Introduction
Epilepsy is a common disease. According to the World
Health Organization, the average prevalence rate of epilepsy
globally is 4-10%, with a rate as high as 6-10% in developing
countries (1,2). At present, the world has witnessed
~50 million patients with epilepsy, 90% of these in developing
countries (3). In China, there
are ~9 million patients with epilepsy; additionally, there are
~400,000-700,000 novel cases. Typically, ~30% of patients with
epilepsy have developed into refractory epilepsy as a result of
lack of effective prevention and treatment measures (4). Various antiepileptic drug therapies
are available at present; however, satisfactory curative effect can
hardly be achieved. Apart from genetic and environmental factors,
the dysregulation of gene expression may result in expression
alterations in genes associated with the pathogenesis of tolerant
epilepsy (5). Such alterations
stimulate mossy fiber sprouting, synaptic reorganization, neuron
necrosis and regeneration, which may contribute to the abnormal
excitatory loop and ultimately result in repeated attack of
refractory epilepsy (6).
A recent study have demonstrated that microRNA
(miRNA) may affect the protein translation at the
post-transcriptional level (7).
miRNA is a class of noncoding and endogenous single-stranded small
molecule RNA ~22nt in length (8).
Primary miRNA (pri-miRNA) is first produced under the function of
RNA polymerase from genome DNA (9), which is subsequently digested into
the precursor miRNA with hairpin-like structure by RNA enzyme III
family in the cell nucleus. The precursor is later transferred to
the cytoplasm by exportin-5 and cut into double-strand miRNA by
Dicer combined enzyme agent. The double-strand miRNA is unlocked by
a helicase, which subsequently forms the mature single-stranded
miRNA. The mature miRNA may subsequently selectively bind with
RNA-induced silencing complex (RISC) to form the RISC complex. This
complex then exhibits base complementarity pairing association with
the 3′ terminal untranslated region (UTR) of the target mRNA.
Therefore, it may directly hydrolyze or restrain the target gene
mRNA to regulate its expression (10). Computer prediction analyses and
experimental studies to date have proved that a single miRNA is
able to regulate multiple target mRNAs (9). Additionally, one-half of mRNAs in
mammalian cells are regulated by one or more miRNAs. For these
reasons, miRNA-associated transcription regulation has attracted
wide scientific attention in the field of life sciences (8).
Acetylation and deacetylation are important
processes in gene expression regulation and post-transcriptional
protein modification (11).
Silent mating-type information regulation 2 (Sir2) is the first
discovered NAD-dependent histone deacetylase. It may prolong the
replicative life span of saccharomycetes (12). Silent information regulator 1
(sirtuin 1 or SIRT1) is the mammalian histone deacetylase
homologous to Sir2 (12). It is
highly conserved in developmental stages and extensively expressed
in all organs. SIRT1 can catalyze histones and a variety of
non-histone proteins. Thereby, it may regulate gene expression and
protein activity. It is a key enzyme during all physiological
processes and is involved in a variety of biological effects. These
biological effects include chromatin remodeling, transcription
suppression, inflammatory response, energy metabolism, cell
survival and apoptosis. A previous study has demonstrated that
miR-183, miR-135a, miR-125b, miR-30c and miR-128 were upregulated
at the seizure-associated phases and in patients with temporal lobe
epilepsy (13). The present study
aimed to evaluate the association between anti-miR-128 expression
and antiepileptic treatment.
Materials and methods
Seizure induction
Adult male Sprague-Dawley rats (6-7 weeks; 180-230g)
were obtained from the Laboratory Animal Center of Taishan Medical
University (Tai'an, China) and kept in a controlled standard
environment (22±2°C; 50-60% humidity) with food and water available
ad libitum and a 12-h light/dark cycle. The rats were
randomly divided into two groups: Control group (n=6), and epilepsy
model group (n=8). All rats were anesthetized using 35 mg/kg
pentobarbital sodium. Epilepsy model rats were injected
subcutaneously with kainic acid (5 mg/kg), and 10 min later they
received a second injection of kainic acid (2.5 mg/kg) (14). The control group rats were
subcutaneously injected with normal saline. After the induction of
epilepsy at 3 days, rats were sacrificed using decollation under 35
mg/kg pentobarbital sodium. The present study was approved by the
Institutional Animal Care and Ethics Committee of Liaocheng
People's Hospital (Liaocheng, China).
Cell culture and transfections
Rat pheochromacytoma (PC12) cells were obtained from
The Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China) and cultured in Dulbecco's modified Eagle's
medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing 10% fetal bovine serum (Gibco; Thermo Fisher Scientific,
Inc.), in 5% CO2 and 95% air. Anti-miR-128 mimics
(5′-AGAGACCGGUUCACGGUGAUU-3′), miR-128 mimics
(5′-UCACAGUGAACCGGUCUCUUU-3′) and negative control mimics
(5′-UUCUCCGAACGUGUCACGUTT-3′) were acquired from Sangon Biotech
Co., Ltd. (Shanghai, China). PC12 cells (1×106 cells)
were transfected with 100 ng miR-128 mimics, 100 ng anti-miR-128
mimics or 100 ng negative control mimics using
Lipofectamine® 2000 reagent (Thermo Fisher Scientific,
Inc.). After 4 h of transfection, the medium was changed and the
cells were used for subsequent experimentation.
Cell proliferation assay and lactate
dehydrogenase (LDH) activity levels
MTT reagent was added into cells and cultivated for
4 h at 37°C. Dimethyl sulfoxide was subsequently added and
cultivated for another 20 min at 37°C. Absorbance was measured at
492 nm. LDH activity levels were measured using an LDH activity kit
(cat. no. C0016; Beyotime Institute of Biotechnology, Haimen,
China) and absorbance was measured at 450 nm.
Apoptosis rate
Cell was washed with PBS and centrifuged at 2,000 ×
g for 10 min at 4°C. Cells were fixed with 4% paraformaldehyde for
15 min and stained with Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (cat. no. 556570; BD Biosciences, San Jose,
CA, USA) for 10 min in the dark at room temperature. The cell
apoptosis rate was measured using a flow cytometer (BD Biosciences)
and analyzed using FlowJo 7.6.1 (FlowJo, LLC, Ashland, OR,
USA).
Caspase-3 activity assay
Hippocampi and cells were obtained and washed with
PBS, lysed with radioimmunoprecipitation buffer (Beyotime Institute
of Biotechnology) and protein concentrations were determined using
a bicinchoninic acid protein (BCA) protein assay kit (Beyotime
Institute of Biotechnology). A total of 5 µg protein from
each sample was incubated with Caspase 3 and 9 activity kit (cat.
nos. C1116 and C1158; Beyotime Institute of Biotechnology) reagents
for 2 h at 37°C, according to the manufacturer's protocol.
Absorbance was measured at 405 nm.
Reverse transcription-quantitative
polymerase chain reaction analysis
Total RNA was isolated from brain tissues using
RNAiso Plus (Takara Biotechnology Co., Ltd., Dalian, China),
according to the manufacturer's protocol. Total RNA (1 µg)
was synthesized into cDNA using PrimeScript RT Reagent kit with
gDNA eraser (Takara Biotechnology Co., Ltd.) at 42°C for 30 min and
82°C for 10 sec. A CFX96 Real-Time System (Bio-Rad Laboratories,
Inc., Hercules, CA, USA) was used to run the qPCR reactions with
SYBR Premix Ex Taq II (Takara Biotechnology Co., Ltd.). The primers
were: miR-128, forward 5′-GGCTCACAGTGAACCGG-3′ and reverse
5′-GTGCAGGGTCCGAGGT-3′; and U6, forward 5′-CTCGCTTCGGCAGCACA-3′ and
reverse 5′-AACGCTTCACGAATTTGCGT-3′. The thermocycling conditions
were as follows: 94°C for 45 sec, followed by 40 cycles of 95°C for
30 sec and 60°C for 30 sec, and a final step at 72°C for 30 sec.
miR-128 expression was quantified using the 2−ΔΔCq
method (15).
