Introduction
Skin scars could be divided into several categories,
for example, hypertrophic scar (HS), keloid scar and atrophic scar
(or sunken scar). Among these, HS, a benign hyperproliferative
growth of dermal collagen, originates from unbalanced fibroblast
cellular dynamics that result from an elevated proliferation and a
reduced apoptosis of fibroblasts (1). HS is an aberrant healing response,
secondary to traumatic injuries, empyrosis and surgical trauma
(2). The formation of HS has been
demonstrated to be relevant to a proliferative stage of wound
healing, during which dermal tissue hyperplasia and the
over-deposition of extracellular matrix (ECM) proteins derived from
fibroblasts can cause durative fibrosis and inflammation (3).
Fibroblast-to-myofibroblast transformation is a
vital event during wound healing and hypertrophic scar formation
(4). Fibroblasts arise at the
wound site at the ultimate inflammatory stage and the initial
proliferative phase of healing wound (5). Myofibroblasts, which are
differentiated from fibroblasts, are primitively produced in the
local derma and hypodermal tissues around the injured skin in HS
(6) in addition to other origins,
for instance, bone marrow-derived fibrocytes (7), tissue-specific stem cells (8), pericytes and vascular smooth muscle
cells (9) and tubular epithelial
cells with epithelial-mesenchymal transition (EMT) (10). Myofibroblasts in granulation
tissues can lead to HS formation; however, the roles of these
myofibroblasts in the pathogenesis of HS remain unclear.
Transforming growth factor-β (TGF-β) may serve as a
therapeutic target for HS and may be an important regulatory factor
in the process of HS (4).
TGF-β1/Smad2 signaling can promote collagen synthesis and enhance
the proliferation of human HS fibroblasts (11). A previous study revealed that
TGF-β can exert differential temporal effects on HS formation
(12). Moreover, TGF-β1 is able
to induce Smad2 nuclear translocation, finally causing the
transcription of target genes, including collagens I and III
(13). Moreover, the silencing of
Smad2 can prevent type I and III collagen overproduction in HS
fibroblasts (14). Recently,
multiple agents, such as baicalein (15), loureirin B (16), tetrandrine (17) and botulinum toxin type A (BTXA)
(1), have been reported to exert
anti-HS effects by inhibiting TGF-β1 signaling.
Botulinum toxin is an effective neurotoxin generated
from Clostridium botulinum and it has been shown to have
potential for use in the treatment of wounds following trauma,
burns or surgery (18). BTXA is
available for clinical use in a number of countries as its safe and
has effective properties in the treatment of hypertrophic scarring
(20). Rasaii reported that
triamcinolone in combination with BTXA was ineffective for keloid
scars (21), while BTXA was
demonstrated to attenuate HS growth and improve the symptoms of HS
in clinical practice (22).
Currently, an increasing number of studies have demonstrated that
BTXA can alleviate fibrosis by regulating fibroblast behaviors in
HS and capsular contracture. For instance, BTXA can attenuate HS
and capsular contracture by suppressing the phenotypic
transformation of fibroblasts to myofibroblasts, according to in
vitro and in vivo experiments (1,19).
BTXA can markedly reduce the collagen deposition in HS (22-24). Furthermore, BTXA has been
demonstrated to inhibit fibroblast proliferation and to reduce the
level of α-smooth muscle actin (α-SMA) (25,26). Furthermore, BTXA can also regulate
the expression levels of matrix metalloproteinase (MMP)-2 MMP-9,
and collagen types I and III, in addition to inhibiting capsule
formation (27).
HS fibroblasts have been reported to possess
malignant characteristics, including hyperproliferation,
anti-apoptosis and atypical differentiation (28-31). Tansdifferentiation, proliferation
and apoptosis have been demonstrated to be regulated by the
phosphatidylinositol 3-kinase (PI3K)/Akt signaling pathway in HS
fibroblasts (32-34). Moreover, Akt is also involved in
cell motility and ECM deposition (35,36). Phosphatase and tensin homologue
deleted on chromosome ten (PTEN) protein has reported as a tumor
suppressor and it can negatively regulate Akt signaling to inhibit
the proliferation of HS fibroblasts (37). Inactive PTEN expression has been
observed in several fibrotic diseases (38-40). Therefore, HS may be relevant to
the antagonistic association between PTEN and Akt. In the present
study, our aim was to explore the potential mechanisms involved in
PTEN//PI3K/Akt signaling and the effects of BTXA on the phenotypic
transformation of fibroblasts induced by TGF-β1. Our findings are
expected to provide a theoretical basis for the treatment of
HS.
Materials and methods
Fibroblast culture and morphological
identification
Murine L929 fibroblasts (41) (Cat no. CC-Y2049, ATCC) were
maintained in the Dulbecco's minimal Eagle's medium with 10% fetal
bovine serum, penicillin (100 U/ml) and streptomycin (100 mg/ml) at
37°C with 5% CO2 (all purchased from Invitrogen; Thermo
Fisher Scientific). The morphological changes of the fibroblasts
were observed using an inverted microscope (Nikon). For vimentin
identification, the fibroblasts were fixed with 4% paraformaldehyde
for 0.5 h and incubated in 0.5% Triton X-100 (Sigma-Aldrich) for 20
min. After being blocked in bovine serum albumin (BSA; Solarbio),
the fibroblasts were incubated with anti-vimentin antibody
(M00235-1, 1:100; BosterBio) for 1 h at room temperature. The
fibroblasts were then incubated with goat anti-rabbit IgG H&L
secondary antibodies (1:5,000; ab6721; Abcam) for 1 h. The nuclei
were stained with Hoechst 33258 (Beyotime) for 20 min at 37°C in
the dark and the stained fibroblasts were observed under a
fluorescence microscope (Nikon).
Fibroblast treatment and viability
assay
Fibroblasts (2×105) were maintained in
96-well plates for 24 h, and fresh media were then added with
various concentrations of BTXA (0, 0.125, 0.25, 0.5, 1 and 2 UI/ml;
Lanzhou Biochemical Co.). Cell counting kit-8 (CCK-8; Beyotime)
solution (20 µl), with the original concentration in the
kit, was incubated with the fibroblasts for an additional 2 h at
37°C when the fibroblasts were incubated for 12, 24 and 48 h,
respectively. Thereafter, other fresh fibroblasts were incubated
with TGF-β1 (10 ng/ml) (4) for 3
h, and the fibroblasts were subsequently treated with low, medium
and high concentrations of BTXA (0.25, 0.5 and 1 UI/ml,
respectively). Subsequently, 20 µl CCK-8 were incubated with
the fibroblasts for an additional 2 h at 37°C, for the
determination of fibroblast viability. The absorbance of 450 nm was
determined using a multimode detector (BioTek Instruments).
Apoptosis analysis
Apoptosis was detected according to the protocol of
the Annexin V-fluorescein isothiocyanate (FITC)/propidium iodide
(PI) apoptosis detection kit (KeyGEN). The results were analyzed
using a FACSCalibur flow cytometer (BD Biosciences). The cells were
then identified using Annexin V-FITC/PI double fluorescence
staining as follows: Unlabeled, viable cells; PI-stained cells,
necrotic cells; Annexin V-FITC-bounded cells, early apoptotic
cells; and double-labeled cells, late apoptotic cells.
Immunofluorescence assay
Fibroblasts were fixed with 4% paraformaldehyde for
20 min and washed with PBS. After being blocked in bovine serum
albumin (BSA), the fibroblasts were incubated first with anti-α-SMA
antibody (A03744, 1:200; BosterBio) overnight at 4°C and then with
Cy3-conjugated goat anti-rabbit secondary antibodies (BA1032,
1:500; Beyotime) for 0.5 h. Subsequently, the fibroblasts were
stained with 4′,6-diamidino-2-phenylindole (DAPI; Beyotime) and
images were captured under a fluorescence microscopy (Nikon).
Methylation-specific PCR (MSP)
DNA was isolated using the DNeasy tissue kit
(Qiagen) and then analyzed by MSP using bisulfite-modified DNA. A
total of 1 µg purified DNA was treated according to the
CpGenome DNA Modification kit (Intergen). CpGenome Universal
Methylated DNA (Intergen) and normal mouse fibroblast DNA served as
methylated and unmethylated controls, respectively. The
amplification for methylated (M) PTEN (forward, 5′-TTG ATT
AAC GCG GTT AGT TAG TTC-3′ and reverse, 5′-AAC GCA
TAT CCT ACC GCA ATA C-3′) and unmethylated (U) PTEN
(forward, 5′-GTG TTG ATT AAT GTG GTT AGT TAG TTT-3′
and reverse, 5′-CCA AAC ACA TAT CCT ACC ACA ATA C-3′)
was performed under the following conditions: At 94°C for 2 min,
then 36 cycles at 94°C for 30 sec, at 54°C for 30 sec and at 72°C
for 45 sec, and finally 7 min at 72°C. PCR products were analyzed
on a 2% agarose gel with ethidium bromide.
