Introduction
As an independent risk factor, aging contributes to
cardiovascular morbidity and mortality (1,2).
The characteristic changes of vascular aging include thickening and
stiffness of the vessel wall, which are considered to be largely
due to the senescence of vascular endothelial cells (ECs) (3,4).
Senescent cells usually exhibit a diminished proliferative
potential; in particular, the senescent state of cells is often
accompanied with the secretion of proteins that may promote the
senescent phenotype and adversely affect local tissue environment
(5,6). It has demonstrated that vascular ECs
are crucial for maintaining vascular homeostasis (7). ECs may affect adjacent smooth muscle
cells, and exert dominant regulatory effects on embryonic
vascularization and postnatal vascular remodeling (8,9).
Thus, EC senescence is a critical factor in aging-related vascular
disorders.
Notch signaling has attracted increasing attention
due to its key role in the regulation of events ranging from
vascular formation to vascular aging (10,11). During the development of embryonic
vasculature, the expression of Notch signaling molecules, including
Jagged1 and Notch1, determines the arterial identity of the vessel
(11). In postnatal artery
remodeling, members of the Jagged/Notch gene families are expressed
in injured arteries, which regulate cell phenotype via alterations
in cell-matrix and cell-cell interactions (12). Our previous study
demonstrated that conditional knockdown of Jagged1 expression in EC
caused thickening of the vessel wall in mice (8). However, the underlying mechanism
remains largely unknown.
It has been established that human atherogenesis is
initiated during fetal development, although and the progression
into atherosclerosis usually takes decades. With aging, the
incidence and severity of coronary artery atherosclerosis increases
(1,13). To further elucidate the role of
Jagged1 in aging-related vascular disorders, the differentially
expressed secretory protein genes regulated by Jagged1 in human
coronary arterial ECs (HCAECs) were screened. One of the regulated
secretory proteins, insulin-like growth factor-binding protein 1
(IGFBP1), was identified as a factor of interest. IGFBP1 is a
30-kDa protein that has been implicated in metabolic homeostasis.
Higher IGFBP1 levels were previously found to be associated with
lower metabolic risk (14). Its
impact on the progress of macro-vascular diseases in diabetes
attracted much attention (15);
however, its role in aging-related atherosclerosis in non-diabetes
remains largely unknown. The aim of the present study was to
determine whether there is a correlation between IGFBP1 with aging
and the severity of coronary artery diseases, and whether IGFBP1
exerts an anti-senescence effect on cultured HCAECs.
Materials and methods
Patient enrollment and study design
A total of 112 patients aged >18 years with
clinically diagnosed acute coronary syndrome according to current
guidelines were consecutively enrolled at Xinqiao Hospital between
July 2014 and July 2016. Major exclusion criteria included the
following: Diabetes or impaired glucose tolerance, obesity,
hypertension, dyslipid-emia, infection, and impaired liver or renal
function. Patients with previous percutaneous coronary intervention
or coronary artery bypass grafting were also excluded. Of the 112
enrolled subjects, the age of the patients ranged between 26 and 83
years old, and 66 cases (58.93%) were male. Patients aged ≤65 years
old were included in the young group (n=50, mean age, 51.70±6.65
years) and patients aged >65 years old were included in the old
group (n=62, mean age, 69.11±5.69 years). All the patients
underwent coronary angiography (CAG) according to standard
techniques. Fasting peripheral venous blood was collected from all
patients for the IGFBP1 assay. This study was approved by the
Institutional Ethics Committee of Xinqiao Hospital (approval no.
2014044), and the investigation conformed to the principles
outlined in the Declaration of Helsinki. Written informed consent
was obtained from all enrolled subjects.
Assessment of the severity of coronary
arterial lesions
The severity of coronary artery stenosis was
assessed by at least two experienced interventional cardiologists.
Critical coronary artery disease (CAD) was defined as a segment
with ≥50% stenosis in any major epicardial artery or any important
branch of a major epicardial coronary artery. The severity of CAD
was assessed by the number of diseased vessels and the value of the
Synergy between PCI with TAXUS and Cardiac Surgery (SYNTAX) score.
The SYNTAX score was calculated using the online updated calculator
version (http://www.syntaxscore.com), and each
coronary lesion with a stenosis diameter of ≥50% in vessels ≥1.5 mm
in diameter was scored. Patients were categorized as mild (scores
≤22) vs. moderate and severe stenosis (scores ≥23).
Cell culture, transfection of small
interfering RNA (siRNA) and microarray analysis
HCAEC cultures were obtained from PromoCell GmbH.
HCAECs were cultured in endo-thelial growth medium (EGM, Lonza
Group, Ltd.) with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) and incubated at 37°C in a humidified atmosphere
with 5% CO2, according to manufacturer's protocol. HCAEC
of passages 5-7 were transfected at 50% confluence with 20 nM siRNA
of Jagged1 (siJagged1, Shanghai GenePharma Co., Ltd.) or with 20 nM
control siRNA (siControl, Shanghai GenePharma Co., Ltd.; Table SI)
in Opti-MEM I Reduced Serum Medium (Gibco; Thermo Fisher
Scientific, Inc.), using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). After 4 h of
transfection, the medium was replaced with fresh complete media.
The cells were maintained for 48 h before total RNA was extracted
and analyzed with Human Whole Genome OneArray V6.2 (HOA6.2; Phalanx
Biotech Group) by BGI Sequencing Co., Ltd. as previously reported
(16) HOA6.2 contains 32,679 DNA
oligonucleotide probes, and each probe is a 60-mer designed in the
sense direction. Among the probes, 31,741 correspond to the
annotated genes in RefSeq v51and Ensembl r65 database. Detailed
description of microarray procedures and whole genome gene and
probe lists are available from http://www.phalanxbiotech.com/tech_support/general.php.
Cell senescence and proliferation
assay
As previously reported, in vitro endothelial
culture may serve as a marker for in vivo aging (17). Passaging may induce the senescence
of cultured ECs (18).
Approximately 3 days were required for the early passages of ECs
(<6 passages) to reach confluence. However, the majority of the
cells became non-mitotic after >10 passages (18). In our culture system, HCAECs of
passage 5 (P5) were used as young and of passage 20 (P20) were used
as old cells for comparison. Briefly, HCAECs were inoculated in
6-well plates (Corning, Inc.) in EGM with 10% FBS and incubated at
37°C in a humidified atmosphere with 5% CO2. After 24 h,
the medium was changed with fresh serum-free medium and cultured at
37°C for another 24 h. HCAECs were then treated with
H2O2 (0, 100 μmol/l) for 4 h, followed
by treatment with IGFBP1 (0, 5, 50 and 100 ng/ml; RayBiotech) or
IGFBP1 plus LY294002 (10 μmol/l, Sigma Aldrich; Merck KGaA)
in EGM (Lonza Group, Ltd.) with 0.5% FBS (Gibco; Thermo Fisher
Scientific, Inc.) at 37°C for 48 h. Subsequently, the HCAECs were
harvested for cell senescence and proliferation assays. The
senescence-associated β-gal was determined with a corresponding kit
according to the manufacturer's protocol (Sigma-Aldrich; Merck
KGaA). The proliferation assays were performed using a Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.) and
tritiated thymidine (3H-TdR) incorporation (Shanghai
Nuclear Institute) according to the manufacturer's protocols, as
previously described (9).