Dual luciferase assay
293 cells were purchased from Shanghai Cell Bank of
Chinese Academy of Sciences and cultured in Dulbecco's modified
Eagle's medium (Gibco; Thermo Fisher Scientific, Inc.) containing
10% fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.), in
5% CO2 and 95% air. The pGL3-PLK2 (Invitrogen; Thermo
Fisher Scientific, Inc.)-wild-type 3′-UTR and the mutant 3′-UTR of
SIRT1 were obtained from Shanghai GeneChem Co., Ltd. (Shanghai,
China). The wild-type SIRT1-3′-UTR-pGL3-PLK2 (Shanghai GeneChem
Co., Ltd.), the mutant SIRT1-3′-UTR-pGL3-PLK2 (Shanghai GeneChem
Co., Ltd.) and miR-128 mimics were co-transfected into 293 cells
(1×106 cells/ml) using Lipofectamine® 2000
reagent (Thermo Fisher Scientific, Inc.). After transfection for 24
h, luciferase intensity was measured using a dual-luciferase assay
kit (Promega Corporation, Madison, WI, USA) by a GloMax 20/20
luminometer (Promega Corporation). The absolute values of firefly
luminescence were normalized to those of Renilla luciferase
activity.
Hematoxylin and eosin (H&E)
staining
The brain tissues were harvested, fixed in 4%
paraformaldehyde for 1-2 days at room temperature, processed and
embedded into paraffin blocks. The brain tissue sections (10
µm) were dipped into gradient ethanol (75-100%) and stained
with H&E for 10 min at room temperature. The stained sections
were observed with an upright light microscope (magnification, ×50;
E600FN; Nikon Corporation, Tokyo, Japan).
Western blot analysis
Total proteins were extracted from brain tissues
using a whole protein extraction kit (Nanjing KeyGen Biotech Co.,
Ltd., Nanjing, China). Total protein was quantified using a BCA
assay kit (Beyotime Institute of Biotechnology). Total protein (50
µg) was separated on 10-12% SDS-PAGE and electrophoretically
transferred to polyvinylidene fluoride membranes. Membranes were
blocked in 5% bovine serum albumin (BSA; Beyotime Institute of
Biotechnology) in TBS containing 0.1% Tween-20 (TBST) at room
temperature for 2 h. The membranes were subsequently incubated in
TBST at 4°C overnight with the following primary antibodies (all
from Santa Cruz Biotechnology, Inc., Dallas, TX, USA): SIRT1 (cat.
no. sc-135791; 1:1,000), tumor protein p53 (cat. no. sc-47698;
1:1,000), BCL2 associated X (Bax; cat. no. sc-6236; 1:1,000),
Cytochrome c (cat. no. sc-13561; 1:1,000) and GAPDH (cat.
no. sc-32233; 1:2,000). Following three washes with TBST, the
membranes were incubated with horseradish peroxidase-conjugated
secondary antibody (cat. no. sc-2004; 1:5,000; Santa Cruz
Biotechnology, Inc.) at room temperature for 2 h. Finally, protein
signals were visualized using an Enhanced Chemiluminescence kit
(Pierce; Thermo Fisher Scientific, Inc.) and analyzed using
Image_Lab_3.0 (Bio-Rad Laboratories, Inc.).
Immunofluorescence analysis
Cells were washed with PBS and fixed with 4%
paraformaldehyde for 15 min at room temperature. They were
subsequently blocked with 5% BSA and 0.1% Tris-X100 for 1 h at room
temperature. Cells were incubated at 4°C overnight with a primary
antibody targeting SIRT1 (cat. no. sc-135791; 1:100; Santa Cruz
Biotechnology, Inc.), followed by a secondary goat anti-rabbit
immunoglobulin G-CFL 555 antibody (cat. no. sc-362272; 1:100; Santa
Cruz Biotechnology, Inc.) for 1 h at room temperature. The nuclei
were counterstained with DAPI for 30 min in the dark at room
temperature. Finally, the samples were washed with PBS and observed
by fluorescence microscopy (magnification, ×200).
Statistical analysis
All data are expressed as mean ± standard deviation
(n=3) using SPSS 20.0 (IBM, Corp., Armonk, NY, USA). The
statistical significance of the differences was evaluated by
unpaired t-test (two-tailed) or one-way analysis of variance with
Tukey's multiple comparison tests. P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression of miR-128 in rats with
epilepsy
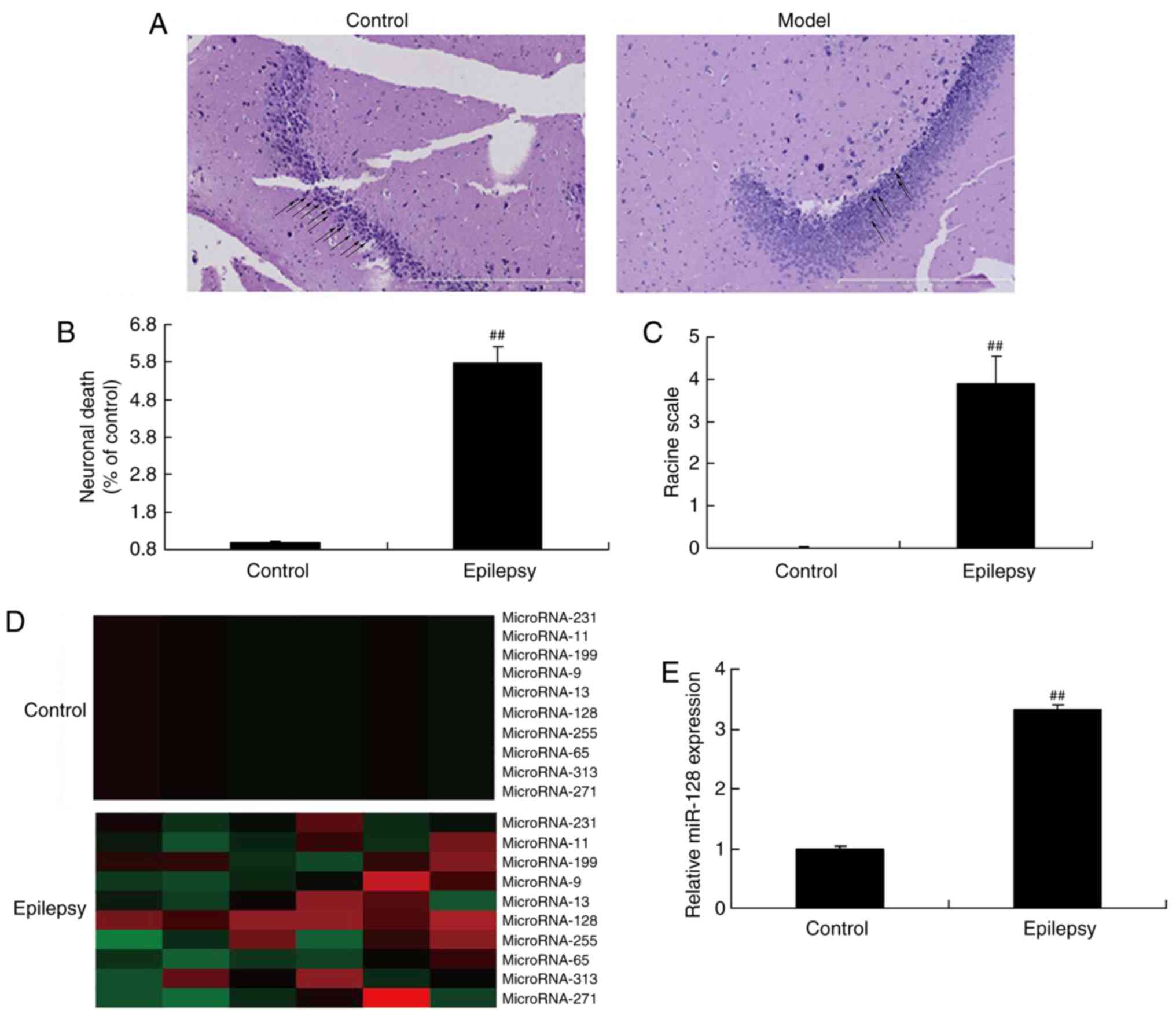
Firstly, the difference in miR-128 expression
between the brain tissues of the epilepsy model rats and normal
control rats was examined. The results indicated that the number of
nerve corpuscles in epilepsy rat model was decreased compared with
the control group (normal rats; Fig.
1A), similar to results from a previous study (16). Neuronal death and racine scale
were increased in the epilepsy rat model, compared with the control
group (normal rats; Fig. 1B and
C), which demonstrated that epilepsy was successfully
established in the rats. qPCR was performed in the present study to
detect the miR-128 expression levels. In addition, miR-128
expression levels in the brains of the epilepsy rats were
significantly higher compared with the normal control rats
(Fig. 1D).
