Introduction
Bone quality is maintained by the balance between
osteoclastic bone resorption and osteoblastic bone formation. The
disruption of this balance leads to bone metabolic diseases, such
as osteoporosis, arthritis, periodontitis and Paget's disease
(1). Osteoporosis is a metabolic
bone disease characterized by low bone mass, whereby the
microstructure of the bone tissue is destroyed, increasing the
probability of fractures. In recent years, an increase in the
elderly population has led to a high incidence of osteoporosis,
which is a serious health concern (2). In order to treat osteoporosis, bone
resorption inhibitors, such as bisphosphonates and denosumab have
been developed (3). However,
patients treated with these inhibitors experience severe
side-effects, including uterine cancer, breast cancer and
osteonecrosis (4,5). In addition, as patients with
osteoporosis usually present with significant bone loss, promoting
osteoblast activity is vital. For this reason, it is necessary to
identify natural products that inhibit osteoclast differentiation
and promote osteoblast differentiation.
Osteoblasts are derived from mesenchymal stem cells
and are located in regions of bone regeneration. Bone morphogenetic
protein 2 (BMP-2) and runt-related transcription factor 2 (RUNX2)
signaling pathways are known to be important in mediating
osteoblast differentiation and activation (6). Activated RUNX2 regulates the
expression of osteogenic genes, including alkaline phosphatase
(ALP), osteonectin (OSN), osteopontin (OPN), type I collagen (COL1)
and bone sialoprotein (BSP), which leads to the successful
differentiation of osteoblasts (7). Receptor activator of nuclear factor
κ-B ligand (RANKL) is a tumor necrosis factor (TNF)-associated,
activation-induced cytokine expressed in osteoblasts that is
essential for osteoclastogenesis (8). RANKL binds to RANK on the surface of
osteoclast precursor cells, and RANK then stimulates
mitogen-activated protein kinases (MAPKs), nuclear factor (NF)-κB
and c-Fos to induce nuclear factor of activated T cells 1 (NFATc1)
(9). NFATc1 is essential for
osteoclast differentiation and regulates the expression of various
osteoclast-associated genes, such as tartrate-resistant acid
phosphatase (TRAP), matrix metallopeptidase-9 (MMP-9), cathepsin K
(Ctsk), osteoclast-associated immunoglobulin-like receptor (OSCAR),
c-src, c-myc, osteoclast stimulatory trans-membrane (OC-STAMP) and
ATPase H+ transporting V0 subunit d2 (ATP6v0d2) (10).
Leonurus sibiricus L. (LS), also known as
motherwort, is currently used as a medicinal plant in various
regions, such as East Asia, Europe and the USA. LS is traditionally
used in the treatment of a variety of female-related conditions,
including menstrual pain, dysmenorrhea and amenorrhea. For this
reason, it is known as 'IG-MO-CHO' in Korean, which translates into
'herb that is beneficial to mother' (11). In addition, LS is used for the
treatment of renal diseases, such as acute nephritis and kidney
stones (12). In oriental
medicine, the kidney is considered to be the key organ of bone
control (13). Therefore,
kidney-enhancing medications, such as LS, have been used for the
treatment of weakened bones. Previous studies have demonstrated
that LS exerts numerous biological activities, such as
anti-inflammatory, antioxidant (14), anti-bacterial (15) and angiogenic effects (16). Among these, the anti-inflammatory
and antioxidant effects have been demonstrated to be associated
with the treatment of bone metabolic disease (17). Therefore, the authors of the
present study hypothesized that LS may exert a positive effect on
bone metabolism. However, to the best of our knowledge, no studies
published to date have investigated the effects of LS on osteoblast
differentiation and osteoclastogenesis.
In the present study, the effects of LS on
osteoblast differentiation and osteoclast differentiation, which
play a major role in bone metabolism, were investigated. The
effects of LS in an in vivo model of lipopolysaccharide
(LPS)-induced osteoporosis were also examined.
Materials and methods
Reagents
Minimum essential Eagle's medium, α-modification
(α-MEM), fetal bovine serum (FBS) and penicillin/streptomycin (P/S)
were supplied by Gibco; Thermo Fisher Scientific, Inc. Dulbecco's
modified Eagle's medium (DMEM) was procured from Welgene, Inc.
Alizarin Red S was obtained from Duksan Co., Ltd. RANKL was
purchased from Peprotech, Inc. Rutin, dimethylsulfoxide (DMSO) and
the TRAP assay kit were purchased from Sigma-Aldrich; Merck KGaA.
Osteo strip well plates were purchased from Corning Inc.
Anti-RUNX2, anti-BMP-2, anti-Ctsk and anti-MMP-9 antibodies were
purchased from Abcam. Anti-phosphorylated (p)-extracellular
signal-regulated kinase 1/2 (ERK1/2), anti-ERK1/2, anti-p-c-Jun
N-terminal kinase (JNK), anti-JNK, anti-p-p38, anti-p38, anti-NF-κB
and anti-p-NF-κB antibodies were supplied by Cell Signaling
Technology, Inc. Anti-NFATc1 was procured from BD Biosciences, and
anti-c-Fos, anti-actin and anti-lamin B antibodies were purchased
from Santa Cruz Biotechnology, Inc. Secondary antibodies were
procured from Jackson ImmunoResearch Laboratories, Inc. The reverse
transcriptase kit and SYBR-Green solution was supplied by
Invitrogen; Thermo Fisher Scientific, Inc. Taq polymerase was
purchased from Kapa Biosystems; Roche Diagnostics. PCR primers were
purchased from Genotech Corp. All of the chemicals used in the
experiments were analytical grade for cell culture.
Preparation of LS
LS was purchased from Omniherb and was certified by
Professor Yungmin Bu at the Herbology Laboratory, College of Korean
Medicine. Voucher specimens of the plants used in the current study
were stored at the Department of Anatomy Herbarium (ref. no.
KHU-ANA-Et010). LS was soaked in 80% ethanol for 1 week before the
extract was filtered through filter paper, and then concentrated
under reduced pressure and lyophilized to obtain a powder (yield,
7.83%). The extracts were stored at -20°C until required. Prior to
use in the in vitro experiments, LS was filtered through a sterile
filter (pore size, 0.22 µm), diluted in DMSO and not treated
with >0.1% of the total volume of cell culture medium.
High-performance liquid chromatography
(HPLC) analysis
To quantitatively evaluate the LS extract, HPLC was
performed using rutin; a reference compound of LS. Rutin (standard
stock solution, 1,000 µg/ml) were prepared in methanol. LS
and rutin were analyzed using an A Waters 2695 system equipped with
a Waters 2487 Dual λ absorbance detector. Separation was achieved
using a C18 guard column with Xbridge-C18 (250×6 mm, 5 µm).
All solvents were filtered through a 45-µm filter and the
elution time was 0-30 min. The binary mobile phase was acetonitrile
(A) and water (B) at a composition of 10% A from 0 to 10 min and
50% A from 10 to 30 min. The flow rate was 1.0 ml/min and rutin was
detected at 254 nm.
Cell culture and assessment of cell
viability
C57BL/6 mouse calvaria MC3T3-E1 cells were purchased
from the American Type Culture Collection (ATCC), and the RAW 264.7
murine macrophage cell line was purchased from the Korean Cell Line
Bank. The MC3T3-E1 cells were cultured in α-MEM (without ascorbic
acid) supplemented with 10% FBS and 1% P/S, and RAW 264.7 cells
were cultured in DMEM supplemented with 10% FBS and 1% P/S. Cell
lines were maintained in a cell incubator at 37°C and 5%
CO2. To analyze cytotoxicity levels, MC3T3-E1 cells
(1×104 cells/well) treated with or without LS for 3, 7
and 14 days, or RAW 264.7 cells (5×103 cells/well)
treated with or without LS and RANKL for 1 and 5 days, were seeded
in 96-well-plates and then treated with MTS solution for 2 h. The
absorbance was subsequently measured using an enzyme-linked
immunosorbent assay (ELISA) reader (Versamax; Molecular Devices,
LLC) at a wavelength of 490 nm.
Von Kossa and Alizarin Red S
staining
The MC3T3-E1 cells were seeded in 6-well-plates at
5×104 cells/well and incubated in differentiation medium
consisting of α-MEM, 25 µg/ml ascorbic acid and 10 mM
β-glycerophosphate with or without LS for 21 days. The culture
medium was refreshed every 2 days. For Von Kossa staining, the
nodules were first fixed in 80% Et-OH and washed 3 times with
deionized water The fixed nodules were incubated at room
temperature with 1% sliver nitrate for 40 min under ultraviolet
light prior to incubation with 5% sodium thiosulfate for 10 min.
The dried plate was imaged using a camera. For Alizarin Red S
staining, formed mineralized nodules were fixed with 80% Et-OH and
then stained with Alizarin Red S solution at room temperature for 3
min. The stained nodules were washed 3 times with Dulbecco's
phosphate-buffered saline (Gibco; Thermo Fisher Scientific, Inc.).
The formed nodules were subsequently photographed with a camera
before the dye was extracted by the addition of 10 mM sodium
phosphate (pH 7.0) diluted in 10% cetylpyridinium chloride for 15
min. The extracted dyes were measured measured using an ELISA
reader at an absor-bance of 570 nm.
Western blot analysis
The cells were lysed using radioimmunoprecipitation
assay buffer (50 mM Tris-Cl, 150 mM NaCl, 1% NP-40, 0.5% sodium
deoxycholate and 0.1% SDS) with protease inhibitor and phosphatase
inhibitor 2 and 3 cocktails (Sigma Aldrich; Merck KGaA). Protein
concentrations were determined using a bicinchoninic acid assay kit
using bovine serum albumin as the standard. An equivalent quantity
of protein for each sample (10-30 µg) was separated by 10%
SDS-PAGE and transferred to nitrocellulose membranes via
electrophoresis. The membranes were blocked with 5% skim milk and
incubated with specific primary antibodies against RUNX2 (cat. no.
ab76956; dilution, 1:1,000), BMP-2 (cat. no. ab14933; dilution,
1:1,000), β-actin (cat. no. sc-8432; dilution, 1:500), p-ERK (cat.
no. 4370S; dilution, 1:1,000), ERK (cat. no. 4695S; dilution,
1:1,000), p-JNK (cat. no. 4668S; dilution, 1:1,000), JNK (cat. no.
9258S; dilution, 1:1,000), p-p38 (cat. no. 4511S; dilution, 1:500),
p38 (cat. no. 9212L; dilution, 1:1,000), NF-κB (cat. no. 8242S;
dilution, 1:1,000), p-NF-κB (cat. no. 3033S; dilution, 1:500),
lamin B (cat. no. sc6216; dilution, 1:1,000), NFATc1 (cat. no.
556602; dilution, 1:1,000), c-Fos (cat. no. sc-447; dilution,
1:200), MMP-9 (cat. no. ab38898; dilution, 1:1,000) and Ctsk (cat.
no. ab19027; dilution, 1:1,000) at 4°C overnight. The membranes
were then incubated with secondary antibodies (cat. no.
111-035-045, 115-035-062; dilution, 1:10,000) at room temperature
for 1 h and chemiluminescence was measured using enhanced
chemiluminescence substrate (Santa Cruz Biotechnology, Inc.). Band
densities were quantified using ImageJ software version 1.51j8
(National Institutes of Health).
