Introduction
Sepsis refers to 'life-threatening organ dysfunction
due to a dysregulated host response to infection' (1). The heart is one of the most
frequently affected organs during sepsis, and the damage is
primarily manifested in the structure and function of cardiac
endothelial cells (ECs) (2). ECs
are widely distributed throughout the body (3). ECs function as protective barriers
of blood vessels and are involved in apoptosis, shedding,
permeability changes and uncontrolled secretion of various
cytokines and inflammatory factors (3). Endothelial damage and dysfunction is
characterized by enhanced inflammation and reduced nitric oxide
(NO) bioavailability (4,5).
Dysfunction in the ECs determines the occurrence and
development of sepsis and is therefore the pathological basis for
sepsis (2). Sepsis and multiple
organ dysfunction (MODS) caused by the dysfunction of ECs is
usually accompanied by the following four events: i) Changes in
vascular endothelial permeability and loss of barrier function; ii)
changes in the blood levels of EC-associated biomarkers, including
intercellular adhesion molecule 1 (ICAM-1), vascular endothelial
growth factor (VEGF) and Von Willebrand Factor (vWF); iii)
over-activation of the inflammatory response; and iv) NO levels
influencing the blood supply in the myocardium and the contraction
and diastolic function of ECs (6).
Neuregulin-1 (NRG-1) is a member of the epidermal
growth factor family (7). NRG-1
receptors include epidermal growth factor receptor (ErbB)1, ErbB2,
ErbB3 and ErbB4, the last three types all being associated with EC
protection (7). NRG-1/ErbB
signaling pathways are involved in the regulation of EC migration,
proliferation, survival and adhesion (8). One previous study revealed that
NRG-1 overexpression exerts cardioprotective effects against
ischemic injury in cultured cardiomyocytes (9). NRG-1/ErbB signaling may be regulated
to attenuate anoxia-induced injuries in cardiac ECs. Furthermore,
NRG-1/ErbB signaling has been demonstrated to serve an important
function in paracrine and autocrine signaling in the heart, in
anti-apoptosis, and in the regulation of myocyte and EC functions
(10). In vivo and in
vitro experiments have demonstrated that NRG-1 may promote
myocardial cell proliferation, inhibit cardiomyocyte apoptosis and
protect cardiac ECs; however, the corresponding molecular
mechanisms remain yet to be elucidated (11).
NRG-1 exerts protection effect via multiple targets
(12). In previous years, it has
been reported that NRG-1 may improve myocardial dysfunction caused
by sepsis by alleviating inflammation and oxidative stress
(12). NRG-1 was demonstrated to
increase hemodynamics acutely and improve left ventricular (LV)
systolic function (13). However,
it has not been reported whether NRG-1 may enhance cardiac function
by protecting cardiac vascular ECs from injury in sepsis-induced
cardio-myopathy. Considering that the protective mechanism
underlying NRG-1 function shares certain common features with the
pathway of EC injury in sepsis, it was hypothesized that NRG-1
serves a function in strengthening cardiac function by protecting
ECs during sepsis.
Materials and methods
Animal and specimen collection
Male Sprague Dawley rats (weight, 300-400 g; age, 10
weeks; N=36; Hunan SJA Laboratory Animal Co., Ltd., Hunan, China)
were used following one week of acclimation. Briefly, rats were
randomly divided into the following three groups: Sham,
lipopolysaccharide (LPS) and NRG-1, and each group was divided into
two subgroups (24 and 48 h treatment times). Recombinant human
NRG-1 (rhNRG-1; cat. no. 10658-H08H; Sino Biological Inc., Wayne,
PA, USA) or saline solution was injected into the tail vein at 5
and 24 h subsequent to model establishment. For the LPS group, the
model was established by an intraperitoneal injection of 10 mg/kg
LPS (cat. no. L2880; Sigma Aldrich; Merck KGaA, Darmstadt,
Germany); for the sham group, the same volume of saline was
injected intraperitoneally at the same time. For the survival
assessment, all rats were observed at 6-h intervals for 48 h. At
the end of the hemodynamic measurement, blood samples were
collected through the inferior vena cava and transferred to sealed
tubes. Following 15-20 min, the blood samples were centrifuged at
1,810 × g for 15 min at 4°C and stored at −80°C until measurement.
Subsequent to collecting the blood samples, cardiac tissue samples
were selected for biochemical and histopathological analysis. The
left ventricular myocardium was transferred to a cryotube and
stored at −80°C until Ras homolog family member A
(RhoA)/Rho-associated protein kinase 1 (ROCK1) analysis, while the
right ventricular myocardium was removed and fixed at 37°C for at
least 48 h in 10% paraformaldehyde solution for histopathological
and immunofluorescence analyses. The animal experiments and
procedures were ethically approved by the Animal Care and Use
Committee of Renmin Hospital of Wuhan University (Wuhan, China).
All animals were euthanized with carbon dioxide (displacement rate
in the chamber, 20% volume/min) and then they spines were
dislocated to confirm death. All protocols were performed in
accordance with the US National Institutes of Health guide for the
care and use of Laboratory animals (National Institutes of Health
Publication revised in 2011) (14).
Assessment of cardiac function
At 24 or 48 h following surgery, the rats were
anesthetized again with 2% sodium pentobarbital in saline (40
mg/kg, intraperitoneally) and placed on a warming pad (37°C).
Following tracheal intubation, the rats were placed on
positive-pressure ventilation using a rodent ventilator with a
respiratory rate of 70 and a tidal volume six times the weight of
the animal. Subsequent to exposing the right carotid artery, a
catheter was inserted into the right common carotid artery to
quantify the arterial blood pressure using a Labchart system
(ADInstruments, Bella Vista, Australia). Once the heart rate and
mean arterial pressure (MAP) were measured, the catheter was
introduced into the left ventricle via the right carotid artery to
monitor the LV systolic pressure (LVSP), LV end-diastolic pressure
(LVEDP), LV +dp/dt max (maximum rate of the increase of left
ventricular pressure) and LV -dp/dt max (maximum rate of the
decrease of left ventricular pressure).
Vascular permeability assay
The structural damage to the cardiac vasculature and
vascular permeability were evaluated by measuring vWF expression
using immunofluorescence staining. The paraffin sections
(5-µm-thick) were deparaffinized using conventional xylene
and absolute ethanol at 37°C, placed in a repair box filled with
EDTA antigen repair buffer (pH 8.0) and permeabilized with
phosphate-buffered saline at 92-96°C for 15 min. In this process,
excessive evaporation of the buffer was prevented and no occurred.
After natural cooling, the slides were placed in PBS (pH 7.4) and
washed three times in a decolorizing shaking bed for 5 min each
time. The slides were blocked with BSA (cat. no. G5001; Servicebio,
Inc., Wuhan, China) for 30 min at 37°C. Sections were immunostained
by incubation with primary antibodies against vWF (1:200; cat. no.
GB11020; Servicebio, Inc.) at 4°C overnight, then incubated with
CY3-conjugated goat anti-rabbit secondary antibodies (1:300; cat.
no. GB21303; Servicebio, Inc.) at 37°C for 50 min and subsequently
incubated with DAPI (cat. no. G1012; Servicebio, Inc.) at 37°C for
10 min. The slides were photographed using a fluorescence
microscope (Nikon Corporation, Tokyo, Japan). The percentage of vWF
present on the endothelial lining was calculated for each image.
The images were analyzed using ImageJ software (version d 1.47;
National Institutes of Health, Bethesda, MD, USA).
Measurement of endothelial-associated and
oxidative stress biomarkers
ICAM-1 and VEGF enzyme-linked immunosorbent assay
(ELISA) kits (cat. nos. CSB-E04576r and CSB-E04757r, respectively;
Cusabio, Wuhan, China) were used to measure the serum levels of the
endothelial biomarkers in blood samples, including leukocyte
adhesion molecule ICAM-1 and VEGF. The NO contents of the rats in
each group were measured following the reductase method using a NO
kit (cat. no. A012; Jiancheng Bioengineering Institute, Nanjing,
China). The assays were all performed according to the
manufacturer's protocols. Each treatment and the corresponding
control were analyzed in the same run.
Observation of myocardial tissues and
cells ultrastructure
The structures of myocardial cells and tissues were
observed by transmission electron microscopy and hematoxylineosin
(H&E) staining, respectively.
The myocardial tissues were sliced into
1-mm3 pieces and fixed with 2.5% glutaraldehyde in
phosphate buffer for 2.5 h at 4°C and then re-fixed in 1% osmium
tetroxide in phosphate buffer for 2 h at 4°C. The tissues were
dehydrated in a graded series of ethanol solutions (50, 70, 80, 90.
95 and 100), and then immersed in a mixture of acetone and epoxy
resin twice at 37°C (2:1 for 3 h the first time, 1:2 for overnight
the second time). Finally, the tissues were embedded in epoxy
resin-filled capsules and heated at 70°C overnight. Ultrathin
sections (60-80 nm) were sliced and observed using a transmission
electron microscope (TECNAI G2 20 TWIN; FEI; Thermo Fisher
Scientific, Inc., Waltham, MA, USA).
