Introduction
Atherosclerosis (AS) is a pathological condition
characterized by artery narrowing, and is associated with high
morbidity and mortality rates worldwide. Although the detailed
pathogenic mechanisms remain to be elucidated, endothelial
dysfunction (ED) is the initiating event of AS (1,2),
which can alter the homeostasis of the cardiovascular system, with
accompanying changes in cell morphology and function. The canonical
Wnt/β-catenin pathway plays a crucial role in cell proliferation,
adhesion and other physiological processes (3), and has been implicated in various
conditions, such as inflammatory disease, fibrotic disease and
multiple cancer types (4,5). Previous studies have suggested that
this pathway is also related to the development of AS (6,7).
Indeed, Wnt/β-catenin participates in the inflammatory response,
which may cause ED (8). Moreover,
previous findings have demonstrated that sphingomyelin synthase 2
overexpression may activate the Wnt/β-catenin pathway and
potentially lead to ED (9,10).
AS has also been associated with endoplasmic
reticulum (ER) stress, which is related to numerous cellular
biological functions (11). ER
stress is induced by unfolded proteins when homeostasis of the ER
is disrupted by adverse conditions, such as hyperlipidaemia and
oxidative stress (12,13). Animal experiments have shown that
the expression levels of the ER stress-associated proteins 78 kDa
glucose-regulated protein (GRP78), phosphorylated
(phospho)-PRKR-like endoplasmic reticulum kinase,
phosphoserine/threonine-protein kinase/endoribonuclease IRE1,
cyclic AMP-dependent transcription factor ATF-6α (ATF6) and
C/EBP-homologous protein (CHOP) were increased in apolipoprotein
E-knockout mice. In addition, many atherogenic risk factors can
activate ER stress in the initial stages of AS, further aggravating
ED and AS (14,15). Moreover, Hong et al
(16) proposed ER stress as an
important contributing factor to ED during AS development and
progression. Thus, ER stress might contribute to AS by promoting
ED.
Simvastatin is a 3-hydroxy-3-methylglutaryl-coenzyme
A reductase inhibitor, and its ability to reduce cholesterol and
blood lipids is clinically exploited to lower the risk of
cardiovascular events (17).
However, its effects on endothelial cells under oxidative stress
conditions and the possible underlying mechanisms are still
unclear. In addition, statins can decrease the rate of neural
apoptosis and promote neural recovery by activating the
Wnt/β-catenin pathway (18).
Notably, in functional recovery from traumatic brain injury,
simvastatin reinforces neurogenesis via the regulation of
isoprenoid synthesis and the activation of Wnt signalling (19). Most of these previous results
support the beneficial effects of simvastatin. However, the present
study revealed that high doses of simvastatin (≥1 µM) may
even induce ED. Simvastatin regulates the Wnt/β-catenin pathway in
nerve cells; however, its effect on the Wnt/β-catenin pathway in
relation to ER stress contributing to ED is not clear. To examine
the potential impact of simvastatin on ED, in the present study,
human umbilical vascular endothelial cells (HUVECs) were subjected
to H2O2-induced oxidative stress in the
presence of simvastatin and/or the Wnt/β-catenin inhibitor
salinomycin, and the level of ED, as well as the expression of
proteins related to the Wnt/β-catenin pathway and ER stress, were
evaluated. These findings may contribute to gaining an improved
understanding of the molecular and cellular mechanisms regulating
AS, and offer an insight into the risks and mechanisms of action of
the clinical use of statins, while highlighting new targets for
treatment and prevention.
Materials and methods
Cell culture
HUVECs (Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences) were cultivated in Dulbecco's modified
Eagle's medium containing penicillin and streptomycin (100 U/ml and
0.1 mg/ml, respectively; Beijing Solarbio Science & Technology
Co., Ltd.) plus 10% certified foetal bovine serum (FBS; Biological
Industries). In addition, THP-1 cells (Cell Bank of Type Culture
Collection of the Chinese Academy of Sciences) were used in a
co-culture experiment to assess the effect of the treatments on the
adhesion ability of HUVECs, which may reflect the degree of ED.
THP-1 cells were cultured in RPMI-1640 (Beijing Solarbio Science
& Technology Co., Ltd.) plus 10% FBS. All cells were cultured
at 37°C in a 5% CO2 atmosphere.
Treatment of HUVECs with simvastatin and
salinomycin
The cells were cultured as described above, and then
treated with 1 µM simvastatin (Dalian Meilun Biology
Technology Co., Ltd.) and/or 10 µM salinomycin
(MedChemExpress) for 24 h. Next, the cells were treated with
H2O2 (500 µM) for another 24 h to
induce oxidative stress. Therefore, the following four treatment
groups were created: Control (H2O2); Sim
(Simvastatin+H2O2); Sal
(Salinomycin+H2O2); and Sim+Sal
(Simvastatin+Salinomycin+H2O2). Supernatants
and cells were collected for further analysis. Each treatment group
consisted of six replicate wells, and each experiment was performed
in triplicate. All cells were cultured at 37°C in a 5%
CO2 atmosphere.
Cell viability assay
HUVECs were seeded in 96-well plates at 80%
confluence and were exposed to different concentrations of
simvastatin (0, 1, 2 and 4 µM) for 24 h. The cells were also
treated with simvastatin (1 µM) and/or salinomycin (10
µM) for 24 h, and then incubated with
H2O2 (500 µM) for another 24 h. To
assess cell viability, 20 µl MTT (5 mg/ml; cat. no. M8180;
Beijing Solarbio Bioscience & Technology Co., Ltd.) was added
to each well and the cells were further incubated for 4 h. The
medium was removed, 150 µl dimethyl sulfoxide was added to
each well, and the plates were agitated for 15 min. The absorbance
was measured at 490 nm. All cells were cultured at 37°C in a 5%
CO2 atmosphere.
Acridine orange (AO) and ethidium bromide
(EB) staining
HUVECs were added into 24-well plates, grown to 80%
confluence, and the aforementioned treatments were applied. The
cells were stained with 5 µl AO and 5 µl EB (Solarbio
Life Science) for 5 min at room temperature in the dark. The dual
stain was then removed, and the cells washed with PBS three times.
Fluorescence was observed under a fluorescence microscope
(magnification, ×20; Olympus IX71; Olympus Corporation).
Measurement of the lactate dehydrogenase
(LDH) level in the medium, intracellular superoxide dismutase
(SOD), nitric oxide synthase (NOS) activity and malondialdehyde
(MDA) content
HUVECs in 6-well plates at a confluence of 80% were
treated in the aforementioned manner, and the medium was collected
for the measurement of LDH release levels (cat. no. A020-1; Nanjing
Jiancheng Bioengineering Institute) based on the absorbance
measured at 450 nm, indicating the extent of cell injury. The cells
were harvested and centrifuged (1,520 × g; 5 min; room
temperature). Homogenates were obtained after cell breakage by
sonication, set at an overall duration of 5 min and performed at a
frequency of 20 kHZ every 30 sec for 5 sec at 4°C. SOD (cat. no.
A001-1; Nanjing Jiancheng Bioengineering Institute) and nitric
oxide synthase NOS (cat. no. A014-1; Nanjing Jiancheng
Bioengineering Institute) activities, as well as the MDA (cat. no.
A003-1; Nanjing Jiancheng Bioengineering Institute) intracellular
content were determined using the respective kits, according to the
manufacturer's protocol, based on the absorbance measured at 560
nm.
Cell adhesion assay
After treatment with simvastatin and/or salinomycin,
followed by exposure to H2O2, THP-1 monocytic
cells were added to the medium of HUVECs at a density of
5×103/well, and culturing was continued for 30 min.
Next, the cells were washed twice with PBS so that only the THP-1
cells adhering to the HUVECs remained in the wells. Finally, THP-1
cells on HUVECs were counted under a phase-contrast inverted
microscope (magnification, ×20; Olympus IX71; Olympus Corporation)
to assess the cell adhesion ability. The average number of THP-1
cells was obtained from at least three replicate experiments.
