Introduction
Non-alcoholic fatty liver disease (NAFLD) refers to
the excessive accumulation of liver fat according to a histological
analysis identifying fatty degeneration in >5% of hepatocytes or
a fat fraction density >5.6%, as evaluated by proton magnetic
resonance spectroscopy (1H-MRS) or quantitative fat/water selective
magnetic resonance imaging (MRI), that leads to subsequent liver
injury, balloon-like degeneration, and chronic inflammatory
infiltration with or without fibrosis and without excessive alcohol
intake (male, <30 g/day, female <20 g/day) or other known
causes. NAFLD includes non-alcoholic fatty liver (NAFL) and
non-alcoholic steatohepatitis (NASH) (1,2)
and has become an epidemic chronic liver disease worldwide
(3). However, no specific
therapeutic drugs have been developed following decades of
research. The reason for this may be related to several factors,
such as the long latency of NAFLD, an incomplete understanding of
the pathogenesis of NAFLD (4,5),
and the involvement of multiple factors. Further exploration of the
molecular mechanisms of NAFLD will contribute to the future
development of drugs for the treatment of NAFLD.
The WNT signaling pathway has been classified into
the classical and non-classical pathways according to whether the
accumulation of β-catenin in the nucleus is necessary or not. The
classical WNT/β-catenin signaling pathway has been recognized as a
key regulator of adipose differentiation and exerts anti-lipid
formation and anti-inflammatory effects. By contrast, the
non-classical WNT signaling pathway promotes fat formation, lipid
accumulation and inflammation. An imbalance in the WNT signaling
pathways has been closely associated with NAFLD. When the classical
WNT/β-catenin signaling pathway is activated, the expression of
CCAAT enhancer binding protein α (C/EBP-α) and peroxisome
prolif-erator activated receptor γ (PPAR-γ) is inhibited by
β-catenin, which in turn inhibits the differentiation of
preadipocytes (Fig. 1) (4). A mutation in low-density
lipoprotein-related receptor 6 (LRP6; LRP6R611C), a
common coreceptor of the classical WNT/β-catenin signaling pathway,
induces lipid accumulation in the liver through the nutritional
sensory pathway [insulin-like growth factor 1 (IGF1)/AKT/mammalian
target of rapamycin (mTOR)/sterol regulatory element-binding
transcription factor (SREBP)1/2] (Fig. 1). Homozygotic LRP6R611C
(LRP6mut/mut) mice have been shown to exhibit NASH with
fibrosis (6,7). Wnt3a is a canonical Wnt ligand
(4). As an inhibitor of
inflammatory processes, the Wnt3a inhibition of the non-canonical
Wnt pathway reduces lipid accumulation and inflammation, but has
almost no effect in enhancing the canonical Wnt pathway (7). Along with reducing the influence of
the risk factors of NAFLD, it is reasonable to believe that
inhibiting the classical WNT/β-catenin regulatory signaling pathway
is indispensable in promoting the pathogenesis of NAFLD, and this
may be achieved by upregulating PPAR-γ and SREBP1c.
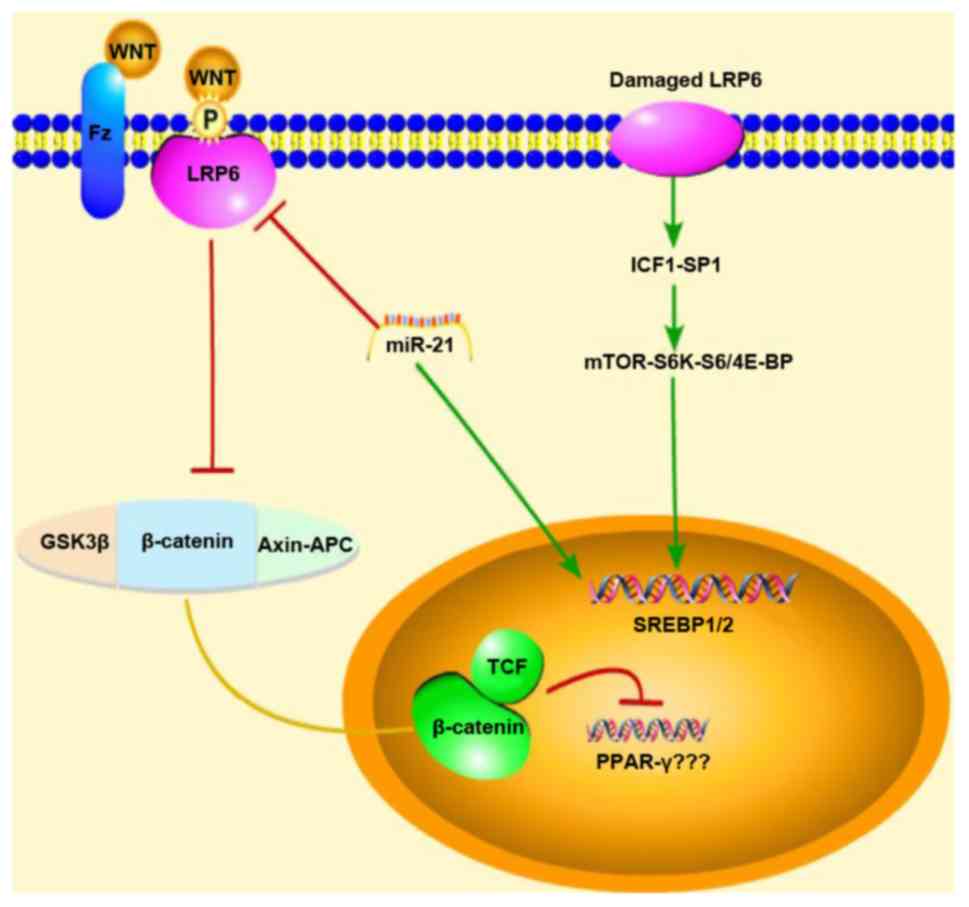 | Figure 1Schematic diagram of the mechanisms
of action of miR-21 showing the regulation of the WNT
signaling pathway in NAFLD mice. Red arrows represent inhibition,
green arrows represent promotion, and the yellow line represents
migration from the cytoplasm into the nucleus When the classical
WNT/β-catenin signal is activated, the expression of PPAR-γ is
inhibited; however, our experimental results were reversed, and
thus, we do not know association between PPAR-γ and the
WNT/β-catenin signaling pathway. Thus, due to this uncertainty,
question marks (???) were added next to PPAR-γ in the figure.
NAFLD, non-alcoholic fatty liver disease; GSK3β, glycogen synthase
kinase-3β; LRP6, low-density lipoprotein-related receptor 6;
SREBP1/2, sterol regulatory element-binding transcription factor
1/2; IGF1, insulin-like growth factor 1; mTOR, mammalian target of
rapamycin. |
MicroRNAs (miRNAs or miRs) are a class of
endogenous, single-stranded, non-coding small RNAs with a length of
~19-25 nucleotides that regulate gene expression by inhibiting
translation, promoting the cleavage of mRNAs or targeting promoter
regions (8). miR-21 was
one of the earliest discovered human miRNAs. In the pathogenesis of
NAFLD, miR-21 has been shown to participate in liver lipid
metabolism through a variety of targets, including fatty acid
binding protein 7 (FABP7) and 3-hydroxy-3-methylglutaryl coenzyme A
reductase (HMGCR), and to contribute to NASH via peroxi-some
proliferator activated receptor α (PPAR-α). It can also
participate in NAFLD through SMAD7, phosphate and tension homolog
(PTEN), HMG-box transcription factor 1D (HBP1D) and other targets
(8-13). Moreover, our research group
previously demonstrated that the expression of LRP6 was inhibited
in HepG2 cells transfected with a miR-21 mimic and that LRP6
was a target of miR-21. We also observed that the synthesis
of triglycerides was decreased through the inhibition of
miR-21 through RNA interference, suggesting that
miR-21 may be involved in lipid synthesis and metabolism by
interacting with the WNT/β-catenin signaling pathway to further
participate in the pathogenesis and pathology of NAFLD (14). This was the first time, to the
best of our knowledge, that miR-21 was linked to the
WNT/β-catenin signaling pathway. Therefore, in order to explore the
regulatory role of miR-21 in the WNT/β-catenin signaling
pathway in NAFLD in mice, the objective of this study was to
explore the role of the molecular regulatory network of
miR-21 in the pathogenesis of NAFLD and to elucidate the
pathological mechanisms underlying NAFLD.
