Introduction
Bladder cancer (BC) is the ninth most common cancer
globally, with an annual incidence of 430,000 cases (1,2).
It is a complex disease associated with high mortality rates
without appropriate treatment (3). BC includes non-muscle-invasive BC
(NMIBC) and muscle-invasive BC (MIBC); ~10-15% of patients with
NMIBC will progress to MIBC (4).
Currently, there is no ideal treatment for high-grade invasive BC.
Radical cystectomy and chemotherapy are alternative treatment
options; however, both significantly reduce patient quality of life
and survival time (5,6). Therefore, to prevent BC progression
more effectively, the mechanisms of BC progression must be
investigated.
Circular RNAs (circRNAs) are 'covalently closed,
single-stranded transcripts comprising many RNA species' that are
ubiquitous throughout molecular biology (7). Although circRNAs have been observed
in eukaryotic cells for decades, they were mainly perceived as
products of splicing errors (8).
circRNAs are not easily separated from other RNA species by size or
electrophoretic mobility (9),
hence they are rarely studied intensively. Recent developments in
high-throughput deep sequencing and computational approaches have
piqued interest in single-stranded circRNA (10).
In the past few years, considerable efforts have
been devoted to circRNAs; it has been revealed that they are
closely related to the risk of atherosclerotic vascular disease and
neurological disorders, among others (11-13). Furthermore, an increasing number
of studies have shown that circRNAs participate in tumorigenesis,
metastasis, and other malignant cancer processes (14-18). Memczak et al (19) and Hansen et al (13) reported that circRNAs containing
microRNA (miRNA/miR) response elements could interact with miRNAs
as competitive endogenous RNAs (ceRNAs) to regulate the expression
of target mRNAs. Since then, accumulating circRNA dysregulation was
found to be associated with BC by functioning as ceRNAs (20,21). High-throughput sequencing and
transcriptional analysis have been carried out to elucidate the
association between circRNAs and BC, revealing considerable circRNA
expression in BC (22,23). Information regarding circRNA
expression in different grades of BC remains very limited;
therefore, we investigated differences in the circRNA expression
profiles of different grades of BC and normal urothelial cells to
identify novel targets for the diagnosis and treatment of BC.
In the present study, we sequenced tissues from
different grades of BC and normal controls to define their circRNA
expression profiles. We focused on hsa_circ_0137606, which was
significantly downregulated in BC, finding that it could suppress
BC cell proliferation and metastasis by sponging miR-1231. We aimed
to provide potential therapeutic targets for MIBC.
Materials and methods
Tissue collection
Our study was conducted according to the
recommendations of the Declaration of Helsinki and was approved by
the Ethics Committee of Nanjing Drum Tower Hospital and the
Affiliated Hospital of Nanjing University Medical School. Patients
with a history of other cancers, preoperative chemotherapy, or
radiotherapy were excluded. Each patient provided written informed
consent before tissue samples were collected. A total of 13
patients were employed for tissue collection (age 48-74; 5 males, 8
females), comprising high-grade BC, low-grade BC and a normal
control. All samples were placed in frozen storage tubes with
RNAlater (Thermo Fisher Scientific, Inc.), immediately frozen in
liquid nitrogen, and stored at -80°C. Three pairs of tissue
specimens were randomly chosen for high-throughput sequencing.
RNA extraction
TRIzol® (Thermo Fisher Scientific, Inc.)
was used to extract total RNA from the paired cancer and normal
control tissues according to the manufacturer's instructions. RNA
purity and concentration were checked by A260/A230 (>1.6) and OD
A260/A280 (>1.8). Quality and yield were assessed using an
Agilent 2100 Bioanalyzer (Agilent Technologies, Inc.) and RNA 6000
Nano Lab Chip Kit (Agilent Technologies, Inc.).
Sequencing process and analysis
Whole transcriptome library preparation and deep
sequencing were conducted by Biomarker Technologies Co, Ltd. Total
RNA quality and purity were determined using an ultra-micro
spectrophotometer (optical density 260 nm, NanoDrop; Thermo Fisher
Scientific, Inc.) and circRNA sequencing libraries were constructed
according to the manufacturer's recommendations. IlluminaHiSeq 4000
sequencing (Illumina, Inc.) was used to sequence the libraries.
FastQC was used to check the raw data. After the data had been
filtered, clean circRNA reads were mapped on to the human reference
genome (release hg19). Spliced reads per billion mapping was used
to quantify circRNA expression levels. DESeq R package was used to
identify the significantly dysregulated circRNAs with cut-off
criteria: P<0.05 and |log2 fold change|>1.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
After RNA was isolated from the 13 tissue specimen
pairs, cDNA was synthesized using M-MLV reverse transcriptase
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. qPCR (ABI VII7 PCR System, Applied
Biosystems; Thermo Fisher Scientific, Inc.) was conducted in a 20
μl reaction volume (10 μl SYBR Green Master Mix, 0.8
μl PCR Forward Primer (10 μM), 0.8 μl PCR
Reverse Primer (10 μM), 0.4 μl ROX, 2 μl cDNA,
and 6 μl nuclease-free water) with the following protocol:
Initiation at 95°C for 5 min, followed by 40 cycles of 95°C (5 sec)
and 60°C (34 sec). GAPDH was used as a reference and reactions were
performed in three independent wells. The 2-ΔΔCq method
(24) was used to calculate
relative RNA expression levels. Primers sequences are presented in
Table SI.
Bioinformatics analysis
GO and KEGG pathway analyses were performed to
explore the biological functions of dysregu-lated circRNAs. The R
package clusterProfiler (25) was
used to analyze biological processes, cellular components and
molecular functions enriched for circRNA-derived genes.
Hypergeometric testing was performed in the enrichment analysis to
identify GO entries significantly enriched compared with the whole
genome. The R package clusterProfiler was used for KEGG pathway
analysis to explore the biological pathways in which the
dysregulated circRNAs participate. Metascape analysis was performed
using online tool metascape (www.metascape.org). Protein-protein-interaction (PPI)
network was constructed using online tool STRING (https://string-db.org/). Online tool miRTarBase
(http://mirtarbase.mbc.nctu.edu.tw/php/index.php) was
used to investigate the miRNA-mRNA interactions.
Cell transfection with small interfering
(si)RNA
Hsa_ circ_0137606 was specifically knocked down
using siRNA (5′-GGC AGC TGA TGT GCT CAT CTT-3′) designed by
CircInteractome (26) (http://circinteractome.nia.nih.gov) and
synthesized (Shanghai GenePharma Co., Ltd.) to target the
hsa_circ_0137606 back-splice junction. Scramble siRNA (5′-CCG UGC
TGA TGT GCT CAT CTT-3′) was the negative control (NC). T24 cells
were transfected with 100 pmol of siRNA using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) and harvested after 48 or 72 h for RT-qPCR or
other experiments.
Cell culture
Human BC cell lines T24 and normal Urinary
epithelial cells SV-HUC-1 were purchased from American Type Culture
Collection. SV-HUC-1 cells were cultured in Dulbecco's Modified
Eagles medium (Gibco; Thermo Fisher Scientific, Inc.); T24 cells
were cultured with RPMI 1640 (Gibco; Thermo Fisher Scientific,
Inc.), containing 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 mg/ml streptomycin.
All these cell lines were maintained at 37°C with 5% CO2
in a humidified incubator.
Dual-luciferase reporter assay
Hsa_circ_0137606-wild type (WT) and
hsa_circ_0137606-mutant (Mut) were constructed using pGL3-Basic
luciferase vectors (Promega Corporation) and transfected into BC
cells with or without NC or miR-1231 mimics, respectively. After 48
h, a dual-luciferase reporter assay kit (Promega Corporation) was
used to determine lucif-erase activity. Renilla luciferase
was used for normalization.
The luciferase activities were measured using a
luciferase assay kit (Promega Corporation). Three independent
experiments were performed in triplicate.