Reverse transcription-semi-quantitative
polymerase chain reaction (PCR)
Total RNA was extracted using TRIzol reagent (Takara
Bio, Inc.) according to the manufacturer's protocol. A total of 2
µg total RNA was reverse transcribed into cDNA using a
reverse transcriptase enzyme (Invitrogen; Thermo Fisher Scientific,
Inc.). RT-PCR was performed using the C1000 Touch™ Thermal Cycler
(Bio-Rad, Laboratories, Inc.) and Taq polymerase (Kapa Biosystems;
Roche Diagnostics). The thermal cycling parameters were as follows:
30 sec at 94°C (denaturation), 30 sec at 53-58°C (annealing) and 30
sec at 72°C (extension). The temperature and number of cycles for
each gene are listed in Table I.
The final PCR mixtures were electrophoresed on agarose gels stained
with SYBR-Green (Invitrogen; Thermo Fisher Scientific, Inc.) and
bands were quantified using ImageJ software version 1.51j8.
 | Table IPrimer sequences used for reverse
transcription semi-quantitative PCR. |
Table I
Primer sequences used for reverse
transcription semi-quantitative PCR.
| Gene name | Sequence | Accession no. | Base pair | Annealing
temperature | Cycle |
|---|
| Runx2 | F:
CGGCCCTCCCTGAACTCT | NM_001145920.2 | 75 | 60°C | 38 |
| R:
TGCCTGCCTGGGATCTGTA |
| Alpl
(ALP) | F:
CGGGACTGGTACTCGGATAA | NM_001287172.1 | 208 | 55°C | 42 |
| R:
TGAGATCCAGGCCATCTAGC |
| Sparc
(OSN) | F:
AAACATGGCAAGGTGTGTGA | NM_001290817.1 | 217 | 54°C | 35 |
| R:
TGCATGGTCCGATGTAGTC |
| Spp1
(OPN) | F:
TCTGATGAGACCGTCACTGC | NM_001290377.1 | 170 | 53°C | 38 |
| R:
AGGTCCTCATCTGTGGCATC |
| Col1a1
(COL1) | F:
GCTCCTCTTAGGGGCCACT | NM_007742.4 | 103 | 60°C | 38 |
| R:
CCACGTCTCACCATTGGGG |
| Ibsp
(BSP) | F:
AAAGTGAAGGAAAGCGACGA | NM_008318.3 | 215 | 53°C | 40 |
| R:
GTTCCTTCTGCACCTGCTTC |
| Nfatc1 | F:
TGCTCCTCCTCCTGCTGCTC | NM_198429.2 | 480 | 58°C | 32 |
| R:
CGTCTTCCACCTCCACGTCG |
| Fos
(c-Fos) | F:
ATGGGCTCTCCTGTCAACAC | NM_010234.3 | 480 | 58°C | 33 |
| R:
GGCTGCCAAAATAAACTCCA |
| Mmp-9 | F:
CGACTTTTGTGGTCTTCCCC | NM_013599.4 | 258 | 58°C | 30 |
| R:
TGAAGGTTTGGAATCGACCC |
| Ctsk | F:
AGGCGGCTATATGACCACTG | NM_007802.4 | 403 | 58°C | 26 |
| R:
CCGAGCCAAGAGAGCATATC |
| Tnfrsf11a
(RANK) | F:
AAACCTTGGACCAACTGCAC | NM_009399.3 | 377 | 53°C | 35 |
| R:
ACCATCTTCTCCTCCCHAGT |
| Acp5
(TRAP) | F:
ACTTCCCCAGCCCTTACTACCG | NM_007388.3 | 381 | 58°C | 30 |
| R:
TCAGCACATAGCCCACACCG |
| Oscar | F:
CTGCTGGTAACGGATCAGCTCCCCAGA | NM_001290377.1 | 310 | 53°C | 35 |
| R:
CCAAGGAGCCAGAACCTTCGAAACT |
| Src
(c-src) | F:
TCCAGGCTGAGGAGTGGTACTTTGG | NM_001025395.2 | 306 | 64°C | 40 |
| R:
ATACGGTAGTGAGGCGGTGACACAG |
| Myc
(c-myc) | F:
CACCAGCAGCGACTCTGAAGAAGAG | NM_001177352.1 | 505 | 64°C | 40 |
| R:
AGAGGTGAGCTTGTGCTCGTCTGC |
| Spi1
(PU-1) | F:
CTTCCCTTATCAAACCTTGTC | NM_011355.2 | 398 | 55°C | 30 |
| R:
AGGTGAGCTTCTTCTTGACTT |
| Ocstamp
(OC-Stamp) | F:
AGCTGTAGCCTGGGCTCAGAAG | NM_029021.1 | 196 | 64°C | 45 |
| R:
AGCCTGTGGTAGATGACAGTCGTG |
|
Atp6v0d2 | F:
ATGGGGCCTTGCAAAAGAAATCTG | NM_175406.3 | 504 | 58°C | 30 |
| R:
CGACAGCGTCAAACAAAGGCTTGTA |
| Gapdh | F:
ACTTTGTCAAGCTCATTTCC | NM_008084.3 | 267 | 58°C | 30 |
| R:
TGCAGCGAACTTTATTGATG |
TRAP and pit formation assay
To generate osteoclasts from RAW 264.7 cell
cultures, the RAW 264.7 cells were first seeded at a density of
5×103 cells/well in 96-well-plates containing
differentiation medium (α-MEM containing 100 ng/ml RANKL with or
without LS) and incubated in a cell incubator at 37°C for 5 days.
The culture medium was refreshed on days 2 and 4. The
differentiated cells were fixed with 10% formalin for 10 min and
stained using the TRAP kit according to the manufacturer's
protocol. The cells were then transferred to a new plate and an
equal volume of TRAP solution [4.93 mg p-nitrophenyl phosphate
(PNPP) in 0.5 M 750 ml acetate solution, mixed with 150 ml tartrate
acid solution] was added followed by incubation at 37°C for 30 min.
The reaction was then inhibited by the addition of 0.5 M NaOH, and
the absorbance was measured using an ELISA reader at a wavelength
of 405 nm. To evaluate bone resorption activity, RAW 264.7 cells
(5×103 cells/well) were cultured in osteo strip well
plates containing differentiation medium (α-MEM containing 100
ng/ml RANKL with or without LS) and incubated for 5 days. The cell
culture medium was refreshed on days 2 and 4. Following
differentiation, the medium was removed and the cells were
dissolved with NaClO before the plate was allowed to dry. The
number of TRAP-positive cells and the area of the pit formation
were measured using ImageJ software.
F-actin ring immunofluorescence
assay
To evaluate F-actin ring formation, the RAW 264.7
cells (5×103 cells/well) were cultured in 96-well plates
containing differentiation medium (α-MEM containing 100 ng/ml RANKL
with or without LS) and incubated in a cell incubator at 37°C for 5
days. The differentiated cells were fixed with 4% paraformaldehyde
for 20 min and permeabilized with 0.1% Triton X-100 in PBS for 5
min. The cells were then stained using the Actistain™ 488
Fluorescent Phalloidin (Cytoskeleton Inc.) at room temperature in
the dark for 30 min. The cells were washed with PBS, and the nuclei
were then counterstained with 4′,6-diamidino-2-phenyl-indole (DAPI,
Sigma Aldrich; Merck KGaA). The formation of the actin ring was
captured using an immunofluorescence microscope (Cellena;
Logosbio).
Animal experiments
Institute of Cancer Research (ICR) CD-1 mice (male;
age, 4 weeks; weighing, 27-29 g) were purchased from Nara Biotech,
Co., Ltd. Animal experiments proceeded with permission from the
Kyunghee University Animal Committee [ref. no. KHUASP(SE)-15-095].
The mice were maintained under 12 h light/dark cycle at 22-24°C at
a relative humidity of 55-55% with free access to food and water.
The mice were allowed to acclimatize to the laboratory conditions
at the animal breeding facility for 1 week. The animal model of
osteoporosis was established by an intraperitoneal injection of
LPS, as previously described (18,19). The mice were divided into the
following 3 groups (n=6/group): Group 1, mice treated with 100
µl PBS via intraperitoneal injection and orally administered
with distilled water daily; group 2, mice treated with LPS (5 mg/kg
body weight) via intraperitoneal injection and orally administered
with distilled water daily; and group 3, mice treated with LPS via
intraperitoneal injection and orally administered with LS at a
concentration of 100 mg/kg daily. PBS or LPS administration were
carried out on days 1 and 4. The humane endpoint of this experiment
was as follows: Dirty hair and eye discharge, self-injury and
anxiety, vomiting and hemoptysis, inactivity, anxiety and
headache). No abnormal signs that signified the humane endpoints of
the experiment were observed from any of the mice during the
experiment. All mice were sacrificed on day 9 and the right femurs
were extracted and analyzed by micro-computed tomography (micro-CT;
SkyScan1176; SkyScan; Bruker Corp.). Bone volume/total volume
(BV/TV), trabecular pattern factor (Tb.pf) and structure model
index (SMI) calculations were performed using NRecon software
(SkyScan version 1.6.10.1; SkyScan; Bruker Corp.).
Statistical analysis
The experiments were repeated at least 3 times. The
results are presented as the means ± standard error of the mean.
Statistical comparisons were performed one-way ANOVA tests,
followed by Dunnett's post hoc analysis. A value of P<0.05 was
considered to indicate a statistically significant difference.
Results
Quantitative analysis of the LS
extract
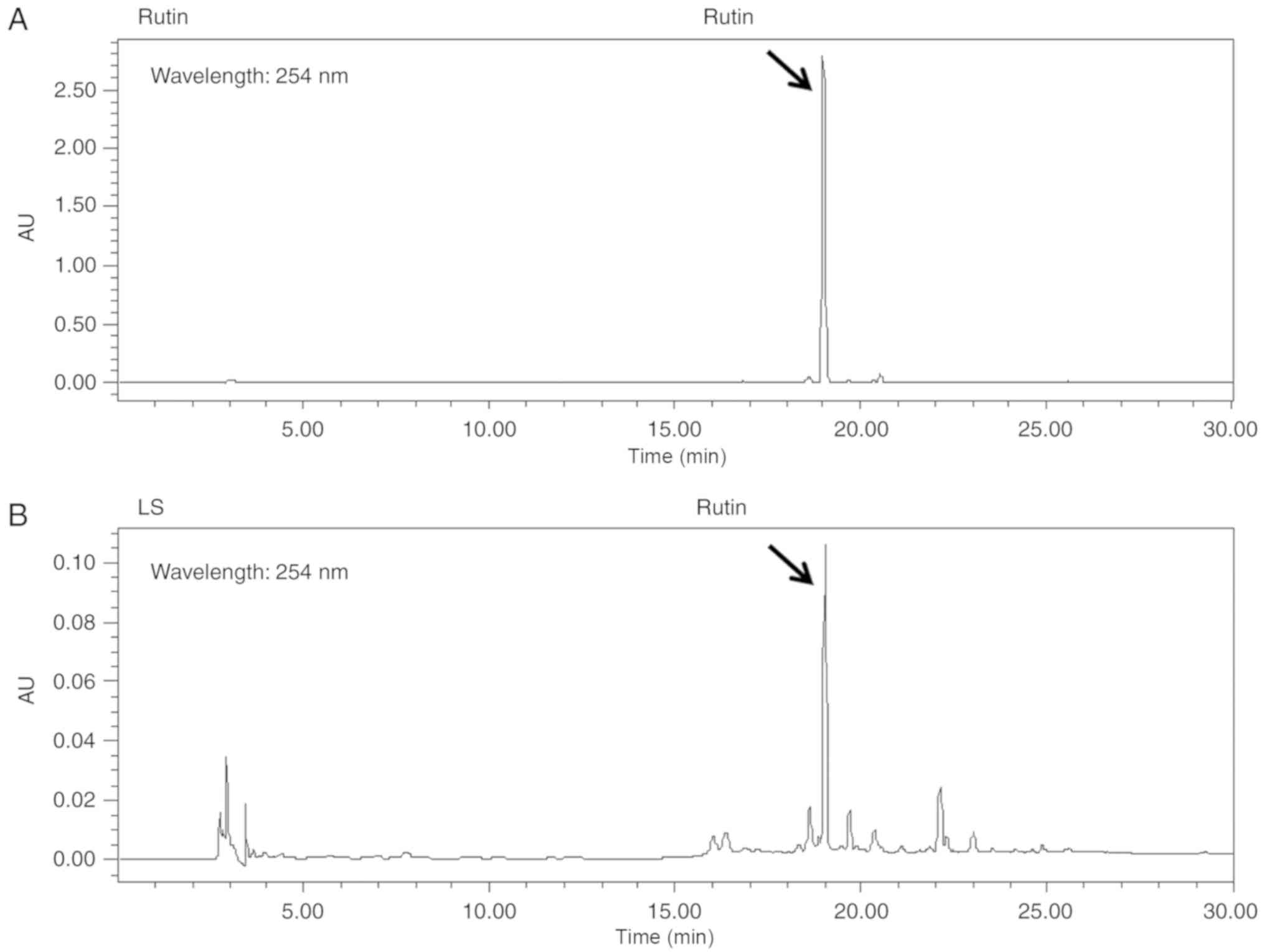
Rutin is a bioactive marker compound used for the
validation of LS. The chromatogram of the ethanol extract from LS
demonstrated that a number of peaks were detected at a retention
time of 0 and 30 min, and rutin was identified at the same
retention time as the standards (Fig.