Materials and methods
Mouse model
All mice were male C57BL/6J mice (6 weeks
old, n=15, 22.60±1.24 g, purchased from Chengdu Dashuo Laboratory
Animal Co., Ltd.; http://www.cd-dossy.cn/) that were bred at the
Laboratory Animal Center of Southwest Medical University
(http://dwzx.swmu.edu.cn/) and allowed to
acclimatize to their environment for 1 week. All animals received
care according to the guidelines of the Institutional Animal Care
and Use Committee of Southwest Medical University (Luzhou, China),
and the experiment was approved by the Experimental Animal Ethics
Committee of Southwest Medical University (application acceptance
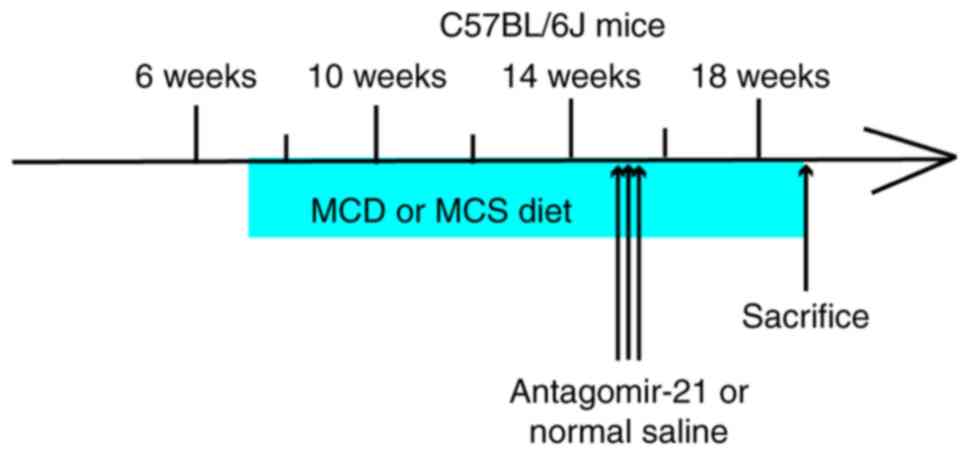
no. 20180521-11). C57BL/6J mice (n=9) were fed a methionine- and
choline-deficient diet (MCD, Trophic Animal Feed High-Tech Co.,
Ltd., http://www.trophic.cn/) to establish
NAFLD; after 4 weeks, 3 mice (15.03±0.75 g) without manifestations
of peritonitis were sacrificed by cervical dislocation following an
intraperitoneal injection of 10% chloral hydrate (400 mg/kg), and
the liver tissues were removed for hematoxylin & eosin
(H&E) staining to confirm the successful establishment of the
model. Subsequently, the remaining mice were divided into 2 groups
of 3 mice in each. Antagomir-21 (antagomir-21 group, n=3, 8 mg/kg
5-UCA ACA CUG UCU GUA GAU CUA-3 (10), purchased from Shanghai Genepharma
Pharmaceutical Technology Co. Ltd., http://www.genepharma.bioon.com.cn/) or the same dose
of saline (control group, n=3), was injected through the tail vein
at 15 weeks of age once a day for 3 consecutive days. The C57BL/6J
wild-type mice (normal group, n=6) were fed a methionine- and
choline-sufficient diet (MCS, Trophic Animal Feed High-Tech Co.
Ltd., http://www.trophic.cn/). After 4 weeks,
3 mice (26.7±0.76 g) without manifestations of peritonitis were
randomly selected, anesthetized and sacrificed (using the same
method as described above). The liver tissues were removed for
H&E staining as a NAFLD control. At 15 weeks of age, the same
dose of saline was injected into the tail vein once a day for 3
consecutive days (Fig. 2).
All mice without manifestations of peritonitis
(antagomir-21 group, 12.63±0.72 g; control group, 13.4±0.69 g;
normal group, 30.13±5.51 g) were anesthetized with 10% chloral
hydrate (400 mg/kg) at the age of 19 weeks. Once the mice no longer
exhibited a corneal reflex or pain responses, blood (~0.5 ml) was
collected from the eyeball and the heart through a needle inserted
obliquely into the heart at a 45° angle (in the location with the
most obvious heartbeat) to obtain blood. After collecting the
blood, the mice died within 1 min from continuous cardiac arrest.
The collected blood was allowed to settle for 30 min and
centrifuged for 20 min at 2,500 x g at 4°C. The supernatant was
collected following centrifugation and stored at -80°C for blood
lipid and aminotransferase detection. The liver was separated
quickly and weighed.
Approximately 400 mg of liver tissue were frozen and
stored at -80°C for PCR and western blot analysis. Approximately
300 mg of liver tissue were fixed in 4% formaldehyde for H&E
staining and immunohistochemical analysis.
Biochemical analysis
The levels of alanine aminotransferase (ALT),
aspartate aminotransferase (AST), triglycerides (TG), total
cholesterol (TC) and low-density lipoprotein (LDL) were detected
with the DR-200Bs enzyme labeling instrument (Diatek Co.,
http://www.diateklab.com/) according to the
manufacturer's instructions (Nanjing Jiancheng Bioengineering
Institute., http://www.njjcbio.com/).
Histological analysis
Paraffin-embedded liver sections (3-µm-thick)
were stained with H&E (sent to the Department of Pathology,
Affiliated Hospital of Southwest Medical University). The
H&E-stained sections were observed under an optical microscope
(Olympus, http://www.olympus.com.cn/). The NAS
scores of the H&E-stained sections were determined by a
pathologist who was blinded to the mouse groupings and according to
the following criteria (15): i)
Steatosis: 0-3 points, <5, 5-33, 33-66 and ≥66%; ii)
intralobular inflammation (counting of necrotic foci at x20
magnification): 0-3 points, none, <2, 2-4, ≥4; and iii)
hepatocyte ballooning degeneration change: 0-2 points, none, rare,
more common. NASH was excluded by a score of 0-2; NASH could be
considered based on a score of 3-4, and NASH would likely be
diagnosed based on a score of 5-8.
Western blot analysis
The liver tissues were rinsed 2-3 times with
precooled PBS buffer to remove blood, cut into small sections, and
placed in a homogenizer (Wuhan Aspen Biotechnology Co., Ltd.). A
10-fold volume of tissue protein extraction reagent (Wuhan Aspen
Biotechnology Co., Ltd.) with protease inhibitor (Wuhan Aspen
Biotechnology Co., Ltd.) was added and thoroughly homogenized, and
the homogenate was incubated in an ice bath for 30 min. The
supernatant protein concentration was determined by a BCA protein
concentration assay kit (Wuhan Aspen Biotechnology Co., Ltd.)
following centrifugation at 4°C and 13,000 x g for 5 min. The
sample size was determined according to the sample concentration,
and the total protein in each sample was 40 µg. The protein
samples were electrophoretically transferred onto PVDF membranes
(Shanghai Millipore Filter Material Co., Ltd., http://millipore.org.cn/search/) after adding an
appropriate amount of 5X protein sample buffer (Wuhan Aspen
Biotechnology Co., Ltd.) and boiling the samples in a water bath at
95-100°C for 5 min, in which protein samples were separated using
8-10% SDS-PAGE and the percentage of Tween-20 in TBST was 0.1%. The
PVDF membranes were activated with methanol prior to use. The
membranes were sealed and incubated at room temperature for 1 h.
The sealing fluid was removed, and the membranes were incubated
with primary antibody overnight at 4°C. The diluted primary
antibody (Table I) was recovered,
and the membranes were washed 3 times with TBST for 5 min each
time. The diluted secondary antibody (Table II) was added, and the membranes
were incubated at room temperature for 30 min. The membranes were
washed in TBST 4 times on a shaker at room temperature for 5 min
each time. Freshly mixed ECL solution (Wuhan Aspen Biotechnology
Co., Ltd.) was added onto the protein side of the membranes, and
the membranes were exposed in a darkroom. The exposure conditions
were adjusted to achieve different light intensities prior to
developing the film. The photographic films (Eastman Kodak,
http://www.kodak.com.cn/) were scanned into
files, and the AlphaEaseFC software processing system (Alpha
Innotech Corporation, http://www.alphainnotech.com) was used to analyze the
optical density of the target bands.
 | Table IDetails of primary antibodies used in
western blot analysis. |
Table I
Details of primary antibodies used in
western blot analysis.
| Name of first
antibody | Origin | Manufacturer | Cat. no. | Dilution
method | Dilution ratio |
|---|
| GAPDH | Rabbit | Abcam | ab37168 | 5% evaporated
milk | 1:10,000 |
| LRP6 | Rabbit | Abcam | ab134146 | 5% evaporated
milk | 1:500 |
| GSK3β | Mouse | Abcam | ab93926 | 5% evaporated
milk | 1:1,000 |
| p-β-catenin | Rabbit | Cell Signaling
Technology | #4176 | 5% BSA | 1:1,000 |
| β-catenin | Rabbit | Abcam | ab32572 | 5% evaporated
milk | 1:3,000 |
| PPAR-γ | Rabbit | Abcam | ab209350 | 5% evaporated
milk | 1:500 |
| Table IIDetails of secondary antibodies used
in western blot analysis. |
Table II
Details of secondary antibodies used
in western blot analysis.
