effects
(12). Transfection with HO-1 can
improve the transformation and anti-oxidative capacity of BMMSCs
(13), thereby enhancing the
activity of BMMSCs and prolonging the duration in which BMMSCs
serve a role (14).
CXC-chemokine receptor 3 (CXCR3) is a G
protein-coupled seven-subunit transmembrane receptor expressed in
damaged parenchymal cells in lesions from multiple organs, and in
inflammatory cells, including activated lymphocytes, macrophages
and dendritic cells (15). CXCR3
receptors bind to their specific ligands [monokine induced by
interferon (IFN)-γ, IFN-γ-inducible protein 10 and IFN-γ-inducible
T-cell α chemoattractant], and recruit CXCR3-expressing cells to
the site of injury (16,17). It has been shown that, following
transfection with the CXCR3 gene, BMMSCs are chemotactically
directed to migrate to a transplanted intestine; this was shown by
the increased homing of BMMSCs and inhibited chemotaxis of T cells
to transplanted intestine, resulting in protective effects on the
transplanted small bowel (18).
In our preliminary study, HO-1 and CXCR3 genes were transfected
into BMMSCs in vitro, and subsequently transfused into a rat
model of rejection to small bowel transplantation. The results
showed that the HO-1 and CXCR3 gene-modified BMMSCs were
significantly increased in the damaged sites, and the function of
the damaged intestine quickly recovered (18). However, the mechanism by which the
recovery of intestinal function was achieved remains to be
elucidated and requires further investigation.
It has been found that mitogen-activated protein
kinases (MAPKs) are important intracellular signal transduction
systems that serve an important role in the development of
inflammatory responses (19)
induced by trauma (20),
infection (21) and
ischemia-reperfusion (22). The
activation of the p38-MAPK pathway is important in cellular damage,
including the production of proinflammatory cytokines, cytoskeletal
remodeling and cellular apoptosis (23). However, whether the p38-MAPK
signaling pathway is involved in the recovery of transplanted
intestinal function by BMMSCs has not been reported. In addition,
tumor necrosis factor (TNF)-α has been shown to be involved in the
recovery of transplanted intestinal function by CXCR3/HO-1
gene-modified BMMSCs in rats (18). The rat intestinal epithelial crypt
cell line-6 (IEC-6) cell line is derived from the intestinal crypt
of normal rats, which can be applied to the in vitro
investigation of intestinal mucosal injury (24). Therefore, in the present study, an
in vitro model of damaged intestinal epithelial cells was
established using IEC-6 cells treated with TNF-α to investigate the
effects of CXCR3/HO-1 gene-modified BMMSCs on intestinal epithelial
cell damage.
Materials and methods
Acquisition and transfection of
BMMSCs
All animals were provided by the Experimental Animal
Center of the Academy of Military Medical Sciences (Beijing, China)
and were housed individually in standard animal facilities at
18-26°C and 40-70% relative humidity with a 12-h light/dark cycle,
and were provided with commercially available chow and tap water
ad libitum. All experiments involving animals followed the
experimental animal ethical regulations, and were approved by the
Ethics Committee of Tianjin First Central Hospital (Tianjin,
China). Healthy male Lewis rats (n=15) aged 2-3 weeks and weighing
40-60 g were sacrificed by cervical dislocation and soaked in 75%
alcohol for 5-10 min. The bone marrow cells were cultured in
DMEM-F12 (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
medium containing 10% fetal bovine serum (FBS; Biowest, Loire
Valley, France) and the third generation of the cultured cells was
prepared for further use (18).
The third generation BMMSCs were induced to differentiate into
adipocytes and osteoblasts, and subjected to phenotypic assessment
(25). The third generation
BMMSCs were inoculated into a 75-mm culture flask with
5×106 cells/flask containing 20 ml complete medium
(DMEM-F12 containing 10% FBS). The culture solution was discarded
following culture for 24 h. Subsequently, 5 ml of each culture
medium was added to the different BMMSC groups as follows: MSCs
with culture medium only; Ad/MSCs with medium containing
adenovirus; Ad-HO/MSCs with medium containing the HO-1 transgene;
Ad-CXCR3/MSCs with medium containing the CXCR3 transgene; Ad-(CXCR3
+ HO)/MSCs with medium containing the HO-1 and CXCR3 transgenes
(all Shanghai Genechem Co., Ltd.). The BMMSC to transgene ratio was
1:10 (MOI=10). Following incubation for 6 h, the medium containing
the transfection solution was discarded and 20 ml complete medium
was added for 48 h. Fluorescent protein expression was observed
under a fluorescence microscope (Nikon Corporation, Tokyo, Japan,)
in the dark. These cells were used in subsequent experiments.
In vitro model of intestinal cell
damage
The IEC-6 cells (National Infrastructure of Cell
Line Resource, Beijing, China) were inoculated in a six-well plate
at a density of 2×105 cells/well. After 24 h, the
culture medium was discarded, and the IEC-6 cells were cultured in
complete RPMI-1640 medium containing 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) and 100 ng/ml TNF-α (PeproTech, Inc., Rocky Hill,
NJ, USA) for 48 h to establish the model of damaged IEC-6 cells
(TNF-α/IEC-6) (26). Rat
lymphocytes were obtained using the method described by Yin et
al (18).
The cells were divided into nine experimental
groups: i) Control group, untreated IEC-6 cells; ii) Model group,
TNF-α/IEC-6; iii) L group, TNF-α/IEC-6 + lymphocytes; iv) S + L
group, p38 inhibitor + TNF-α/IEC-6 + lymphocytes; v) MSCs + L
group, BMMSCs + TNF-α/IEC-6 + lymphocytes; vi) Ad/MSCs + L group,
Ad/BMMSCs + TNF-α/IEC-6 + lymphocytes; vii) Ad-HO/MSCs + L group,
Ad-HO-1/BMMSCs + TNF-α/IEC-6 + lymphocytes; viii) Ad-CXCR3/MSCs + L
group, Ad-CXCR3/BMMSCs + TNF-α/IEC-6 +lymphocytes; and ix)
Ad-(CXCR3 + HO)/MSCs + L group, Ad-(CXCR3 + HO-1)/BMMSCs +
TNF-α/IEC-6 + lymphocytes. The TNF-α/IEC-6 cells were prepared in
the lower Transwell (Corning Inc., Corning, NY, USA) layer, whereas
the BMMSCs (1×106 cells/well) and lymphocyte
(5×106 cells/well) were added to the upper layer of the
Transwell chamber. The cells were co-cultured for 24 h and then
collected following the experiment.
Chemotaxis
The experimentally-treated Transwell chambers were
fixed (anhydrous methanol: Glacial acetic acid 3:1) for 30 min,
stained with a 2% crystal violet dye solution for 30 min and washed
with phosphate-buffered saline (PBS). The upper layer of cells was
removed with a cotton swab, peeled off and placed on a slide, fixed
with neutral gum, and then observed under a Ti2-E inverted
microscope, (Nikon Corporation). The TNF-α/IEC-6 cells were
prepared in a 35-mm diameter well and added to a Transwell chamber
containing Ad/MSCs, Ad-CXCR3,/MSCs or Ad-(CXCR3 + HO)/MSCs. The
green fluorescent protein (GFP) signal was locked with a living
cell workstation microscope, and GFP-expressing BMMSCs located 5
µm below the Transwell membrane were observed to record the
time at which when BMMSCs began to appear.
Cell viability
The BMMSCs and IEC-6 cells in each of the groups
were trypsinized and adjusted to a density of 1×105
cells/ml. To each well, 200 µl of the cell suspension was
added according to the different groups, and 200 µl complete
medium containing 10% FBS was added to the blank control wells. The
cells were incubated at 37°C for 24 h, following which 10 µl
of the CCK-8 solution (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan) was added to each well. Following culture for 3 h,
the absorbance was measured at 450 nm with an enzyme standard
instrument (BioTek Synergy 2, BioTek Instruments, Inc., Winooski,
VT, USA) to calculate the cellular viability.
Immunohistochemistry
Third passage BM MSCs were placed into six-well
plates containing a cell slide, at a density of 2.5×105
cells/well. After 24 h, transfection was performed, and different
BMMSCs were obtained. The culture solution was discarded, the
BMMSCs were washed with PBS, fixed with 4% paraformaldehyde for 30
min, washed with PBS, and treated with 1% NP40 (both Beijing
Solarbio Science and Technology Co., Ltd., Beijing, China) for 30
min to rupture the cell membrane. Following washing with PBS and
blocking with 10% goat serum (Minghai, Lanzhou, China) for 30 min,
each slide was then stained with a mixture of anti-HO-1 mouse IgG
(1:100; cat. no. ab13248; Abcam, Cambridge, UK) and anti-CXCR3
rabbit IgG (1:100; cat. no. bs-2209R; Affinity Bioreagents, Inc.,
Golden, CO, USA) at 4°C overnight. Following allowing the slides to
recover to room temperature for 30 min and washing via dipping in
PBS, all slides were stained with a secondary antibody mixture of
Alexa Fluor 594 (1:100; cat. no. ZF-0513; OriGene Technologies,
Inc., Beijing, China)-conjugated goat anti-mouse IgG and Alexa
Fluor 488-conjugated goat anti-rabbit IgG (1:100; cat. no. ZF-0511;
OriGene Technologies, Inc.). The slides were incubated at 37°C for
1 h, washed with PBS, stained with 4′,6-diamidino-2-phe-nylindole
(10 µg/ml; Beijing Solarbio Science and Technology Co.,
Ltd.) for 2 min, rinsed with running water, and observed under a
laser confocal microscope (Nikon Corporation). The IEC-6 cells were
cultured on the slide in advance, and different IEC-6 groups were
obtained following exposure to different treatments. The steps were
the same as above, and the slides were stained separately with the
following antibodies: Antizonula occludens-1 (ZO-1) rabbit IgG
(1:100; cat. no. 21773-1-AP; ProteinTech Group, Inc., Chicago, IL,
USA), followed by Alexa Fluor 488-conjugated goat anti-rabbit IgG
(1:50), or the anti-phosphorylated (p)-C/EBP homologous protein-10
(p-CHOP10) rabbit IgG (1:100; cat. no. bs-51772; BIOSS, Beijing,
China), followed by secondary Alexa Fluor 594-horseradish
peroxidase (HRP)-conjugated goat anti-rabbit IgG (1:50). Staining
of apoptosis was performed for each IEC-6 group using TUNEL kits
(Roche Diagnostics, Basel, Switzerland) according to the
manufacturer's protocol.
The paraffin-fixed small intestine tissues were
immunohistochemically stained. The tissue sections were incubated
at 70°C for 50 min, deparaffinized with xylene, hydrated with
gradient alcohol, and citrated for 15 min at 95°C, and then blocked
with goat serum for 1 h. The paraffin sections were stained with
anti-p-CHOP10 rabbit IgG (1:100) overnight at 4°C. Following
incubation at room temperature for 30 min, the sections were
stained with HRP (Thermo Fisher Scientific, Inc.) for 1 h at 37°C.
Following DAB staining, hematoxylin counterstaining, alcohol
dehydration and transparency, the tissue sections were fixed with
neutral gum and were observed under the Eclipse Ni-U positive
fluorescence microscope (Nikon Corporation).
Western blot analysis
The BMMSCs from each of the groups were exposed to
500 µl RIPA lysis buffer on ice for 30 min. Following
centrifugation at 13,000 × g for 5 min at 4°C, the supernatant was
obtained to assess the concentrations of various proteins. Protein
determination was performed using a bicinchoninic acid assay (Wuhan
Boster Biological Technology, Ltd., Wuhan, China). SDS-PAGE (10%)
was used for HO-1 and CXCR3. A total of 30 µg proteins/well
in the gels underwent electrophoresis, and were transferred onto a
nitrocellulose membrane (Beijing Solarbio Science and Technology
Co., Ltd.) and blocked with skimmed milk (BD Biosciences, Franklin
Lakes, NJ, USA) for 2 h at room temperature, following which the
membranes were stained with anti-HO-1 mouse IgG (1:400) or
anti-CXCR3 rabbit IgG (1:800) or β-actin rabbit IgG (1:2,000; cat.
no. 21338; SAB, College Park, Maryland, United States) at 4°C
overnight. The membranes were washed three times with TBST, stained
with goat anti-rabbit IgG (1:1,000; cat. no. zb2301) or goat
anti-mouse IgG (1:1,000; cat. no. zb2305; both OriGene
Technologies, Inc.), and incubated at room temperature for 2 h.