Western blot analysis
Total proteins were extracted from the cells using
radioimmunoprecipitation assay buffer (Beijing Solarbio Science
& Technology Co., Ltd.), quantified using a bicinchoninic acid
kit, and ~60 µg protein/lane was separated via SDS-PAGE on
an 8-12% gel. Polyvinylidene fluoride membranes (Immobilon-P; EMD
Millipore) were used to transfer the separated proteins. Blocking
was performed with 5% skimmed milk or 10% bovine serum albumin
(Beijing Solarbio Science & Technology Co., Ltd.) dissolved in
TBS with 0.05% Tween-20 (TBST) for 1 h at room temperature. Next,
the following primary antibodies were added to TBST, and incubated
for at least 8 h at 4°C: Antibodies against the
apoptosis-associated proteins Bcl-2 (cat. no. 60178-1-Ig; 1:1,000;
ProteinTech Group, Inc.) and Bax (cat. no. 505992-2-Ig; 1:2,000;
ProteinTech Group, Inc.); the adhesion molecules intercellular cell
adhesion molecule 1 (ICAM-1; cat. no. 10831-1-AP; 1:1,000;
ProteinTech Group, Inc.), vascular cell adhesion protein 1 (VCAM-1;
cat. no. WL02474; 1:500; Wanleibio Co., Ltd.) and monocyte
chemoattractant protein 1 (MCP-1; cat. no. WL01755; 1:1,000;
Wanleibio Co., Ltd.); the Wnt/β-catenin pathway-associated proteins
β-catenin (cat. no. 51067-2-AP; 1:2,000; ProteinTech Group, Inc.),
phospho-β-catenin (cat. no. DF2989; 1:1,000; ProteinTech Group,
Inc.), low-density lipoprotein receptor-related protein 6 (LRP6;
cat. no. A13325; 1:1,000; ABclonal Biotech Co., Ltd.) and
phospho-LRP6 (cat. no. abs140173; 1:1,000; Absin Bioscience, Inc.);
and the ER stress-associated proteins GRP78 (cat. no. 66574-1-Ig;
1:5,000; ProteinTech Group, Inc.), CHOP (cat. no. WL00880; 1:800;
Wanleibio Co., Ltd.) and ATF6 (cat. no. Wl02407; 1:800; Wanleibio
Co., Ltd.). Protein content was normalized using GAPDH (cat. no.
HRP-60004; 1:8,000; ProteinTech Group, Inc.) as the internal
control. The membrane was subsequently washed three times and
incubated with horseradish peroxidase-conjugated anti-rabbit (cat.
no. BA1054; 1:8,000; Boster Biological Technology) and anti-mouse
(cat. no. SA0000I-I; 1:8,000; ProteinTech Group, Inc.) secondary
antibodies in TBST for 1 h at room temperature. Following three
additional washes, bands were detected by enhanced
chemiluminescence (cat. no. WLA006c; Wanleibio Co., Ltd.) and
autoradiography (Chemiluminescence Imaging system; version 5.1;
Bio-Rad Laboratories, Inc.). All experiments were performed in
triplicate. The results were assessed using Image Lab software
(version 5.1; Bio-Rad Laboratories, Inc.).
Statistical analysis
The data were analyzed using GraphPad Prism 6.0
(GraphPad Software, Inc.). Significant differences between groups
were determined using one-way ANOVA followed by Tukey's post-hoc
analysis. Data are presented as the mean ± SD from at least three
independent experiments. P<0.05 was considered to indicate a
statistically significant difference.
Results
Effect of simvastatin on HUVECs and its
possible mechanisms
To study the impact of simvastatin on the viability
of HUVECs pre-exposed to oxidative stress, two different
concentration series were tested. At the lower concentrations of
0.2 µM and 0.4 µM, simvastatin treatment resulted in
a decrease in extracellular LDH levels, which serve as an indicator
of cell rupture, by 47 and 57%, respectively, compared to the LDH
levels observed in control cells (20). However, the levels of LDH release
increased by 10% after treatment with 0.8 µM simvastatin
(P<0.001; n=3) compared to the control group (Fig. 1B). This demonstrated that
relatively low doses of simvastatin reduced cell damage, whereas
slightly higher doses exacerbated it. Similar dose-dependent
effects of simvastatin were observed with respect to Bax and Bcl-2
expression (Fig. 1A; P<0.05;
n=3), indicating that higher doses of simvastatin (≥0.8 µM),
as demonstrated in Fig. 1D, were
associated with a pro-apoptotic expression pattern. The MTT assay
showed that HUVEC viability decreased by 17, 32 and 65% after
treatment with 1, 2 and 4 µM simvastatin, respectively,
compared to that of untreated cells (Fig. 1C; P<0.001; n=6). Again, the
expression levels of Bax and Bcl-2 proteins reflected the degree of
cell damage at the same doses of simvastatin. Specifically, Bax
expression increased by 26, 57 and 81%, and Bcl-2 expression
decreased by 18, 33 and 60% after treatment with 1, 2 and 4
µM simvastatin, respectively (Fig. 1D; P<0.05; n=3). Since 1
µM simvastatin caused significant HUVEC injury, this
concentration was used in the following experiments.
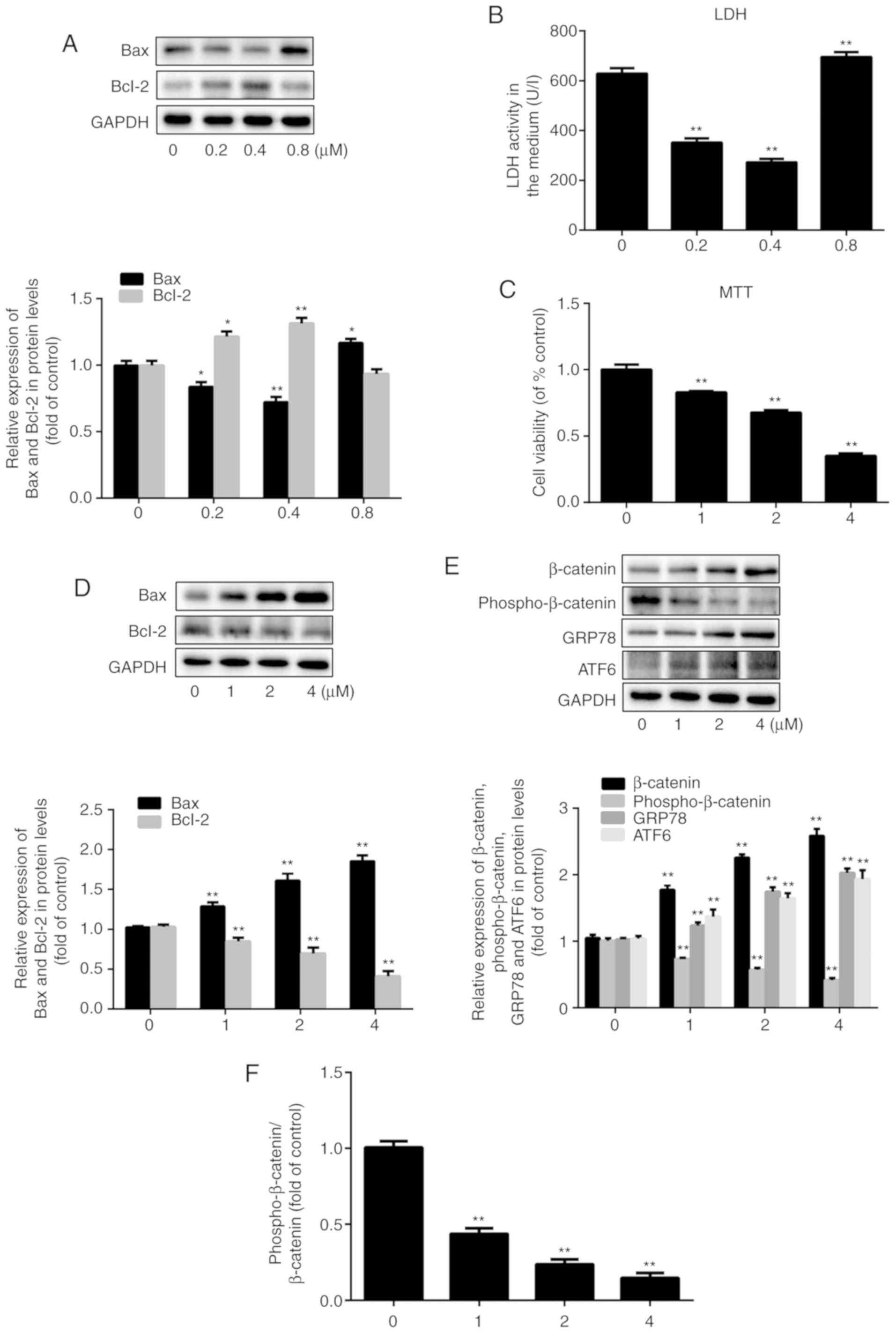 | Figure 1Effect of simvastatin on HUVECs and
its possible mechanisms. HUVECs were treated with simvastatin in
two concentration series (0, 0.2, 0.4, and 0.8 µM; 0, 1, 2,
and 4 µM) for 24 h. (A) Western blot analysis of the
expression levels of Bax and Bcl-2 after exposure to simvastatin at
0-0.8 µM. (B) Assessment of endothelial dysfunction based on
LDH release. (C) Cell viability tested by the MTT assay. (D)
Western blot analysis of the expression levels of Bax and Bcl-2
after exposure to simvastatin at 0-4 µM. (E) Western blot
analysis of the expression levels of β-catenin, phospho-β-catenin,
GRP78 and ATF6. (F) The ratio of phospho-β-catenin/β-catenin. All
values are presented as the mean ± SD; n=3 (in A, B, D, E and F)
and n=6 (in C). *P<0.05 or **P<0.01 vs.