| Name of secondary
antibody | Manufacturer | Cat. no. | Dilution
method | Dilution ratio |
|---|
| HRP-goat anti
rabbit | KPL | 074-1506 | 5% evaporated
milk | 1:10,000 |
| HRP-goat anti
mouse | KPL | 074-1806 | 5% evaporated
milk | 1:10,000 |
RNA isolation and RT-qPCR analysis
Total RNA was extracted from the liver tissues using
a TRIzol reagent kit (Invitrogen™, Semerfly Technology Co., Ltd.,
https://www.thermofisher.com/),
quantified by a SYBR® Premix Ex Taq™ kit (Takara
Biomedical Technology Co., Ltd., http://www.takara.com.cn/) according to the
manufacturer's instructions and stored at -80°C. The first-strand
cDNA of miR-21 was synthesized using the M-MLV reverse
transcriptase kit (Invitrogen™, Semerfly Technology Co., Ltd.,
https://www.thermofisher.com/) according
to the manufacturer's protocol. The synthesis of the first-strand
cDNA for other genes, such as SREBP1c, fatty acid synthase
(FAS), adenosine 5-monophosphate (AMP)-activated protein
kinase α (AMPKα) and carnitine palmitoyl transferase 1α
(CPT1α), was performed using the PrimeScriptTM RT Reagent
kit with gDNA Eraser (Takara Biomedical Technology Co., Ltd.,
http://www.takara.com.cn/) according to the
manufacturer's protocol. The sequences of the primers are presented
in Table III. qPCR was
performed on a Life Technologies StepOneTM Real-Time PCR
instrument, and each sample was assayed by using 3 replicate wells
with the SYBR® Premix Ex TaqTM kit (Takara Biomedical
Technology Co., Ltd., http://www.takara.com.cn/). The reaction procedure was
as follows: Predenaturation, 95°C for 1 min; 40 cycles of 95°C, 15
sec → 58°C, 20 sec → 72°C, 45 sec; melting curve, 60°C → 95°C, 20
sec per 1°C of temperature. The reaction conditions were as
follows: 2X qPCR Mix, 5.0 µl; primer working dilution (2.5
µM), 1.0 µl; template, 1.0 µl;
ddH2O, 2.8 µl; Rox, 0.2 µl. The
corresponding gene expression and relative mRNA expression were
evaluated by the 2-ΔΔCq method and the determination of
the geometric mean (16).
| Table IIISequences of mouse primers used for
RT-qPCR. |
Table III
Sequences of mouse primers used for
RT-qPCR.
| Primer name | Primer
sequence | Product length
(bp) |
|---|
| M-GAPDH | | |
| Forward |
5′-TGAAGGGTGGAGCCAAAAG-3′ | 227 |
| Reverse |
5′-AGTCTTCTGGGTGGCAGTGAT-3′ | |
| M-SREBP-1c | | |
| Forward |
5′-ACAGACAAACTGCCCATCCA-3′ | 223 |
| Reverse |
5′-GCAAGAAGCGGATGTAGTCG-3′ | |
| M-FAS | | |
| Forward |
5′-ATCTGGGCTGTCCTGCCTCT-3′ | 116 |
| Reverse |
5′-TTATCAGTTTCACGAACCCGC-3′ | |
| M-AMPKα | | |
| Forward |
5′-GATGATGACCATGTGCCAACTC-3′ | 270 |
| Reverse |
5′-CTCCGAACACTCGAACTTCTCAC-3′ | |
| M-CPT1α | | |
| Forward |
5′-CATGATTGCAAAGATCAATCGG-3′ | 141 |
| Reverse |
5′-AGCACCTTCAGCGAGTAGCG-3′ | |
| U6 | | |
| RT-primer |
5′-AACGCTTCACGAATTTGCGT-3′ | |
| Forward |
5′-CTCGCTTCGGCAGCACAT-3′ | |
| Reverse |
5′-AACGCTTCACGAATTTGCGT-3′ | |
|
mmu-miR-21 | | |
|
RT-primer |
5'-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTCAACATC-3′ | |
| Forward |
5′-ACGGCTTATCAGACTGATGTTGA-3′ | |
| Reverse |
5′-CTCAACTGGTGTCGTGGAGTC-3′ | |
Immunohistochemistry
Paraffin-embedded sections were placed in a 65°C
oven for 2 h, dewaxed in water, and washed 3 times with PBS (Wuhan
Aspen Biotechnology Co., Ltd.) for 5 min each time. The slices were
placed in EDTA buffer (Wuhan Aspen Biotechnology Co., Ltd. China)
for microwave antigen retrieval, and the microwave was set at
medium power until a boil was reached, after which it was set at
low power until a boil was reached at intervals of 10 min. After
allowing them to cool, the cells were washed with PBS 3 times for 5
min each time. The sections were placed in 3% hydrogen peroxide
solution (Sinopharm Chemical Reagent Co., Ltd., https://www.sinoreagent.com/) and incubated for 10 min
at room temperature in the dark. The sections were washed 3 times
with PBS for 5 min each time and incubated in 5% BSA (Shanghai
Roche Pharmaceutical Co., Ltd., https://www.roche.com/) for 20 min after drying. The
BSA solution was removed, and ~50 µl of diluted primary
antibody (Table IV) was added to
each section to cover the tissue, which was incubated at 4°C
overnight. After washing with PBS 3 times for 5 min each time,
50-100 µl of secondary antibody for the corresponding
species (Table V) was added to
each section, which was incubated for 50 min at 37°C. The sections
were then washed with PBS 3 times for 5 min each time. After
removing the PBS solution, 50-100 µl of freshly prepared DAB
solution (Beijing Zhongshang Jinqiao Biotechnology Co., Ltd.,
http://www.zsbio.com/) was added to each section,
and the color development was controlled under a microscope
(OLYMPUS, Japan, https://www.olympusglobal.com). After the color was
completely developed, the sections were rinsed with distilled water
or tap water, counterstained with hematoxylin (Wuhan Aspen
Biotechnology Co., Ltd. China) differentiated with 1% hydrochloric
acid in alcohol (approximately 1 sec), rinsed with tap water until
the ammonia water turned blue, and then rinsed with water again.
The sections were dehydrated with an alcohol gradient (Sinopharm
Chemical Reagent Co., Ltd., https://www.sinoreagent.com/) consisting of 75, 90 and
100%, alcohol (10 min each time) and dried, after which they were
treated with transparent xylene (Sinopharm Chemical Reagent Co.,
Ltd., https://www.sinoreagent.com/) and
neutral gum (Sinopharm Chemical Reagent Co., Ltd., https://www.sinoreagent.com/) and sealed. The images
were magnified at x400 under a microscope (Olympus, https://www.olympus-global.com).
| Table IVDetails of the primary antibodies
used for immunohistochemistry. |
Table IV
Details of the primary antibodies
used for immunohistochemistry.
| Name | Species | Manufacturer | Cat. no. | Dilution ratio |
|---|
| LRP6 | Goat | Abcam | ab24386 | 1:200 |
| β-catenin | Rabbit | Abcam | ab32572 | 1:200 |
| PPAR-γ | Rabbit | Sanying | 16643-1-AP | 1:300 |
| GSK-3β | Rabbit | Cell Signaling
Technology | #12456S | 1:400 |
| p-β-catenin | Rabbit | Cell Signaling
Technology | #4176 | 1:100 |
| Table VDetails of the secondary antibodies
used for immunohistochemistry. |
Table V
Details of the secondary antibodies
used for immunohistochemistry.
| Name | Manufacturer | Cat. no. | Dilution ratio |
|---|
| HRP-labeled rabbit
anti-goat | Aspen | AS-1108 | 1:200 |
| HRP-labeled goat
anti-rabbit | Aspen | AS-1107 | 1:200 |
Statistical analysis
The quantitative data were analyzed by SPSS 23.0.
All the results are expressed as the means ± SD. One-way ANOVA was
used to compare the differences among the groups, followed by the
LSD post hoc test. A value of P<0.05 was considered to indicate
statistically significant differences. The graphics were designed
with GraphPad Prism 7 Software (GraphPad Software Inc., http://www.graphpad.com/).
