Introduction
Acute lung injury (ALI) is a leading cause of acute
respiratory failure (1). ALI is
characterized by extreme inflammation, the release of
pro-inflammatory cytokines, excessive neutrophil infiltration and
lung endothelial/epithelial cell injury, resulting in edema and gas
exchange deterioration (2).
However, the clinical mortality rate of the severe form of ALI,
acute respiratory distress syndrome (ARDS), remains >40.0%
(1). Therefore, more effective
therapeutic strategies for ARDS are urgently required.
Macrophages, the principal immune cells in the
lungs, produce inflammatory molecules and carry out vital functions
in the molecular mechanisms of ALI, such as boosting neutrophil
infiltration and triggering inflammatory reactions (3). Neutrophils trigger the release of
pro-inflammatory cytokines, such as tumor necrosis factor (TNF)-α,
interleukin (IL)-1β and IL-6 (4).
These pro-inflammatory cytokines induce the production of oxidants,
which are associated with the activation of nuclear factor
κ-light-chain-enhancer of activated B cells (NF-κB), eventually
contributing to ALI (5,6). Accordingly, oxidative stress is
increased in lipopolysaccha-ride (LPS)-induced ALI (7). The transcription factor, nuclear
factor-erythroid 2 related factor 2 (Nrf2), plays a critical role
in protection against ALI by inducing the expression of antioxidant
and detoxifying enzymes and proteins (8). For example, it has been reported
that Nrf2 attenuates ALI and inflammation by supressing Toll-like
receptor (TLR)4 and Akt signaling (9).
Artemisinin is isolated from Artemisia annua,
a Chinese traditional medicinal herb. Artesunate is a water-soluble
hemisuccinate derivative of artemisinin. Studies have reported that
artesunate inhibits ischemia-reperfusion-induced lung inflammation
and LPS-induced ALI (10,11). Dihydroartemisinin (DHA), the major
active metabolite of artemisinin or arte-sunate, is an effective
and widely distributed anti-malarial drug with good absorption
(12,13). DHA is more stable and ten times
more effective than artesunate (14). Recent studies have demonstrated
that DHA not only exerts an anti-malarial effect, but also exerts
anticancer, anti-organizational fibrosis and anti-neuronal cell
death effects (15-17). However, whether DHA can attenuate
ALI and affect NF-κB signaling activation in macrophages remains
unclear.
The present study thus hypothesized that DHA may
attenuate LPS-induced ALI and evaluated the effects of DHA on
LPS-treated macrophages to elucidate the mechanisms through which
DHA attenuates LPS-induced ALI.
Materials and methods
Ethics statement
The Ethics Committee of the Center for Scientific
Research with Animal Models at Central South University (Changsha,
China) approved the experiments, which were performed in accordance
with the guidelines of the National Institutes of Health. Mice were
anesthetized with pentobarbital sodium (80 mg/kg, intraperitoneal
injection), and all necessary efforts were taken to minimize
suffering prior to the experiments.
Animal experiments
Male adult C57bl/6 mice were kept in
climate-controlled quarters with a 12-h light/dark cycle and a
relative humidity of 40-60%, and were provided with food and water
ad libitum at a temperature of 25°C. The mice were housed
for 1 week for environmental adaptation prior to experimentation.
The mice were randomly divided into 4 groups (weight, 20-25 g; age,
8 weeks; n=24 in each group) as follows: i) The control group; ii)
ALI group; iii) DHA group; and iv) ALI + DHA group. The ALI model
was induced by the intratracheal injection of LPS (E. coli
O111:B4; 5 mg/kg; Sigma-Aldrich; Merck KGaA) in 50 µl saline
as described in our previous studies (18,19). Mice in the control group received
saline. Mice in the DHA group received DHA (75 mg/kg;
Sigma-Aldrich; Merck KGaA) only. Mice in the LPS + DHA group were
treated with DHA (75 mg/kg) via intragastric administration 1 h
prior to LPS administration. A total of 12 h following the LPS
administration, the mice were anesthetized by an intraperitoneal
(i.p.) injection of sodium pentobarbital (80 mg/kg) and blood was
collected for further analysis. Following the collection of lung
tissue, the mice were then sacrificed by an i.p. injection of 200
mg/kg sodium pentobarbital.
Survival experiment
For survival analysis, another 80 mice were randomly
divided into 4 groups as follows: i) The control group; ii) ALI
group; iii) DHA group; and iv) ALI + DHA group (n=20 in each
group). The mice were treated with LPS at a lethal dose (25 mg/kg,
intratracheal) (20-23). Mice in the DHA group received DHA
(75 mg/kg; Sigma-Aldrich; Merck KGaA) only. Mice in the LPS + DHA
group were treated with DHA (75 mg/kg) via intragastric
administration 2 h following the adminstration of LPS. The survival
rate was monitored every 6 h, as previously described (19).
Histological analysis
Lung tissue was excised and immersed in 4%
paraformaldehyde for 24 h at 4°C. Paraffin-embedded lung tissue was
sectioned at a thickness of 4 µm and was then stained with
haematoxylin and eosin (cat. no. G1120; Solarbio) for 5 min at room
temperature for pathological analysis. According to a previous
study, the severity of inflammation was graded between 0 and 4 as
follows: 1, <25% lung involvement; 2, 25-49% lung involvement;
3, 50-75% lung involvement; and 4, >75% lung involvement
(24). Lung injury was scored by
3 pathologists blinded to the treatments.
Bronchoalveolar lavage fluid (BALF)
acquisition and analysis
BALF was collected by lavaging the lungs with 1 ml
PBS 3 times and was centrifuged at 800 x g for 5 min at 4°C. Total
cells, macrophages and neutrophils were counted with a
hemocytometer following Wright-Giemsa staining for 10 min at room
temperature. The cell-free supernatant of BALF was harvested, and
total protein content was determined using a bicinchoninic acid
protein assay kit (Thermo Fisher Scientific, Inc.).
Lung wet-to-dry weight ratio
The lungs were excised from the mice and blood was
removed by blotting the tissue with filter papers until dry. After
weighing (wet weight), the lungs were placed in an incubator at
60°C for 48 h and then weighed again (dry weight). The wet-to-dry
ratio of the lungs was calculated to reflect edema.
Lactate dehydrogenase (LDH) activity
assay
LDH activity in BALF was determined with an LDH
Cytotoxicity assay kit (cat. no. A020; Nanjing Jiancheng
Bioengineering Institute) according to the manufacturer's protocol.
The absorbance at 490 nm was measured using a microplate reader
(Thermo Fisher Scientific, Inc.).
Measurement of myeloperoxidase (MPO),
malondialde- hyde (MDA), superoxide dismutase (SOD) and glutathione
(GSH) levels
The MPO, MDA, SOD and GSH levels were detected using
the related kits (MPO: cat. no. A044; MDA: cat. no. A003; SOD: cat.
no. A001; GSH: cat. no. A005; Nanjing Jiancheng Bioengineering
Institute). All procedures were performed according to the
manufacturer's protocols.