1).
LS treatment significantly increases
osteoblast differentiation and promotes calcified nodule
formation
LS at a concentration of 400 and 800 µg/ml
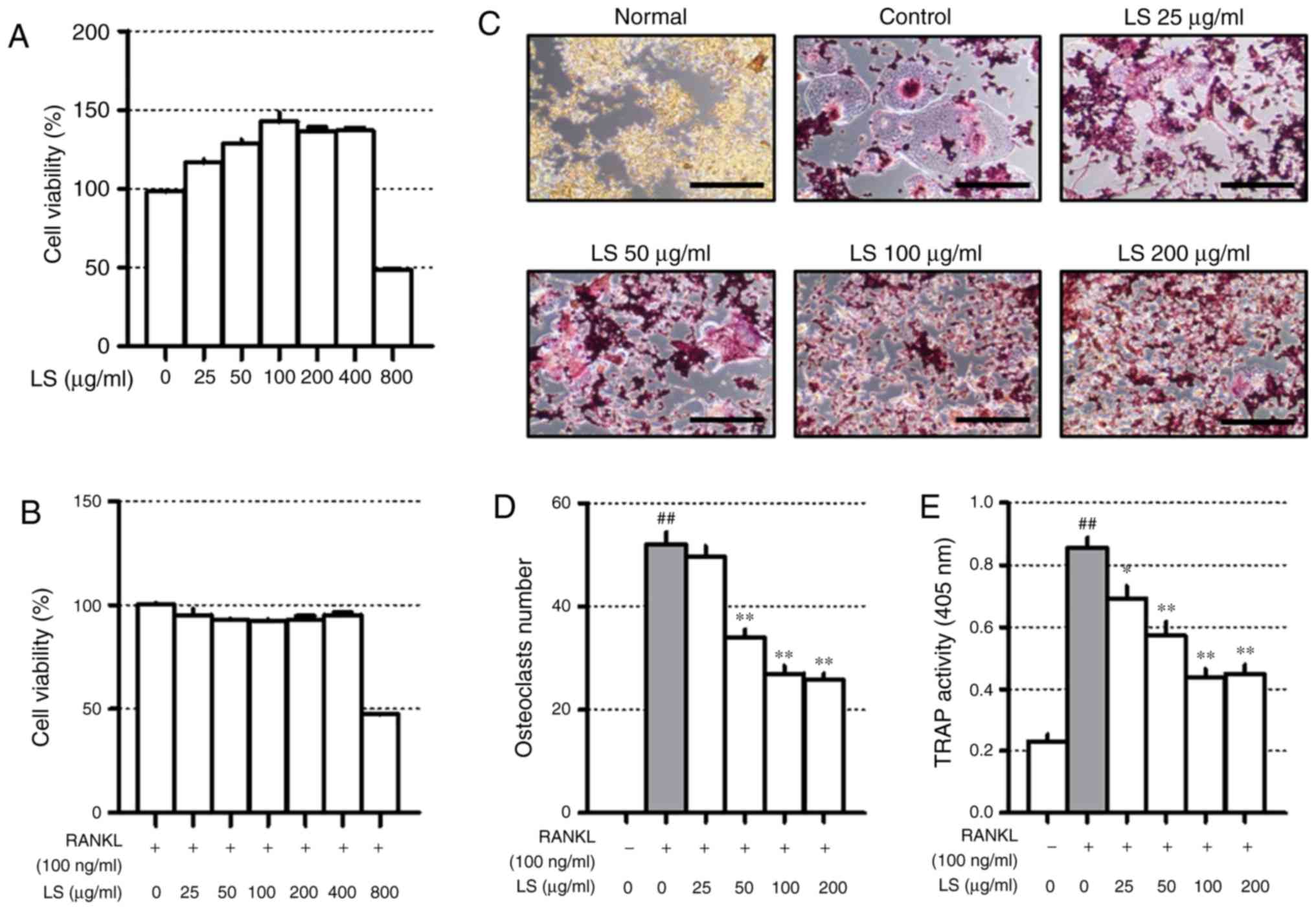
was toxic to the MC3T3-E1 cells for 3, 7 and 14 days (Fig. 2A). Subsequently, to investigate
the effects of LS on osteoblast differentiation, the MC3T3-E1 cells
were treated with LS, ascorbic acid and β-glycerophosphate.
Following 14 days of culture, Von Kossa and Alizarin Red S staining
confirmed that calcified nodules were formed at an earlier stage in
the LS treatment group when compared with the differentiated
medium-treated group (Fig. 2B and
C). At 17 and 21 days, LS enhanced osteoblast differentiation,
as indicated by the increase in mineralized nodule formation. In
addition, the absorbance of the extracted Alizarin Red S dye was
increased by LS treatment (Fig.
2D). No significant difference was observed in osteoblast
differentiation with LS at 50 to 200 µg/ml. Thus, the
highest concentration was determined to be 50 µg/ml (data
not shown). To determine the mechanisms of action of LS as regards
osteoblast differentiation, the expression levels of transcription
factors and osteogenic genes were analyzed in LS-treated
osteoblasts. BMP-2 and RUNX2 are major transcription factors
involved in osteoblast differentiation, and LS was observed to
increase the expression levels of these genes (Fig. 2E and F). In addition, LS promoted
the expression of osteogenic genes, including RUNX2, ALP
(Alpl), OPN (Sparc), OSN (Spp1) and COL1
(Col1a1). BSP (Ibsp) expression increased with LS
treatment, although the difference was not statistically
significant (Fig. 2G and H).
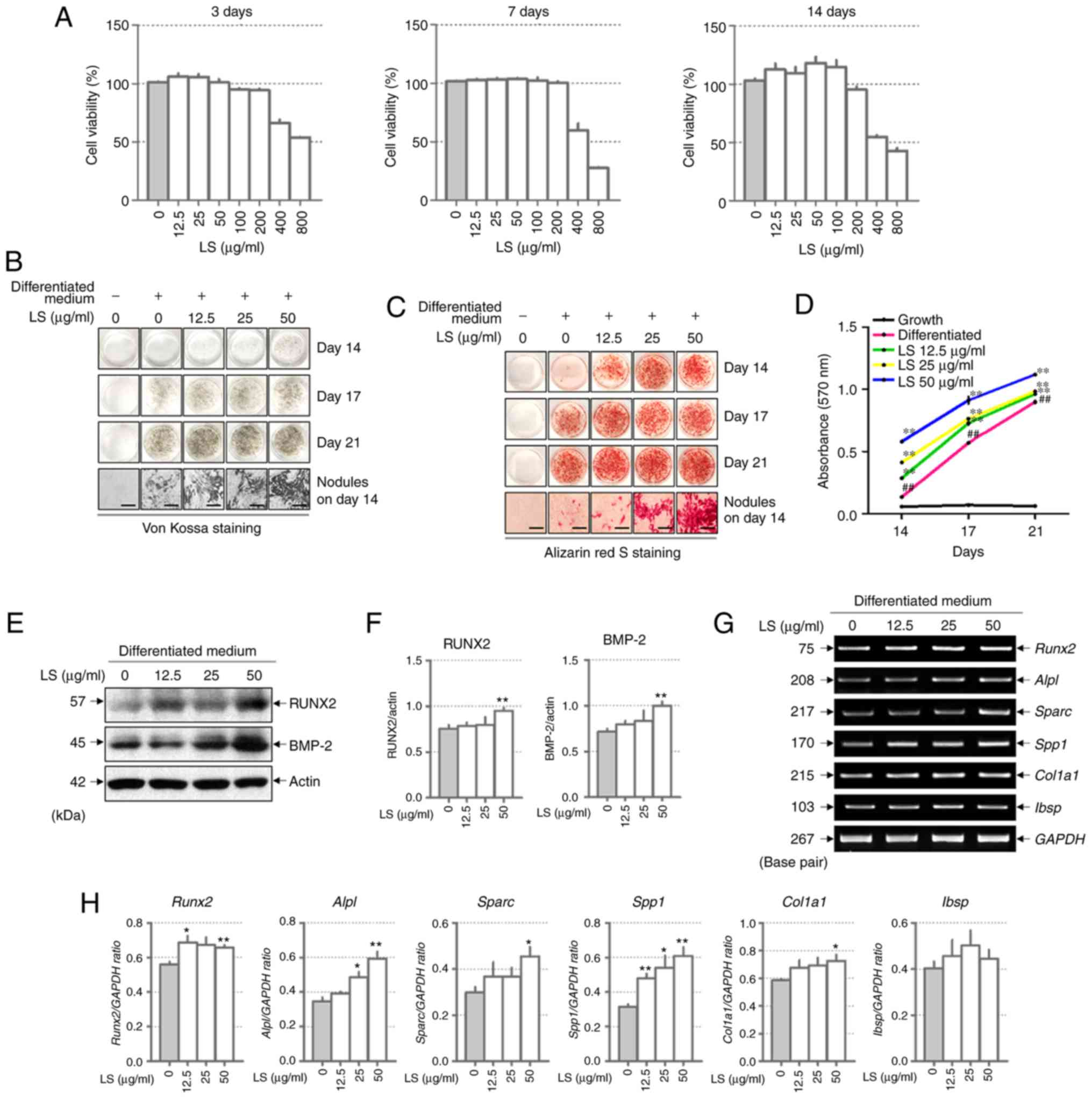 | Figure 2Effect of LS on osteoblast
differentiation. (A) Cytotoxicity of LS used in osteoblast
promotion assays was confirmed using an MTS assay for 3, 7 and 14
days. Calcified nodules produced by osteoblasts were stained with
(B) Von Kossa and (C) Alizarin Red S (×100 magnification; scale
bar, 200 µm). (D) Alizarin Red S stain was extracted from
the cells and quantified by measuring the absorbance at 570 nm. The
effect of LS on the expression of transcription factors involved in
osteoblast differentiation was also determined. (E) Protein
expression of RUNX2 and BMP-2 were examined by western blot
analysis. (F) The protein expression levels of each marker were
normalized to actin. (G) mRNA levels of RUNX2, ALP (alpl),
OPN (Spp1), OSN (Sparc), COL1 (Col1a1) and BSP
(Ibsp) were analyzed using reverse transcription
semi-quantitative PCR. (H) The mRNA expression levels of each
factor was normalized to GAPDH. The results are presented as the
means ± standard error of the mean (n=3). ##P<0.01
vs. growth cells; *P<0.05 and **P<0.01
vs. differentiated cells. LS, Leonurus sibiricus L.; RUNX2,
runt-related transcription factor 2; BMP-2, bone morphogenetic
protein 2; ALP, alkaline phosphatase; OPN, osteopontin; OSN,
osteonectin; COL1, type I collagen; BSP, bone sialoprotein. |
LS inhibits osteoclast differentiation at
non-cytotoxic concentrations
Before confirming the osteoclast experiments, the
toxicity of LS in the RAW 264.7 cells was confirmed. A high
concentration (800 µg/ml) of LS reduced cell viability to
below 50% and the concentrations of LS used in the osteoclast
experiments did not affect the level of cytotoxicity (Fig. 3A and B). Thus, the possibility
that the inhibition of osteoclast formation was due to cytotoxicity
was excluded. To confirm the inhibitory effects of LS on
osteoclastogenesis, TRAP staining and pit formation assays were
performed. As demonstrated in Fig.