Samples were fixed in 4% formalin for at least 48 h
at 37°C. Subsequent to fixation, each tissue sample was processed
routinely and embedded in paraffin wax and serially sectioned at 4
µm thickness. The sections were stained with H&E at 37°C
(hematoxylin staining for 5 min and eosin staining for 3 min), and
analyzed under an optical microscope (Nikon Corporation); 5-6
fields were photographed randomly under a magnification of
×100.
Apoptosis of myocardial cells
analysis
The paraffin sections were deparaffinized, incubated
with the rupture solution for 30 min at 37°C, and subsequently
reagent 1 (TdT) and reagent 2 (dUTP) were added at a ratio of 2:29
using the terminal deoxynucleotidyl-transferase-mediated dUTP nick
end labeling assay kit (cat. no. 11684817910; Roche Diagnostics,
Basel, Switzerland). DAPI was used to counterstain the cell nuclei
for 5 min at 37°C. Images of the sections were observed and
captured under an inverted fluorescent microscope under a
magnification of ×400. The TUNEL kit was used according to the
manufacturer's protocol. The apoptosis rate was calculated as the
number of apoptotic cells/total number of cells ×100%.
Measurement of inflammatory factors
Serum levels of tumor necrosis factor α (TNF-α) and
interleukin-6 (IL-6) were detected by ELISA using TNF-α ELISA kit
(cat. no. 88-7340; Thermo Fisher Scientific, Inc.) and IL-6 kit
[cat. no. EK3061; MultiSciences (LIANKE) biotech, Co., Ltd.,
Hangzhou, China]. The protocol was strictly performed according to
the manufacturer's protocol.
Cell culture and treatment
H9c2 cardiomyoblasts were purchased from iCell
Bioscience Inc. (San Francisco, CA, USA) and were cultured in
Dulbecco's modified Eagle's medium (cat. no. SH30021.01; Hyclone;
GE Healthcare Life Sciences, Logan, UT, USA) supplemented with 10%
fetal bovine serum (cat. no. 10099-141; Gibco; Thermo Fisher
Scientific, Inc.), penicillin (100 U/ml) and streptomycin (100
mg/ml; cat. no. SV30010.01; Hyclone; GE Healthcare Life Sciences).
The H9c2 cells were challenged with LPS (10 µg/ml) at 37°C
for 24 and 48 h. rhNRG-1 (1 µg/ml) was added to the cells at
5 h in the NRG-1 group treated with LPS for 24 h, or at 5 and 24 h
in the NRG-1 group treated with LPS for 48 h. The culture medium
and cells were collected for western blot analysis.
RhoA and ROCK1 western blot protein
analysis
A 10-fold volume of RIPA lysate (cat. no. G2002;
Servicebio Inc.) was added 100 mg heart tissues; H9c2 cells were
incubated with an appropriate volume RIPA lysate. Heart tissues
were homogenized and lysed at −20°C for 60 sec and H9c2 cells were
homogenized and lysed at −20°C for 15 min, the supernatants of
lysates were obtained by centrifugation at 12,000 × g for 0.5 h at
4°C. After protein concentration was determined with a BCA protein
assay kit (cat. no. G2026; Servicebio, Inc.), aliquots of protein
(40 µg per lane) were separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis on a 12% gel. Proteins
were transferred onto polyvinylidene fluoride membranes (EMD
Millipore, Billerica, MA, USA) for 1.5 h, and the membranes were
blocked with 5% nonfat milk in tris-buffered saline with 0.05%
Tween-20 for 1 h at 37°C. After being blocked with 5% non-fat dry
milk for 1 h at 37°C, the primary antibodies used included
polyclonal antibodies for RhoA (dilution 1:1,000; cat. no. 2117S)
and ROCK1 (dilution 1:1,000; cat. no. 4035S; both Cell Signaling
Technology, Inc., Danvers, MA, USA) at 4°C overnight. The membrane
was also incubated with GAPDH primary antibodies (dilution
1:10,000; cat. no. GB12002; Servicebio Inc.) at 37°C for 30 min.
Subsequent to incubation with horseradish peroxidase-conjugated
goat anti-rabbit (cat. no. GB23303) or goat anti-rat (cat. no.
GB23302) secondary antibodies (dilution 1:3,000). After washing
three times with TBST (cat. no. G0001; both Servicebio, Inc.) at
37°C for 30 min, the proteins were immersed in enhanced
chemiluminescence solution (cat. no. G2014; Servicebio, Inc.) for 1
min and imaged in a chemical scanner (V300; EPSON, Seiko Epson
Corp., Japan). GAPDH was used as a loading control. The bands were
analyzed using ImageJ software (version d 1.47).
Statistical analysis
Data were presented as the mean ± standard deviation
and analyzed using SPSS 24.0 software (IBM Corp., Armonk, NY, USA).
Multiple groups were analyzed using two-way analysis of variance
with Bonferroni's post-hoc test. Survival rates were expressed as
the percentage of live animals, and survival curves were generated
using the Kaplan-Meier method. Survival assessments were compared
using Gehan-Breslow-Wilcoxon tests. P<0.05 was considered to
indicate a statistically significant difference.
Results
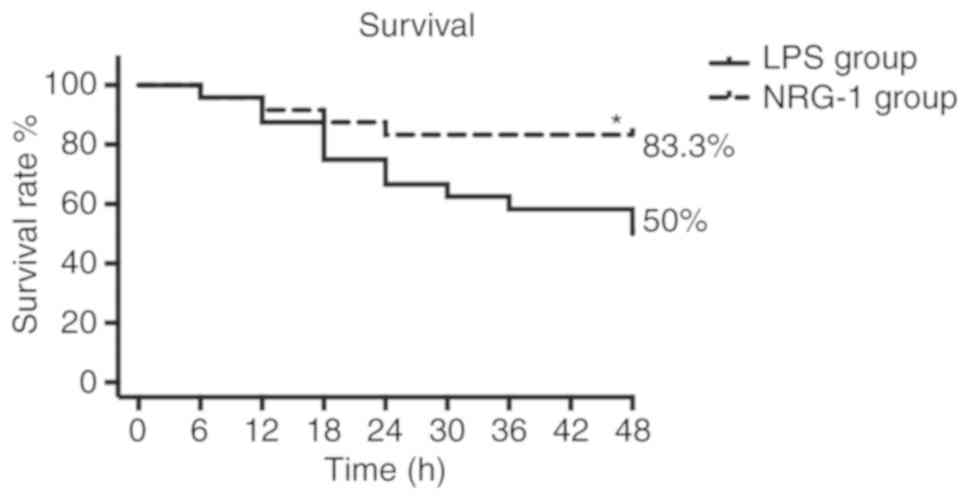
Experimental model and survival
rates
Rats presented with reduced activity, diarrhea,
piloerection and exudation around the eyes at 4 h after LPS
induction. Conversely, the sham group exhibited no detectable signs
of disease. The survival rates were evaluated at 48 h after LPS
injection. Rats in the sham group demonstrated a 100% survival rate
(n=12). Rats in the LPS group demonstrated survival rates of 87.5%
(21 out of 24) at 12 h and 66.7% (16 of 24) at 24 h, which
progressively dropped to 50.0% (14 of 24) at 48 h. The
administration of NRG-1 significantly increased the survival rate
in the NRG-1 group to 83.3% (20 out of 24) at 48 h compared with
the LPS group (P<0.05; Fig.
1). The above results indicated that NRG-1 effectively
abrogated the decrease in survival rate of rats with sepsis.