Primary peritoneal macrophages
Male C57bl/6 mice (8-week-old, n=20) were used to
extract primary peritoneal macrophages. At 3 days following the
intraperitoneal injection of 3 ml 3% thioglycolate (Sigma-Aldrich;
Merck KGaA), the animals were euthanized by CO2
inhalation (flow rate of CO2, 20%) and peritoneal
macrophages were harvested by peritoneal lavage with cooled
RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc.). Cells were
collected by centrifugation (room temperature, 1,200 g, 10 min) and
resuspended with culture medium. Cells were plated into 6- or
12-well plates (1x106 cells/well) and the culture medium
was discarded 2 h later. After being allowed to rest overnight, the
cells were treated with DHA (20 µM) for 1 h followed by
treatment with LPS (100 ng/ml) for a further 6 h at 37°C. To
investigate the effects of ML385 (an Nrf2 inhibitor; MedChemExpres)
on macrophages, primary macrophages were pre-treated with ML385 (20
µM) for 30 min, and the cells were then treated with DHA (20
µM) for 1 h followed by treatment with LPS (100 ng/ml) for a
further 6 h.
Immunofluorescence
Primary peritoneal macrophages were plated on
polylysine-coated coverslips. Following fixing with 4%
paraformaldehyde at 4°C and permeabilization with Triton X-100,
macrophages were incubated overnight at 4°C with anti-CD68 antibody
(cat. no. ab125212; 1:50; Abcam) and anti-F4/80 antibody (cat. no.
ab6640; 1:50; Abcam). FITC (cat. no. ab6717; 1:200; Abcam) and Cy3
(cat. no. ab6953; 1:200; Abcam)-conjugated secondary antibody were
then applied for 1 h at room temperature. Following nuclear
staining with DAPI (cat. no. C0065; Solarbio), the macrophages were
viewed under a fluorescence microscope (Thermo Fisher Scientific,
Inc.).
Cytokine measurements
The levels of TNF-α, IL-1β and IL-6 in mouse sera
and the culture supernatants of primary peritoneal macrophages were
detected using enzyme-linked immunosor-bent assay (ELISA) kits
(TNF-α, cat. no. BMS607-3; IL-1β, cat. no. BMS6002; IL-6, cat. no.
BMS603-2; Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions.
Total RNA extraction and reverse
transcription-quantitative polymerase chain reaction
Total RNA was extracted from the lung tissue or
macrophages using RNAiso reagent (cat. no. 5301100; Takara Bio,
Inc.). Total RNA (1 µg) was used for the synthesis of cDNA
using a PrimeScript RT Reagent kit with gDNA Eraser (cat. no.
RR047A; Takara Bio, Inc.). qPCRs were run using SYBR-Green
Real-time PCR Master mix (Thermo Fisher Scientific, Inc.) on a
Bio-Rad real-time PCR system (CFX96 Touch™; Bio-Rad Laboratories,
Inc.). The amplification conditions were as follows:
Pre-degeneration at 95°C for 10 min, then 40 cycles of denaturing
at 95°C for 15 sec, annealing at 60°C for 30 sec and extension at
72°C for 30 sec, and a final extension at 72°C for 5 min. Relative
fold expression levels were normalized to GAPDH and calculated
using the 2-∆∆Cq method (25). The primer sequences were as
follows: TNF-α forward, ACA GCA AGG GAC TAG CCA GGA G and reverse,
GGA GTG CCT CTT CTG CCA GT; IL-1β forward, GGG CCT CAA AGG AAA GAA
TC and reverse, TAC CAG TTG GGG AAC TCT GC; IL-6 forward, CTG GGG
ATG TCT GTA GCT CA and reverse, CTG TGA AGT CTC CTC TCC GG; and
β-actin forward, TCT TTG CAG CTC CTT CGT TG and reverse, TCC TTC
TGA CCC ATT CCC AC.
Western blot analysis
The lung tissues or cell samples were lysed in RIPA
buffer (cat. no. P0013K; Beyotime) at 4°C for 30 min, and the total
proteins were quantified using a BCA kit (cat. no. P0010;
Beyotime). Briefly, 30 µg proteins were separated by 12%
SDS-PAGE gels and transferred onto polyvinylidene difluoride
membranes (EMD Millipore). The membranes were blocked with 5%
fat-free milk for 2 h and then probed at 4°C overnight with the
following primary antibodies: Anti-phosphorylated(p)-I-κB (1:1,000;
cat. no. 2859; Cell Signaling Technology, Inc.), anti-I-κB
(1:1,000; cat. no. 4812; Cell Signaling Technology, Inc.),
anti-Nrf2 (1:1,000; cat. no. YT3189; ImmunoWay), anti-heme
oxygenase 1 (HO-1; 1:1,000; cat. no. 43966; Cell Signaling
Technology, Inc.), anti-p65 (1:1,000; cat. no. 10745; Proteintech),
anti-β-actin (1:1,000; cat. no. 4970; Cell Signaling Technology,
Inc.) and anti-p-p65 (1:1,000; cat. no. 3033; Cell Signaling
Technology, Inc). After washing with TBST 3 times, the membranes
were incubated with secondary antibodies (1:7,500; cat. no. ab6721;
Abcam) at room temperature for 1 h.
Enhanced chemiluminescence (EMD Millipore) was used
to detect the protein content. Images were obtained using a
ChemiDoc XRS system (Bio-Rad Laboratories, Inc.).
Measurement of ROS production
The lung tissues were homogenized and stained with
50 µM of DCFH-DA (cat. no. S0033; Beyotime) at 37°C in the
dark for 30 min. DCF fluorescence intensities were detected by a
multi-detection reader (Thermo Fisher Scientific, Inc.) at an
excitation and emission wavelength of 485 and 535 nm.
Statistical analysis
Data were analyzed using SPSS 19.0 software (SPSS,
Inc.). All data are expressed as the means ± standard error of the
mean. Means were compared by two-way ANOVA followed by Tukey's
post-hoc test to assess significance. Survival analysis was carried
out using the Kaplan-Meier log-rank test. P<0.05 was considered
to indicate a statistically significant difference.
Results
DHA attenuates lung tissue injury in mice
with LPS-induced ALI
The lung tissues of the control mice were with
intact alveoli, while the lung tissues from the mice with ALI
exhibited obvious pulmonary edema, alveolar disarray and
inflammatory cell infiltration in the alveolar cavity. Following
treatment with DHA, the pathological changes induced by LPS in the
lungs were attenuated (Fig. 1A).
DHA treatment also significantly reduced the lung injury score and
LDH activity in the BALF of mice with ALI (Fig. 1B and C). LPS significantly
increased the lung wet-to-dry weight ratio and the total protein
content in BALF, and these were reduced by DHA treatment (Fig. 1D and E), indicating that DHA
attenuated LPS-induced edema. In addition, it was identified that
DHA treatment significantly decreased the numbers of total cells,
macrophages and neutrophils in the BALF of mice with LPS-induced
ALI (Fig. 1F-H). It was also
revealed that DHA treatment significantly reduced MPO activity in
the lungs of mice with LPS-induced ALI (Fig. 1I). These results indicate that DHA
attenuates the lung injury induced by LPS in mice.