Results
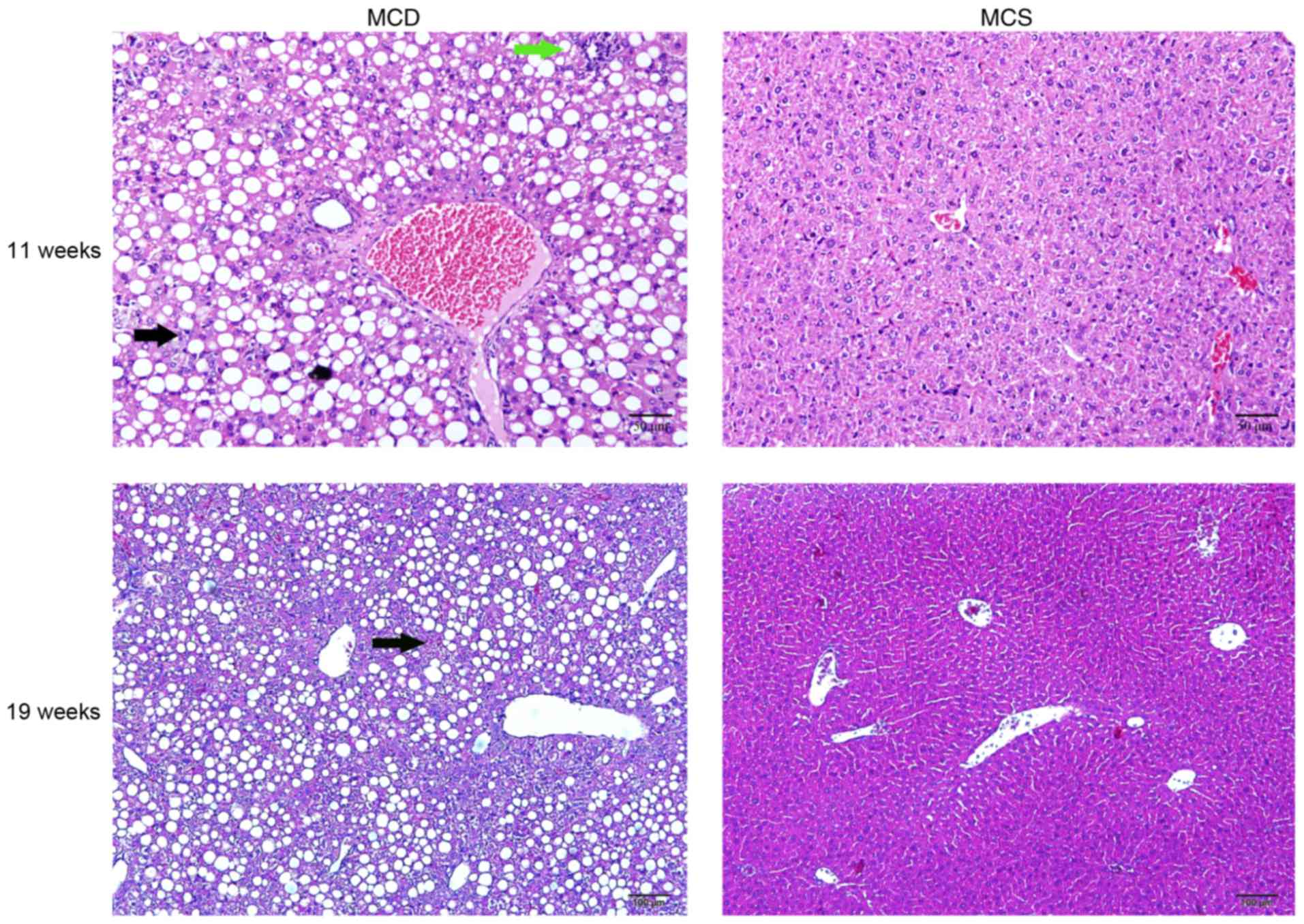
Establishment of NAFLD by feeding
C57BL/6J mice an MCD diet
After 4 weeks, the livers from the C57BL/6J
mice fed an MCD diet were examined by H&E staining, which
revealed an increase in steatosis and inflammatory cell
infiltration in the MCD C57BL/6J group compared with the
normal group, and the normal structure of the hepatic lobules was
disordered. However, hepatic fibrosis was not observed. After
continuously feeding the mice an MCD diet until they reached 19
weeks of age, hepatic fibrosis was not observed (Fig. 3).
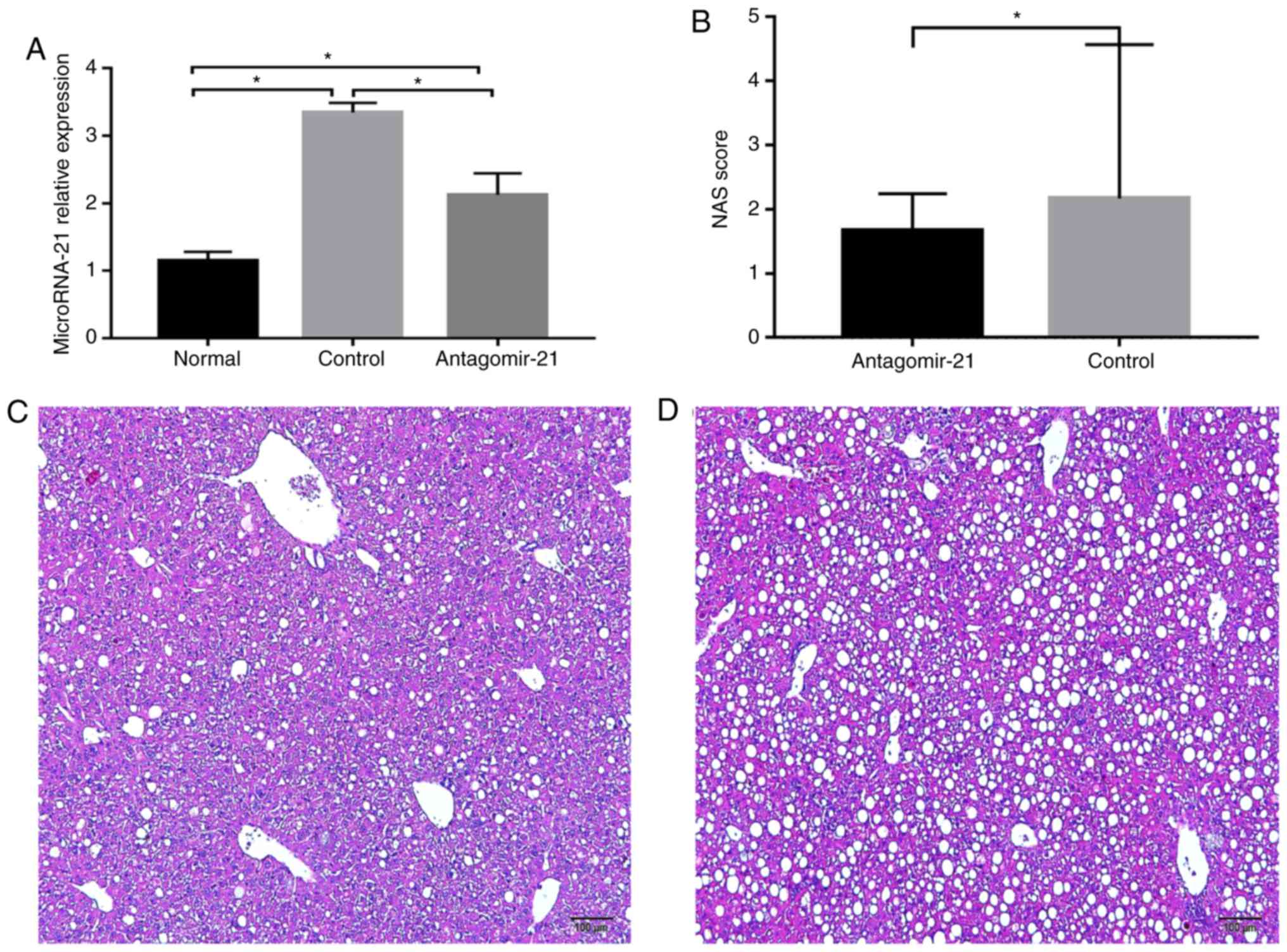
Expression of miR-21 is significantly
increased in C57BL/6J mice fed an MCD diet
After feeding the mice the MCD diet, the expression
of miR-21 in the livers of mice in each group was detected,
and miR-21 expression was markedly increased in the mice fed
the MCD diet (control group) compared with that in the MCS group
(normal group) at 19 weeks of age. Furthermore, antagomir-21 was
injected via the tail vein, and the level of miR-21 in the
livers of the mice was markedly lower than that in the control
group (Fig. 4A). In addition,
hepatic steatosis and inflammation in the mice fed the MCD and
injected with antagomir-21 (Fig.
4C) were significantly reduced compared with the control group
(Fig. 4D).