respective 0 µM group. HUVEC, human umbilical vein
endothelial cell; LDH, lactate dehydrogenase; phosphor,
phosphorylated; GRP78, 78 kDa glucose-regulated protein; ATF6,
cyclic AMP-dependent transcription factor ATF-6α. |
Next, the effect of simvastatin on the Wnt/β-catenin
pathway was examined by evaluating the expression of β-catenin and
phospho-β-catenin, which are indicative of pathway activation and
suppression, respectively (21-23). The β-catenin expression level was
increased by 69, 115 and 146%, while the phospho-β-catenin level
was decreased by 27, 43 and 59% after treatment with 1, 2 and 4
µM simvastatin, respectively (Fig. 1E, P<0.001; n=3). Thus,
simvastatin promoted the activation of the Wnt/β-catenin pathway.
Moreover, the protein expression levels of the ER stress-related
markers GRP78 and ATF6 increased in a simvastatin dose-dependent
manner, which confirmed that ER stress was also involved in
simvastatin-induced ED (Fig. 1E;
P<0.001; n=3). The ratio of phospho-β-catenin/total β-catenin
was decreased along with the increase in the concentration of
simvastatin. The ratio was decreased to 44, 24 and 15% of the
control value after treatment with 1, 2, and 4 µM
simvastatin (Fig. 1F; P<0.001;
n=3).
Simvastatin promotes ED by activating the
Wnt/β-catenin pathway
To verify whether simvastatin promoted HUVEC
dysfunction through the Wnt/β-catenin pathway, HUVECs were treated
with simvastatin, salinomycin or both for 24 h, and then exposed to
H2O2 for an additional 24 h. Bax expression
increased by 23% and decreased by 25% after treatment with
simvastatin and salinomycin, respectively (Fig. 2A; P<0.05 and P<0.001; n=3).
Compared to the combined treatment (Sim+Sal), simvastatin alone
increased Bax expression by 37%, while salinomycin alone decreased
the Bax level by 16%. Bcl-2 expression showed the opposite pattern
of change (Fig. 2A; P<0.05 or
P<0.001; n=3). Furthermore, the apoptosis rate HUVECs was
measured by AO/EB staining. The results confirmed the same
conclusion (Fig. 2B; P<0.05 or
P0.001; n=3); compared with the control group, the apoptosis of
HUVECS in the Sim group was significantly increased while that in
the Sal group was significantly decreased.
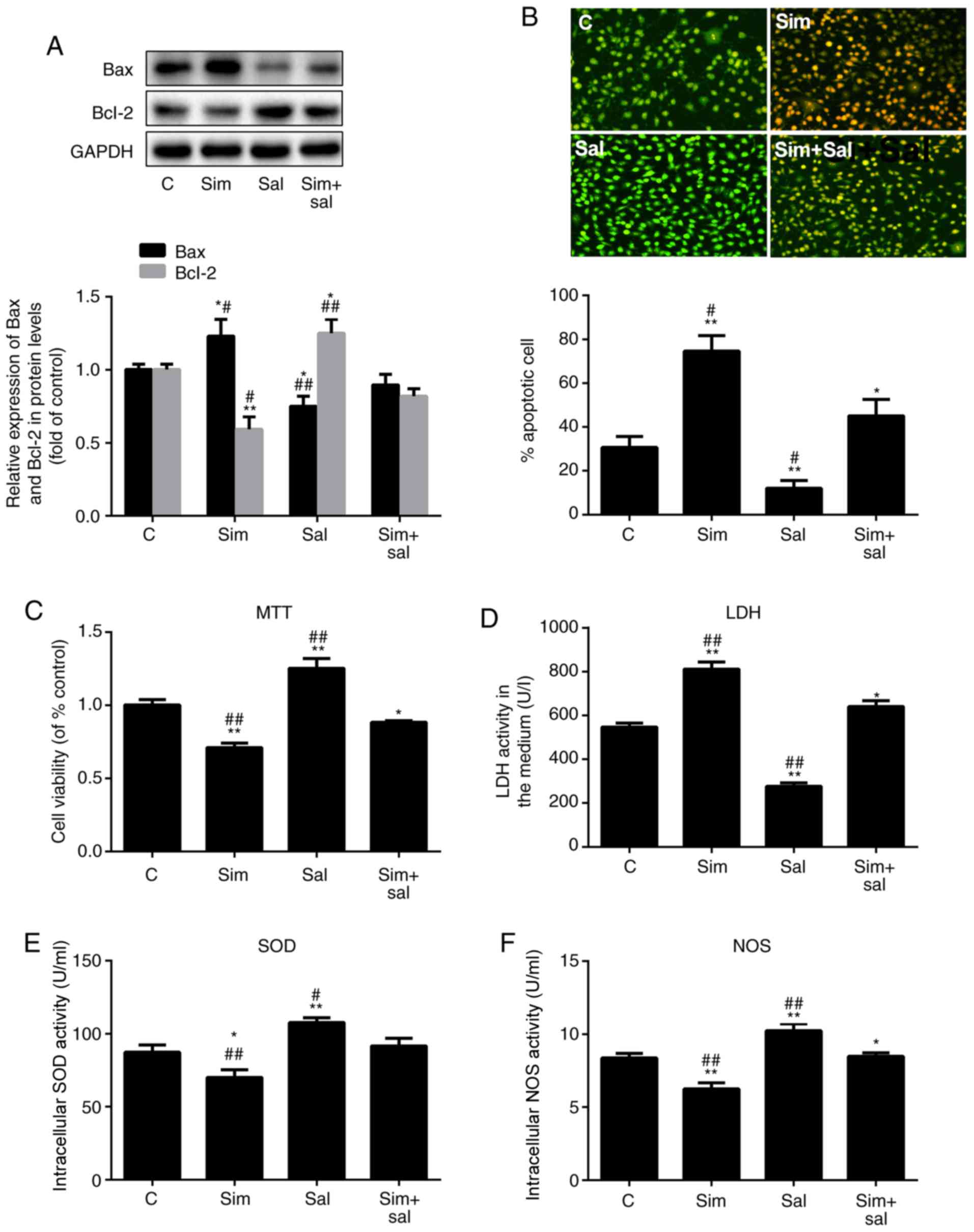 | Figure 2Simvastatin promotes ED by inducing
the Wnt/β-catenin pathway. (A) Western blot analysis of Bax and
Bcl-2 protein levels. (B) Acridine orange/ethidium bromide staining
of human umbilical vein endothelial cells; magnification, ×20. The
ED level of cells was reflected by the (C) MTT assay, (D) LDH
levels, (E) SOD activity and (F) NOS activity. All values are
presented as the mean ± SD. *P<0.05,
**P<0.01 vs. C; #P<0.05,
##P<0.01 vs. Sim + Sal. n=3 (in A, B, D, E and F) and
n=6 (in C). Sim, simvastatin; Sal, salinomycin; LDH, lactate
dehydrogenase; ED, endothelial dysfunction; NOS, nitric oxide
synthase; SOD, superoxide dismutase; C, control. |
The effects of simvastatin on cell viability were
also reversed by salinomycin (Fig.