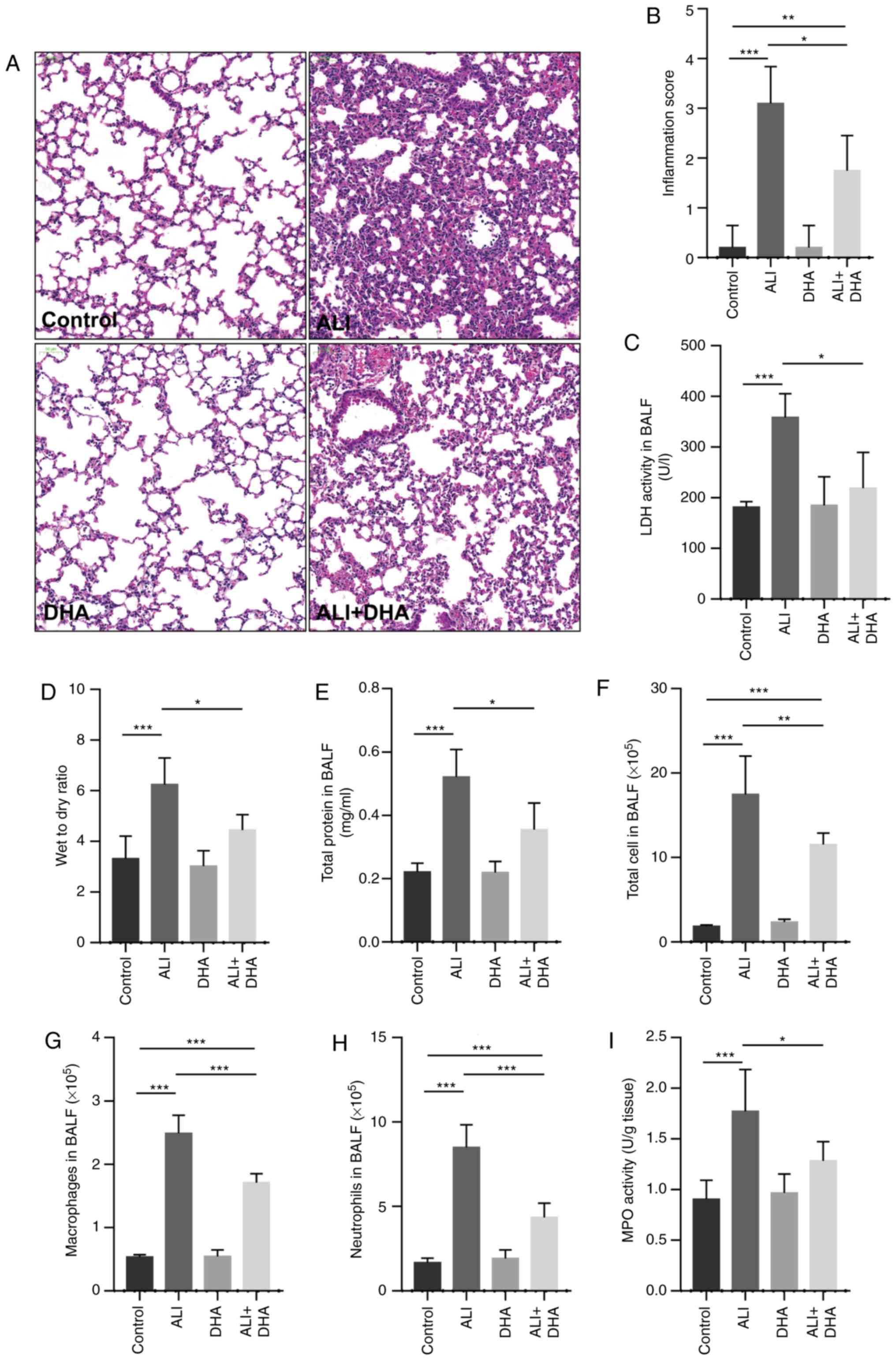 | Figure 1DHA attenuates lung tissue injury in
mice with LPS-induced ALI. Mice were treated with saline or DHA (75
mg/kg). After 1 h, mice were administered with LPS (5 mg/kg,
intratracheal) or saline for 12 h. (A) Lung histopathological
changes were detected by hematoxylin and eosin staining (scale bar,
100 µm) and (B) lung inflammation was scored (n=4-7). (C)
LDH activity in BALF was determined to assess lung tissue damage
(n=4-7). (D) Lung wet-to-dry ratio and (E) total protein in BALF
were measured to determine lung edema (n=6-9). The numbers of (F)
total cells, (G) macrophages and (H) neutrophils in BALF were
determined (n=5-7). (I) MPO activity in lung tissue was determined
(n=6-8). Data are expressed as the means ± standard error of the
mean. *P<0.05, **P<0.01,
***P<0.001. DHA, dihydroartemisinin; LPS,
lipopolysaccharide; ALI, acute lung injury; BALF, bronchoalveolar
lavage fluid; MPO, myeloperoxidase; LDH, lactate dehydrogenase. |
DHA reduces the inflammatory response and
oxidative stress in the lungs of LPS-exposed mice
IL-1β, TNF-α and IL-6 are pro-inflammatory cytokines
that are critical to the development of ALI (26). The results of the present study
demonstrated that mice with ALI exhibited a significant increase in
the mRNA levels of IL-1β, TNF-α and IL-6 in the lungs following LPS
administration; however, these effects were significantly
suppressed by DHA treatment (Fig.
2A-C). Accordingly, DHA reduced the protein levels of IL-1β,
TNF-α and IL-6 in the serum of mice with LPS-induced ALI (Fig. 2D-F). LPS induced a significant
increase in ROS generation and MDA content in the lungs, which was
signifi-cantly suppressed by DHA treatment (Fig. 2G and H). By contrast, SOD activity
and GSH content were decreased in the ALI group and were partially
restored by treatment with DHA (Fig.
2I and J). These results indicate that DHA protects against
LPS-induced lung injury by reducing inflammation and oxidative
stress.