3C, the number of TRAP-positive osteoclasts was increased
following stimulation of the cells with RANKL, and LS decreased the
number and size of osteoclasts in a dose-dependent manner (Fig. 3D). In addition, LS inhibited TRAP
activity in the differentiation medium (Fig. 3E). No significant difference was
observed in the inhibitory effects on osteoclastogenesis with LS at
100 to 400 µg/ml. Thus, the highest concentration was
determined to be 100 µg/ml.
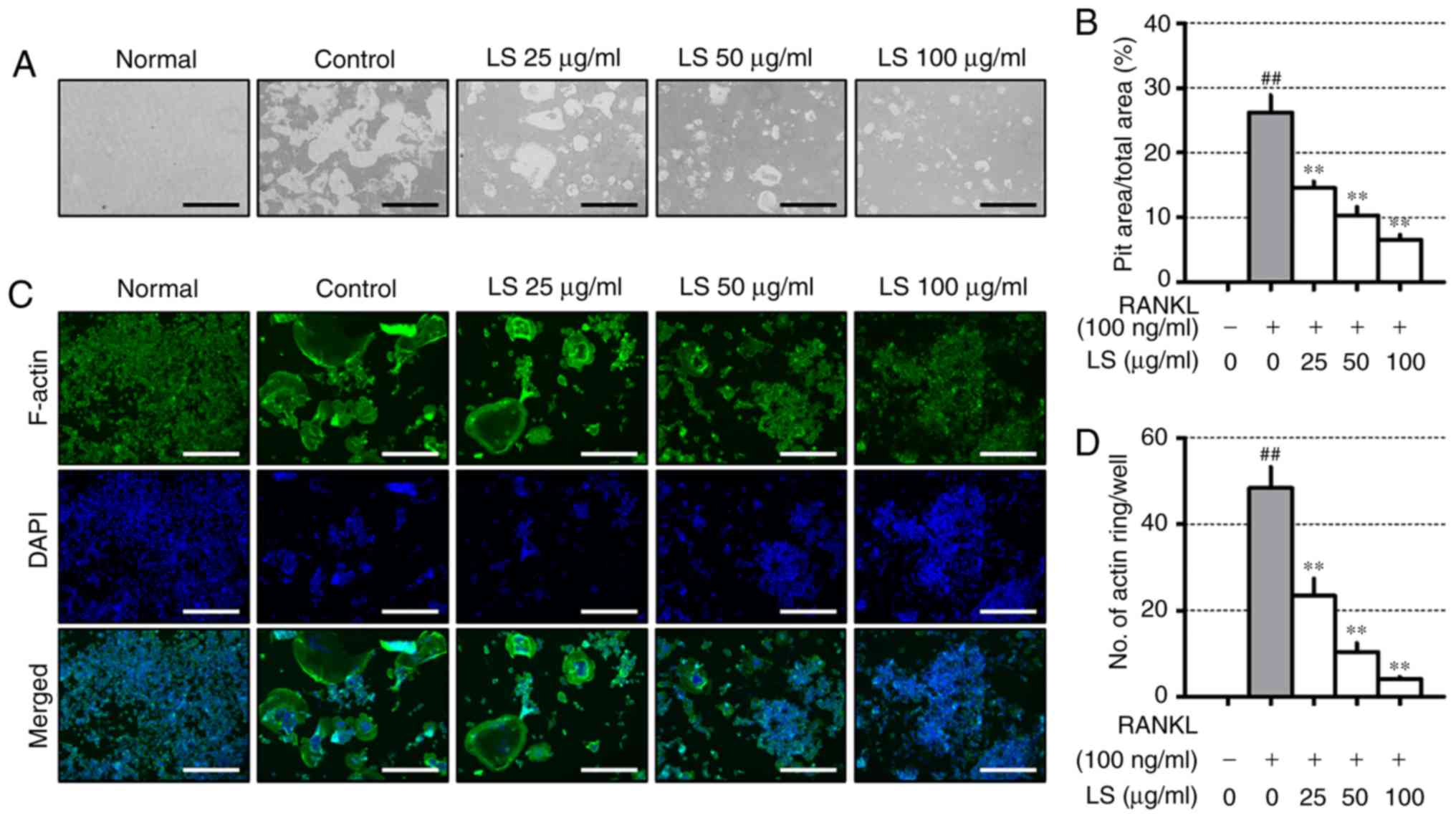
LS inhibits bone resorptive activity
To evaluate the inhibitory effects of LS on
resorptive activity, differentiated osteoclasts formed resorption
pits through calcium absorption (Fig.
4A), and the area of the resorption pit was reduced by LS
treatment (Fig. 4B). RANKL
stimulation increased the formation of the well-polarized F-actin
ring (Fig. 4C). However, LS was
observed to decrease both the size and number of F-actin ring
structures (Fig. 4D).
LS inhibits the activation of NF-κB and
the phosphorylation of MAPKs
When osteoclast differentiation is induced by RANKL,
the NF-κB and MAPKs signaling pathways are essential. RANKL
stimulation increases the protein levels of NF-κB and p-NF-κB. In
this study, LS was observed to decrease the translocation of NF-κB
to the nucleus and inhibit the phosphorylation of NF-κB (Fig. 5A and B). In addition, LS treatment
significantly inhibited the RANKL-induced phosphorylation of
ERK1/2, JNK and p38 in a concentration-dependent manner (Fig. 5C and D).
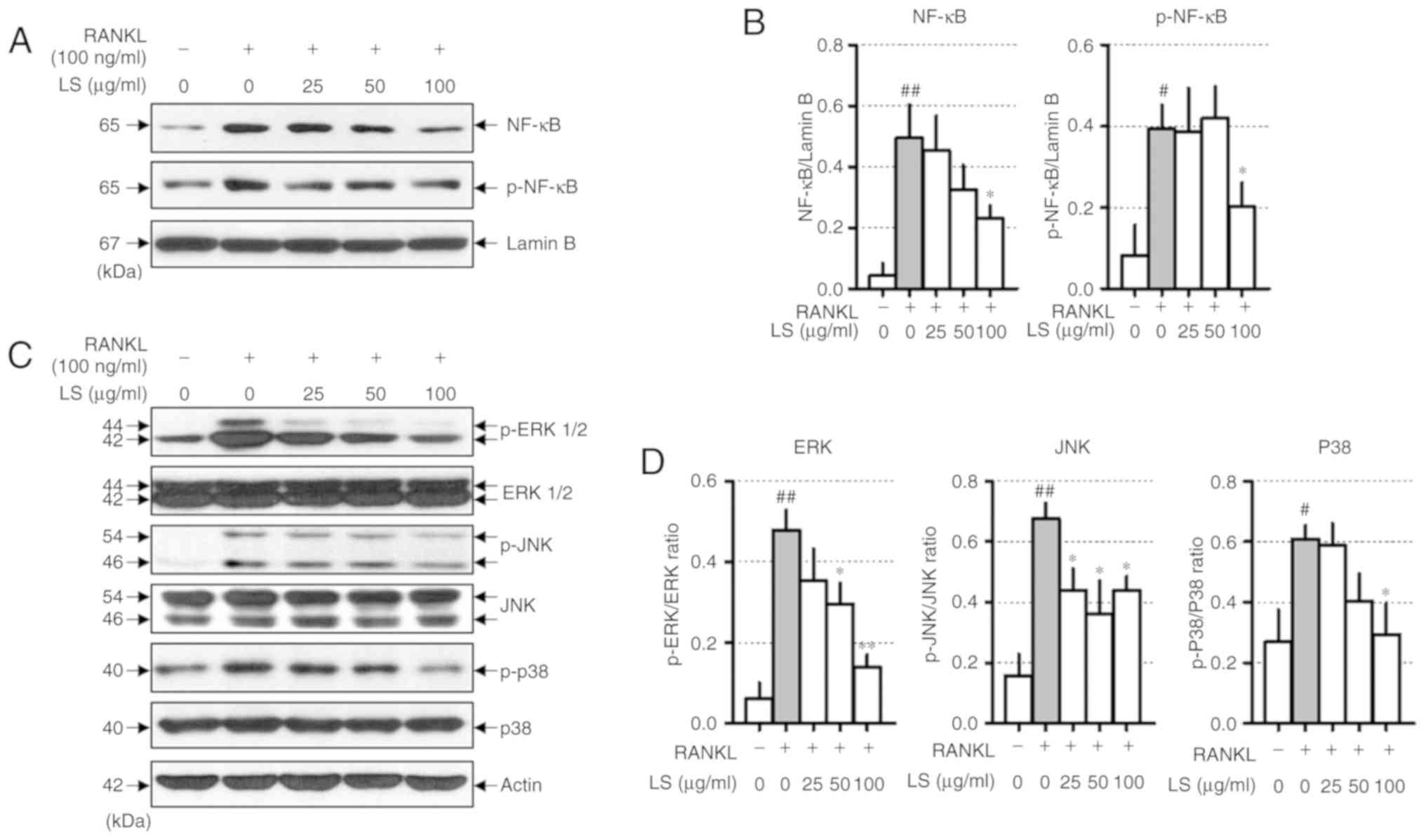 | Figure 5Effect of LS on the expression of
NF-κB and MAPK signaling pathways. (A) Protein expression of NF-κB
and p-NF-κB were measured by western blot analysis. (B) NF-κB and
p-NF-κB expression was normalized to lamin B. (C) Protein
expression of MAPKs were measured by western blot. (D) The
expression of p-ERK1/2, p-JNK and p-p38 were normalized to total
ERK1/2, JNK and p38, respectively. The results are presented as the
means ± standard error of the mean (n=3). #P<0.05 and
##P<0.01 vs. normal; *P<0.05 and
**P<0.01 vs. RANKL-induced control. LS, Leonurus
sibiricus L.; NF-κB, nuclear factor-κB; MAPKs,
mitogen-activated protein kinases; p-, phosphorylated; ERK1/2,
extracellular signal-regulated kinase 1/2; JNK, c-Jun N-terminal