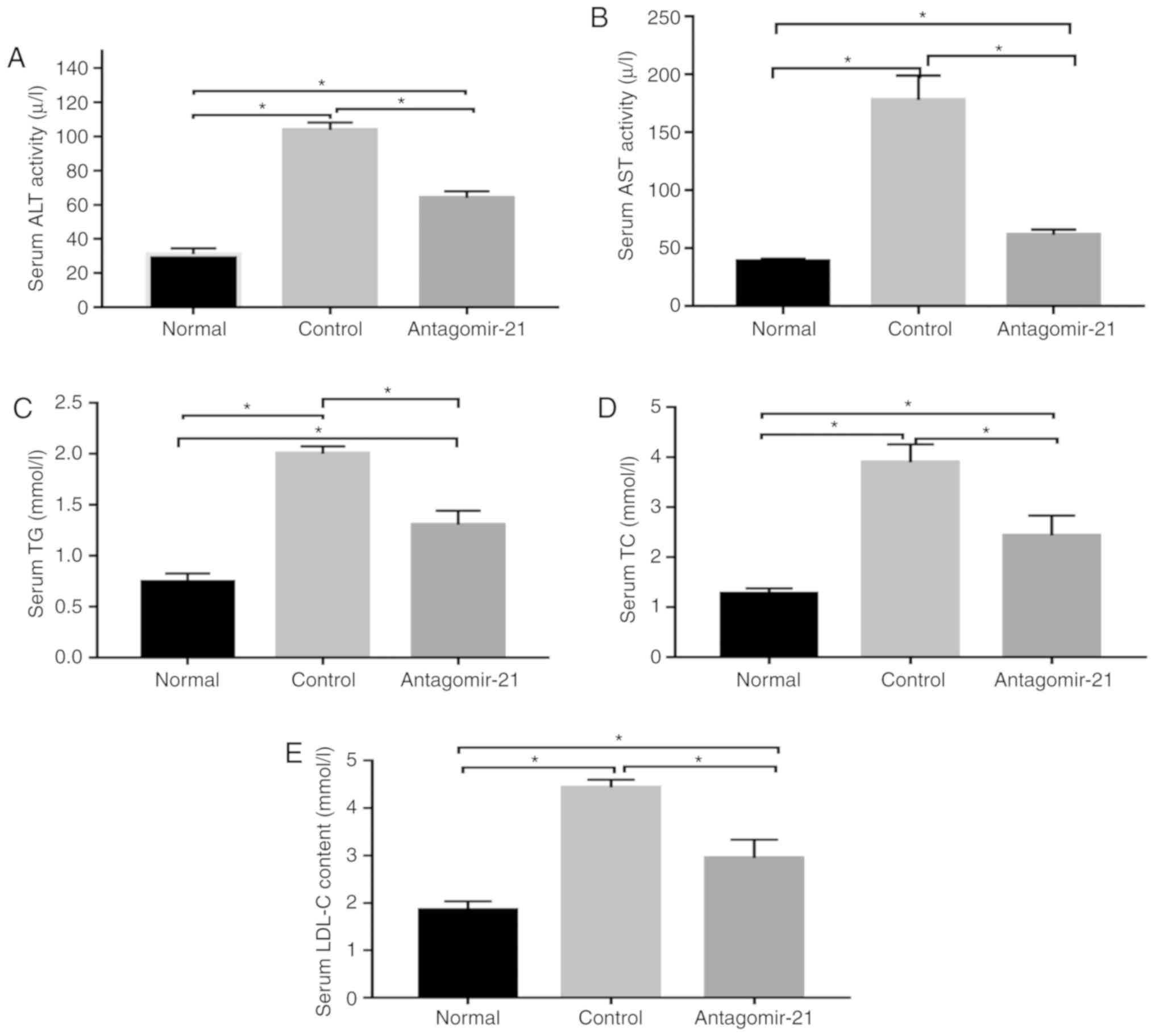
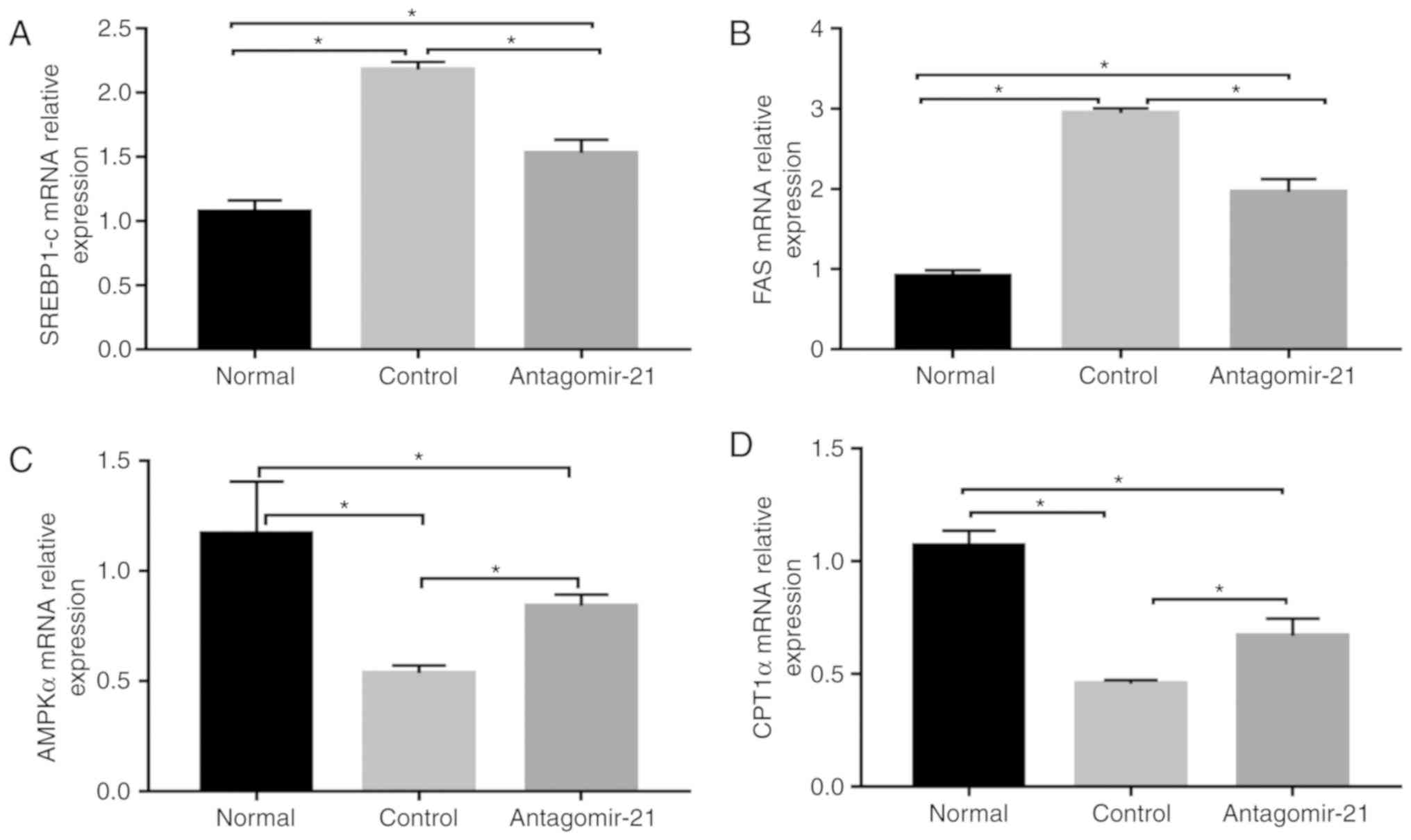
Inhibition of miR-21 expression improves
lipogenesis and transaminase levels in C57BL/6J mice fed an MCD
diet
In mice fed the MCD diet, the levels of serum lipids
(TG, TC and LDL), transaminases (ALT and AST) and genes related to
lipid synthesis (SREBP1c and FAS) were signifi-cantly
higher than those in the normal mice fed the MCS diet, while the
expression of genes related to lipid oxidation (AMPKα and
CPT1α) in the MCD group was relatively lower than that in
the normal group. Following the injection of antagomir-21, the
blood lipid and transaminase levels were improved, the expression
of lipid synthesis-related genes was inhibited, however, the
expression of lipid oxidation genes was increased (Figs. 5 and 6).