IGFBP1 assay
The levels of circulating IGFBP1 in the enrolled
subjects, as well as in the culture supernatant of HCAECs, were
measured using ELISA. Briefly, ~3 ml of blood sample was collected
in a vacuum tube from each patient after an overnight fast. The
blood sample was allowed to clot for 30-60 min at room temperature,
and then each clotted sample was centrifuged at 1,500 x g for 10
min at 4°C. The serum samples were then frozen at -70°C until the
concentration of IGFBP1 was measured. In HCAEC culture, following
transfecting with Jagged1 siRNA or siControl, the ECs were cultured
with serum-free medium for 48 h and the supernatants were collected
and stored as described above. The levels of IGFBP1 (sensitivity: 5
pg/ml, cat. no. ELH-IGFBP1, RayBiotech) were then assayed according
to the manufacturer's instructions. Data are expressed as units of
ng/ml.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from cultured HCAECs with an
RNAiso kit according to the manufacturer's instruction (Takara Bio,
Inc.). RT was performed using the PrimeScript® RT
reagent kit (Takara Bio, Inc.) and qPCR was performed using the
SYBR® Premix Ex Taq™ (Takara Bio, Inc.) in a 7500 Fast
Real Time PCR System (Applied Biosystems; Thermo Fisher Scientific,
Inc.) according to the manufacturers' protocols, respectively. The
reaction condition for qPCR included two stages. Stage 1, Initial
denaturation: 95°C for 30 sec; stage2, 95°C for 5 sec and 60°C for
34 sec with 40 cycles. The specific primers of IGFBP1, Jagged1 and
GAPDH were designed by Fulengene (Table SII). The 2-ΔΔCq
method was used to determine the relative mRNA expression of target
genes after normalization to housekeeping gene GAPDH as previously
reported (19).
Western blot analysis
Whole-cell lysates were prepared from cultured
HCAECs with radioimmunoprecipitation assay buffer (Santa Cruz
Biotechnology, Inc.), and the lysates were then separated via 10%
SDS-PAGE, transferred to polyvinylidene fluoride membranes, and
probed with antibodies: Rabbit anti-Jagged1 (1:1,000; cat. no.
ab109536, Abcam), rabbit anti-IGFBP1 (1:1,000; cat. no. ab181141;
Abcam), rabbit anti-Akt (1:1,000; cat. no. 4691, Cell Signaling
Technology, Inc.), rabbit anti phosphorylated (p)-Akt (1:1,000;
cat. no. 4060, Cell Signaling Technology, Inc.) and rabbit
anti-βactin (1:1,000; cat. no. ab8227; Abcam) by standard
procedures.
Statistical analysis
Statistical analysis was performed using SPSS
software, version 20.0 (IBM Corp.). Continuous variables were
summarized as mean ± standard deviation or medians and
interquartile ranges, and all categorical variables were expressed
as proportions. Two groups were compared with independent samples
t test. Datasets containing three or more groups were first
analyzed by one-way ANOVA, and significance between any two groups
was further analyzed by post hoc test. For the post hoc tests, the
least significant difference test was used for comparing three
groups and a Bonferroni test was used for comparing four groups. If
all comparisons were compared with the control group, the Dunnett's
post hoc test was used. Correlations between IGFBP1 level and
severity of coronary arterial lesions were assessed by calculating
the Spearman's correlation coefficients. P<0.05 was considered
to indicate a statistically significant difference.
Results
IGFBP1 is a secretory protein regulated
by Jagged1 in HCAECs
siRNA-mediated knockdown demonstrated that
inhibition of Jagged1 expression (Fig. 1A) in HCAECs led to the
upregulation of 17 and the downregulation of 78 genes by >3-fold
(Fig. 1B). Among the
downregulated genes, IGFBP1 encodes a secretory protein and has
been reported to be associated with cardiovascular disease risk and
formation of vascular lesions (20,21). The downregulation of IGFBP1 mRNA
was further confirmed by the reduced protein expression of IGFBP1,
both in HCAECs (0.498±0.060 vs. 0.330±0.019; P<0.01) and in the
culture supernatant (197.93±20.81 pg/ml vs. 86.93±13.35 pg/ml;
P<0.01) (Fig. 1C-F).
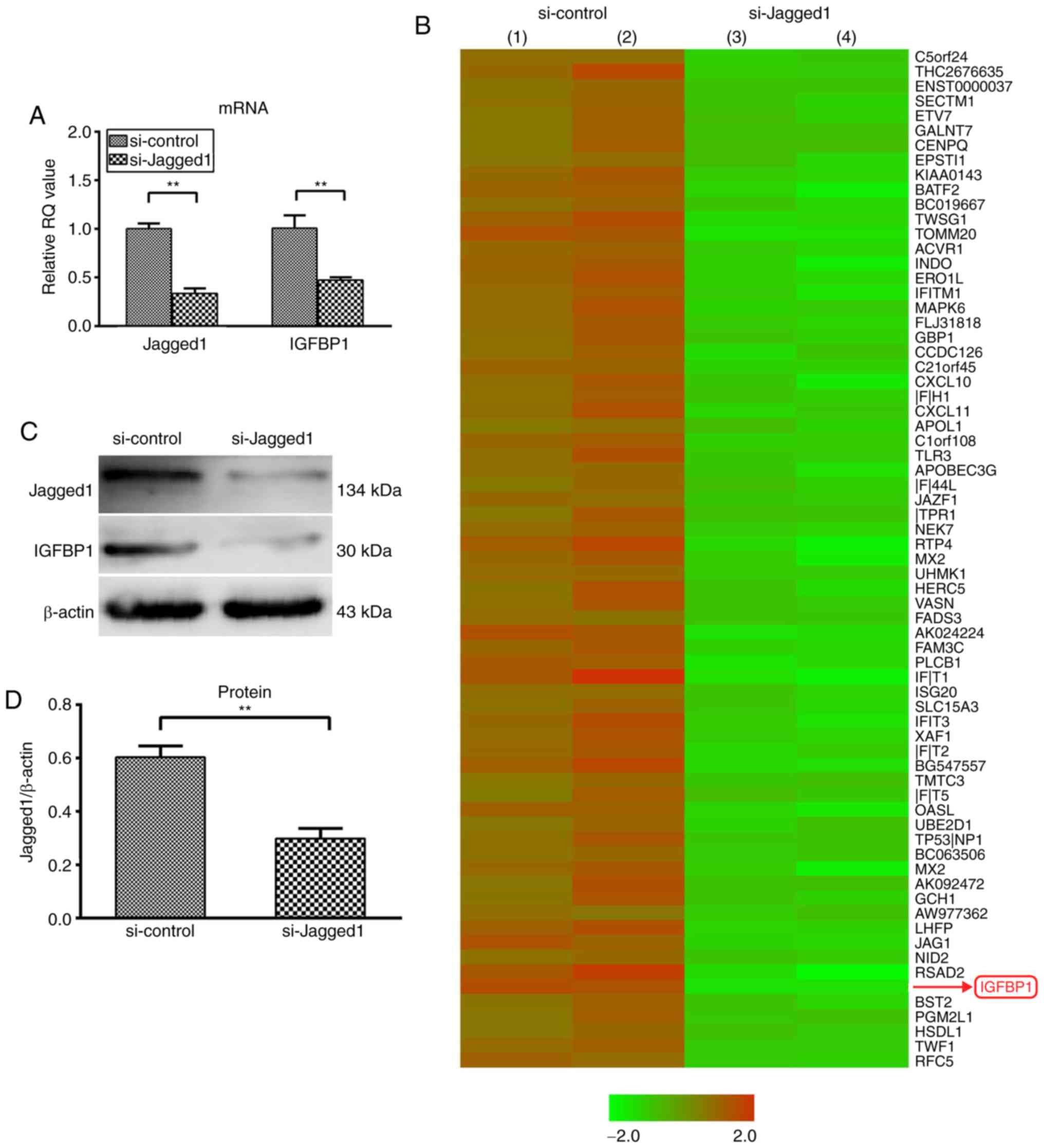 | Figure 1Knockdown of Jagged1 decreased the
expression of IGFBP1 in HCAEC and in the supernatant. (A) Reverse
transcription-quantitative polymerase chain reaction analysis of
the relative mRNA expression of Jagged1 and IGFBP1 in HCAEC 48 h
after transfection with siJagged1 or siControl. One of two
independent experiments, each with three biological replicates, is
shown. (B) Microarray screening of genes regulated by Jagged1 in
HCAECs 48 h after trans-fection with siJagged1 or siControl. Red
represented upregulation and green indicated downregulation.
