Introduction
Glioblastoma (GBM) is the most malignant type of
brain tumor in humans, and ~3,000 incident cases are diagnosed in
Germany every year. Even with the highest level of care and use of
novel therapeutic strategies, including tumor treating fields, the
median survival of GBM patients is <20 months (1), and survival times have only
marginally improved during the previous decade. Therefore, the
development of novel therapeutic strategies is necessary. The high
levels of malignancy of GBM is a result of its characteristic
behaviors, including immunosuppression, and high levels of
resistance towards irradiation, most chemotherapeutics and hypoxia.
In addition, the tumor exhibits extensive neo-angiogenesis and
grows invasively into the healthy brain tissue, making complete
surgical resection impossible. Brain tumor initiating cells
(BTICs), also termed glioma stem cell like cells, are an extremely
resistant subpopulation of GBM cells that are considered to be
responsible for tumor initiation and recurrence (2). It has also been demonstrated that
temozolomide (TMZ), the standard chemotherapeutic drug used to
treat GBM, leads to a dedifferentiation of GBM cells towards a more
stem cell like phenotype (3).
Involved in this processes are signaling cascades including the
PI3K/AKT and RAS/mitogen active protein kinase pathway, which are
constitutively activated in GBM cells (4,5). A
downstream substrate of these pathways is Y-box binding protein-1
(YB-1), a protein that exhibits both transcriptional and
translational activity (6,7).
In a number of tumor types, YB-1 expression is correlated with
malignancy and progression (8-10).
Cellular stress, for example by DNA damage induced by irradiation
or chemotherapy, induces several post-translational YB-1
modifications including phos-phorylation, acetylation or
SUMOylation. These modifications are suggested to promote its
nuclear translocation (11-14). Subsequently, nuclear YB-1
activates the expression of multi-drug resistance
protein/P-glycoproteins, classical mediators of resistance towards
tumor treatment (15).
YB-1 is involved in the replication of therapeutic
adenoviruses. The YB-1-dependent oncolytic adenovirus XVir-N-31
developed by this study group, also termed Ad-Delo3-RGD, replicates
in tumor cells harboring nuclear YB-1 (16,17). In mice bearing R28-cell-derived
glioma, a single intratumoral injection of XVir-N-31 prolonged
survival rates (18). In ~60% of
BTICs, YB-1 is upregulated and located primarily in the nucleus,
which correlates with good virus replication and virus-mediated
lysis of these cells. However, in ~40% of BTICs, YB-1 is only
slightly expressed and located primarily in the cytoplasm, which
mitigates virus replication 100-1,000-fold (18). We demonstrated previously that
irradiation of established cancer cell lines leads to the
translocation of YB-1 into the nucleus (6,19).
Whether this also occurs in BTICs has not yet been identified. The
present study evaluated whether a therapeutically relevant
irradiation dose induced the translocation of YB-1 into the
nucleus, promoted XVir-N-31 replication and subsequently oncolysis
of BTICs in vitro and in a BTIC-derived mouse glioma model,
with aim of developing a clinically relevant therapeutic strategy
that combines irradiation and XVir-N-31-based oncovirotherapy for
patients with recurrent glioma.
Materials and methods
Cell culture
BTIC R11, R28 and R49 cell lines, kindly provided by
Dr C. Beier (University Hospital Regensburg, Center for Brain
Tumors, Regensburg, Germany), were obtained from patients with
primary GBM, as described previously (20), and were maintained as tumor
spheres in stem cell-permissive Dulbecco's modified Eagle's
medium/F12 medium (Merck KGaA) supplemented with human recombinant
epidermal growth factor (EGF; BD Biosciences), human recombinant
basic fibroblast growth factor (bFGF; R&D Systems Europe,
Ltd.), human leukemia inhibitory factor (EMD Millipore; 20 ng/ml
each) and 2% B27 supplement (Thermo Fisher Scientific, Inc.) for
preservation of the original molecular characteristics of the
tumors and to minimize differentiation. R28mCherry BTIC
were generated by transduction of R28 cells with Lenti-mCherry
(BioCat GmbH) followed by selection with puromycin. In brief,
4×105 cells were infected with pre-made lentiviral
particles encoding the fluorescent protein mCherry, at multiplicity
of infection (MOI) 5. At 24 h post-infection, the medium was
changed to fresh growth medium. At 48 h post-infection, puromycin
(2 µg/ml; Merck KGaA) was added. Selected surviving cells
were monitored for mCherry fluorescence at ×50 magnification using
a Zeiss Axio Imager Z1 fluorescence microscope (Carl Zeiss AG). For
adenoviral infection, BTIC spheres were separated into single cells
by trypsinization. Irradiation of BTICs and R28-tumor-sphere
containing brain slices was performed in a Gammacell GC40 device
[Nordion (Canada) Inc.].
Brain tissue slices and BTIC implantation.
Written informed consent was obtained from all patients, allowing
spare tissue from resective surgery to be included in the present
study. Ethical approval was obtained from the Ethics Commission of
Tübingen (approval no. 338/2016A). Tissues from healthy cortex
areas were obtained during surgery of patients with epilepsy.
Tissue preparation was performed according to previously published
protocols (21). The tissue was
carefully micro-dissected and resected with only minimal use of
bipolar forceps to ensure tissue integrity, transferred into
ice-cold artificial cerebrospinal fluid (aCSF: 110 mM choline
chloride, 26 mM NaHCO3, 10 mM D-glucose, 11.6 mM
Na-ascorbate, 7 mM MgCl2, 3.1 mM Na-pyruvate, 2.5 mM
KCl, 1.25 mM NaH2PO4, 0.5 mM
CaCl2) equilibrated with carbogene (95% O2,
5% CO2) and immediately transported to the laboratory.
Tissue was kept submerged in cool and carbogenated aCSF at all
times. Following removal of the pia, tissue chunks were trimmed
perpendicular to the cortical surface and 250-350 µm thick
slices were prepared using a Microm HM 650V vibratome (Thermo
Fisher Scientific, Inc.). Following slicing of the cortical tissue,
slices were cut into several evenly sized pieces (~5.0×2.0 mm).
Subsequently, the slices were transferred onto culture membranes of
uncoated 30 mm Millicell-CM tissue culture inserts with 0.4
µm pores (Merck KGaA) and maintained in 6-well culture
dishes (BD Biosciences). For long-term culture the plates were
stored in at 37°C, 5% CO2 and 100% humidity in human CSF
isolated from non-tumor patients, as described previously (22). Neurons were stained for 1 h at
37°C with NeuO (0.25 µM; STEMCELL Technologies, Inc.). Then,
the NeuO-containing CSF was removed and fresh CSF was added. To
calculate the total amount of cells in a tissue slice, nuclei were
stained with DAPI (Abcam), according to the manufacturer's
protocol. Z-stacks were obtained microscopically using a Zeiss Axio
Imager Z1 fluorescence microscope (Carl Zeiss AG) at ×50
magnification. Nuclei were counted using the ImageJ software
(version win63, Fiji) (23). For
BTIC implantation into the tissue slices, R28mCherry
spheres measuring 20-50 cells in size were used. Implantation was
performed manually under microscopic observation using a 10
µl pipette tip. Slices and spheres were grown in human CSF
supplemented with recombinant human bFGF and EGF (20 ng/ml each)
for up to 23 days. The CSF was changed every 3rd day. Images were
captured using Zeiss AxioVision (version SE64 Rel. 4.8.) and ZEN2
software (version 1.0; Carl Zeiss AG) on a Zeiss Axio Imager Z1
fluorescence microscope (Carl Zeiss AG) at ×50 or ×100
magnification. Quantification of tumor growth was performed using
Image J software (version win64, Fiji) (23). If the tumor area reached the
borders of the photographically detectable area, the highest
measurable value was used for quantification.