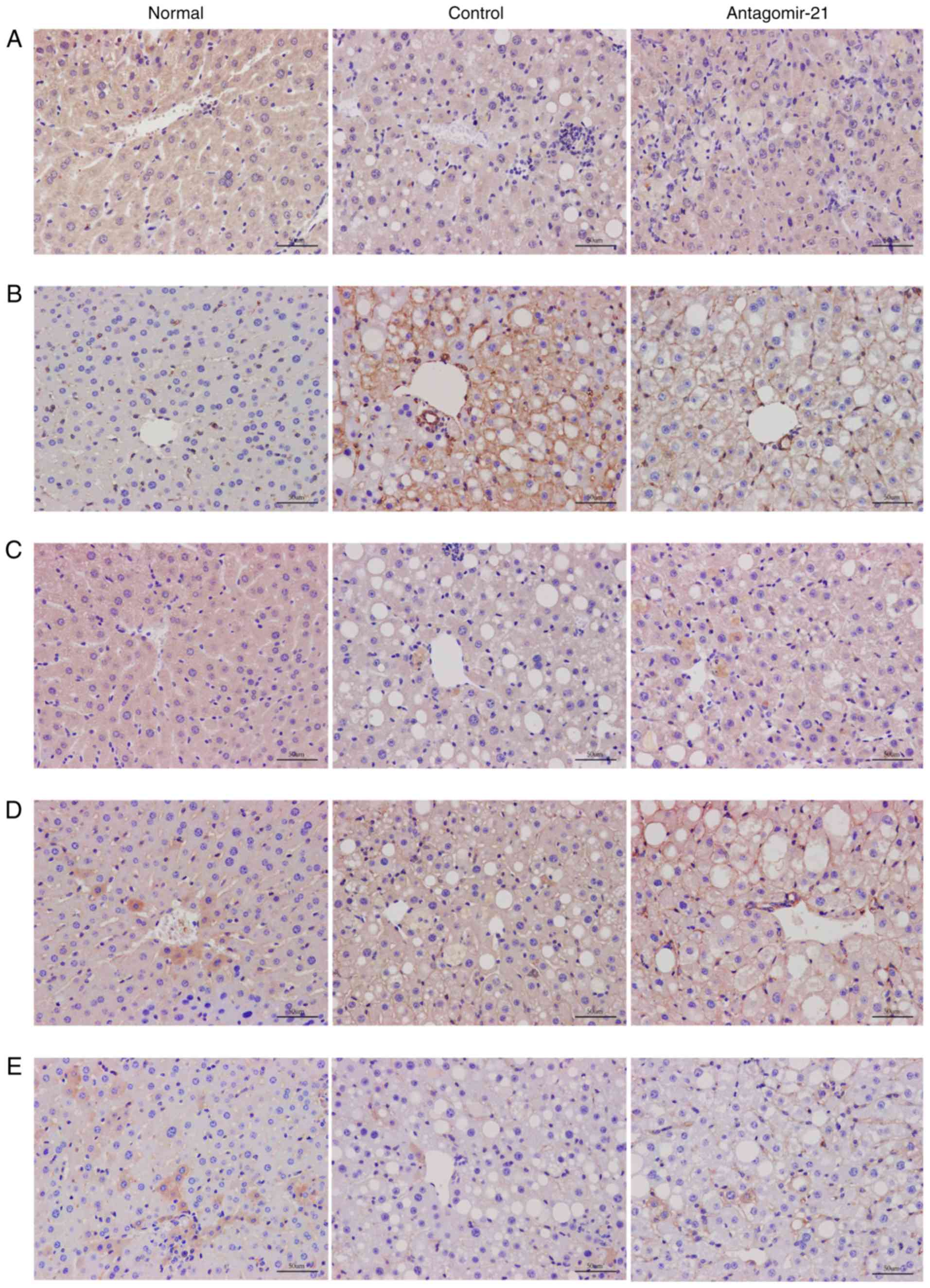
Inhibition of MIR-21 expression activates
the WNT/β-catenin signaling pathway in NAFLD
To validate the regulatory role of miR-21 in
the WNT signaling pathway in mice with NAFLD, antagomir-21 was
injected via the tail vein into mice fed the MCD diet, and the same
amount of saline was used as a control. Western blot analysis and
immunohistochemical staining were used to detect the expression of
classical WNT/β-catenin signaling pathway-related proteins in the
liver. By inhibiting the expression of miR-21, the protein
levels of LRP6 were increased compared to those in the control
group. Moreover, the expression of glycogen synthase kinase-3β
(GSK3β), a downstream degradation complex protein, was reduced, and
the accumulation of β-catenin was increased in the cytoplasm.
Furthermore, the activity of PPAR-γ, a downstream target of the
WNT/β-catenin signaling pathway, was not inhibited following the
activation of the WNT signaling pathway, but was in fact increased
(Figs. 7 and 8).
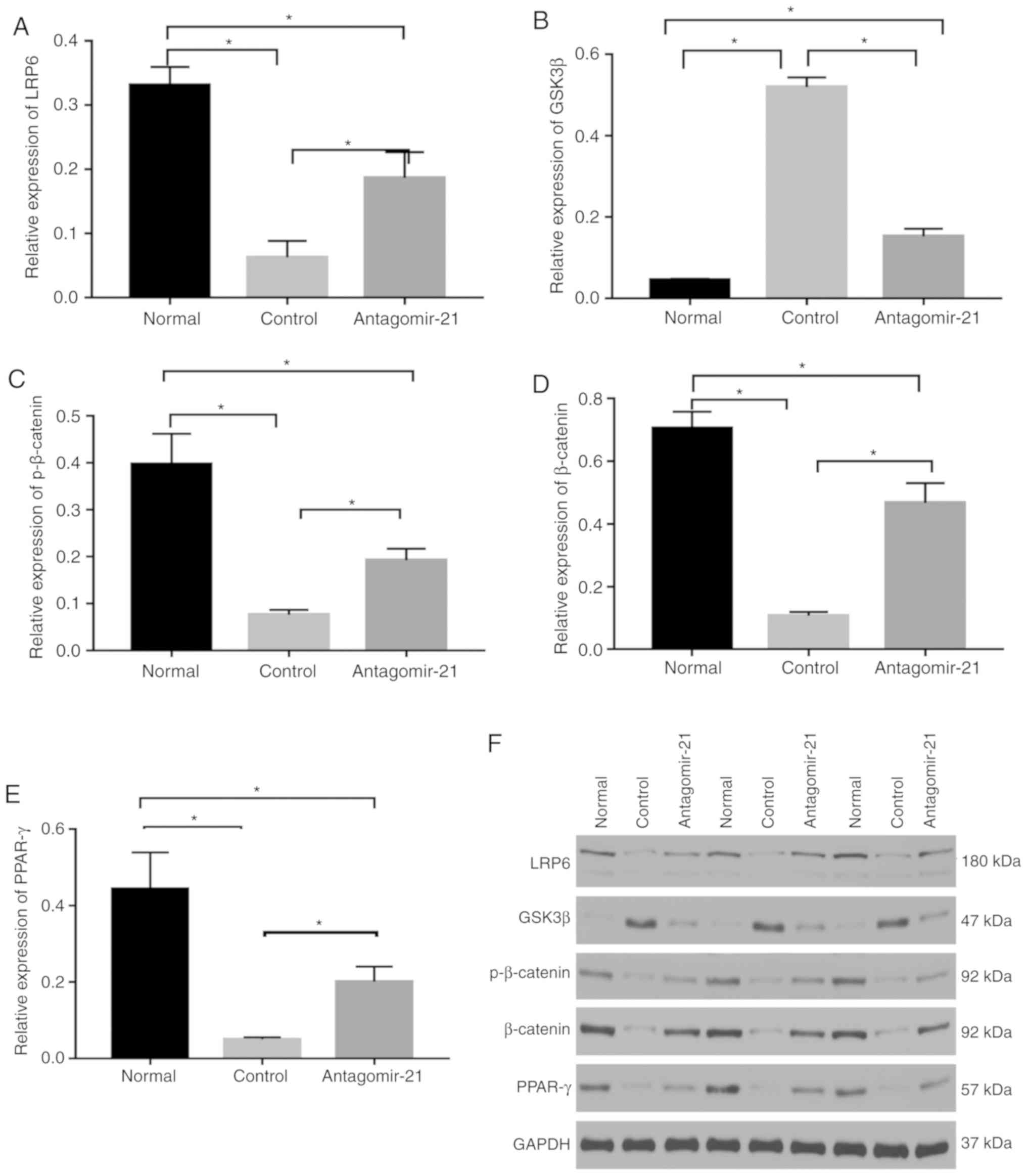 | Figure 7Regulatory effects of mi-21 on
the WNT/β-catenin signaling pathway in NAFLD. Western blot analysis
was performed to compare protein levels. (A) The levels of LRP6
protein were inhibited in mice fed the MCD diet, which was
upregulated by antagomir-21. (B) Protein levels of GSK3β were
higher in the control group and lower in the antagomir-21 group.
(C) Protein levels of p-β-catenin were lower in the control group
and higher in the antagomir-21 group. (D) Protein levels of
β-catenin were lower in the control group and higher in the
antagomir-21 group. (E) Protein levels of PPAR-γ were lower in the
control group and higher in the antagomir-21 group. (F) Expression
levels of LRP6, pβ-catenin, β-catenin, PPAR-γ were inhibited, and
the expression of GSK3β was increased in mice fed the MCD diet;
antagomir-21 administration exerted opposite effects.
*P<0.05. NAFLD, non-alcoholic fatty liver disease;
MCD, methionine and choline-deficient; GSK3β, glycogen synthase
kinase-3β; LRP6, low-density lipoprotein-related receptor 6;
PPAR-γ, peroxisome proliferator-activated receptor γ. |
Discussion
The MCD diet has commonly been used for the
establishment of NAFLD models, and steatosis and steatohepatitis
are usually evident after feeding for 4 weeks (17). Due to the simplicity and rapid
development of the MCD model, we used the MCD diet to establish a
NAFLD model, and we observed evidence of NAFLD in our experiments.