DNA methyltransferase (DNMT)
activity
The nuclear protein was extracted from the
fibroblasts using the Nuclear Extract kit (Active Motif). Total
DNMT activity was measured according to the protocol of the EpiQuik
DNMT Activity/Inhibition Assay Ultra kit (Epigentek).
RNA extraction, cDNA synthesis and
reverse transcription- quantitative PCR (RT-qPCR)
Total RNA was extracted from the mouse fibroblasts
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific). In
brief, TRIzol reagent and chloroform were added to the samples and
mixed for 5 min, and the supernatant was recovered by
centrifugation, at 10,000 x g, at 4°C for 20 min. The supernatant
was then incubated with an equal volume of isopropyl alcohol and
then centrifuged for 10 min. After dislodging the supernatants, 75%
ethanol was added to wash the precipitate and the RNA was eluted
with nuclease-free water. The purity and content of the reverse
transcription was determined using a NanoDrop 2000
spectrophotometer (Thermo Fisher Scientific). cDNA was obtained by
RNA with mixture in the PrimeScriptTM 1st Strand cDNA
Synthesis kit (Takara). The reactions (primers are listed in
Table I) were conducted with a
LightCycler system (Roche Applied Science) using the following
parameters: at 95°C for 10 min, 40 cycles at 95°C for 15 sec and at
60°C for 60 sec. GAPDH was used as the internal control and the
results were calculated using the 2−ΔΔCq method
(42).
 | Table ISequences of primers used for RT-qPCR
assays. |
Table I
Sequences of primers used for RT-qPCR
assays.
| Gene name | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| Collagen I |
ACTGTCTTGCCCCAAGTTCC |
TGGGCATCTGGTTTAGCCTT |
| Collagen III |
ACGTAGATGAATTGGGATGCAG |
GGGTTGGGGCAGTCTAGTG |
| α-SMA |
AGGGGAATGAAAAGCCGGAA |
TAGGATATGCCTGGGGGTC |
| MMP-2 |
ACCATCGAGACCATGCGG |
CTCCCCCAACACCAGTGC |
| MMP-9 |
TTCTGCCCTACCCGAGTGGA |
CATAGTGGGAGGTGCTGTCGG |
| PTEN |
AATTCCCAGTCAGAGGCGCTATGT |
GATTGCAAGTTCCGCCACTGAACA |
| DNMT1 |
AGTGCAAGGCGTGCAAAGATATGG |
TGGGTGATGGCATCTCTGACACAT |
| DNMT3a |
GCCGAATTGTGTCTTGGTGGATGACA CC |
TGGTGGAATGCACTGCAGAAGGA |
| DNMT3b |
TTCAGTGACCAGTCCTCAGACACGAA |
TCAGAAGGCTGGAGACCTCCCTCTT |
| GAPDH |
GTAGAGGCAGGGATGATGTTCT |
CTTTGGTATCGTGGAAGGACTC |
Western blot analysis
The fibroblasts were lysed in lysis buffer
(Beyotime) and centrifuged for supernatant recovery. The protein
concentrations were determined using a BCA kit (Beyotime). After
being separated by 10% SDS-PAGE, the proteins (20 µg) were
transferred onto nitrocellulose membranes (Millipore) and blocked
with 5% non-fat dry milk for 1 h at room temperature. The membranes
were then first incubated at 4°C overnight with anti-PTEN (1:200;
cat. no. 9188), anti-DNMT1 (1:1,000; cat. no. 5032), anti-DNMT3a
(1:1,000; cat. no. 49768), anti-DNMT3b (1:1,000; cat. no. 48488),
anti-PI3K (1:1,000; cat. no. 4255), anti-p-PI3K (1:1,000; cat. no.
4228), anti-Akt (1:1,000; cat. no. 4685), anti-p-Akt (1:1,000; cat.
no. 4056), anti-MMP-2 (1:1,000; cat. no. 87809) (all from Cell
Signaling Technology), anti-α-SMA (1:350; cat. no. A03744,
BosterBio), anti-MMP-9 (1:500; sc-21733), anti-caspase-3 (1:200;
sc-271759), anti-collagen I (1:1,000; sc-376350), anti-collagen III
(1:1,000; sc-271249) and anti-β-actin (1:1,000; sc-58673) (all from
Santa Cruz Technology) antibodies and then with secondary
antibodies [goat anti-rabbit IgG H&L (HRP) (1:5,000; ab6721,
Abcam)] for 2 h at room temperature. The proteins were visualized
using an ECL kit (Amersham Biosciences, Germany), and analyzed with
the Bio-Rad ChemiDoc™ XRS+ System and Image Lab™ Software version
4.1 (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
All results are presented as the means ± standard
deviation. All statistical analyses were performed with SPSS 17.0
software (SPSS, Inc.), and the differences between two groups were
analyzed by a t-test or by one-way ANOVA with Dunnett's post hoc
test for multiple group comparisons. P<0.05 was considered to
indicate a statistically significant difference.
Results
Effect of BTXA on the viability of mouse
L929 fibroblasts
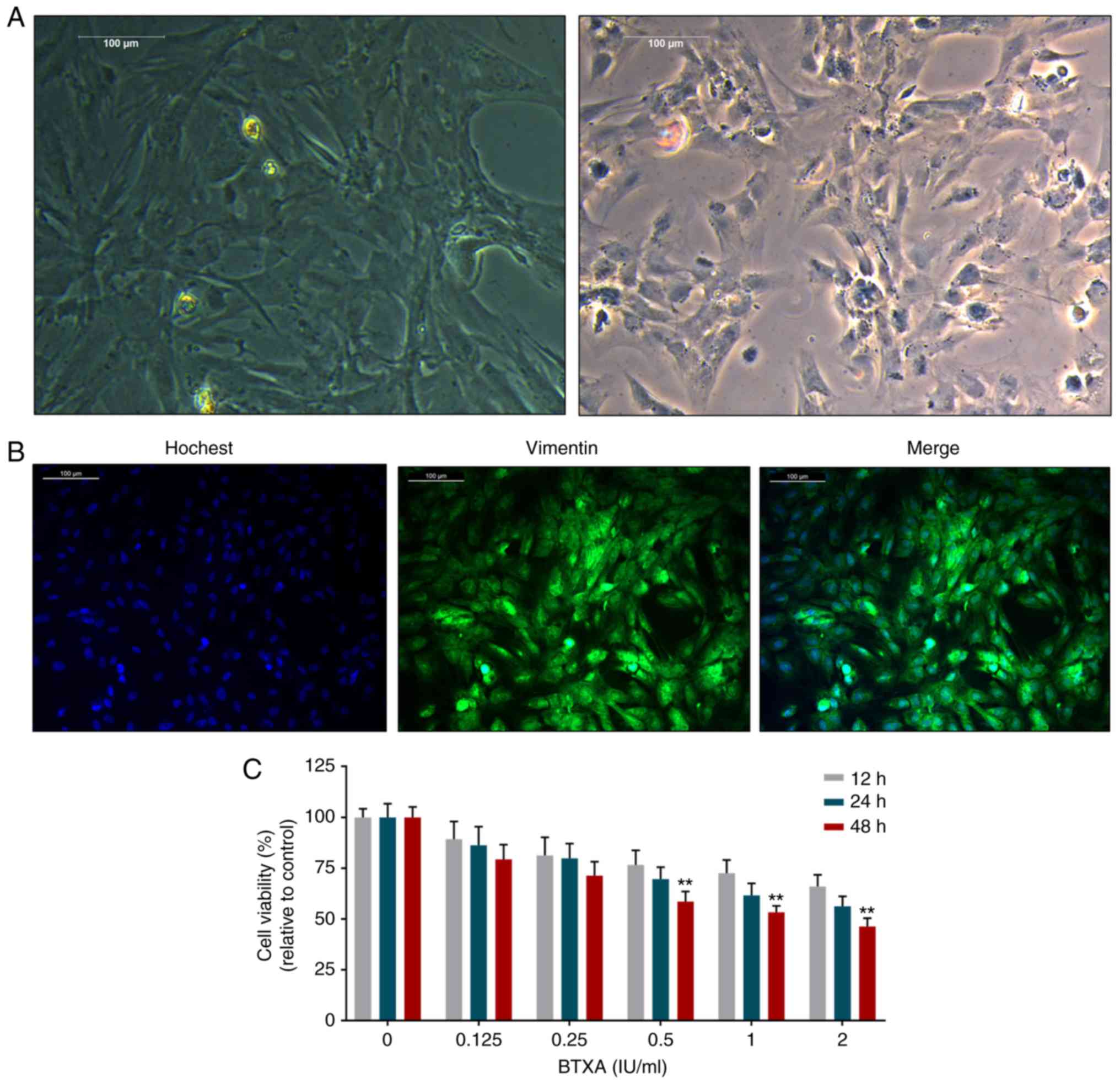
The fibroblasts exhibited spindle-like, ellipse and
irregular shapes with large nuclei and transparent cytoplasms
(Fig. 1A). Moreover, vimentin
expression was significantly positive in the fibroblasts (Fig. 1B). However, BTXA was found to
inhibit the viability of the fibroblasts in a dose- and
time-dependent manner (Fig.