Effect of miR-128 on apoptosis in
epilepsy
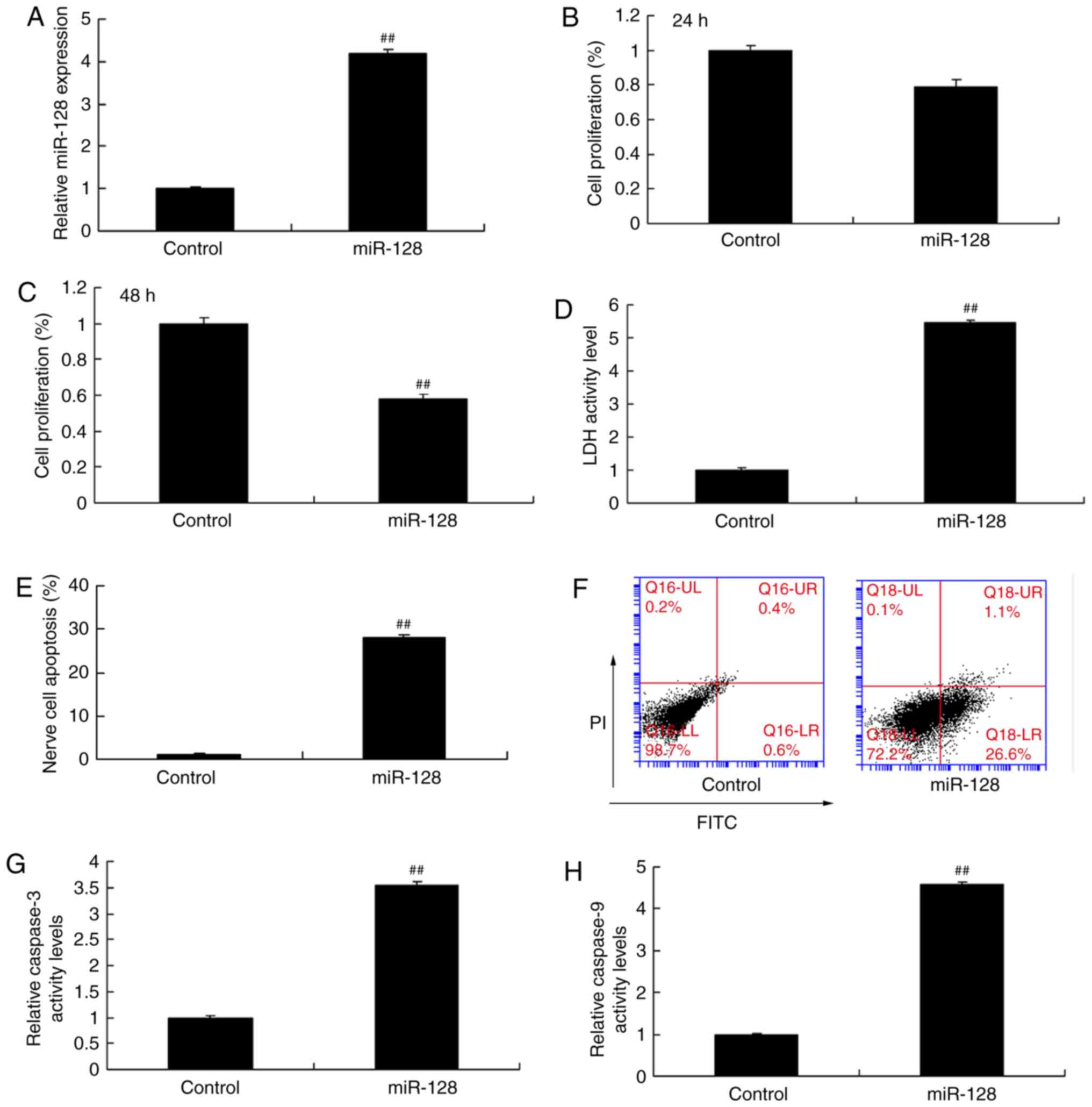
The effect of miR-128 overexpression was further
examined in vitro. Transfection of miR-128 mimics was used
to upregulate the expression of miR-128 in PC12 cells (Fig. 2A). The results from an MTT assay
demonstrated that overexpression of miR-128 significantly inhibited
cell proliferation at 48 h. Additionally, miR-128 overexpression
increased LDH activity (Fig. 2D),
promoted nerve cell apoptosis (Fig.
2E and F), and enhanced caspase-3/9 activity (Fig. 2G and H) compared with the control
group. Thus, the present results suggested that the function of
miR-128 might be associated with nerve cell apoptosis.
Effect of anti-miR-128 on apoptosis in
epilepsy
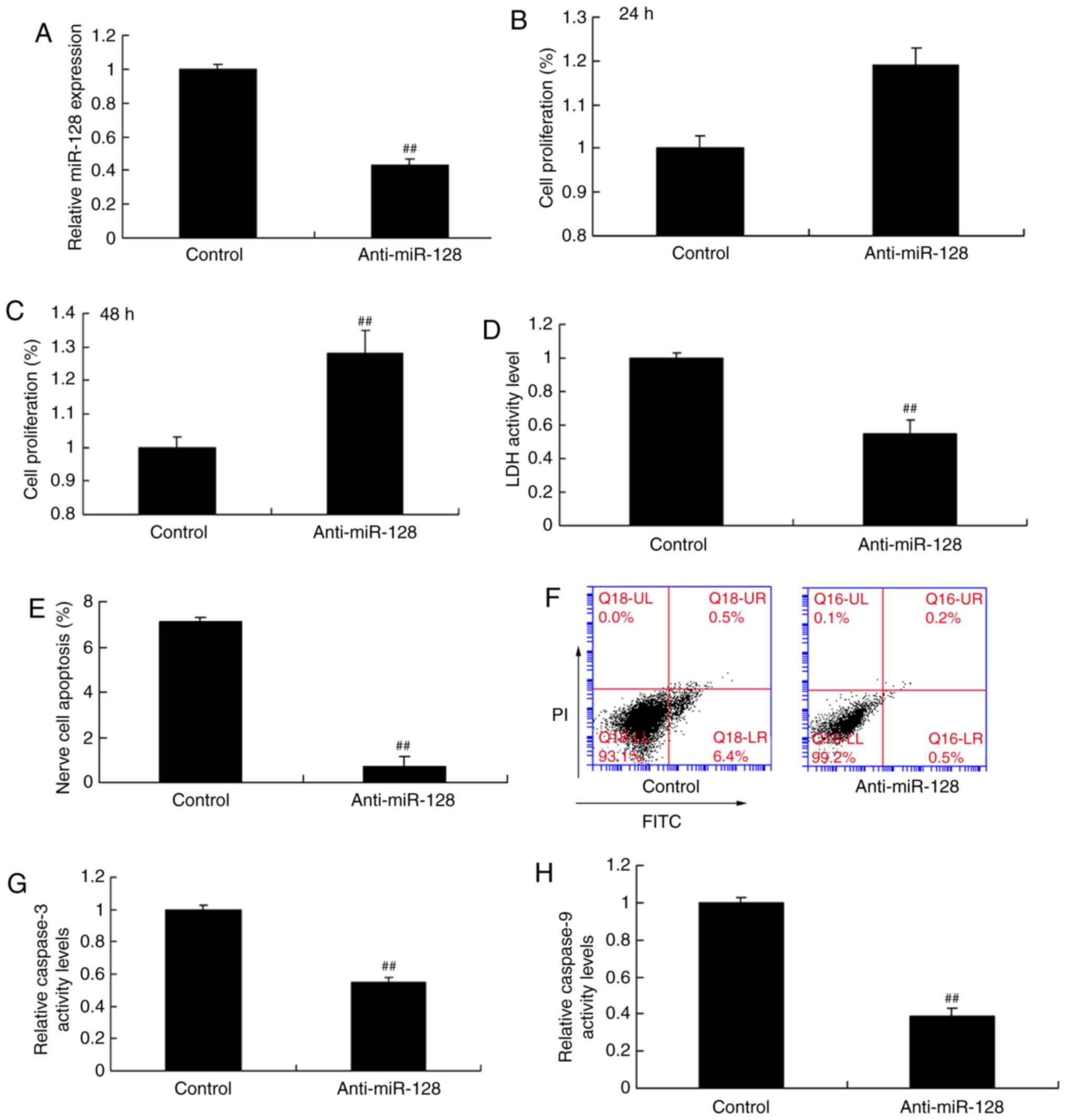
The effects of anti-miR-128 on regulating nerve cell
apoptosis were additionally examined. As illustrated in Fig. 3A, transfection of PC12 cells with
anti-miR-128 mimics significantly downregulated miR-128 expression
compared with the control group. miR-128 expression inhibition
promoted cell proliferation at 48 h (Fig. 3B and C), while it decreased LDH
activity (Fig. 3D), nerve cell
apoptosis (Fig. 3E and F) and
caspase-3/9 activity (Fig. 3G and
H) compared with the control group.