Representative images of two independent experiments, each with two
biological replicates. (C) Representative image of western blot
detection of Jagged1 and IGFBP1 protein in HCAEC 48 h after
transfection with siJagged1 or siControl. Quantification of western
blot band density of (D) Jagged1 and (E) IGFBP1. One of two
independent experiments, each with three biological replicates, is
shown. (F) Quantification of IGFBP1 protein by ELISA in the culture
supernatant of HCAECs 48 h after transfection with siJagged1 or
siControl. One of two independent experiments, each with three
biological replicates, is shown. For A, D, E and F, data are
presented as the mean ± standard deviation; **P<0.01.
IGFBP1, insulin-like growth factor-binding protein 1; HCAECs, human
coronary arterial endothelial cells; si, small interfering RNA. |
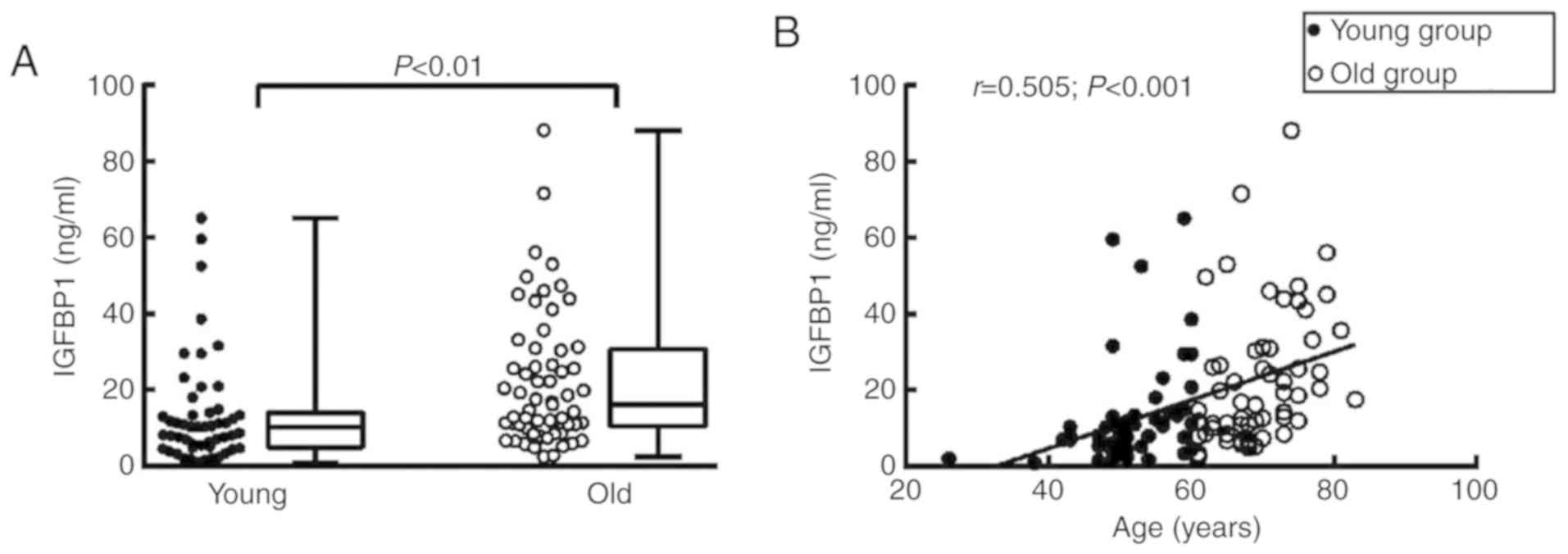
Circulating IGFBP1 levels are positively
correlated with patient age
To elucidate the role of IGFBP1 in aging-related
coronary atherosclerosis, the association of circulating IGFBP1
levels with age was first examined. The basic clinical data of the
enrolled subjects are shown in Table
I. Of the 112 enrolled subjects, the circulating level of
IGFBP1 in the old group [median, 15.41 ng/ml; interquartile range
(IQR), 9.64-30.54 ng/ml] was higher compared with that in the young
group (median, 10.30 ng/ml; IQR, 5.14-14.25 ng/ml) (Table II; Fig. 2A). The detected IGFBP1 level was
positively correlated with the age of all enrolled subjects
(r=0.505, P<0.001) (Fig.
2B).
 | Table IBasic data of enrolled subjects. |
Table I
Basic data of enrolled subjects.
| Variables | Total subjects
(n=112) |
|---|
| Age, years | 61.34±10.63 |
| Male sex, n
(%) | 66 (58.93) |
| BMI,
kg/m2 | 23.78±2.79 |
| Cigarette smoking,
n (%) | 48 (42.86) |
| Blood pressure, n
(%) | 54 (48.21) |
| Systolic,
mmHg | 132.56±18.21 |
| Diastolic,
mmHg | 76.59±11.34 |
| Lipids | 35 (31.25) |
| TC, mmol/l | 4.19±0.98 |
| TG, mmol/l | 1.27
(0.96-1.48) |
| ApoB, g/l | 0.77±0.21 |
| HDL, mmol/l | 1.10±0.28 |
| LDL, mmol/l | 2.55±0.71 |
| Medications | |
| Anti-platelets, n
(%) | 95 (84.82) |
| ACEIs or ARBs, n
(%) | 75 (66.96) |
| β-blockers, n
(%) | 76 (67.86) |
| Calcium channel
blockers, n (%) | 22 (19.64) |
| Statins, n
(%) | 95 (84.82) |
| Diuretics, n
(%) | 14 (12.50) |
| Coronary artery
disease, n (%) | 67 (59.82) |
| Multi-vessel
lesions | 41 (36.61) |
| Single-vessel
lesions | 26 (23.21) |
| Glucose,
mmol/l | 4.72±0.60 |
| BNP, pg/ml | 28.95
(10.23-64.23) |
| IGFBP1, ng/ml | 11.85
(7.11-24.67) |
| Table IIBasic clinical data of young and old
patients. |
Table II
Basic clinical data of young and old
patients.