1C).
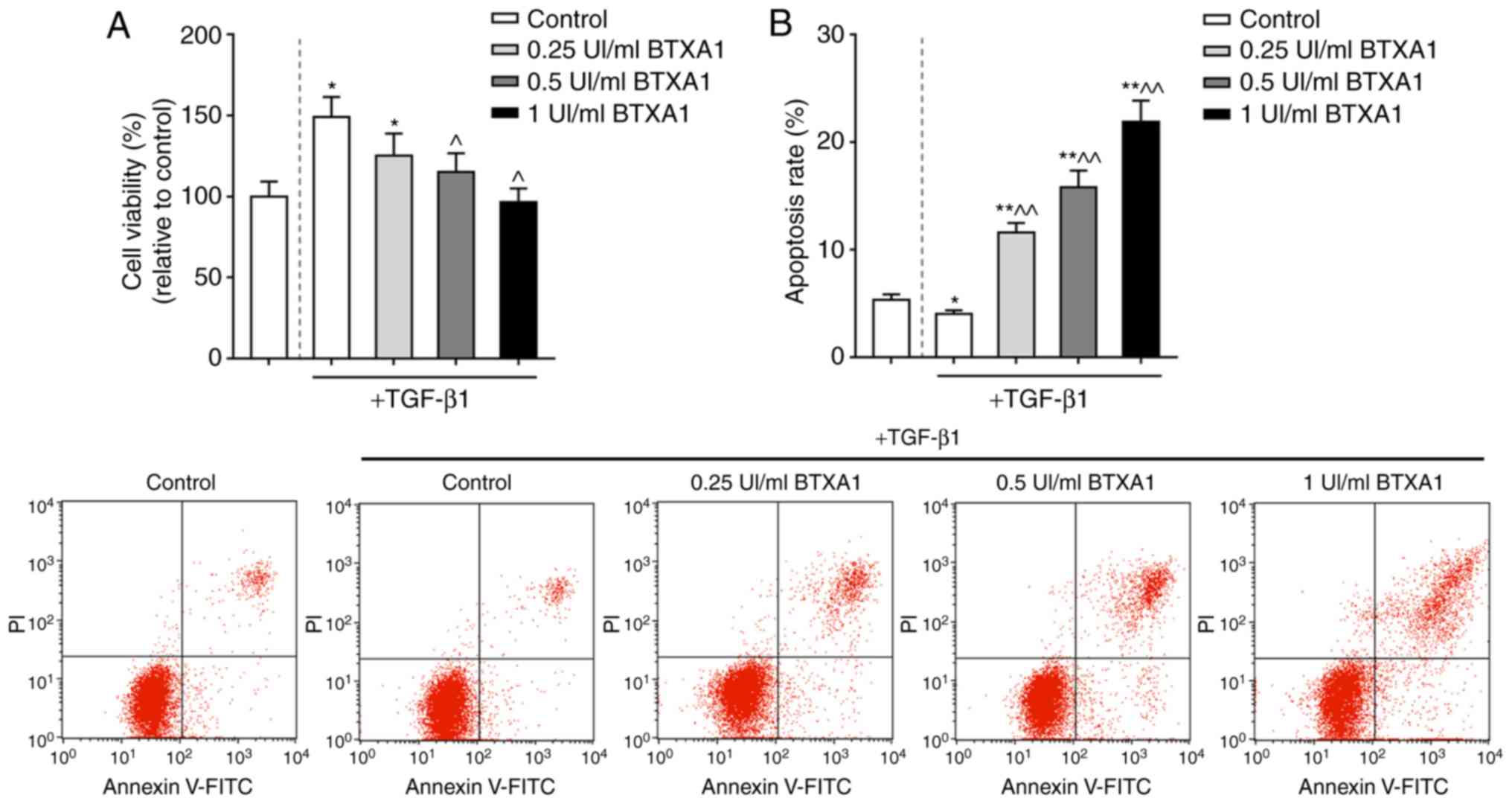
TGF-β1 is a crucial factor for the induction of the
transformation of fibroblasts into myofibroblasts. In this study,
the proliferation of fibroblasts treated with TGF-β1 was markedly
increased. However, the increased viability of the fibroblasts
induced by TGF-β1 was significantly inhibited by BTXA in a
concentration-dependent manner (P<0.05; Fig. 2A). Moreover, the apoptosis of the
fibroblasts was markedly increased by BTXA (P<0.01; Fig. 2B).
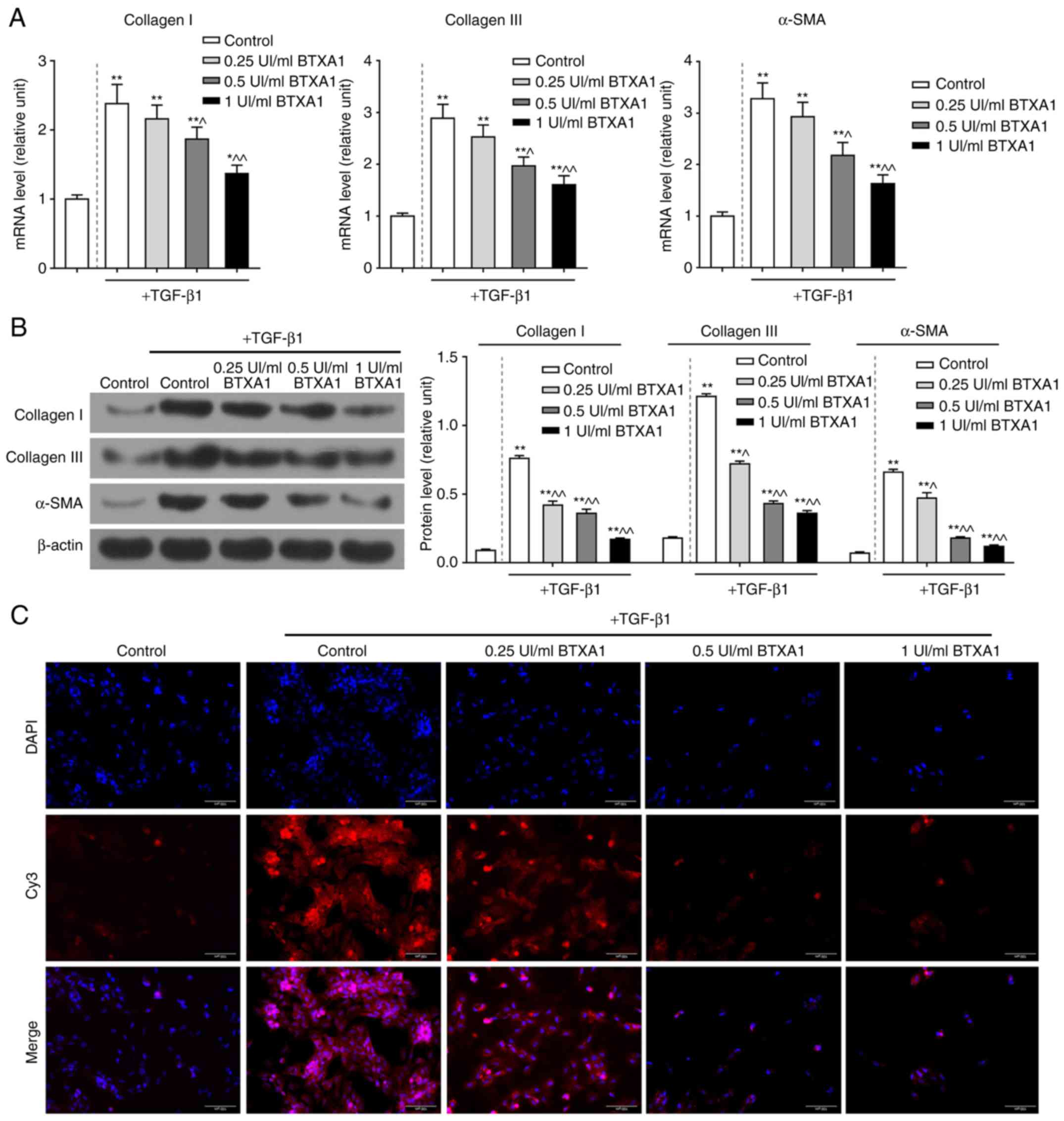
BTXA prevents over-deposition of ECM
components in fibroblasts
The mRNA expression levels of collagen I, collagen
III and α-SMA were notably enhanced by TGF-β1; however, the
increased mRNA levels of these molecules were evidently reduced by
treatment with gradient concentrations of BTXA (P<0.01; Fig. 3A). Similarly, BTXA suppressed the
high protein levels of collagen I, collagen III and α-SMA induced
by TGF-β1 (P<0.01; Fig. 3B).