2C; P<0.05 or P<0.001; n=6). Notably, the LDH leakage
increased by 45% after treatment with simvastatin and decreased by
49% after exposure to salinomycin compared to that of control cells
(Fig. 2D; P<0.05 or
P<0.001; n=3). The effects of simvastatin and salinomycin on the
intracellular levels of SOD (Fig.
2E; P<0.05 or P<0.001; n=3) and NOS (Fig. 2F; P<0.05 or P<0.001; n=3)
were consistent with the aforementioned results. Therefore, these
results indicated that 1 µM simvastatin promoted HUVEC
apoptosis and inhibited cell viability by activating the
Wnt/β-catenin pathway.
Simvastatin increases HUVEC adhesion
ability by activating the Wnt/β-catenin pathway
HUVEC adhesion ability to THP-1 cells may reflect
the degree of ED. To investigate the effect of the experimental
conditions on HUVEC adhesion ability, the number of THP-1 cells
adhering to HUVEC and the expression of adhesion molecules were
measured. As shown in Fig. 3A,
the number of HUVEC-attached THP-1 cells was increased by 47% and
decreased by 22% after treatment with simvastatin and salinomycin,
respectively, compared to control cells (P<0.001; n=3).
Moreover, treatment with simvastatin and salinomycin alone resulted
in a 32% increase and a 31% decrease in the number of
HUVEC-attached THP-1 cells, respectively, compared to the combined
treatment (P<0.001; n=3). Consistent results were obtained in
terms of the protein levels of the adhesion markers VCAM-1, ICAM-1
and MCP-1, which were increased by 43, 17 and 26%, respectively,
after treatment with simvastatin, and were decreased by 31, 38 and
17%, respectively, after exposure to salinomycin, compared to those
of control cells (Fig. 3B;
P<0.05 or P<0.001; n=3). Collectively, these results suggest
that simvastatin promoted the adhesion ability of HUVECs through
the Wnt/β-catenin pathway.
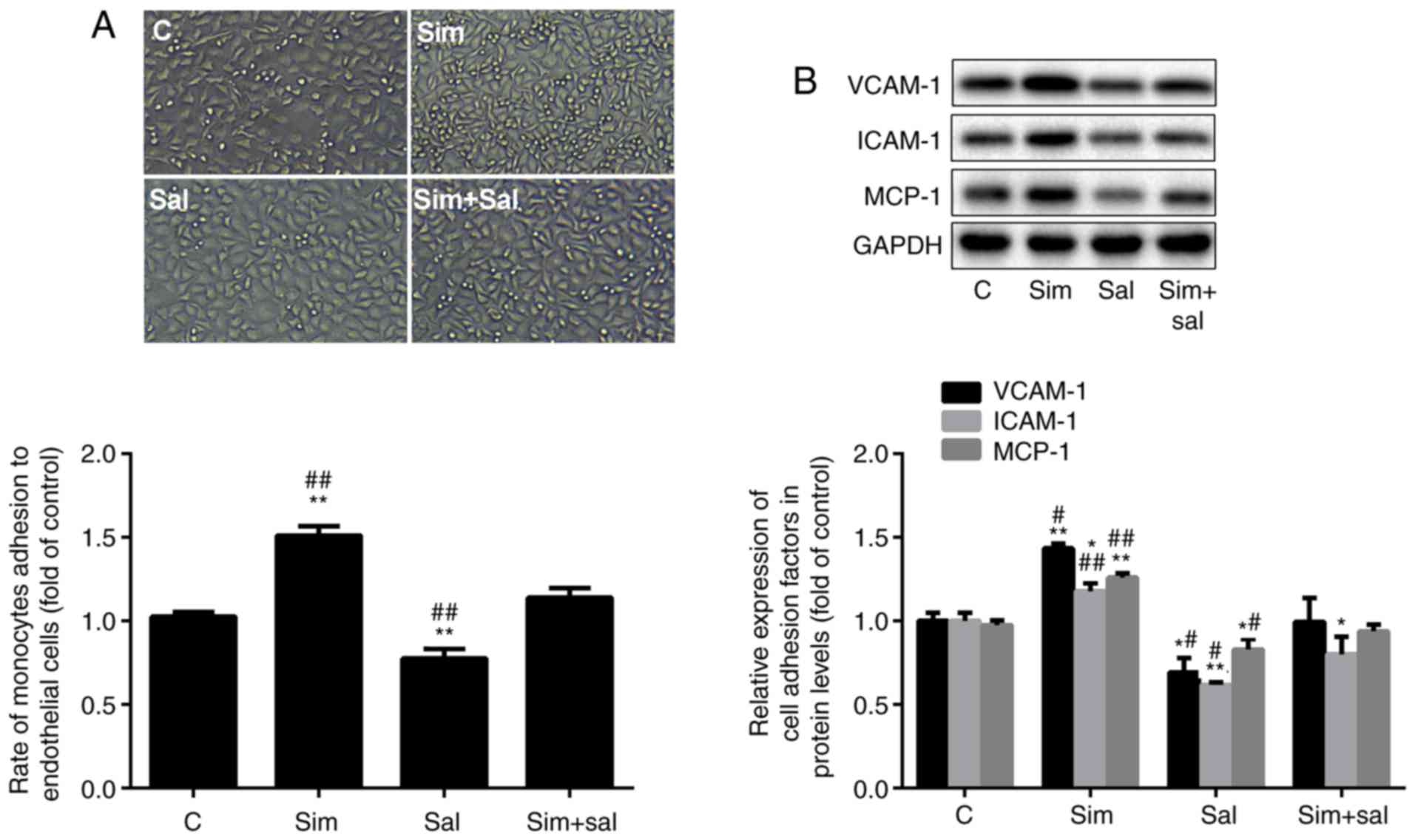 | Figure 3Simvastatin increases the adhesion
ability of HUVECs by inducing the Wnt/β-catenin pathway. (A)
Adhesion rate of HUVECs to THP-1 cells; magnification, ×20. (B)
Western blot analysis of VCAM-1, ICAM-1 and MCP-1 protein levels
(mean ± SD). *P<0.05, **P<0.01 vs. C;
#P<0.05, ##P<0.01 vs. Sim + Sal. n=3.