Virus preparation and infection
XVir-N-31 was prepared, purified and titrated as
previously described (18). The
transduction efficacy of R11, R28 and R49 BTICs was determined
using a green fluorescent protein (GFP)-expressing adenovirus
(Ad-RGD-GFP) that possessed the same RGD-motif-modified capsid as
XVir-N-31. Infections were performed by adding the appropriate
amount of virus into the growth medium. At 48 h post-infection, the
cells were separated into single cells by trypzination, and
analyzed for GFP expression by flow cytometry. Analyses were
performed on a Cyan ADP flow cytometer (Beckman Coulter, Inc.)
using the Summit software (version 4.3; Beckman Coulter, Inc.).
The MOI of BTICs using XVir-N-31 was optimized for
each in vitro experiment. Infections were performed, as
described above. After 72 h, cells and medium were collated and
freeze-thaw lysates were prepared. Viral titers in these lysates
were determined in 293 cells (Microbix Biosystems, Inc.) using the
Adeno-Rapid-X-Titration kit (Takara Bio Europe SAS).
Western blot analysis
Total cell lysates were prepared as described
previously (18). Nuclear and
cytoplasmic protein lysates were prepared according to the protocol
described by Toulany et al (6). Western blot analysis was performed
as previously described (18).
The following antibodies were used: Anti-YB-1 (1:1,000, cat. no.
SC-101198, Santa Cruz Biotechnology, Inc.; or cat. no. 9744, Cell
Signaling Technology, Inc.), anti-Histone H3 (1:10,000; cat. no.
39163; Active Motif), anti-calreticulin (1:5,000; cat. no. C7492;
Sigma-Aldrich; Merck KGaA) and GAPDH (1:1,000; cat. no. MAB374;
Merck KGaA). Immunoreactive specific proteins were detected using
the CLARITY Western ECL substrate on a ChemiDoc MP imaging device
using the ImageLab 5.1 software (all from Bio-Rad Laboratories,
Inc.). For quantification of YB-1 expression, Image J (version
win64; Fiji) (23) was used.
Intracranial tumor model
A total of 1×105 R28 cells were implanted
under peritoneally applied anesthesia (0.05 mg/kg fentanyl, 5 mg/ml
midazolam and 0.5 mg/kg medetomin) (24) in the right striatum of 5-6 week
old female Rj:NMRI Foxn1nu/Foxn1nu nude mice
(Janvier Labs) using a mouse stereotactic device with automated
infusion pump (Stoelting Co) and a 10 µl Hamilton syringe
(Type 701SN; Medchrom Medizinischer Chromatographiebedarf GmbH).
Carprofen (5 mg/kg) was injected subcutaneously as an analgesic.
Following surgery, anesthesia was antagonized by a subcutaneous
injection of naloxon (1.2 mg/kg), flumazenil (0.5 mg/kg) and
atipamezol (2.5 mg/kg). Mice were housed in IVC cages under
standard pathogen-free conditions in the animal facility of the
Hertie Institute (Tübingen, Germany). Animal health and behavior
was monitored 3 times per week. At day 11 after tumor cell
implantation, the 30 tumor cell-injected mice were randomly split
into four groups (n=5-7 animals per group). A total of two groups
were tumor-irradiated with 3 Gy (6 MV photons) using a LINAC linear
accelerator as described previously (25) and received an intratumoral
injection of either PBS (sham) or 1.5×108 infectious
units (IFU) XVir-N-31 the next day. One group received only
XVir-N-31 and one group received an intracranial PBS injection
(sham). The mice were euthanized by lethal CO2
inhalation (10-30 volume percentage discharge of CO2/per
minute in their homing cage) at the onset of tumor-related
symptoms. Human endpoints were defined according to the score sheet
of regional council approval N12/14 and include the early onset of
hemi- or paraplegia, paroxism, gait disorder, or loss of body
weight of >19%. Euthanasia was confirmed by the lack of
respiration and faded eye color. After 150 days the experiment was
terminated by euthanasia of the surviving animals. All animal
research was performed in accordance with the German Animal Welfare
Act and its guidelines for the care and use of laboratory animals
(https://www.gesetze-im-internet.de/tierschg/BJNR012770972.html)
and was approved by the regional council of Tübingen
(Regierungspräsidium Tübingen, approval no. N12/14).
Statistical analysis
For the in vitro assays, results were
calculated as means ± standard deviation (SD) and compared using
the one-way analysis of variance, followed by Bonferroni post-hoc
correction for multiple testing using GraphPad Prism software
(version 7.04; GraphPad Software, Inc.). If not otherwise
indicated, the data were from at least 3 repeat experiments. For
the in vivo assays, survival and mean or median survival
times were analysed using the Kaplan-Meier method and compared
using a log-rank test with JMP v13 software (SAS Institute Inc.),
followed by Bonferroni correction for multiple testing. P<0.05
was considered to indicate a statistically significant
difference.
Results
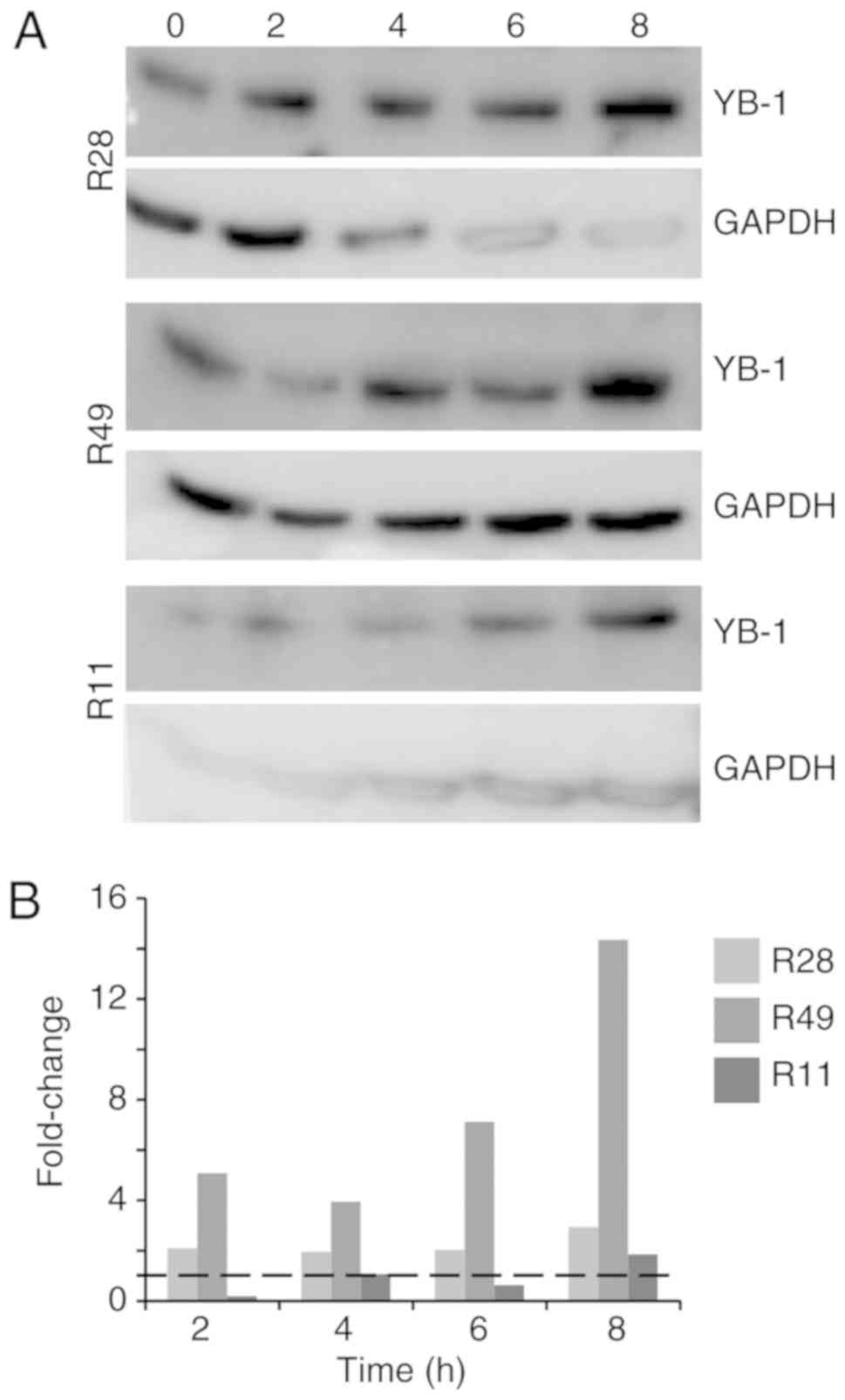
Irradiation induces YB-1 expression, its
nuclear translocation and promotes XVir-N-31 production in
BTICs
For established glioma and breast cancer cell lines,
it has been demonstrated that irradiation leads to the
translocation of YB-1 into the nucleus (6,19).