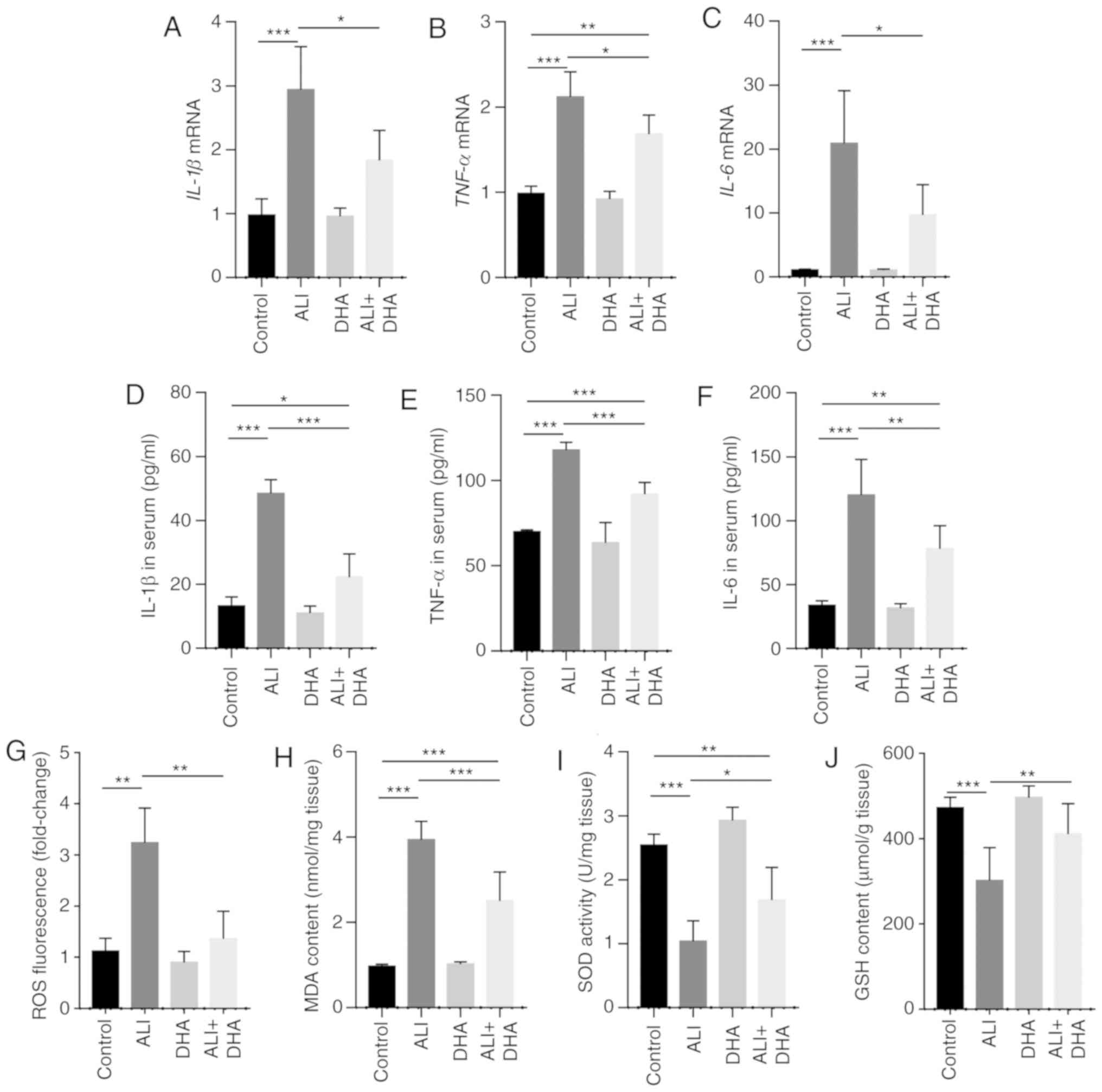 | Figure 2DHA reduces the inflammatory response
and oxidative stress in the lungs of LPS-administered mice. Mice
were treated with saline or DHA (75 mg/kg). After 1 h, mice were
treated with LPS (5 mg/kg, intratracheal) or saline for 12 h. (A)
IL-1β, (B) TNF-α and (C) IL-6 mRNA levels in the lungs were
determined by RT-qPCR (n=6, 7). (D) IL-1β, (E) TNF-α, and (F) IL-6
protein levels in the serum were determined by ELISA (n=4-6). (G)
ROS production, (H) MDA content, (I) SOD activity and (J) GSH
content in lung tissue were determined (n=6-8). Data are expressed
as the means ± standard error of the mean. *P<0.05,
**P<0.01, ***P<0.001. DHA,
dihydroartemisinin; LPS, lipopolysaccharide; ROS, reactive oxygen
species; MDA, malondialdehyde; SOD, superoxide dismutase; GSH,
glutathione; RT-qPCR, reverse transcription-quantitative polymerase
chain reaction. |
DHA inhibits inflammatory cytokine
release and oxidative stress induced by LPS in primary
macrophages
Treatment with DHA (20 µM) suppressed the
increase in the IL-1β, TNF-α and IL-6 mRNA expression levels in
primary macrophages and protein levels in the culture supernatant,
which were induced by LPS (100 ng/ml) (Fig. 3A-G). Furthermore, ROS generation
and MDA levels were increased in the macrophages treated with LPS,
whereas these responses were significantly suppressed by DHA
(Fig. 3H and I). In addition, DHA
treatment partially restored the decreased SOD activity and GSH
content in macrophages exposed to LPS (Fig. 3J and K). Collectively, these
findings suggest that DHA reduces the LPS-induced inflammatory
response and oxidative stress in primary macrophages.