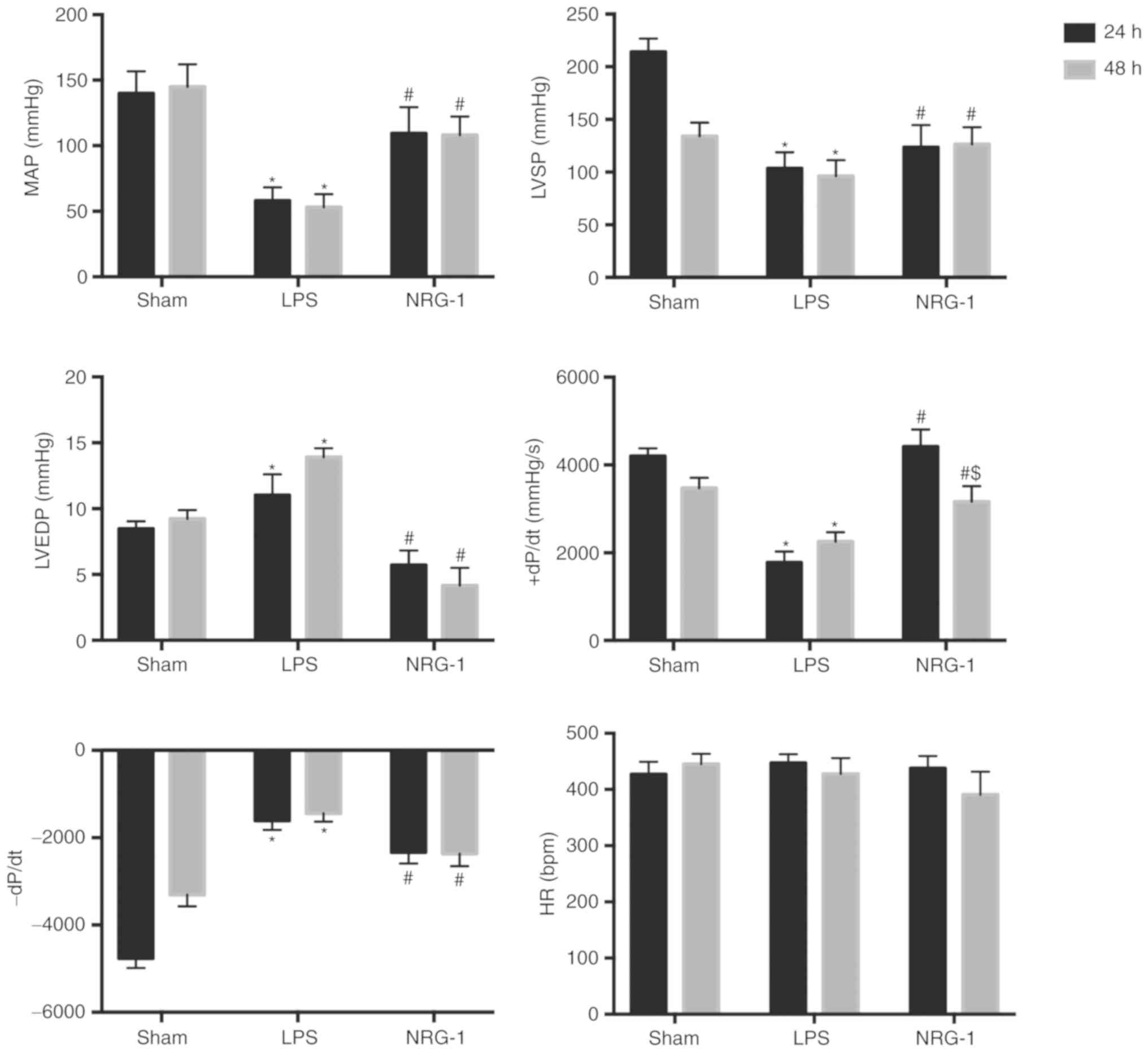
rhNRG-1 improves cardiac function
Cardiac function was evaluated by invasive
hemodynamic measurements at 24 and 48 h after sepsis induction.
LPS-induced sepsis was revealed to significantly alter the indices
of cardiovascular performance compared with the sham group
(P<0.05; Fig. 2). The
differences in heart rate among the three groups were not
statistically significant (P>0.05). Compared with rats in the
sham group, the rats in the LPS group exhibited significantly lower
values of MAP, LVSP and ±dp/dt max, in addition to increased LVEDP
(P<0.05). By contrast, subsequent to the administration of
rhNRG-1, the hemodynamic abnormalities mentioned above were
significantly attenuated (P<0.05). The rats in the 48 h group
exhibited slightly greater improvement in cardiovascular
performance compared with the 24 h group (Table I).
 | Table ICardiac function indices of rats in
each treatment group. |
Table I
Cardiac function indices of rats in
each treatment group.
| HR (bpm) | MAP (mmHg) | LVSP (mmHg) | LVEDP (mmHg) |
+dP/dtmax (mmHg/s) |
−dP/dtmax (mmHg/s) |
|---|
| Sham 24 h | 427.43±21.70 | 146.64±11.01 | 214.20±5.12 | 8.47±0.33 | 4,265.60±41.56 |
−4,902.61±13.61 |
| Sham 48 h | 445.36±18.12 | 144.94±6.98 | 133.90±5.23 | 9.23±0.38 | 3,603.92±33.50 |
−3,338.87±125.67 |
| LPS 24 h | 447.51±15.29 | 58.09±4.12a | 103.55±6.28a | 12.69±0.05a |
1,972.14±18.36a |
−1,776.90±85.46a |
| LPS 48 h | 427.76±28.26 | 53.03±4.10a | 95.87±6.22a | 13.91±0.40a |
2,250.74±126.44a |
−1,407.43±87.29a |
| NRG-1 24 h | 437.91±21.67 | 109.35±8.17b |
128.56±12.98b | 5.72±0.64b |
4,744.99±226.84b |
−2,401.84±92.66b |
| NRG-1 48 h | 390.86±40.95 | 111.40±7.63b | 126.05±6.69b | 4.16±0.78b |
3,163.68±203.58b,c |
−2,435.89±116.65b |
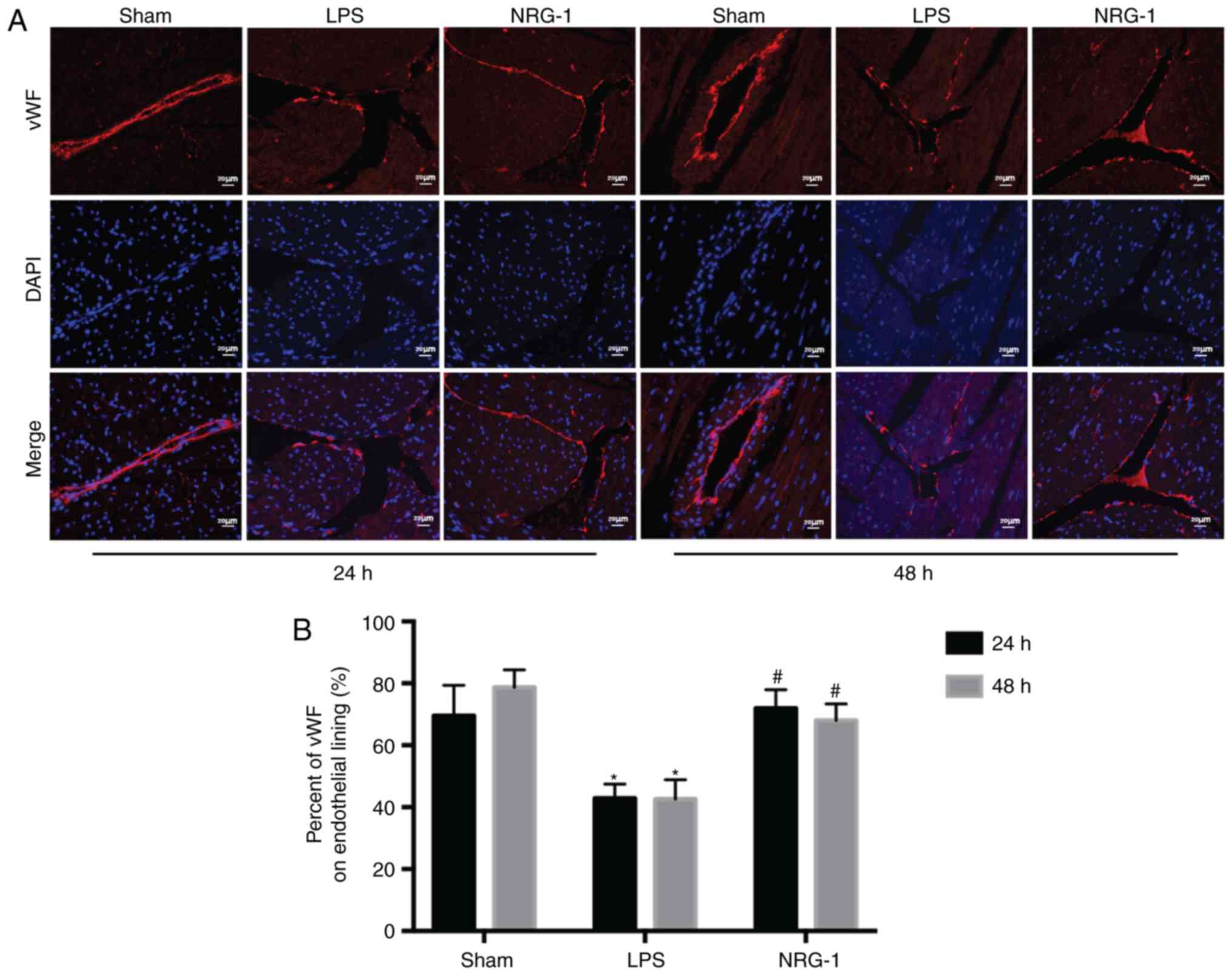
rhNRG-1 decreases cardiac vascular
permeability
The vascular endothelium was partially exfoliated
and vascular permeability significantly increased in the sepsis
group compared with the sham group (P<0.05); the observed
alterations were significantly attenuated in the NRG-1 group
(P<0.05). Staining patterns for vWF exhibiting respective
degrees of damage are illustrated in Fig. 3.