kinase; RANKL, receptor activator of nuclear factor κ-B ligand. |
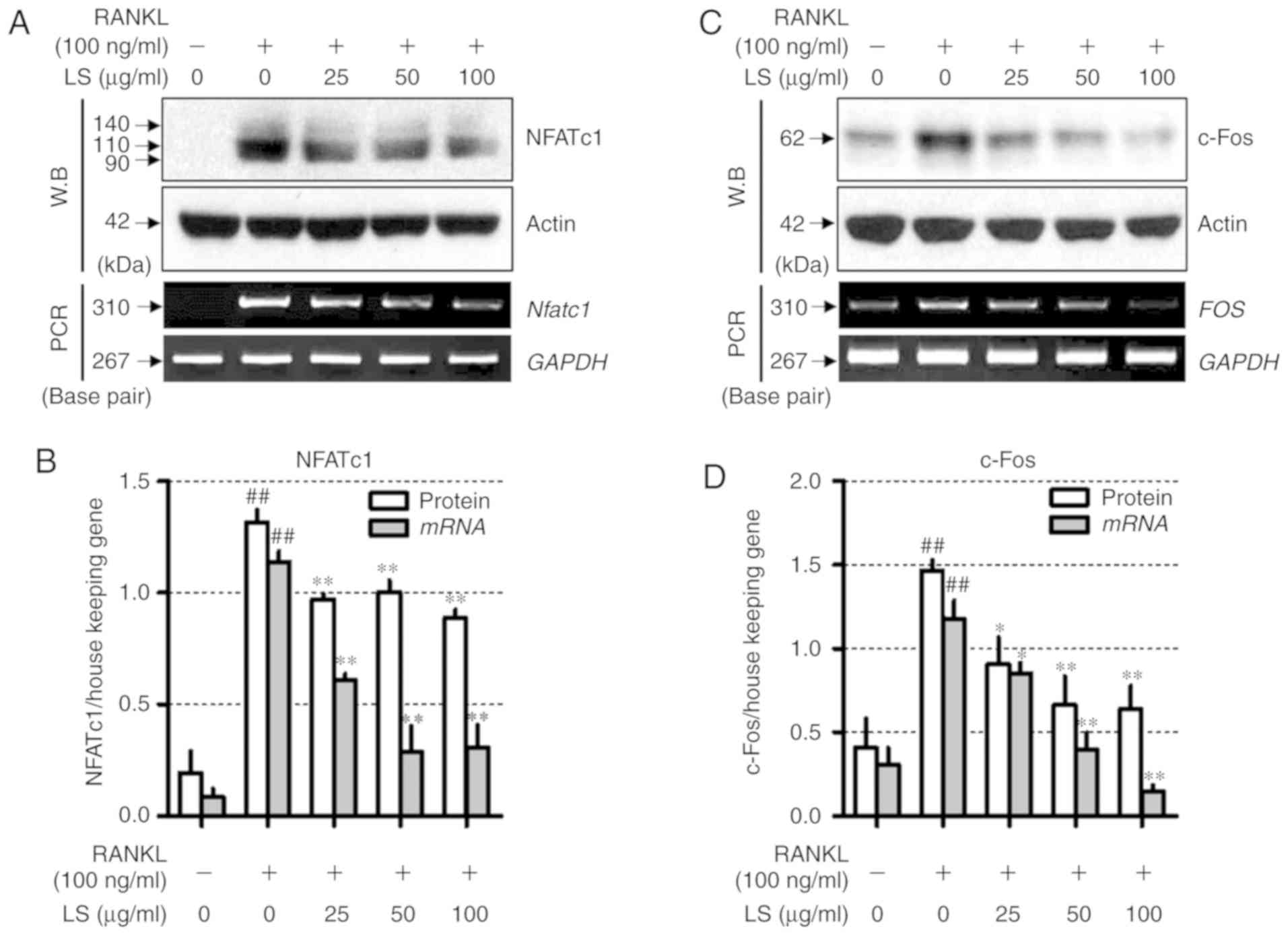
LS inhibits the expression of essential
transcription factors involved in osteoclastogenesis
To determine the mechanisms underlying LS-mediated
osteoclast inhibition, the expression of NFATc1 and c-Fos was
measured by western blot analysis and reverse
transcription-semi-quantitative PCR. As demonstrated in Fig. 6, RANKL stimulation increased the
protein and mRNA levels of Nfatc1 and FOS, whereas LS
suppressed the expression of these factors. In addition, LS did not
affect the expression of housekeeping genes, such as actin and
GAPDH.
LS inhibits the expression of bone
resorption markers and osteoclastogenesis-associated markers
To examine whether LS inhibits bone
resorption-related enzymes, the effects of LS on the expression of
MMP-9 and Ctsk were examined by western blot analysis and reverse
transcription-semi-quantitative PCR. As demonstrated in Fig. 7A-D, RANKL stimulation induced the
protein and mRNA expression levels of Mmp-9 and Ctsk,
whereas LS markedly suppressed the expression of these bone
resorption-related enzymes. In addition, the effects of LS on
markers involved in osteoclast differentiation were analyzed. As
demonstrated in Fig. 7E and F,
the expression of RANK (Tnfrsf11a), TRAP (Acp5),
OSCAR, c-src, c-myc, PU-1 (Spi1), OC-STAMP and ATP6v0d2 was
increased by RANKL treatment, whereas LS significantly inhibited
the expression of these genes.
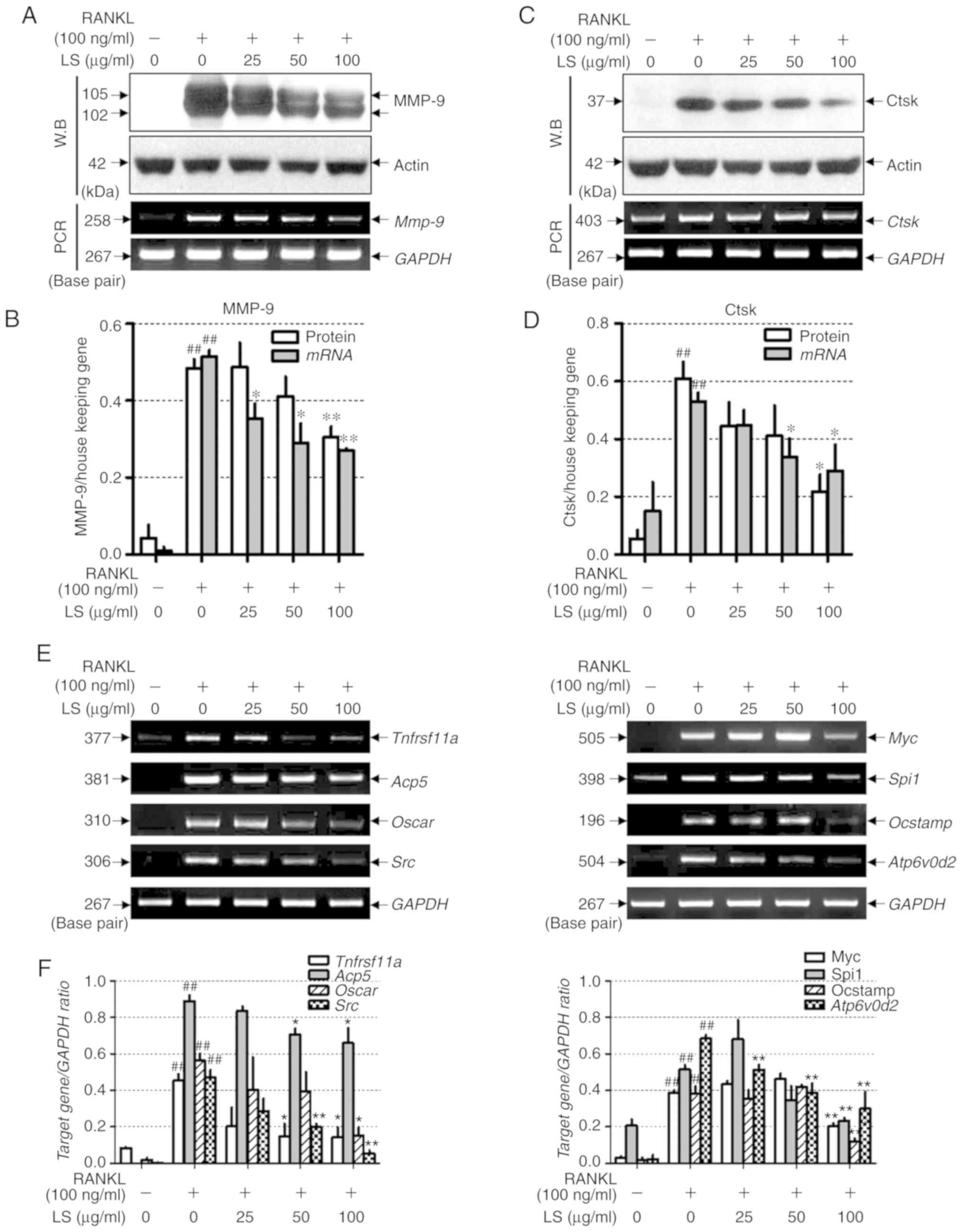 | Figure 7Effect of LS on the expression of
osteoclastic bone resorption enzymes. (A) Expression of MMP-9 was
measured by western blot analysis and RT-PCR analysis. (B) Protein
and mRNA expression levels of MMP-9 were normalized to actin and
GAPDH, respectively. (C) Expression of Ctsk was detected by western
blot and reverse transcription semi-quantitative PCR. (D) Protein
and mRNA expression of Ctsk was normalized to actin and GAPDH,
respectively. (E) mRNA levels of RANK (Tnfrsf11a), TRAP
(Acp5), OSCAR, c-src (Src), c-myc (Myc), PU-1 (Spi1),
OC-STAMP and ATP6v0d2 were analyzed by reverse transcription
semi-quantitative PCR on day 4. (F) The mRNA expression levels of
each factor were normalized to GAPDH. The results are presented as
the means ± standard error of the mean (n=3).