HUVEC, human umbilical vein endothelial cell; VCAM-1, vascular cell
adhesion protein 1; ICAM-1, intercellular adhesion molecule 1;
MCP-1, monocyte chemoattractant protein 1; C, control; Sim,
simvastatin; Sal, salinomycin. |
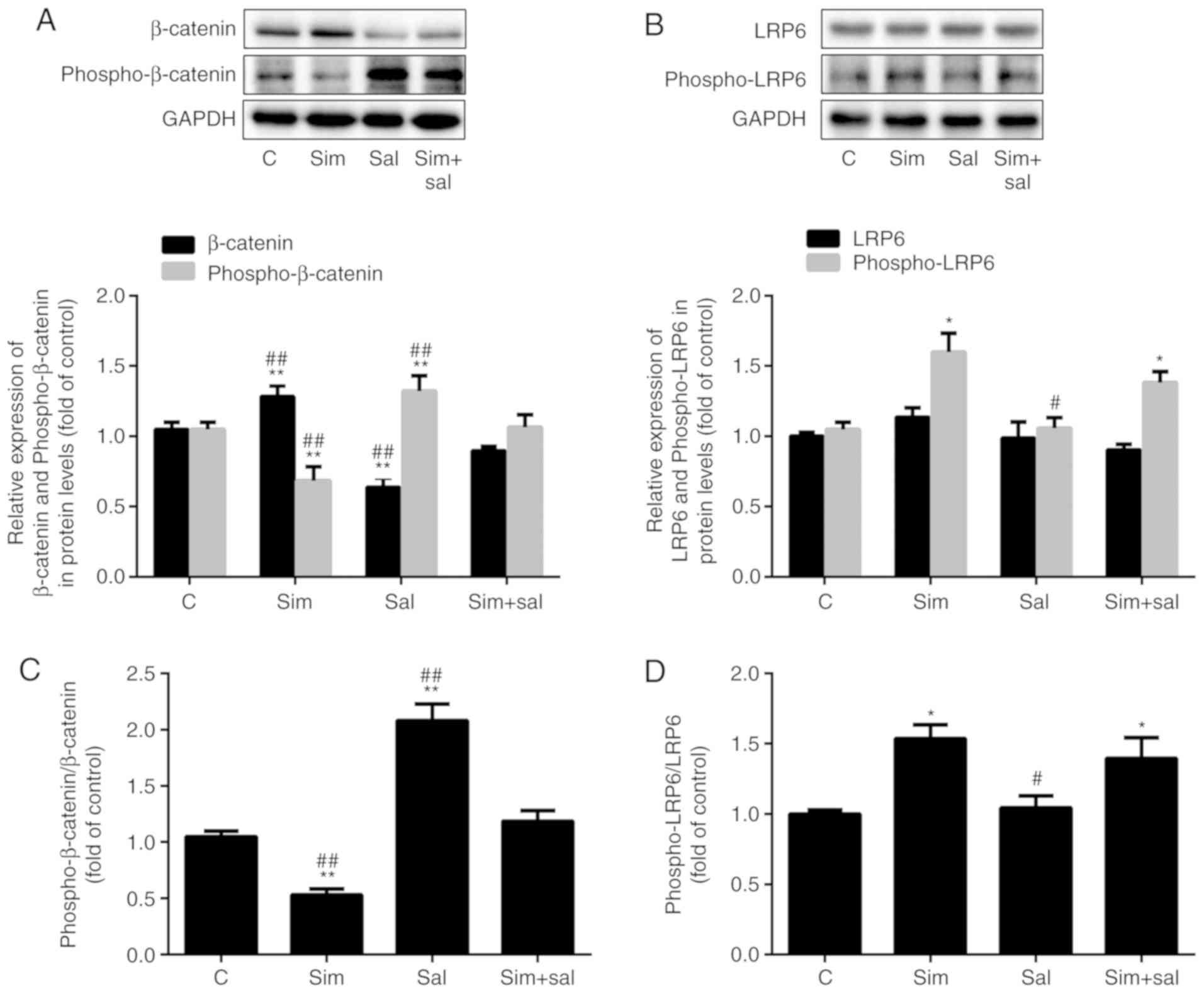
Simvastatin activates the Wnt/β-catenin
pathway by enhancing LRP6 phosphorylation
The levels of β-catenin were elevated after
treatment with simvastatin and were attenuated by salinomycin
(Fig. 4A; P<0.001; n=3).
Specifically, β-catenin protein expression was increased by 22% and
decreased by 40% after treatment with simvastatin and salinomycin,
respectively. In addition, simvastatin decreased the level of
phospho-β-catenin by 35%, while salinomycin treatment increased the
phospho-β-catenin level by 26% compared to that of control cells.
Compared to cells receiving the combined treatment, the
phospho-β-catenin content was 36% lower in cells treated with
simvastatin alone and was 24% higher in those exposed to
salinomycin alone. Analysis of the ratio of phospho-β-catenin/total
β-catenin produced similar results. As the ratio decreased by 53%
after treatment with simvastatin and increased by 90% with
salinomycin compared to that of control cells (Fig. 4B; P<0.001; n=3). The combined
treatment mitigated the effects of simvastatin and salinomycin, as
there was no statistical difference compared to the control group.
Simvastatin affects cholesterol synthesis. Cholesterol is an
important component of lipid rafts and the phosphorylation of LRP6,
an upstream element of the Wnt/β-catenin pathway, is affected by
lipid rafts (24). Notably, the
expression level of phospho-LRP6 was increased by 52% after
treatment with simvastatin (Fig.
4C; P<0.05; n=3), whereas no significant differences were
observed in total LRP6 expression. However, the ratio of
phospho-LRP6/LRP6 changed significantly. Specifically, the ratio
increased by 54 and 39% after treatment with simvastatin and the
combined treatment, respectively, compared to the control cells
(Fig. 4D; P<0.05 or
P<0.001; n=3).
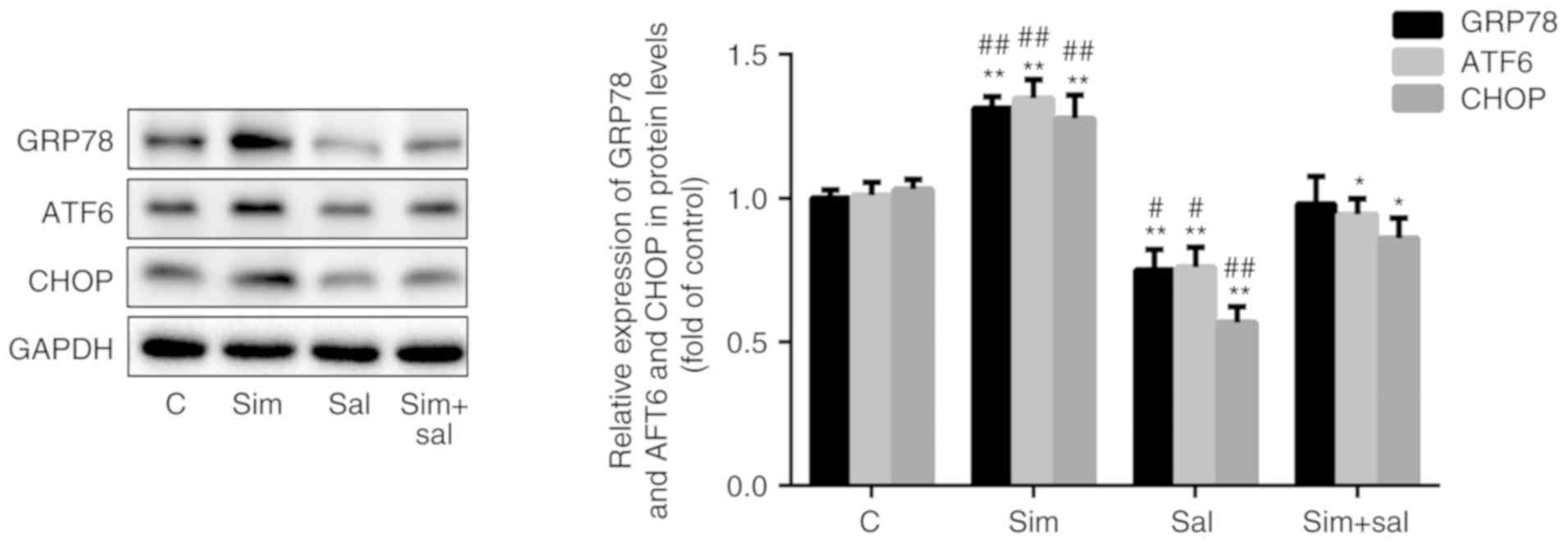
Simvastatin augments ER stress by
activating the Wnt/β-catenin pathway
To verify the impact of simvastatin on ER stress,
the expression of relevant protein markers was evaluated.