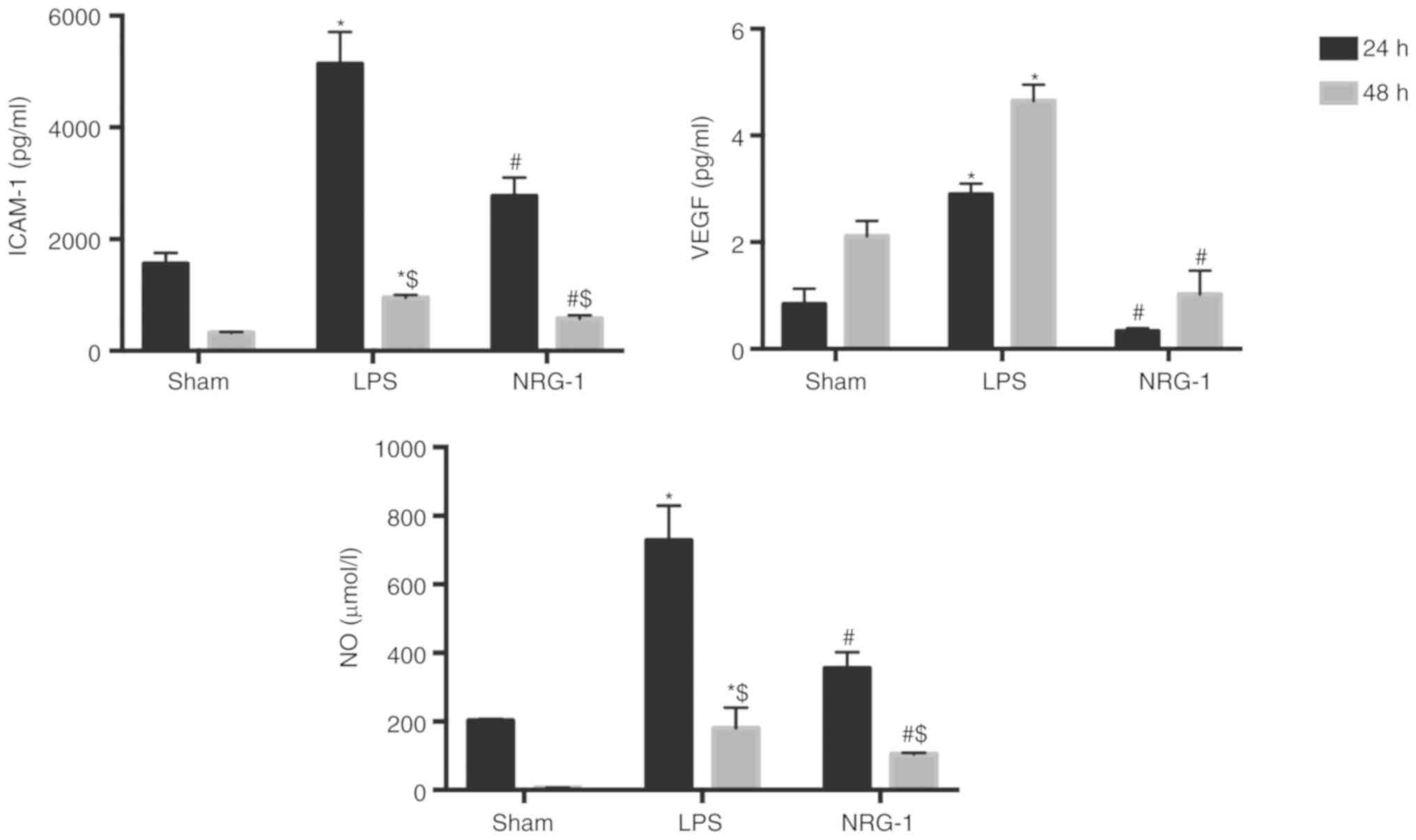
rhNRG-1 influences levels of endothelial
and oxidative stress biomarkers
The biomarkers used to evaluate the activation or
functional impairment of ECs included ICAM-1 and VEGF. The further
sequence of pathogenesis of endothelial dysfunction in sepsis is
the overproduction of NO (15).
The serum levels of ICAM-1, VEGF and NO in rats in the sepsis group
were significantly higher compared with the sham group (P<0.05),
whereas rhNRG-1 treatment was able to prevent these increases, and
ICAM-1 and NO levels were significantly decreased at 48 h of
treatment compared with 24 h (Fig.
4; Table II).
| Table IISerum levels of ICAM-1, VEGF and NO
in rats from each treatment group. |
Table II
Serum levels of ICAM-1, VEGF and NO
in rats from each treatment group.
| Variables | ICAM-1 (pg/ml) | VEGF (pg/ml) | NO (pg/ml) |
|---|
| Sham 24 h | 1,564.74±94.41 | 0.84±0.28 | 203.27±1.81 |
| Sham 48 h | 322.92±7.92 | 1.78±0.61 | 6.57±0.38 |
| LPS 24 h |
5,139.85±284.15a | 3.11±0.04a |
679.05±78.59a |
| LPS 48 h |
950.41±25.23a,b | 4.65±0.30a |
180.40±30.08a,b |
| NRG-1 24 h |
2,772.18±164.45c | 0.30±0.04c |
355.14±23.41c |
| NRG-1 48 h |
578.57±28.97b,c | 1.02±0.45c | 104.67±2.13b,c |
rhNRG-1 ameliorates cardiac injury in
septic rats
Based on the histological results. rats in the LPS
group exhibited disorderly arranged myocardial fibers, unclear cell
boundaries, interstitial edema and inflammatory cell infiltration
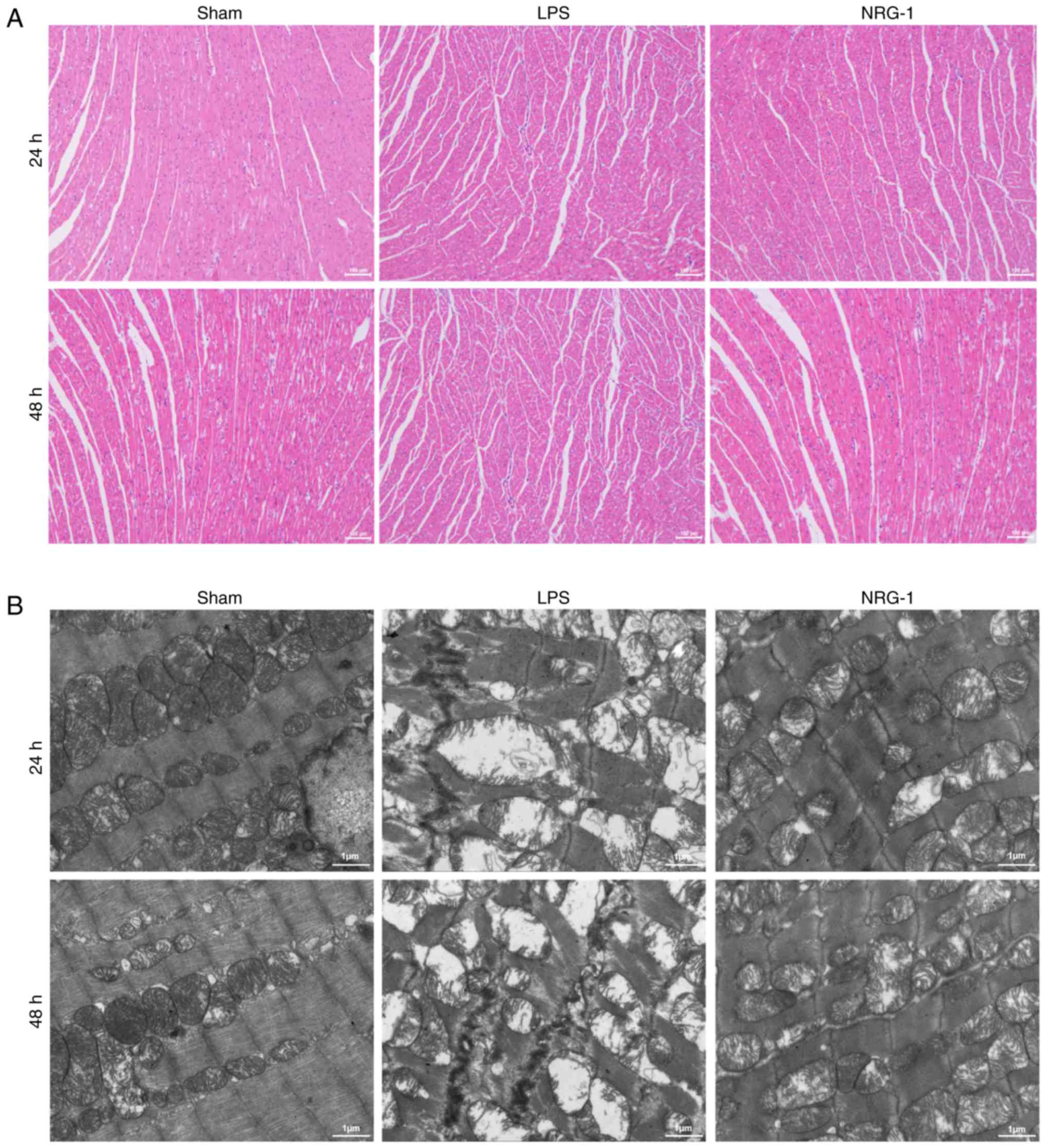
when compared with the rats in the sham group (Fig. 5A). Cytological results indicated
that at 24 h after LPS-induced sepsis, the distances between the
myofibrils of the myocardium were widened and even broken or
dissolved, mitochondrial swelling of the volume of mitochondria
increased and the electron density of the matrix decreased; and in
addition, matrix particles decreased or disappeared, the cristae
shortened, were decreased and their edges shifted and the damage
was aggravated with the progression of sepsis (Fig. 5B). rhNRG-1 treatment markedly
reduced the aforementioned changes.