The present study examined whether this was also true in
TMZ-resistant BTICs (18). Using
3 different BTIC lines, we identified an upregulation of total YB-1
in the cells 8 h after irradiation, however to a different extent
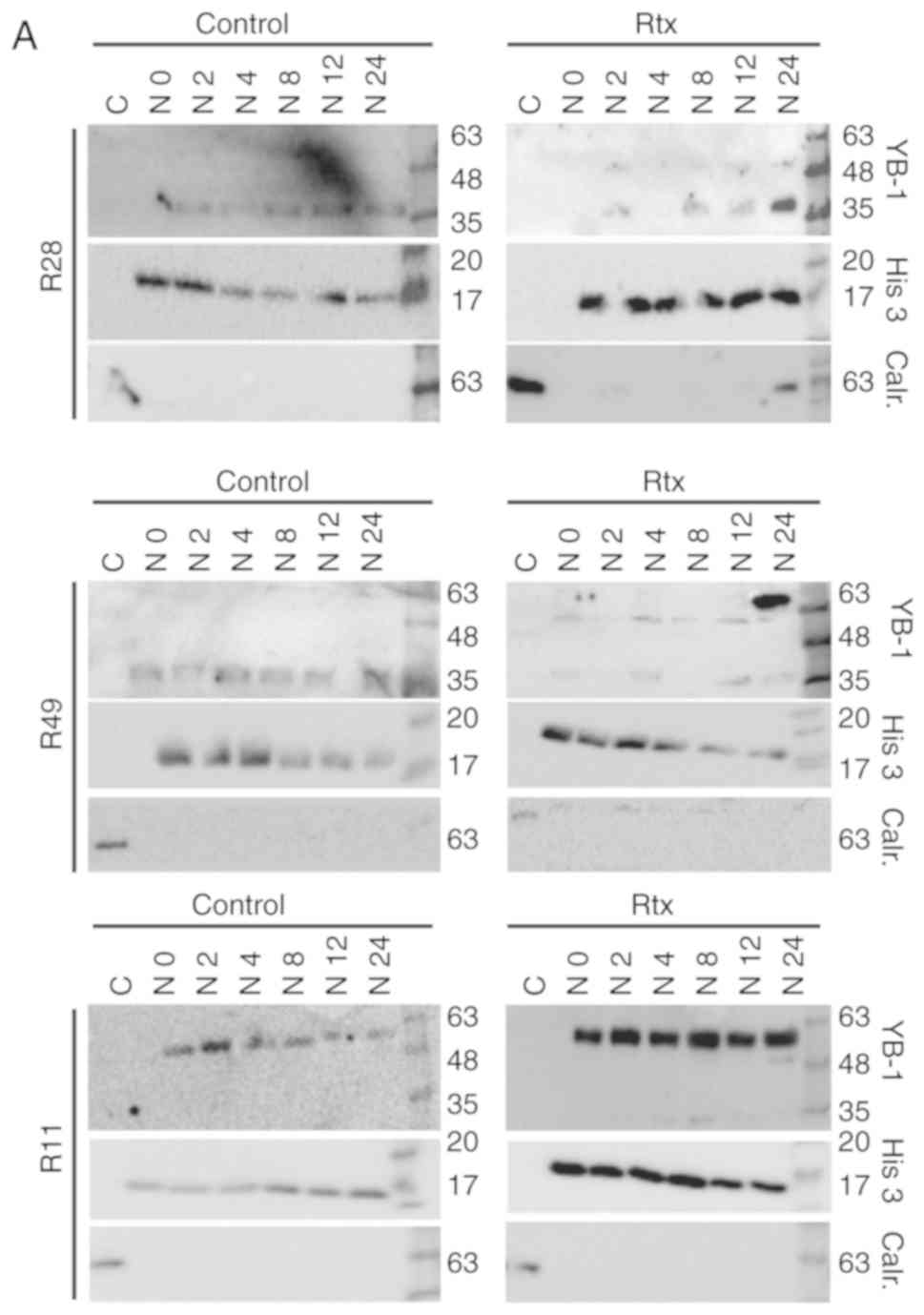
(Fig. 1). Using nuclear and
cytoplasmic cell extracts, an additional, higher molecular weight
YB-1 band (YB-160 kD) was detected in the nuclear
fraction of irradiated R28 and R49 cells, but not in non-irradiated
control cells. In the R11 cells, this band was present in the
nuclear extracts of both irradiated and non-irradiated cells. In
R49 cells, an additional YB-1 protein variant (YB-165
kD) was detected at a high level at 24 h after irradiation
(Fig. 2A). As it has been
established that under stress stimuli, like irradiation, the YB-1
protein will be post-translationally modified, all YB-1 bands were
used to quantify the amount of nuclear YB-1 (Fig. 2B). Nuclear YB-1 levels were
enhanced in both irradiated R28 and R49 cells. However, doses of
only 2 Gy were sufficient to relocate YB-1 into the nucleus in R28
cells whilst doses of 6 Gy were required for nuclear translocation
of YB-1 in R49 cells (Fig. 2 and
data not shown). Notably, in R49 cells, YB-165 kD, a
highly post-translationally modified YB-1, was prominent at high
levels and was not observed in R11 nor in R28 cells. No relevant
elevated nuclear YB-1 levels and no YB-165 kD bands were
observed in R11 cells, which harbor a high basal level of nuclear
YB-1 in contrast to R28 and R49 cells (Fig. 2). R11 and R28 cells were also
irradiated using a dose of 6 Gy. However, this irradiation dose did
not increase the level of nuclear YB-1 in R11 nor in R28 cells, and
even induced cell death in R28 cells (data not shown).