Effect of miR-128 on regulating the SIRT1
signaling pathway
Subsequently, the mechanism of miR-128 in regulating
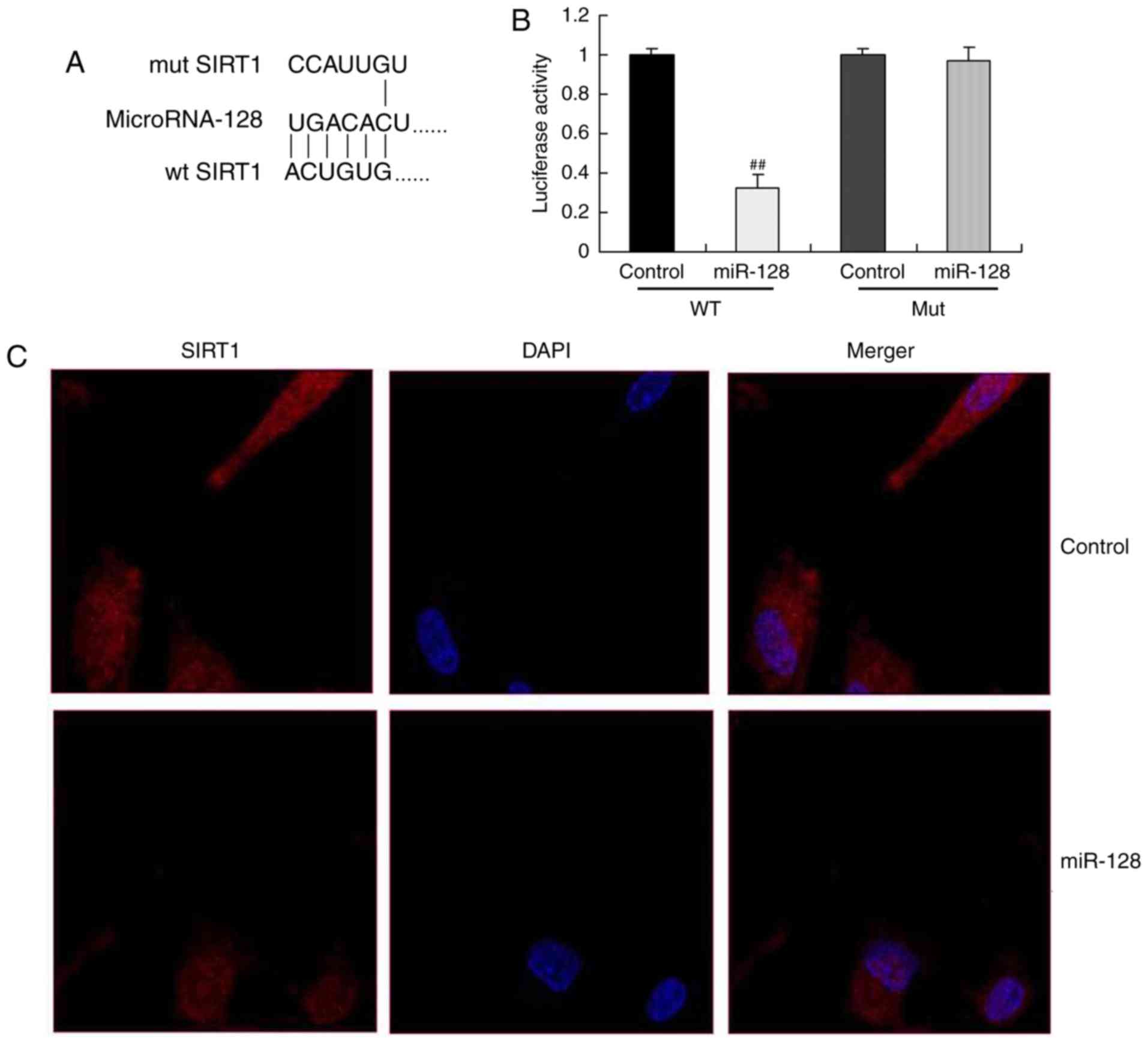
nerve cell apoptosis in epilepsy was evaluated. As illustrated in
Fig. 4A and B, the miR-128
binding site was deleted from the wild-type UTR SIRT1 3′UTR and the
luciferase activity of miR-128 was reduced in miR-128+wild-type
UTR, compared with the control mimics+wild-type UTR group; however,
the luciferase activity remained unaltered in the control
mimics+mutUTR or miR-128+mutUTR, which suggested that the miR-127
was binding to the wild-type UTR of SIRT1. In addition,
immunofluorescence analysis revealed that miR-128 mimics
transfection suppressed SIRT1 protein expression in vitro
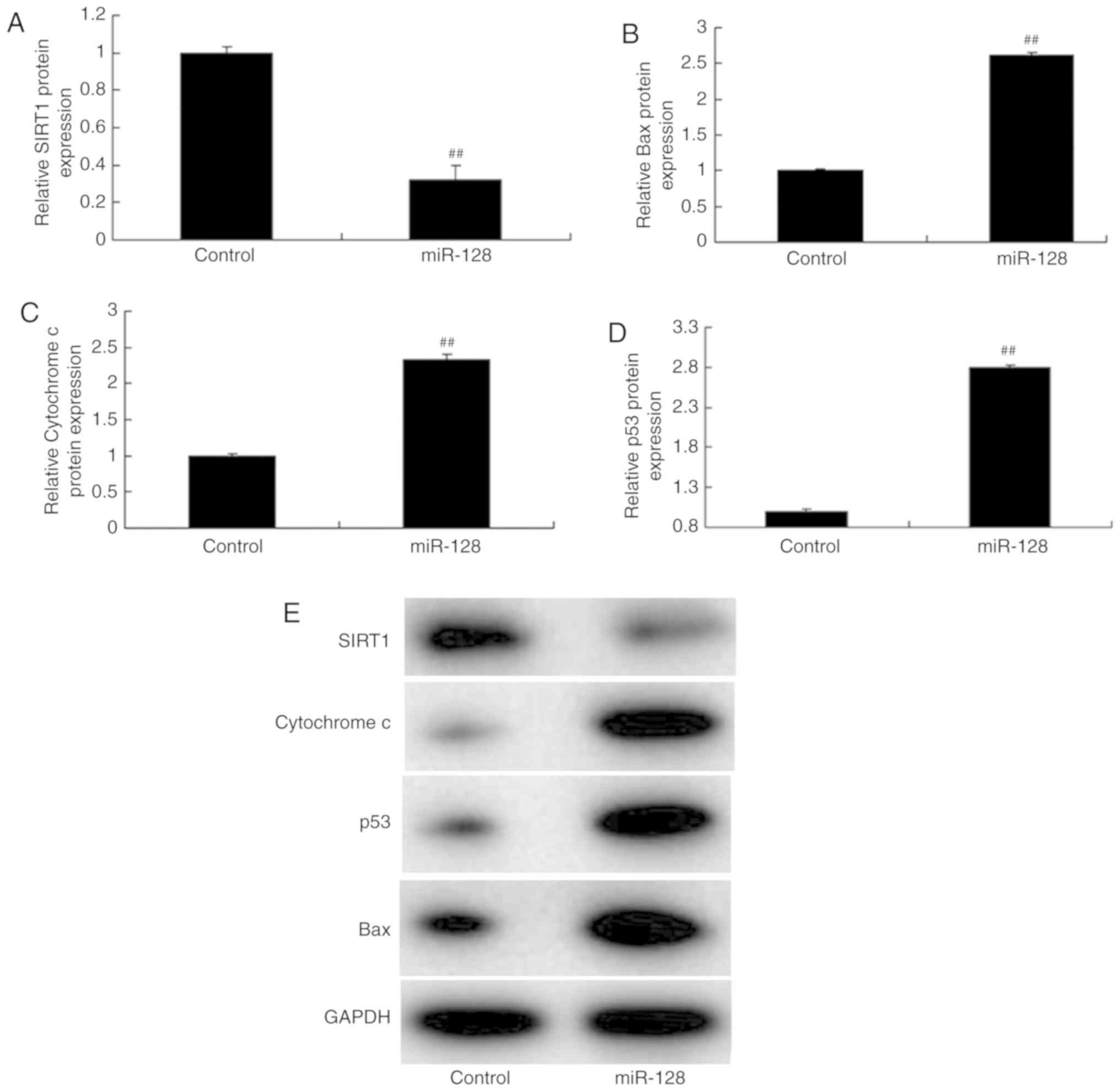
compared with the cells transfected with control mimics (Fig. 4C). The results of Fig. 5A-E demonstrated that upregulation
of miR-128 decreased SIRT1 protein expression, while it upregulated
the protein expression of p53, Bax and Cytochrome c in vitro
compared with the control group. By contrast, downregulation of
miR-128 by anti-miR-128 mimics transfection induced SIRT1 protein
expression, and suppressed the protein expression of p53, Bax and
Cytochrome c compared with the control group (Fig. 5F-J). In conclusion, the present
results suggested that the SIRT1 signaling pathway might be an
important mediator of the anti-epileptic effect of miR-128 on
apoptosis of PC12 cells in vitro.