Furthermore, α-SMA was found to be acutely expressed in fibroblasts
by treatment with TGF-β1; however, α-SMA expression was markedly
inhibited by treatment with medium and high concentrations of BTXA
(Fig. 3C).
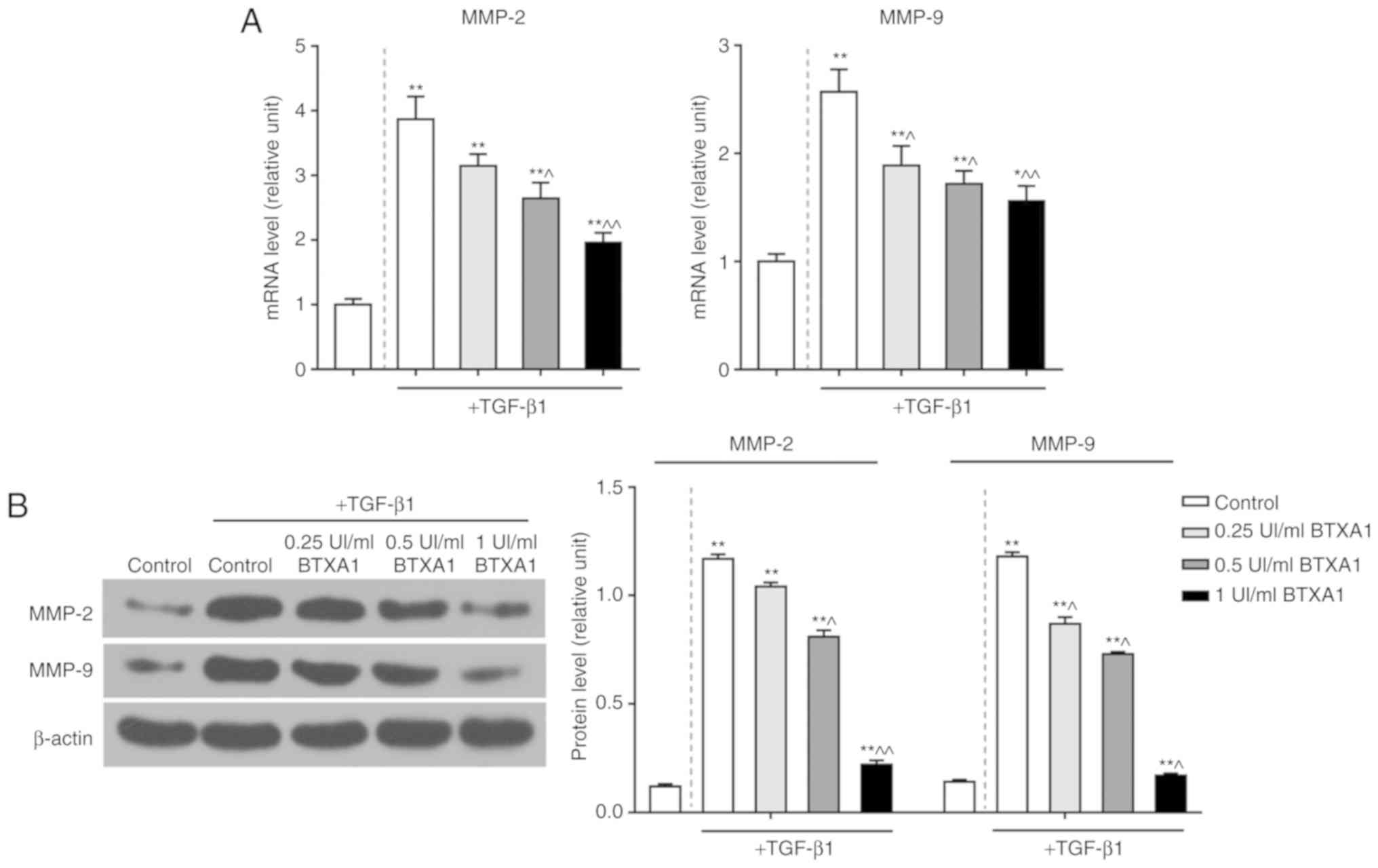
BTXA suppresses the expression of MMP-2
and MMP-9
To explore the potential role of BTXA in EMT, we
detected the levels of MMP-2 and MMP-9. TGF-β1 significantly
enhanced the mRNA expression levels of MMP-2 and MMP-9. However,
BTXA inhibited these expression levels in a dose-dependent manner
(P<0.01; Fig. 4A). Moreover,
the elevated protein levels of MMP-2 and MMP-9 induced by TGF-β1
were also notably suppressed by BTXA (P<0.01; Fig. 4B).
BTXA enhances PTEN expression and
inhibits apoptosis
The expression of PTEN was significantly reduced by
TGF-β1, while BTXA markedly increased PTEN expression at the
transcriptional and translational level (P<0.01; Fig. 5A and B). Additionally, the protein
level of cleaved caspase-3 was notably inhibited by TGF-β1,
whereas, BTXA markedly increased cleaved caspase-3 expression. The
expression of cleaved caspase-3 in the fibroblasts treated with a
high concentration of BTXA (1 UI/ml) did not differ significantly
from that in the untreated fibroblasts. Nevertheless, the
expression of pro-caspase-3 exhibited an opposite trend (P<0.01;
Fig. 5B).
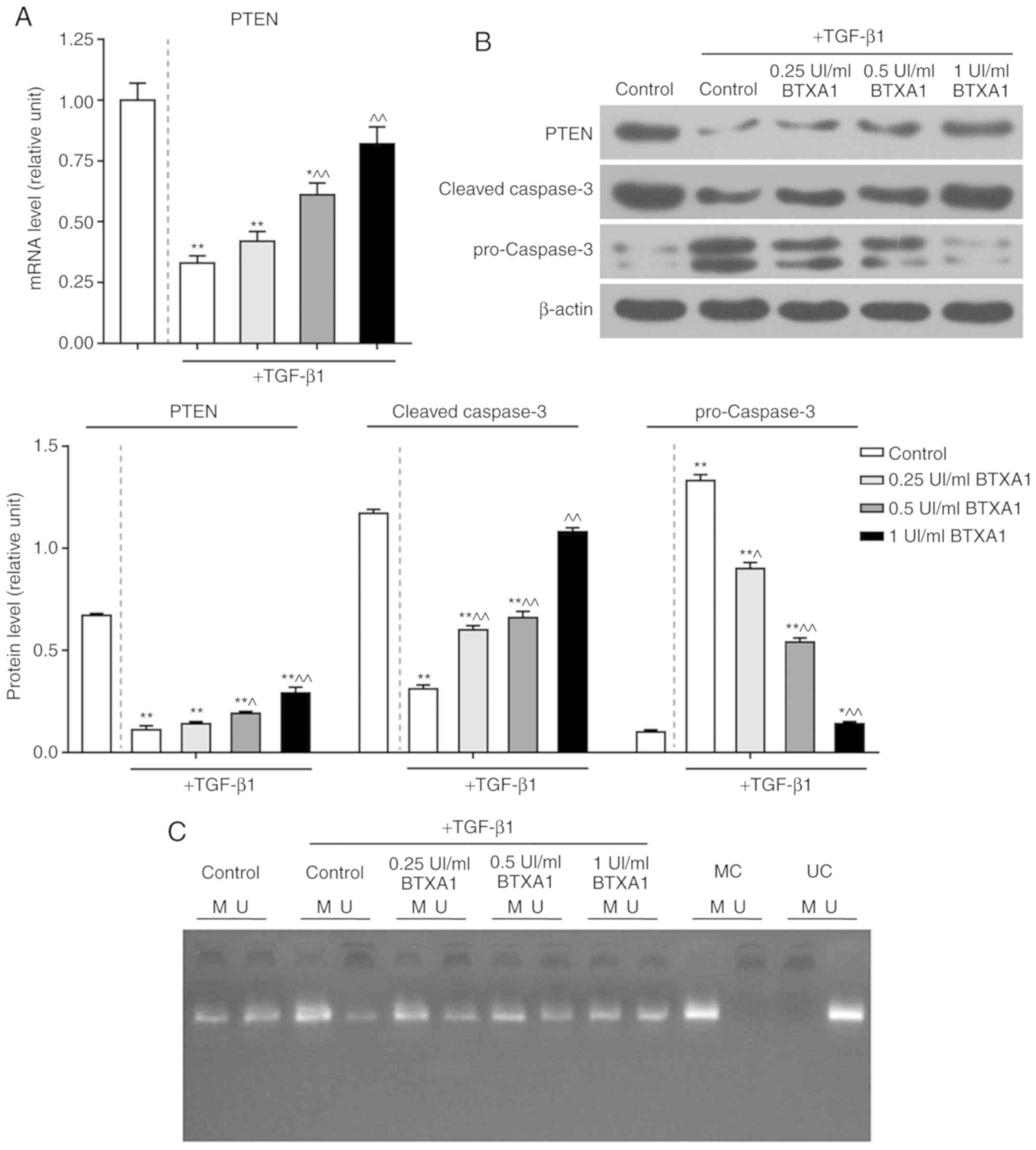 | Figure 5Effect of BTXA on PTEN and caspase-3
expression, as well as PTEN methylation. (A) Effect of treatment of
fibroblasts, which were cultured with or without 10 ng/ml of
TGF-β1, with various concentrations (0.25, 0.5 and 1 UI/ml) of BTXA
on the mRNA expression of PTEN, as determined by RT-qPCR. (B)
Effects of BTXA on the protein levels of PTEN, pro-caspase-3 and
cleaved-caspase-3, as detected by western blot analysis. (C) PTEN
methylation was measured by methylation-specific PCR (MSP). M,
methylated; U, unmethylated; MC, methylated control; UC,
unmethylated control. β-actin was used as an internal control.