Specifically, simvastatin increased the expression of the ER
stress-related proteins GRP78, ATF6 and CHOP by 31, 33 and 24%,
respectively, while salinomycin reduced the levels of these
proteins by 25, 25 and 45%, respectively, compared to those of
control cells. As expected, the combined treatment with simvastatin
and salinomycin resulted in higher levels of ER stress markers than
those observed with salinomycin treatment alone, but lower levels
than those observed with simvastatin treatment alone (Fig. 5; P<0.05 or P<0.001; n=3).
These findings suggested that simvastatin promoted ER stress via
the Wnt/β-catenin pathway.
Discussion
Simvastatin, a commonly used lipid-lowering drug,
plays an anti-atherosclerotic role by reducing the level of blood
lipids (25). However, its
effects on endothelial cells under oxidative stress conditions and
the possible underlying mechanisms are still unclear. The results
of the present study demonstrated that when the concentration of
simvastatin was <0.8 µM,
H2O2-induced endothelial dysfunction was
significantly reduced (Fig. 1A and
B). However, at 1 µM, simvastatin significantly enhanced
ED (Fig. 1C and D) and increased
the β-catenin/phospho-β-catenin ratio, as well as the expression of
ER stress markers (Fig. 1E and
F). Therefore, it may be suggested that relatively high doses
of simvastatin (≥1 µM) may induce ED by activating the
Wnt/β-catenin pathway and ER stress.
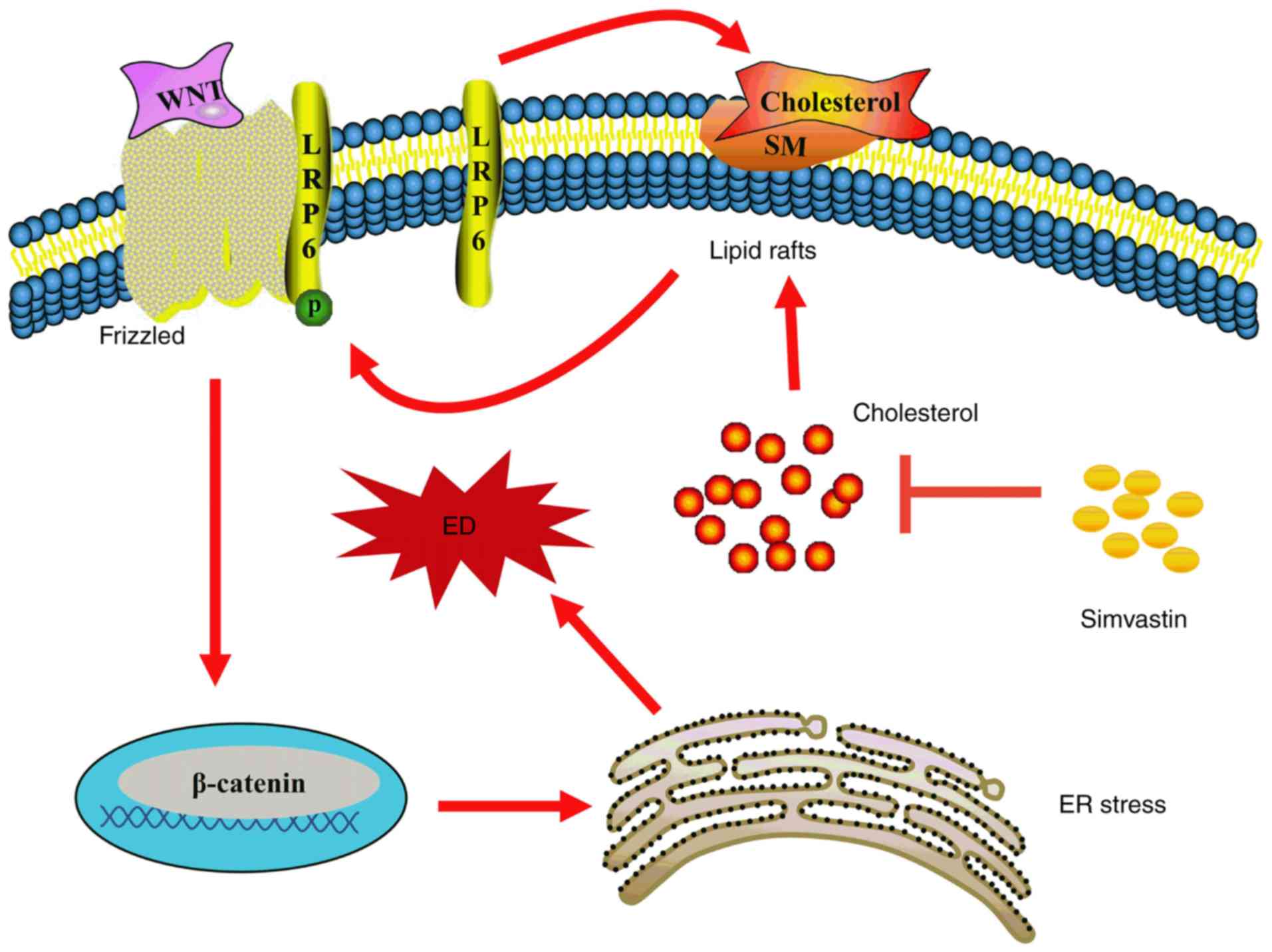
In the canonical Wnt/β-catenin pathway, LRP6 is
phosphorylated and combines with Wnt molecules and Frizzled on the
lipid raft, then delivers the signal downstream, thereby recruiting
Dishevelled homolog proteins under the membrane and facilitating
the phosphorylation of glycogen synthase kinase 3β; this results in
large-scale accumulation of β-catenin (24,26). Lipid rafts are also termed
microdomains, and are mainly anchored by cholesterol and
sphingolipids (27); they can
affect the localization of numerous receptors and signalling
proteins, and therefore have a strong influence on multiple signal
transduction cascades. For instance, lipid rafts play a vital role
in lipopolysaccharide receptor (Toll-like receptor 4) and
transforming growth factor-β receptor-mediated signal transduction
(28,29). Activation of the Wnt/β-catenin
pathway is facilitated by the localization of LRP6 in lipid rafts
(30-32). However, methyl-β-cyclodextrin
leads to the downregulation of LRP6 and β-catenin expression by
depleting cholesterol and disrupting the structure of lipid rafts
(31). Conversely, cholesterol
supplementation upregulates the expression of LRP6 and β-catenin
(33). In fact, in our previous
study, 0.2 µM simvastatin had no effect on the expression or
phosphorylation of LRP6, but reduced intracellular cholesterol
deposition and inhibited ER stress (34). However, in the present study, 1
µM simvastatin was used to inhibit cholesterol biosynthesis,
increase LRP6 phosphorylation, and activate the Wnt/β-catenin
pathway by decreasing β-catenin phosphorylation and degradation
(Fig. 4). Consequently, the
accumulated functional β-catenin in the cytoplasm may be
translocated to the nucleus and activate transcription factors such
as those in the TCF/lymphoid enhancer-binding factor (LEF) family
(35). Importantly, salinomycin,
an inhibitor of the Wnt/β-catenin pathway, reversed these
simvastatin-induced changes (Fig.