SIRT1 promotion reverses the effects of
miR-128
It was hypothesized in the present study that the
SIRT1 signaling pathway was involved in the effect of miR-128.
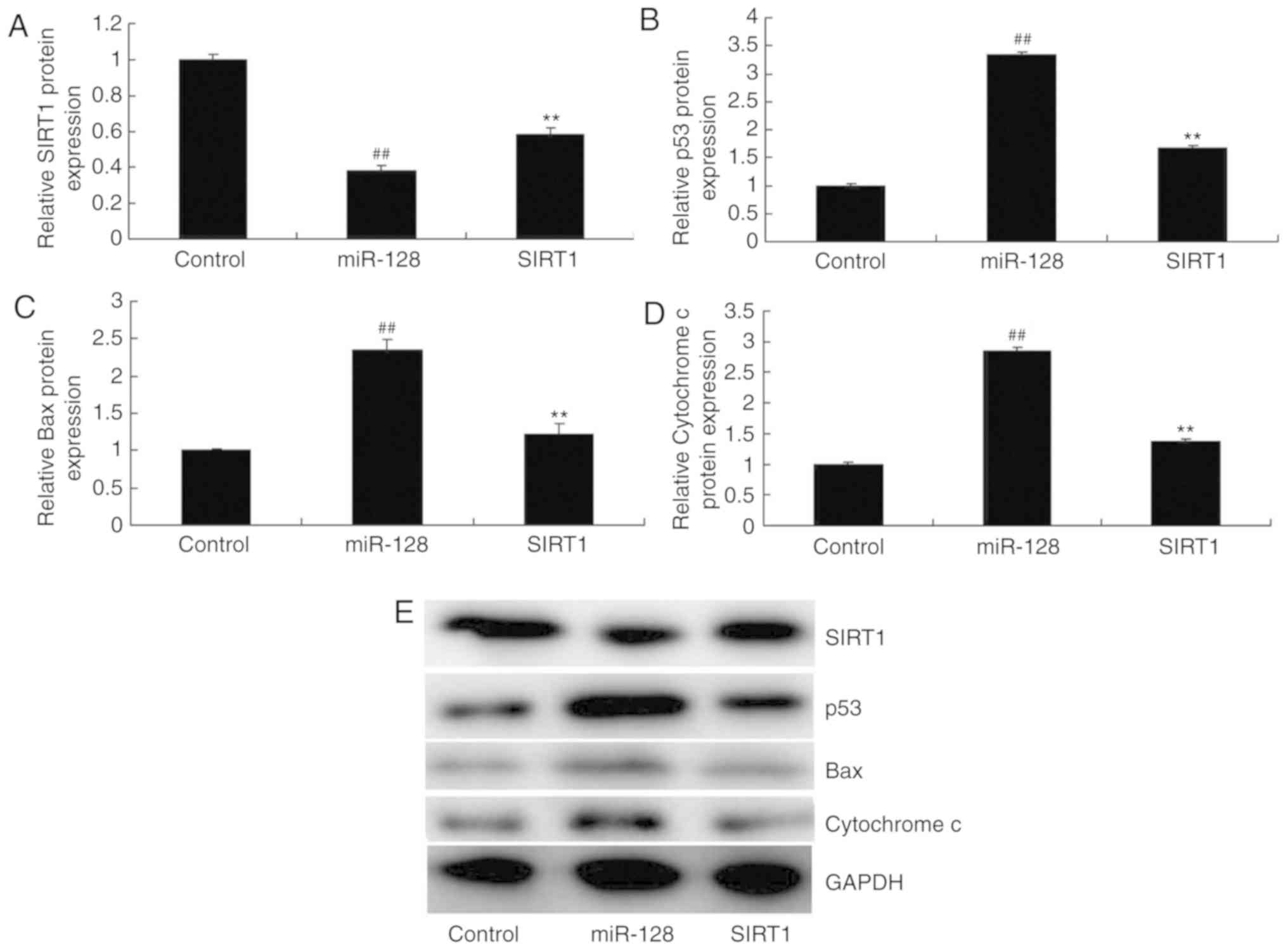
Consequently, the SIRT1 signaling pathway was examined by western
blotting following incubation of the cells with CAY10602, a SIRT1
agonist. Western blotting results demonstrated that the SIRT1
agonist could induce the protein expression of SIRT1, while it
suppressed p53, Bax and Cytochrome c expression in PC12
cells compared with the miR-128 alone group (Fig. 6). In addition, incubation with the
SIRT1 agonist inhibited the effect of miR-128 on cell proliferation
(Fig. 7A), while it increased LDH
activity (Fig. 7B), nerve cell
apoptosis (Fig. 7C and D) and
caspase-3/9 activity (Fig. 7E and
F) in vitro compared with the miR-128 alone group.
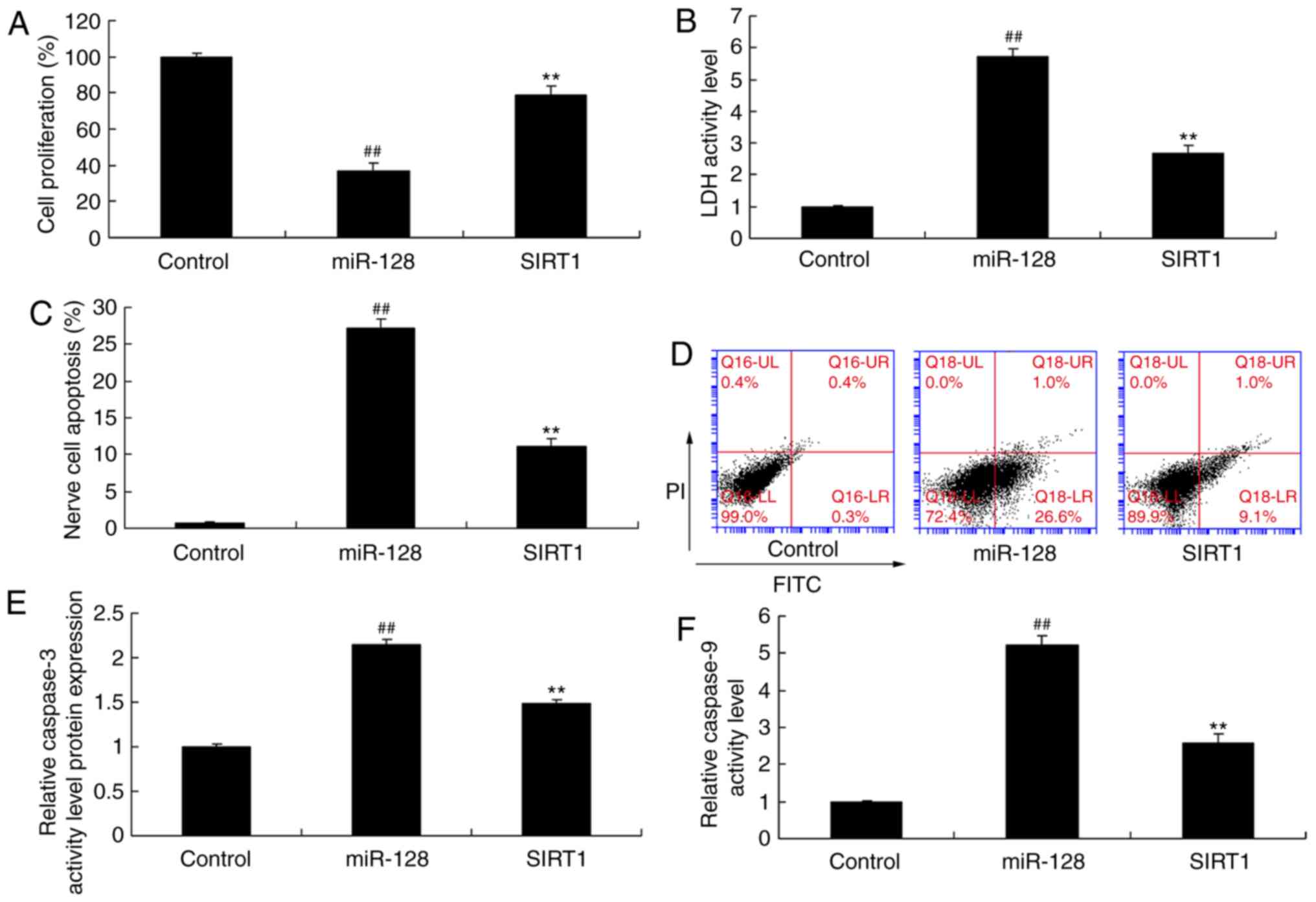 | Figure 7SIRT1 promotion reverses the function
of anti-miR-128 on apoptosis. (A) Cell proliferation, (B) LDH
activity levels, (C and D) apoptosis rate, and (E and F) caspase-3
and caspase-9 activity levels were measured in cells treated with
control, or miR-128 mimics alone, or miR-128 mimics and a SIRT1
agonist. ##P<0.01 compared with control group;
**P<0.01 compared with miR-128 group. SIRT1, sirtuin
1; LDH, lactate dehydrogenase; FITC, fluorescein isothiocyanate;
PI, propidium iodide; miR, microRNA. |