| Variables | Young group (≤65
years) (n=50) | Old group (>65
years) (n=62) | P-value |
|---|
| Age, years | 51.70±6.65 | 69.11±5.69 | <0.001 |
| Male sex, n
(%) | 28 (56.00) | 38 (61.29) | 0.572 |
| BMI,
kg/m2 | 24.11±2.67 | 23.51±2.89 | 0.257 |
| Cigarette smoking,
n (%) | 22 (44.00) | 26 (41.94) | 0.826 |
| Blood pressure, n
(%) | 22 (44.00) | 32 (51.61) | 0.423 |
| SBP, mmHg | 131.58±18.13 | 133.35±18.39 | 0.610 |
| DBP, mmHg | 78.70±11.90 | 74.89±10.67 | 0.077 |
| Lipids | 19 (38.00) | 16 (25.81) | 0.166 |
| TC, mmol/l | 4.37±1.06 | 4.04±0.88 | 0.069 |
| TG, mmol/l | 1.42
(1.04-1.97) | 1.15
(0.91-1.49) | 0.022 |
| ApoB, g/l | 0.81±0.24 | 0.73±0.18 | 0.039 |
| HDL, mmol/l | 1.07±0.27 | 1.13±0.29 | 0.306 |
| LDL, mmol/l | 2.73±0.78 | 2.40±0.63 | 0.014 |
| Medications | | | |
| Antiplatelets, n
(%) | 42 (84.00) | 53 (85.48) | 0.828 |
| ACEIs or ARBs, n
(%) | 32 (64.00) | 43 (69.35) | 0.549 |
| β-blockers, n
(%) | 35 (70.00) | 41 (66.13) | 0.663 |
| CCB, n (%) | 9 (18.00) | 13 (20.97) | 0.694 |
| Statins, n
(%) | 43 (86.00) | 52 (83.87) | 0.755 |
| Diuretics, n
(%) | 4 (8.00) | 10 (16.13) | 0.196 |
| Coronary artery
disease, n (%) | 28 (56.00) | 39 (62.90) | 0.459 |
| Multi-vessel
lesions, n (%) | 13 (26.00) | 28 (45.16) | 0.036 |
| Single-vessel
lesions, n (%) | 15 (30.00) | 11 (17.74) | 0.127 |
| Glucose,
mmol/l | 4.68±0.56 | 4.75±0.64 | 0.599 |
| BNP, pg/ml | 19.05
(5.65-43.15) | 42.05
(13.95-79.00) | 0.006 |
| IGFBP1 (ng/ml) | 10.30
(5.14-14.25) | 15.41
(9.64-30.54) | 0.001 |
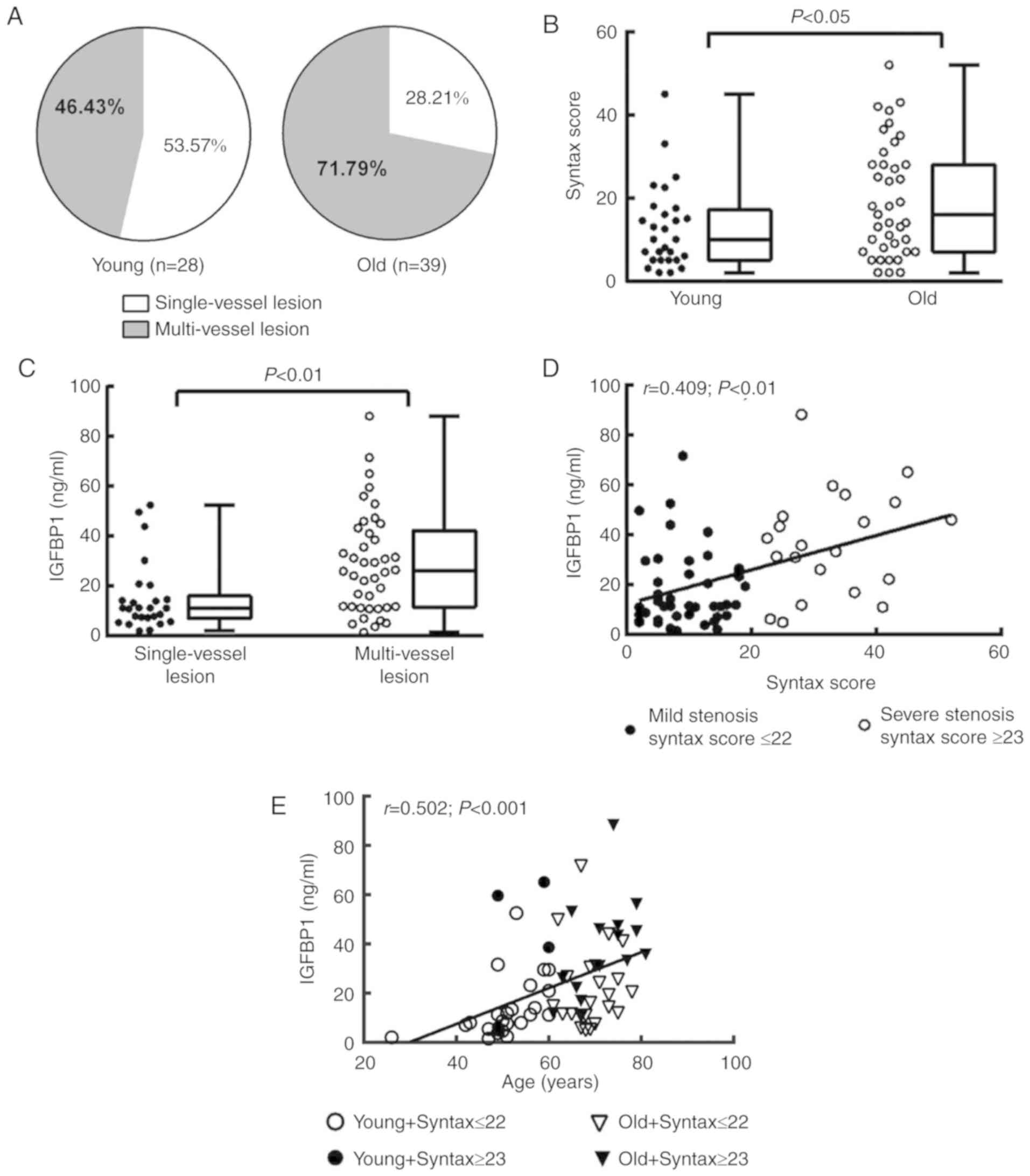
Association between IGFBP1 level and
aging-related coronary atherosclerosis
We then investigated the associations between the
severity of coronary arterial lesions and circulating IGFBP1 levels
in each age group. Of the 112 enrolled subjects, 67 (59.82%) were
diagnosed by CAG with critical CAD and 45 cases (40.18%) with
non-critical CAD (Table I). Among
patients with critical CAD, 28 cases (41.79%, data not shown) were
in the young group and 39 cases (58.21%, data not shown) were in
the old group (Fig. 3A, Table II). The incidence of multi-vessel
disease and SYNTAX score were significantly higher in old critical
CAD patients compared with those in young critical CAD patients,
suggesting a link between aging and the severity of coronary
arterial lesions (Fig. 3A and
B).
In addition, among the 67 patients with critical
CAD, the circulating level of IGFBP1 was significantly higher in
patients with multi-vessel lesions (26.08 ng/ml; IQR, 11.63-42.26
ng/ml) compared with those with single-vessel lesions (11.26 ng/ml;
IQR, 7.01-16.13 ng/ml, P<0.01), and IGFBP1 was positively
correlated with SYNTAX score (Fig. 3C
and D). Consistently, the circulating level of IGFBP1 was
positively correlated with age in critical CAD patients. In
particular, among age-comparable patients with critical CAD, the
circulating level of IGFBP1 was increased in patients with higher
SYNTAX scores (Fig. 3E),
suggesting a potential link between circulating IGFBP1 level and
aging-related severity of coronary atherosclerosis.
IGFBP1 protects HCAEC against
senescence
To explore the role of increased IGFBP1 level in
aging-related coronary arterial lesions, we further examined the
effects of IGFBP1 on the senescence of HCAECs. P5 HCAECs were used
as young cells and P20 as old cells for comparison. We found that
very few β-gal-positive cells were present in the P5 EC cultures,
while a number of strongly β-gal-positive cells was found in the
P20 cultures. The ratio of β-gal-positive cells was significantly
higher among P20 cells compared with that among P5 cells at
baseline conditions (30.20±4.82% vs. 5.40±2.07%, respectively;
P<0.001) (Fig. 4A and B).
These findings suggested that P5 cells were in a relatively
juvenile status while P20 cells more senescent. Following treatment
with 50 ng/ml IGFBP1 for 48 h, the senescence among P5 cells did
not change (5.60±1.52% vs. 4.40±1.67%; P=0.269). Instead, IGFBP1
significantly decreased the ratio of β-gal-positive cells among P20
cells (29.60±5.27% vs. 12.80±3.35%; P<0.001) (Fig. 4A and B).