Cell Counting Kit-8 (CCK-8) proliferation
assay. Transfected cells were cultured in 96-well plates (1,000
cells/well) for 0, 24, 48, 72 or 96 h. According to the
manufacturer's protocols, a CCK-8 (Dojindo Molecular Technologies,
Inc.) was used to detect cell proliferation. A total of 10
μl of CCK-8 solution was added to each well of the 96-well
plate and incubated at 37°C for 2 h. The absorbance (450 nm) was
measured using a Sunrise Microplate Reader (Tecan Group, Ltd.).
Three independent experiments were performed in triplicate.
Transwell assays
A 24-well Transwell chamber (Costar; Corning, Inc.)
precoated with or without Matrigel was used to detect cell
migratory or invasive abilities, according to the manufacturer's
instructions. Cells were cultured in the upper chamber (pre-coated
with Matrigel for the invasion assay) with 200 ml serum-free media.
The lower chamber was filled with 600 ml of 20% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.) and RPMI 1640 medium. After
48 h, the cells that had migrated to the lower chamber were fixed
using formaldehyde (4%, room temperature, 1 h) and stained with
0.1% crystal violet (room temperature, 20 min). The cells were
observed and photographed using Axio Observer D1 microscope
(magnification, ×100 times, Zeiss AG). Three independent
experiments were performed in triplicate.
Statistical analysis
SPSS 21.0 (IBM Corp.) and GraphPad Prism (GraphPad
Software, Inc.) was selected for data analysis and plotting. The
measured data were presented as the mean ± standard deviation. Each
experiment was repeated three times. The differences between groups
were assessed by a Student's t test and one-way ANOVA. Multiple
comparison between the groups was performed using a
Student-newman-Keuls post hoc-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
circRNA expression profiles for different
grades of BC and normal controls
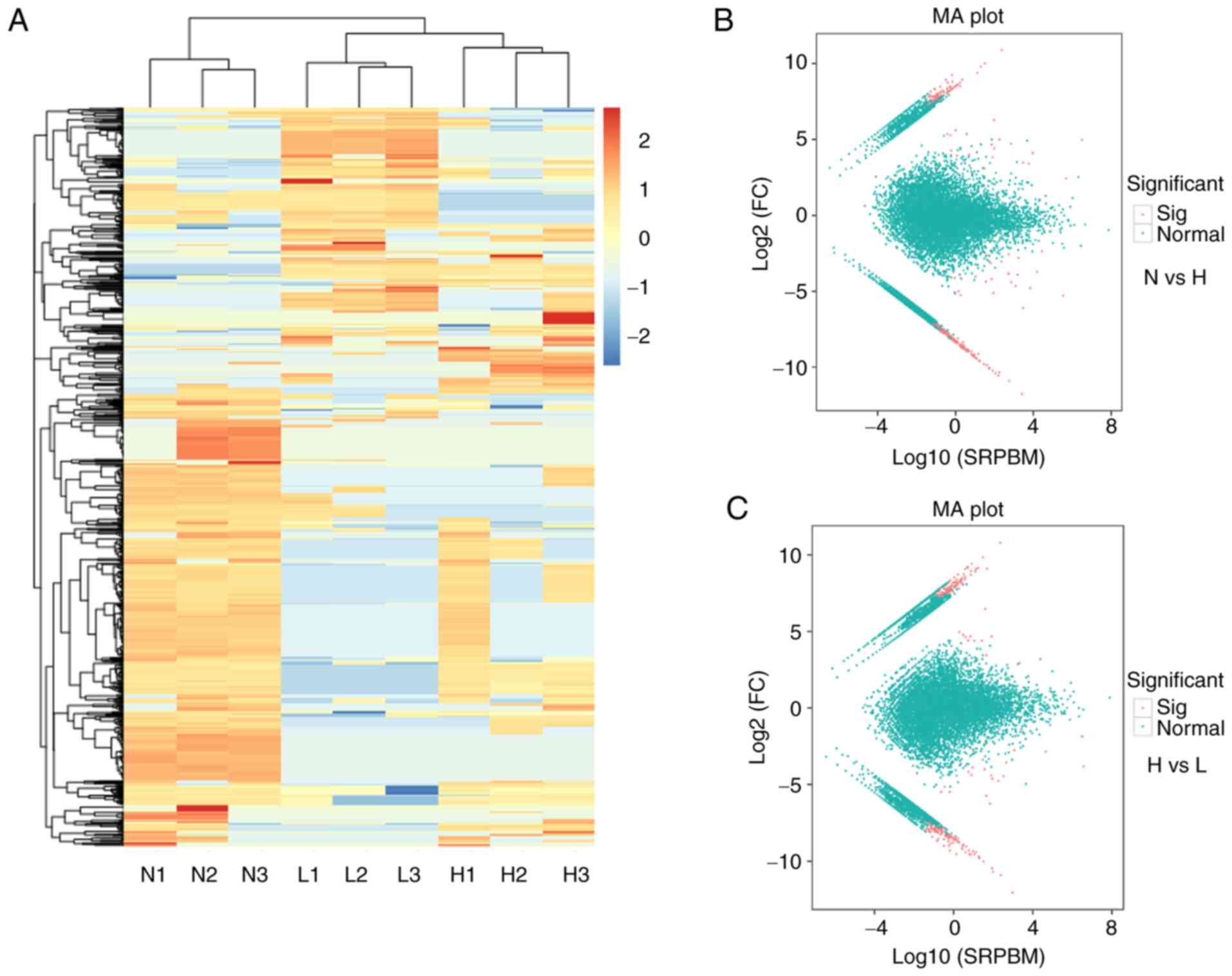
Heatmaps (Fig. 1A
and B) and MA plots (Fig. 1C and
D) were used to demonstrate variation in circRNA expression. In
high-grade BC tissues, 316 circRNAs were dysregulated compared with
NCs (205 upregulated and 111 downregulated) and 244 circRNAs were
dysregulated compared with low-grade BC tissues (109 upregulated
and 135 downregu-lated). Further analysis indicated that 42
dysregulated circRNAs overlapped between the two groups (Table I). These differentially expressed
circRNAs will be valuable in future studies to reveal the
physiopathological mechanisms of BC progression.
 | Table IOverlapped differential circRNAs in
two groups. |
Table I
Overlapped differential circRNAs in
two groups.