Following treatment with a developing solution (EMD Millipore,
Billerica, MA, USA) the membranes were exposed (ProteinSimple, San
Jose, CA, USA), the expression levels of HO-1 and CXCR3 in the
different groups was calculated using a relative quantitative
analysis method using Alphaview SA software 3.4.0.0
(ProteinSimple). The IEC-6 cells in the different groups and
transplanted small bowel on day 7 post-transplantation were
collected for protein extraction using the same steps as those
described above. The primary antibodies used were as follows:
Anti-GAPDH rabbit IgG (1:2,000; cat. no. 21612; SAB),
anti-proliferating cell nuclear antigen (PCNA) mouse IgG (1:500;
cat. no. EM111201; Abcam), anti-ZO-1 rabbit IgG (1:1,000),
anti-p-mixed lineage kinases 3 (p-MLK3) rabbit IgG (1:800; cat. no.
ab191530; Abcam), anti-MLK3 rabbit IgG (1:500; cat. no. 11996-1-AP;
ProteinTech Group, Inc.), anti-p MAPK kinase 3 (p-MKK3) rabbit IgG
(1:1,000; cat. no. 12280; Abcam), anti-MKK3 rabbit IgG (1:500; cat.
no. 13898-1-AP; ProteinTech Group, Inc.), anti-p-p38 rabbit IgG
(1:800; cat. no. ab47363; Abcam), anti-p38 rabbit IgG (1:500; cat.
no. 9212S; Cell Signaling Technology, Inc., Danvers, MA, USA),
anti-p-CHOP10 rabbit IgG (1:500), anti-CHOP10 rabbit IgG (1:300,
cat. no. 15204-1-AP; ProteinTech Group, Inc.), anti-p-MEF2C rabbit
IgG (1:500; cat. no. bs-5479R; BIOSS) and anti-MEF2C rabbit IgG
(1:500; cat. no. 10056-1-AP; ProteinTech Group, Inc.) respectively.
The secondary antibodies were HRP-conjugated goat anti-rabbit IgG
(1:2,000) and goat anti-mouse IgG (1:2,000), which were diluted
with skimmed milk (BD Biosciences). The abundance of the target
protein was calculated relative to the abundance of the internal
control protein, GAPDH, using Alphaview SA software
(ProteinSimple).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
The different groups of BMMSCs were added to 1 ml
TRIzol to extract the total RNA and determine the RNA concentration
and purity. An RT-PCR kit (Takara Bio, Inc., Osaka, Japan) was used
to reverse transcribe the RNA into cDNA. Following dilution, the
cDNA was treated with an amplification kit (Takara Bio, Inc.) for
PCR, 2 µl cDNA was added to 20 µl of a PCR reaction
system, following the manufacturer's protocol exactly to observe
the SYBR-Green fluorescence signal. The primer sequences of the
target genes are listed in Table
I. The mRNA expression levels of HO-1 and CXCR3 were calculated
as the relative quantity, using GAPDH as the reference. The
relative expression of the target genes in each of the groups was
calculated as the ratio of the relative level of the target gene to
that of the GAPDH gene, using the Cq value as the statistical
parameter (27). The primer
sequences of the target genes are listed in Table I. The IEC-6 cells and the
transplanted small bowel on day 7 post-transplantation were
processed using the same steps as those described above. The
relative mRNA expression levels of PCNA, ZO-1, MLK3, MKK3, P38,
activating transcription factor 2 (ATF2), CHOP10 and MEF2C were
also calculated using the relative quantity method (Table I).
 | Table IPrimer sequences for reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I
Primer sequences for reverse
transcription-quantitative polymerase chain reaction analysis.
| Gene | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| HO-1 |
CTGGCTCTTTTCTTGG |
ATGGTCAGAACATGGAC |
| CXCR3 |
TCATGGCCTACTGCTATGC |
CGACTTGGCCACGTCTAC |
| GAPDH |
CCGTATCGGACGCCTGGTTAC |
GCCGTGGGTAGAGTCATACTGGAAC |
| MLK3 |
TCCCAGACTCAGATCCCTTCTG CC |
AGTGTATGCTATGCCTCCTC |
| MKK3 |
CACCCGTTCTTCACCTTGCAC |
ACTTGGGACAGCTAGTTGCGAG |
| P38 |
CCCAGCAGTCCTATCCACG |
TCTCCCTTTGTTCGGTTTGC |
| ATF2 |
TGAGTTGGCAAGTCCATTCG |
GCTATCCTGGTGAGTTGTTTCTAC |
| CHOP10 |
CTTGCTGAAGAGAACGAG |
CATGTGCACTGGAGATTAC |
| MEF2C |
GGGTCACCGTAGGCATAGAG |
CGACAAAGTCCAGCTTATGC |
| PCNA |
AACTTGGAATCCCAGAACAGG C |
TGTAGGAGACAGTGGAGTGGC |
| ZO-1 |
TGAGCCTTGAACTTTGACCTC |
GAAATCGTGCTGATGTGCC |
Flow cytometry
The BMMSCs in each of the groups were trypsinized
and adjusted to a density of 5×105 cells/100 µl
for further examination. The blank control tube of BMMSCs received
no antibodies, whereas the other tubes received one of the three
following antibody combinations: i) 0.625 µl anti-CD29-PE
rat IgG (cat. no. 102207; BioLegend, Inc., San Diego, CA, USA) with
2.5 µl anti-CD34-FITC rat IgG (cat. no. sc-7324; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) rat IgG; ii) 0.625 µl
anti-CD45-PE rat IgG (cat. no. 202224) with 0.25 µl
anti-CD90-FITC rat IgG (cat. no. 202503); or iii) 0.625 µl
anti-RT1A-PE rat IgG (cat. no. 205208) with 0.25 µl
anti-RT1B-FITC rat IgG (cat. no. 205305; all BioLegend, Inc.). The
BMMSCs were incubated at 4°C for 40 min, resuspended in PBS,
centrifuged (300 × g for 5 min at 4°C), fixed in 200 µl of
2% paraformaldehyde, and then analyzed by flow cytometry C6 (BD
Biosciences). The lymphocytes in each of the groups were collected
and adjusted to a density of 1×106 cells/100 µl.
Each sample received either 1.25 µl anti-CD3-APC rat IgG
(cat. no. 17-0030) with 0.625 µl anti-CD25-PE rat IgG (cat.
no. 12-0390; both eBioscience; Thermo Fisher Scientific, Inc.) or
1.25 µl anti-CD3-APC rat IgG with 0.625 µl
anti-CD71-FITC rat IgG (cat. no. 204405; BioLegend, Inc.), and the
lymphocytes were then incubated at 4°C for 40 min, resuspended in
PBS, centrifuged (500 × g for 5 min at 4°C), fixed in 200 µl
of 2% paraformaldehyde, and then analyzed by flow cytometry C6 (BD
Biosciences).
ELISA
The supernatants from each of the groups of cells
were collected following centrifugation and stored at −80°C.
Interleukin (IL)-2 and IFN-γ kits (R&D Systems, Inc.
Minneapolis, MN, USA) were used to assess the concentrations of
cytokines, in accordance with the manufacturer's protocol.
Animal model
The small bowel transplantation donors were healthy
male Brown Norway rats (aged 6-8 weeks, weighing 180-200 g, n=40),
and the recipients for small bowel transplantation were healthy
male Lewis rats (aged 6-8 weeks, weighing 180-200 g, n=70). All
animals were provided by the Experimental Animal Center of the
Academy of Military Medical Sciences and standard rat food was
provided ad libitum. The experimental animals were kept at
23°C with 50% humidity and a 12 h light/dark cycle for 2 weeks,
with free access to water and food, and regular replacement of
bedding prior to the experiments. All experiments on animals
followed the experimental animal ethical regulations, and were
approved by the Ethics Committee of Tianjin First Central Hospital.
The small bowel transplantation model used was established using
the method described by Yin et al (18). The rats were divided into six
groups: i) NSBT group, sham-operated without small bowel
transplantation; ii) IsoT group, received an isogeneic
transplantation of the small bowel from genetically identical hosts
(Lewis); iii) NS group, injected intravenously with 1 ml sterile
normal saline (NS; 0.9% sodium chloride solution) from the dorsal
penile vein; iv) MSCs group, injected with a single-cell suspension
including 5×106 BMMSCs; v) Ad-HO/MSCs group, injected
with a single-cell suspension including 5×106
Ad-HO-1/MSCs; and vi) Ad-(CXCR3 + HO)/MSCs group, injected with a
single-cell suspension of 5×106 Ad-(CXCR3 + HO-1)/MSCs.
On day 7 post-small bowel transplantation, samples from each of the
groups were acquired and analyzed.
Statistical analysis
SPSS statistical software, version 17.0 (SPSS, Inc.,
Chicago, IL, USA) was used for all statistical analysis. Normally
distributed data are presented as the mean ± standard deviation.
The significance of differences between groups were assessed using
Student's t-test (single comparisons) or one-way analysis of
variance with Least Significant Difference and Student-Newman-Keuls
post hoc comparison. P<0.05 was considered to indicate a
statistically significant difference. GraphPad Prism 5.0 software
(GraphPad Software, Inc., La Jolla, CA, USA) was used to plot data
for presentation.
Results
Verification of BMMSCs transfected with
Ad, HO-1, CXCR3, and CXCR3 + HO-1
In terms of morphological aspects, the third
generation BMMSCs typically exhibited a spindle shape and it was
not possible to differentiate them into adipocytes and osteoblasts.
The positivity of the expression of the extracellular markers CD29,
CD90 and RT1A on BMMSCs was >95% (18). The morphological changes of the
Ad/MSCs, Ad-HO/MSC Ad-CXCR3/MSCs and Ad-(HO + CXCR3)/MSCs were not
marked different compared with those of BMMSCs (untreated control),
and the cells remained spindle-shaped (Fig. 1A).
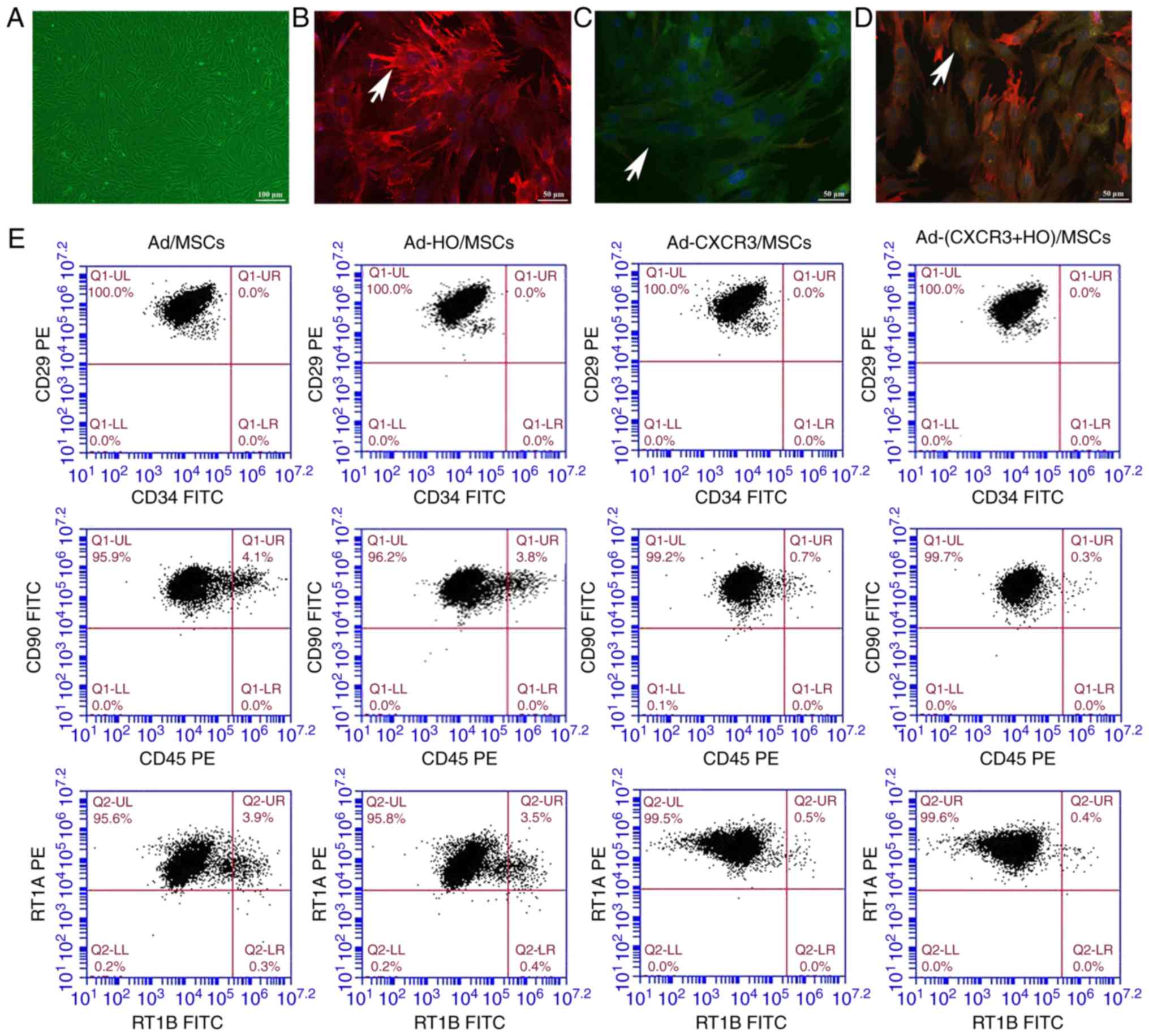 | Figure 1Morphology, phenotype, gene
expression and viability of the different groups of BMMSCs. (A)
Cellular morphology of Ad-MSCs (scale bar, 100 µm). The red
fluorescence indicates HO-1 protein, the green fluorescence
indicates CXCR3 protein and the blue fluorescence indicates the
nucleus. (B) Positive expression of HO-1 protein was high in
Ad-HO/MSCs (indicated by white arrows), (C) Positive expression of
CXCR3 protein was high in Ad-CXCR3/MSCs (indicated by white
arrows); scale bar, 50 µm. (D) Positive expression of both
HO-1 and CXCR3 proteins was high in Ad-(CXCR3 + HO)/MSCs (indicated
by white arrows); scale bar, 50 µm. (E) Changes in cell
surface markers of BMMSCs transfected with different genes. BMMSCs,
bone marrow mesenchymal stem cells; Ad, adenovirus; CXCR3,
CXC-chemokine receptor CXCR3; HO-1, heme oxygenase-1. |
For the identification of gene expression, low
positive expression of the HO-1 and CXCR3 proteins was in observed
in the BMMSCs and Ad/MSCs. The protein expression of HO-1 was high
and that of CXCR3 protein was low in the Ad-HO MSCs (Fig. 1B). The protein expression of CXCR3
was high and that of HO-1 was low in the Ad-CXCR3/MSCs (Fig. 1C). The protein expression of HO-1
and CXCR3 were high in the Ad-(HO + CXCR3)/MSCs (Fig. 1D).