Dotted line separation represents whether or not fibroblast were
treated with TGF-β1. Data are shown as the means ± SD, n=3.
*P<0.05 and **P<0.01 vs. control
without TGF-β1; ^P<0.05 and ^^P<0.01
vs. control with TGF-β1. BTXA, botulinum toxin type A; TGF-β1,
transforming growth factor-β1; PTEN, phosphatase and tensin homolog
deleted on chromosome ten. |
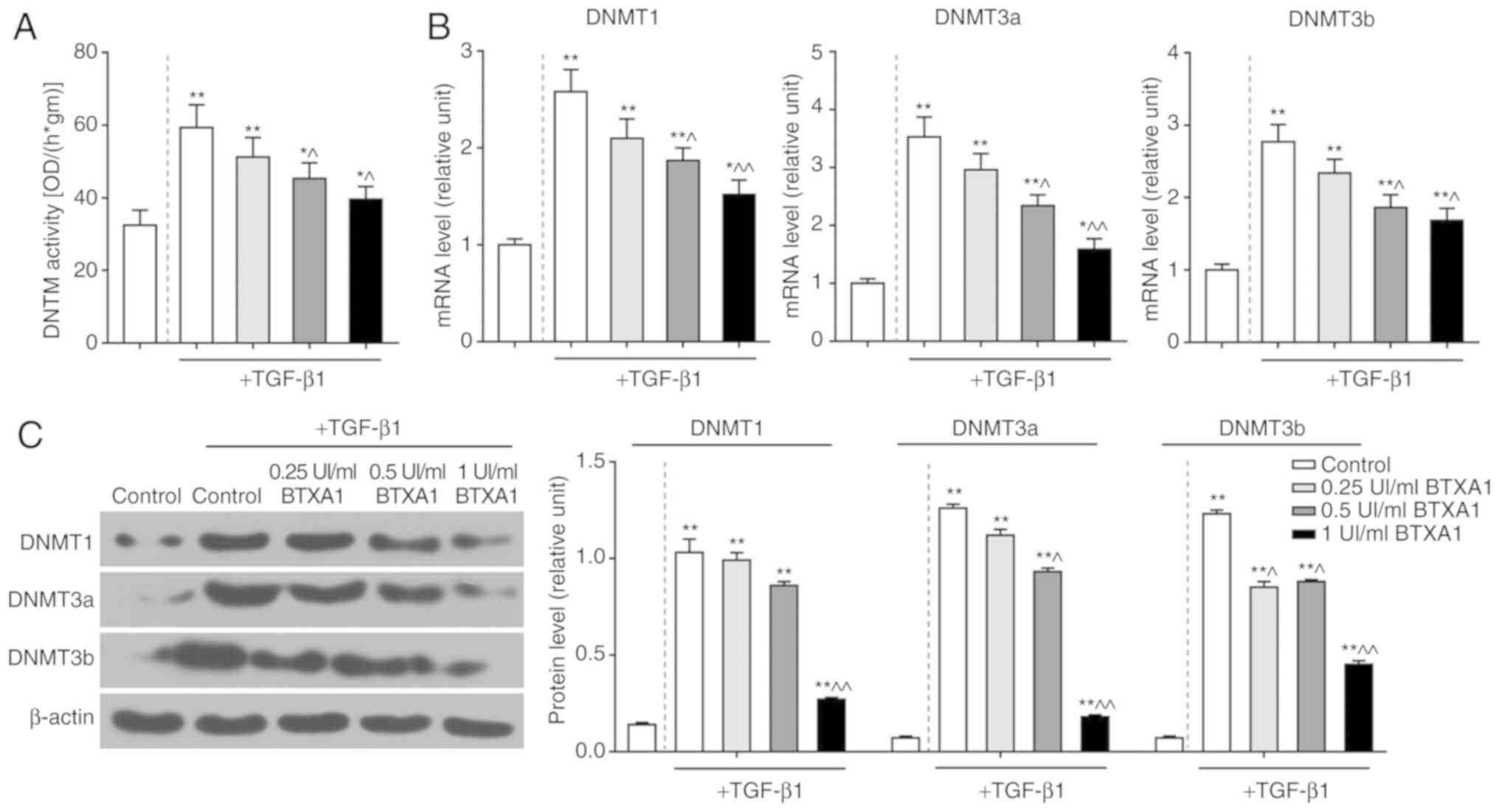
BTXA blocks PTEN methylation
To further elucidate the mechanisms of action of
BTXA in fibroblasts, we measured the methylation level of PTEN.
PTEN methylation was induced by TGF-β1, but prevented by BTXA
(Fig. 5C). Moreover, we detected
DNMT activity and determined the expression levels of several genes
related to DNA methylation. DNMT activity was intensified by
TGF-β1, and was attenuated by BTXA (P<0.05; Fig. 6A). Moreover, the enhanced mRNA and
protein levels of DNMT1, DNMT3a and DNMT3b induced by TGF-β1 were
evidently decreased by BTXA (P<0.05; Fig. 6B and C).