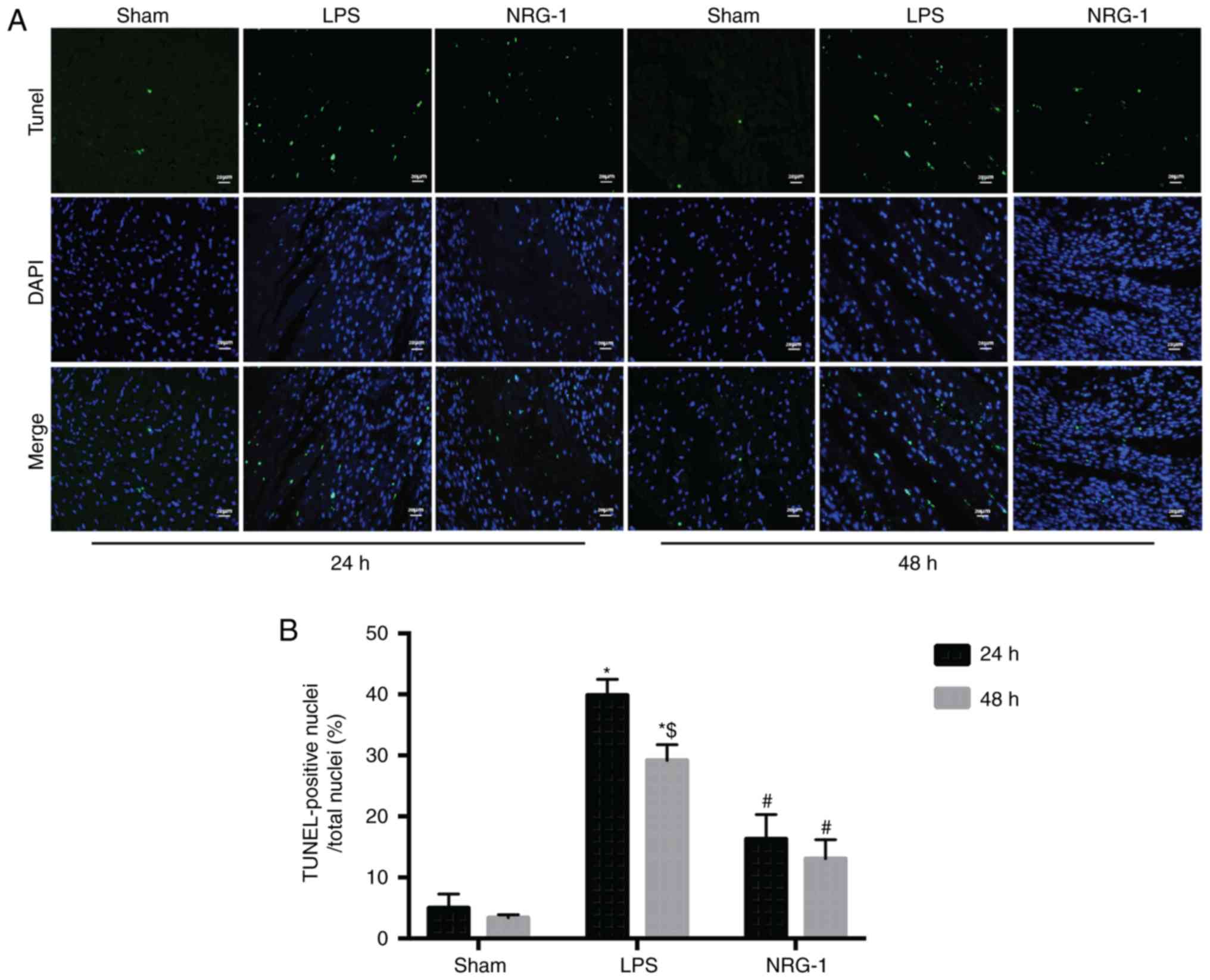
rhNRG-1 reduces cardiac apoptotic cells
in septic rats
LPS-induced sepsis significantly increased the
percentage of apoptotic cardiomyocytes compared with the sham group
(P<0.05). The observed increase was significantly attenuated in
the NRG-1 group (P<0.05); thus, overall, treatment with rhNRG-1
exhibited an anti-apoptotic effect (Fig. 6). The percentage of apoptotic
cells in the LPS 48 h group were significantly reduced compared
with those in the LPS 24 h group (P<0.05), However, rats in the
24 and 48 h groups exhibited no significant differences in the
therapeutic effects of rhNRG-1 (P>0.05).
rhNRG-1 suppresses the proinflammatory
response
rhNRG-1 is known to exert an anti-inflammatory
effect in the nervous system (7).
The preliminary experiment in the present study revealed the
inhibitory effect of rhNRG-1 on the serum levels of inflammatory
cytokines in the septic rat model at 24 h after sepsis induction.
The levels of TNF-α and IL-6 continued to increase within 48 h but
were partially restored to original levels by rhNRG-1 treatment at
48 h after sepsis induction (Table
III).
| Table IIILevels of TNF-α and IL-6 in the sera
of rats for each treatment group. |
Table III
Levels of TNF-α and IL-6 in the sera
of rats for each treatment group.
| TNF-α (pg/ml) | IL-6 (pg/ml) |
|---|
| Sham 24 h | 3.08±0.66 | 3.19±1.11 |
| Sham 48 h | 7.72±0.70 | 10.41±1.23 |
| LPS 24 h | 13.56±3.32 |
491.66±77.86a |
| LPS 48 h | 31.28±5.11a,b |
1,209.35±28.21a,b |
| NRG-1 24 h | 4.77±0.23 | 21.30±6.35c |
| NRG-1 48 h | 3.32±0.64c |
596.27±53.75b,c |
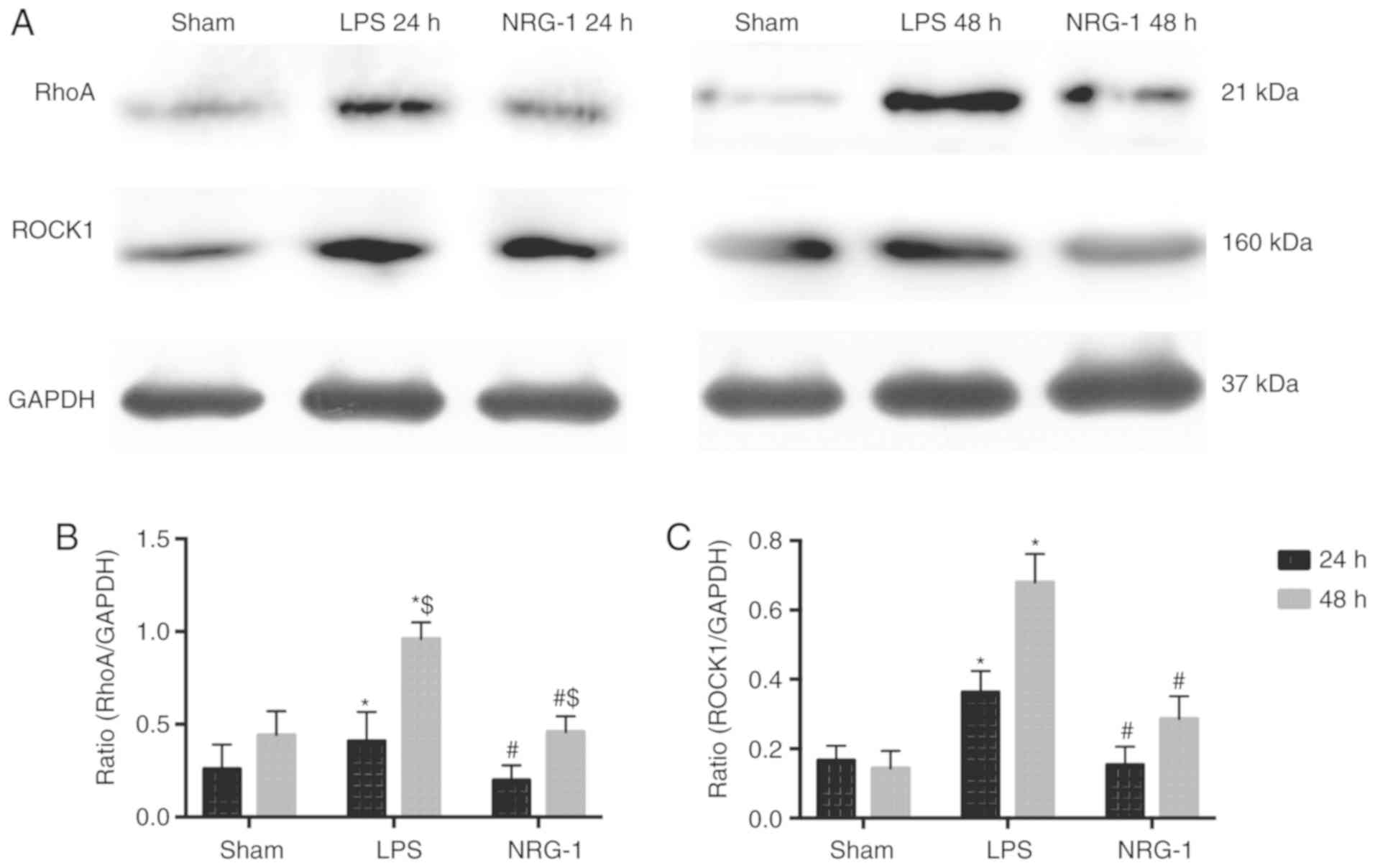
rhNRG-1 inhibits the activation of
RhoA/ROCK signaling
To investigate whether angiogenic factors are
associated with myocardial endothelial protection induced by NRG-1
treatment, the present study examined the RhoA/ROCK signaling
systems using western blot analysis in vivo (Fig. 7) and in vitro (Fig. 8). Protein expression levels of
RhoA and ROCK1 were revealed to be significantly upregulated in the
LPS group compared with those in the sham group (P<0.05).