##P<0.01 vs. normal; *P<0.05 and
**P<0.01 vs. RANKL-induced control. LS, Leonurus
sibiricus L.; MMP-9, matrix metallopeptidase-9; Ctsk, cathepsin
K; RANK, receptor activator of nuclear factor κ; TRAP,
tartrate-resistant acid phosphatase; OSCAR, osteoclast-associated
immunoglobulin-like receptor; OC-STAMP, osteoclast stimulatory
transmembrane protein; ATP6v0d2, ATPase H+ transporting V0 subunit
d2; RANKL, receptor activator of nuclear factor κ-B ligand. |
Administration of LS inhibits LPS-induced
inflammatory bone loss
The results presented thus far indicated that LS
activated osteoblast differentiation and inhibited osteoclast
differentiation. To confirm these in vitro results, the
potential inhibitory effects of LS on bone loss in vivo were
investigated. The concentration of LS administered in this study
was determined as follows: In oriental medicine, based on a typical
adult weight of 60 kg, a single dose of LS is 8 g and corresponding
to 0.6184 g (yield, 7.73%) of LS extract. Therefore, 10.3 mg of LS
should be administered per 1 kg. In previous studies, mice are
known to have a 7-fold higher metabolism than humans (20), and an LS dose of approximately 72
mg/kg is appropriate. In this study, LS was administered at a
slightly higher concentration than that administered to humans. To
achieve this, a mouse model of endotoxin-induced bone destruction
was first generated (21,22). The following day after LPS
administration, the weight of the mice was reduced, but this was
recovered. In addition, no marked changes in body weight due to LS
were observed (Fig. S1). As
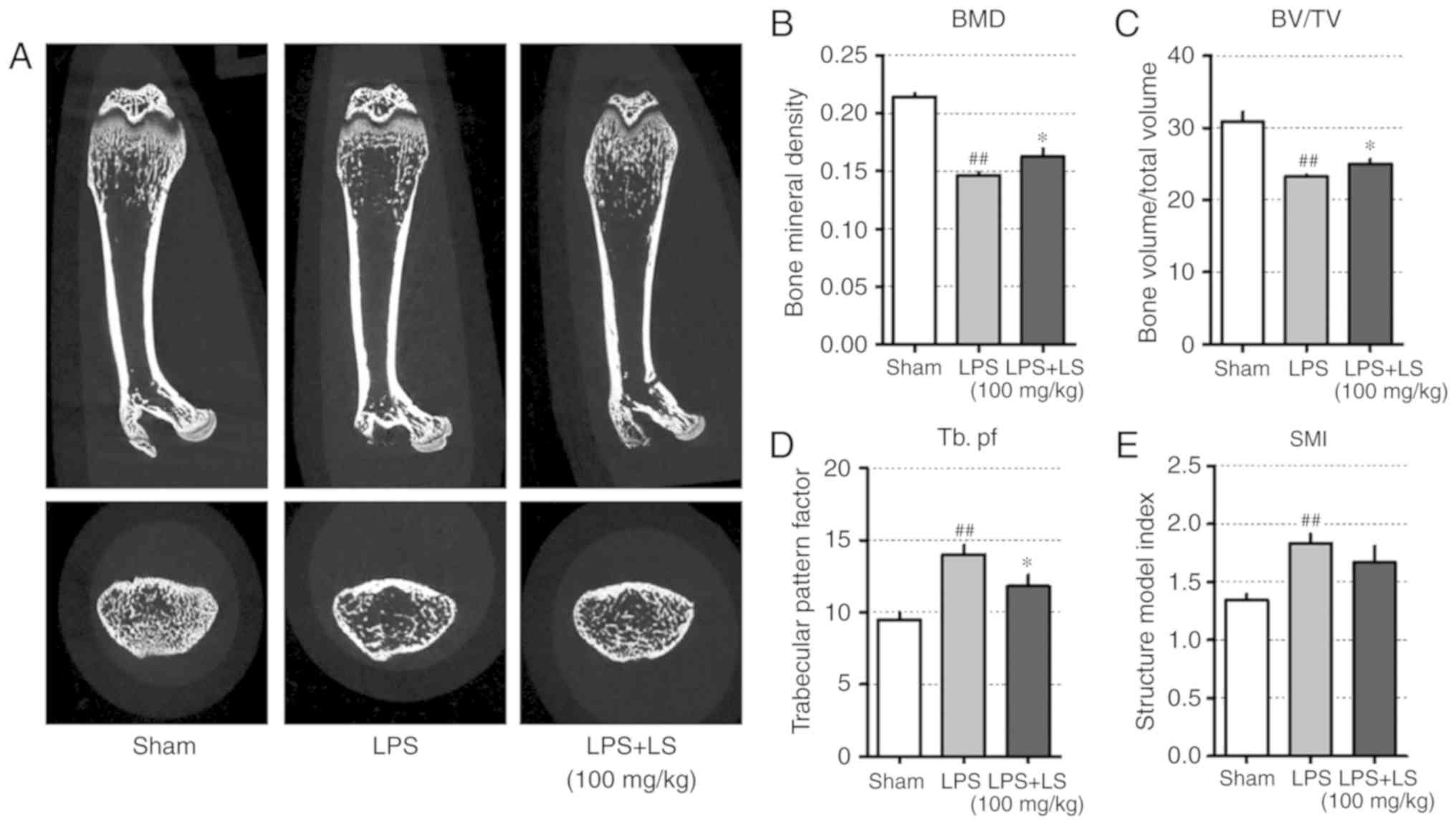
demonstrated in Fig. 8A, micro-CT
images of the femur indicated a decrease in trabecular bone;
however, the administration of LS significantly inhibited
LPS-induced bone loss. Based on the radiographic results, the
effect of LS on bone microstructure was also analyzed. This
revealed that LPS administration reduced bone mineral density (BMD)
and BV/TV, whereas the administration of LS significantly inhibited
the LPS-induced reduction in BMD and BV/TV. In addition, the
LPS-induced increase in Tb.pf was suppressed by LPS treatment.
Similarly, SMI increased with LPS administration, which was
suppressed by treatment with LS; however, the difference was not
statistically significant (Fig.
8B-E).
Based on the results of this study, LS promoted
osteogenic gene expression by upregulating RUNX2 in MC3T3-E1 cells.
In addition, LS inhibited the bone resorption gene by inhibiting
the expression of NFATc1/c-Fos in RANKL-induced RAW 264.7 cells. As
a result, it exerted an anti-osteoporotic effect in mice with
LPS-induced bone less (Fig.
9).
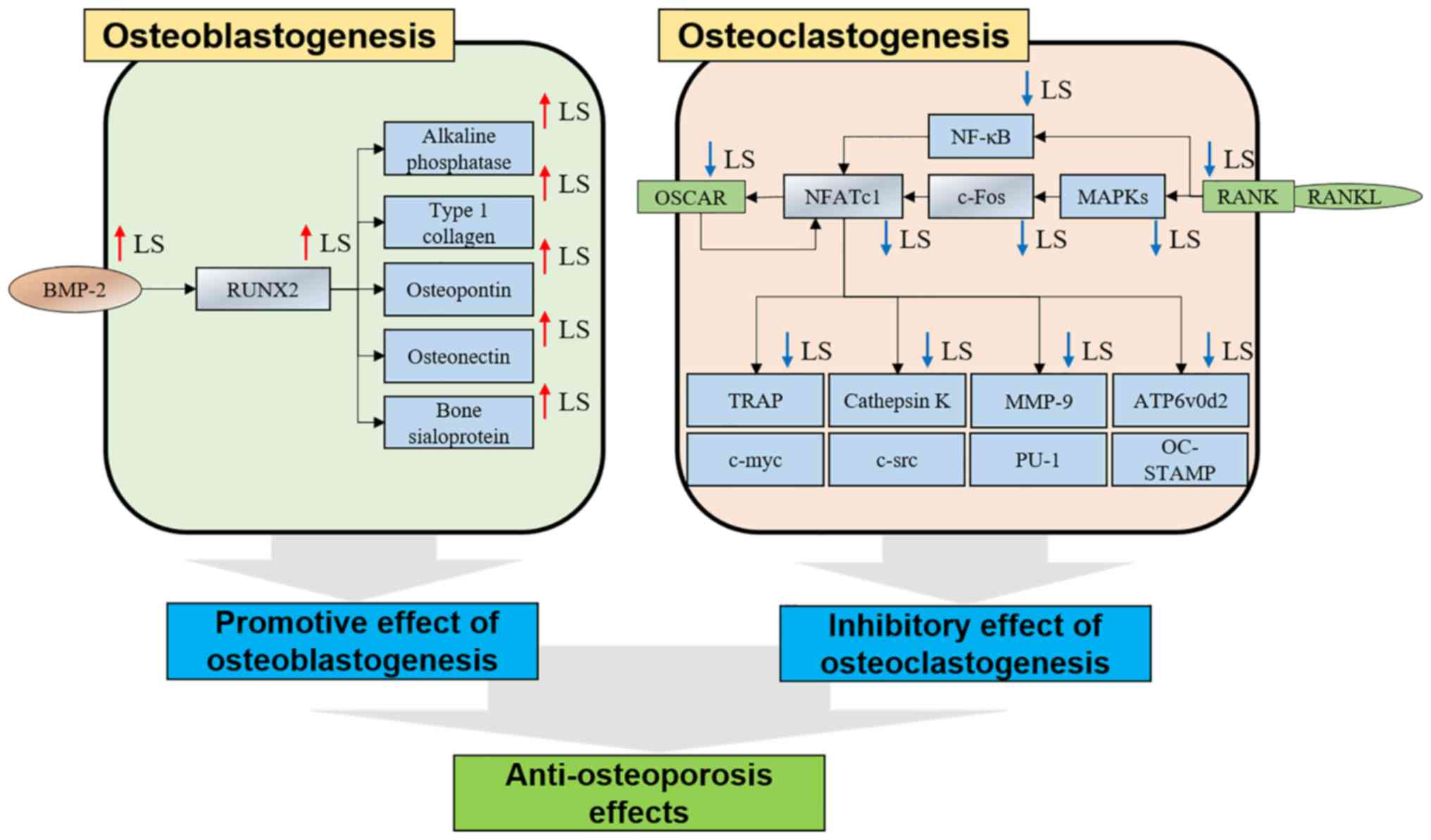 | Figure 9Mechanisms of osteoblasts and
osteoclasts in LS. LS, Leonurus sibiricus L. BMP-2, bone
morphogenetic protein 2; RUNX2, Runt-related transcription factor
2; OSCAR, osteoclast-associated immunoglobulin-like receptor;
NFATc1, nuclear factor of activated T-cells 1; TRAP,
tartrate-resistant acid phosphatase; RANKL, receptor activator of
nuclear factor-κB ligand; MAPK, mitogen-activated protein kinase;
NF-κB, nuclear factor-κB; MMP-9, matrix metallopeptidase-9;
ATP6v0d2, ATPase H+ transporting V0 subunit d2. |
Discussion
Bone metabolic disease is characterized by the
abnormal activity of osteoclasts. To solve this problem,
researchers have investigated natural products that activate
osteoblasts or exert inhibitory effects on osteoclastogenesis
(23). Although the ethanol
extract of LS on osteoporosis has not been studied to date, the
study by Yang et al demonstrated that leonurine
hydrochloride, which is component of LS, promoted osteogenic
differentiation (24). However,
their study did not proceed onto investigating osteoclastogenesis.
In osteoblast differentiation, RUNX2 induction through the
Wnt/b-catenin mechanism is the main focus of research. On the
contrary, this study investigated the effects of LS on osteoblasts
and osteoclasts, and the promotion of osteoblast differentiation
was the result of the upregulation of BMP-2 and RUNX2. To the best
of our knowledge, this study is also the first LS study of bone
destruction due to chronic inflammation. Two cell models were used
to determine the effects of LS on osteoblasts and osteoclasts.
MC3T3-E1 cells are derived from the bone/calvaria of C57BL/6 mice
and are a well-known osteoblast-like cell line. These cells are
useful to the research of the molecular mechanisms of osteoblasts
and provide a model for studying bone cell differentiation and
proliferation (25). The RAW
264.7 murine cell line is suitable for establishing a RANKL-induced
osteoclast experimental model (26).
Mineralization nodules are one of the biomarkers
that determine osteoblast maturation, and can be detected by Von
Kossa and Alizarin S Red staining (27,28). In the current study, the formation
of calcified nodules following induction by ascorbic acid and
β-glycerophosphate was increased by LS treatment. BMP-2 is a
transcription factor that regulates typical osteoblast
differentiation and bone formation (29). Once activated, it translocates
into the nucleus to induce the expression of bone matrix proteins,
such as RUNX2, ALP, BSP and COL1 (6). Among these, RUNX2 is essential for
osteoblast differentiation, and is detected in pre-osteoblasts,
where its expression increases as the cell develops into an
immature osteoblast (7). Some
studies have demonstrated that natural extracts can upregulate
BMP-2/RUNX2 and promote osteoblast differentiation (30,31). ALP and BSP is involved in
early-stage molecular events during osteoblast differentiation and
is known to upregulate the initial mineralization stage (32,33). OPN is known as a non-collagenous
major bone matrix protein, and it is expressed when osteoblasts and
osteocytes form bone, and plays an important role in bone
regeneration. In particular, OPN enables osteoclasts to attach to
the bone (34). Recently, the
overexpression of OPN was observed to inhibit the expression of
BMP-2-induced ALP and BSP, ultimately inhibiting osteoblast
differentiation and mineralization (35). OSN is a glycoprotein that attaches
to calcium in bone. This occurs when osteoblasts produce bone and
initiate mineralization (36). In
OSN-null mice, the number of osteoblasts and the rate of bone
formation are decreased (37).