| circRNA | Position | Gene symbol | log2 cold
change | P-value |
|---|
| H vs. L | H vs. N | H vs. L | H vs. N |
|---|
|
hsa_circ_0020840 |
chr11:3418346-3418636 |
TCONS_l2_00004347 | 10.019 | 10.017 | 0.0103 | 0.0079 |
|
hsa_circ_0103136 |
chr15:24369721-24426402 | TCONS_00023304 | 9.809 | 9.808 | 0.0122 | 0.0094 |
|
hsa_circ_0078816 |
chr6_cox_hap2:2834337-2836058 | None | 9.398 | 9.388 | 0.0161 | 0.0127 |
|
hsa_circ_0030392 |
chr13:64608479-64608670 |
TCONS_l2_00007568 | 8.846 | 8.817 | 0.0065 | 0.0047 |
|
hsa_circ_0071818 |
chr5:9318483-9380135 | SEMA5A | 8.744 | 8.742 | 0.0301 | 0.0242 |
|
hsa_circ_0006329 |
chr5:9421788-9437964 | SEMA5A | 8.559 | 8.557 | 0.0359 | 0.0289 |
|
hsa_circ_0002099 |
chr5:9190378-9238002 | SEMA5A | 8.530 | 8.529 | 0.0370 | 0.0295 |
|
hsa_circ_0084606 |
chr8:62531536-62566219 | ASPH | 8.521 | 8.511 | 0.0357 | 0.0287 |
|
hsa_circ_0136698 |
chr8:51415337-51449368 | SNTG1 | 8.487 | 8.485 | 0.0384 | 0.0309 |
|
hsa_circ_0099364 |
chr12:83250788-83324324 | TMTC2 | 8.472 | 8.470 | 0.0391 | 0.0313 |
|
hsa_circ_0071820 |
chr5:9337824-9380135 | SEMA5A | 8.472 | 8.470 | 0.0390 | 0.0312 |
|
hsa_circ_0081963 |
chr7:111127293-111161505 | IMMP2L | 8.451 | 4.722 | 0.0128 | 0.0476 |
|
hsa_circ_0063865 |
chr22:48081948-48082955 | None | 8.395 | 8.347 | 0.0426 | 0.0359 |
|
hsa_circ_0009130 |
chr19:1147307-1154401 | SBNO2 | 8.330 | 8.328 | 0.0442 | 0.0360 |
|
hsa_circ_0114699 |
chr20:13550153-13568017 | TASP1 | 8.292 | 8.274 | 0.0027 | 0.0017 |
|
hsa_circ_0007632 |
chr20:34312491-34313077 | RBM39 | 8.255 | 8.222 | 0.0145 | 0.0110 |
|
hsa_circ_0088274 |
chr9:119976636-119977021 | ASTN2 | 8.064 | 8.032 | 0.0072 | 0.0048 |
|
hsa_circ_0115417 |
chr20:4913100-4951565 | SLC23A2 | 8.052 | 8.023 | 0.0253 | 0.0191 |
|
hsa_circ_0140725 |
chrY:14493981-14518736 | GYG2P1 | 8.042 | 8.039 | 0.0301 | 0.0221 |
|
hsa_circ_0023256 |
chr11:68529002-68530229 | CPT1A | 7.956 | 7.913 | 0.0344 | 0.0278 |
|
hsa_circ_0001377 |
chr3:195605123-195615477 | TNK2 | 7.912 | 7.901 | 0.0375 | 0.0285 |
|
hsa_circ_0139039 |
chr9:81035195-81038128 | None | 7.891 | 7.872 | 0.0363 | 0.0279 |
|
hsa_circ_0007071 |
chr5:43122140-43139411 | ZNF131 | 7.850 | 7.812 | 0.0430 | 0.0347 |
|
hsa_circ_0082583 |
chr7:138209985-138223548 | TRIM24 | 7.846 | 7.818 | 0.0406 | 0.0317 |
|
hsa_circ_0006423 |
chr1:94140169-94140497 | BCAR3 | 7.840 | 7.814 | 0.0409 | 0.0318 |
|
hsa_circ_0128896 |
chr5:34922321-34923339 | BRIX1 | 7.495 | 7.482 | 0.0322 | 0.0227 |
|
hsa_circ_0124765 |
chr3:8977554-9000685 | RAD18 | 7.349 | 7.323 | 0.0442 | 0.0328 |
|
hsa_circ_0007766 |
chr17:37864573-37866734 | ERBB2 | 3.801 | 4.970 | 0.0124 | 0.0015 |
|
hsa_circ_0072654 |
chr5:64084777-64100213 | CWC27 | -7.504 | -8.169 | 0.0403 | 0.0017 |
|
hsa_circ_0109103 |
chr19:13246013-13247219 | NACC1 | -7.520 | -7.334 | 0.0330 | 0.0347 |
|
hsa_circ_0004113 |
chr3:66293626-66313803 | SLC25A26 | -7.595 | -7.791 | 0.0267 | 0.0336 |
|
hsa_circ_0102172 |
chr14:56083234-56086030 | KTN1 | -7.677 | -7.347 | 0.0189 | 0.0302 |
|
hsa_circ_0008510 |
chr9:86279944-86292876 | UBQLN1 | -7.719 | -8.367 | 0.0166 | 0.0010 |
|
hsa_circ_0008426 |
chr4:129919028-129925031 | SCLT1 | -7.796 | -8.642 | 0.0447 | 0.0004 |
|
hsa_circ_0099708 |
chr13:100191699-100196249 | TM9SF2 | -7.863 | -8.315 | 0.0112 | 0.0013 |
|
hsa_circ_0096402 |
chr11:73418460-73429763 | RAB6A | -8.185 | -9.166 | 0.0180 | 0.0001 |
|
hsa_circ_0005004 |
chr7:44714009-44714867 | OGDH | -8.217 | -8.650 | 0.0028 | 0.0004 |
|
hsa_circ_0005746 |
chr19:55756487-55757046 | PPP6R1 | -8.332 | -7.899 | 0.0018 | 0.0060 |
|
hsa_circ_0125309 |
chr4:129042980-129043343 | LARP1B | -8.344 | -7.240 | 0.0024 | 0.0406 |
|
hsa_circ_0005429 |
chr14:67768105-67770316 | MPP5 | -8.379 | -8.285 | 0.0019 | 0.0015 |
|
hsa_circ_0006784 |
chr7:151960100-152012423 | MLL3 | -9.080 | -8.338 | 0.0003 | 0.0011 |
|
hsa_circ_0001882 |
chr9:114148656-114154104 | KIAA0368 | -9.098 | -10.335 | 0.0214 | 0.0005 |
|
hsa_circ_0137606 |
chr9:107513236-107521452 | NIPSNAP3A | -9.204 | -8.101 | 0.0213 | 0.0494 |
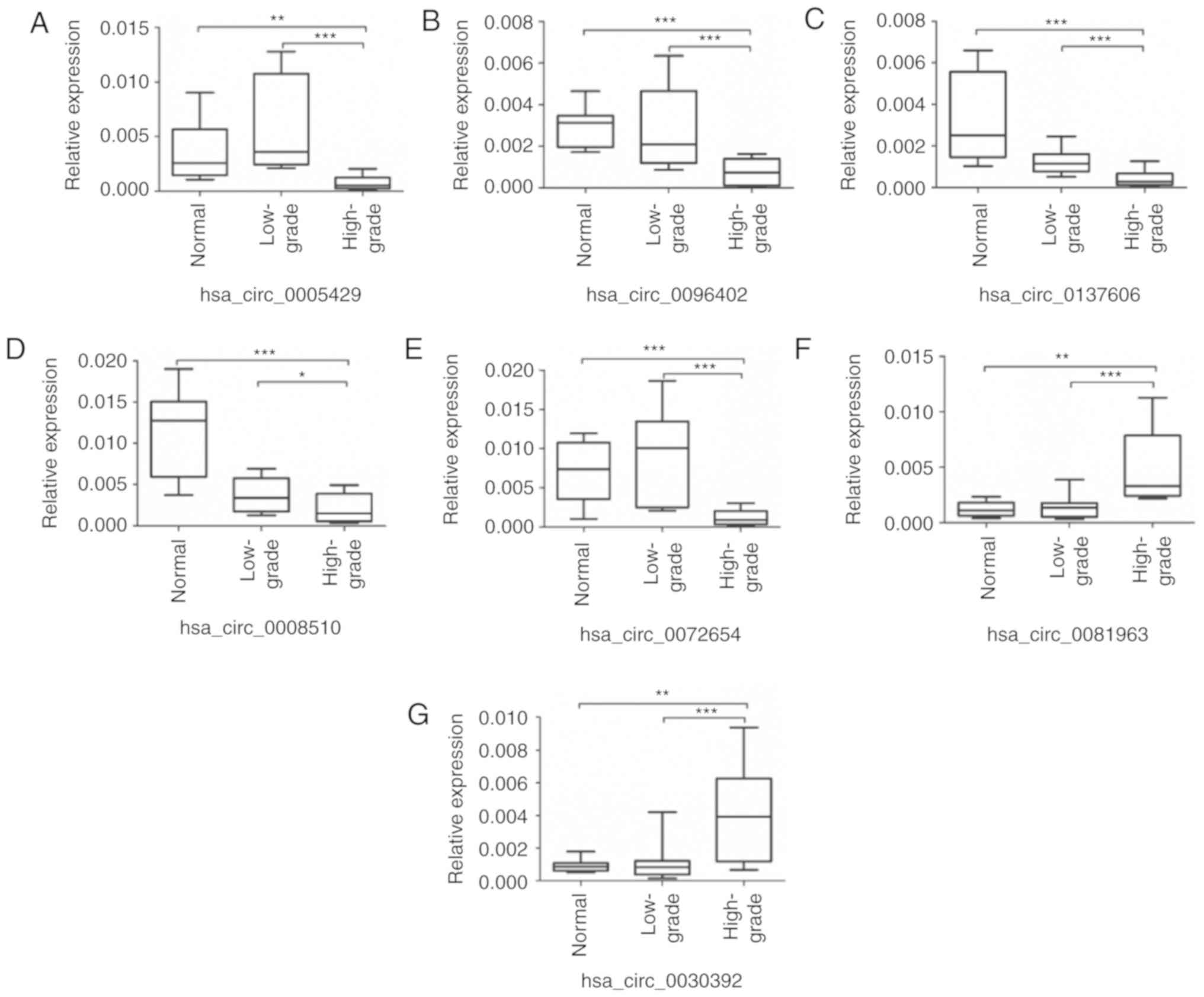
RT-qPCR validation of the differentially
expressed circRNAs
To validate the sequencing data, 10 pairs of tissue
specimens were collected for RT-qPCR analysis. A total of seven
circRNAs were randomly selected for RT-qPCR quantification. As
shown in Fig. 2A-G, the RT-qPCR
results were consistent with the expression profiles of our
high-throughput sequencing data, in which, hsa_circ_0137606
exhibited a high degree of downregulation.