4). It may be inferred that simvastatin was able to regulate
the Wnt/β-catenin pathway by affecting the phosphorylation of LRP6
on lipid rafts (Fig. 6).
The involvement of the Wnt/β-catenin pathway in ED
has also been demonstrated in previous studies (10,36,37). Treatment of HUVECs with 1
µM simvastatin led to a significant decrease in the
intracellular activities of SOD and NOS (Fig. 2E and F). In addition, cell
integrity was markedly destroyed, as demonstrated by the
significant increase in the release of LDH into the medium
(Fig. 2D). Furthermore, the
decreased cell viability, as demonstrated in Fig. 2C, and changes in the expression of
the apoptosis-related proteins (Bax and Bcl-2) indicated that 1
µM simvastatin can exacerbate the injury to HUVECs under
oxidative stress conditions (Fig.
2A). The result of AO/EB staining also revealed that
simvastatin was able to significantly promote the apoptosis of
HUVECS under oxidative stress (Fig.
2B). In addition, this concentration of simvastatin clearly
increased the number of THP-1 cells adhering to HUVECs, as well as
the expression of adhesion molecules VCAM-1, ICAM-1 and MCP-1
(Fig. 3A and B). Salinomycin was
able to reverse all these effects. These findings collectively
demonstrated that high-dose simvastatin enhanced
H2O2-mediated ED via activation of the
Wnt/β-catenin pathway and the promotion of ER stress.
ER stress is characterized by the altered morphology
and impaired function of the ER, and results from endogenous or
exogenous stimuli that may lead to the accumulation of misfolded
proteins in the ER. The main markers of ER stress are increased
expression levels of GRP78, CHOP and ATF6 (12). Moreover, previous studies have
shown that the Wnt/β-catenin pathway is able to regulate ER stress
(32,38). For instance, inhibition of this
pathway was shown to induce ER stress, leading to apoptosis in
cancer cells (38). Furthermore,
Zhang et al (39) proposed
that inhibition of the Wnt/β-catenin pathway could cause
ATF6-related ER stress in preadipocytes by promoting β-catenin
degradation and leading to the inhibition of ATF6 functions, which
are regulated by transcription factor LEF1, thus indicating a
negative association between the Wnt/β-catenin pathway and ER
stress. By contrast, the present study showed that in HUVECs,
activation of the Wnt/β-catenin pathway induced by high-dose
simvastatin was positively correlated with ER stress (Fig. 5). Furthermore, inhibition of the
Wnt/β-catenin pathway has been demonstrated to promote apoptosis
(38), whereas activation of this
pathway has been shown to contribute to injury in HUVECs (10,26,37). Thus, the Wnt/β-catenin pathway may
be closely associated with endothelial cell dysfunction, and its
involvement in ER stress is plausible.
Many reports believe that classical pathways, such
as the PI3K/Akt, isoprenoid and small G protein pathways, are
protective to endothelial cells. Therefore, further investigation
of these pathways may provide a greater understanding of the
results of the present study. Furthermore, animal experiments or
clinical studies may contribute further knowledge in relation to
the mechanism (34,40,41). Overall, the present results
revealed a novel, alternative mechanism of action of simvastatin,
demonstrating that it can promote LRP6 phosphorylation, thereby
activating the Wnt/β-catenin pathway and promoting ER stress,
ultimately enhancing H2O2-induced HUVEC
dysfunction (Fig. 6). In
conclusion, this finding highlights the potential toxicity to
endothelial cells of simvastatin at high concentrations, as well as
its possible mechanism, providing a theoretical basis for better
clinical control of the drug concentration and prevention of the
potential toxicity and side effects of simvastatin.
Abbreviations:
|
AS
|
atherosclerosis
|
|
ED
|
endothelial dysfunction
|
|
ER
|
endoplasmic reticulum
|
|
FBS
|
foetal bovine serum
|
|
HUVECs
|
human umbilical vascular endothelial
cells
|
|
LDH
|
lactate dehydrogenase
|
|
MDA
|
malondialdehyde
|
|
NOS
|
nitric oxide synthase
|
|
SOD
|
superoxide dismutase
|
|
TBST
|
Tris-buffered saline with 0.05%
Tween-20
|
|
AO
|
acridine orange
|
|
EB
|
ethidium bromide
|
Acknowledgments
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81560151)
and the Jiangxi Provincial Department of Science and Technology
(grant no. 20181BAB205022).
Availability of data and materials
The analysed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
NY conceived the study and designed the experiments.
ZH and XD performed the experiments and data analysis. YW, LH, and
LW made substantial contributions to interpretation of data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Frostegård J: Immunity, atherosclerosis
and cardiovascular disease. BMC Med. 11:1172013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shah P, Bajaj S, Virk H, Bikkina M and
Shamoon F: Rapid progression of coronary atherosclerosis: A review.
Thrombosis. 2015:6349832015. View Article : Google Scholar
|
|
3
|
Kerr GE, Young JC, Horvay K, Abud HE and
Loveland KL: Regulated Wnt/beta-catenin signaling sustains adult
spermatogenesis in mice. Biol Reprod. 90:32014. View Article : Google Scholar
|
|
4
|
Clevers H and Nusse R: Wnt/β-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Polakis P: Drugging Wnt signalling in
cancer. EMBO J. 31:2737–2746. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gelfand BD, Meller J, Pryor AW, Kahn M,
Bortz PD, Wamhoff BR and Blackman BR: Hemodynamic activation of
beta-catenin and T-cell-specific transcription factor signaling in
vascular endothelium regulates fibronectin expression. Arterioscler
Thromb Vasc Biol. 31:1625–1633. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ueland T, Otterdal K, Lekva T, Halvorsen
B, Gabrielsen A, Sandberg WJ, Paulsson-Berne G, Pedersen TM,
Folkersen L, Gullestad L, et al: Dickkopf-1 enhances inflammatory
interaction between platelets and endothelial cells and shows
increased expression in atherosclerosis. Arterioscler Thromb Vasc
Biol. 29:1228–1234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tsaousi A, Mill C and George SJ: The Wnt
pathways in vascular disease: Lessons from vascular development.
Curr Opin Lipidol. 22:350–357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zerlin M, Julius MA and Kitajewski J:
Wnt/Frizzled signaling in angiogenesis. Angiogenesis. 11:63–69.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang P, Hua L, Hou H, Du X, He Z, Liu M,
Hu X and Yan N: Sphingomyelin synthase 2 promotes
H2O2-induced endothelial dysfunction by
activating the Wnt/β-catenin signaling pathway. Int J Mol Med.
42:3344–3354. 2018.PubMed/NCBI
|
|
11
|
Shinozaki S, Chiba T, Kokame K, Miyata T,
Kaneko E and Shimokado K: A deficiency of Herp, an endoplasmic
reticulum stress protein, suppresses atherosclerosis in ApoE
knockout mice by attenuating inflammatory responses. PLoS One.
8:e752492013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sozen E, Karademir B and Ozer NK: Basic
mechanisms in endoplasmic reticulum stress and relation to
cardiovascular diseases. Free Radic Biol Med. 78:30–41. 2015.
View Article : Google Scholar
|
|
13
|
Huang A, Patel S, McAlpine CS and Werstuck
GH: The role of endoplasmic reticulum stress-glycogen synthase
kinase-3 signaling in atherogenesis. Int J Mol Sci. 19:E16072018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Halleskog C, Mulder J, Dahlström J, Mackie
K, Schulte G, Hortobágyi T, Tanila H, Kumar Puli L, Färber K and
Harkany T: WNT signaling in activated microglia is proinflammatory.