Phenotypically, the positive expression of CD29,
CD90 and RT1A on the BMMSCs exposed to different treatments were
>95%, with no significant difference between the five groups
(Fig. 1E). The negative
expression of CD34, CD45 and RT1B exposed to different treatments
were >95%, with no significant difference between the five
groups (Fig. 1E).
The viability of the BMMSCs transduced with the
different genes was similar to that of the normal BMMSC (Fig. S1A-F). The protein and mRNA
concentrations of HO-1 in the Ad-HO/MSC and Ad-(HO + CXCR3)/MSCs
were significantly higher than those in the BMMSCs, Ad/MSCs and
Ad-CXCR3/MSC (P<0.05; Fig.
S1G). The protein and mRNA concentrations of CXCR3 in the
Ad-CXCR3/MSCs and Ad-(HO + CXCR3)/MSC were significantly higher
than those in the BMMSCs, Ad/MSCs and Ad-HO/MSCs (P<0.05;
Fig. S1H). These results suggest
that the CXCR3, HO-1 and HO-1 + CXCR3 genes were successfully
transfected into BMMSCs, did not affect the morphology of the
BMMSCs and exhibited no toxic effects.
Establishment of the TNF-α/IEC-6 model,
IEC-6 cell viability and BMMSC chemotaxis
The protein and mRNA levels of the tight junction
protein, ZO-1, in the injured intestinal epithelium model
(TNF-α/IEC-6, referred to as the Model group) were significantly
lower than those in the normal IEC-6 cells (P<0.05; Fig. 2A). This suggests that the model of
IEC-6 injury was successfully established, as indicated by the
presence of damaged intestinal epithelial cells.
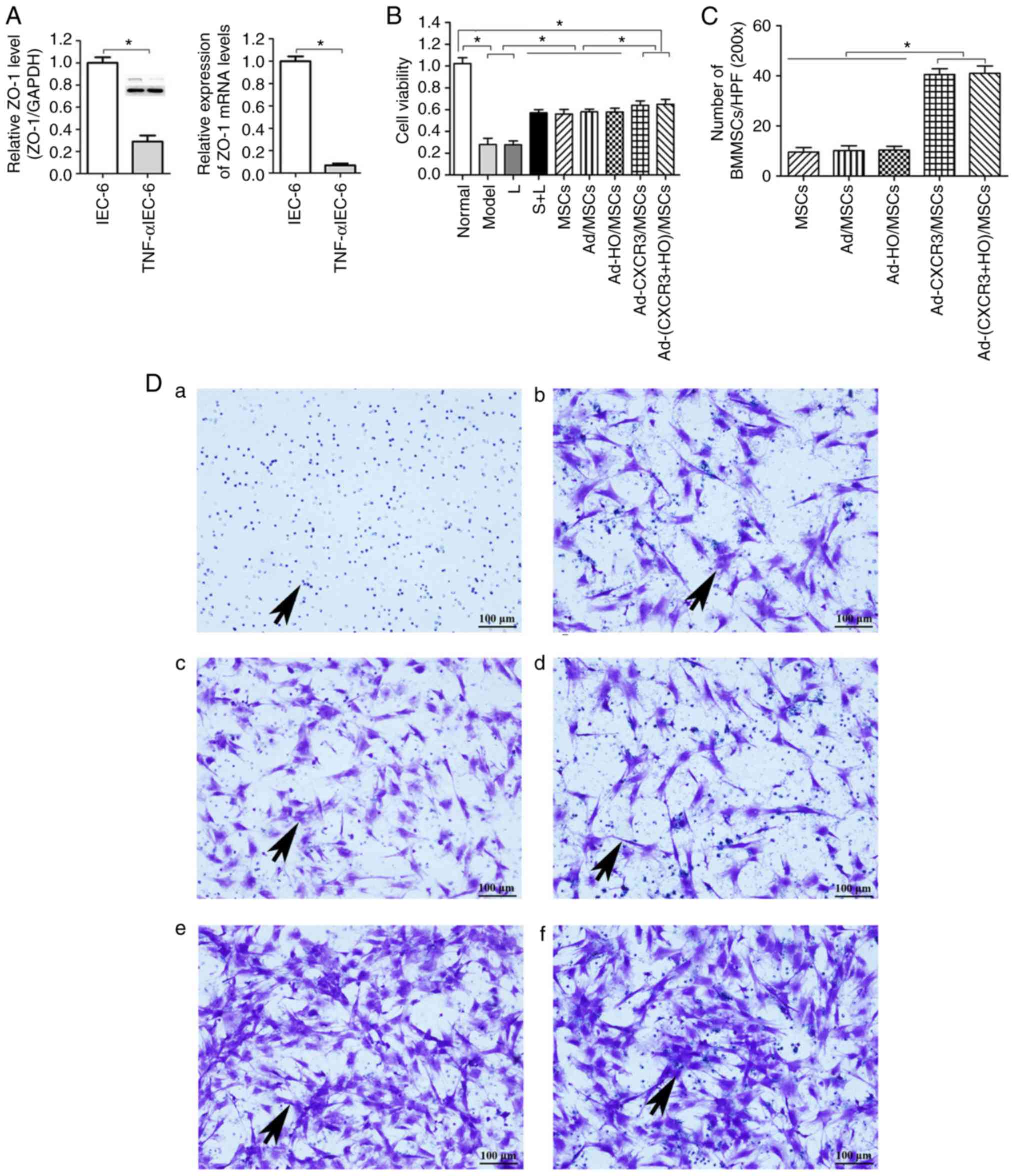 | Figure 2Validation of the TNF-α/IEC-6 model,
IEC-6 cell viability and BMMSC chemotaxis. (A) Protein and mRNA
content of ZO-1 in IEC-6 cells and TNF-α/IEC-6. (B) IEC-6 viability
in each of the groups was detected. (C) BMMSC migration; the number
of migrated BMMSCs in the Ad-CXCR3/MSCs + L and Ad-(CXCR3 +
HO)/MSCs + L groups were all higher than in the other groups. (D)
Migrated BMMSCs across Transwell membranes (indicated by black
arrows): (a) L group, (b) MSC + L group, (c) Ad/MSCs + L group, (d)
Ad-HO/MSCs + L group, (e) Ad-CXCR3/MSCs + L group, and (f)
Ad-(CXCR3 + HO)/MSCs + L (scale bar, 100 µm). (E) A large
number of CXCR3-modified BMMSCs migrated together at 1 h, whereas a
(F) small number of BMMSCs without the CXCR3 gene modified began to
migrate at 4 h (indicated by white arrows; scale bar, 50
µm). *P<0.05. IEC-6, intestinal epithelial
crypt cell line-6; ZO-1, zonula occludens-1; TNF-α; tumor necrosis
factor-α; BMMSCs, bone marrow mesenchymal stem cells; Ad,
adenovirus; CXCR3, CXC-chemokine receptor CXCR3; HO-1, heme
oxygenase-1; L, lymphocytes; S, S203580. |
IEC-6 viability in the Model and L groups was
similar and significantly lower than that in the Normal group
(P<0.05). The viability of IEC-6 in the S + L, MSCs + L, Ad/MSCs
+ L, and Ad-HO/MSCs + L groups were similar, which were lower than
the that in the Normal group (P<0.05) and significantly higher
than that in the Model and L groups (P<0.05). IEC-6 viability in
the Ad-CXCR3/MSCs + L and Ad-(CXCR3 + HO)/MSCs + L groups were
similar (P<0.05), which was lower than that in the Normal group
and significantly higher than that in all other groups (P<0.05;
Fig. 2B). These results suggest
that BMMSCs improved the viability of injured IEC-6 cells. The
effects of the HO-1 gene-modified Ad-HO-1/BMMSCs on IEC-6 cells was
similar to that of the BMMSCs, whereas the effects of the CXCR3
gene-modified Ad-CXCR3/BMMSCs and Ad-(HO-1 + CXCR3)/BMMSCs were
more substantial.
Following co-culture for 24 h, the numbers of
migrated BMMSCs (Fig. 2C) in the
Ad-CXCR3/MSCs + L and Ad-(CXCR3 + HO)/MSCs + L groups were
significantly higher than the numbers in the L, MSCs + L, Ad/MSCs +
L and Ad-HO/MSCs + L groups (Fig.
2Da-f), P<0.05). Only lymphocytes migrated in the L group
and lymphocytes appear as blue dots (Fig. 2Da). There was no significant
difference in the number of migrated BMMSCs between the MSCs + L,
Ad/MSCs + L and Ad-HO/MSCs + L groups, and the BMMSCs had a
polygonal shape (Fig. 2D). A
large number of BMMSCs migrated at 1 h in the Ad-CXCR3/MSCs + L and
Ad-(CXCR3 + HO)/MSCs + L (Fig.
2E) groups, and gradually increased over time. Few BMMSCs had
migrated at 1 h in the Ad/MSCs + L and Ad-HO/MSCs + L groups, and a
small number of BMMSCs began to migrate at 4 h (Fig. 2F). These results indicate that the
CXCR3 gene was capable of increasing the chemotactic ability of
BMMSCs, allowing the rapid migration of a large number of cells
through the Transwell membrane, which were able to reach the
damaged IEC-6 cells where they mediated their effector
function.
Effect of BMMSCs on tight junction
proteins and the apoptosis of damaged IEC-6 cells
The positive expression of ZO-1 (assessed via green
fluorescence in Fig. 3Aa-j) was
significant in the Normal group. The positive expression of ZO-1
decreased significantly in the Model group, and the intercellular
tight junctions were severely damaged. The positive expression of
ZO-1 also decreased significantly in the L group, exhibiting
destruction similar to that in the Model group. The positive
expression of ZO-1 increased significantly in the S + L, MSCs + L,
Ad-MSCs + L and Ad-HO/MSCs + L groups, with similar results
observed in these four groups. The positive expression of ZO-1
increased the most markedly in the Ad-CXCR3/MSCs + L and Ad-(CXCR3
+ HO)/MSCs + L groups (Fig. 3A).