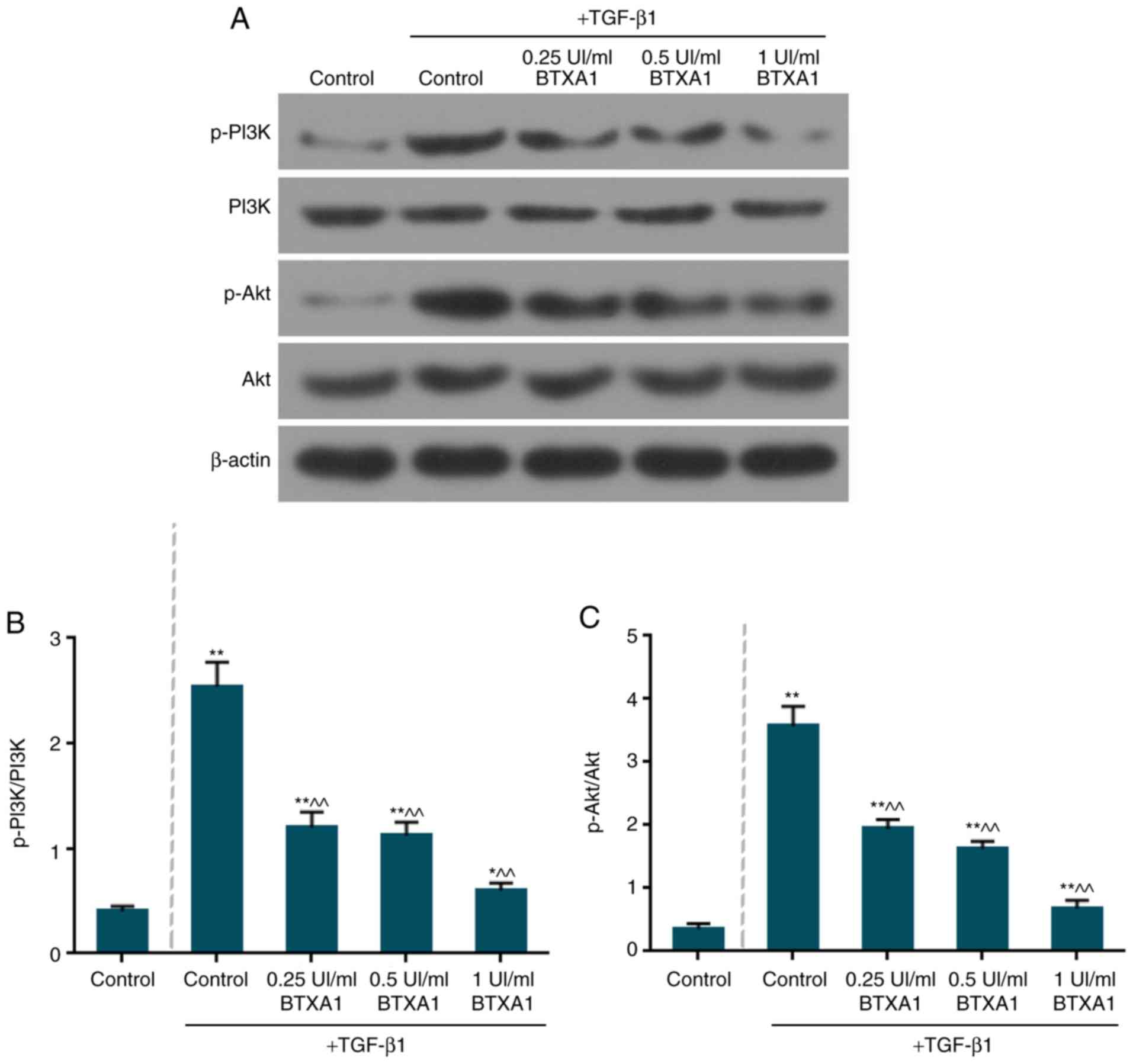
BTXA inactivates the phosphorylation of
PI3K and Akt
The expression levels of total PI3K and Akt remained
unaltered in the fibroblasts (Fig.
7A). However, the levels of phosphorylated (p)-PI3K/PI3K and
p-Akt/Akt were markedly increased by TGF-β1 (Fig. 7B and C). Moreover, BTXA notably
suppressed the ratio of p-PI3K/PI3K and p-Akt/Akt (P<0.01).
Discussion
BTXA has been widely applied in clinical therapies
including hyperhidrosis, spasticity, facial muscular hypertrophy
and muscular contraction in humans (43). Recently, studies have demonstrated
that BTXA is involved in the pathogenesis of dermal fibrosis. BTXA
can effectively prevent the differentiation of fibroblasts into
myofibroblasts by inhibiting α-SMA expression (19). Moreover, BTXA is able to notably
suppress the proliferation of fibroblasts in scar tissue by
inhibiting α-SMA and myosin II expression (25). BTXA has also been demonstrated to
attenuate HS by preventing collagen deposition (23,26). In this study, BTXA inhibited the
expression levels of ECM-related molecules and suppressed PTEN
methylation. We also found that PI3K/Akt was involved in the
regulatory mechanisms of BTXA in mouse fibroblasts.
The formation and remodeling of HS has been revealed
to be relevant to aberrant fibroblast proliferation and
differentiation (44). In this
study, we found that BTXA inhibited fibroblast viability in a
dose-dependent manner. Furthermore, TGF-β has been shown to be
associated with HS. For instance, a TGF-β1 inhibitor has been
demonstrated to alleviate scars and improve HS morphological
characteristics in xenograft mice with human HS (45). The antagonist peptide of TGF-β has
been reported to improve the fibrotic behaviors of human
fibroblasts derived from HS (46). In this study, TGF-β1 enhanced
fibroblast viability and inhibited apoptosis. However, BTXA
partially reversed the effects of TGF-β1 on fibroblasts.
The accumulated fibroblasts and myofibroblast caused
by proliferation, activation and differentiation can induce massive
collagen deposition, resulting in ECM deposition below the derma.
These activated fibroblasts can be identified by an increased
expression of α-SMA (43). α-SMA
expression is relevant to the phenotypic transformation of
fibroblasts into myofibroblasts during the wound healing process
(44). The decreased ratio of
collagen I/collagen III has been found in human HS (47). Furthermore, collagen I expression
in human HS-derived fibroblasts has been shown to be significantly
suppressed by the knockdown of TGF-β receptor I and wound scars
were also shown to be decreased in rabbits (48). In this study, the elevated
expression levels of collagen I, collagen III and α-SMA induced by
TGF-β1 were suppressed by BTXA. Additionally, fibroblasts have been
reported to degrade fibrin clots by generating MMPs and their
inhibitors (TIMPs), leading to a disorder of matrix formation and
degradation (49). The enhanced
expression levels of MMP-2 and MMP-9 have been shown to be
associated with decreased levels of collagen I and collagen III in
HS tissues (50). In this study,
we found that TGF-β1 significantly increased the MMP-2 and MMP-9
expression at the transcriptional and translational levels.
However, BTXA treatment markedly altered these trends.
PTEN has been reported to inhibit the proliferation
and functions of HS fibroblasts (37). A recent study revealed that PTEN
overexpression was involved in the inhibition of glial scar
formation (51). In this study,
we also found that PTEN expression was inhibited by TGF-β1 and was
upregulated by BTXA in fibroblasts, indicating that BTXA may
suppress HS formation in vitro. Moreover, PTEN is also a key
regulator of apoptosis (52). The
apoptosis is detected based on the activation of caspase-3, which
occurs in apoptotic death (53).
In this study, we found that the pro-caspase-3 expression was
elevated, while the cleaved-caspase-3 level was decreased by
TGF-β1. BTXA reversed this expression pattern. These results
suggest that BTXA promotes fibroblast apoptosis.
Abnormal promoter methylation often accounts for the
transcriptional inactivation of various molecules associated with
apoptosis and tumor suppression (54). DNMTs can catalyze the methylation
of CpG dinucleotides, which are involved in DNA methylation. It has
been demonstrated that the decreased expression of PTEN is
accompanied by an enhanced level of DNMT1, which is associated with
PTEN promoter hypermethylation (55). Moreover, DNMT3a and DNMT3b can
methylate non-methylated DNA as de novo methyltransferases
(56). In this study, the high
DNMT activity induced by TGF-β1 was significantly reduced by BTXA.
Additionally, the levels of DNMT1, DNMT3a and DNMT3b were also
inhibited by BTXA, suggesting that BTXA may prevent PTEN
methylation.
The PI3K/AKT pathway has been reported to be
negatively regulated by PTEN (57). The reduced expression of PTEN has
been reported to activate the PTEN/AKT pathway and to be associated
with the pathogenesis of HS (37). The activation of the PI3K/AKT
pathway has been demonstrated to promote cell viability and inhibit
apoptosis (58). The abnormal
PI3K/AKT pathway activation may result in multiple diseases,
including HS (59). Moreover, the
activated PI3K/AKT pathway can enhance the accumulation of dermal
fibroblasts (60). In this study,
we found that the expression levels of p-PI3K and p-Akt were
significantly increased by TGF-β1. However, BTXA can inhibit the
phosphorylation of the PI3K and Akt, and this expression pattern is
opposite to PTEN expression, indirectly suggesting that the loss of
PTEN can activate the PI3K/AKT pathway (61).
In this study, there were still some limitations;
for example, the lack of fibroblasts derived from HS in animal
models or derived from skin, the absence of one more cell line and
the lack of verification experiments with the application of the
corresponding PTEN/PI3K/Akt pathway inhibitor. Therefore, further
studies on the effects of BTXA on scars of other nature, as well as
studies using animal models treated with BTXA, and one more cell
line or skin-derived cell line with validation experiments
including a signaling inhibitor are required in order to validate
our findings.
In conclusion, the findings of this study
demonstrated that BTXA inhibited the viability and promoted the
apoptosis of fibroblasts induced by TGF-β1. The enhanced expression
levels of molecules associated with ECM and EMT, which were induced
by TGF-β1, were suppressed by BTXA. Furthermore, PTEN methylation
triggered by TGF-β1 was prevented by BTXA and the activities of
DNMTs were also suppressed. Moreover, BTXA blocked the
phosphorylation of PI3K and Akt. The findings of this study provide
a molecular basis for the role of BTXA in the fibroblast phenotypic
transformation and a theoretical basis for HS treatment.