H2O2 is a potent senescence inducer. To
further understand the role of IGFBP1 in EC senescence and
oxidative stimulation in aging, we next examined the protective
effects of IGFBP1 against H2O2-induced
senescence in P20 HCAECs. Following incubation with 100
μmol/l of H2O2 for 4 h, the ratio of
β-gal-positive cells increased significantly; however, the
ensuing treatment with IGFBP1 at 50 ng/ml for 48 h signifi-cantly
reversed this increase (84.60±6.15% vs. 44.00±4.30%; P<0.001) in
HCAECs (Fig. 4A and C). The
senescent cells often exhibit reduced proliferative potential.
Consistent with the anti-senescence effect of IGFBP1, we found that
IGFBP1 also promoted the proliferation of HCAECs in a
concentration-dependent manner, as confirmed in the assays for
CCK-8 and 3H-TdR incorporation (Fig. 4D and E).
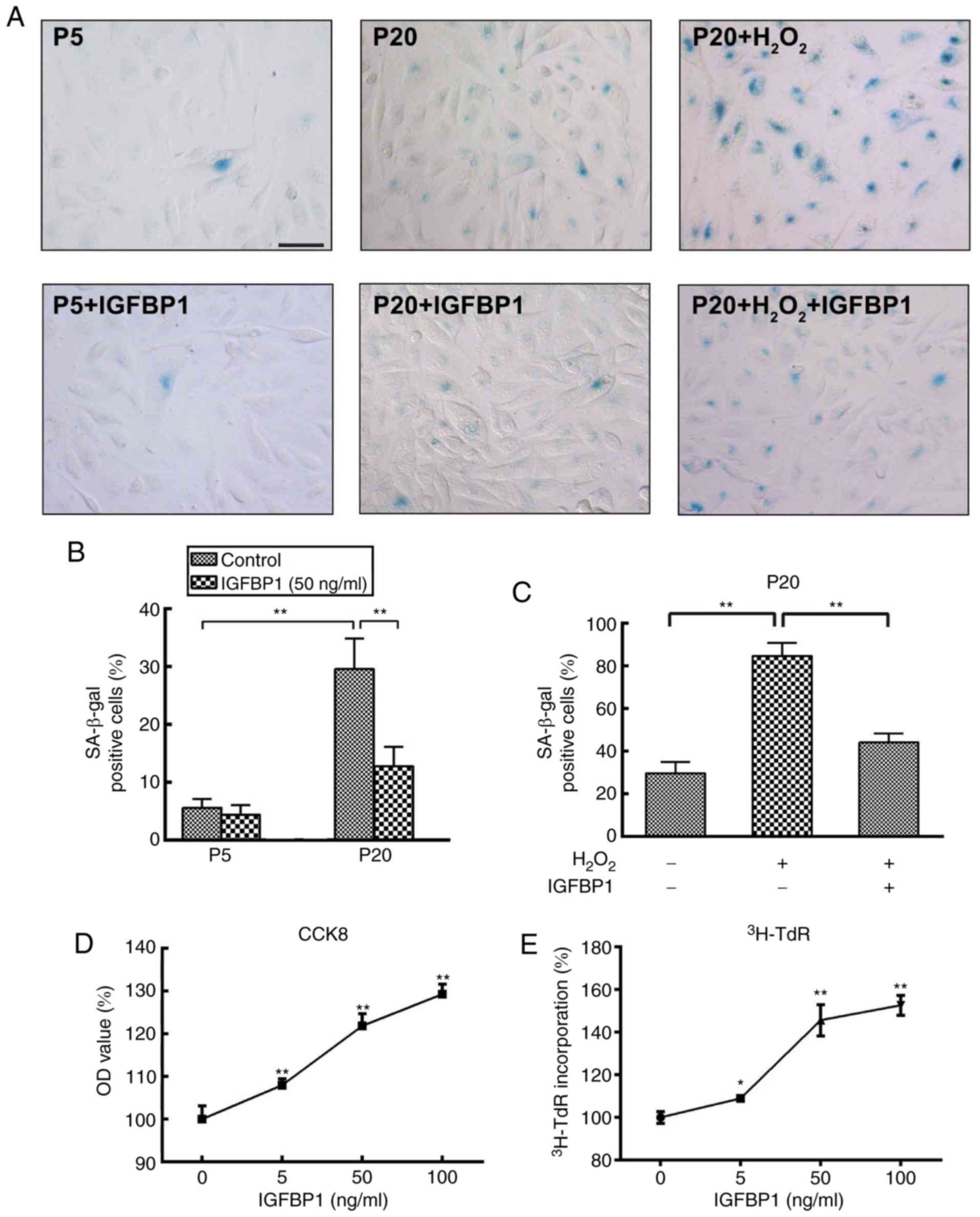 | Figure 4Effects of IGFBP1 on the senescence
and proliferation of HCAECs. (A) Representative images of β-gal
staining in HCAECs. P5 and P20 HCAECs were used as young cells and
as old cells, respectively. HCAEC were treated with
H2O2 (0, 100 μmol/l) for 4 h, followed
by treatment with IGFBP1 at 50 ng/ml for 48 h. One of two
independent experiments is shown. Scale bar, 50 μm. (B)
Quantification of β-gal-positive cells in P5 and P20 HCAECs at
baseline or treated with IGFBP1 at 50 ng/ml for 48 h. (C)
Quantification of β-gal-positive cells in HCAECs of passage 20
treated with H2O2 (100 μmol/l) for 4
h, followed by treatment with IGFBP1 at 50 ng/ml for 48 h. (D)
Quantification of HCAEC proliferation by CCK-8 assay after
treatment with IGFBP1 (0, 5, 50 and 100 ng/ml) for 48 h. (E)
Quantification of HCAEC proliferation by 3H-TdR
incorporation after treatment with IGFBP1 (0, 5, 50 and 100 ng/ml)
for 48 h. For B, C, D and E, one of two independent experiments,
each with three biological replicates, is shown. Data are presented
as the mean ± standard deviation, *P<0.05,
**P<0.01 vs. the control or as indicated. IGFBP1, insulin-like
growth factor-binding protein 1; CCK-8, Cell Counting Kit-8;
HCAECs, human coronary arterial endothelial cells; OD, optical
density; P5, passage 5; P20, passage 20; 3H-TdR,
tritiated thymidine. |
Inhibition of Akt signaling reverses the
protective effects of IGFBP1 on the senescence of HCAECs
Akt signaling regulates the senescence and
proliferation of ECs. We further studied the role of Akt signaling
in IGFBP1-regulated senescence and the proliferation of HCAECs. We
observed that IGFBP1 did not affect the total level of Akt protein
in HCAECs. IGFBP1 did not significantly affect the amount of
phosphorylated Akt (p-Akt) in the 5 ng/ml group when compared with
the control group (0 ng/ml). However, p-Akt levels were
significantly upregulated in the 50 and 100 ng/ml groups in a
concentration-dependent manner (Fig.
5A). The upregulation of p-Akt by IGFBP1 was blocked by
LY294002 (10 μmol/l), a known Akt inhibitor (Fig. 5B). In accordance with this
observation, LY294002 intervention also reversed the protective
effects of IGFBP1 at baseline and
H2O2-induced conditions, as shown by the
number of β-gal-positive senescent cells (Fig. 5C-E), and CCK-8 viability (Fig. 5F) and 3H-TdR
incorporation (Fig. 5G) assays of
cultured HCAECs.