As one of the miRNAs associated with the pathogenesis and
progression of NAFLD, miR-21 has exhibited alterations in
its expression associated with NAFLD. Ahn et al found that
the expression level of miR-21 in the livers of mice with
steatosis induced by a high-fat diet (49.29% of total fat calories)
was lower than that in the control group, and that the expression
of miR-21 was also downregulated in vitro by
cultivating Hepa 1-6 cells in saturated fatty acids, including
stearic acid (SA) (8), and this
is consistent with the results observed in insulin-resistant and
diabetic mice with NAFL induced by a high-fat diet (18) and in patients with NAFLD (9). However, other studies have
demonstrated that serum miR-21 levels in patients with NAFL
are significantly higher than those in healthy individuals and are
increased with the increasing severity of fatty liver, although the
correlation has not been found to be statistically signifi-cant
(19). A previous study by Becker
et al found that there was no significant difference in the
expression of miR-21 in serum from patients with NAFL and
healthy individuals (20). In
this study, we established a model of NAFLD by feeding
C57BL/6J mice an MCD diet and found that the expression of
miR-21 in the liver was increased by ~3-fold compared to
that in the liver in the normal group that was fed the MCS diet.
The discrepancy in the expression of miR-21 in different
research models of NAFLD could be explained by the experimental
results of Loyer et al, who found that miR-21 was
mainly expressed in inflammatory cells and bile duct cells, but was
not expressed as much in the liver (10).
In the model of NAFLD in this study, the livers of
mice exhibited evidence of steatosis, the infiltration of
inflammatory cells and damage to hepatocytes. The inhibition of
miR-21 expression in mice alleviated steatosis and
inflammation due to NAFLD, which is consistent with other research
results (10,21). Transaminase and lipid levels in
the blood were also improved, and the expression of lipid
metabolism genes was also ameliorated following the inhibition of
miR-21. Antagomir-21 could be used as a treatment method.
Thus far, whether therapy targeting miR-21 can alleviate
liver fibrosis has not yet been determined. miR-21 may
participate in the process of liver fibrosis through its effects on
multiple targets by inhibiting SMAD7 via the TGF-β/SMAD7 signaling
pathway (11) and activating the
extracellular signal-regulated kinase 1 (ERK1) signaling pathway by
inhibiting SPROUTY2 (SPRY2) to promote epithelial-to-mesenchymal
transition (EMT) (22). However,
the results from the study by Caviglia et al demonstrated
that the inhibition of miR-21 did not prevent the
development of liver fibrosis (23).
LRP6 participates in the endocytosis of lipoproteins
(6). It has been recognized that
the dysfunction of LRP6, such as that caused by the LRP6 gene
mutation LRP6R611C (R611C: rs121918313), is involved in
dyslipidemia and NAFLD. LRP6R611C mice exhibit
hyperlipidemia and liver lipid accumulation, lipid synthesis genes,
such as SREBP1c and SREBP2, and regulated downstream
lipases, including acetyl coenzyme A carboxylase, FAS,
SCD1, diglyceryl transferase 1 and the elongation of very
long chain fatty acids [fatty acid elongase (ELOVL)] family
members, are increased in LRP6R611C mice (6). Similarly, Montazeri-Najafabady et
al recently demonstrated that the LRP6V1062I
polymorphism (V1062I: rs2302685) was associated with an increased
risk of hyperlipidemia in Iranian children and adolescents, which
also increased the risk of elevated total cholesterol, TG, LDL and
non-HDL levels (24). By using
rmWnt3a, the blood lipid levels of LRP6R611C mice and
the enzymes involved in lipid de novo synthesis can be
normal-ized (25). These results
indicate that LRP6 can be utilized as a therapeutic target for the
treatment of NAFLD by regulating the WNT/β-catenin signaling
pathway. In a previous study by our group, the overexpression of
miR-21 in hepatocytes inhibited the expression of LRP6
(14). In the mouse experiments
in this study, we observed that LRP6 expression was increased,
which activated the WNT/β-catenin signaling pathway, decreased the
degradation of β-catenin and caused more β-catenin to translo-cate
to the nucleus to regulate the expression of target genes, such as
PPAR-γ, following the inhibition of the expression of miR-21
in the livers of mice (Fig. 1).
However, in contrast to previous research (6), in this study, the level of PPAR-γ
was increased compared with that in the control group. This may be
related to the regulation of miR-21. Some studies have shown
that PPAR-γ is negatively associated with the expression of
miR-21 (18), however,
whether it is a target of miR-21 has yet to be determined.
PPAR-γ can induce preadipocyte differentiation, promote
adipo-genesis, induce adipocytes to absorb and store free fatty
acids, and promote the transfer of liver fat to adipocytes, at the
same time, PPAR-γ activation alleviates inflammation via its
negative interaction with nuclear factor-κB (NF-κB) and signal
transducer and activator of transcription (STAT), and promotes
macrophage cell transition to M2 to inhibit the development of
NAFLD (26), for example, the
agonist, rosiglitazone, has been shown to prevent NASH progression
in animal models induced by dietary methionine choline deficiency.
The antagonistic expression of miR-21 may alleviate NASH
steatosis and inflammation by regulating the WNT/β-catenin
signaling pathway in addition to negatively regulating other
targets, such as PPAR-α (10).
In addition to PPAR-γ, miR-21 may also
regulate SREBP1c via the WNT signaling pathway.
SREBP1c promotes the de novo synthesis of fat and
adipogenesis. SREBP1/2 is normally retained in the
endoplasmic reticulum through Insig1 and Insig2, activated by AKT,
transported to the Golgi apparatus via SREBP cleavage-activating
protein (SCAP) to induce protein hydrolysis and maturation,
and then ectopically transferred to the nucleus, in which it binds
to target genes (25).
miR-21 regulates the expression of SREBP1c through
the Hbp1-p53-Srebp1c pathway (13). LRP6R611C also
participates in the progression of NAFLD through the nutritional
IGF1-AKT-mTOR-SREBP1/2 pathway. In this study, we also
verified the expression of SREBP1c and the expression level
of the downstream target, FAS. After miR-21
expression was reduced, LRP6 was activated, and SREBP1c was
inhibited. The expression levels of the CPT1α and
AMPKα genes, which are related to lipolysis, were increased,
and thereby NAFLD was attenuated. Therefore, miR-21 can
regulate the expression of SREBP1c-related lipid metabolism
genes through the WNT/β-catenin signaling pathway. Due to limited
funding, this study did not examine the effects of inhibiting
miR-21, while antagonizing LRP6 expression or activating
LRP6 expression with other drugs in NAFLD. Simultaneously, if
miR-21 knockout mice can be used instead of miR-21
inhibitors to treat mice, perhaps better results would be
obtained.
Further limitations to this study include the small
numbers of mice, the simple design of the experiment and the lack
of an appropriate control. However, in spite of these, this study
demonstrates that the inhibition of miR-21 expression in
mice attenuated NASH steatosis and inflammation, partly by
targeting the coreceptor LRP6 and activating the WNT/β-catenin
signaling pathway. This finding was consistent with that of a
previous cell study (14), which
further supported our hypothesis that miR-21 may participate
in the pathogenesis of NAFLD via its regulation of the
WNT/β-catenin signaling pathway; this has enriched our
understanding of the molecular mechanisms of miR-21 in
NAFLD. This finding also supported the possibility of inhibiting
MIR-21 expression as a treatment for NAFLD. Due to the
complexity of the pathogenesis of NAFLD and the influence of
lifestyle habits, single-target therapy has not achieved
satisfactory results. miRNAs have multitarget characteristics and
can be regulated by multiple miRNAs. Although the inhibition of
miR-21 alone may not achieve satisfactory results, the
targeting of miR-21 could also enhance the therapeutic
efficacy of other drugs, such as a combination of miR-21 and
the farne-soid X receptor (FXR) agonist obeticholic acid (21), and the complex molecular mechanism
network underlying the involvement of miR-21 in NAFLD
warrants further investigation. In the future, multitarget drug
combinations may become an option for NAFLD treatment.
Abbreviations:
|
NAFLD
|
non-alcoholic fatty liver disease
|
|
1H-MRS
|
proton magnetic resonance
spectroscopy
|
|
MRI
|
magnetic resonance imaging
|
|
NAFL
|
non-alcoholic fatty liver
|
|
NASH
|
non-alcoholic steatohepatitis
|
|
C/EBPα
|
CCAAT enhancer-binding protein α
|
|
PPAR-γ
|
peroxisome proliferator-activated
receptor γ
|
|
LRP6
|
low-density lipoprotein-related
receptor 6
|
|
SREBP1c
|
sterol regulatory element-binding
transcription factor 1c
|
|
miR-21
|
microRNA-21
|
|
PPAR-α
|
peroxisome proliferator activated
receptor α
|
|
FABP7
|
fatty acid binding protein 7
|
|
HMGCR
|
3-hydroxy-3-methylglutaryl coenzyme A
reductase
|
|
PTEN
|
phosphate and tension homolog
|
|
HBP1D
|
HMG-box transcription factor 1D
|
|
MCD
|
methionine and choline-deficient
|
|
MCS
|
methionine- and
choline-sufficient
|
|
ALT
|
serum alanine aminotransferase
|
|
AST
|
aspartate aminotransferase
|
|
TG
|
triglyceride
|
|
TC
|
total cholesterol
|
|
LDL
|
low-density lipoprotein
|
|
FAS
|
fatty acid synthase
|
|
AMPKα
|
adenosine 5-monophosphate
(AMP)-activated protein kinase α
|
|
CPT1α
|
carnitine palmitoyl transferase
1α
|
|
GAPDH
|
glyceraldehyde-3-phosphate
dehydrogenase
|
|
GSK3β
|
glycogen synthase kinase-3β
|
|
SA
|
stearic acid
|
|
ERK1
|
extracellular signal-regulated kinase
1
|
|
SPRY2
|
Sprouty2
|
|
EMT
|
epithelial-to-mesenchymal
transition
|
|
ELOVL
|
fatty acid elongase
|
|
NF-κB
|
nuclear factor-κB
|
|
STAT
|
signal transducer and activator of
transcription
|
|
SCAP
|
SREBP cleavage-activating protein
|
|
FXR
|
farnesoid X receptor
|
Acknowledgements
Not applicable.