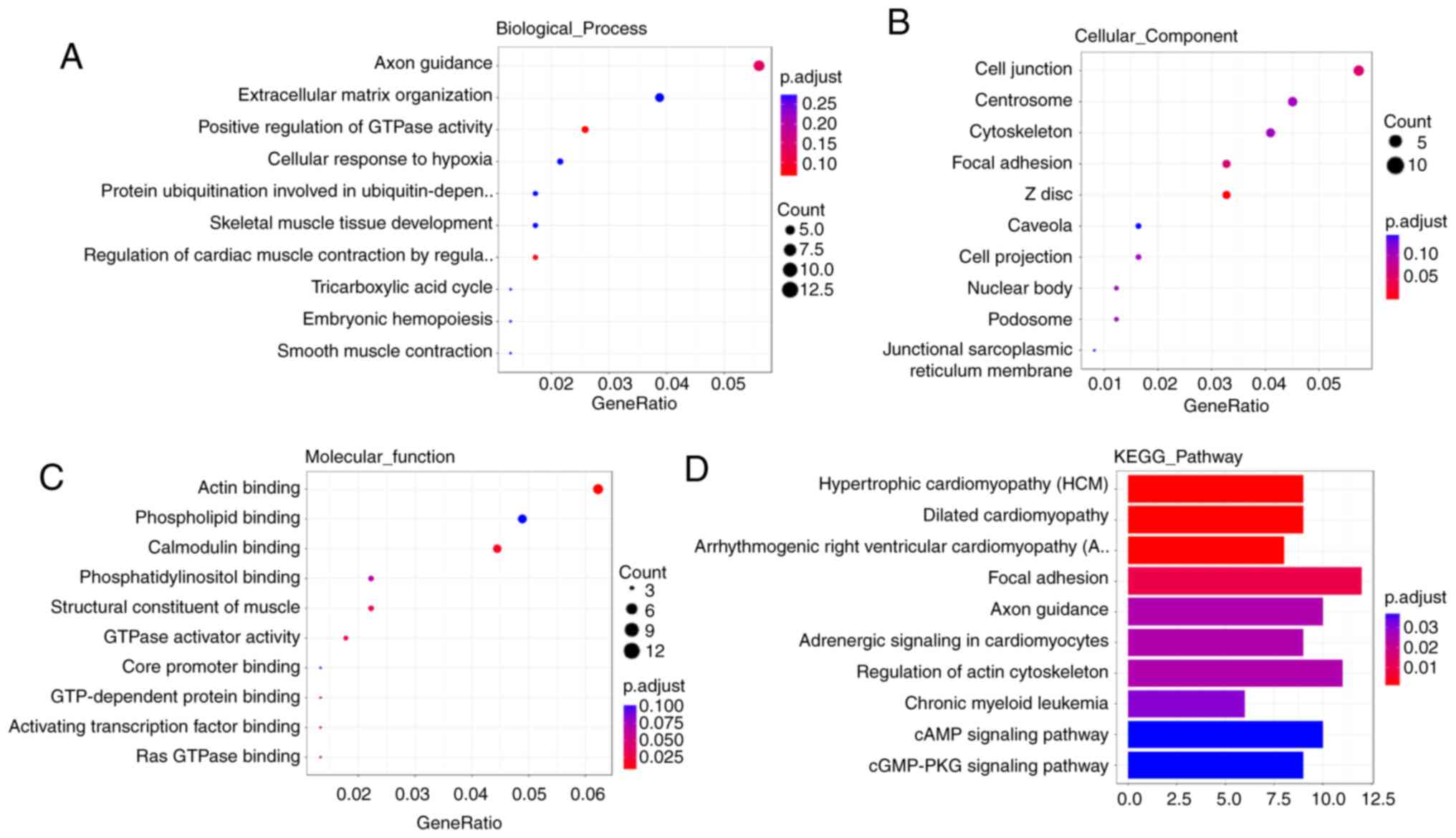
GO and KEGG pathway analyses
Studies have revealed that circRNAs can exert
biological functions by regulating neighboring coding genes
(27-29). GO and KEGG pathway analyses were
performed on dysregulated circRNAs from the two groups to
investigate the mechanisms involved in BC tumorigenesis and
progression.
GO analysis of the H vs. N groups demonstrated that
the dysregulated circRNAs were enriched for 'extracellular matrix
organization' and 'positive GTPase activity' regulation biological
processes, 'cell junction' and 'focal adhesion' cellular
components, and 'GTPase activity' molecular functions. The top 10
significantly enriched GO terms for the two groups are shown in
Figs. 3A-C and S1A-C, whilst
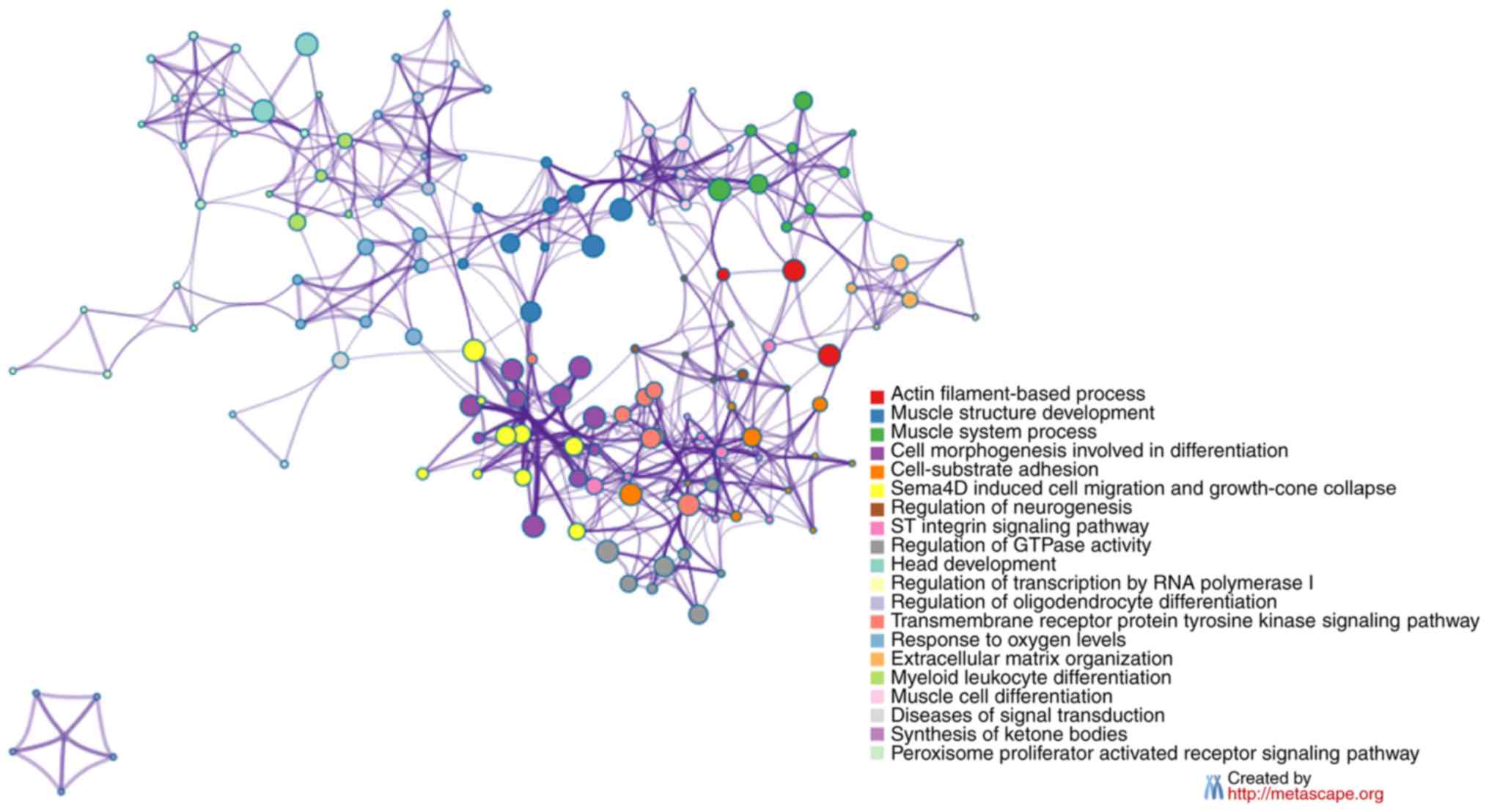
their top 10 enriched KEGG pathways are presented in Figs. 3D and S1D. The enrichment network
of the dysregulated circRNAs was constructed using metascape
(Figs. 4 and S2) (30).