Glia. 59:119–131. 2011. View Article : Google Scholar
|
|
15
|
Amodio G, Moltedo O, Faraonio R and
Remondelli P: Targeting the endoplasmic reticulum unfolded protein
response to counteract the oxidative stress-induced endothelial
dysfunction. Oxid Med Cell Longev. 2018:49462892018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hong J, Kim K, Park E, Lee J, Markofski
MM, Marrelli SP and Park Y: Exercise ameliorates endoplasmic
reticulum stress-mediated vascular dysfunction in mesenteric
arteries in atherosclerosis. Sci Rep. 8:79382018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Garshick M and Underberg JA: The use of
primary prevention statin therapy in those predisposed to
atherosclerosis. Curr Atheroscler Rep. 19:482017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gao K, Shen Z, Yuan Y, Han D, Song C, Guo
Y and Mei X: Simvastatin inhibits neural cell apoptosis and
promotes loco-motor recovery via activation of Wnt/β-catenin
signaling pathway after spinal cord injury. J Neurochem.
138:139–149. 2016. View Article : Google Scholar :
|
|
19
|
Robin NC, Agoston Z, Biechele TL, James
RG, Berndt JD and Moon RT: Simvastatin promotes adult hippocampal
neurogenesis by enhancing Wnt/β-catenin signaling. Stem Cell
Reports. 2:9–17. 2013. View Article : Google Scholar
|
|
20
|
Duan J, Yu Y, Li Y, Yu Y, Li Y, Zhou X,
Huang P and Sun Z: Toxic effect of silica nanoparticles on
endothelial cells through DNA damage response via Chk1-dependent
G2/M checkpoint. PLoS One. 8:e620872013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Valenta T, Hausmann G and Basler K: The
many faces and functions of β-catenin. EMBO J. 31:2714–2736. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li Y, Qin X, Li P, Zhang H, Lin T, Miao Z
and Ma S: Isobavachalcone isolated from Psoralea corylifolia
inhibits cell proliferation and induces apoptosis via inhibiting
the AKT/GSK-3β/β-catenin pathway in colorectal cancer cells. Drug
Des Devel Ther. 13:1449–1460. 2019. View Article : Google Scholar :
|
|
23
|
Wang LR, Kim SH and Baek SS: Effects of
treadmill exercise on the anxiety-like behavior through modulation
of GSK3β/β-catenin signaling in the maternal separation rat pup. J
Exerc Rehabil. 15:206–212. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Duvetorp A, Olsen RS, Nyström H, Skarstedt
M, Dienus O, Mrowietz U, Söderman J and Seifert O: Expression of
low-density lipoprotein-related receptors 5 and 6 (LRP5/6) in
psoriasis skin. Exp Dermatol. 26:1033–1038. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Harisa GI, Alomrani AH and Badran MM:
Simvastatin-loaded nanostructured lipid carriers attenuate the
atherogenic risk of erythrocytes in hyperlipidemic rats. Eur J
Pharm Sci. 96:62–71. 2017. View Article : Google Scholar
|
|
26
|
Yu X, Yan N, Li Z, Hua Y and Chen W: FGF19
sustains the high proliferative ability of keratinocytes in
psoriasis through the regulation of Wnt/GSK-3β/β-catenin signaling
via FGFR4. Clin Exp Pharmacol Physiol. 46:761–769. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chung CL, Wang SW, Sun WC, Shu CW, Kao YC,
Shiao MS and Chen CL: Sorafenib suppresses TGF-β responses by
inducing caveolae/lipid raft-mediated internalization/degradation
of cell-surface type II TGF-β receptors: Implications in
development of effective adjunctive therapy for hepatocellular
carcinoma. Biochem Pharmacol. 154:39–53. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Özhan G, Sezgin E, Wehner D, Pfister AS,
Kühl SJ, Kagermeier- Schenk B, Kühl M, Schwille P and Weidinger G:
Lypd6 enhances Wnt/β-catenin signaling by promoting Lrp6
phosphorylation in raft plasma membrane domains. Dev Cell.
26:331–345. 2013. View Article : Google Scholar
|
|
29
|
Li Z, Yang Y, Gao Y, Wu X, Yang X, Zhu Y,
Yang H, Wu L, Yang C and Song L: Elevated expression of flotillin-1
is associated with lymph node metastasis and poor prognosis in
early-stage cervical cancer. Am J Cancer Res. 6:38–50. 2015.
|
|
30
|
Haack F, Lemcke H, Ewald R, Rharass T and
Uhrmacher AM: Spatio-temporal model of endogenous ROS and
raft-dependent WNT/beta-catenin signaling driving cell fate
commitment in human neural progenitor cells. PLoS Comput Biol.
11:e10041062015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Badana AK, Chintala M, Gavara MM, Naik S,
Kumari S, Kappala VR, Iska BR and Malla RR: Lipid rafts disruption
induces apoptosis by attenuating expression of LRP6 and survivin in
triple negative breast cancer. Biomed Pharmacother. 97:359–368.
2018. View Article : Google Scholar
|
|
32
|
Cao L, Lei H, Chang MZ, Liu ZQ and Bie XH:
Down-regulation of 14-3-3β exerts anti-cancer effects through
inducing ER stress in human glioma U87 cells: Involvement of
CHOP-Wnt pathway. Biochem Biophys Res Commun. 462:389–395. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jia X, Chen Y, Zhao X, Lv C and Yan J:
Oncolytic vaccinia virus inhibits human hepatocellular carcinoma
MHCC97-H cell proliferation via endoplasmic reticulum stress,
autophagy and Wnt pathways. J Gene Med. 18:211–219. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
He Z, He X, Liu M, Hua L, Wang T, Liu Q,
Chen L and Yan N: Simvastatin attenuates
H2O2-induced endothelial cell dysfunction by
reducing endoplasmic reticulum stress. Molecules. 24:E17822019.
View Article : Google Scholar
|
|
35
|
Sun H, Wang X, Liu K, Guo M, Zhang Y, Ying
QL and Ye S: β-catenin coordinates with Jup and the TCF1/GATA6 axis
to regulate human embryonic stem cell fate. Dev Biol. 431:272–281.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kim J, Kim J, Kim DW, Ha Y, Ihm MH, Kim H,
Song K and Lee I: Wnt5a induces endothelial inflammation via
beta-catenin-independent signaling. J Immunol. 185:1274–1282. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vikram A, Kim YR, Kumar S, Naqvi A,
Hoffman TA, Kumar A, Miller FJ Jr, Kim CS and Irani K: Canonical
Wnt signaling induces vascular endothelial dysfunction via p66 Shc
-regulated reactive oxygen species. Arterioscler Thromb Vasc Biol.
34:2301–2309. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Martinez-Font E, Felipe-Abrio I,
Calabuig-Fariñas S, Ramos R, Terrasa J, Vögler O, Alemany R,
Martín-Broto J and Obrador- Hevia A: Disruption of TCF/β-catenin
binding impairs Wnt signaling and induces apoptosis in soft tissue
sarcoma cells. Mol Cancer Ther. 16:1166–1176. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang Z, Wu S, Muhammad S, Ren Q and Sun
C: miR-103/107 promote ER stress mediated apoptosis via targeting
the Wnt3a/β-catenin/ATF6 pathway in preadipocytes. J Lipid Res.
59:843–853. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Karlson BW, Palmer MK, Nicholls SJ,
Lundman P and Barter PJ: Doses of rosuvastatin, atorvastatin and
simvastatin that induce equal reductions in LDL-C and non-HDL-C:
Results from the VOYAGER meta-analysis. Eur J Prev Cardiol.
23:744–747. 2016. View Article : Google Scholar
|
|
41
|
Barale C, Frascaroli C, Cavalot F and
Russo I: Hypercholesterolemia impairs the Glucagon-like peptide 1
action on platelets: Effects of a lipid-lowering treatment with
simvastatin. Thromb Res. 180:74–85. 2019. View Article : Google Scholar : PubMed/NCBI
|