Funding
The study was supported by a grant from the
Affiliated Hospital of Southwestern Medical University Research
Project-General Project in 2017 (grant no. 17172), which was
awarded to Dr Chun Yang who was responsible for the funding for
this study.
Availability of data and materials
All data generated or analyzed during this study
are included in this published article.
Authors' contributions
XMW, XYW and CPL were involved in the conception,
design and review the manuscript in the study. XMW and XYW were
involved in the writing and revising of the original draft. XMW,
XYW and YMH were involved in data collection and statistical
analysis. XC, LS and MHL were responsible for experimental method
operation, resource procurement and fund management. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All animals received humane care according to the
guidelines of the Institutional Animal Care and Use Committee of
Southwest Medical University, and the experiment was approved by
the Experimental Animal Ethics Committee of Southwest Medical
University (application acceptance no. 20180521-11).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hardy T, Oakley F, Anstee QM and Day CP:
Nonalcoholic fatty liver disease: Pathogenesis and disease
spectrum. Annu Rev Pathol. 11:451–496. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cai J, Zhang XJ and Li H: Progress and
challenges in the prevention and control of nonalcoholic fatty
liver disease. Med Res Rev. 39:328–348. 2019. View Article : Google Scholar
|
|
3
|
Fiorucci S, Biagioli M and Distrutti E:
Future trends in the treatment of non-alcoholic steatohepatitis.
Pharmacol Res. 134:289–298. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ackers I and Malgor R: Interrelationship
of canonical and non-canonical Wnt signalling pathways in chronic
metabolic diseases. Diab Vasc Dis Res. 15:3–13. 2018. View Article : Google Scholar :
|
|
5
|
Green CJ, Parry SA, Gunn PJ, Ceresa CDL,
Rosqvist F, Piché ME and Hodson L: Studying non-alcoholic fatty
liver disease: The ins and outs of in vivo, ex vivo and in vitro
human models. Horm Mol Biol Clin Investig Aug. 11:2018.Epub ahead
of print.
|
|
6
|
Go GW: Low-density lipoprotein
receptor-related protein 6 (LRP6) is a novel nutritional
therapeutic target for hyperlip-idemia, non-alcoholic fatty liver
disease, and atherosclerosis. Nutrients. 7:4453–4464. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang S, Song K, Srivastava R, Dong C, Go
GW, Li N, Iwakiri Y and Mani A: Nonalcoholic fatty liver disease
induced by noncanonical Wnt and its rescue by Wnt3a. FASEB J.
29:3436–3445. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ahn J, Lee H, Jung CH and Ha T: Lycopene
inhibits hepatic steatosis via microRNA-21-induced downregulation
of fatty acid-binding protein 7 in mice fed a high-fat diet. Mol
Nutr Food Res. 56:1665–1674. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sun C, Huang F, Liu X, Xiao X, Yang M, Hu
G, Liu H and Liao L: miR-21 regulates triglyceride and cholesterol
metabolism in non-alcoholic fatty liver disease by targeting HMGCR.
Int J Mol Med. 35:847–853. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Loyer X, Paradis V, Hénique C, Vion AC,
Colnot N, Guerin CL, Devue C, On S, Scetbun J, Romain M, et al:
Liver microRNA-21 is overexpressed in non-alcoholic steatohepatitis
and contributes to the disease in experimental models by inhibiting
PPARα expression. Gut. 65:1882–1894. 2016. View Article : Google Scholar
|
|
11
|
Dattaroy D, Pourhoseini S, Das S, Alhasson
F, Seth RK, Nagarkatti M, Michelotti GA, Diehl AM and Chatterjee S:
Micro-RNA 21 inhibition of SMAD7 enhances fibrogenesis via
leptin-mediated NADPH oxidase in experimental and human
nonalcoholic steatohepatitis. Am J Physiol Gastrointest Liver
Physiol. 308:G298–G312. 2015. View Article : Google Scholar :
|
|
12
|
Wei J, Feng L, Li Z, Xu G and Fan X:
MicroRNA-21 activates hepatic stellate cells via PTEN/Akt
signaling. Biomed Pharmacother. 67:387–392. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu H, Ng R, Chen X, Steer CJ and Song G:
MicroRNA-21 is a potential link between non-alcoholic fatty liver
disease and hepa-tocellular carcinoma via modulation of the
HBP1-p53-Srebp1c pathway. Gut. 65:1850–1860. 2016. View Article : Google Scholar
|
|
14
|
Li CP, Li HJ, Nie J, Chen X and Zhou X:
Mutation of miR-21 targets endogenous lipoprotein receptor-related
protein 6 and nonalcoholic fatty liver disease. Am J Transl Res.
9:715–721. 2017.PubMed/NCBI
|
|
15
|
Kleiner DE, Brunt EM, Van Natta M, Behling
C, Contos MJ, Cummings OW, Ferrell LD, Liu YC, Torbenson MS,
Unalp-Arida A, et al: Design and validation of a histological
scoring system for nonalcoholic fatty liver disease. Hepatology.
41:1313–1321. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
17
|
Kong M, Chen X, Xu H, Wenping, Fang M and
Xu Y: Hepatocyte-specific deletion of Brg1 alleviates
methionine-and-choline-deficient diet (MCD) induced non-alcoholic
steatohepatitis in mice. Biochem Biophys Res Commun. 503:344–351.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao XY and Shao K: Roles of microRNA-21
in the pathogenesis of insulin resistance and diabetic
mellitus-induced non-alcoholic fatty liver disease. Zhongguo Yi Xue
Ke Xue Yuan Xue Bao. 38:144–149. 2016.In Chinese. PubMed/NCBI
|
|
19
|
Yamada H, Suzuki K, Ichino N, Ando Y,
Sawada A, Osakabe K, Sugimoto K, Ohashi K, Teradaira R, Inoue T, et
al: Associations between circulating microRNAs (miR-21, miR-34a,
miR-122 and miR-451) and non-alcoholic fatty liver. Clin Chim Acta.
424:99–103. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Becker PP, Rau M, Schmitt J, Malsch C,
Hammer C, Bantel H, Müllhaupt B and Geier A: Performance of serum
microRNAs -122, -192 and -21 as biomarkers in patients with
non-alcoholic steatohepatitis. PLoS One. 10:e01426612015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rodrigues PM, Afonso MB, Simão AL,
Carvalho CC, Trindade A, Duarte A, Borralho PM, Machado MV,
Cortez-Pinto H, Rodrigues CM and Castro RE: miR-21 ablation and
obeticholic acid ameliorate nonalcoholic steatohepatitis in mice.
Cell Death Dis. 8:e27482017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu K, Ye C, Lin L, Chu Y, Ji M, Dai W,
Zeng X and Lin Y: Inhibiting miR-21 attenuates experimental hepatic
fibrosis by suppressing both the ERK1 pathway in HSC and hepatocyte
EMT. Clin Sci (Lond). 130:1469–1480. 2016. View Article : Google Scholar
|
|
23
|
Caviglia JM, Yan J, Jang MK, Gwak GY, Affo
S, Yu L, Olinga P, Friedman RA, Chen X and Schwabe RF: MicroRNA-21
and Dicer are dispensable for hepatic stellate cell activation and
the development of liver fibrosis. Hepatology. 67:2414–2429. 2018.
View Article : Google Scholar :
|
|
24
|
Montazeri-Najafabady N, Dabbaghmanesh MH
and Mohammadian Amiri R: The association of LRP6 rs2302685 (V1062I)
polymorphism with the risk of hyperlipidemia in Iranian children
and adolescents. Ann Hum Genet. 82:382–388. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Go GW, Srivastava R, Hernandez-Ono A, Gang
G, Smith SB, Booth CJ, Ginsberg HN and Mani A: The combined
hyperlip-idemia caused by impaired Wnt-LRP6 signaling is reversed
by Wnt3a rescue. Cell Metab. 19:209–220. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Silva AKS and Peixoto CA: Role of
peroxisome proliferator-activated receptors in non-alcoholic fatty
liver disease inflammation. Cell Mol Life Sci. 75:2951–2961. 2018.
View Article : Google Scholar : PubMed/NCBI
|