COL1 expression is typically upregulated in the osteoblast
differentiation and mineralization stages (38). The results of the current study
demonstrated that LS upregulated the expression of BMP-2 and RUNX2,
and markedly increased the expression of ALP, BSP, COL1, OPN and
OSN. These results indicate that the effect of LS on promoting
osteoblasts and the formation of mineralization nodules may be
mediated via the activation of BMP-2, RUNX2 and osteogenic
genes.
TRAP is known as an osteoclast phenotype marker and
TRAP staining is the standard method used to detect osteoclast
differentiation (39). In
addition, TRAP expression is associated with differentiated
osteoclast migration and also effects osteoclast differentiation
and maturation. Mice lacking the TRAP gene are known to develop
osteoporosis and display irregular bone microstructures (40). In the present study, LS inhibited
RANKL-induced osteoclast differentiation and activity. The pit
formation assay is used to measure the bone resorption ability of
osteoclasts and an F-actin ring is the most visible feature of
mature osteoclasts (41). The
results of the current study also demonstrated that LS
significantly inhibited pit formation in RANKL-induced osteoclasts.
Taken together, these results suggest that LS inhibits osteoclast
differentiation and impairs the function of mature osteoclasts.
NF-κB translocates to the nucleus via the binding of
RANK-RANKL and is known to play a key role in the early stages of
osteoclast development. Rats lacking NF-κB1 and NF-κB2 are unable
to differentiate into osteoclasts, which leads to osteopetrosis in
rats (42). In addition to the
NF-κB signaling pathway, the MAPK signaling pathway is also known
to play an important role in osteoclast differentiation. ERK is
associated with osteoclast survival (43), and JNK-deficient rats are unable
to differentiate into osteoclasts (44). Furthermore, p38 affects early
osteoclastogenesis and plays an important role in the expression of
Ctsk (45,46). The present study confirmed that LS
inhibited NF-κB expression and its phosphorylation in the nucleus.
In addition, LPS suppressed the MAPK signaling pathway by
inhibiting the phosphorylation of ERK, JNK and p38.
c-Fos is an established group of essential
transcription factors involved in osteoclast differentiation
(47). c-Fos, which is induced by
binding of RANKL-RANKL, directly activates the expression of
NFATc1. Mice deficient in c-fos exhibit defective osteoclast
differentiation, which leads to osteopetrosis. NFATc1 is the master
transcription factor involved in osteoclast differentiation;
NFATc1-deficient embryonic stem cells are unable to differentiate
into osteoclasts via RANKL (9).
In addition, the ectopic expression of NFATc1 successfully promotes
osteoclast differentiation even in the absence of RANKL (48). In the present study, LS inhibited
the protein and gene expression levels of NFATc1 and c-Fos. This
suggests that the inhibitory effect of LS on osteoclast
differentiation may be due to the suppression of the essential
transcription factors, NFATc1 and c-Fos.
As LS strongly inhibited NFATc1, it can thus be
hypothesized that LS may also suppress the expression of
NFATc1-regulated osteoclast-associated genes, including TRAP,
MMP-9, Ctsk, ATP6v0d2 and OSCAR (9). MMP-9 and Ctsk are enzymes that are
released when osteoclasts undergo bone resorption. MMP-9 is
expressed in the early differentiation phase of osteoclasts and
when mature osteoclasts absorb bone (49). In addition, Ctsk is essential for
bone resorption, and osteoclasts extracted from Ctsk-deficient mice
are unable to absorb bone (50).
In the present study, LS was demonstrated to suppress osteoclast
function by significantly inhibiting bone resorption-related
enzymes. c-src is essential for ruffled border formation (51), c-myc is expressed early in
osteoclast differentiation and induces other effector genes
(52). PU-1 is induced by RANKL
stimulation and up-regulates p38 and NFATc1 (53). In this study, LS inhibited the
gene expression levels of c-src, c-myc and PU-1. OSCAR is
specifically expressed in early stage osteoclasts or in mature
osteoclasts, and induces calcium activation to assist in the
expression of RANKL-mediated NFATc1 (54). Similar to these results, the
majority of osteoclast inhibition studies have reported that
osteoclast-related genes are decreased by reduction of c-Fos and
NFATc1 (55,56). OC-STAMP and ATP6v0d2 are expressed
when the osteoclast precursor cells are fused, and when it is
deficient, differentiation into osteoclasts is not completed.
Moreover, these genes are essential for formation of F-actin ring
(57,58). In the current study, LS suppressed
the expression of ATP6v0d2 and OSCAR. These results suggest that LS
inhibits MMP-9, Ctsk, OSCAR, c-src, c-myc, PU-1, OC-STAMP and
ATP6v0d2 which serve important roles in osteoclast differentiation
and activation via inhibition of NFATc1 (Fig. 9).
LPS-induced mouse models are commonly used to verify
bone loss and bone destruction in chronic inflammatory conditions
(59). However, the mechanisms
underlying the activation of osteoclasts by LPS in vivo are
unclear. LPS-induced inflammatory cytokines, such as tumor necrosis
factor-α, interleukin (IL)-1β and IL-6 are known to influence
osteoclast differentiation (22).
As demonstrated by the radiographic results in the current study,
LS treatment significantly inhibited bone loss induced by LPS. This
suggests that LS suppresses bone loss induced by LPS, and that this
inhibitory effect may be due to the LS-mediated inhibition of
osteoclast differentiation and osteoblast induction.
In conclusion, the effects of LS treatment on bone
metabolism in the current study revealed a number of important
results: i) LS was not cytotoxic to the MC3T3-E1 and RAW 264.7
cells; ii) LS enhanced osteoblast cells, leading to increased
mineralization nodules; iii) LS induced the expression of
osteogenic genes and RUNX2 and BMP-2 in osteoblast precursor cells;
iv) LS inhibited osteoclast differentiation and function; v) LS
suppressed the expression of NFATc1 and c-Fos; vi) LS suppressed
the expression of MMP-9, CTK, OSCAR, c-src, c-myc, PU-1, OC-STAMP
and ATP6v0d2; vii) LS regulated bone loss in inflammatory
osteoporosis induced by LPS. Based on these results, LS may present
a potential alternative for the treatment of bone metabolic
diseases.
However, this study had the following limitations.
i) The estrogenic activity of phenol-red in medium used for in
vitro experiments was not considered (60). ii) It is also known that LS has
estrogenic activity. Since estrogenic activity can promote
osteoblast differentiation or inhibit osteoclast differentiation
(61), in the future, cell
experiments using phenol red-free medium and measurement of
estrogen hormone changes in animal model should be studied. iii)
The safety and side-effects of LS have not yet been elucidated
(62,63). In this experiment, LS did not
affect the body weight and liver weight of the animals; however,
further studies on high-dose and long-term administration are
warranted. iv) Quantitative PCR was not used for mRNA analysis.
Therefore, the amount of expression of the gene was not known, and
the increase/decrease relative to control was found. v) Considering
that LS is traditionally used in female-related diseases, it would
be valuable to examine the effects of LS in post-menopausal
osteoporosis models in the future.
Supplementary Materials
Funding
This study was supported by the National Research
Foundation of Korea (NRF) grant funded by the Korea government
(MSIT, 2017R1A2B4010163) and the Ministry of Education
(2017R1D1A1B03028505).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article or are available from the
corresponding author on reasonable request.
Authors' contributions
YS and HSJ conceptualized the study. JHK and MK
performed all experiments and verified the analytical data. JHK and
HSJ contributed to the statistical analysis and helped interpret
the results. YS supervised the experiments in discussion with JHK
and MK. JHK and MK wrote the manuscript. All authors have read and
agreed on the final manuscript.
Ethics approval and consent to
participate
All animal experiments proceeded with permission of
the Kyunghee University Animal Committee [ref. no.
KHUASP(SE)-15-095].
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Feng X and McDonald JM: Disorders of bone
remodeling. Annu Rev Pathol. 6:121–145. 2011. View Article : Google Scholar
|
|
2
|
Colon-Emeric CS and Saag KG: Osteoporotic
fractures in older adults. Best Pract Res Clin Rheumatol.
20:695–706. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Augoulea A, Tsakonas E, Triantafyllopoulos
I, Rizos D, Armeni E, Tsoltos N, Tournis S, Deligeoroglou E,
Antoniou A and Lambrinoudaki I: Comparative effects of denosumab or
bisphosphonate treatment on bone mineral density and calcium
metabolism in postmenopausal women. J Musculoskelet Neuronal
Interact. 17:444–449. 2017.PubMed/NCBI
|
|
4
|
Kennel KA and Drake MT: Adverse effects of
bisphosphonates: Implications for osteoporosis management. Mayo
Clin Proc. 84:632–637; quiz 638. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Miyazaki T, Tokimura F and Tanaka S: A
review of denosumab for the treatment of osteoporosis. Patient
Prefer Adhere. 8:463–471. 2014. View Article : Google Scholar
|
|
6
|
Liu T, Gao Y, Sakamoto K, Minamizato T,
Furukawa K, Tsukazaki T, Shibata Y, Bessho K, Komori T and
Yamaguchi A: BMP-2 promotes differentiation of osteoblasts and
chondroblasts in Runx2-deficient cell lines. J Cell Physiol.
211:728–735. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Komori T: Regulation of osteoblast
differentiation by Runx2. Adv Exp Med Biol. 658:43–49. 2010.
View Article : Google Scholar
|
|
8
|
Boyce BF and Xing L: Functions of
RANKL/RANK/OPG in bone modeling and remodeling. Arch Biochem
Biophys. 473:139–146. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim JH and Kim N: Regulation of NFATc1 in
osteoclast differentiation. J Bone Metab. 21:233–241. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Herbology Editorial Committee of Korean
Medicine: Herbology Younglimsa, Seoul. 460–461. 2004.In Korean.
|
|
12
|
Kim DH, Kim HM, Ryu JH, Um JY and Kim SC:
Korean Medical Pharmacology. 3rd edition. Shinnil Books; Seoul: pp.
507–511. 2010, In Korean.
|
|
13
|
Ju DH, Liu MJ, Zhao HY and Wang J:
Mechanisms of 'kidney governing bones' theory in traditional
Chinese medicine. Front Med. 8:389–393. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Oliveira AS, Cercato LM, de Santana Souza
MT, Melo AJO, Lima BDS, Duarte MC and Araujo AAS: The ethanol
extract of Leonurus sibiricus L. induces antioxidant,
antinociceptive and topical anti-inflammatory effects. J
Ethnopharmacol. 206:144–151. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sitarek P, Rijo P, Garcia C, Skała E,
Kalemba D, Białas AJ, Szemraj J, Pytel D, Toma M, Wysokińska H and
Śliwiński T: Antibacterial, anti-inflammatory, antioxidant, and
antiproliferative properties of essential oils from hairy and
normal roots of Leonurus sibiricus L. and their chemical
composition Oxid Med Cell Longev. 2017:73840612017.
|
|
16
|
He YL, Shi JY, Peng C, Hu LJ, Liu J, Zhou
QM, Guo L and Xiong L: Angiogenic effect of motherwort (Leonurus
japonicus) alkaloids and toxicity of motherwort essential oil on
zebrafish embryos. Fitoterapia. 128:36–42. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ginaldi L, Di Benedetto MC and De Martinis
M: Osteoporosis, inflammation and ageing. Immun Ageing. 2:142005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim JY, Cheon YH, Kwak SC, Baek JM, Yoon
KH, Lee MS and Oh J: Emodin regulates bone remodeling by inhibiting
osteoclastogenesis and stimulating osteoblast formation. J Bone
Miner Res. 29:1541–1553. 2014. View Article : Google Scholar
|
|
19
|
Kim KJ, Yeon JT, Choi SW, Moon SH, Ryu BJ,
Yu R, Park SJ, Kim SH and Son YJ: Decursin inhibits
osteoclastogenesis by downregulating NFATc1 and blocking fusion of
pre-osteoclasts. Bone. 81:208–216. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tschop MH, Speakman JR, Arch JR, Auwerx J,
Bruning JC, Chan L, Eckel RH, Farese RV Jr, Galgani JE, Hambly C,
et al: A guide to analysis of mouse energy metabolism. Nat Methods.