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request.
Authors' contributions
XZ and DL made substantial contributions to the
conception and design of the study. SN and SY were involved in data
acquisition, data analysis and interpretation. HJ and SY performed
the experiments, drafted the article or critically revised it for
important intellectual content. All authors have read and approved
the final manuscript and agree to be accountable for all aspects of
the work in ensuring that questions related to the accuracy or
integrity of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Xiao Z, Zhang F, Lin W, Zhang M and Liu Y:
Effect of botulinum toxin type A on transforming growth factor
beta1 in fibroblasts derived from hypertrophic scar: A preliminary
report. Aesthetic Plast Surg. 34:424–427. 2010. View Article : Google Scholar
|
|
2
|
Butzelaar L, Ulrich MM, Mink van der Molen
AB, Niessen FB and Beelen RH: Currently known risk factors for
hypertrophic skin scarring: A review. J Plast Reconstr Aesthet
Surg. 69:163–169. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xue M and Jackson CJ: Extracellular matrix
reorganization during wound healing and its impact on abnormal
scarring. Adv Wound Care (New Rochelle). 4:119–136. 2015.
View Article : Google Scholar
|
|
4
|
Liu J, Wang Y, Pan Q, Su Y, Zhang Z, Han
J, Zhu X, Tang C and Hu D: Wnt/β-catenin pathway forms a negative
feedback loop during TGF- β1 induced human normal skin
fibroblast-to-myofibroblast transition. J Dermatol Sci. 65:38–49.
2012. View Article : Google Scholar
|
|
5
|
Chun Q, ZhiYong W, Fei S and XiQiao W:
Dynamic biological changes in fibroblasts during hypertrophic scar
formation and regression. Int Wound J. 13:257–262. 2016. View Article : Google Scholar
|
|
6
|
Sarrazy V, Billet F, Micallef L, Coulomb B
and Desmoulière A: Mechanisms of pathological scarring: Role of
myofibroblasts and current developments. Wound Repair Regen.
19(Suppl 1): s10–s15. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Curran TA and Ghahary A: Evidence of a
role for fibrocyte and keratinocyte-like cells in the formation of
hypertrophic scars. J Burn Care Res. 34:227–231. 2013. View Article : Google Scholar
|
|
8
|
Ding J, Ma Z, Shankowsky HA, Medina A and
Tredget EE: Deep dermal fibroblast profibrotic characteristics are
enhanced by bone marrow-derived mesenchymal stem cells. Wound
Repair Regen. 21:448–455. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gökçinar-Yagci B, Uçkan-Çetinkaya D and
Çelebi-Saltik B: Pericytes: Properties, functions and applications
in tissue engineering. Stem Cell Rev. 11:549–559. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yan C, Grimm WA, Garner WL, Qin L, Travis
T, Tan N and Han YP: Epithelial to mesenchymal transition in human
skin wound healing is induced by tumor necrosis factor-alpha
through bone morphogenic protein-2. Am J Pathol. 176:2247–2258.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang X, Chu J, Wen CJ, Fu SB, Qian YL, Wo
Y, Wang C and Wang DR: Functional characterization of TRAP1-like
protein involved in modulating fibrotic processes mediated by
TGF-β/Smad signaling in hypertrophic scar fibroblasts. Exp Cell
Res. 332:202–211. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lu L, Saulis AS, Liu WR, Roy NK, Chao JD,
Ledbetter S and Mustoe TA: The temporal effects of anti-TGF-beta1,
2, and 3 monoclonal antibody on wound healing and hypertrophic scar
formation. J Am Coll Surg. 201:391–397. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pakyari M, Farrokhi A, Maharlooei MK and
Ghahary A: Critical role of transforming growth factor beta in
different phases of wound healing. Adv Wound Care (New Rochelle).
2:215–224. 2013. View Article : Google Scholar
|
|
14
|
Yin L, Zhao X, Ji S, He C, Wang G, Tang C,
Gu S and Yin C: The use of gene activated matrix to mediate
effective SMAD2 gene silencing against hypertrophic scar.
Biomaterials. 35:2488–2498. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang YF, Zhou SZ, Cheng XY, Yi B, Shan
SZ, Wang J and Li QF: Baicalein attenuates hypertrophic scar
formation via inhibition of the transforming growth
factor-β/Smad2/3 signalling pathway. Br J Dermatol. 174:120–130.
2016. View Article : Google Scholar
|
|
16
|
Bai X, He T, Liu J, Wang Y, Fan L, Tao K,
Shi J, Tang C, Su L and Hu D: Loureirin B inhibits fibroblast
proliferation and extracellular matrix deposition in hypertrophic
scar via TGF-β/Smad pathway. Exp Dermatol. 24:355–360. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zunwen L, Shizhen Z, Dewu L, Yungui M and
Pu N: Effect of tetrandrine on the TGF-β-induced smad signal
transduction pathway in human hypertrophic scar fibroblasts in
vitro. Burns. 38:404–413. 2012. View Article : Google Scholar
|
|
18
|
Omranifard M, Heidari M, Farajzadegan Z,
Niktabar MR and Motamedi N: Botulinum toxin and burn induces
contracture. Arch Plast Surg. 43:609–611. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jeong HS, Lee BH, Sung HM, Park SY, Ahn
DK, Jung MS and Suh IS: Effect of botulinum toxin type A on
differentiation of fibroblasts derived from scar tissue. Plast
Reconstr Surg. 136:171e–178e. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gauglitz GG, Bureik D, Dombrowski Y,
Pavicic T, Ruzicka T and Schauber J: Botulinum toxin A for the
treatment of keloids. Skin Pharmacol Physiol. 25:313–318. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rasaii S, Sohrabian N, Gianfaldoni S,
Hadibarhaghtalab M, Pazyar N, Bakhshaeekia A, Lotti T,
Ramirez-Pacheco LA, Lange CS, Matta J, et al: Intralesional
triamcinolone alone or in combination with botulinium toxin A is
ineffective for the treatment of formed keloid scar: A double blind
controlled pilot study. Dermatol Ther. 32:e127812019. View Article : Google Scholar
|
|
22
|
Xiao Z, Zhang F and Cui Z: Treatment of
hypertrophic scars with intralesional botulinum toxin type A
injections: A preliminary report. Aesthetic Plast Surg. 33:409–412.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xiao Z and Qu G: Effects of botulinum
toxin type a on collagen deposition in hypertrophic scars.
Molecules. 17:2169–2177. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee SD, Yi MH, Kim DW, Lee Y, Choi Y and
Oh SH: The effect of botulinum neurotoxin type A on capsule
formation around silicone implants: The in vivo and in vitro study.
Int Wound J. 13:65–71. 2016. View Article : Google Scholar
|
|
25
|
Chen M, Yan T, Ma K, Lai L, Liu C, Liang L
and Fu X: Botulinum toxin type A inhibits α-smooth muscle actin and
myosin II expression in fibroblasts derived from scar contracture.
Ann Plast Surg. 77:e46–e49. 2016. View Article : Google Scholar
|
|
26
|
Xiao Z, Zhang M, Liu Y and Ren L:
Botulinum toxin type a inhibits connective tissue growth factor
expression in fibroblasts derived from hypertrophic scar. Aesthetic
Plast Surg. 35:802–807. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kim S, Ahn M, Piao Y, Ha Y, Choi DK, Yi
MH, Shin N, Kim DW and Oh SH: Effect of botulinum toxin type A on
TGF-beta/Smad pathway signaling: Implications for silicone-induced
capsule formation. Plast Reconstr Surg. 138:821e–829e. 2016.
View Article : Google Scholar
|
|
28
|
Li Y, Zhang J, Zhou Q, Wang H, Xie S, Yang
X, Ji P, Zhang W, He T, Liu Y, et al: Linagliptin inhibits high
glucose-induced transdifferentiation of hypertrophic scar-derived
fibroblasts to myofibroblasts via IGF/Akt/mTOR signalling pathway.
Exp Dermatol. 28:19–27. 2019. View Article : Google Scholar
|
|
29
|
Liu B, Guo Z and Gao W: miR-181b-5p
promotes proliferation and inhibits apoptosis of hypertrophic scar
fibroblasts through regulating the MEK/ERK/p21 pathway. Exp Ther
Med. 17:1537–1544. 2019.PubMed/NCBI
|
|
30
|
Zhang Q, Guo B, Hui Q, Chang P and Tao K:
miR-137 inhibits proliferation and metastasis of hypertrophic scar
fibroblasts via targeting pleiotrophin. Cell Physiol Biochem.