The concentrations of ZO-1 protein in the Model and L groups did
not differ significantly, and both were significantly lower than
that in the Normal group (P<0.05). The protein concentrations of
ZO-1 in the S + L, MSCs + L, Ad/MSCs + L and Ad-HO/MSCs + L groups
were higher than those in the Model and L groups (P<0.05), and
all were lower than that in the Normal group (P<0.05). There
were no statistically significant differences among the S + L, MSCs
+ L, Ad/MSCs + L and Ad-HO/MSCs + L groups. The Ad-CXCR3/MSCs + L
and Ad-(CXCR3 + HO)/MSCs + L groups exhibited a similar protein
concentration of ZO-1, which was lower than that in the Normal
group (P<0.05), and significantly higher than that in all other
groups (P<0.05; Fig.
3Aa-j).
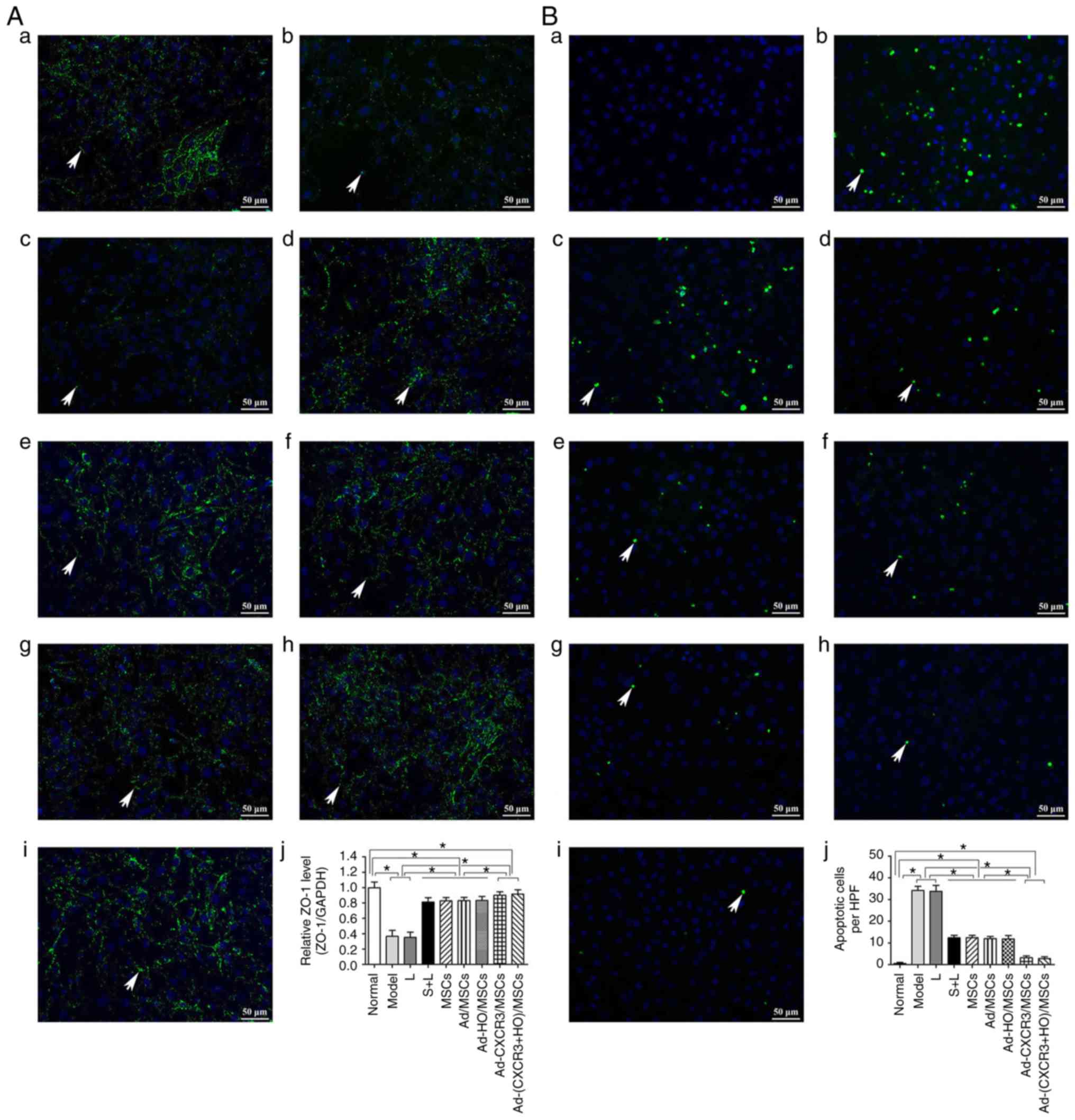 | Figure 3Expression of tight junction protein,
ZO-1, and IEC6 apoptosis. (A) Expression of ZO-1 in each group,
with the cell nucleus shown in blue and tight junction protein,
ZO-1 (indicated by white arrows) shown in green. (a) Normal, (b)
Model, (c) L, (d) S + L, (e) MSCs + L, (f) Ad/MSCs + L, (g)
Ad-HO/MSCs + L, (h) Ad-CXCR3/MSCs + L and (i) Ad-(CXCR3 + HO)/MSCs
+ L groups (scale bar, 50 µm); (j) protein expression of
ZO-1 in each group. (B) Apoptosis of IEC-6 cells in each of groups,
with the cell nucleus shown in blue and apoptotic bodies (indicated
by white arrows) shown in green. (a) Normal, (b) Model, (c) L, (d)
S + L, (e) MSCs + L, (f) Ad/MSCs + L, (g) Ad-HO/MSCs + L, (h)
Ad-CXCR3/MSCs + L, and (i) Ad-(CXCR3 + HO)/MSCs + L groups (scale
bar, 50 µm); (j) apoptosis of IEC-6 cells.
*P<0.05. IEC-6, intestinal epithelial crypt cell
line-6; ZO-1, zonula occludens-1; MSCs, bone marrow mesenchymal
stem cells; Ad, adenovirus; CXCR3, CXC-chemokine receptor CXCR3;
HO-1, heme oxygenase-1; L, lymphocytes; S, S203580. |
No apoptosis of IEC-6 cells was detected in the
Normal group, however, levels of apoptosis in the Model and L
groups were increased significantly, and were higher than apoptosis
in the S + L, MSCs + L, Ad/MSCs + L and Ad-HO/MSCs + L groups
(P<0.05). However, no significant differences were observed
among these four groups. The levels of IEC-6 apoptosis in the
Ad-CXCR3/MSCs + L and Ad-(CXCR3 + HO)/MSCs + L groups did not
differ significantly (P=0.471), but they were significantly lower
than that in all other groups, with the exception of the Normal
group (P<0.05; Fig.
3Ba-j).
These results suggest that the expression of the
intestinal tight junction protein, ZO-1, decreased and apoptosis
increased when the IEC-6 cells were damaged. In addition, the
BMMSCs increased the expression of ZO-1 and decreased apoptosis of
the damaged IEC6 cells. The effects of the Ad-HO/MSCs were similar
to those of the BMMSCs, and the effects of the Ad-CXCR3/MSCs and
Ad-(CXCR3 + HO)/MSCs were more marked than those of the Ad-HO/MSCs
and BMMSCs.
Lymphocyte activity in each treatment
group
The proportions of CD3+ CD25+
and CD3+ CD71+ within the CD3+
lymphocyte population were relatively low in the normal rat
lymphocytes. The proportions of CD3+ CD25+
and CD3+ CD71+ cells in the L group were
significantly higher than those of the other groups (P<0.05).
The proportions of CD3+CD25+ cells and
CD3+CD71+ cells in the Ad-CXCR3/MSCs + L and
Ad-(CXCR3 + HO)/MSCs + L groups were similar to those in the normal
rat lymphocytes (P>0.05), but significantly lower than those in
the MSCs + L, Ad-MSCs + L and Ad-HO/MSCs + L groups (P<0.05;
Fig. 4A-J). Increased
concentrations of IL-2 and IFN-γ indicate enhanced lymphocyte
activity, and vice versa. The concentrations of IL-2 and IFN-γ in
the L group were significantly higher than those in the other
groups (P<0.05). The concentrations of IL-2 and IFN-γ were
decreased and at similar levels in the MSCs + L, Ad-MSCs + L and
Ad-HO/MSCs + L groups, which were higher than those in the Normal
and Model groups (P<0.05). The concentrations of IL-2 and IFN-γ
in the Ad-CXCR3/MSCs + L and Ad-(CXCR3 + HO)/MSCs + L groups were
decreased significantly and were lower than those observed in the
other groups (P<0.05; Fig. 4K and
L).
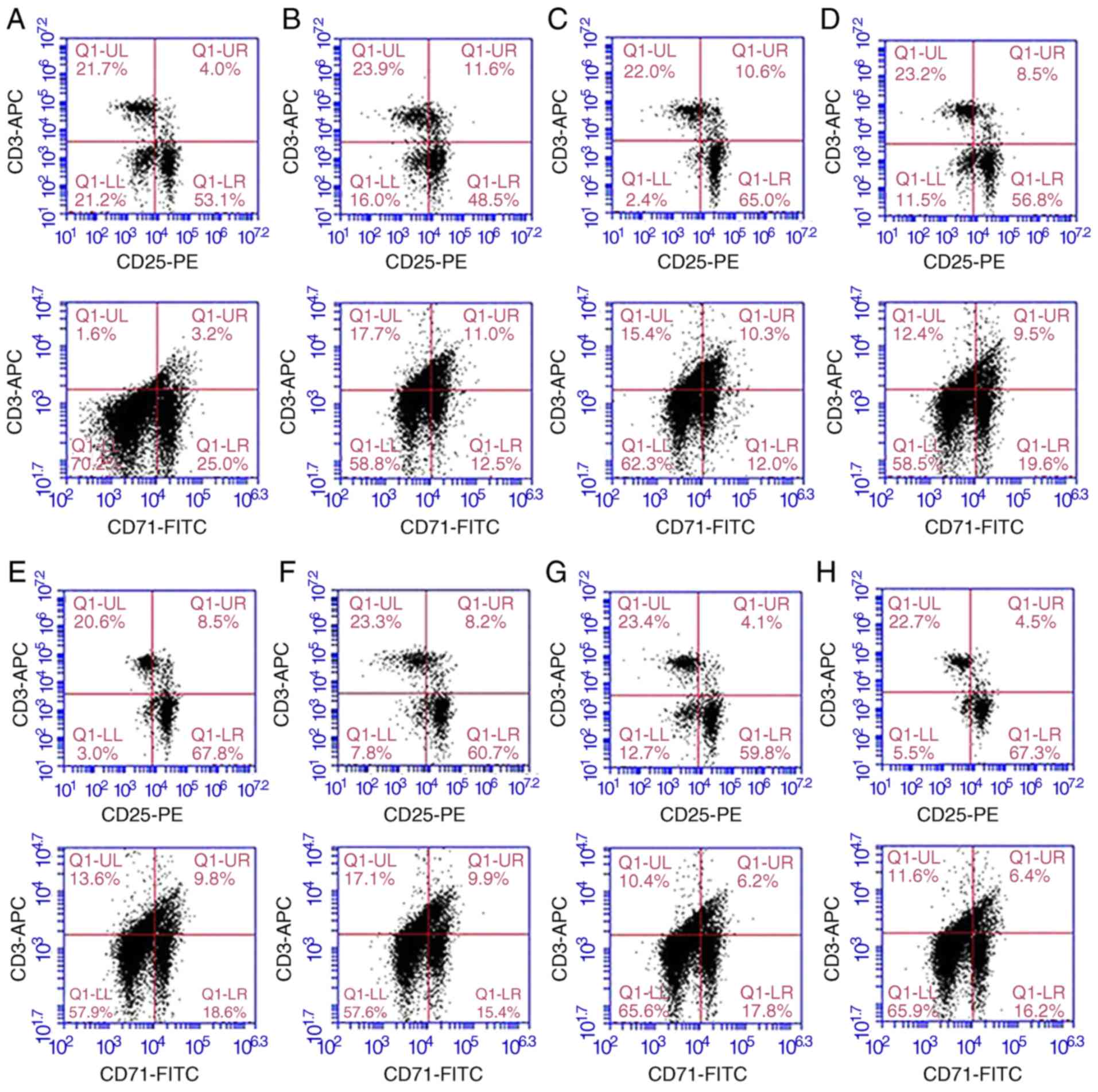 | Figure 4Proportions of
CD3+CD25+ and
CD3+CD71+, and supernatant concentrations of
IL-2 and IFN-γ in the different groups of T lymphocytes.