PPI network construction and target gene
prediction
To investigate the potential associations between
circRNAs and BC progression, we constructed a
protein-protein-interaction (PPI) network based on the circRNA
genes dysregulated in both the H vs. N and H vs. L groups. As shown
in Fig. S3, receptor
tyrosine-protein kinase erbB-2 precursor (ERBB2(exhibited the
highest degree of connectivity.
Hsa_circ_0137606 knockdown promotes BC
cell proliferation and metastasis
The expression of hsa_circ_0137606 was validated in
normal uroepithelial cells (SV-HUC1) and different BC cell lines
(Fig. 5A), revealing its
significant downregulation in different BC cells compared with
SV-HUC1 cells. To investigate its biological function in BC, we
knocked down hsa_circ_0137606 in T24 cells. RT-qPCR revealed that
hsa_circ_0137606 expression was significantly decreased in the
hsa_circ_0137606-knockdown group compared with the NC group after
transfection (Fig. 5B). We
performed a CCK-8 assay to explore the effect of hsa_circ_0137606
on cell proliferation; hsa_circ_0137606 knockdown led to
significant increases in BC cell proliferation compared with the NC
group at 72 and 96 h (Fig. 5C).
The Transwell assays revealed that hsa_circ_0137606 knockdown
significantly promoted BC cell migration (Fig. 5D) and invasion (Fig. 5E) compared with the NC group. In
summary, hsa_circ_0137606 knockdown promoted the proliferation and
metastasis of BC cells in vitro.
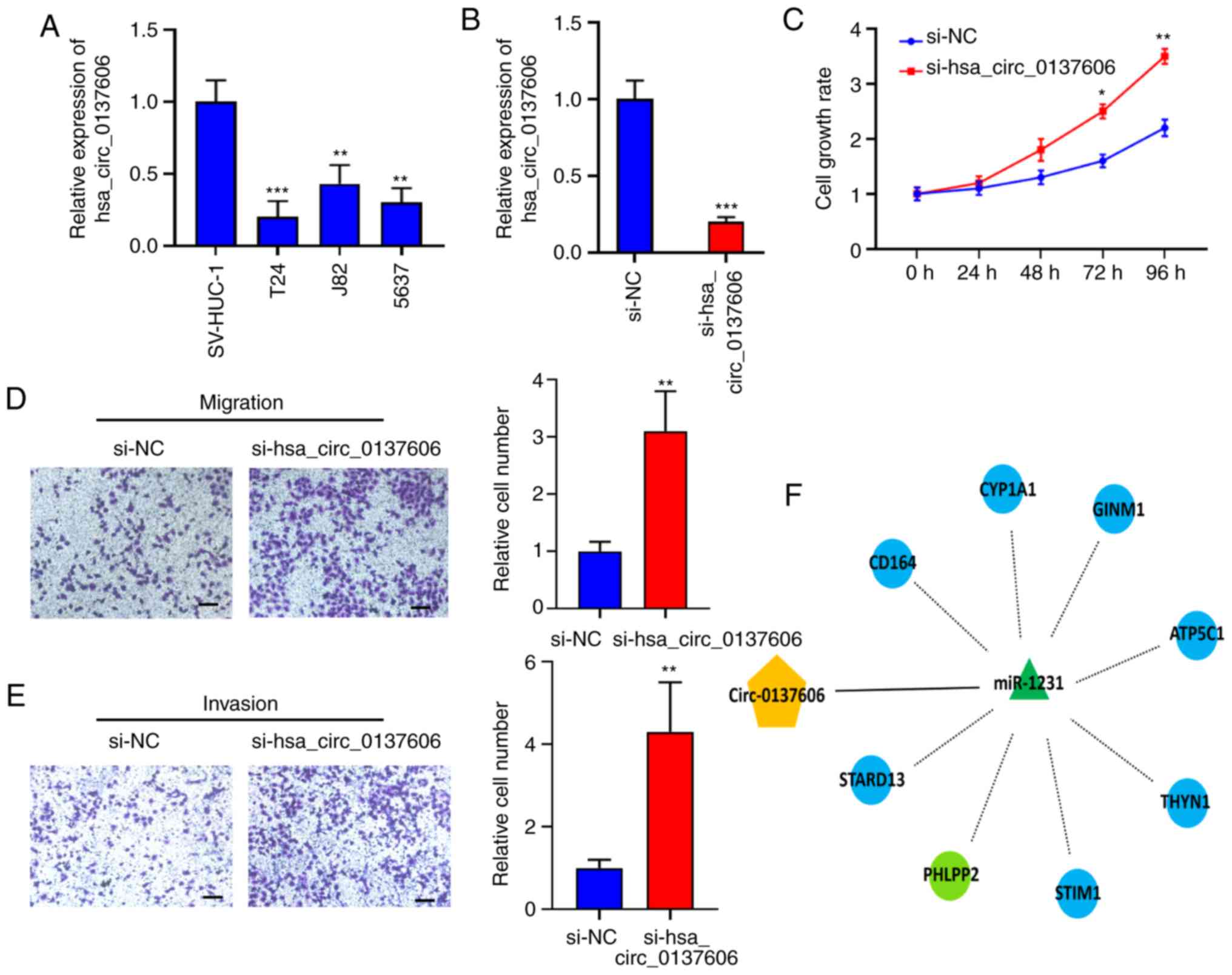 | Figure 5Hsa_circ_0137606 knockdown suppresses
BC cell proliferation, migration and invasion. (A) Hsa_circ_0137606
expression in normal uroepithelial cell and different BC cell lines
detected with reverse transcription-quantitative polymerase chain
reaction. (B) Hsa_circ_0137606 knockdown in T24 cells. (C) Cell
Counting Kit-8 assay of proliferation in the knockdown and control
groups. (D) Transwell assay of migration in the knockdown and
control groups. (E) Transwell assay of invasion in the knockdown
and control groups. Scale bar, 125 μm (F) Candidate target
genes predicted by the Circ-interactome of miR-1231 in bladder
cancer. Data were shown as the mean ± standard deviation,
*P<0.05, **P<0.01 and
***P<0.001 vs. si-NC. Hsa, homo sapiens; NC,
negative control; si, small interfering RNA; circRNA, circular RNA;
ATP5C1, ATP, synthase F1 subunit gamma; CD164, cluster of
differentiation 164; CYP1A1, cytochrome P450 1A1; GINM1,
glycoprotein integral membrane 1; miR, microRNA; PHLPP2, PH domain
and leucine rich repeat protein phosphatase 2; STIM1, stromal
interaction molecule 1; STARD13, StAR related lipid transfer domain
containing 13; THYN1, thymocyte nuclear protein 1. |
Using bioinformatics analysis, we predicted
potential target genes that miR-1231 may bind to and regulate
during BC progression; the miRNA-mRNA interactions were validated
via miRTarBase. As presented in Fig.