Inhibition of p53 reduces the effects of
miR-128
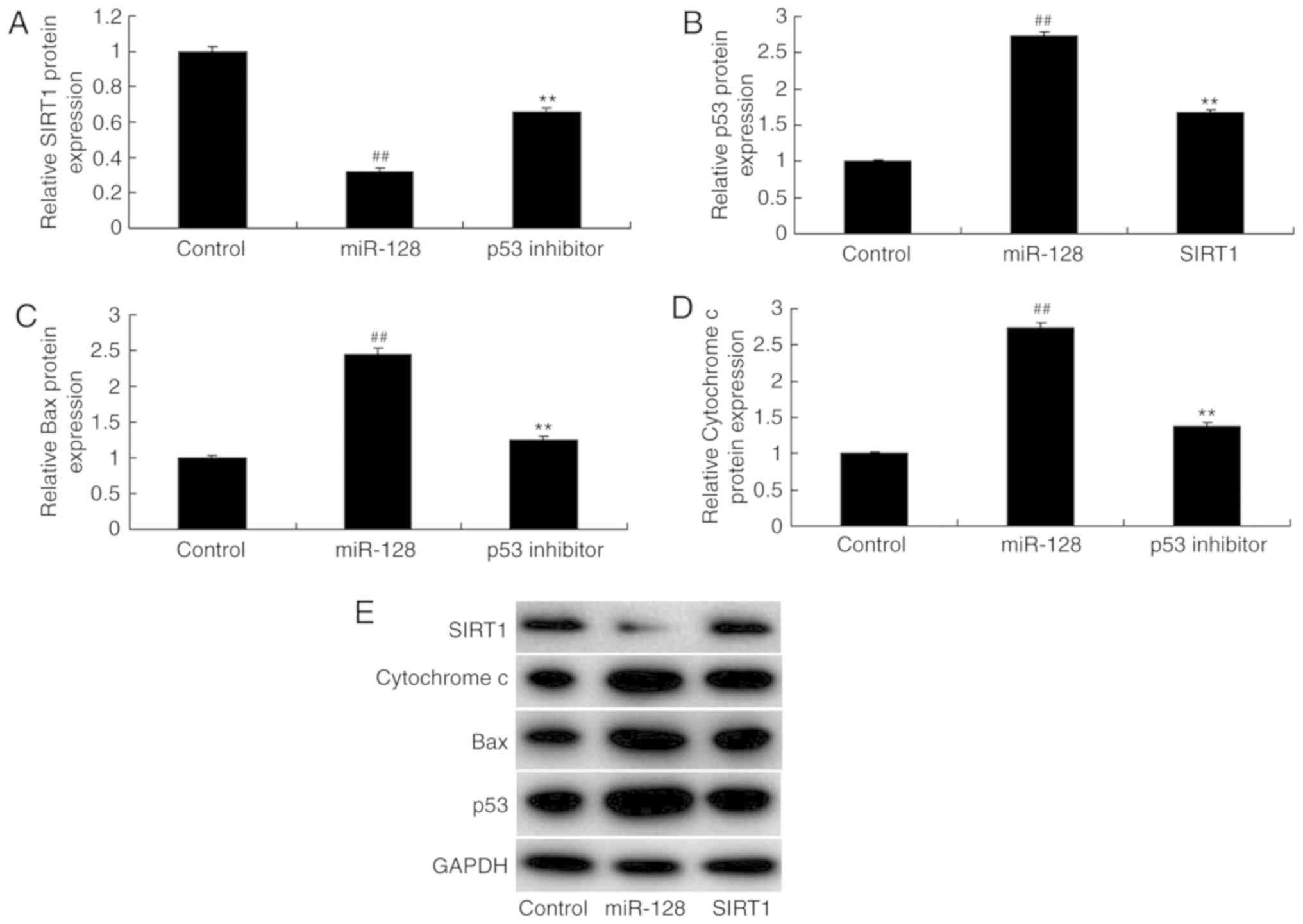
To determine whether the p53 signaling pathway was
functionally related to the homeostatic changes in the effect of
miR-128, cells were treated with p53 inhibitor and apoptosis was
measured. The results indicated that the p53 inhibitor
significantly inhibited p53, Bax and Cytochrome c compared
with the miR-128 alone group (Fig.
8). Compared with the miR-128 group, p53 inhibition
additionally suppressed the effects of miR-128 on inhibiting cell
proliferation and promoting LDH activity, nerve cell apoptosis and
caspase-3/9 activity (Fig.
9).
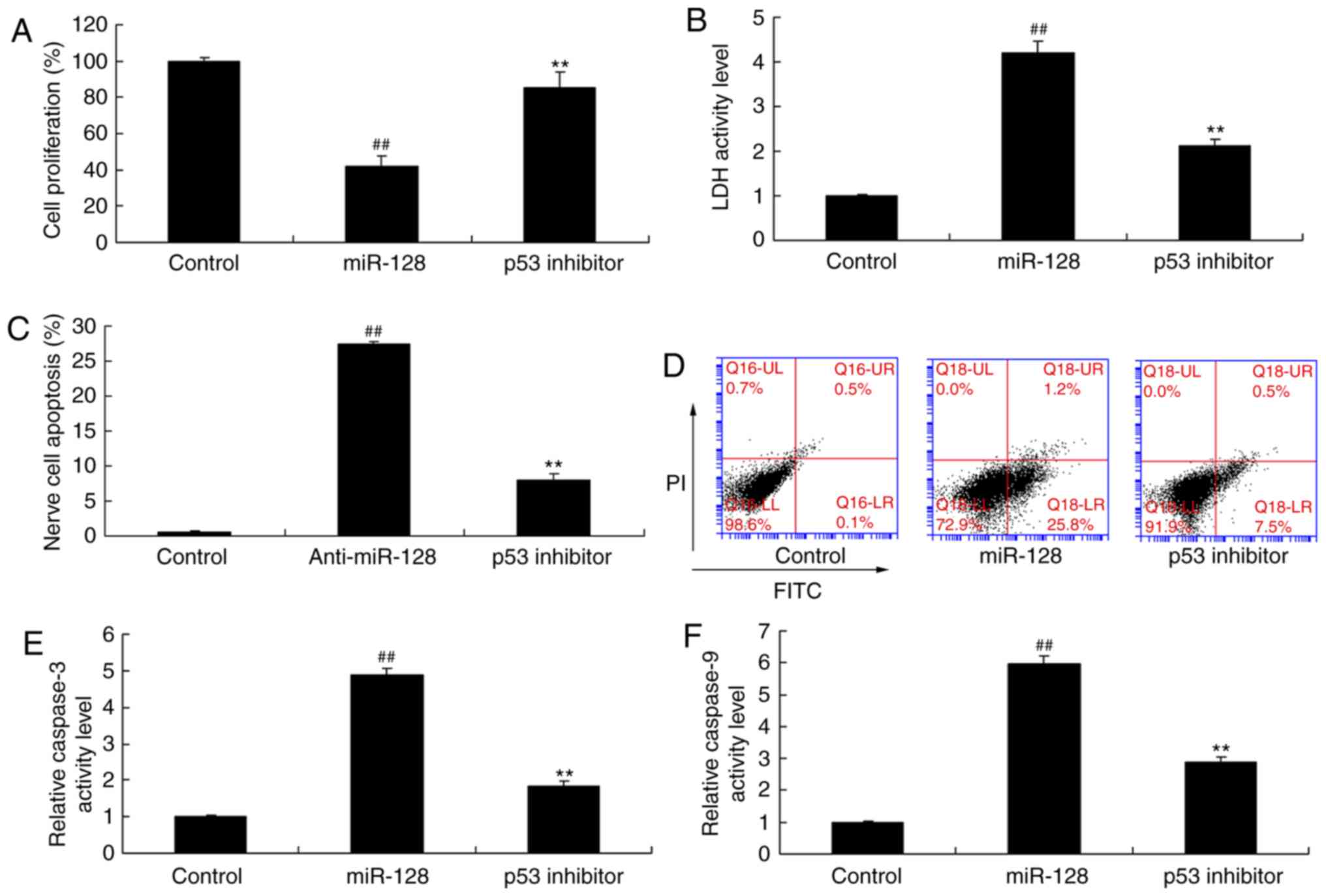 | Figure 9p53 inhibition reverses the function
of anti-miR-128 on apoptosis. (A) Cell proliferation, (B) LDH
activity levels, (C and D) apoptosis rate, and (E and F) caspase-3
and caspase-9 activity levels were measured in cells treated with
control, or miR-128 mimics alone, or miR-128 mimics and a p53
inhibitor. ##P<0.01 compared with control group;
**P<0.01 compared with miR-128 group. SIRT1, sirtuin
1; LDH, lactate dehydrogenase; FITC, fluorescein isothiocyanate;
PI, propidium iodide; miR, microRNA. |
Discussion
Epilepsy is a common nervous system disease, the
morbidity of which is ~25/1,000 in the general population (6). It severely affects the life, work
and learning of patients, which has thus become a chronic central
nervous system disease (5).