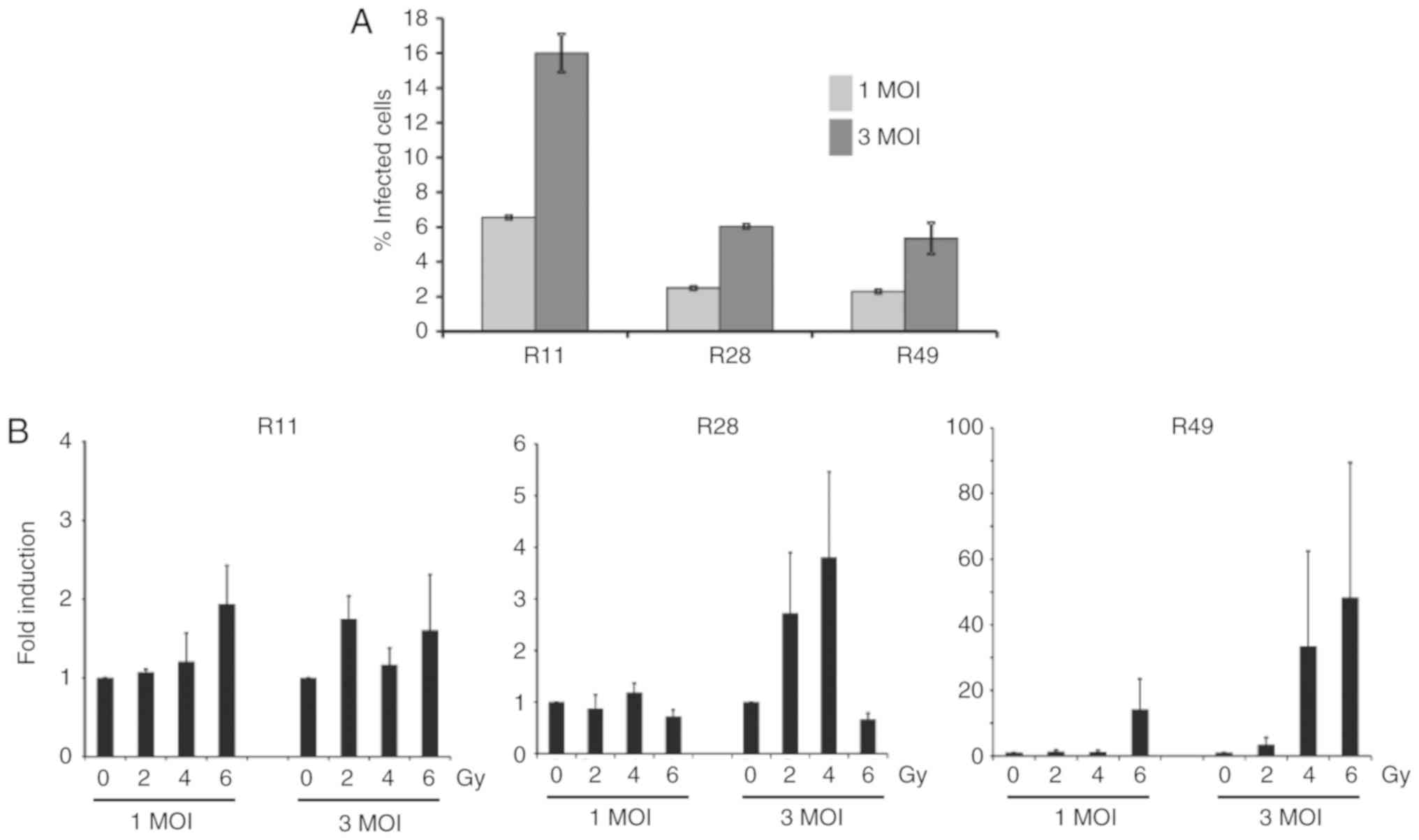
To determine the effect of nuclear YB-1 on XVir-N-31
production, the adenoviral transduction efficacy of glioma stem
cells was first examined, which was 6.55±0.1% at 1 MOI and
16.0±1.1% at 3 MOI for R11 cells, 2.5±0.1% at 1 MOI and 6.05±0.1%
at 3 MOI for R28 cells and 2.3±0.1% at 1 MOI and 5.35±0.9% for R49
cells (Fig. 3A). To examine
whether irradiation enhanced XVir-N-31 production, the cells were
irradiated 24 h prior to XVir-N-31 infection. For infection, only
the aforementioned low infection rates were used, as it has been
established that XVir-N-31 replicates efficiently in glioma cells
at a dose of 5 MOI (19).
Although the levels were not significant due to a high variability
in virus production in the independent experiments, the nuclear
translocation of YB-1 in R28 and R49 cells, and in particular the
level of nuclear YB-165 kD, correlated well with the
enhanced production of XVir-N-31 in these cells. Whilst combined
irradiation and infection of R28 and R49 cells using 1 MOI of
XVir-N-31 exhibited almost no enhancement in virus production, the
increase of XVir-N-31 production in irradiated R49 cells infected
with 3 MOI was 1.6-130-fold, depending on the irradiation dose, and
was superior to that observed in R28 cells (1.2-7-fold; Fig. 3B; Table I). In R28 cells, a dose of 6 Gy of
irradiation led to cell death (data not shown), paralleled by a
decrease in virus production (Fig.
3). As expected, in irradiated R11 BTICs that exhibited high
basal nuclear YB-1 expression but no relevant irradiated-mediated
nuclear transfer of YB-1, no significant enhancement of XVir-N-31
production was detected.
 | Table IXVir-N-31 production in irradiated
brain tumor initiating cells. |
Table I
XVir-N-31 production in irradiated
brain tumor initiating cells.
| 2 | 4 | 6 | 2 | 4 | 6 | Gy |
|---|
| Cell lines | 1 | 1 | 1 | 3 | 3 | 3 | MOI |
|---|
| R28 | 0.4-1.4 | 0.8-1.4 | 0.5-0.9 | 1.2-5.0 | 1.6-7.0 | 0.5-0.9 | - |
| R49 | 0.5-2.1 | 0.4-1.2 | 0.4-31.8 | 1.0-7.5 | 1.6-91.4 | 4.9-130.4 | - |
| R11 | 1.0-1.3 | 0.6-1.8 | 1.1-2.8 | 1.4-2.0 | 0.8-1.5 | 0.7-3.0 | - |