5F, the top eight predicted target genes that miR-1231 may
regulate during BC progression were identified.
Hsa_circ_0137606 suppresses BC
proliferation and metastasis by sponging miR-1231
circRNAs can act as miRNA sponges to regulate cells;
therefore, we performed bioinformatics analysis and predicted the
miRNAs that hsa_circ_0137606 could act as a sponge for; miR-1248,
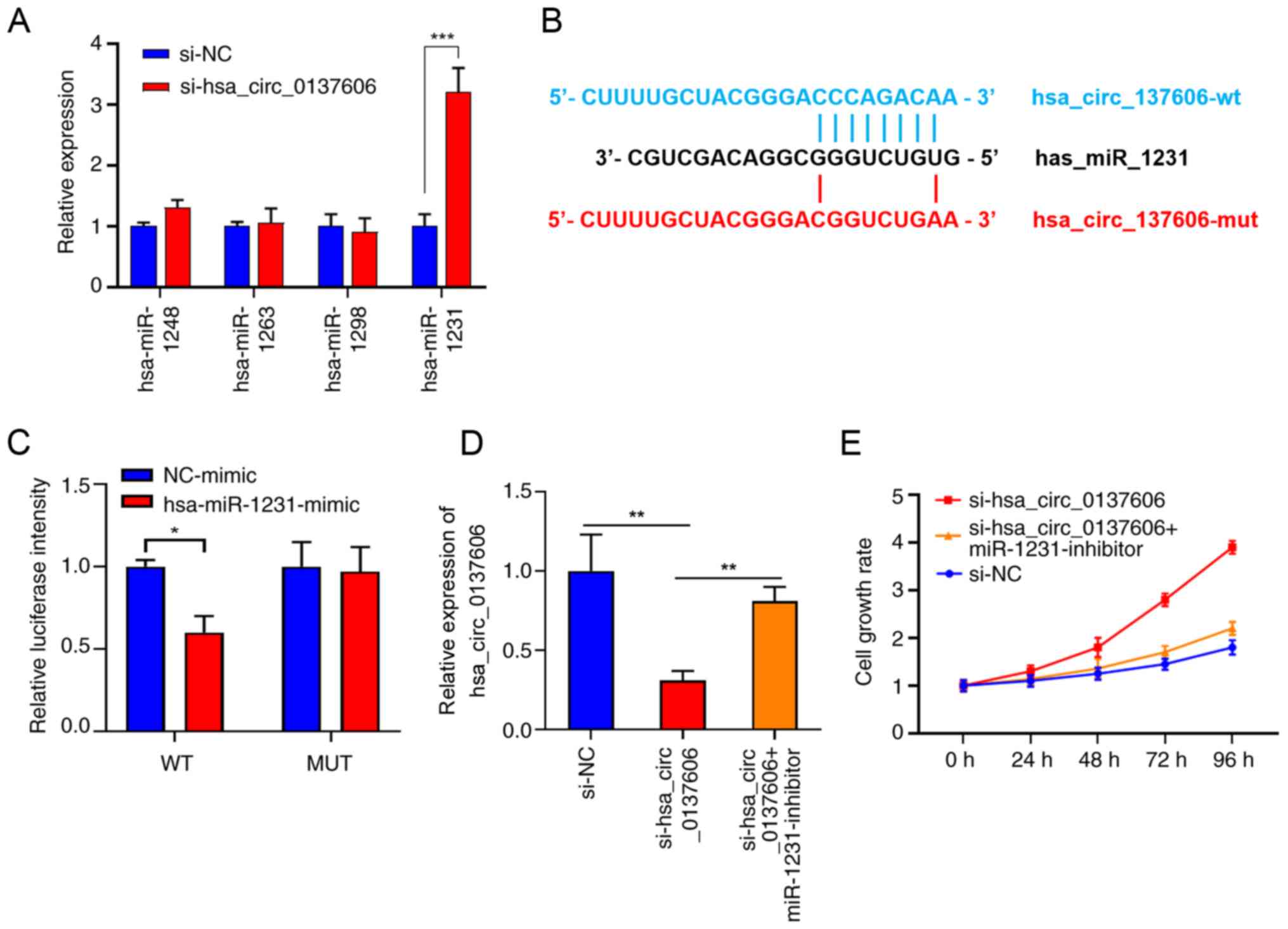
miR-1263, miR-1298 and miR-1231 were the four most likely miRNAs.
To validate this prediction, we determined the expression of the
four miRNAs in the hsa_circ_0137606-knockdown and control groups,
finding that only miR-1231 was significantly increased in the
knockdown group (Fig. 6A).
Luciferase assays were performed to further investigate the role of
miR-1231, revealing that hsa_circ_0137606 could bind to miR-1231 as
its sponge (Fig. 6B and C).
Rescue experiments showed that miR-1231 inhibitor treatment in
hsa_circ_0137606-silenced cells significantly rescued their
proliferation (Fig. 6D and E),
suggesting that hsa_circ_0137606 suppresses BC proliferation and
metastasis by sponging miR-1231.
Discussion
Since the breakthrough in sequencing technology in
2013 (19), numerous circRNAs
have been discovered and many studies have been conducted to
investigate their potential functions in disease, particularly
cancer (31-34). The mechanism of a few circRNAs in
BC tumorigenesis have been revealed (35,36); however, the relationship between
circRNAs and the process by which low-grade BC progresses to
high-grade BC remains unclear. Therefore, we performed
high-throughput sequencing to detect dysregulated circRNAs in
different grades of BC and normal controls.
A total of 316 and 244 dysregulated circRNAs were
discovered in the H vs. N and H vs. L groups, respectively. Among
these, circADAMTS14 has been reported to suppress hepato-cellular
carcinoma progression by competitively combining with microRNA-572
(37). Moreover, Xu et al
(36) found that circPTK2
participates in the proliferation and migration of BC cells. Our
further analysis revealed that 42 circRNAs overlapped between the H
vs. N and H vs. L groups.
GO and KEGG pathway analysis were performed to
investigate the potential molecular mechanisms of these
dysregulated circRNAs. GO analysis revealed that 'GTPase activity
regulation' was significantly enriched. GTPases have been strongly
implicated in cancer (38); a
previous study revealed that dynamin 2 GTPase contributes to the
formation of invadopodia, which play an important role in invasive
BC cells (12). Liu et al
(20) also found that ANXA7
GTPase activity could markedly affect prostate cancer metastasis,
indicating that these dysregulated circRNAs could participate in
the progression of BC by regulating GTPase activity. Additionally,
pathway analysis revealed that 'actin cytoskeleton regulation',
'focal adhesion', and 'cGMP-PKG signaling pathway' were enriched.
Peng et al (39) and
Ohishi et al (40) found
that the actin cytoskeleton serves a crucial role in cancer
invasion. The focal adhesion pathway is also involved in cancer
cell invasion (41), migration
(42), and therapy resistance
(23). Consequently, we
speculated that the dysregulated circRNAs we detected predominantly
participate in BC progression via such pathways.
To further investigate the association between
circRNAs and BC, we constructed a PPI network based on the circRNA
gene symbols dysregulated in both the H vs. N and H vs. L groups.
We found that ERBB2, which serves a key role in the
development of breast cancer (43), had the highest degree of
connectivity. Previous studies have revealed that ERBB2
could be involved in BC (44,45), indicating that the differently
expressed circRNAs may affect BC progression by regulating
ERBB2.