9:57–63. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mizutani H, Ishihara Y, Izawa A, Fujihara
Y, Kobayashi S, Gotou H, Okabe E, Takeda H, Ozawa Y, Kamiya Y, et
al: Lipopolysaccharide of Aggregatibacter actinomycetemcomitans
up-regulates inflammatory cytokines, prostaglandin E2 synthesis and
osteoclast formation in interleukin-1 receptor antagonist-deficient
mice. J Periodontal Res. 48:748–756. 2013.PubMed/NCBI
|
|
22
|
Nason R, Jung JY and Chole RA:
Lipopolysaccharide-induced osteoclastogenesis from mononuclear
precursors: A mechanism for osteolysis in chronic otitis. J Assoc
Res Otolaryngol. 10:151–160. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim JH, Kim EY, Lee B, Min JH, Song DU,
Lim JM, Eom JW, Yeom M, Jung HS and Sohn Y: The effects of Lycii
Radicis Cortex on RANKL-induced osteoclast differentiation and
activation in RAW 264.7 cells. Int J Mol Med. 37:649–658. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang L, Liu S, Mu S, Man X, Ba G, Guo R,
Li Y, Zhou L, Yang L and Fu Q: Leonurine hydrochloride promotes
osteogenic differentiation and increases osteoblastic bone
formation in ovariectomized mice by Wnt/β-catenin pathway. Biochem
Biophys Res Commun. 504:941–948. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang D, Christensen K, Chawla K, Xiao G,
Krebsbach PH and Franceschi RT: Isolation and characterization of
MC3T3-E1 preosteoblast subclones with distinct in vitro and in vivo
differentiation/mineralization potential. J Bone and Miner Res.
14:893–903. 1999. View Article : Google Scholar
|
|
26
|
Collin-Osdoby P and Osdoby P:
RANKL-mediated osteoclast formation from murine RAW 264.7 cells.
Methods Mol Biol. 816:187–202. 2012. View Article : Google Scholar
|
|
27
|
Bills CE, Eisenberg H and Pallante SL:
Complexes of organic acids with calcium phosphate: The von Kossa
stain as a clue to the composition of bone mineral. Johns Hopkins
Med J. 128:194–207. 1971.PubMed/NCBI
|
|
28
|
Gregory CA, Gunn WG, Peister A and Prockop
DJ: An Alizarin red-based assay of mineralization by adherent cells
in culture: Comparison with cetylpyridinium chloride extraction.
Anal Biochem. 329:77–84. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ogasawara T, Kawaguchi H, Jinno S, Hoshi
K, Itaka K, Takato T, Nakamura K and Okayama H: Bone morphogenetic
protein 2-induced osteoblast differentiation requires Smadmediated
down-regulation of Cdk6. Mol Cell Biol. 24:6560–6568. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jao HY, Hsu JD, Lee YR, Lo CS and Lee HJ:
Mulberry water extract regulates the osteoblast/osteoclast balance
in an ovariectomic rat model. Food Funct. 7:4753–4763. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shim KS, Lee CJ, Yim NH, Gu MJ and Ma JY:
Alpinia officinarum stimulates osteoblast mineralization and
inhibits osteoclast differentiation. Am J Chin Med. 44:1255–1271.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Golub EE, Harrison G, Taylor AG, Camper S
and Shapiro IM: The role of alkaline phosphatase in cartilage
mineralization. Bone Miner. 17:273–278. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gordon JA, Tye CE, Sampaio AV, Underhill
TM, Hunter GK and Goldberg HA: Bone sialoprotein expression
enhances osteoblast differentiation and matrix mineralization in
vitro. Bone. 41:462–473. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Morinobu M, Ishijima M, Rittling SR, Tsuji
K, Yamamoto H, Nifuji A, Denhardt DT and Noda M: Osteopontin
expression in osteoblasts and osteocytes during bone formation
under mechanical stress in the calvarial suture in vivo. J Bone
Miner Res. 18:1706–1715. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang W, Carlsen B, Rudkin G, Berry M,
Ishida K, Yamaguchi DT and Miller TA: Osteopontin is a negative
regulator of proliferation and differentiation in MC3T3-E1
pre-osteoblastic cells. Bone. 34:799–808. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rosset EM and Bradshaw AD:
SPARC/osteonectin in mineralized tissue. Matrix Biol. 52-54:78–87.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Delany AM, Kalajzic I, Bradshaw AD, Sage
EH and Canalis E: Osteonectinnull mutation compromises osteoblast
formation, maturation, and survival. Endocrinology. 144:2588–2596.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hurley MM, Abreu C, Harrison JR, Lichtler
AC, Raisz LG and Kream BE: Basic fibroblast growth factor inhibits
type I collagen gene expression in osteoblastic MC3T3-E1 cells. J
Biol Chem. 268:5588–5593. 1993.PubMed/NCBI
|
|
39
|
Hayman AR: Tartrate-resistant acid
phosphatase (TRAP) and the osteoclast/immune cell dichotomy.
Autoimmunity. 41:218–223. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sheu TJ, Schwarz EM, Martinez DA, O'Keefe
RJ, Rosier RN, Zuscik MJ and Puzas JE: A phage display technique
identifies a novel regulator of cell differentiation. J Biol Chem.
278:438–443. 2003. View Article : Google Scholar
|
|
41
|
Marchisio PC, Cirillo D, Naldini L,
Primavera MV, Teti A and Zambonin-Zallone A: Cell-substratum
interaction of cultured avian osteoclasts is mediated by specific
adhesion structures. J Cell Biol. 99:1696–1705. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Iotsova V, Caamano J, Loy J, Yang Y, Lewin
A and Bravo R: Osteopetrosis in mice lacking NF-kappaB1 and
NF-kappaB2. Nat Med. 3:1285–1289. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Miyazaki T, Katagiri H, Kanegae Y,
Takayanagi H, Sawada Y, Yamamoto A, Pando MP, Asano T, Verma IM,
Oda H, et al: Reciprocal role of ERK and NF-kappaB pathways in
survival and activation of osteoclasts. J Cell Biol. 148:333–342.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
David JP, Sabapathy K, Hoffmann O,
Idarraga MH and Wagner EF: JNK1 modulates osteoclastogenesis
through both c-Jun phosphorylation-dependent and -independent
mechanisms. J Cell Sci. 115:4317–4325. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Matsumoto M, Kogawa M, Wada S, Takayanagi
H, Tsujimoto M, Katayama S, Hisatake K and Nogi Y: Essential role
of p38 mitogen-activated protein kinase in cathepsin K gene
expression during osteoclastogenesis through association of NFATc1
and PU.1. J Biol Chem. 279:45969–45979. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Matsumoto M, Sudo T, Saito T, Osada H and
Tsujimoto M: Involvement of p38 mitogen-activated protein kinase
signaling pathway in osteoclastogenesis mediated by receptor
activator of NF-kappa B ligand (RANKL). J Biol Chem.
275:31155–31161. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Grigoriadis AE, Wang ZQ, Cecchini MG,
Hofstetter W, Felix R, Fleisch HA and Wagner EF: c-Fos: A key
regulator of osteoclast-macrophage lineage determination and bone
remodeling. Science. 266:443–448. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Takayanagi H, Kim S, Koga T, Nishina H,
Isshiki M, Yoshida H, Saiura A, Isobe M, Yokochi T, Inoue J, et al:
Induction and activation of the transcription factor NFATc1 (NFAT2)
integrate RANKL signaling in terminal differentiation of
osteoclasts. Dev Cell. 3:889–901. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sundaram K, Nishimura R, Senn J, Youssef
RF, London SD and Reddy SV: RANK ligand signaling modulates the
matrix metal-loproteinase-9 gene expression during osteoclast
differentiation. Exp Cell Res. 313:168–178. 2007. View Article : Google Scholar
|
|
50
|
Troen BR: The role of cathepsin K in
normal bone resorption. Drug News Perspect. 17:19–28. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Miyazaki T, Tanaka S, Sanjay A and Baron
R: The role of c-Src kinase in the regulation of osteoclast
function. Mod Rheumatol. 16:68–74. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Battaglino R, Kim D, Fu J, Vaage B, Fu XY
and Stashenko P: C-myc is required for osteoclast differentiation.
J Bone and Miner Res. 17:763–773. 2002. View Article : Google Scholar
|
|
53
|
Sharma SM, Bronisz A, Hu R, Patel K,
Mansky KC, Sif S and Ostrowski MC: MITF and PU.1 recruit p38 MAPK
and NFATc1 to target genes during osteoclast differentiation. J
Biol Chem. 282:15921–15929. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kim JH, Kim K, Jin HM, Youn BU, Song I,
Choi HS and Kim N: Upstream stimulatory factors regulate OSCAR gene
expression in RANKL-mediated osteoclast differentiation. J Mol
Biol. 383:502–511. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Choi BY, Park CH, Na YH, Bai HW, Cho JY
and Chung BY: Inhibition of RANKL-induced osteoclast
differentiation through the downregulation of c-Fos and NFATc1 by
Eremochloa ophiuroides (centipedegrass) extract. Mol Med Rep.
13:4014–4022. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Han SY and Kim YK: Berberine suppresses
RANKL-induced osteoclast differentiation by inhibiting c-fos and
NFATc1 expression. Am J Chin Med. 47:439–455. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lee SH, Rho J, Jeong D, Sul JY, Kim T, Kim
N, Kang JS, Miyamoto T, Suda T, Lee SK, et al: v-ATPase V0 subunit
d2-deficient mice exhibit impaired osteoclast fusion and increased
bone formation. Nat Med. 12:1403–1409. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yang M, Birnbaum MJ, MacKay CA,
Mason-Savas A, Thompson B and Odgren PR: Osteoclast stimulatory
transmembrane protein (OC-STAMP), a novel protein induced by RANKL
that promotes osteoclast differentiation. J Cell Physiol.
215:497–505. 2008. View Article : Google Scholar
|
|
59
|
Miyaura C, Inada M, Matsumoto C, Ohshiba
T, Uozumi N, Shimizu T and Ito A: An essential role of cytosolic
phospho-lipase A2alpha in prostaglandin E2-mediated bone resorption
associated with inflammation. J Exp Med. 197:1303–1310. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Welshons WV, Wolf MF, Murphy CS and Jordan
VC: Estrogenic activity of phenol red. Mol Cell Endocrinol.
57:169–178. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Khosla S, Oursler MJ and Monroe DG:
Estrogen and the skeleton. Trends Endocrinol Metab. 23:576–581.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Geller SE and Studee L: Contemporary
alternatives to plant estrogens for menopause. Maturitas. 55(Suppl
1): pp. S3–S13. 2006, View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Tao J, Zhang P, Liu G, Yan H, Bu X, Ma Z,
Wang N, Wang G and Jia W: Cytotoxicity of Chinese motherwort
(YiMuCao) aqueous ethanol extract is non-apoptotic and estrogen
receptor independent on human breast cancer cells. J
Ethnopharmacol. 122:234–239. 2009. View Article : Google Scholar : PubMed/NCBI
|