Irradiation in combination with XVir-N-31
infection significantly decreases the growth of
R28mCherry spheres implanted in human cortical brain
slices
To analyze the effect of a combined irradiation and
XVir-N-31-based oncovirotherapy in therapy-resistant BTICs, an
organotypic in vitro glioma model was established, in which
R28mCherry spheres were implanted in human cortical
brain tissue slices. The slices were irradiated with either 3 Gy,
infected with 5×107 IFU of XVir-N-31 or were irradiated
and infected 24 h later. Tumor sphere growth was quantified over a
period of 14 days. A longer period of quantification was not
possible, as most tumor sphere sizes expanded the microscopic
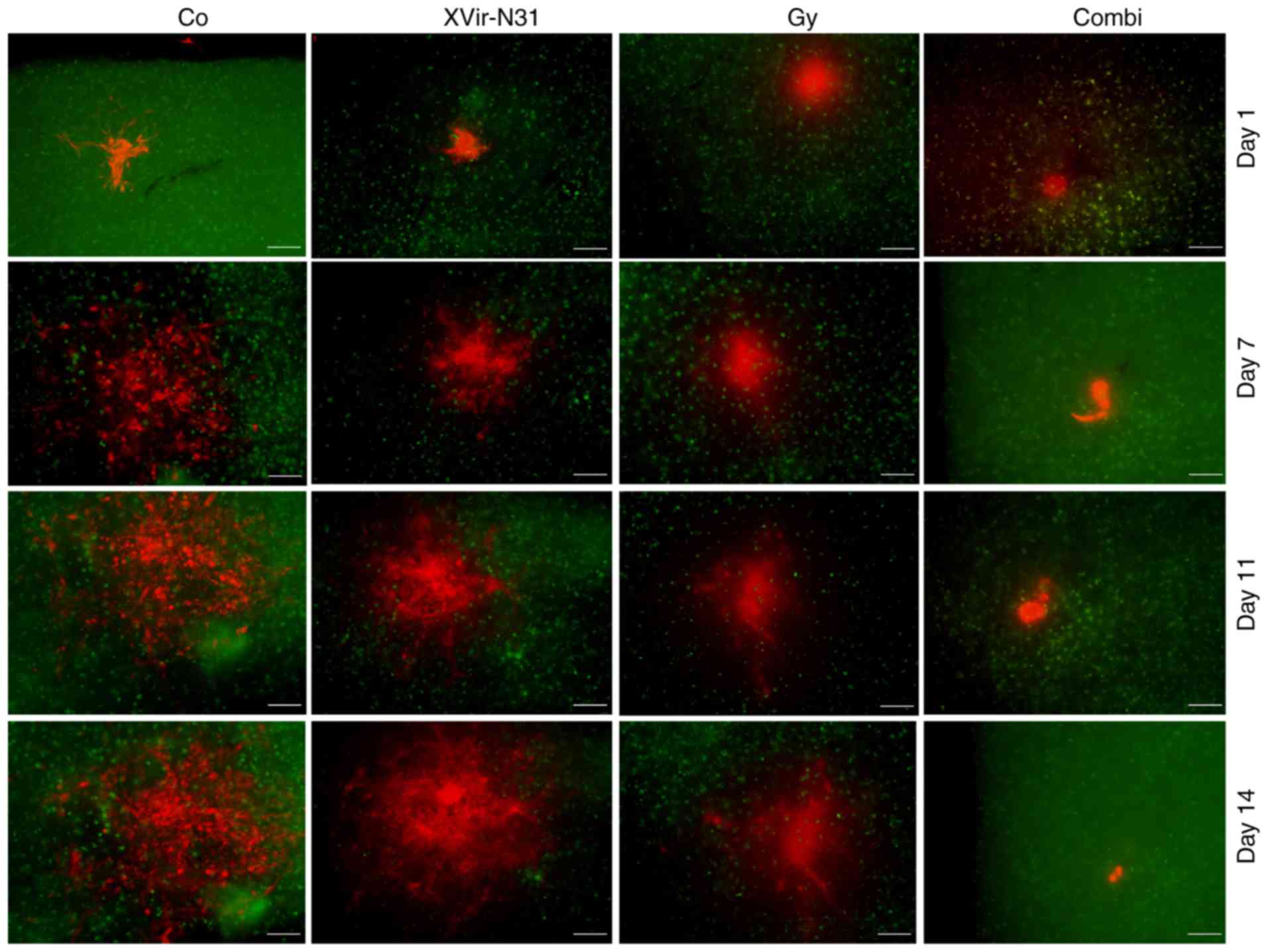
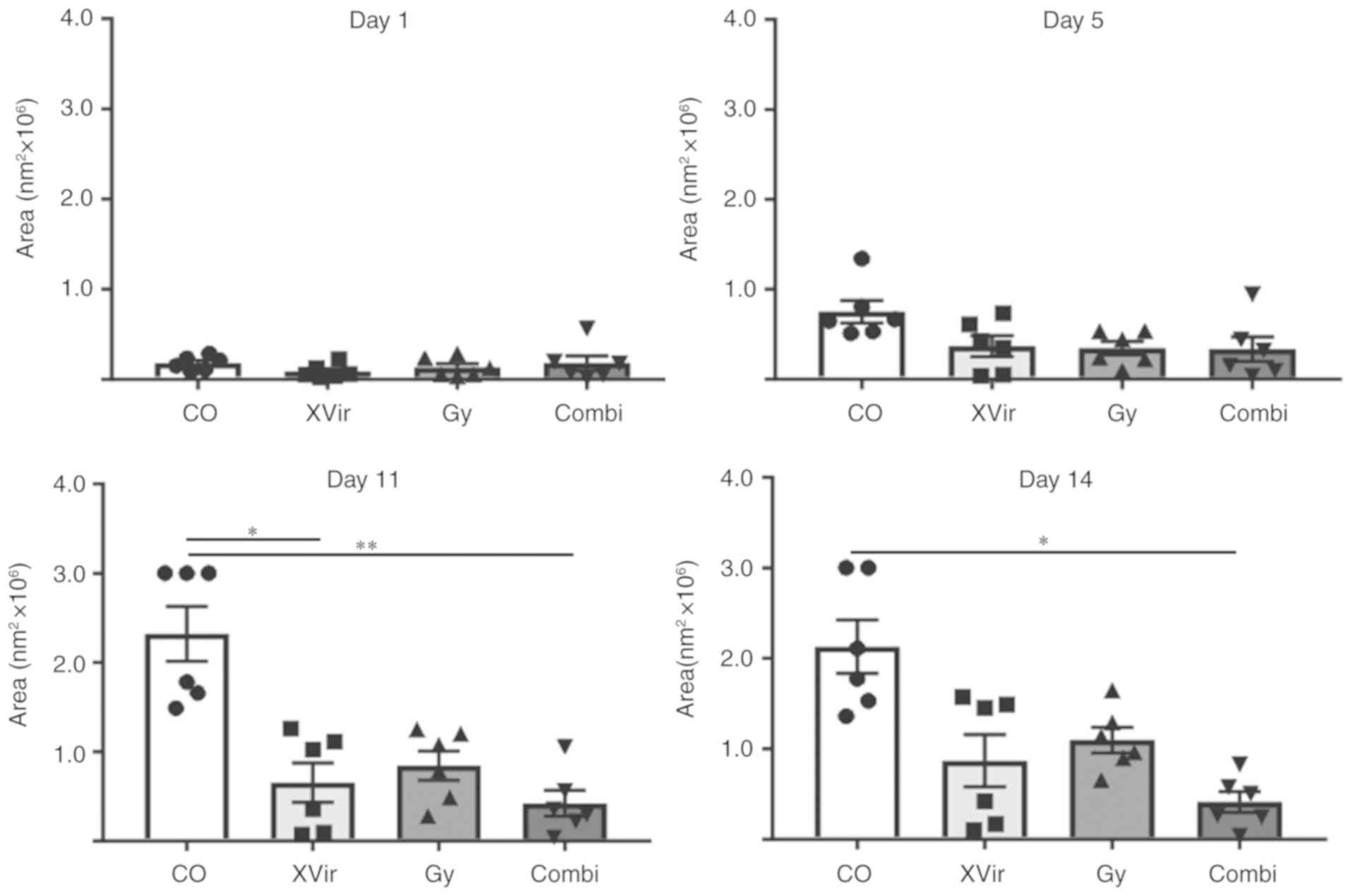
observation area after this time point. As indicated in Figs. 4 and 5, all (6/6) untreated control
R28mCherry tumor spheres grew quickly, whilst
irradiation or infection alone resulted in a decrease in sphere
growth rate. By single infection with XVir-N-31, the growth of 3/6
spheres was almost completely inhibited, whilst 3/6 spheres
continued to grow, but to a lesser extent compared with the
controls. Nevertheless, in this group a number of tumor cells
exhibited the typical characteristic of an oncolytic adenovirus
infection, including decreased cell size and a rounded cell shape,
indicating that these cells were dying (Fig. 6). When irradiation was used as a
single treatment, all spheres (6/6) were delayed in growth.
However, surviving tumor cells grew in a highly invasive manner
into the brain tissue (Fig. 6).
By contrast, combined therapy was very efficient and significantly
delayed tumor sphere growth; 1/6 spheres was nearly completely
eradicated upon treatment, 2/6 spheres exhibited a highly delayed
growth and 3/6 grew slowly. The morphology of these slow-growing
spheres was therefore assessed. As indicated in Fig. 6, in these spheres many cells
exhibited the characteristics of oncolysis, however, a population
of non-infected viable tumor cells that continued to proliferate
was visible.