Therefore, investigating the pathogenesis of epilepsy is an
important topic in neuroscience currently (5). In the present study, miR-128
expression was demonstrated to be upregulated in an epilepsy rat
model compared with normal rats, while the upregulation of miR-128
induced nerve cell apoptosis in vitro. Only one cell line,
the PC12 cell line, was used in the present study, therefore
further studies are required in the future to verify the results in
additional cell lines and models.
miRNA are able to regulate protein synthesis at the
translation level. A previous study demonstrated that a number of
miRNAs in the brain are closely associated with synaptic
reorganization (8). Dendritic
spine exists in small protuberance with prominent dendrite. It has
become the extruding components of the primary excitatory synapse,
the dynamic alteration of which is an important form of synaptic
reorganization (10).
Anti-miR-128 is a miRNA with brain specificity, which may function
in the dendrite of nerve cells and negatively regulate the size of
dendritic spine (17). The
present study presented evidence that anti-miR-128 expression in an
epilepsy rat model was higher compared with normal rats. A previous
study has reported that miR-183, miR-135a, miR-125b, miR-30c and
miR-128 are upregulated at the seizure-associated phases and in
patients with temporal lobe epilepsy (13). The present results demonstrated
that downregulation of miR-128 promoted cell proliferation at 48 h,
while it reduced LDH activity, nerve cell apoptosis and caspase-3/9
activity in nerve cells in vitro.
SIRT1 is mainly expressed in neurons in the nervous
system, such as cortex, hippocampus, cerebellum and hypothalamus
(11). It serves a vital role in
neuron apoptosis and differentiation, cognitive function and
synaptic plasticity (11). SIRT1
exhibits protective effects on multiple acute and chronic nervous
system diseases (18). These
diseases include cerebral ischemia, Wallerian degeneration,
Huntington's disease, axonal injury, Alzheimer's disease,
Parkinson's disease, multiple sclerosis and amyotrophic lateral
sclerosis (18). Accumulating
evidence has indicated that SIRT1 is closely associated with
epilepsy (18). Specifically,
Resveratrol, the SIRT1 agonist, has anti-epileptic effect on
kainite- and FeCl3-induced epileptic animal models (19). The present findings suggested that
miR-128 directly binds and regulates SIRT1 expression. In addition,
upregulation of miR-128 induced SIRT1 protein expression, while it
suppressed the protein expression of p53, Bax and Cytochrome c
in vitro. Promotion of SIRT1 reduced the effect of miR-128 on
nerve cells in vitro. A previous study indicated that
miR-128 was able to promote apoptosis in human cancer via the
p53/Bak axis through SIRT1 (20),
whereas miR-128 has additionally been demonstrated to target SIRT1
in glioma subtypes (21).
The p53 gene is located on human chromosome 17p13.1.
It is the most common pro-apoptotic gene that serves a key role in
neural apoptosis (22). p53 is
highly expressed in the case of neural apoptosis (22). By contrast, the application of a
p53 gene blocker can suppress apoptosis. p53 protein
immunoreactivity was remarkably enhanced in hippocampus samples
that were surgically removed from patients with refractory temporal
lobe epilepsy (23). In addition,
the Bax gene is one of the important regulatory genes during cell
apoptosis (24). Bax is located
on the mitochondrial outer membrane (25). It can exert its apoptotic effect
through inhibiting tBid insertion, Bax transposition and Bax/Bax
oligomerization on the mitochondrial membrane (25). The present study demonstrated that
p53 inhibition reduced the effect of miR-128 on epilepsy. A
previous study has additionally suggested that miR-128 promotes
apoptosis in human cancers via the p53/Bak axis through SIRT1
(20).
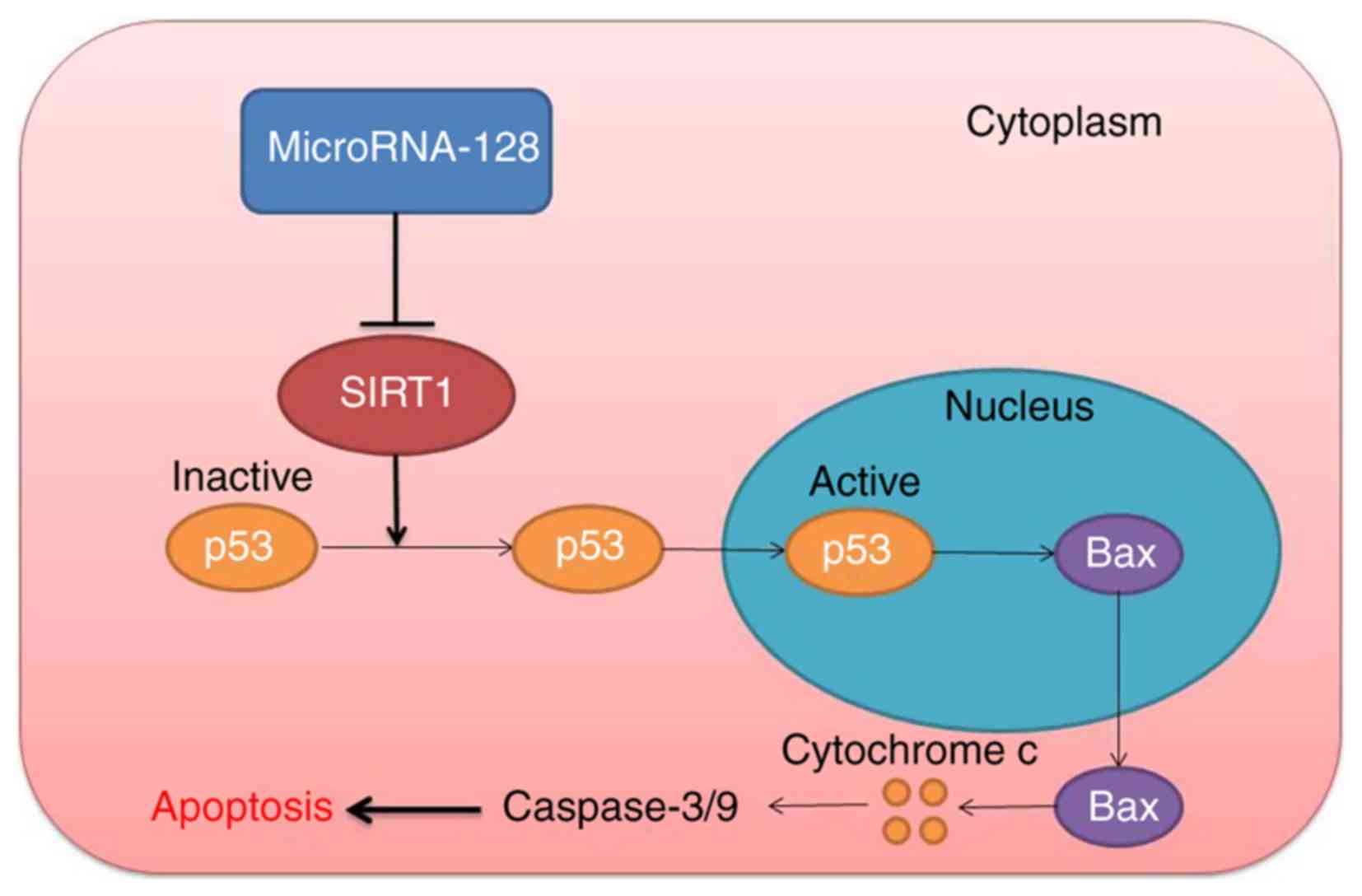
Finally, the present results demonstrated that
miR-128 levels were increased in epilepsy in vivo, and
downregulation of miR-128 induced nerve cell apoptosis through the
SIRT1/p53/Bax/Cytochrome c/caspase signaling pathway
(Fig. 10). Therefore,
upregulation of miR-128 may promote apoptosis in epilepsy model
in vivo and in vitro through the
SIRT1/p53/Bax/Cytochrome c/caspase signaling pathway.