Proportions of CD3+ CD25+ and CD3+
CD71+ in (A) normal rat lymphocytes, and the (B) L, (C)
S + L, (D) MSCs + L, (E) Ad/MSCs + L, (F) Ad-HO/MSCs + L, (G)
Ad-CXCR3/MSCs + L and (H) Ad-(CXCR3 + HO)/MSCs + L groups. The
proportions of CD3+ CD25+ and CD3+
CD71+ within the CD3+ lymphocyte population
were relatively low in the normal rat lymphocytes (4.0 and 3.2%,
respectively). Expression was high (11.6 and 11.0%) in the L group,
and in the S + L group (10.6% and 10.3%). Expression was
significantly lower in the MSCs + L, Ad/MSCs + L and Ad-HO/MSCs + L
groups compared with that in the L group (P<0.05). (I) The
proportions of CD3+CD25+ cells in the
Ad-CXCR3/MSCs + L and Ad-(CXCR3 + HO)/MSCs + L group were 4.1 and
4.5%, respectively, and those of (J)
CD3+CD71+ cells were 6.2 and 6.4%,
respectively, which were similar to those in normal lymphocytes
(P>0.05, but were significantly lower compared with those in
other groups (P<0.05). Concentrations of (K) IL-2 and (L) IFN-γ
were relatively low in the normal group, significantly higher in
the model group (P<0.05), and similar in the Ad-CXCR3/MSCs + L
and Ad-(CXCR3 + HO)/MSCs + L groups, which were higher than those
in the Normal group (P<0.05), but significantly lower than in
the other groups (P<0.05). *P<0.05. MSCs, bone
marrow mesenchymal stem cells; Ad, adenovirus; CXCR3, CXC-chemokine
receptor CXCR3; HO-1, heme oxygenase-1; L, lymphocytes; S, S203580;
IL-2, interleukin-2; IFN-γ, interferon-γ. |
These results indicate that BMMSCs inhibited the
activation of lymphocytes, and the effects of the Ad-HO/MSCs were
similar to those of BMMSCs. The inhibitory effects of the CXCR3
gene-modified BMMSCs were more marked, which may be attributed to
the ability of Ad-CXCR3/MSCs and Ad-(CXCR3 + HO)/MSCs to compete
with preliminary activated T lymphocytes for CXCR3 ligands. This
results in the inability of T lymphocytes to bind with these
ligands, inhibiting further T lymphocyte activation, and decreasing
the secretion of inflammatory cytokines.
P38-MAPK pathway-related protein and mRNA
changes in the IEC-6 cells of the treatment groups
The present study subsequently analyzed the p38-MAPK
upstream phosphory-lated proteins, p-MLK3, p-MKK3 and p-p38, and
downstream phosphorylated proteins p-ATF2, p-CHOP10 and p-MEF2C in
the MAPK pathway, and PCNA protein of the target cells. The protein
and mRNA expression levels of p-MLK3 and p-MKK3 were high in all
experimental groups, with the exception of the Normal group;
however, there was no difference among these groups (Fig. 5A and B). These results suggest
that the upstream proteins in the p38-MAPK pathway are not involved
in the repair of damaged IEC-6 cells by the genetically modified
BMMSCs.
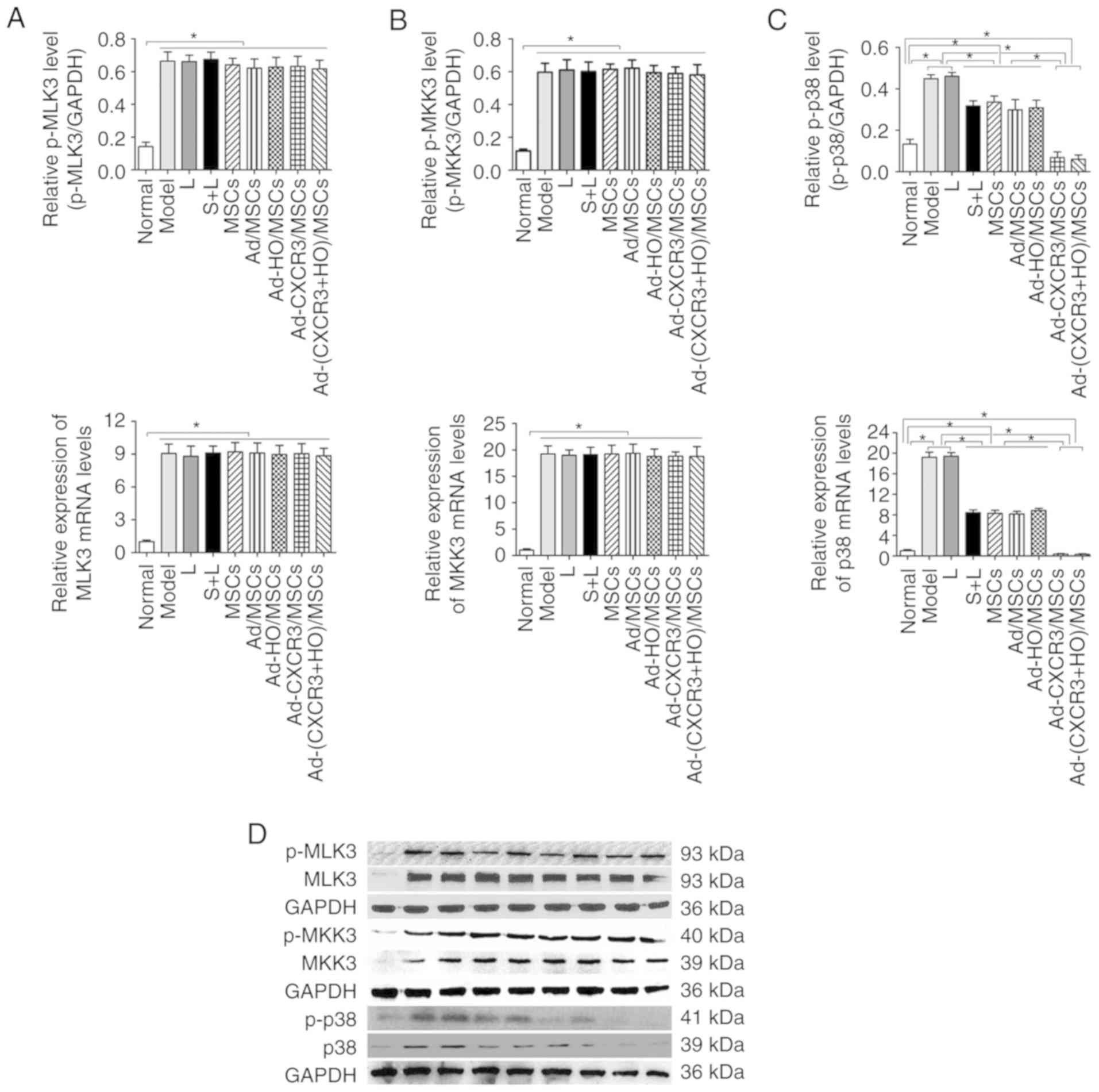 | Figure 5Relative expression levels of p-MLK3,
MLK3, p-MKK3, MKK3, p-p38 and p38 in each of the treatment groups.
The protein and mRNA expression levels of (A) MLK3 and (B) MKK3
were high in all experimental groups, with the exception of the
Normal group, and the levels of p-MKK3 and p-MLK3 were
significantly higher than that in the Normal group (P<0.05). (C)
Protein and mRNA expression levels of p38 were highest in the Model
and L groups, which were significantly higher than in all other
groups (P<0.05). The expression levels of p38 and p-p38 in the
Ad-CXCR3/MSCs + L and Ad-(CXCR3 + HO)/MSCs + L groups were the
lowest, which were significantly lower than those of the other
experimental groups (P<0.05). *P<0.05. (D)
Expression of p-MLK3, MLK3, p-MKK3, MKK3, p-p38 and p38 in each of
the treatment groups as demonstrated by western blotting. MSCs,
bone marrow mesenchymal stem cells; Ad, adenovirus; CXCR3,
CXC-chemokine receptor CXCR3; HO-1, heme oxygenase-1; L,
lymphocytes; S, S203580; MLK3, mixed lineage kinases 3; MKK3,
mitogen-activated protein kinase kinase 3; p-, phosphorylated. |
The protein and mRNA expression levels of p-p38,
p-ATF2, p-CHOP10 and p-MEF2C were high in the Model and L groups,
and these levels were higher than those in all other groups
(P<0.05). Following use of the p38 MAPK inhibitor, S203580 (S +
L group), the expression levels of p-p38 and associated downstream
molecules were significantly decreased to levels lower than those
in the Model and L groups (P<0.05) (Figs. 5C and 6A-E). The protein and mRNA expression
levels of p-p38, p-ATF2, p-CHOP10 and p-MEF2C in the Ad-CXCR3/MSCs
+ L and Ad-(CXCR3 + HO)/MSCs + L groups decreased, and were
significantly lower than all other experimental groups (P<0.05);
however, there was no significant difference between these two
groups. The results of the S + L, MSCs + L, Ad-MSCs + L, and
Ad-HO/MSCs + L groups exhibited no statistically significant
difference (Fig. 5C and 6A-E). The protein, p-CHOP10, was
histochemically stained (Fig.
6F). The positive expression of p-CHOP10 in the Model and L
groups was significantly higher than that in the other groups
(P<0.05). The S + L, MSCs + L, Ad/MSCs + L and Ad-HO/MSCs + L
groups exhibited similar positive expression of p-CHOP10, the level
of which was higher than that in the Ad-CXCR3/MSCs + L and
Ad-(CXCR3 + HO)/MSCs + L groups (P<0.05); the latter two groups
were not significantly different (P=0.242; Fig. 6F). These results indicate that, as
an inhibitor of p38 protein phosphorylation, S203580 inhibits the
activation of the p38-MAPK pathway, thereby significantly
inhibiting the function of downstream protein expression without
significantly altering the expression of the upstream protein in
the pathway. As BMMSCs exhibited effects similar to that of
S203580, it was hypothesized that they exert a reparative effect on
damaged IEC-6 cells through the p38 MAPK pathway. The effects of
the HO-1 gene-modified BMMSCs were similar to those of BMMSCs,
suggesting that HO-1 provided no advantage. In addition, the
CXCR3-modified BMMSCs demonstrated more marked effects the BMMSCs,
indicating that the CXCR3 gene synergistically enhanced the
reparative capacity of the BMMSCs.