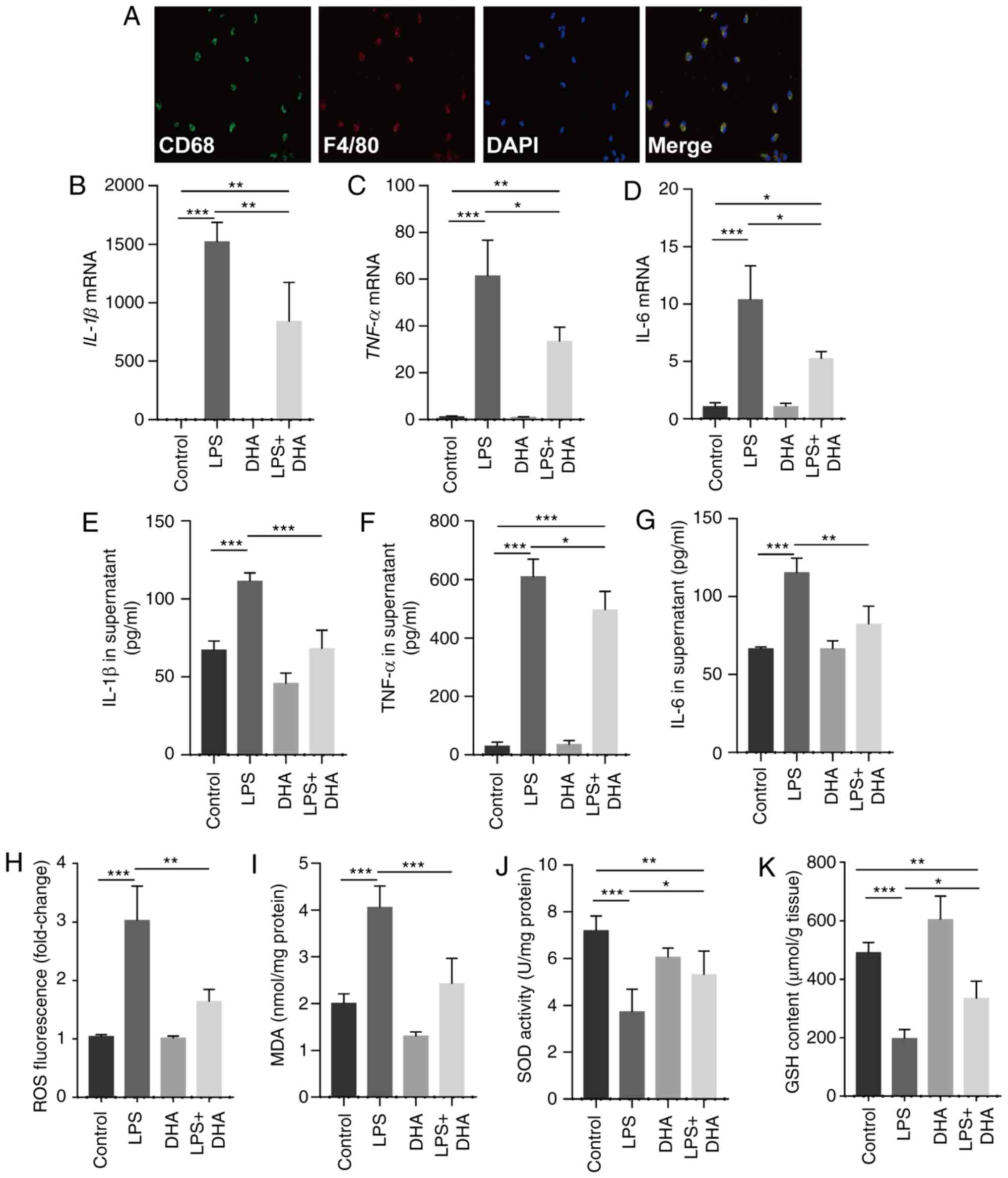 | Figure 3DHA inhibits pro-inflammatory
cytokine release and oxidative stress induced by LPS in primary
macrophages. LPS (100 ng/ml)-treated primary macrophages were
treated with DHA (20 µM) for 6 h. (A) Representative images
of immunofluorescence analyses performed on primary macrophages
using anti-CD68 (green) and anti-F4/80 (red) antibodies. The mRNA
expression levels of (B) IL-1β, (C) TNF-α and (D) IL-6 in primary
macrophages were determined (n=4-6). (E) IL-1β, (F) TNF-α and (G)
IL-6 protein levels in the culture supernatant of primary
macrophages were determined (n=4-6). (H) ROS production, (I) MDA
content, (J) SOD activity and (K) GSH content in primary
macrophages were determined (n=4-6). Data are expressed as the
means ± standard error of the mean. *P<0.05,
**P<0.01, ***P<0.001. DHA,
dihydroartemisinin; LPS, lipopolysaccharide; ROS, reactive oxygen
species; MDA, malondialdehyde; SOD, superoxide dismutase; GSH,
glutathione. |
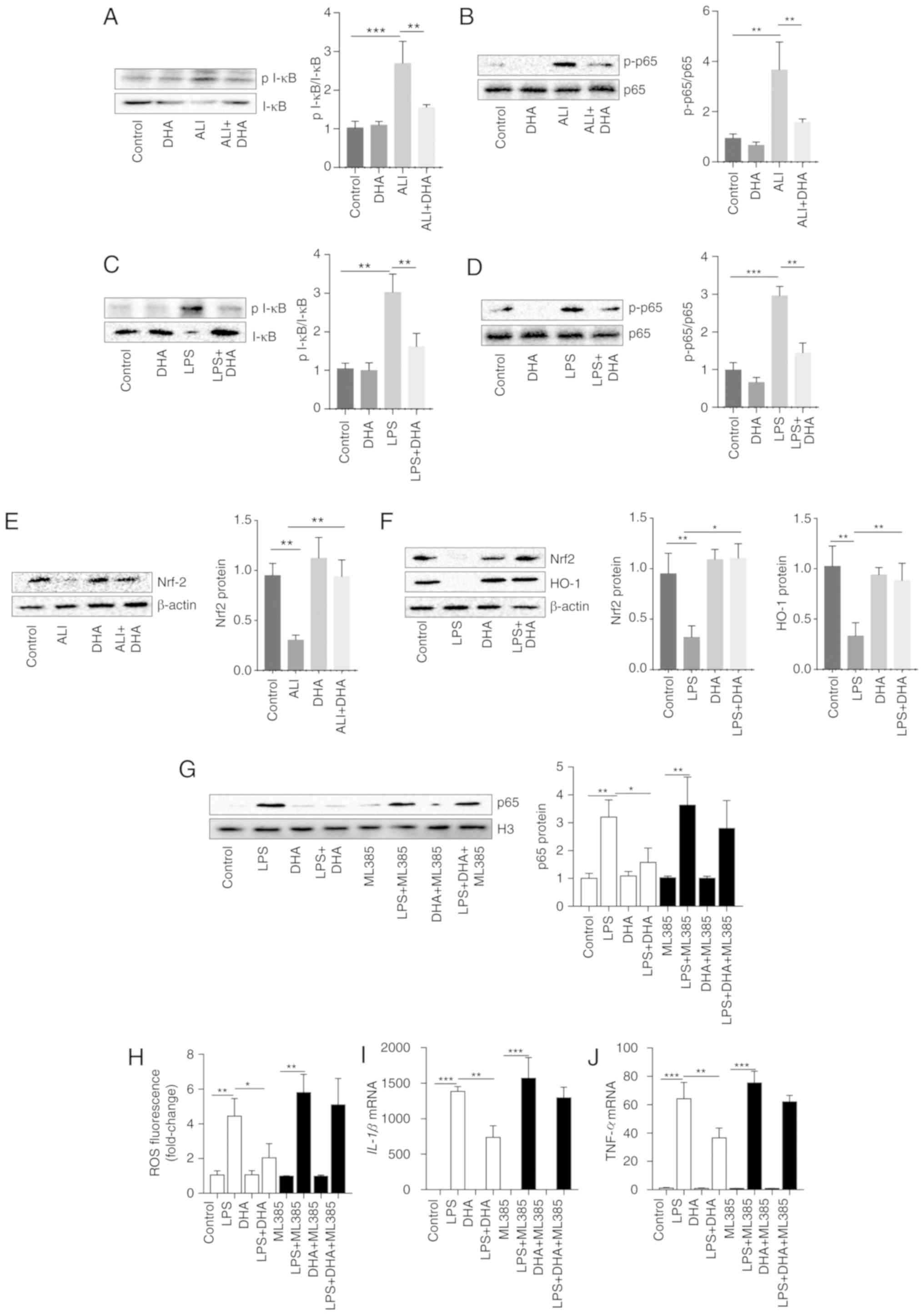
Inhibition of the NF-κB pathway by DHA is
dependent on Nrf2 in primary macrophages
DHA inhibited I-κB degradation and reduced the
increase in p-p65 expression induced by LPS in the lung tissue of
mice and primary macrophages (Fig.