Irradiation in combination with XVir-N-31
infection prolongs the median survival of R28 tumor-bearing nude
mice
The promising in vitro results prompted the
analysis of the therapeutic effect of a combined XVir-N-31-based
oncoviro-irradiation therapy in mice. For this purpose, R28 BTICs
were intracranially implanted into the striatum of nude mice. The
mice were treated as follows: i) Sham (no irradiation +
intratumoral injection of PBS); ii) irradiation (3 Gy tumor
irradiation) + intratumoral PBS injection 24 h later; iii)
XVir-N-31 (no irradiation + intratumoral injection of
1.5×108 IFU XVir-N-31). This virus amount represents
one-half of the recently determined effective therapeutic dose
(18); iv) Combi [3 Gy tumor
irradiation + an intratumoral XVir-N-31 injection
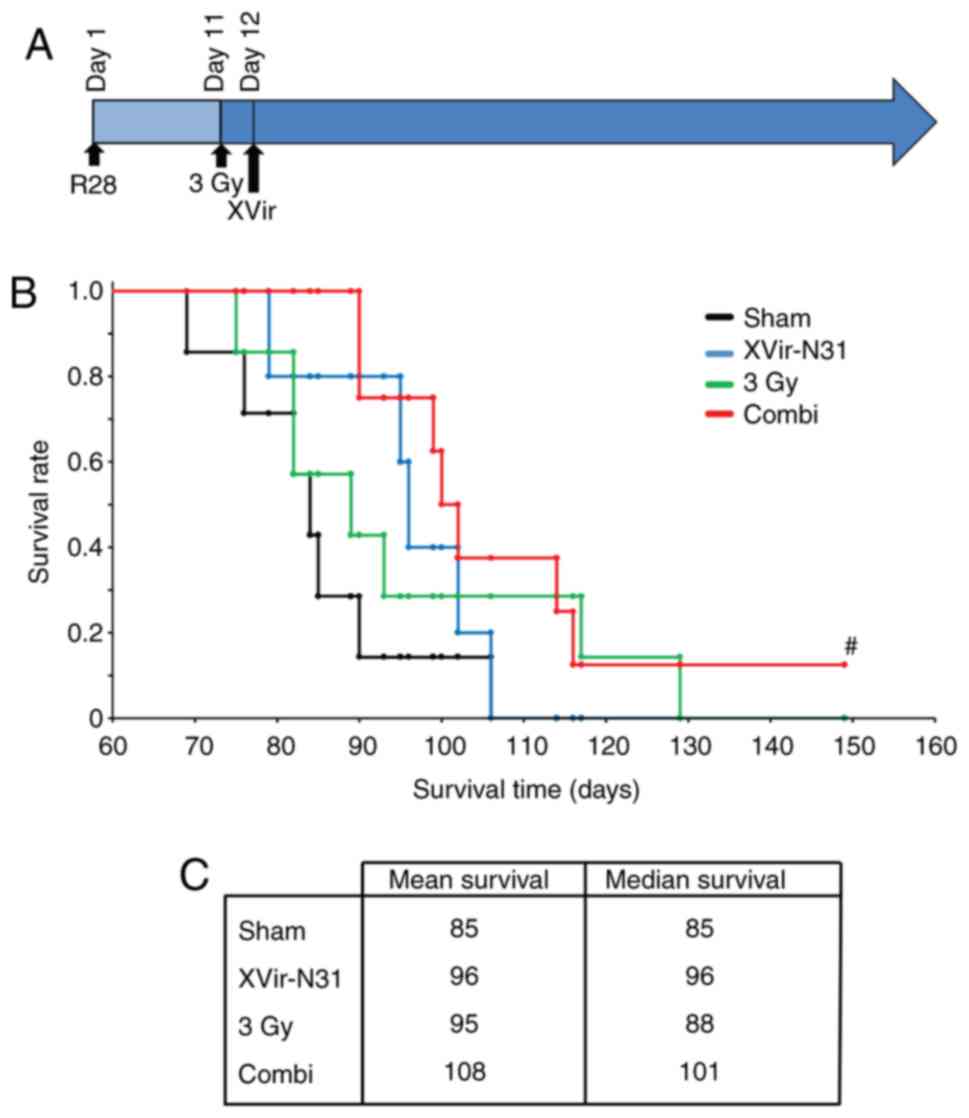
(1.5×108 IFU) 24 h later; Fig. 7A]. A suboptimal dose of XVir-N-31
was selected for the XVir-N-31 and Combi groups to identify any
possible augmentation of the oncolytic effect of XVir-N-31 by tumor
irradiation prior to infection. Large tumors were observed visually
in the sham treated mice (data not shown), and these animals
succumbed at around day 85. The mean survival of the single
treatment groups was increased compared with that of the controls,
but was almost the same for both mono-therapies (95 days for
irradiation and 96 days for XVir-N-31 infection groups,
respectively). The median survival of the virus-treatment group was
improved (96 days) compared with that of the irradiated group (88
days), but did not reach significance. As expected, based on
previous data, oncovirotherapy using a suboptimal amount of
infectious XVir-N-31 particles (1.5×108 IFU) did not
reach the median survival of ~120 days, as was observed previously
when using the double amount of XVir-N-31 (3×108 IFU)
(18). Nevertheless, there was a
trend towards a prolonged survival in the group of mice subjected
to tumor irradiation followed by oncovirotherapy, although this did
not reach significance. The median survival in this group was 101
days, and the mean survival 108 days. One mouse within this group
did not exhibit any tumor-associated symptoms, even at a period of
150 days after tumor implantation. This mouse was sacrificed, but
no visible tumors were detected (data not shown).
Discussion
The results of the present study demonstrated that
irradiation of BTICs enhanced the lytic activity of the
YB-1-dependent oncolytic adenovirus XVir-N-31. BTICs are highly
therapy-resistant cells that exhibit stem cell characteristics, and
have been proposed as the primordial cells responsible for disease
initiation and its recurrence (2). Due to the fact that not many
therapeutic options are available to efficiently treat recurrent
glioma, the results of the present study may have substantial
consequences on the development of future combined treatment
strategies. We previously determined the effects of XVir-N-31 in
BTICs and demonstrated that XVir-N-31 replicated in BTICs. In a
BTIC-derived mouse glioma model, treatment of mice with a single
intratu-moral injection of XVir-N-31 significantly prolonged their
survival (18). XVir-N-31
replication is dependent on the presence of nuclear YB-1 (19). YB-1 is primarily expressed in all
high grade glioma, conveys resistance towards TMZ, the standard
chemotherapeutic drug to treat GBM, and serves as a prognostic
biomarker for tumor progression (18,26,27). However, in some BTICs only a small
amount of YB-1 was identified to be nuclear (18), therefore virus replication is
suboptimal. In previous years, several studies have demonstrated
that YB-1, in response to genotoxic stress, becomes
post-translationally modified and is transferred from the cytoplasm
into the nucleus. This in turn enhances the replication and
oncolytic effect of XVir-N-31 in established tumor cell lines and
experimental glioma models (6,19,28).
Since oncovirotherapy is still not a standardized
therapy option to treat glioma, it is expected to be primarily used
as second line therapy for recurrent glioma. As it has been
established that the irradiation of established glioma cell lines
induced the translocation of YB-1 into the nucleus and supported
XVir-N-31 replication, we hypothesized that irradiation prior to
oncovirotherapy may augment the therapeutic effect of a
XVir-N-31-based oncovirotherapy in patients with recurrent glioma.
To examine this hypothesis, the effect of irradiation on the
subcellular localization of YB-1 in BTICs, the primary target cells
in and drivers of recurrent glioma, was investigated. Elevated
nuclear YB-1 was detected in R28 and R49 BTICs, however, in R11
cells no irradiation-mediated nuclear translocation of YB-1 was
observed. In the present study, in contrast to R28 and R49 BTICs,
R11 cells were observed to possess a high basal level of nuclear
YB-1. This observation is not concordant with previous data that
revealed the presence of both cytoplasmic and nuclear YB-1 in R11
cells (18), but this may be
explained by the use of different YB-1 antibodies between these
studies. Nevertheless, the high basal level of nuclear YB-1 in R11
BTICs may explain why these cells possess a higher basal XVir-N-31
replication rate compared with the R28 and R49 cells, why there is
no additional increase in nuclear YB-1 levels detected following
irradiation of these cells, and why irradiation did not enhance
XVir-N-31 production.
In the present study, R49 cells required a higher
irradiation dose (6 Gy) to initiate the transfer of YB-1 into the
nucleus. In concordance with the observed elevated nuclear level of
YB-1 in R49 and R28 BTICs, irradiation prior to infection
facilitated XVir-N-31-production in these cells up to 130-fold.
However, induction of virus production was variable. As YB-1 serves
as a stress protein, its posttranslational modification and nuclear
translocation, and consequently its activity as a transcription
factor that is necessary for XVir-N-31 replication, is very
sensitive to external conditions. Additionally, virus production
itself is a complex process including transcription of viral
proteins, DNA replication, and the self-assembly of viral proteins
and the genome to create infectious viral particles. This may
explain the variability of virus production observed in the
irradiated R28 and R49 cells.