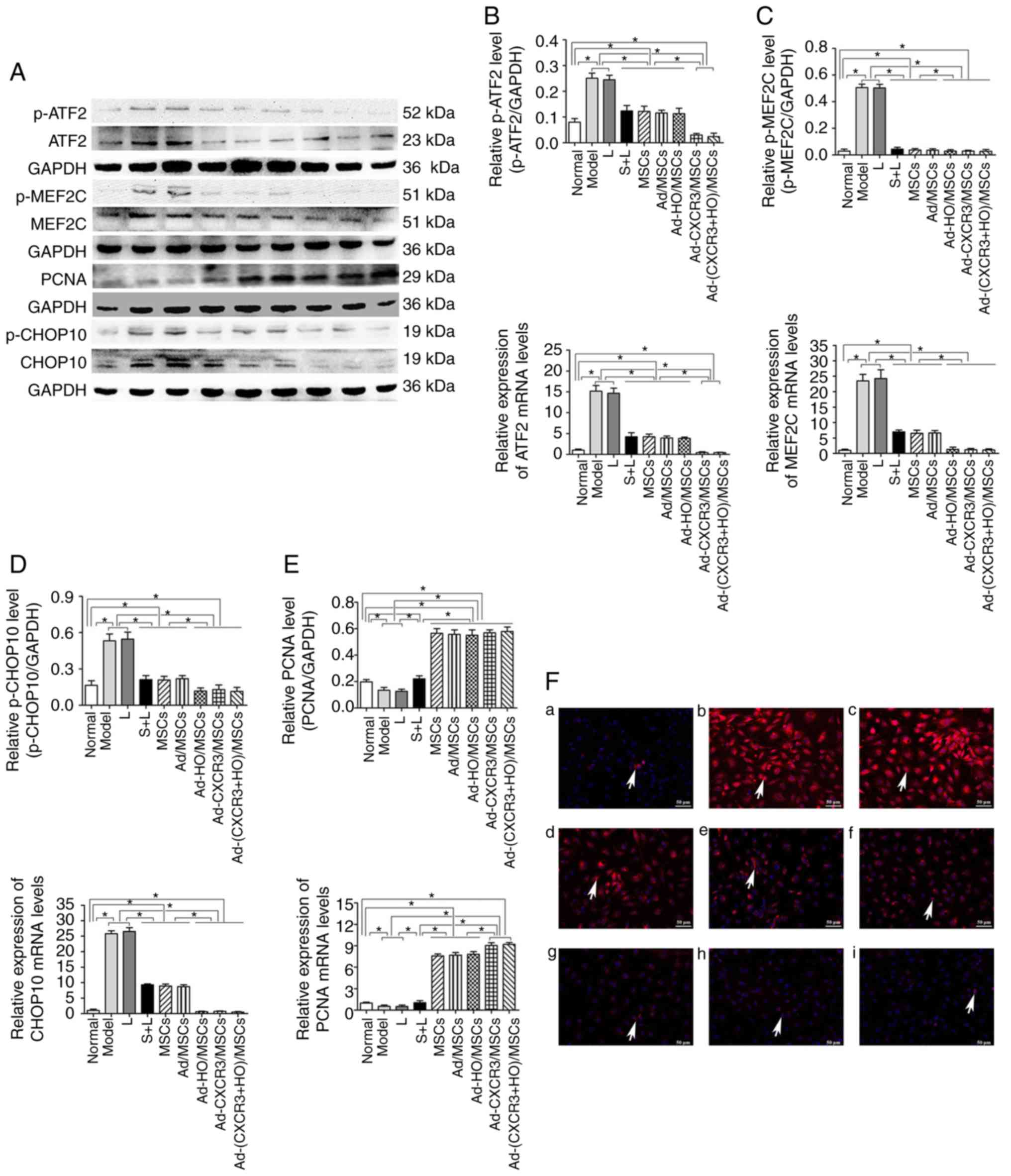 | Figure 6Relative expression of p-ATF2, ATF2,
p-MEF2C, MEF2C, PCNA, p-CHOP10 and CHOP10, and p-CHOP10 positivity
in each treatment group. (A) Expression of p-ATF2, ATF2, p-MEF2C,
MEF2C, PCNA, p-CHOP10 and CHOP10 in each treatment group as
demonstrated by western blotting. Protein and mRNA expression of
(B) ATF2 to p-ATF2, (C) MEF2C to p-MEF2C, (D) PCNA and (E) CHOP10
to p-CHOP10 in each group. (F) p-CHOP10 (indicated by white arrows)
exhibited low positive rate in the (a) Normal group but was
expressed at high levels when the IEC-6 cells were damaged: (b)
Model and (c) L groups. Compared with the (d) S + L, (e) MSCs + L,
and (f) Ad-MSCs + L groups, p-CHOP10-positivity was significant
lower in the (g) Ad-HO/MSCs + L, (h) Ad-CXCR3/MSCs + L and (i)
Ad-(CXCR3 + HO)/MSCs + L groups. Scale bar, 50 µm.
*P<0.05. MSCs, bone marrow mesenchymal stem cells;
Ad, adenovirus; CXCR3, CXC-chemokine receptor CXCR3; HO-1, heme
oxygenase-1; L, lymphocytes; S, S203580; ATF2, activating
transcription factor 2; MEF2C, myocyte enhancer factor type 2C;
PCNA, proliferating cell nuclear antigen; CHOP10, C/EBP homologous
protein-10; p-, phosphorylated. |
The protein and mRNA expression levels of PCNA in
the IEC-6 cells of the Normal group were higher than those in the
Model and L groups (P<0.05); the expression levels in these
latter two groups were lower than in all the other experimental
groups (P<0.05). The protein and mRNA expression levels of PCNA
in the IEC-6 cells of the MSCs + L, Ad/MSCs + L and Ad-HO/MSCs + L
groups were similar, which were higher than that in the S + L
group. The Ad-CXCR3/MSCs + L and Ad-(CXCR3 + HO)/MSCs + L groups
exhibited similar results, exhibiting higher expression than in the
other groups (P<0.05; Fig.
6E). These results suggest that, as a downstream target protein
in the p38-MAPK pathway, the expression of PCNA was influenced by
p38-MAPK signaling. The effects of the HO-1 gene-modified BMMSCs
were similar to those of BMMSCs, indicating that HO-1 provided no
advantage. In addition, the CXCR3-modified BMMSCs exhibited more
marked effects than the BMMSCs.
Expression of p38-MAPK-related proteins
in a rejection model of small bowel transplantation
The protective effects of the Ad-(CXCR3 + HO)/MSCs
and Ad-HO/MSCs groups on the damaged IEC-6 cells were the most
marked on post-operative day 7 in the rejection model of small
bowel transplantation. Therefore, p38-MAPK-related proteins were
detected in the transplanted intestinal tissues of rats on day 7
post-transplantation. The results showed that the upstream proteins
p-MLK3, p-MKK3 and p-p38, and downstream proteins p-ATF2, p-CHOP10
and p-MEF2C of the p38-MAPK pathway, and the target protein, PCNA,
exhibited low expression in the Normal and IsoT groups, which were
significantly different from all other groups (P<0.05; Fig. 7A-I). p-MLK3 and p-MKK3 were
expressed at high levels in the NS, MSCs, Ad-HO/MSCs, and Ad-(CXCR3
+ HO)/MSCs groups (Fig. 7A and
B). The expression of p-p38 was significantly different among
the groups as follows: Expression in the Ad-(CXCR3 + HO)/MSCs group
was significantly lower than that in all other experimental groups
(P<0.05); expression in the Ad-HO/MSCs group was lower than that
in the MSCs and NS (P<0.05) groups; and expression in the MSCs
group was lower than that in the NS group (P<0.05; Fig. 7C). The expression levels of
p-ATF2, p-CHOP10 and p-MEF2C in the Ad-(CXCR3 + HO)/MSCs were
significantly lower than those in the Ad-HO/MSCs (P<0.05),
whereas levels in the Ad-HO/MSCs were lower than those in the MSCs
(P<0.05), and levels in the NS group were the highest (Fig. 7D-F). The expression of PCNA was
significantly higher in the Ad-(CXCR3 + HO)/MSCs group compared
with that in the other experimental groups (P<0.05). The
expression of PCNA in the Ad-HO/MSCs group was higher than that in
the MSCs and NS groups (P<0.05). The MSCs group exhibited higher
expression of PCNA than the NS group (P<0.05) (Fig. 7G). The expression of ZO-1 was
significantly higher in the NBST and IsoT groups compared with that
in the other experimental groups. In the Ad-(CXCR3 + HO)/MSCs group
was higher than that in the Ad-HO/MSCs, MSCs and NS groups
(P<0.05). In the Ad-HO/MSCs was higher than that in the MSCs and
NS groups (P<0.05). The expression of ZO-1 in the NS groups was
the lowest. (Fig. 7H). The
results of the above proteins detected by western blot are
summarized in Fig. 7I. The
positive expression of p-CHOP10 in the Ad-(CXCR3 + HO)/MSCs group
was significantly lower than that in the other groups (Fig. 7J). These results suggest that
BMMSCs may repair the damaged intestinal epithelial cells via the
p38-MAPK pathway. The HO-1 gene-modified BMMSCs showed more marked
protective effects than the BMMSCs; however, the effects of BMMSCs
co-modified with the CXCR3 and HO-1 genes were the most marked.
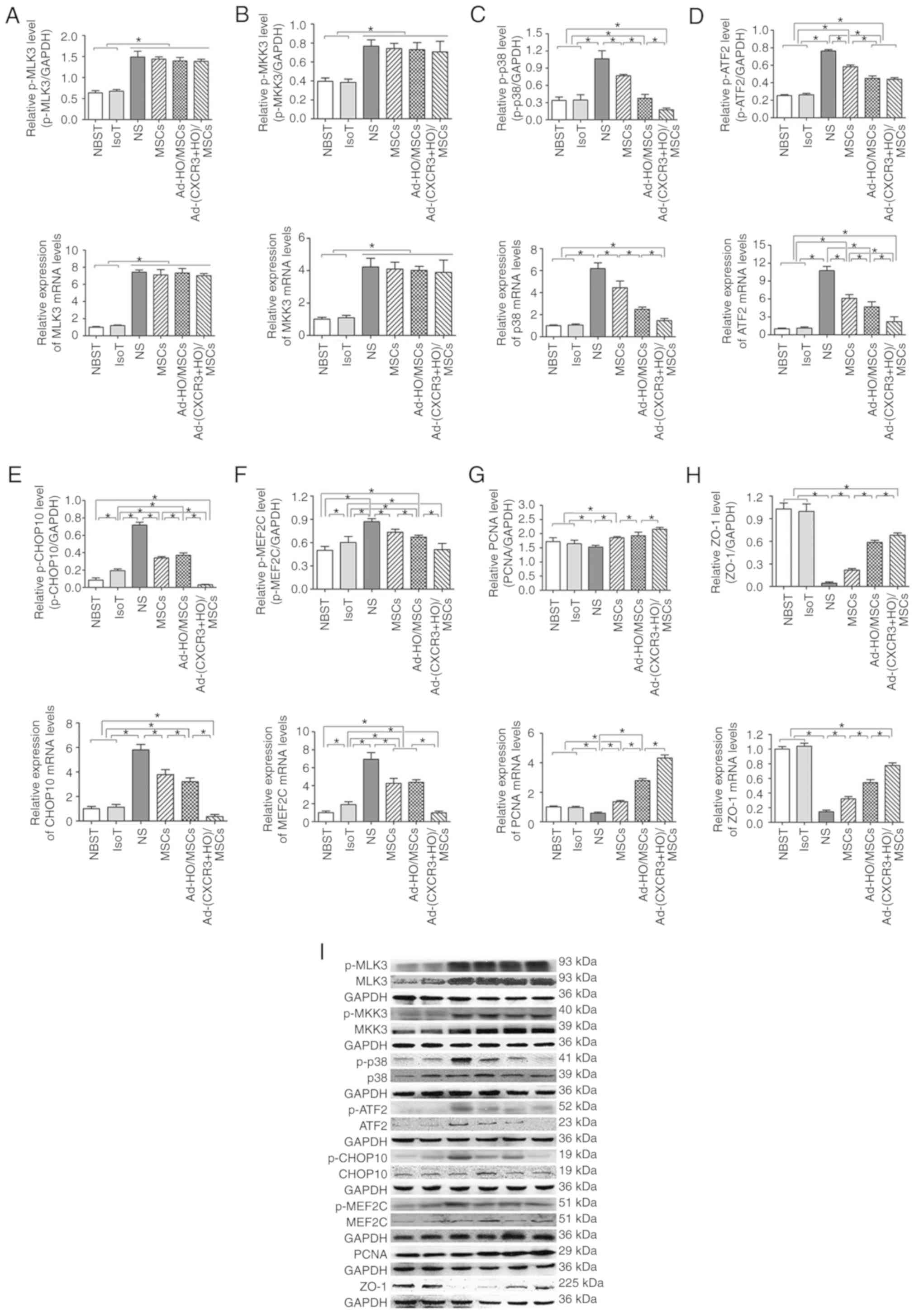 | Figure 7p38-MAPK pathway-related protein and
mRNA expression, and immunohistochemical expression of p-CHOP10 on
day 7 post-small bowel transplantation. Protein and mRNA expression
of (A) MLK3 to p-MLK3, (B) MKK3 to p-MKK3, (C) p38 to p-p38, (D)
ATF2 to p-ATF2, (E) CHOP10 to p-CHOP10, (F) MEF2C to p-MEF2C, (G)
PCNA and (H) ZO-1 in each group. (I) Expression of related protein
of the p38-MAPK pathway as demonstrated by western blotting.
*P<0.05.. (J) Immunohistochemical expression of
p-CHOP10 on day 7 post-small bowel transplantation: (a) NBST, (b)
IsoT, (c) NS, (d) MSCs, (e) Ad-HO/MSCs and (f) Ad-(CXCR3 + HO)/MSCs
groups. Expression was significantly lower in the Normal group than
in the other groups, with the exception of the Ad-(CXCR3 + HO)/MSCs
group (P<0.05); expression in the IsoT group was lower than that
in the NS, MSCs and Ad-HO/MSCs groups (P<0.05). Expression in
the Ad-HO/MSCs group was lower than that in the MSCs and NS groups
(P<0.05). Expression in the NS group was the highest, and
expression in the Ad-(CXCR3 + HO)/MSCs group was significantly
lower than that in all experimental groups. MSCs, bone marrow
mesenchymal stem cells; Ad, adenovirus; CXCR3, CXC-chemokine
receptor CXCR3; HO-1, heme oxygenase-1; L, lymphocytes; S, S203580;
IsoT, isogeneic transplantation of the small bowel from genetically
identical host; NS, injected intravenously with normal saline (0.9%
sodium chloride solution) CHOP10, CHOP10, C/EBP homologous
protein-10; MLK3, mixed lineage kinases 3; MKK3, mitogen-activated
protein kinase kinase 3; ATF2, activating transcription factor 2;
MEF2C, myocyte enhancer factor type 2C; PCNA, proliferating cell
nuclear antigen; p-, phosphorylated. |
Discussion
Stem cells are pluripotent cells with the ability to
self-replicate. BMMSCs are one of the most representative types of
stem cells. BMMSCs have the properties of proliferation,
differentiation, and immune-modulation in vitro, and can
promote tissue repair in damaged tissues (28,29). BMMSCs can also secrete soluble
cytokines through paracrine mechanisms to reduce inflammatory
reactions and enhance reparative effects (30,31). BMMSCs exert inhibitory effects on
the rejection response following small bowel transplantation, and
reduce intestinal damage (9).