49:985–995. 2018. View Article : Google Scholar
|
|
31
|
Lim CP, Phan TT, Lim IJ and Cao X: Stat3
contributes to keloid pathogenesis via promoting collagen
production, cell proliferation and migration. Oncogene.
25:5416–5425. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liao WT, Yu HS, Arbiser JL, Hong CH,
Govindarajan B, Chai CY, Shan WJ, Lin YF, Chen GS and Lee CH:
Enhanced MCP-1 release by keloid CD14+ cells augments fibroblast
proliferation: Role of MCP-1 and Akt pathway in keloids. Exp
Dermatol. 19:e142–e150. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kulkarni AA, Thatcher TH, Olsen KC,
Maggirwar SB, Phipps RP and Sime PJ: PPAR-γ ligands repress
TGFβ-induced myofibroblast differentiation by targeting the
PI3K/Akt pathway: Implications for therapy of fibrosis. PLoS One.
6:e159092011. View Article : Google Scholar
|
|
34
|
Paterno J, Vial IN, Wong VW, Rustad KC,
Sorkin M, Shi Y, Bhatt KA, Thangarajah H, Glotzbach JP and Gurtner
GC: Akt-mediated mechanotransduction in murine fibroblasts during
hypertrophic scar formation. Wound Repair Regen. 19:49–58. 2011.
View Article : Google Scholar
|
|
35
|
Dey JH, Bianchi F, Voshol J, Bonenfant D,
Oakeley EJ and Hynes NE: Targeting fibroblast growth factor
receptors blocks PI3K/AKT signaling, induces apoptosis, and impairs
mammary tumor outgrowth and metastasis. Cancer Res. 70:4151–4162.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Deng B, Yang X, Liu J, He F, Zhu Z and
Zhang C: Focal adhesion kinase mediates TGF-beta1-induced renal
tubular epithelial-to-mesenchymal transition in vitro. Mol Cell
Biochem. 340:21–29. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Guo L, Chen L, Bi S, Chai L, Wang Z, Cao
C, Tao L and Li S: PTEN inhibits proliferation and functions of
hypertrophic scar fibroblasts. Mol Cell Biochem. 361:161–168. 2012.
View Article : Google Scholar
|
|
38
|
Takashima M, Parsons CJ, Ikejima K,
Watanabe S, White ES and Rippe RA: The tumor suppressor protein
PTEN inhibits rat hepatic stellate cell activation. J
Gastroenterol. 44:847–855. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
White ES, Thannickal VJ, Carskadon SL,
Dickie EG, Livant DL, Markwart S, Toews GB and Arenberg DA:
Integrin alpha4beta1 regulates migration across basement membranes
by lung fibroblasts: A role for phosphatase and tensin homologue
deleted on chromosome 10. Am J Respir Crit Care Med. 168:436–442.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Larsson O, Diebold D, Fan D, Peterson M,
Nho RS, Bitterman PB and Henke CA: Fibrotic myofibroblasts manifest
genome-wide derangements of translational control. PLoS One.
3:e32202008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mori T, Okumura M, Matsuura M, Ueno K,
Tokura S, Okamoto Y, Minami S and Fujinaga T: Effects of chitin and
its derivatives on the proliferation and cytokine production of
fibroblasts in vitro. Biomaterials. 18:947–951. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
43
|
Jiang B, Zu W, Xu J, Xiong Z, Zhang Y, Gao
S, Ge S and Zhang L: Botulinum toxin type A relieves
sternocleidomastoid muscle fibrosis in congenital muscular
torticollis. Int J Biol Macromol. 112:1014–1020. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhao JC, Zhang BR, Hong L, Shi K, Wu WW
and Yu JA: Extracorporeal shock wave therapy with low-energy flux
density inhibits hypertrophic scar formation in an animal model.
Int J Mol Med. 41:1931–1938. 2018.PubMed/NCBI
|
|
45
|
Qiu SS, Dotor J and Hontanilla B: Effect
of P144® (Anti-TGF-β) in an 'In Vivo' human hypertrophic scar model
in nude mice. PLoS One. 10:e01444892015. View Article : Google Scholar
|
|
46
|
Wang X, Gao Z, Wu X, Zhang W, Zhou G and
Liu W: Inhibitory effect of TGF-β peptide antagonist on the
fibrotic phenotype of human hypertrophic scar fibroblasts. Pharm
Biol. 54:1189–1197. 2016.
|
|
47
|
Oliveira GV, Hawkins HK, Chinkes D, Burke
A, Tavares AL, Ramos-e-Silva M, Albrecht TB, Kitten GT and Herndon
DN: Hypertrophic versus non hypertrophic scars compared by
immu-nohistochemistry and laser confocal microscopy: Type I and III
collagens. Int Wound J. 6:445–452. 2009. View Article : Google Scholar
|
|
48
|
Wang YW, Liou NH, Cherng JH, Chang SJ, Ma
KH, Fu E, Liu JC and Dai NT: siRNA-targeting transforming growth
factor-β type I receptor reduces wound scarring and extracellular
matrix deposition of scar tissue. J Invest Dermatol. 134:2016–2025.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ulrich D, Ulrich F, Unglaub F, Piatkowski
A and Pallua N: Matrix metalloproteinases and tissue inhibitors of
metalloproteinases in patients with different types of scars and
keloids. J Plast Reconstr Aesthet Surg. 63:1015–1021. 2010.
View Article : Google Scholar
|
|
50
|
Huang D, Liu Y, Huang Y, Xie Y, Shen K,
Zhang D and Mou Y: Mechanical compression upregulates MMP9 through
SMAD3 but not SMAD2 modulation in hypertrophic scar fibroblasts.
Connect Tissue Res. 55:391–396. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Luan Y, Chen M and Zhou L: MiR-17 targets
PTEN and facilitates glial scar formation after spinal cord
injuries via the PI3K/Akt/mTOR pathway. Brain Res Bull. 128:68–75.
2017. View Article : Google Scholar
|
|
52
|
Pi WF, Guo XJ, Su LP and Xu WG:
Troglitazone upregulates PTEN expression and induces the apoptosis
of pulmonary artery smooth muscle cells under hypoxic conditions.
Int J Mol Med. 32:1101–1109. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yan YT, Li SD, Li C, Xiong YX, Lu XH, Zhou
XF, Yang LQ, Pu LJ and Luo HY: Panax notoginsenoside saponins Rb1
regulates the expressions of Akt mTOR/PTEN signals in the
hippocampus after focal cerebral ischemia in rats. Behav Brain Res.
345:83–92. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Sanders YY, Pardo A, Selman M, Nuovo GJ,
Tollefsbol TO, Siegal GP and Hagood JS: Thy-1 promoter
hypermethylation: A novel epigenetic pathogenic mechanism in
pulmonary fibrosis. Am J Respir Cell Mol Biol. 39:610–618. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Bian EB, Huang C, Ma TT, Tao H, Zhang H,
Cheng C, Lv XW and Li J: DNMT1-mediated PTEN hypermethylation
confers hepatic stellate cell activation and liver fibrogenesis in
rats. Toxicol Appl Pharmacol. 264:13–22. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang X, Wang Z, Wang Q, Wang H, Liang H
and Liu D: Epigenetic modification differences between fetal
fibroblast cells and mesenchymal stem cells of the Arbas Cashmere
goat. Res Vet Sci. 114:363–369. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhu G, Chai J, Ma L, Duan H and Zhang H:
Downregulated microRNA-32 expression induced by high glucose
inhibits cell cycle progression via PTEN upregulation and Akt
inactivation in bone marrow-derived mesenchymal stem cells. Biochem
Biophys Res Commun. 433:526–531. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kitagishi Y and Matsuda S: Diets involved
in PPAR and PI3K/AKT/PTEN pathway may contribute to neuroprotection
in a traumatic brain injury. Alzheimers Res Ther. 5:422013.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang Y, Yao X, Jiang C, Yue J, Guan J,
Cheng H, Hajirashid M, Wang Y and Fan L: Expression of PI3K, PTEN
and Akt in small intestinal adenocarcinoma detected by quantum
dots-based immunofluorescence technology. Cancer Biomark.
13:299–305. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Pericacho M, Velasco S, Prieto M, Llano E,
López-Novoa JM and Rodríguez-Barbero A: Endoglin haploinsufficiency
promotes fibroblast accumulation during wound healing through Akt
activation. PLoS One. 8:e546872013. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Seront E, Pinto A, Bouzin C, Bertrand L,
Machiels JP and Feron O: PTEN deficiency is associated with reduced
sensitivity to mTOR inhibitor in human bladder cancer through the
unhampered feedback loop driving PI3K/Akt activation. Br J Cancer.
109:1586–1592. 2013. View Article : Google Scholar : PubMed/NCBI
|