To validate our results, we performed RT-qPCR on
seven circRNAs in independent tissues; the novel circRNA,
hsa_circ_0137606, was selected for further analysis. RT-qPCR was
performed on normal uroepithelial cells and different BC cell lines
to validate its expression, finding it to be significantly
downregulated in BC cells (particularly T24 cells). Functional
in vitro experiments demonstrated that hsa_circ_0137606
could suppress BC cell proliferation, migration and invasion. A
previous study (46) indicated
that circRNAs containing miRNA-binding sites could act as cellular
miRNA sponges. By binding miRNA, they could prevent its inhibitory
effect on target genes and thus indirectly regulate their
expression. By using bioinformatics and RT-qPCR analyses, we
predicted that hsa_circ_0137606 could act as a sponge for miR-1231,
with luciferase reporter assays further validating this finding.
Rescue experiments showed that inhibiting miR-1231 significantly
rescued hsa_circ_0137606 knockdown-induced proliferation, whilst
previous studies have shown that miR-1231 has a role in multiple
malignant tumors (47,48). Furthermore, we used miRTarBase to
predict the eight most likely target genes of miR-1231, one of
which was PH domain and leucine rich repeat protein phosphatase 2
(PHLPP2I) which has been shown to be involved in BC
progression as an miRNA target gene (49,50); thus, PHLPP2 may be a
promising target of miR-1231 in BC. However, some limitations
should be mentioned in this study. First, the number of samples for
sequencing was rather low, more samples should be employed for
future detection to verify our results. Secondly, when the
expression of hsa_circ_0137606 was explored in different BC cell
lines, we did not select a non-bladder cell as control group.
Non-bladder control cells could improve the reliability of the
results. In conclusion, we used high-throughput sequencing to
identify aberrantly expressed circRNAs in different grades of BC.
Using bioinformatics analysis, we found that these dysreg-ulated
circRNAs could synergistically contribute towards BC progression.
Furthermore, we revealed that hsa_circ_0137606, which is
significantly downregulated in BC, could suppress BC proliferation
and metastasis by sponging miR-1231. This study suggests that
hsa_circ_0137606 could be an effective therapeutic target for
BC.
Supplementary Data
Acknowledgments
Not applicable.
Abbreviations:
|
BC
|
bladder cancer
|
|
ceRNAs
|
competitive endogenous RNAs
|
|
PPI
|
protein-protein interaction
|
|
NMIBC
|
non-muscle-invasive bladder cancer
|
|
MIBC
|
muscle-invasive bladder cancer
|
Funding
The present study was financially supported by the
National Natural Science Foundation of China (grant nos. 81772712
and 81702569) and the Natural Science Foundation of Jiangsu
Province (grant no. BK20170151).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WL made substantial contributions to the design of
the present study; YC and JZ acquired the data. ZS and WM performed
the experiments. LY analyzed and interpreted the data; KX and XY
were involved in drafting the manuscript and revising it critically
for important intellectual content.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Nanjing Drum Tower Hospital and the Affiliated
Hospital of Nanjing University Medical School. Written informed
consent was obtained from patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Antoni S, Ferlay J, Soerjomataram I, Znaor
A, Jemal A and Bray F: Bladder cancer incidence and mortality: A
global overview and recent trends. Eur Urol. 71:96–108. 2017.
View Article : Google Scholar
|
|
2
|
Cumberbatch MGK, Jubber I, Black PC,
Esperto F, Figueroa JD, Kamat AM, Kiemeney L, Lotan Y, Pang K,
Silverman DT, et al: Epidemiology of bladder cancer: A systematic
review and contemporary update of risk factors in 2018. Eur Urol.
74:784–795. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kamat AM, Hahn NM, Efstathiou JA, Lerner
SP, Malmström PU, Choi W, Guo CC, Lotan Y and Kassouf W: Bladder
cancer. Lancet. 388:2796–2810. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Soukup V, Čapoun O, Cohen D, Hernández V,
Babjuk M, Burger M, Compérat E, Gontero P, Lam T, MacLennan S, et
al: Prognostic performance and reproducibility of the 1973 and
2004/2016 world health organization grading classification systems
in non-muscle-invasive bladder cancer: A European association of
urology non-muscle invasive bladder cancer guidelines panel
systematic review. Eur Urol. 72:801–813. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Funt SA and Rosenberg JE: Systemic,
perioperative management of muscle-invasive bladder cancer and
future horizons. Nat Rev Clin Oncol. 14:221–234. 2017. View Article : Google Scholar
|
|
6
|
Sargos P, Baumann BC, Eapen L,
Christodouleas J, Bahl A, Murthy V, Efstathiou J, Fonteyne V,
Ballas L, Zaghloul M, et al: Risk factors for loco-regional
recurrence after radical cystectomy of muscle-invasive bladder
cancer: A systematic-review and framework for adjuvant
radiotherapy. Cancer Treat Rev. 70:88–97. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer. J Clin. 66:115–132. 2016.
|
|
8
|
Vicens Q and Westhof E: Biogenesis of
circular RNAs. Cell. 159:13–14. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jeck WR and Sharpless NE: Detecting and
characterizing circular RNAs. Nat Biotechnol. 32:453–461. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li X, Yang L and Chen LL: The biogenesis,
functions, and challenges of circular RNAs. Mol Cell. 71:428–442.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Holdt LM, Stahringer A, Sass K, Pichler G,
Kulak NA, Wilfert W, Kohlmaier A, Herbst A, Northoff BH, Nicolaou
A, et al: Circular non-coding RNA ANRIL modulates ribosomal RNA
maturation and atherosclerosis in humans. Nat Commun. 7:124292016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen F, Chen X, Yang D, Che X, Wang J, Li
X, Zhang Z, Wang Q, Zheng W, Wang L, et al: Isoquercitrin inhibits
bladder cancer progression in vivo and in vitro by regulating the
PI3K/Akt and PKC signaling pathways. Oncol Rep. 36:165–172. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Han D, Li J, Wang H, Su X, Hou J, Gu Y,
Qian C, Lin Y, Liu X, Huang M, et al: Circular RNA circMTO1 acts as
the sponge of microRNA-9 to suppress hepatocellular carcinoma
progression. Hepatology. 66:1151–1164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu J, Xu QG, Wang ZG, Yang Y, Zhang L, Ma
JZ, Sun SH, Yang F and Zhou WP: Circular RNA cSMARCA5 inhibits
growth and metastasis in hepatocellular carcinoma. J Hepatol.
68:1214–1227. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen X, Chen RX, Wei WS, Li YH, Feng ZH,
Tan L, Chen JW, Yuan GJ, Chen SL, Guo SJ, et al: PRMT5 circular RNA
promotes metastasis of urothelial carcinoma of the bladder through
sponging miR-30c to induce epithelial-mesenchymal transition. Clin
Cancer Res. 24:6319–6330. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen Y, Yang F, Fang E, Xiao W, Mei H, Li
H, Li D, Song H, Wang J, Hong M, et al: Circular RNA circAGO2
drives cancer progression through facilitating HuR-repressed
functions of AGO2-miRNA complexes. Cell Death Differ. 26:1346–1364.
2019. View Article : Google Scholar
|
|
18
|
Weng W, Wei Q, Toden S, Yoshida K,
Nagasaka T, Fujiwara T, Cai S, Qin H, Ma Y and Goel A: Circular RNA
ciRS-7-A promising prognostic biomarker and a potential therapeutic
target in colorectal cancer. Clin Cancer Res. 23:3918–3928. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH and
Munschauer M: et al: Circular RNAs are a large class of animal RNAs
with regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu S, Li X, Lin Z, Su L, Yan S, Zhao B
and Miao J: SEC-induced activation of ANXA7 GTPase suppresses
prostate cancer metastasis. Cancer Lett. 416:11–23. 2018.
View Article : Google Scholar :
|
|
21
|
Li Y, Zheng F, Xiao X, Xie F, Tao D, Huang
C, Liu D, Wang M, Wang L, Zeng F and Jiang G: CircHIPK3 sponges
miR-558 to suppress heparanase expression in bladder cancer cells.