Therefore, miR-128 may have potential as an antiepileptic target in
the clinic.
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
WWW designed the experiment. DZC, YLC, XFY, MZ and
YYY performed the experiments. WWW and DZC analyzed the data. WWW
wrote the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Protocols involving animals in the present study
were approved by the Institutional Animal Care and Ethics Committee
of Liaocheng People's Hospital (Liaocheng, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Freitas-Lima P, Alexandre V Jr, Pereira
LR, Feletti F, Perucca E and Sakamoto AC: Influence of enzyme
inducing antiepileptic drugs on the pharmacokinetics of
levetiracetam in patients with epilepsy. Epilepsy Res. 94:117–120.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Abd El Naby SA and Naguib YM:
Sociodemographic, electro-physiological, and biochemical profiles
in children with attention deficit hyperactivity disorder and/or
epilepsy. Behav Neurol. 2018:89328172018. View Article : Google Scholar
|
|
3
|
Angus-Leppan H: Diagnosing epilepsy in
neurology clinics: A prospective study. Seizure. 17:431–436. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Phabphal K, Geater A, Limapichat K,
Sathirapanya P, Setthawatcharawanich S and Leelawattana R: Effect
of switching hepatic enzyme-inducer antiepileptic drug to
levetiracetam on bone mineral density, 25 hydroxyvitamin D, and
parathyroid hormone in young adult patients with epilepsy.
Epilepsia. 54:e94–e98. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ryvlin P, Gilliam FG, Nguyen DK, Colicchio
G, Iudice A, Tinuper P, Zamponi N, Aguglia U, Wagner L, Minotti L,
et al: The long-term effect of vagus nerve stimulation on quality
of life in patients with pharmacoresistant focal epilepsy: The
PuLsE (Open Prospective Randomized Long-term Effectiveness) trial.
Epilepsia. 55:893–900. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dalal K, Devarajan E, Pandey RM, Subbiah V
and Tripathi M: Role of reflexology and antiepileptic drugs in
managing intractable epilepsy-a randomized controlled trial. Forsch
Komplementmed. 20:104–111. 2013. View Article : Google Scholar
|
|
7
|
Reschke CR and Henshall DC: microRNA and
Epilepsy. Adv Exp Med Biol. 888:41–70. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Moon J, Lee ST, Choi J, Jung KH, Yang H,
Khalid A, Kim JM, Park KI, Shin JW, Ban JJ, et al: Unique
behavioral characteristics and microRNA signatures in a drug
resistant epilepsy model. PLoS One. 9:e856172014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Srivastava A, Dixit AB, Banerjee J,
Tripathi M and Sarat Chandra P: Role of inflammation and its miRNA
based regulation in epilepsy: Implications for therapy. Clin Chim
Acta. 452:1–9. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li MM, Li XM, Zheng XP, Yu JT and Tan L:
MicroRNAs dysregulation in epilepsy. Brain Res. 1584:94–104. 2014.
View Article : Google Scholar
|
|
11
|
Hall AM, Brennan GP, Nguyen TM,
Singh-Taylor A, Mun HS, Sargious MJ and Baram TZ: The Role of Sirt1
in Epileptogenesis. eNeuro. 4:ENEURO.0301–16.2017. 2017. View Article : Google Scholar
|
|
12
|
Wang D, Li Z, Zhang Y, Wang G, Wei M, Hu
Y, Ma S, Jiang Y, Che N, Wang X, et al: Targeting of
microRNA-199a-5p protects against pilocarpine-induced status
epilepticus and seizure damage via SIRT1-p53 cascade. Epilepsia.
57:706–716. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Alsharafi W and Xiao B: Dynamic expression
of MicroRNAs (183, 135a, 125b, 128, 30c and 27a) in the rat
pilocarpine model and temporal lobe epilepsy patients. CNS Neurol
Disord Drug Targets. 14:1096–1102. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang L, Liang L, Yang T, Qiao Y, Xia Y,
Liu L, Li C, Lu P and Jiang X: A pilot clinical study of apatinib
plus irinotecan in patients with recurrent high-grade glioma:
Clinical trial/experimental study. Medicine (Baltimore).
96:e90532017. View Article : Google Scholar
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
16
|
Chen L, Zheng H and Zhang S: Involvement
of upregulation of miR-210 in a rat epilepsy model. Neuropsychiatr
Dis Treat. 12:1731–1737. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang XM, Jia RH, Wei D, Cui WY and Jiang
W: MiR-134 blockade prevents status epilepticus like-activity and
is neuro-protective in cultured hippocampal neurons. Neurosci Lett.
572:20–25. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang W, Zhang J, Li Y, Yang X, He Y, Li T,
Ren F, Zhang J and Lin R: Divalproex sodium enhances the
anti-leukemic effects of imatinib in chronic myeloid leukemia cells
partly through SIRT1. Cancer Lett. 356:791–799. 2015. View Article : Google Scholar
|
|
19
|
Qian C, Jin J, Chen J, Li J, Yu X, Mo H
and Chen G: SIRT1 activation by resveratrol reduces brain edema and
neuronal apoptosis in an experimental rat subarachnoid hemorrhage
model. Mol Med Rep. 16:9627–9635. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Adlakha YK and Saini N: miR-128 exerts
pro-apoptotic effect in a p53 transcription-dependent and
-independent manner via PUMA-Bak axis. Cell Death Dis. 4:e5422013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lages E, Guttin A, El Atifi M, Ramus C,
Ipas H, Dupré I, Rolland D, Salon C, Godfraind C, deFraipont F, et
al: MicroRNA and target protein patterns reveal physiopathological
features of glioma subtypes. PLoS One. 6:e206002011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Engel T, Murphy BM, Schindler CK and
Henshall DC: Elevated p53 and lower MDM2 expression in hippocampus
from patients with intractable temporal lobe epilepsy. Epilepsy
Res. 77:151–156. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Araki T, Shinoda S, Schindler CK, Quan-Lan
J, Meller R, Taki W, Simon RP and Henshall DC: Expression,
interaction, and proteolysis of death-associated protein kinase and
p53 within vulnerable and resistant hippocampal subfields following
seizures. Hippocampus. 14:326–336. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao H, Lin G, Shi M, Gao J, Wang Y, Wang
H, Sun H and Cao Y: The mechanism of neurogenic pulmonary edema in
epilepsy. J Physiol Sci. 64:65–72. 2014. View Article : Google Scholar
|
|
25
|
Rabie T, Mühlhofer W, Bruckner T, Schwab
A, Bauer AT, Zimmermann M, Bonke D, Marti HH and Schenkel J:
Transient protective effect of B-vitamins in experimental epilepsy
in the mouse brain. J Mol Neurosci. 41:74–79. 2010. View Article : Google Scholar
|