However, rats in NRG-1 group exhibited a significant reduction in
RhoA and ROCK1 protein levels compared with rats in the LPS group
(P<0.05; Fig. 7).
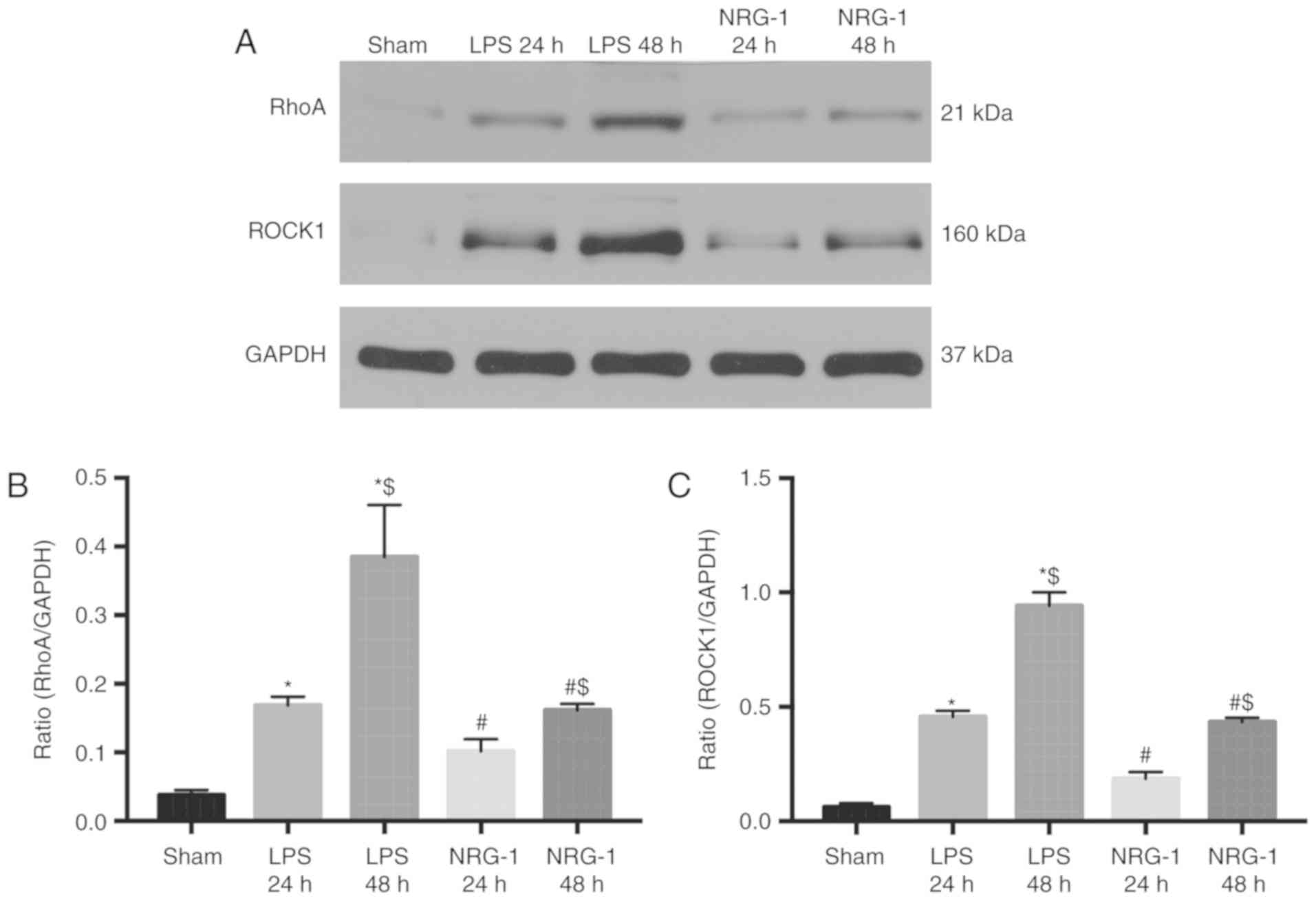
To further verify this hypothesis, the present study
used H9c2 cardiomyoblasts to demonstrate these mechanisms (Fig. 8). The significantly higher
activation of RhoA and ROCK1 signaling were observed in LPS-treated
cells compared with the sham group, while NRG-1 significantly
inhibited this activation compared with the LPS group (P<0.05).
This result is consistent with the result of the in vivo
experiment. In addition, RhoA and ROCK1 proteins were more
significantly activated in the 48 h group compared with the 24 h
group (P<0.05), and NRG-1 was able to significantly inhibit the
activation of RhoA and ROCK in the two groups (P<0.05).
Discussion
Over 70% of all sepsis-associated cases of mortality
are attributed to organ failure, in which the function of
myocardial dysfunction is widely recognized (16). Changes in endothelial function and
vascular tension associated with sepsis inevitably affect cardiac
function (17). Endothelial
dysfunction is a necessary component of the hemodynamic collapse
caused by sepsis. As a result, single drug therapy is often
insufficient to effectively reverse the organ dysfunction caused by
sepsis (17). Therefore,
therapies should be combined with anti-inflammatory and immune
regulation and vascular endothelial function regulation (18). Effectively reducing the production
of cell adhesion molecules and other vascular endothelial
biomarkers in blood may serve a strong protective function against
vascular endothelial function (19).
In the present study, LPS was selected to induce
sepsis in a rat model. LPS is able to directly damage vascular ECs.
On one hand, the effects of histamine, bradykinin and leukotriene
cause the contraction of vascular ECs; on the other hand, the
release of a series of myocardial depressant substances, including
TNF-α and IL-6, may cause the remodeling of the EC skeleton
(18). These two effects are able
to cause an increase in vascular permeability (18). It has been revealed that LPS is
able to induce the release various inflammatory mediators and
reactive oxygen species by leukocytes, and tissue hypoxia induced
by shock may promote the apoptosis of ECs (19). Apoptotic cells release ICAM-1 and
vascular cell adhesion protein 1 (VCAM-1), which may aggravate the
inflammatory reaction and impair the function of ECs (20). LPS results in the release of a
large number of inflammatory factors and EC adhesion molecules
(including ICAM-1 and VCAM-1) by ECs (18). In turn, this process causes
further cell damage and dysfunction, resulting in the loss of
vascular EC barrier function, myocardial ischemia, hypoxia and even
disseminated intravascular coagulation and ultimately, multiple
organ dysfunction, shock and mortality (18,19). In an LPS-induced injury model of
human umbilical vein ECs, IL-6 and TNF levels were substantially
upregulated subsequent to LPS stimulation, and EC injury was
attenuated following glucoside treatment. At the same time, the
levels of inflammatory factors expressed in ECs were reduced
(21). The results of the present
study revealed that the serum levels of the pro-inflammatory
cytokines TNF-α and IL-6 were partially restored following rhNRG-1
treatment. These preliminary results demonstrate the inhibitory
effect of rhNRG-1 on the immune inflammatory response.
The biomarkers that are used at present to evaluate
the activation or functional impairment of ECs include ICAM-1,
VCAM-1, VEGF and vWF (18-21).
Biomarker levels are positively associated with the degree of
injury of ECs (18-21). Previous studies have demonstrated
that 72 h after septic induction in rats, the serum levels of
ICAM-1 and vWF were significantly upregulated; however, simvastatin
treatment alleviated the injury of vascular ECs (22,23). The results of the present study
indicated that ICAM-1 and VEGF levels were significantly
upregulated in the LPS group compared with those in the sham group
(P<0.05). However, the observed increase was reversed by rhNRG-1
treatment. During sepsis, the levels of the endocrine vasodilator
NO increase, resulting in abnormal vasoconstriction and relaxation,
insufficient circulatory perfusion, tissue ischemia and hypoxia,
which further amplify septic shock (15). These processes are closely
associated with cardiac dysfunction (15). The present results revealed that
NO levels were significantly reduced following rhNRG-1 treatment
compared with the LPS group (P<0.05).
Increased permeability and loss of barrier function
are the core features that characterize EC dysfunction during
sepsis (3), thereby causing organ
dysfunction and further triggering MODS. Yano et al
(24) revealed that in the LPS
model, the vascular permeabilities of the lungs and kidneys are
substantially increased. The present study measured vWF levels to
assess the integrity of the endothelium, given that it is abundant
in the ECs and that the circulating level of vWF has been used as a
biomarker for endothelial damage/dysfunction in various disease
states, including systemic inflammatory response syndrome,
atherosclerosis, hypertension and diabetes (25). Kwon et al (26) demonstrated that patients exhibited
weaker vWF immunostaining along the endothelial lining compared
with those who recovered from acute respiratory failure. Similarly,
vWF levels around the endothelial lining were higher in the NRG-1
group. The present study demonstrated that vWF immunostaining along
the endothelial lining was significantly decreased in the LPS group
compared with that in the sham group; however, the observed
decrease was improved by rhNRG-1 treatment.