HO-1 gene modification can enhance the ability of BMMSCs to
tolerate hypoxia-reoxygenation injury, and enhance BMMSC viability
and proliferation (4,5,13,14). Chemokine receptor CXCR3 is
typically expressed in the parenchyma cells of diseased organs and
activated inflammatory cells (15). Upon binding specifically to their
ligands, CXCR3 receptors induce the chemotaxis of lymphocytes to
damaged sites (16,17,32), and further activate lymphocytes to
produce a pro-inflammatory response through the regulation of the T
helper cell signaling pathway (33). Due to the specific binding of the
receptor to its ligand, BMMSCs that overexpress the CXCR3 gene are
able to compete with such activated lymphocytes for the CXCR3
ligand. This allows for a large number of activated lymphocytes to
remain unbound to the CXCR3 receptor, thereby inhibiting the
positive feedback mechanism required for a continued effector
response. Additional lymphocytes cannot be recruited to the damaged
sites, leading to the reduced secretion of inflammatory cytokines
and therefore, reduced cellular injury. Our previous study
demonstrated that the efficiency and survival duration of BMMSCs
were significantly improved following modification by the HO-1 and
CXCR3 genes in animal models of small bowel transplantation
(18). However, the mechanism by
which BMMSCs repair damaged intestinal cells remains to be
elucidated. Therefore, the present study used an ex vivo
model of damaged intestinal epithelial cells to model in
vivo responses.
The experimental model used standard BMMSCs as
previously described (34,35).
Following transfection with the HO-1 gene and/or CXCR3 gene, the
BMMSCs maintained their functionality, and the viability assessment
confirmed that the HO-1 gene and CXCR3 gene did not have any toxic
effects. In the rejection model of small bowel transplantation,
TNF-α increased significantly (9), inducing damage to the intestinal
epithelial cells (36); thus, the
present study used undifferentiated IEC-6 cells to simulate the
in vivo intestinal mucosal environment (24). The results demonstrated that the
expression of the tight junction protein (ZO-1) in IEC-6 cells
treated with TNF-α was significantly decreased and the number of
apoptotic cells was significantly increased. This finding indicates
that the establishment of an in vitro model of damaged
intestinal epithelial cells using TNF-α and IEC-6 cells had been
successful. The BMMSCs increased the expression of ZO-1 and the
proliferation of IEC-6 cells, and decreased IEC-6 apoptosis.
Further investigation revealed that the protective effects of
Ad-(CXCR3 + HO)/MSCs on damaged intestinal epithelial cells was
more marked than that of the Ad-HO/MSCs and BMMSCs, which was
consistent with the conclusions of the in vivo model
(18).
The chemokine receptor, CXCR3, is expressed on the
surface of activated lymphocytes in vivo (15). Therefore, when the CXCR3 receptor
specifically binds with its ligand, CXCR3-expressing cells can be
recruited into the site of injury (16,17). CXCR3-overexpressing BMMSCs compete
with activated T lymphocytes for the CXCR3 ligand, inhibiting
further activation of T lymphocytes and significantly reducing the
inflammatory response (33). CD25
and CD71 are characteristic surface markers of lymphocyte
activation, exhibit low expression during steady-state conditions
and become expressed at high levels following activation (37-39). IL-2 (40) and IFN-γ (41) are sensitive cytokines that can
reflect the activity of T cells. The results of the present study
showed that the expression levels of
CD3+CD25+ and CD3+CD71+
were significantly decreased and those of IL-2 and IFN-γ were
significantly reduced following repair by Ad-CXCR3/MSCs or
Ad-(CXCR3 + HO)/MSCs. This finding confirmed that CXCR3
gene-modified BMMSCs are important in the inhibition of lymphocyte
activation. In addition, CXCR3 gene-modified BMMSCs demonstrated
the highest level of mobility and chemotaxis, which suggested that
an increased number of BMMSCs arriving to the damaged site and
decreased activity of T lymphocytes was the major cause of the
enhanced reparative effects. Therefore, the specific mechanism by
which this occurs warrants further investigation.
Studies have shown that the p38-MAPK pathway serves
an important role in the sequence of events inflammatory responses
caused by trauma (19), infection
(20) and ischemia-reperfusion
(21). MLK3 and MKK3 are the main
upstream molecules of the p38-MAPK pathway, and can phosphorylate
the p38-MAPK protein following activation. The phosphorylation of
p38-MAPK influences the activation of the downstream molecules,
ATF2, CHOP10 and MEF2C, further affecting the proliferation,
differentiation and cytokine synthesis of cells (42,43). In the present study, the
expression of p-p38 MAPK protein in damaged IEC-6 cells was
significantly increased, suggesting that this signaling pathway was
activated. However, the expression of the upstream molecules p-MLK3
and p-MKK3 in the p38-MAPK pathway of the different groups was not
statistically significant, indicating that the BMMSCs did not have
an effect on the upstream molecules in the p38-MAPK pathway. When
treated with a p38 inhibitor or BMMSCs, the expression of p-p38
MAPK significantly decreased, indicating that both the inhibitor
and BMMSCs inhibited the phosphorylation of p38 MAPK. The
downstream molecules p-ATF2, p-CHOP10 and p-MEF2C in the p38-MAPK
pathway were significantly decreased in all groups treated with
BMMSCs, and the decreases in the Ad-CXCR3/MSCs and Ad-(CXCR3 +
HO)/MSCs treatment groups were the most marked. These results
suggest that p38-MAPK and the downstream molecules of the p38-MAPK
pathway are involved in the repair of damaged intestinal epithelial
cells by BMMSCs.
ATF2 (44,45), CHOP-10 (46-48) and MEF2C (49,50) have physiological roles in the
control of cellular growth, differentiation and apoptosis (44-50). Their target molecules include
PCNA, apoptotic bodies and ZO-1 protein. The present study
demonstrated that activation of the p38-MAPK signaling pathway
reduced the expression of ZO-1 and PCNA, and significantly
increased apoptosis. When signaling was inhibited by inhibitors or
BMMSCs, the expression levels of ZO-1 and PCNA were increased, and
apoptosis was decreased. This effect was the most marked in the
Ad-CXCR3/MSCs and Ad-(CXCR3 + HO)/MSCs groups. These results
indicate that the downstream constituents of the p38-MAPK pathway
are involved in the repair of damaged intestinal epithelial cells
by CXCR3 and HO-1 gene-modified BMMSCs in vitro.
In addition, it was found that the effects of
Ad-CXCR3/MSCs were more marked than those of simple MSCs on POD 7
following small bowel transplantation in the preliminary experiment
(18), suggesting that Ad-CXCR3
was associated with significant chemotactic effects. The median
survival rate and pathological changes of the rats in the
Ad-CXCR3/MSCs group and the MSCs group were similar on day 7
following small bowel transplantation (Figs. S2 and S3). As the present study focused on
elucidating the role of the double gene-modified BMMSCs, six groups
were assessed, not including the Ad-CXCR3/MSCs group. p-38
MAPK-related molecules were assessed on day 7 following small bowel
transplantation in different treatment groups of an in vivo
rejection model (NSBT, IsoT, NS, MSCs, Ad-HO-1/MSC, and Ad-(CXCR3 +
HO)/MSCs). Compared with the BMMSCs, Ad-(CXCR3 + HO)/MSCs
significantly decreased the expression of p-38-MAPK, and the
downstream molecules p-ATF2, p-CHOP-10 and p-MEF2C of the p-38-MAPK
pathway. the expression of target molecules (e.g., decreased
apoptosis) were also reduced (18), whereas the expression levels of
PCNA and ZO-1 were increased. The expression of upstream molecules
did not differ between the various treatment groups, which verified
the results of the in vitro experiments.
The effects of Ad-HO/MSCs in an ex vivo model
were similar to that of BMMSCs, whereas the effects of Ad-HO/MSCs
were more marked than BMMSCs in the rat model of small bowel
transplantation, which may be associated with the action time of
the different stem cells. As lymphocytes were added in the
experimental groups, their survival rate declined after 24 h.
Therefore, the investigation was limited to 24 h in vitro,
when the stem cells remained highly active; however, the HO-1 gene
may not have been fully effective. Using a rat model of small bowel
transplantation, the results were analyzed 7 days following small
bowel transplantation, when the activity of BMMSCs was
significantly reduced and that of Ad-HO/MSCs remained high. The
CXCR3 gene enhanced the chemotactic ability of the BMMSCs, enabling
a higher number of stem cells to reach damaged sites more rapidly
to exert reparative effects. The gene expression of CXCR3 only
exhibited a chemotactic effect in the absence of lymphocyte
involvement. However, in the presence of lymphocytes, CXCR3-gene
modified BMMSCs were able to compete with lymphocytes for the CXCR3
ligands. This inhibited lymphocyte activation, reduced the
secretion of inflammatory cytokines and further decreased the
extent of cellular damage. The process of stem cell reparative
effects is complex, which may involve the combined action of
multiple pathways. However, only the p38-MAPK pathway was involved
in transplantation repair in the present study. Therefore, as the
effects of stem cells have not been comprehensively examined,
future investigations aim to examine whether Wnt/β-catenin and
other pathways are also involved in regeneration and
reparation.
In conclusion, CXCR3 and HO-1 double gene-modified
BMMSCs and CXCR3 gene-modified BMMSCs demonstrated significant
reparative effects on damaged intestinal epithelial cells in
vitro. The gene expression of CXCR3 enhanced the chemotactic
ability of BMMSCs, which induced the early and rapid recruitment of
BMMSCs overexpressing this gene to the damaged site. It was also
demonstrated that BMMSCs exert this effect via p38-MAPK pathway.
Taken together, these findings provide an experimental basis for
the dual gene therapy of stem cells.
Supplementary Materials
Abbreviations:
|
CXCR3
|
CXC- chemokine receptor CXCR3
|
|
BMMSCs
|
bone marrow mesenchymal stem cells
|
|
HO-1
|
heme oxygenase-1
|
|
IEC- 6
|
intestinal epithelial crypt cell
line-6
|
|
TNF-α
|
tumor necrosis factor-α
|
|
GFP
|
green fluorescence protein
|
|
p38-MAPK
|
p38 mitogen activated protein
kinase
|
|
p-C HOP10
|
anti-phosphorylated-C /EBP homologous
protein-10
|
|
p-MLK3
|
phosphorylated‑mixed lineage kinases
3
|
|
p-MKK3
|
phosphorylated-mitogen-activated
protein kinase kinase 3
|
|
p-MEF2C
|
phosphorylated-myocyte enhancer
factor type 2C
|
|
IFN-γ
|
interferon-γ
|
|
IL-2
|
interleukin-2
|
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81670574, 81441022
and 81270528); the Tianjin Clinical Research Center for Organ
Transplantation Project (15ZXLCSY00070) and the Natural Science
Foundation of Tianjin, China (grant nos. 08JCYBJC08400,
11JCZDJC27800 and 12JCZDJC25200).
Availability of data and materials
All data generated or analyzed during this study
are included in this published article.
Authors' contributions
MY and ZS performed the research, analyzed the
data, and wrote and revised the manuscript; MY, WZ and LY performed
the research; WZ participated in analyzing the data; HS designed
the research, participated in revision of the manuscript. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
All experiments involving animals followed the
experimental animal ethical regulations, and were approved by the
Ethics Committee of Tianjin First Central Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Ruiz P, Kato T and Tzakis A: Current
status of transplantation of the small intestine. Transplantation.
83:1–6. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Koo J, Dawson DW, Dry S, French SW, Naini
BV and Wang HL: Allograft biopsy findings in patients with small
bowel transplantation. Clin Transplant. 30:1433–1439. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Libby P and Pober JS: Chronic rejection.