4A-D). As shown in Fig. 4E and
F, DHA activated Nrf2 and HO-1, which were suppressed by LPS in
lung tissue and macrophages. To determine the role of Nrf2 in the
protective effects of DHA, Nrf2 was blocked in primary macrophages
using the Nrf2 inhibitor, ML385. It was identified that the
inhibition of NF-kB/p65 translocation by DHA was markedly
diminished by ML385 treatment (Fig.
4G). Furthermore, the LPS-induced increase in ROS production,
and in the IL-1β and TNF-α mRNA levels was not suppressed by DHA in
primary macrophages pre-treated with ML385 (Fig. 4H-J). These results indicate that
Nrf2 is vital for the protective effects of DHA against LPS-induced
ALI.
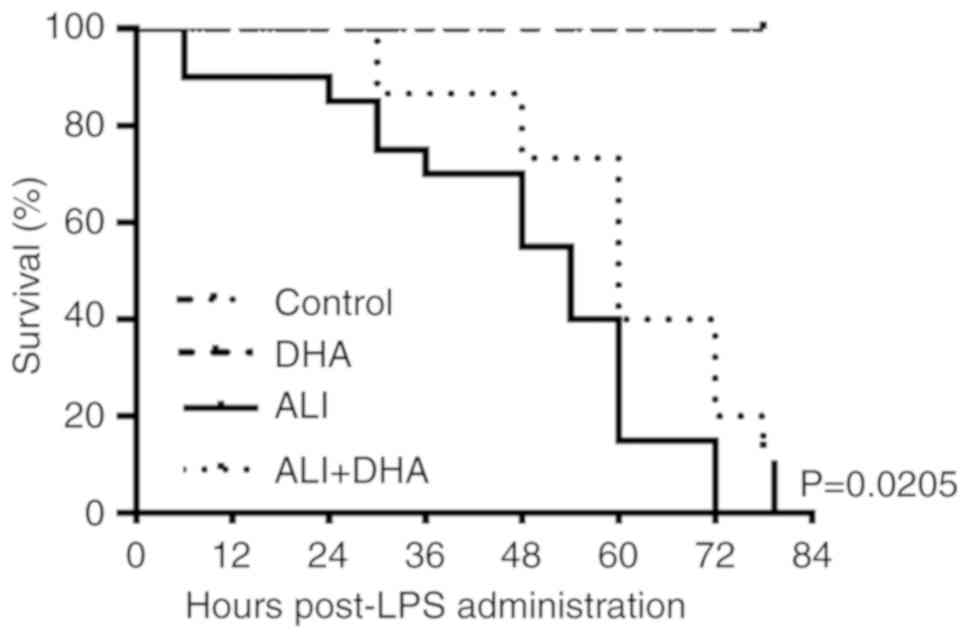
Therapeutic effect of DHA on LPS-induced
ALI mice
Clinically, pharmacotherapy does not commence until
there is an approved diagnosis of ALI. In this study, the
therapeutic effects of DHA on mice with ALI were subsequently
examined. It was identified that treatment with DHA 2 h
post-exposure to LPS significantly improved the survival rate of
mice with LPS-induced ALI (Fig.
5). This result suggests a therapeutic effect of DHA against
ALI in mice.
Discussion
Arteminsinin and its derivative, artesunate, have
been reported to exert protective effects against lung inflammation
(10,11), which indicates the potential use
of arteminsinin and its derivative in therapy. As a more effective
agent than artesunate, DHA has been reported to exert
anti-inflammatory and anti-fibrotic effects against
bleomycin-induced pulmonary fibrosis in rats (27). The present study first reported
that DHA attenuated lung tissue injury in a murine model of
LPS-induced ALI and suppressed macrophage activation induced by
LPS. Notably, it was identified that treatment with DHA 2 h
post-exposure to LPS significantly improved the survival rate of
LPS-exposed mice, indicating a therapeutic effect of DHA against
ALI. Mechanistically, it was determined that the DHA-mediated
suppression of inflammatory injury was dependent on Nrf2. The
present findings suggest the use of DHA as a potential therapeutic
agent for patients with ALI in the future.
Nrf2 plays a critical role in the regulation of
oxida-tive stress, which is the key pathogenic mechanism of ALI
(28-30). Generally, Nrf2 exists as a complex
with Keap1 in the cytoplasm. When cells are sensitized to ROS, Nrf2
is released from the complex and translocates to the nucleus,
promoting the expression of antioxidants, such as HO-1 and SOD
(31). A deficiency in Nrf2
results in severe lung injury induced by ischemia-reperfusion or
LPS (9,32). The present study also demonstrated
that Nrf2 expression was downregulated in the lungs of mice with
ALI. Furthermore, DHA increased the expression of downstream Nrf2,
SOD and HO-1, indicating the activation of Nrf2. Additionally,
inhibition of Nrf2 abolished the protective effects of DHA,
indicating a role of Nrf2 in the therapeutic effects of DHA. The
Nrf2/ARE signaling pathway also regulates the expression of
anti-inflammatory genes and inhibits the progression of
inflammation (33). Nrf2
negatively regulates LPS-induced NF-κB signaling activation
(29). The present study
demonstrated that Nrf2 blockade markedly diminished the inhibition
of NF-κB/p65 translocation induced by DHA in macrophages, which
indicated that DHA inhibits the NF-κB pathway in a Nrf2-dependent
manner in macrophages.
The NF-κB pathway is a key target for the
development of anti-inflammatory agents (34). Numerous natural products have been
screened for anti-inflammatory activities by inhibiting NF-κB
(35-37). Artemisinin significantly inhibits
NF-κB activation by suppressing the phosphorylation and degradation
of I-κBα and p65 nuclear translocation (38). Artesunate has been reported to
suppress LPS-induced TLR4 expression and NF-κB activation in lung
tissue and to upregulate Nrf2 and HO-1 expression in lung tissue
in vivo (11). However,
whether DHA can affect the activation of NF-κB and the underlying
mechanisms in macrophages remain unclear. The present study first
reported that DHA significantly mitigated NF-κB pathway activation
in the lungs of ALI mice and in primary macrophages exposed to LPS.
It has also been reported that DHA inhibits the NF-κB pathway in
rat chondrocytes (39) and tumor
cell invasion (40). While the
exact mechanisms remain unclear, the present study provides a novel
mechanism through which DHA inhibits the NF-κB pathway by
activating Nrf2. This indicates that DHA is a potential
anti-inflammatory and anti-oxidative agent.