Notably, in the R49 cells, which exhibited superior
XVir-N-31 production following irradiation compared with the R28
cells, the presence of nuclear YB-165 kD, which was
suggested to represent a highly post-translationally modified
version of YB-1, was detected. We hypothesized that not only the
nuclear translocation of YB-1, but also its post-translational
modification, is important for its function as a transcription
factor necessary for XVir-N-31 replication. Whether this
modification includes only phosphorylation or also acetylation or
SUMOylation (11-14), or even presently unknown
modifications, requires additional studies. In summary, the data
from the present study demonstrated a direct association between
nuclear (modified) YB-1 and XVir-N-31-replication and its
production in established tumor cell lines, which was in
concordance with our previous observations (19).
Subsequently, the therapeutic effect of a combined
XVir-N31-based oncoviro-radiotherapy was assessed in human cortical
brain slice cultured harboring R28mCherry glioma
spheroids as well as in R28 glioma bearing mice. Infection of slice
cultures was performed with a suboptimal dose of XVir-N-31
(5×107 IFU) to determine the synergistic effects of
irradiation and virus infection. The specific time point of virus
infection (24 h after irradiation) was selected as elevated levels
of nuclear (post-translationally modified) YB-1 were detectable
after this period. Due to the ability of XVir-N-31 to also infect
neurons, glial cells and astrocytes, the virus dose selected only
equated to ~0.1 IFU/cell. Therefore, the therapeutic effect of the
XVir-N-31-based monotherapy was only moderate. However, a delay in
R28mCherry tumor spheroid growth was detectable even at
this concentration. A total of 3/6 analyzed spheroids exhibited a
marked delay in growth, and the others a moderate delay. The
variance in growth rates may not only be caused by virus infection,
but also by the variable size of the implanted spheroids. When
examined more closely, an oncolytic cytopathic effect and dying
BTICs were observed in the XVir-N-31-infected tissues, as indicated
by a rounded cell shape and the occurrence of cell debris in the
spheroids (Fig. 6). Irradiation
also delayed the growth of BTICs, however no dead cells were
observed and the surviving cells massively invaded the surrounding
brain tissue, an effect that has been described in glioma cells in
the past (29). By contrast,
irradiation in combination with XVir-N-31-infection significantly
decreased R28mCherry tumor spheroid growth. Whilst 1
spheroid did not grow at all, 2 exhibited a massive and 3 a
considerable decrease in growth. This effect appeared to be
comparable to, or even exceed, the effect of irradiation alone. The
structure of spheroids with considerable growth reduction were then
microscopically examined and a number of dead cells and cell debris
were detected. In addition, viable cells that had started to divide
again were observed in these samples, but at a later time point
following treatment (days 17-23). The outgrowth of tumor cells
following combined therapy may be an effect provoked by the low
amount of infectious XVir-N-31 used to treat the spheroids, and the
fact that certain cells may not have been infected at all. If these
cells invade the surrounding brain tissue at a faster rate compared
with virus replication in the adjacent infected tumor cells occurs,
the cancer cells will be protected from virus infection due to a
protective wall of non-neoplastic brain cells that forms a barrier
for virus distribution in which the virus is unable to
replicate.
The therapeutic effects of a XVir-N-31-based
oncovirotherapy were previously determined in vivo in a
mouse glioma model, in which it was demonstrated that
~3×108 IFU XVir-N-31 were required to significantly
decrease tumor growth, leading to a significant survival
prolongation (18). To confirm
the therapeutic effect of a combined oncoviro-radiotherapy, as was
observed in R28mCherry BTIC-derived glioma containing
organotypic brain cultures in vitro, and to demonstrate the
support of tumor irradiation on oncolysis-mediated tumor cell death
and therefore on tumor growth and survival, only a suboptimal
amount of XVir-N-31 (1.5×108 IFU) was intra-tumorally
injected in the disease model in the present study. This virus dose
was in itself not able to significantly prolong survival.
Additionally, radiotherapy was administered prior to infection by a
single 3 Gy tumor irradiation, a dose that is used during
fractional irradiation of patients with glioma (30). Although non-significant, a
prolongation of the mean survival in both monotherapy groups for
10-11 days was observed. The survival rates in the mice that
received the oncoviro-irradiation therapy instead increased by 23
days. The median survival was determined in this group, and the
combined treatment exhibited superior effects compared with the
monotherapies. The effects achieved by the combination of
tumor-irradiation and XVir-N-31-based oncovirotherapy were not
statistically significant, however suboptimal total doses of
irradiation and of XVir-N-31 were used in the therapeutic glioma
mouse model. Therefore, we hypothesize that optimizing the
therapeutic schema, for example repeated irradiation and injection
of a therapeutic relevant dose of XVir-N-31, may result in improved
effects of this combination on tumor growth and survival.
Although the interaction between irradiation and
oncolytic adenoviruses is complex, given the diversity of cellular
responses to irradiation and the diverse oncovirus-mediated
antitumoral immune effects, the irradiation-mediated
post-translational modifications of YB-1 and its nuclear transport,
due to its central role in XVir-N-31-replication, may explain the
superior effects of a combined oncoviro-radiotherapy in the glioma
mouse model established in the present study using
therapy-resistant BTICs. The results from the present study suggest
that irradiation of patients with recurrent glioma prior to
XVir-N-31 based oncovirotherapy should be performed to optimize the
effect of the oncolytic virotherapy in these patients. Finally, as
it is well known that both virus infection and tumor-irradiation
cause a broad-range immune cell activity against tumors (31,32), the combined radio-oncovirotherapy
treatment may in turn support the therapeutic outcome of patients
with recurrent GBM.
Abbreviations:
|
BTIC
|
brain tumor initiating cell
|
|
EGF
|
epithelial growth factor
|
|
bFGF
|
basic fibroblast growth factor
|
|
Gy
|
Gray
|
|
IFU
|
infectious unit
|
|
MOI
|
multiplicity of infection
|
|
TMZ
|
temozolomide
|
|
YB-1
|
Y-box binding protein-1
|
Acknowledgments
Not applicable.
Funding
The present study was supported by grants awarded to
UN by the Else-Übelmesser-Foundation.
Availability of data and materials
The datasets generated during and/or analyzed during
the current study are available from the corresponding author on
reasonable request.
Authors' contribution
RC, NS, HK, SS, SH; TW and UN contributed to the
data collection, analysis and interpretation. PH and UN contributed
to the study design. UN wrote the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
For patient data and material, written informed
consent was obtained from all patients, allowing spare tissue from
resective surgery to be included in the present study. Ethical
approval was obtained from the Ethics Commission of Tübingen
(approval no. 338/2016A). Animal experiments were performed
according to the German Animal Welfare Act and its guidelines for
the care and use of laboratory animals (approval N12/14 of the
regional council Tübingen).
Patient consent for publication
Not applicable.
Competing interests
Per S. Holm is CEO and co-founder of XVir
Therapeutics GmbH, 80335 Munich, Germany. All other authors declare
that they have no competing interests.
References
|
1
|
Stupp R, Taillibert S, Kanner A, Read W,
Steinberg D, Lhermitte B, Toms S, Idbaih A, Ahluwalia MS, Fink K,
et al: Effect of tumor-treating fields plus maintenance
temozolomide vs maintenance temozolomide alone on survival in
patients with glioblastoma: A randomized clinical trial. JAMA.
318:2306–2316. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Auffinger B, Spencer D, Pytel P, Ahmed AU
and Lesniak MS: The role of glioma stem cells in chemotherapy
resistance and glioblastoma multiforme recurrence. Expert Rev
Neurother. 15:741–752. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Auffinger B, Tobias AL, Han Y, Lee G, Guo
D, Dey M, Lesniak MS and Ahmed AU: Conversion of differentiated
cancer cells into cancer stem-like cells in a glioblastoma model
after primary chemotherapy. Cell Death Differ. 21:1119–1131. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Molina JR, Hayashi Y, Stephens C and
Georgescu MM: Invasive glioblastoma cells acquire stemness and
increased Akt activation. Neoplasia. 12:453–463. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sunayama J, Matsuda K, Sato A, Tachibana
K, Suzuki K, Narita Y, Shibui S, Sakurada K, Kayama T, Tomiyama A
and Kitanaka C: Crosstalk between the PI3K/mTOR and MEK/ERK
pathways involved in the maintenance of self-renewal and
tumorigenicity of glioblastoma stem-like cells. Stem Cells.