Immunity. 14:387–397. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang Y, Song HL, Zhang W, Wu BJ, Fu NN,
Dong C and Shen ZY: Hemeoxygenase-1-transduced bone marrow
mesenchymal stem cells in reducing acute rejection and improving
small bowel transplantation outcomes in rats. Stem Cell Res Ther.
7:1642016. View Article : Google Scholar
|
|
5
|
Wu B, Song HL, Yang Y, Yin ML, Zhang BY,
Cao Y, Dong C and Shen ZY: Improvement of liver transplantation
outcome by heme oxygenase-1-transduced bone marrow mesenchymal stem
cells in rats. Stem Cells Int. 2016:92350732016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schu S, Nosov M, O'Flynn L, Shaw G, Treacy
O, Barry F, Murphy M, O'Brien T and Ritter T: Immunogenicity of
allogeneic mesenchymal stem cells. J Cell Mol Med. 16:2094–2103.
2012. View Article : Google Scholar
|
|
7
|
De Miguel MP, Fuentes-Julian S,
Blazquez-Martinez A, Pascual CY, Aller MA, Arias J and
Arnalich-Montiel F: Immunosuppressive properties of mesenchymal
stem cells: Advances and applications. Curr Mol Med. 12:574–591.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Barry FP and Murphy JM: Mesenchymal stem
cells: Clinical applications and biological characterization. Int J
Biochem Cell Biol. 36:568–584. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang Y, Song HL, Zhang W, Wu BJ, Fu NN,
Zheng WP, Dong C and Shen ZY: Reduction of acute rejection by bone
marrow mesenchymal stem cells during rat small bowel
transplantation. Plos One. 9:e1145282014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hodgkinson CP, Gomez JA, Mirotsou M and
Dzau VJ: Genetic engineering of mesenchymal stem cells and its
application in human disease therapy. Hum Gene Ther. 21:1513–1526.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Freyman T, Polin G, Osman H, Crary J, Lu
M, Cheng L, Palasis M and Wilensky RL: A quantitative, randomized
study evaluating three methods of mesenchymal stem cell delivery
following myocardial infarction. Eur Heart J. 27:1114–1122. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yamashita K, Ollinger R, McDaid J,
Sakahama H, Wang H, Tyagi S, Csizmadia E, Smith NR, Soares MP and
Bach FH: Heme oxygenase-1 is essential for and promotes tolerance
to transplanted organs. FASEB J. 20:776–778. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vanella L, Kim DH, Asprinio D, Peterson
SJ, Barbagallo I, Vanella A, Goldstein D, Ikehara S, Kappas A and
Abraham NG: HO-1 expression increases mesenchymal stem cell-derived
osteoblasts but decreases adipocyte lineage. Bone. 46:236–243.
2010. View Article : Google Scholar
|
|
14
|
Zeng B, Lin G, Ren X, Zhang Y and Chen H:
Overexpression of HO-1 on mesenchymal stem cells promotes
angiogenesis and improves myocardial function in infarcted
myocardium. J Biomed Sci. 17:802010. View Article : Google Scholar
|
|
15
|
Jenh CH, Cox MA, Cui L, Reich EP, Sullivan
L, Chen SC, Kinsley D, Qian S, Kim SH, Rosenblum S, et al: A
selective and potent CXCR3 antagonist SCH 546738 attenuates the
development of autoimmune diseases and delays graft rejection. BMC
Immunol. 13:22012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Agostini C, Calabrese F, Rea F, Facco M,
Tosoni A, Loy M, Binotto G, Valente M, Trentin L and Semenzato G:
CXCR3 and its ligand CXCL10 are expressed by inflammatory cells
infiltrating lung allografts and mediate chemotaxis of T cells at
sites of rejection. Am J Pathol. 158:1703–1711. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hancock WW, Wang L, Ye Q, Han R and Lee I:
Chemokines and their receptors as markers of allograft rejection
and targets for immunosuppression. Curr Opin Immunol. 15:479–486.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yin ML, Song HL, Yang Y, Zheng WP, Liu T
and Shen ZY: Effect of CXCR3/HO-1 genes modified bone marrow
mesenchymal stem cells on small bowel transplant rejection. World J
Gastroenterol. 23:4016–4038. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chu W, Li M, Li F, Hu R, Chen Z, Lin J and
Feng H: Immediate splenectomy down-regulates the MAPK-NF-κB
signaling pathway in rat brain after severe traumatic brain injury.
J Trauma Acute Care Surg. 74:1446–1453. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu H, Wang G, Li S, Zhang M, Li H and Wang
K: TNF-α- mediated-p38-dependent signaling pathway contributes to
myocyte apoptosis in rats subjected to surgical trauma. Cell
Physiol Biochem. 35:1454–1466. 2015. View Article : Google Scholar
|
|
21
|
Dai J, Gu L, Su Y, Wang Q, Zhao Y, Chen X,
Deng H, Li W, Wang G and Li K: Inhibition of curcumin on influenza
A virus infection and influenzal pneumonia via oxidative stress,
TLR2/4, p38/JNK MAPK and NF-κB pathways. Int Immunopharmacol.
54:177–187. 2017. View Article : Google Scholar
|
|
22
|
Khan SI, Malhotra RK, Rani N, Sahu AK,
Tomar A, Garg S, Nag TC, Ray R, Ojha S, Arya DS and Bhatia J:
Febuxostat modulates MAPK/NF-κBp65/TNF-α signaling in cardiac
ischemia-reperfusion injury. Oxid Med Cell Longev.
2017:80958252017. View Article : Google Scholar
|
|
23
|
Cuadrado A and Nebreda AR: Mechanisms and
functions of p38 MAPK signalling. Biochem J. 429:403–417. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gao JH, Guo LJ, Huang ZY, Rao JN and Tang
CW: Roles of cellular polyamines in mucosal healing in the
gastrointestinal tract. J Physiol Pharmacol. 64:681–693. 2013.
|
|
25
|
Liu T, Fu NN, Song HL, Wang YL, Wu BJ and
Shen ZY: Suppression of microRNA-203 improves survival of rat bone
marrow mesenchymal stem cells through enhancing PI3K-induced
cellular activation. IUBMB Life. 66:220–227. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cao Y, Wu BJ, Zheng WP, Yin ML, Liu T and
Song HL: Effect of heme oxygenase-1 transduced bone marrow
mesenchymal stem cells on damaged intestinal epithelial cells in
vitro. Cell Biol Int. 41:726–738. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
28
|
Ho MS, Mei SH and Stewart DJ: The
immunomodulatory and therapeutic effects of mesenchymal stromal
cells for acute lung Injury and sepsis. J Cell Physiol.
230:2606–2617. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen J, Li C and Chen L: The role of
microvesicles derived from mesenchymal stem cells in lung diseases.
Biomed Res Int. 2015:9858142015.PubMed/NCBI
|
|
30
|
English K: Mechanisms of mesenchymal
stromal cell immunomodulation. Immunol Cell Biol. 91:19–26. 2013.
View Article : Google Scholar
|
|
31
|
Le Blanc K and Mougiakakos D: Multipotent
mesenchymal stromal cells and the innate immune system. Nat Rev
Immunol. 12:383–396. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Singh AK, Arya RK, Trivedi AK, Sanyal S,
Baral R, Dormond O, Briscoe DM and Datta D: Chemokine receptor
trio: CXCR3, CXCR4 and CXCR7 crosstalk via CXCL11 and CXCL12.
Cytokine Growth Factor Rev. 24:41–49. 2013. View Article : Google Scholar
|
|
33
|
Billottet C, Quemener C and Bikfalvi A:
CXCR3, a double-edged sword in tumor progression and angiogenesis.
Biochim Biophys Acta. 1836:287–295. 2013.PubMed/NCBI
|
|
34
|
Morikawa S, Mabuchi Y, Kubota Y, Nagai Y,
Niibe K, Hiratsu E, Suzuki S, Miyauchi-Hara C, Nagoshi N, Sunabori
T, et al: Prospective identification, isolation, and systemic
transplantation of multipotent mesenchymal stem cells in murine
bone marrow. J Exp Med. 206:2483–2496. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Song K, Huang M, Shi Q, Du T and Cao Y:
Cultivation and identification of rat bone marrow-derived
mesenchymal stem cells. Mol Med Rep. 10:755–760. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zheng XK, Liu CX, Zhai YY, Li LL, Wang XL
and Feng WS: Protection effect of amentoflavone in selaginella
tamariscina against TNF-alpha-induced vascular injury of
endothelial cells. Yao Xue Xue Bao. 48:1503–1509. 2013.In Chinese.
PubMed/NCBI
|
|
37
|
Le Blanc K, Rasmusson I, Götherström C,
Seidel C, Sundberg B, Sundin M, Rosendahl K, Tammik C and Ringdén
O: Mesenchymal stem cells inhibit the expression of CD25
(interleukin-2 receptor) and CD38 on phytohaemagglutinin-activated
lymphocytes. Scand J Immunol. 60:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shipkova M and Wieland E: Surface markers
of lymphocyte activation and markers of cell proliferation. Clin
Chim Acta. 413:1338–1349. 2012. View Article : Google Scholar
|
|
39
|
Daniels TR, Delgado T, Rodriguez JA,
Helguera G and Penichet ML: The transferrin receptor part I:
Biology and targeting with cytotoxic antibodies for the treatment
of cancer. Clin Immunol. 121:144–158. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Elhaik Goldman S, Dotan S, Talias A, Lilo
A, Azriel S, Malka I, Portnoi M, Ohayon A, Kafka D, Ellis R, et al:
Streptococcus pneumoniae fructose-1,6-bisphosphate aldolase, a
protein vaccine candidate, elicits Th1/Th2/Th17-type cytokine
responses in mice. Int J Mol Med. 37:1127–1138. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xiang RL, Mei M, Su YC, Li L, Wang JY and
Wu LL: Visfatin protects rat pancreatic β-cells against
IFN-γ-Induced apoptosis through AMPK and ERK1/2 signaling pathways.
Biomed Environ Sci. 28:169–177. 2015.PubMed/NCBI
|
|
42
|
Duarte S, Shen XD, Fondevila C, Busuttil
RW and Coito AJ: Fibronectin-α4β1 interactions in hepatic cold
ischemia and reperfusion injury: Regulation of MMP-9 and MT1-MMP
via the p38 MAPK pathway. Am J Transplant. 12:2689–2699. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Peng S, Hang N, Liu W, Guo W, Jiang C,
Yang X, Xu Q and Sun Y: Andrographolide sulfonate ameliorates
lipopolysaccharide-induced acute lung injury in mice by
down-regulating MAPK and NF-κB pathways. Acta Pharm Sin B.
6:205–211. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lopez-Bergami P, Lau E and Ronai Z:
Emerging roles of ATF2 and the dynamic AP1 network in cancer. Nat
Rev Cancer. 10:65–76. 2010. View Article : Google Scholar :
|
|
45
|
Lau E and Ronai ZA: ATF2 - at the
crossroad of nuclear and cytosolic functions. J Cell Sci.
125:2815–2824. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Todd DJ, Lee AH and Glimcher LH: The
endoplasmic reticulum stress response in immunity and autoimmunity.
Nat Rev Immunol. 8:663–674. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li Y, Guo Y, Tang J, Jiang J and Chen Z:
New insights into the roles of CHOP-induced apoptosis in ER stress.
Acta Biochim Biophys Sin (Shanghai). 47:146–147. 2015. View Article : Google Scholar
|
|
48
|
He J, Wang C, Sun Y, Lu B, Cui J, Dong N,
Zhang M, Liu Y and Yu B: Exendin-4 protects bone marrow-derived
mesenchymal stem cells against oxygen/glucose and serum
deprivation-induced apoptosis through the activation of the
cAMP/PKA signaling pathway and the attenuation of ER stress. Int J
Mol Med. 37:889–900. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shi GX, Han J and Andres DA: Rin GTPase
couples nerve growth factor signaling to p38 and b-Raf/ERK pathways
to promote neuronal differentiation. J Biol Chem. 280:37599–37609.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wu J, Kubota J, Hirayama J, Nagai Y,
Nishina S, Yokoi T, Asaoka Y, Seo J, Shimizu N, Kajiho H, et al:
P38 mitogen-activated protein kinase controls a switch between
cardiomyocyte and neuronal commitment of murine embryonic stem
cells by activating myocyte enhancer factor 2C-dependent bone
morphogenetic protein 2 transcription. Stem Cells Dev.
19:1723–1734. 2010. View Article : Google Scholar : PubMed/NCBI
|