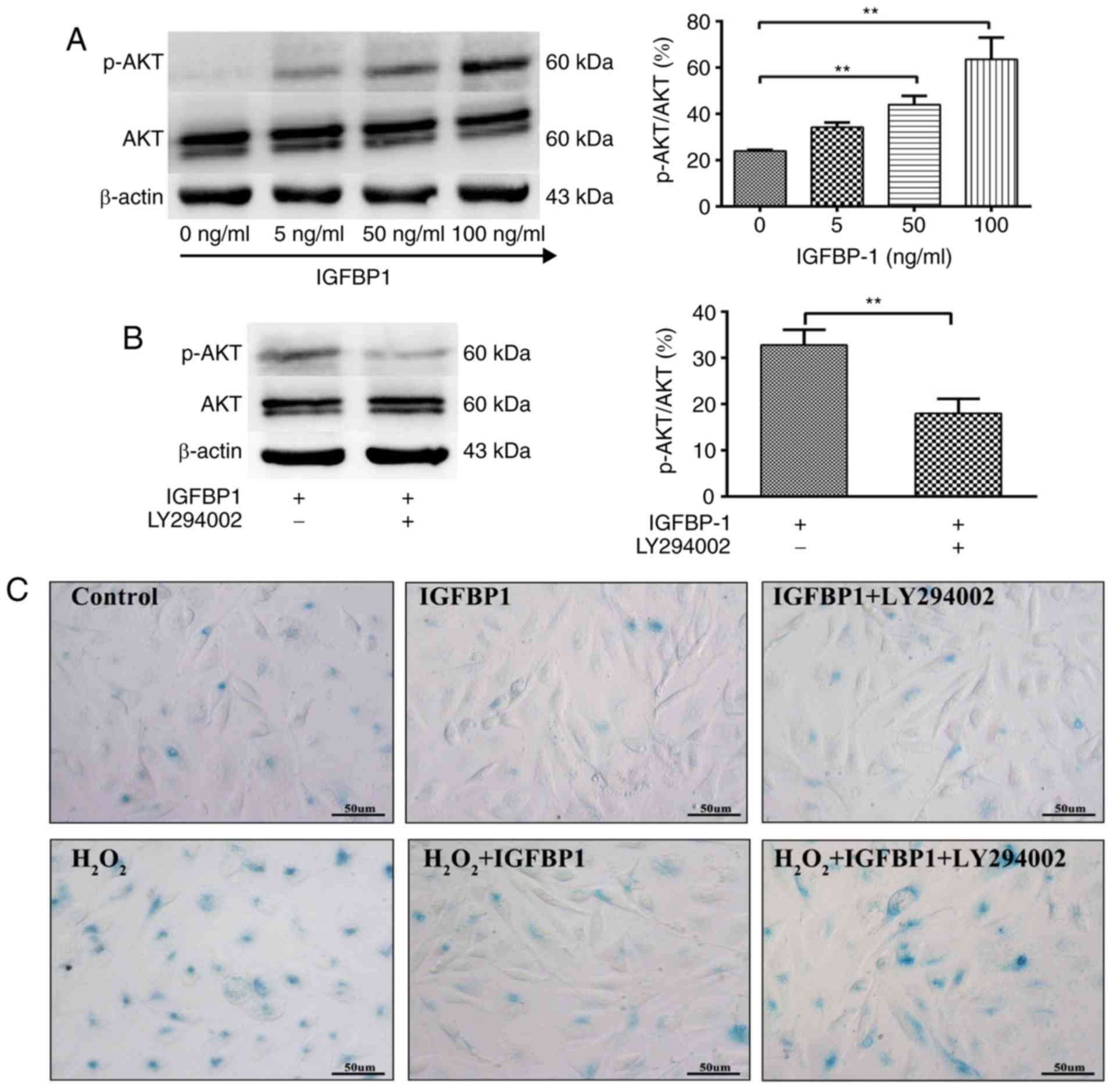 | Figure 5IGFBP1 regulates the senescence and
proliferation of HCAECs through AKT signaling. (A) Western blot
detection of AKT, p-AKT and β-actin in HCAEC 48 h after treatment
with IGFBP1 (0, 5, 50 and 100 ng/ml). (B) Western blot detection of
AKT, p-AKT and β-actin in HCAEC treated with IGFBP1 (50 ng/ml), or
IGFBP1 (50 ng/ml) plus LY294002 (10 μmol/l) for 48 h. For A
and B, left, representative images of western blots; right,
quantification of western blot band density. p-AKT expression was
normalized to both AKT and β-actin, and the quantification of
p-AKT/AKT is shown. (C) Representative images of β-gal staining of
P20 HCAECs. HCAECs were treated with H2O2
(100 μmol/l) for 4 h, followed by treatment with IGFBP1 (50
ng/ml), or IGFBP1 (50 ng/ml) plus LY294002 (10 μmol/l) for
48 h. One of two independent experiments is shown. Scale bar, 50
μm. (D) Quantification of β-gal-positive cells in P20 HCAECs
treated with IGFBP1 (50 ng/ml), or IGFBP1 (50 ng/ml) plus LY294002
(10 μmol/l) for 48 h. (E) Quantification of β-gal-positive
cells in P20 HCAECs treated with H2O2 (100
μmol/l) for 4 h, followed by treatment with IGFBP1 (50
ng/ml), or IGFBP1 (50 ng/ml) plus LY294002 (10 μmol/l) for
48 h. (F) Quantification of proliferation of P20 HCAECs treated
with IGFBP1 (50 ng/ml), or IGFBP1 (50 ng/ml) plus LY294002 (10
μmol/l) for 48 h by a CCK-8 assay. (G) Quantification of
proliferation by 3H-TdR incorporation by P20 HCAECs
treated with IGFBP1 (50 ng/ml), or IGFBP1 (50 ng/ml) plus LY294002
(10 μmol/l) for 48 h. For A, B, D, E, F and G, one of two
independent experiments, each with three biological replicates, is
shown. Data are presented as the mean ± standard deviation.
*P<0.05, **P<0.01. AKT, protein kinase
B; IGFBP1, insulin-like growth factor-binding protein 1; CCK-8,
Cell Counting Kit-8; HCAECs, human coronary arterial endothelial
cells; OD, optical density; p, phosphorylated; P5, passage 5; P20,
passage 20; 3H-TdR, tritiated thymidine; SA-β-gal,
senescence-associated β-gal. |
Discussion
Recent investigations have revealed a potential link
between Notch signaling and aging (22,23). We have previously reported that
the expression of Jagged1 decreased in the endo-thelium of aging
mice, and conditional knockdown of Jagged1 expression in the
endothelium caused thickening of the vessel wall in young mice,
indicating a critical role of Jagged1 in aging-related vascular
changes (8). However, it remains
largely unknown how Jagged1 mediates these aging-related vascular
changes. In the present study, we found that knockdown of Jagged1
altered the secretory function of HCAEC. By using microarray and
protein expression analyses, we identified IGFBP1 as one of the
secretory proteins regulated by Jagged1.
IGFBP1 belongs to the IGFBP family, which comprises
six structurally similar proteins with high affinity to IGFs,
namely IGFBP1 through IGFBP6. IGFBP1 modulates IGF bioactivity
through its high-affinity binding and has been implicated in both
metabolic homeostasis and cell growth (14,15). Low concentrations of IGFBP1 are
associated with insulin resistance and diabetes (20,21). In patients with diabetes, IGFBP1
levels are inversely correlated with cardiovascular disease risk,
carotid artery intima thickening and macrovascular diseases
(24-27). In other cases, IGFBP1 has been
observed to regulate cellular actions independently of IGFs
(28). The circulating level of
IGFBP1 was previously reported to be associated with age and
cardiovascular prognosis (29-31). However, its role in aging CAD
patients without diabetes and insulin resistance has not been
extensively investigated. Therefore, in the present study, we
consecutively enrolled 112 patients who were scheduled for CAG for
clinically diagnosed CAD. Consistently with previous reports
(30-32), we found that circulating IGFBP1
level was positively correlated with age among the tested patients,
and IGFBP1 levels were higher among older patients.