The further sequence of pathogenesis of septic
endothelial dysfunction is the elimination or inhibition of NO
(27). In clinical trials using
patients with sepsis, methylene blue (no scavenger) has been
demonstrated to attenuate hemo-dynamic changes (27). An inducible NO synthase inhibitor
(L-Kanavanin) may prevent the decrease in myocardial contractility
in septic rats (27). NO directly
affects the release of vascular tension and pro-inflammatory
cytokines (27). In addition, NO
may induce the production of apoptotic or anti-apoptotic signals
(27). Numerous cytokines serve
key functions in the inflammatory response and partly regulate
myocardial function through NO generation (3-5). A
study have been documented in patients with left ventricular
dysfunction and septic mice, a decreased adrenergic response is
associated with elevated NO levels (27). All the above-mentioned changes
affect the cardiac function of septic rats. The present results
revealed that NO levels and the number of apoptotic cells were
significantly higher in the LPS group compared with those in the
sham group (P<0.05). The present results suggested that rhNRG-1
attenuates excessive oxidative stress and apoptosis.
The mortality caused by sepsis is most commonly
attributed to cardio-circulatory depression (22). The present study selected rats at
24 and 48 h following LPS-induction to observe, as they reflected
changes in the process of cardiac dysfunction in sepsis. Previous
studies suggest that NRG-1 improves cardiac function and protects
rat cardiomyocytes from cecal ligation and puncture (CLP)-induced
sepsis by suppressing the immune inflammatory response and
inhibiting the excessive activation of the
renin-angiotensin-aldosterone system. NRG-1 was revealed to
increase the survival rates of rats in an CLP-induced sepsis model
at 24 h (21). In the present
study, rats subjected to LPS exhibited cardio-circulatory
depression (impaired MAP, dp/dt and LVP generation; Fig. 2) and histological and cytological
(Fig. 5) changes due to
myocardial cell damage. The results revealed that rhNRG-1 may
alleviate EC dysfunction, in addition to improving the cardiac
function and survival of rats at 48 h after sepsis induction.
NRG-1 is synthesized and released by the endocardial
and cardiac microvascular endothelium (12). A previous study revealed that
NRG-1 was able to improve left ventricular function and increase
capillary density in diabetic cardiomyopathy (DCM) (28). NRG-1 had been demonstrated to
alleviate diastolic dysfunction in animal models of DCM and
angiotensin II-induced injury (28). In vitro experiments
revealed that NRG-1 treatment may upregulate VEGF expression in
human coronary artery smooth muscle cells. The NRG-1/ErbB system is
required for the physiological adaptation of the heart to the
changes of cardiac demand (29,30). One previous study reported that
NRG-1 may improve cardiac function in animal models of heart
disease, including ischemic, dilated and viral cardiomyopathy
(29). NRG-1 exerts beneficial
effects on cardiac function and is known to regulate the metabolic
activity of cardiomyocytes and maintain cardiac homeostasis. For
instance, NRG-1 administration was demonstrated to restore
mitochondrial function and reduce oxidative stress (30). A previous study has confirmed that
rhNRG-1 is able to inhibit the excessive activation of
pro-inflammatory factors in the early stage of sepsis-induced
cardiomyopathy and help prevent myocardial injury in the late stage
via cardio-protective mechanisms (12). However, further studies are
required. ClinicalTrials.gov has also listed a number of cardiac
therapies involving NRG-1 administration (31). Thus, it was hypothesized that
NRG-1 administration may be effective in treating sepsis-induced
cardiomyopathy via its multiple cardio-protective effects.
Activation of the RhoA/ROCK signaling pathway
reduces NO availability, upregulates the expression of the
leukocyte adhesion factor ICAM-1 and participates in various
inflammatory responses (30,32-34). In addition, RhoA/ROCK signaling
causes EC shedding, increases serum levels of vWF and increases the
permeability of the vascular endothelium (35). RhoA activation has been
demonstrated to be involved in oxidative stress (30). ROCK regulates the development of
cardiovascular diseases by inducing the expression of NO synthase
in ECs (32). Insufficient NO
production in vascular ECs results in impaired
endothelium-dependent vasodilation (33). Furthermore, inhibition of the
RhoA/Rho-kinase pathway activates NO production and attenuates
vascular inflammation (34).
NRG-1-induced tumor cell motility is known to negatively regulate
ROCK signaling (36). A previous
study demonstrated that the activation of RhoA/Rho kinase signaling
promotes VEGF-induced in vitro EC migration, angiogenesis
and cytoskeletal changes; by contrast, the inhibition of Rho kinase
prevented mechanical wounding (37). The RhoA/ROCK pathway induces
thrombin-induced ICAM-1 expression via the activation of IκB kinase
(38) and exacerbates LPS-induced
endothelial hyper-permeability by mediating the formation of stress
fibers in ECs. Therefore, inhibition of RhoA/ROCK activation may
improve vascular hyper-permeability in sepsis (39). Next, the present study
investigated the protein expression levels of RhoA and ROCK1 at 24
and 48 h after sepsis (Fig. 7),
and the results revealed that rhNRG-1 is able to suppress the
expression of RhoA and ROCK1 in vivo and in vitro,
indicating that NRG-1 functioned in the regulation of the
anti-inflammatory and anti-oxidative stress action, which is based
on the RhoA/ROCK pathway.
The results of the present study revealed that NRG-1
may alleviate the injury of ECs, improve cardiac function and
protect the integrity of ECs by inhibiting the secretion of EC
adhesion molecules, vascular endothelial growth factor and other
endothelial-associated biomarkers and inflammatory factors. NRG-1
inhibits oxidative stress and apoptosis and strengthens the barrier
function of vascular tissue, thereby improving cardiac function and
increasing survival rates via multiple targets. NRG-1 exerts
protective effects on the vascular endothelium by reducing VEGF
levels, subsequently preventing VEGF-mediated activation of the
RhoA/ROCK signaling pathway. These results suggest that NRG-1 may
improve its secretory function and the permeability of cardiac ECs,
which may be mediated by the RhoA/ROCK signaling pathway. The above
results may serve as the basis for the development of a novel
approach to reverse the impaired vascular ECs in sepsis in future
studies.
At present, the primary treatments for
sepsis-induced cardiac dysfunction primarily focus on hemodynamics,
infections and energy supply (40). The present study revealed that
NRG-1 improved cardiac dysfunction via multiple targets,
alleviating the excessive activation of inflammatory responses and
alleviating the injury of ECs. These observations may serve as a
basis for the development of a novel approach to reverse the
impairment of vascular ECs in sepsis in future studies.
Acknowledgments
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81772044 and
81500255), the Program for Innovative Research Team in Hainan
Province (grant no. 2016CXTD012), the Science Foundation of Wuhan
Health and Family Planning Commission (grant no. WX17B09), the
Natural Science Foundation of Hubei Province (grant no. 2017CFC824)
and Wuhan University Young Teachers Project (grant nos.
2042018kf0199 and 2042017kf0138).
Availability of data and materials
All data generated or analyzed during the current
study are included in this published article.
Authors' contributions
WK, LW and XiW conceived and designed the study. WK,
YC, LZ and HTL conducted the experiments and wrote the manuscript.
WK, SD, FZ and XinW analyzed the data. WK, LW, XiW, LZ and HTL
revised the manuscript. All the authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The animal experiments and procedures were ethically
approved by the Animal Care and Use Committee of Renmin Hospital of
Wuhan University (Wuhan, China). All protocols were performed in
accordance with the US National Institutes of Health guide for the
care and use of Laboratory animals (National Institutes of Health
Publication revised in 2011) (40). All animal studies also complied
with the Animal Research: Reporting of In Vivo Experiments
guidelines and the American Veterinary Medical Association's
euthanasia guidelines of 2013 (41,42).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Seymour CW, Liu VX, Iwashyna TJ,
Brunkhorst FM, Rea TD, Scherag A, Rubenfeld G, Kahn JM,
Shankar-Hari M, Singer M, et al: Assessment of clinical criteria
for sepsis. For the third international consensus definitions for
sepsis and septic shock (Sepsis-3). JAMA. 315:762–774. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Opal SM and van der Poll T: Endothelial
barrier dysfunction in septic shock. J Internal Med. 277:277–293.