28:1930–1939. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Toulany M, Schickfluss TA, Eicheler W,
Kehlbach R, Schittek B and Rodemann HP: Impact of oncogenic K-RAS
on YB-1 phosphorylation induced by ionizing radiation. Breast
Cancer Res. 13:R282011. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sinnberg T, Sauer B, Holm P, Spangler B,
Kuphal S, Bosserhoff A and Schittek B: MAPK and PI3K/AKT mediated
YB-1 activation promotes melanoma cell proliferation which is
counteracted by an autoregulatory loop. Exp Dermatol. 21:265–270.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee Wu J, Yokom C, Jiang D, Cheang H,
Yorida MC, Turbin E, Berquin D, Mertens IM, Iftner PRT, et al:
Disruption of the Y-box binding protein-1 results in suppression of
the epidermal growth factor receptor and HER-2. Cancer Res.
66:4872–4879. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kosnopfel C, Sinnberg T and Schittek B:
Y-box binding protein 1-a prognostic marker and target in tumour
therapy. Eur J Cell Biol. 93:61–70. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schittek B, Psenner K, Sauer B, Meier F,
Iftner T and Garbe C: The increased expression of Y box-binding
protein 1 in melanoma stimulates proliferation and tumor invasion,
antagonizes apoptosis and enhances chemoresistance. Int J Cancer.
120:2110–2118. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Alemasova EE, Pestryakov PE, Sukhanova MV,
Kretov DA, Moor NA, Curmi PA, Ovchinnikov LP and Lavrik OI:
Poly(ADP-ribosyl)ation as a new posttranslational modification of
YB-1. Biochimie. 119:36–44. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tiwari A, Rebholz S, Maier E, Dehghan
Harati M, Zips D, Sers C, Rodemann HP and Toulany M: Stress-induced
phosphorylation of nuclear YB-1 depends on nuclear trafficking of
p90 ribosomal S6 kinase. Int J Mol Sci. 19:pii: E24412018.
View Article : Google Scholar
|
|
13
|
Ewert L, Fischer A, Brandt S, Scurt FG,
Philipsen L, Müller AJ, Girndt M, Zenclussen AC, Lindquist JA,
Gorny X and Mertens PR: Cold shock Y-box binding protein-1
acetylation status in monocytes is associated with systemic
inflammation and vascular damage. Atherosclerosis. 278:156–165.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pagano C, di Martino O, Ruggiero G, Maria
Guarino A, Mueller N, Siauciunaite R, Reischl M, Simon Foulkes N,
Vallone D and Calabrò V: The tumor-associated YB-1 protein: New
player in the circadian control of cell proliferation. Oncotarget.
8:6193–6205. 2017. View Article : Google Scholar :
|
|
15
|
Lage H, Surowiak P and Holm PS: YB-1 as a
potential target in cancer therapy. Pathologe. 2(Suppl 2):
S187–S190. 2008.In German. View Article : Google Scholar
|
|
16
|
Holm PS, Bergmann S, Jurchott K, Lage H,
Brand K, Ladhoff A, Mantwill K, Curiel DT, Dobbelstein M, Dietel M,
et al: YB-1 relocates to the nucleus in adenovirus-infected cells
and facilitates viral replication by inducing E2 gene expression
through the E2 late promoter. J Biol Chem. 277:10427–10434. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Holm PS, Lage H, Bergmann S, Jürchott K,
Glockzin G, Bernshausen A, Mantwill K, Ladhoff A, Wichert A, Mymryk
JS, et al: Multidrug-resistant cancer cells facilitate
E1-independent adenoviral replication: Impact for cancer gene
therapy. Cancer Res. 64:322–328. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mantwill K, Naumann U, Seznec J, Girbinger
V, Lage H, Surowiak P, Beier D, Mittelbronn M, Schlegel J and Holm
PS: YB-1 dependent oncolytic adenovirus efficiently inhibits tumor
growth of glioma cancer stem like cells. J Transl Med. 11:2162013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bieler A, Mantwill K, Holzmüller R,
Jürchott K, Kaszubiak A, Stärk S, Glockzin G, Lage H, Grosu AL,
Gansbacher B and Holm PS: Impact of radiation therapy on the
oncolytic adenovirus dl520: Implications on the treatment of
glioblastoma. Radiother Oncol. 86:419–427. 2008. View Article : Google Scholar
|
|
20
|
Beier D, Hau P, Proescholdt M, Lohmeier A,
Wischhusen J, Oefner PJ, Aigner L, Brawanski A, Bogdahn U and Beier
CP: CD133(+) and CD133(−) glioblastoma-derived cancer stem cells
show differential growth characteristics and molecular profiles.
Cancer Res. 67:4010–4015. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Verhoog MB, Goriounova NA, Obermayer J,
Stroeder J, Hjorth JJ, Testa-Silva G, Baayen JC, de Kock CP,
Meredith RM and Mansvelder HD: Mechanisms underlying the rules for
associative plasticity at adult human neocortical synapses. J
Neurosci. 33:17197–17208. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schwarz N, Hedrich UBS, Schwarz H, P A H,
Dammeier N, Auffenberg E, Bedogni F, Honegger JB, Lerche H, Wuttke
TV and Koch H: Human Cerebrospinal fluid promotes long-term
neuronal viability and network function in human neocortical
organotypic brain slice cultures. Sci Rep. 7:122492017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schindelin J, Arganda-Carreras I, Frise E,
Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S,
Schmid B, et al: Fiji: An open-source platform for biological-image
analysis. Nat Methods. 9:676–682. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Erhardt W, Henke J and Haberstroh J:
Anästhesie und Analgesie beim klein-und heimtier. Schattauer
Verlag; 2004
|
|
25
|
Mutschelknaus L, Azimzadeh O, Heider T,
Winkler K, Vetter M, Kell R, Tapio S, Merl-Pham J, Huber SM, Edalat
L, et al: Radiation alters the cargo of exosomes released from
squamous head and neck cancer cells to promote migration of
recipient cells. Sci Rep. 7:124232017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zheng J, Dong W, Zhang J, Li G and Gong H:
YB-1, a new biomarker of glioma progression, is associated with the
prognosis of glioma patients. Acta Biochim Biophys Sin (Shanghai).
48:318–325. 2016. View Article : Google Scholar
|
|
27
|
Tong H, Zhao K, Zhang J, Zhu J and Xiao J:
YB-1 modulates the drug resistance of glioma cells by activation of
MDM2/p53 pathway. Drug Des Devel Ther. 13:317–326. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu J, Stratford AL, Astanehe A and Dunn
SE: YB-1 is a transcription/translation factor that orchestrates
the oncogenome by hardwiring signal transduction to gene
expression. transl Oncogenomics. 2:49–65. 2007.PubMed/NCBI
|
|
29
|
Wild-Bode C, Weller M, Rimner A, Dichgans
J and Wick W: Sublethal irradiation promotes migration and
invasiveness of glioma cells: Implications for radiotherapy of
human glioblastoma. Cancer Res. 61:2744–2750. 2001.PubMed/NCBI
|
|
30
|
Khan L, Soliman H, Sahgal A, Perry J, Xu W
and Tsao MN: External beam radiation dose escalation for high grade
glioma. Cochrane Database Syst Rev. 19:CD0114752016.
|
|
31
|
Woller N, Gürlevik E, Fleischmann-Mundt B,
Schumacher A, Knocke S, Kloos AM, Saborowski M, Geffers R, Manns
MP, Wirth TC, et al: Viral infection of tumors overcomes resistance
to PD-1-immunotherapy by broadening neoantigenome-directed T-cell
responses. Mol Ther. 23:1630–1640. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sahebjam S, Sharabi A, Lim M, Kesarwani P
and Chinnaiyan P: Immunotherapy and radiation in glioblastoma. J
Neurooncol. 134:531–539. 2017. View Article : Google Scholar : PubMed/NCBI
|