The severity of coronary atherosclerosis is a key
factor affecting the outcome of CAD. Currently, the severity and
complexity of the obstructive coronary lesions in the clinic are
often aggravated by the number of diseased vessels, as well as by
various scoring systems (33).
SYNTAX score is one of the quantitative tools used to measure the
complexity of coronary artery lesions, as well as to guide optimal
revascularization therapy in the clinic (34). Of the 112 enrolled subjects, 67
cases (59.82%) were confirmed as critical CAD. Among the critical
CAD patients, we found that both the incidence of multi-vessel
disease and the SYNTAX scores were higher among older patients,
indicating that aging is an important factor affecting the
development of coronary artery lesions. We also found that the
circulating IGFBP1 level was higher in patients with multi-vessel
disease compared with those with single-vessel diseases. Moreover,
the circulating IGFBP1 level was positively correlated with SYNTAX
scores. In particular, IGFBP1 was higher in patients with higher
SYNTAX scores among the age-comparable CAD patients. CAG is
currently a widely used invasive procedure in the diagnosis of CAD.
Screening and identification of CAD patients by non-invasive
biomarkers prior to a devastating clinical presentation warrants
intensive research (35,36). The data of the present study
indicate a potential association between the elevation of serum
IGFBP1 levels, and the severity and complexity of coronary
atherosclerosis.
To date, the role of IGFBP1 in cardiovascular
prognosis remains controversial. Studies by Rajwani et al
(37) and Borai et al
(38) demonstrated that increased
IGFBP1 level protected against atherosclerosis and a low level of
IGFBP1 may be a marker of coronary disease risk. Wang et al
(39) reported that increased
levels of IGFBP1 were detected in human carotid plaques, which may
play a role in fibro-proliferative processes and contribute to
plaque stability. However, in other cases, there were evidence
suggesting that an increased level of IGFBP1 is usually associated
with the release of inflammatory factors, and is correlated with
poor prognosis and high mortality rate (31,32,34). To confirm the role of IGFBP1 in
aging-related coronary atherosclerosis, we examined the effects of
IGFBP1 on the senescence of HCAEC. IGFBP1 treatment significantly
reduced both the passage- and H2O2-induced
senescence of HCAEC. This anti-senescence effect of IGFBP1 was
blocked by LY294002 through inhibiting the phosphorylation of Akt
in HCAECs. Consistent with its anti-senescence effects, IGFBP1
promoted the proliferation of HCAECs in a concentration-dependent
manner. These data suggest that the anti-senescence effects of
IGFBP1 were mediated through Akt signaling. Vascular ECs serve a
key role in the regulation of vascular remodeling and homeostasis
(40). The data of the present
study indicate that IGFBP1 plays a crucial protective role in
aging-related vascular remodeling. As a natural adaptive measure,
the elevation of IGFBP1 may boost EC regeneration in order to
counter the development of obstructive coronary artery lesions.
However, the final outcome may depend on the interactions between
protective and adverse factors.
The major limitation of the present study was the
relatively small sample of enrolled subjects. A larger randomized
sample across the gender and age range may be more definitive.
Long-term follow-up of the changing pattern of IGFBP1, as well as
its threshold, in the assessment of coronary artery lesions among
individual subjects with different ages warrants further
investigation.
It has been established that aging is an independent
risk factor for atherosclerosis (1,41);
however, how aging causes vascular lesions remains largely unknown.
We observed that, as a Jagged1-regulated secretory protein by EC,
the circulating level of IGFBP1 was positively correlated with age
and the severity of coronary atherosclerosis. Parallel to these
findings, we demonstrated that IGFBP1 protected EC against passage-
and H2O2-induced senescence via Akt signaling
in cell culture studies. Based on these observations, it appears
that IGFBP1 may play a protective role against aging-related
vascular disorders, and it may prove to be a promising biomarker
and intervention target.
Supplementary Data
Acknowledgments
The authors would like to thank Dr Fei Li of the
Center for Lung and Vascular Biology, University of Illinois at
Chicago for his critical revision of the manuscript.
Funding
The present study was supported by the National
Science Foundation of China (grant nos. 91539104 and 81370404) and
the Natural Science Foundation of Shenzhen University General
Hospital (grant no. SUGH2018QD048).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
XW and QZ conceived and designed this study and
wrote the manuscript. WZ, PJ and JH collected the data of the
enrolled patients. WZ, PJ, JH and XW conducted the experiments and
analyzed the data. All the authors have read and approved the final
version of this manuscript for publication.
Ethics approval and consent to
participate
This observational study was approved by the
Institutional Ethics Committee of Xinqiao Hospital (approval no.
2014044), and the investigation conformed to the principles
outlined in the Declaration of Helsinki.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mukamal KJ, Kronmal RA, Tracy RP, Cushman
M and Siscovick DS: Traditional and novel risk factors in older
adults: Cardiovascular risk assessment late in life. . Am J Geriatr
Cardiol. 13:69–80. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu Y, Bloom SI and Donato AJ: The role of
senescence, telomere dysfunction and shelterin in vascular aging.
Microcirculation. 26:pp. e124872019, View Article : Google Scholar
|
|
3
|
Ungvari Z, Tarantini S, Kiss T, Wren JD,
Giles CB, Griffin CT, Murfee WL, Pacher P and Csiszar A:
Endothelial dysfunction and angiogenesis impairment in the ageing
vasculature. Nat Rev Cardiol. 15:555–565. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yildiz O: Vascular smooth muscle and
endothelial functions in aging. Ann N Y Acad Sci. 1100:353–360.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Minamino T and Komuro I: Vascular cell
senescence: Contribution to atherosclerosis. Circ Res. 100:15–26.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yin H and Pickering JG: Cellular
senescence and vascular disease: Novel routes to better
understanding and therapy. . Can J Cardiol. 32:612–623. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
High FA, Lu MM, Pear WS, Loomes KM,
Kaestner KH and Epstein JA: Endothelial expression of the Notch
ligand Jagged1 is required for vascular smooth muscle development.
. Proc Natl Acad Sci USA. 105:1955–1959. 2008. View Article : Google Scholar
|
|
8
|
Wu X, Zou Y, Zhou Q, Huang L, Gong H, Sun
A, Tateno K, Katsube K, Radtke F, Ge J, et al: Role of Jagged1 in
arterial lesions after vascular injury. Arterioscler Thromb Vasc
Biol. 31:2000–2006. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wu X, Zhou Q, Huang L, Sun A, Wang K, Zou
Y and Ge J: Ageing-exaggerated proliferation of vascular smooth
muscle cells is related to attenuation of Jagged1 expression in
endothelial cells. Cardiovasc Res. 77:800–808. 2008. View Article : Google Scholar
|
|
10
|
Tetzlaff F and Fischer A: Control of blood
vessel formation by Notch signaling. Adv Exp Med Biol.
1066:319–338. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gridley T: Notch signaling in vascular
development and physiology. Development. 134:2709–2718. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lindner V, Booth C, Prudovsky I, Small D,
Maciag T and Liaw L: Members of the Jagged/Notch gene families are
expressed in injured arteries and regulate cell phenotype via
alterations in cell matrix and cell-cell interaction. Am J Pathol.