2015. View Article : Google Scholar
|
|
3
|
Boonla O, Kukongviriyapan U, Pakdeechote
P, Kukongviriyapan V, Pannangpetch P, Prachaney P and Greenwald SE:
Curcumin improves endothelial dysfunction and vascular remodeling
in 2K-1C hypertensive rats by raising nitric oxide availability and
reducing oxidative stress. Nitric Oxide. 42:44–53. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Park MH, Heo SJ, Park PJ, Moon SH, Sung
SH, Jeon BT and Lee SH: 6,6′-Bieckol isolated from ecklonia cava
protects oxidative stress through inhibiting expression of ROS and
proinflammatory enzymes in high-glucose-induced human umbilical
vein endothelial cells. Appl Biochem Biotechnol. 174:632–643. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Reverri EJ, Morrissey BM, Cross CE and
Steinberg FM: Inflammation, oxidative stress, and cardiovascular
disease risk factors in adults with cystic fibrosis. Free Radic
Biol Med. 76:261–277. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Leligdowicz A, Richard-Greenblatt M,
Wright J, Crowley VM and Kain KC: Endothelial activation: The
Ang/Tie axis in sepsis. Front Immunol. 9:8382018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu M, Solomon W, Cespedes JC, Wilson NO,
Ford B and Stiles JK: Neuregulin-1 attenuates experimental cerebral
malaria (ECM) pathogenesis by regulating ErbB4/AKT/STAT3 signaling.
J Neuroinflamm. 15:1042018. View Article : Google Scholar
|
|
8
|
Howell MD and Davis AM: Management of
sepsis and septic shock. JAMA. 317:847–848. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xiao J, Li B, Zheng Z, Wang M, Peng J, Li
Y and Li Z: Therapeutic effects of neuregulin-1 gene transduction
in rats with myocardial infarction. Coron Artery Dis. 23:460–468.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Parodi EM and Kuhn B: Signaling between
microvascular endothelium and cardiomyocytes through neuregulin.
Cardiovasc Res. 102:194–204. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Orlandi A and Bennett M: Progenitor
cell-derived smooth muscle cells in vascular disease. Biochem
pharmacol. 79:1706–1713. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou Q, Pan X, Wang L, Wang X and Xiong D:
The protective role of neuregulin-1: A potential therapy for
sepsis-induced cardiomyopathy. Eur J Pharmacol. 788:234–240. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lenihan DJ, Anderson SA, Lenneman CG,
Brittain E, Muldowney JAS III, Mendes L, Zhao PZ, Iaci J, Frohwein
S, Zolty R, et al: A phase I, single ascending dose study of
cimaglermin alfa (neuregulin 1β3) in patients with systolic
dysfunction and heart failure. JACC Basic Transl Sci. 1:576–586.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the Care and Use of Laboratory Animals. 8th ed.
National Academies Press (US); Washington, DC: 2011
|
|
15
|
Liao MH, Shih CC, Tsao CM, Chen SJ and Wu
CC: RhoA/Rho-kinase and nitric oxide in vascular reactivity in rats
with endotoxaemia. PLoS One. 8:e563312013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hotchkiss RS and Karl IE: The
pathophysiology and treatment of sepsis. N Engl J Med. 348:138–150.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Candel FJ, Borges Sá M, Belda S, Bou G,
Del Pozo JL, Estrada O, Ferrer R, González Del Castillo J,
Julián-Jiménez A, Martín-Loeches I, et al: Current aspects in
sepsis approach. Turning things around. Rev Esp Quimioter.
31:298–315. 2018.PubMed/NCBI
|
|
18
|
Lee J, Lee S, Zhang H, Hill MA, Zhang C
and Park Y: Interaction of IL-6 and TNF-α contributes to
endothelial dysfunction in type 2 diabetic mouse hearts. PLoS One.
12:e01871892017. View Article : Google Scholar
|
|
19
|
Qi D, Wang D, Zhang C, Tang X, He J, Zhao
Y, Deng W and Deng X: Vaspin protects against LPS-induced ARDS by
inhibiting inflammation, apoptosisand reactive oxygen species
generation in pulmonary endothelial cells via the Akt/GSK-3β
pathway. Int J Mol Med. 40:1803–1817. 2017.PubMed/NCBI
|
|
20
|
Stefanec T: Endothelial apoptosis: Could
it have a role in the pathogenesis and treatment of disease. Chest.
117:841–854. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ma MM, Li Y, Liu XY, Zhu WW, Ren X, Kong
GQ, Huang X, Wang LP, Luo LQ and Wang XZ: Cyanidin-3-O-Glucoside
ameliorates lipopolysaccharide-induced injury both in vivo and in
vitro suppression of NF-kB and MAPK pathways. Inflammation.
38:1669–1682. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sakuragi T, Lin X, Metz CN, Ojamaa K, Kohn
N, Al-Abed Y and Miller EJ: Lung-derived macrophage migration
inhibitory factor in sepsis induces cardio-circulatory depression.
Surg Infect (Larchmt). 8:29–40. 2007. View Article : Google Scholar
|
|
23
|
Jiang S, Li S, Hu J, Xu X, Wang X, Kang X,
Qi J, Lu X, Wu J, Du Y and Xiao Y: Combined delivery of
angiopoietin-1 gene and simvastatin mediated by anti-intercellular
adhesion molecule-1 antibody-conjugated ternary nanoparticles for
acute lung injury therapy. Nanomedicine. 15:25–36. 2019. View Article : Google Scholar
|
|
24
|
Yano K, Liaw PC, Millington JM, Shih SC,
Okada H, Bodyak N, Kang PM, Toltl L, Belikoff B, Buras J, et al:
Vascular endothelial growth factor is an important determinant of
sepsis morbidity and mortality. J Exp Med. 203:1447–1458. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jacobson SH, Egberg N, Hylander B and
Lundahl J: Correlation between soluble markers of endothelial
dysfunction in patients with renal failure. Am J Nephrol. 22:42–47.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kwon O, Hong SM, Sutton TA and Temm CJ:
Preservation of peritubular capillary endothelial integrity and
increasing pericytes may be critical to recovery from postischemic
acute kidney injury. Am J Physiol Renal Physiol. 295:F351–F359.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Barth E, Radermacher P, Thiemermann C,
Weber S, Georgieff M and Albuszies G: Role of inducible nitric
oxide synthase in the reduced responsiveness of the myocardium to
catecholamines in a hyper dynamic, murine model of septic shock.
Crit Care Med. 34:307–313. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hervent AS, Vandekerckhove L and Keulenaer
GWD: Neuregulin-1 antagonizes myocardial fibrosis and diastolic
dysfunction in angiotensin-II treated mice. Eur Heart J. 34:24–34.
2013. View Article : Google Scholar
|
|
29
|
Turner A, Tsamitros M and Bellomo R:
Myocardial cell injury in septic shock. Crit Care Med.
27:1775–1780. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guo LW, Gao L, Rothschild J, Su B and
Gelman IH: Control of protein kinase C activity, phorbol
ester-induced cytoskeletal remodeling, and cell survival signals by
the scaffolding protein SSeCKS/GRAVIN/AKAP12. J Biol Chem.
286:38356–38366. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Galindo CL, Ryzhov S and Sawyer DB:
Neuregulin as a heart failure therapy and mediator of reverse
remodeling. Curr Heart Fail Rep. 11:40–49. 2015. View Article : Google Scholar
|
|
32
|
Siafakas NM, Antoniou KM and Tzortzaki EG:
Role of angiogenesis and vascular remodeling in chronic obstructive
pulmonary disease. Int J Chron Obstruct Pulmon Dis. 2:453–462.
2007.
|
|
33
|
Mount PF, Kemp BE and Power DA: Regulation
of endothelial and myocardial NO synthesis by multi-site eNOS
phosphorylation. J Mol Cell Cardiol. 42:271–279. 2007. View Article : Google Scholar
|
|
34
|
Sawada N and Liao JK: Targeting eNOS and
beyond: Emerging heterogeneity of the role of endothelial Rho
proteins in stroke protection. Expert Rev Neurother. 9:1171–1186.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li D, Cheng P, Jiang H, Cao T, Wang J, Gao
Y, Lin Y, Wang C, Zhang S, Li J, et al: Vascularization converts
the lineage fate of bone mesenchymal stem cells to endothelial
cells in tissue-engineered bone grafts by modulating FGF2-RhoA/ROCK
signaling. Cell Death Dis. 9:9592018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rana MK and Worthylake RA: Novel mechanism
for negatively regulating Rho-kinase (ROCK) signaling through
Coronin1b protein in neuregulin 1 (NRG-1)-induced tumor cell
motility. J Biol Chem. 287:21836–21845. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
van Nieuw Amerongen GP, Koolwijk P,
Versteilen A and van Hinsbergh VW: Involvement of RhoA/Rho kinase
signaling in VEGF-induced endothelial cell migration and
angiogenesis in vitro. Arterioscler Thromb Vasc Biol. 23:211–217.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Anwar KN, Fazal F, Malik AB and Rahman A:
RhoA/Rho- associated kinase pathway selectively regulates
thrombin-induced intercellular adhesion molecule-1 expression in
endothelial cells via activation of I kappa B kinase beta and
phosphorylation of RelA/p65. J Immunol. 173:6965–6972. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhou Z, Guo F, Dou Y, Tang J and Huan J:
Guanine nucleotide exchange factor-H1 signaling is involved in
lipopolysaccharide- induced endothelial barrier dysfunction.
Surgery. 154:621–631. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen J, Lai J, Yang L, Ruan G, Chaugai S,
Ning Q, Chen C and Wang DW: Trimetazidine prevents
macrophage-mediated septic myocardial dysfunction via activation of
the histone deacetylase sirtuin 1. Br J Pharmacol. 173:545–561.
2016. View Article : Google Scholar
|