EMBO Rep. 18:1646–1659. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li M, Liu Y, Zhang X, Liu J and Wang P:
Transcriptomic analysis of high-throughput sequencing about
circRNA, lncRNA and mRNA in bladder cancer. Gene. 677:189–197.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Eke I and Cordes N: Focal adhesion
signaling and therapy resistance in cancer. Semin Cancer Biol.
31:65–75. 2015. View Article : Google Scholar
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
25
|
Yu G, Wang LG, Han Y and He QY:
clusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chou CH, Shrestha S, Yang CD, Chang NW,
Lin YL, Liao KW, Huang WC, Sun TH, Tu SJ, Lee WH, et al: miRTarBase
update 2018: A resource for experimentally validated
microRNA-target interactions. Nucleic Acids Res. 46(D1): pp.
D296–D302. 2018, View Article : Google Scholar :
|
|
27
|
Yao Y, Chen X, Yang H, Chen W, Qian Y, Yan
Z, Liao T, Yao W, Wu W, Yu T, et al: Hsa_circ_0058124 promotes
papillary thyroid cancer tumorigenesis and invasiveness through the
NOTCH3/GATAD2A axis. J Exp Clin Cancer Res. 38:3182019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu K, Liao X, Gong Y, He J, Zhou JK, Tan
S, Pu W, Huang C, Wei YQ and Peng Y: Circular RNA F-circSR derived
from SLC34A2-ROS1 fusion gene promotes cell migration in non-small
cell lung cancer. Mol Cancer. 18:982019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang L, Long H, Zheng Q, Bo X, Xiao X and
Li B: Circular RNA circRHOT1 promotes hepatocellular carcinoma
progression by initiation of NR2F6 expression. Mol Cancer.
18:1192019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Soonthornvacharin S, Rodriguez-Frandsen A,
Zhou Y, Galvez F, Huffmaster NJ, Tripathi S, Balasubramaniam VR,
Inoue A, de Castro E, Moulton H, et al: Systems-based analysis of
RIG-I-dependent signalling identifies KHSRP as an inhibitor of
RIG-I receptor activation. Nat Microbiol. 2:170222017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kristensen LS, Hansen TB, Veno MT and
Kjems J: Circular RNAs in cancer: Opportunities and challenges in
the field. Oncogene. 37:555–565. 2018. View Article : Google Scholar :
|
|
32
|
Liang WC, Wong CW, Liang PP, Shi M, Cao Y,
Rao ST, Tsui SK, Waye MM, Zhang Q, Fu WM and Zhang JF: Translation
of the circular RNA circβ-catenin promotes liver cancer cell growth
through activation of the Wnt pathway. Genome Biol. 20:842019.
View Article : Google Scholar
|
|
33
|
Braicu C, Zimta AA, Gulei D, Olariu A and
Berindan-Neagoe I: Comprehensive analysis of circular RNAs in
pathological states: Biogenesis, cellular regulation, and
therapeutic relevance. Cell Mol Life Sci. 76:1559–1577. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Vo JN, Cieslik M, Zhang Y, Shukla S, Xiao
L, Zhang Y, Wu YM, Dhanasekaran SM, et al: The landscape of
circular RNA in cancer. Cell. 176:869–881.e13. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xie F, Li Y, Wang M, Huang C, Tao D, Zheng
F, Zhang H, Zeng F, Xiao X and Jiang G: Circular RNA BCRC-3
suppresses bladder cancer proliferation through miR-182-5p/p27
axis. Mol Cancer. 17:1442018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xu ZQ, Yang MG, Liu HJ and Su CQ: Circular
RNA hsa_ circ_0003221 (circPTK2) promotes the proliferation and
migration of bladder cancer cells. J Cell Biochem. 119:3317–3325.
2018. View Article : Google Scholar
|
|
37
|
Song C, Li D, Liu H, Sun H, Liu Z, Zhang L
and Hu Y: The competing endogenous circular RNA ADAMTS14 suppressed
hepatocellular carcinoma progression through regulating
microRNA-572/regulator of calcineurin 1. J Cell Physiol.
234:2460–2470. 2019. View Article : Google Scholar
|
|
38
|
Subramani D and Alahari SK:
Integrin-mediated function of Rab GTPases in cancer progression.
Mol Cancer. 9:3122010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Peng JM, Bera R, Chiou CY, Yu MC, Chen TC,
Chen CW, Wang TR, Chiang WL, Chai SP and Wei Y: et al: Actin
cytoskeleton remodeling drives epithelial-mesenchymal transition
for hepatoma invasion and metastasis in mice. Hepatology.
67:2226–2243. 2018. View Article : Google Scholar
|
|
40
|
Ohishi T, Yoshida H, Katori M, Migita T,
Muramatsu Y, Miyake M, Ishikawa Y, Saiura A, Iemura SI, Natsume T
and Seimiya H: Tankyrase-binding protein TNKS1BP1 regulates actin
cytoskeleton rearrangement and cancer cell invasion. Cancer Res.
77:2328–2338. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Schlienger S, Ramirez RA and Claing A:
ARF1 regulates adhesion of MDA-MB-231 invasive breast cancer cells
through formation of focal adhesions. Cell Signal. 27:403–415.
2015. View Article : Google Scholar
|
|
42
|
Meng F, Saxena S, Liu Y, Joshi B, Wong TH,
Shankar J, Foster LJ, Bernatchez P and Nabi IR: The
phospho-caveolin-1 scaffolding domain dampens force fluctuations in
focal adhesions and promotes cancer cell migration. Mol Biol Cell.
28:2190–2201. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Petry IB, Fieber E, Schmidt M, Gehrmann M,
Gebhard S, Hermes M, Schormann W, Selinski S, Freis E and Schwender
H: et al: ERBB2 induces an antiapoptotic expression pattern of
Bcl-2 family members in node-negative breast cancer. Clin Cancer
Res. 16:451–460. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Junttila TT, Laato M, Vahlberg T,
Söderström KO, Visakorpi T, Isola J and Elenius K: Identification
of patients with transitional cell carcinoma of the bladder
overexpressing ErbB2, ErbB3, or specific ErbB4 isoforms: Real-time
reverse transcription-PCR analysis in estimation of ErbB receptor
status from cancer patients. Clin Cancer Res. 9:5346–5357.
2003.PubMed/NCBI
|
|
45
|
Groenendijk FH, de Jong J, Fransen van de
Putte EE, Michaut M, Schlicker A, Peters D, Velds A, Nieuwland M,
van den Heuvel MM, Kerkhoven RM, et al: ERBB2 mutations
characterize a subgroup of muscle-invasive bladder cancers with
excellent response to neoadjuvant chemotherapy. Eur Urol.
69:384–388. 2016. View Article : Google Scholar
|
|
46
|
Liu G, Huang K, Jie Z, Wu Y, Chen J, Chen
Z, Fang X and Shen S: CircFAT1 sponges miR-375 to promote the
expression of Yes-associated protein 1 in osteosarcoma cells. Mol
Cancer. 17:1702018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhang J, Zhang J, Qiu W, Zhang J, Li Y,
Kong E, Lu A, Xu J and Lu X: MicroRNA-1231 exerts a tumor
suppressor role through regulating the EGFR/PI3K/AKT axis in
glioma. J Neurooncol. 139:547–562. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhou C, Yu Q, Chen L, Wang J, Zheng S and
Zhang J: A miR-1231 binding site polymorphism in the 3′UTR of
IFNAR1 is associated with hepatocellular carcinoma susceptibility.
Gene. 507:95–98. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Mao XP, Zhang LS, Huang B, Zhou SY, Liao
J, Chen LW, Qiu SP and Chen JX: Mir-135a enhances cellular
proliferation through post-transcriptionally regulating PHLPP2 and
FOXO1 in human bladder cancer. J Transl Med. 13:862015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Huang C, Liao X, Jin H, Xie F, Zheng F, Li
J, Zhou C, Jiang G, Wu XR and Huang C: MEG3, as a competing
endogenous RNA, binds with miR-27a to promote PHLPP2 protein
translation and impairs bladder cancer invasion. Mol Ther Nucleic
Acids. 16:51–62. 2019. View Article : Google Scholar : PubMed/NCBI
|