Macrophages are the principal immune cells of
inflammatory molecules in pulmonary tissue and exert a vital
function in the molecular mechanisms of ALI, triggering
inflammation reactions and boosting the infiltration of neutrophils
(3). There is increasing evidence
to suggest that macrophages, which act as the first line of defense
in the lungs, are key factors in the pathogenesis of ALI (41). The depletion of macrophages has
been found to mitigate lung injury significantly at 4 h following
the administration LPS in mice by attenuating neutrophilic
alveolitis and reducing pro-inflammatory cytokines (42). Under the LPS challenge, the
pro-inflammatory M1 alveolar macrophages are mainly derived from
the bone marrow. Those alveolar macrophages are the triggers of the
uncontrolled inflammatory response during ALI. However, it is hard
to harvest a sufficient amount of alveolar macrophages from healthy
mice to conduct an experiment. In this study, the primary
peritoneal macrophages were recruited to the peritoneal cavity by
3% thioglycolate. Thus, these macrophages are also derived from
bone marrow. Notably, the adoptive transfer of peritoneal
macrophages into the lungs results in the expression of certain
alveolar macrophage-specific genes (43). In some studies, primary peritoneal
macrophages are used to investigate the role of macrophages in
lungs (19,44-46). The present study focused on the
role of DHA in the LPS-challenged inflammatory response in
macrophages in vitro. It was found that DHA inhibited
inflammatory cytokine release and oxidative stress induced by LPS
in primary peritoneal macrophages. Collectively, we hypothesized
that primary murine peritoneal macrophages share, at least partly,
the response to LPS-challenge with alveolar macrophages.
The limitations of the present were the following:
First, only the protective effects of DHA against LPS-induced ALI
in mice were examined. To further clarify the effects of DHA on
ALI, the protective effects of DHA in other models of ALI should
also be investigated. Second, the mechanisms underlying the
suppression of ALI by DHA are not completely clear. In addition to
the NF-κB pathway, the effects of DHA on the activation of the
NLRP3 inflammasome should be examined, which is a vital mechanism
underlying the uncontrolled inflammation during ALI (47). Artesunate has been identified to
alleviate renal ischemia-reperfusion-induced lung inflammation by
attenuating the activation of NLRP3 inflammasome (10).
In conclusion, this study demonstrates that DHA
exerts protective and therapeutic effects against LPS-induced ALI
by inhibiting the NF-κB signaling pathway in a Nrf2-dependent
manner. The present findings provide further evidence that DHA may
be a valuable therapeutic candidate for use in the treatment of
ALI.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81370974, 81500056
and 81500065) and the Hunan Provincial Natural Science Foundation
of China (grant no. 2019JJ50785).
Availability of data and materials
The data used to support the findings of this study
are presented in the present study or are available from the
corresponding author upon request.
Authors' contributions
XTH designed and performed most of the experiments,
analyzed and interpreted the data, and wrote the manuscript. WL,
CXH, YaZ, CYZ, and CCS assisted during the acquisition, analysis,
and interpretation of data and revised the manuscript. ZQL and YoZ
assisted with data acquisition and revision of the manuscript. SYT
performed experiments and prepared the manuscript. All authors have
reviewed and approved the final version of the manuscript.
Ethics approval and consent to
participate
The Ethics Committee of the Center for Scientific
Research with Animal Models at Central South University (Changsha,
China) approved the experiments, which were performed in accordance
with the guidelines of the National Institutes of Health.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Pan C, Liu L, Xie JF and Qiu HB: Acute
respiratory distress syndrome: Challenge for diagnosis and therapy.
Chin Med J (Engl). 131:1220–1224. 2018. View Article : Google Scholar
|
|
2
|
Han X, Wu YC, Meng M, Sun QS, Gao SM and
Sun H: Linarin prevents LPS-induced acute lung injury by
suppressing oxidative stress and inflammation via inhibition of
TXNIP/NLRP3 and-NF-κB pathways. Int J Mol Med. 42:1460–1472.
2018.PubMed/NCBI
|
|
3
|
Wu J, Yan Z, Schwartz DE, Yu J, Malik AB
and Hu G: Activation of NLRP3 inflammasome in alveolar macrophages
contributes to mechanical stretch-induced lung inflammation and
injury. J Immunol. 190:3590–3599. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu H, Liang X, Wang D, Zhang H, Liu L,
Chen H, Li Y, Duan Q and Xie K: Combination therapy with nitric
oxide and molecular hydrogen in a murine model of acute lung
injury. Shock. 43:504–511. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Takashima K, Matsushima M, Hashimoto K,
Nose H, Sato M, Hashimoto N, Hasegawa Y and Kawabe T: Protective
effects of intratracheally administered quercetin on
lipopolysaccha-ride-induced acute lung injury. Respir Res.
15:1502014. View Article : Google Scholar
|
|
6
|
Deng G, He H, Chen Z, OuYang L, Xiao X, Ge
J, Xiang B, Jiang S and Cheng S: Lianqinjiedu decoction attenuates
LPS-induced inflammation and acute lung injury in rats via
TLR4/NF-κB pathway. Biomed Pharmacother. 96:148–152. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jiang K, Guo S, Yang C, Yang J, Chen Y,
Shaukat A, Zhao G, Wu H and Deng G: Barbaloin protects against
lipopolysaccharide (LPS)-induced acute lung injury by inhibiting
the ROS-mediated PI3K/AKT/NF-κB pathway. Int Immunopharmacol.
64:140–150. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Walters DM, Cho HY and Kleeberger SR:
Oxidative stress and antioxidants in the pathogenesis of pulmonary
fibrosis: A potential role for Nrf2. Antioxid Redox Signal.
10:321–332. 2008. View Article : Google Scholar
|
|
9
|
Yan J, Li J, Zhang L, Sun Y, Jiang J,
Huang Y, Xu H, Jiang H and Hu R: Nrf2 protects against acute lung
injury and inflammation by modulating TLR4 and Akt signaling. Free
Radic Biol Med. 121:78–85. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu Z, Qu M, Yu L, Song P and Chang Y:
Artesunate inhibits renal ischemia-reperfusion-mediated remote lung
inflammation through attenuating ROS-induced activation of NLRP3
inflammasome. Inflammation. 41:1546–1556. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao D, Zhang J, Xu G and Wang Q:
Artesunate protects LPS-induced acute lung injury by inhibiting
TLR4 expression and inducing Nrf2 activation. Inflammation.
40:798–805. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Keating GM:
Dihydroartemisinin/Piperaquine: A review of its use in the
treatment of uncomplicated Plasmodium falciparum malaria. Drugs.
72:937–961. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Morris CA, Onyamboko MA, Capparelli E,
Koch MA, Atibu J, Lokomba V, Douoguih M, Hemingway-Foday J, Wesche
D, Ryder RW, et al: Population pharmacokinetics of artesunate and
dihydroartemisinin in pregnant and non-pregnant women with malaria.