159:875–883. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rodríguez-Flores M, Rodríguez-Saldaña J,
Cantú-Brito C, Aguirre-García J and Alejandro GG: Prevalence and
severity of atherosclerosis in different arterial territories and
its relation with obesity. Cardiovasc Pathol. 22:332–338. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
van der Kaay D, Deal C, de Kort S,
Willemsen R, Leunissen R, Ester W, Paquette J, van Doorn J and
Hokken-Koelega A: Insulin-like growth factor-binding protein-1:
Serum levels, promoter polymorphism, and associations with
components of the metabolic syndrome in short subjects born small
for gestational age. . J Clin Endocrinol Metab. 94:1386–1392. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hu D, Pawlikowska L, Kanaya A, Hsueh WC,
Colbert L, Newman AB, Satterfield S, Rosen C, Cummings SR, Harris
TB, et al: Serum insulin-like growth factor-1 binding proteins 1
and 2 and mortality in older adults: The health, aging, and body
composition study. J Am Geriatr Soc. 57:1213–1218. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu YL, Chou RH, Shyu WC, Hsieh SC, Wu CS,
Chiang SY, Chang WJ, Chen JN, Tseng YJ, Lin YH, et al:
Smurf2-mediated degradation of EZH2 enhances neuron differentiation
and improves functional recovery after ischaemic stroke. EMBO Mol
Med. 5:531–547. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee MY, Wang Y and Vanhoutte PM:
Senescence of cultured porcine coronary arterial endothelial cells
is associated with accelerated oxidative stress and activation of
NFkB. . J Vasc Res. 47:287–298. 2010. View Article : Google Scholar
|
|
18
|
Shi Q, Aida K, Vandeberg JL and Wang XL:
Passage-dependent changes in baboon endothelial cells-relevance to
in vitro aging. . DNA Cell Biol. 23:502–509. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. . Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
20
|
Leinonen ES, Salonen JT, Salonen RM,
Koistinen RA, Leinonen PJ, Sarna SS and Taskinen MR: Reduced
IGFBP-1 is associated with thickening of the carotid wall in type 2
diabetes. . Diabetes Care. 25:1807–1812. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Heald AH, Siddals KW, Fraser W, Taylor W,
Kaushal K, Morris J, Young RJ, White A and Gibson JM: Low
circulating levels of insulin-like growth factor binding protein-1
(IGFBP-1) are closely associated with the presence of macrovascular
disease and hypertension in type 2 diabetes. . Diabetes.
51:2629–2636. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Quillien A, Moore JC, Shin M, Siekmann AF,
Smith T, Pan L, Moens CB, Parsons MJ and Lawson ND: Distinct Notch
signaling outputs pattern the developing arterial system. .
Development. 141:1544–1552. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Balistreri CR, Madonna R, Melino G and
Caruso C: The emerging role of Notch pathway in ageing: Focus on
the related mechanisms in age-related diseases. . Ageing Res Rev.
29:50–65. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Venkatesh D, Fredette N, Rostama B, Tang
Y, Vary CP, Liaw L and Urs S: RhoA-mediated signaling in
Notch-induced senescence-like growth arrest and endothelial barrier
dysfunction. . Arterioscler Thromb Vasc Biol. 31:876–882. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sandhu MS, Heald AH, Gibson JM,
Cruickshank JK, Dunger DB and Wareham NJ: Circulating
concentrations of insulin-like growth factor-I and development of
glucose intolerance: A prospective observational study. . Lancet.
359:1740–1745. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Heald AH, Cruickshank JK, Riste LK, Cade
JE, Anderson S, Greenhalgh A, Sampayo J, Taylor W, Fraser W, White
A and Gibson JM: Close relation of fasting insulin-like growth
factor binding protein-1 (IGFBP-1) with glucose tolerance and
cardiovascular risk in two populations. Diabetologia. 44:333–339.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lewitt MS, Hilding A, Ostenson CG, Efendic
S, Brismar K and Hall K: Insulin-like growth factor-binding
protein-1 in the prediction and development of type 2 diabetes in
middle-aged Swedish men. . Diabetologia. 51:1135–1145. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wheatcroft SB and Kearney MT:
IGF-dependent and IGF-independent actions of IGF-binding protein-1
and -2: Implications for metabolic homeostasis. . Trends Endocrinol
Metab. 20:153–162. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Janssen JA, Stolk RP, Pols HA, Grobbee DE
and Lamberts SW: Serum total IGF-I, free IGF-I, and IGFB-1 levels
in an elderly population: Relation to cardiovascular risk factors
and disease. . Arterioscler Thromb Vasc Biol. 18:277–282. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nolte AA, Movin M, Lundin H and Salminen
H: IGFBP-1 predicts all-cause mortality in elderly women
independently of IGF-I. Growth Horm IGF Res. 25:281–285. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chisalita SI, Dahlström U, Arnqvist HJ and
Alehagen U: Proinsulin and IGFBP-1 predicts mortality in an elderly
population. Int J Cardiol. 174:260–267. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rutkute K and Nikolova-Karakashian MN:
Regulation of insulin-like growth factor binding protein-1
expression during aging. . Biochem Biophys Res Commun. 361:263–269.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kalkan K, Hamur H, Yildirim E, Ipek E,
Ermis E, Ozturk M, Karal H, Korkmaz AF, Bayantemur M and Demirelli
S: The comparison of angiographic scoring systems with the
predictors of atherosclerosis. . Angiology. 69:158–163. 2018.
View Article : Google Scholar
|
|
34
|
Wykrzykowska JJ, Garg S, Girasis C, de
Vries T, Morel MA, van Es GA, Buszman P, Linke A, Ischinger T,
Klauss V, et al: Value of the SYNTAX score for risk assessment in
the all-comers population of the randomized multicenter LEADERS
(Limus Eluted from A Durable versus ERodable Stent coating) trial.
J Am Coll Cardiol. 56:272–277. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Vogel RA: Biomarkers of high-grade
coronary stenosis: Searching for seventies. J Am Coll Cardiol.
69:1157–1159. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ibrahim NE, Januzzi JL Jr, Magaret CA,
Gaggin HK, Rhyne RF, Gandhi PU, Kelly N, Simon ML, Motiwala SR,
Belcher AM and van Kimmenade RRJ: A clinical and biomarker scoring
system to predict the presence of obstructive coronary artery
disease. . J Am Coll Cardiol. 69:1147–1156. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rajwani A, Ezzat V, Smith J, Yuldasheva
NY, Duncan ER, Gage M, Cubbon RM, Kahn MB, Imrie H, Abbas A, et al:
Increasing circulating IGFBP1 levels improves insulin sensitivity,
promotes nitric oxide production, lowers blood pressure, and
protects against atherosclerosis. Diabetes. 61:915–924. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Borai A, Livingstone C, Ghayour-Mobarhan
M, Abuosa A, Shafi S, Mehta S, Heidari A, Emadzadeh A, Wark G and
Ferns G: Serum insulin-like growth factor binding protein-1
(IGFBP-1) phosphorylation status in subjects with and without
ischaemic heart disease. . Atherosclerosis. 208:593–598. 2010.
View Article : Google Scholar
|
|
39
|
Wang J, Razuvaev A, Folkersen L, Hedin E,
Roy J, Brismar K and Hedin U: The expression of IGFs and IGF
binding proteins in human carotid atherosclerosis, and the possible
role of IGF binding protein-1 in the regulation of smooth muscle
cell proliferation. . Atherosclerosis. 220:102–109. 2012.
View Article : Google Scholar
|
|
40
|
Cahill PA and Redmond EM: Vascular
endothelium-Gatekeeper of vessel health. . Atherosclerosis.
248:97–109. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Head T, Daunert S and Goldschmidt-Clermont
PJ: The aging risk and atherosclerosis: A fresh look at arterial
homeostasis. . Front Genet. 8:2162017. View Article : Google Scholar
|