Malar J. 10:1142011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tu Y: The discovery of artemisinin
(qinghaosu) and gifts from Chinese medicine. Nat Med. 17:1217–1220.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang DX, Qiu J, Zhou HH, Yu Y, Zhou DL, Xu
Y, Zhu MZ, Ge XP, Li JM, Lv CJ, et al: Dihydroartemisinin
alleviates oxidative stress in bleomycin-induced pulmonary
fibrosis. Life Sci. 205:176–183. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin SP, Li W, Winters A, Liu R and Yang
SH: Artemisinin prevents glutamate-induced neuronal cell death via
Akt pathway activation. Front Cell Neurosci. 12:1082018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jiang C, Li S, Li Y and Bai Y: Anticancer
effects of dihydroartemisinin on human esophageal cancer cells in
vivo. Anal Cell Pathol (Amst). 2018:87597452018.
|
|
18
|
Zhou Y, Liu T, Duan JX, Li P, Sun GY, Liu
YP, Zhang J, Dong L, Lee KSS, Hammock BD, et al: Soluble epoxide
hydrolase inhibitor attenuates lipopolysaccharide-induced acute
lung injury and improves survival in mice. Shock. 47:638–645. 2017.
View Article : Google Scholar :
|
|
19
|
Zhong WJ, Yang HH, Guan XX, Xiong JB, Sun
CC, Zhang CY, Luo XQ, Zhang YF, Zhang J, Duan JX, et al: Inhibition
of glycol-ysis alleviates lipopolysaccharide-induced acute lung
injury in a mouse model. J Cell Physiol. 234:4641–4654. 2019.
View Article : Google Scholar
|
|
20
|
Kim KH, Kwun MJ, Choi JY, Ahn KS, Oh SR,
Lee YG, Christman JW, Sadikot RT, Han CW and Joo M: Therapeutic
effect of the tuber of alisma orientale on
lipopolysaccha-ride-induced acute lung injury. Evid-Based
Complement Alternat Med. 2013:8638922013.
|
|
21
|
Suzuki K, Okada H, Takemura G, Takada C,
Kuroda A, Yano H, Zaikokuji R, Morishita K, Tomita H, Oda K, et al:
Neutrophil elastase damages the pulmonary endothelial glycocalyx in
lipopolysaccharide-induced experimental endotoxemia. Am J Pathol.
189:1526–1535. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mahapatra S, Ying L, Ho PP, Kurnellas M,
Rothbard J, Steinman L and Cornfield DN: An amyloidogenic
hexapeptide derived from amylin attenuates inflammation and acute
lung injury in murine sepsis. PLoS One. 13:e01992062018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ueda J, Starr ME, Takahashi H, Du J, Chang
LY, Crapo JD, Evers BM and Saito H: Decreased pulmonary
extracellular superoxide dismutase during systemic inflammation.
Free Radic Biol Med. 45:897–904. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wolthuis EK, Vlaar AP, Choi G, Roelofs JJ,
Juffermans NP and Schultz MJ: Mechanical ventilation using
non-injurious ventilation settings causes lung injury in the
absence of pre-existing lung injury in healthy mice. Crit Care.
13:R12009. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
26
|
Janz DR and Ware LB: Biomarkers of
ALI/ARDS: Pathogenesis, discovery, and relevance to clinical
trials. Semin Respir Crit Care Med. 34:537–548. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang D, Yuan W, Lv C, Li N, Liu T, Wang L,
Sun Y, Qiu X and Fu Q: Dihydroartemisinin supresses inflammation
and fibrosis in bleomycine-induced pulmonary fibrosis in rats. Int
J Clin Exp Pathol. 8:1270–1281. 2015.PubMed/NCBI
|
|
28
|
Zhang HX, Liu SJ, Tang XL, Duan GL, Ni X,
Zhu XY, Liu YJ and Wang CN: H2S attenuates LPS-induced acute lung
injury by reducing oxidative/nitrative stress and inflammation.
Cell Physiol Biochem. 40:1603–1612. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fan L, Fan Y, Liu L, Tao W, Shan X, Dong
Y, Li L, Zhang S and Wang H: Chelerythrine attenuates the
inflammation of lipopolysaccharide-induced acute lung inflammation
through NF-κB signaling pathway mediated by Nrf2. Front Pharmacol.
9:10472018. View Article : Google Scholar
|
|
30
|
Liu Q, Lv H, Wen Z, Ci X and Peng L:
Isoliquiritigenin activates nuclear factor erythroid-2 related
factor 2 to suppress the NOD-like receptor protein 3 inflammasome
and inhibits the NF-κB pathway in macrophages and in acute lung
injury. Front Immunol. 8:15182017. View Article : Google Scholar
|
|
31
|
Yang Y, Willis TL, Button RW, Strang CJ,
Fu Y, Wen X, Grayson PRC, Evans T, Sipthorpe RJ, Roberts SL, et al:
Cytoplasmic DAXX drives SQSTM1/p62 phase condensation to activate
Nrf2-mediated stress response. Nat Commun. 10:37592019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lv H, Liu Q, Wen Z, Feng H, Deng X and Ci
X: Xanthohumol ameliorates lipopolysaccharide (LPS)-induced acute
lung injury via induction of AMPK/GSK3β Nrf2 signal axis. Redox
Biol. 12:311–324. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ahmed SM, Luo L, Namani A, Wang XJ and
Tang X: Nrf2 signaling pathway: Pivotal roles in inflammation.
Biochim Biophysica Acta Mol Basis Dis. 1863:585–597. 2017.
View Article : Google Scholar
|
|
34
|
Shang L, Wang T, Tong D, Kang W, Liang Q
and Ge S: Prolyl hydroxylases positively regulated LPS-induced
inflammation in human gingival fibroblasts via TLR4/MyD88-mediated
AKT/NF-κB and MAPK pathways. Cell Prolif. 51:e125162018. View Article : Google Scholar
|
|
35
|
Huang XT, Li C, Peng XP, Guo J, Yue SJ,
Liu W, Zhao FY, Han JZ, Huang YH, Yang Li, et al: An excessive
increase in glutamate contributes to glucose-toxicity in β-cells
via activation of pancreatic NMDA receptors in rodent diabetes. Sci
Rep. 7:441202017. View Article : Google Scholar
|
|
36
|
Zhang D, Li X, Hu Y, Jiang H, Wu Y, Ding
Y, Yu K, He H, Xu J, Sun L and Qian F: Tabersonine attenuates
lipopolysac-charide-induced acute lung injury via suppressing TRAF6
ubiquitination. Biochem Pharmacol. 154:183–192. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Seo EJ, Fischer N and Efferth T:
Phytochemicals as inhibitors of NF-κB for treatment of Alzheimer's
disease. Pharmacol Res. 129:262–273. 2018. View Article : Google Scholar
|
|
38
|
Wang KS, Li J, Wang Z, Mi C, Ma J, Piao
LX, Xu GH, Li X and Jin X: Artemisinin inhibits inflammatory
response via regulating NF-κB and MAPK signaling pathways.
Immunopharmacol Immunotoxicol. 39:28–36. 2017. View Article : Google Scholar
|
|
39
|
Jiang LB, Meng DH, Lee SM, Liu SH, Xu QT,
Wang Y and Zhang J: Dihydroartemisinin inhibits catabolism in rat
chon-drocytes by activating autophagy via inhibition of the NF-κB
pathway. Sci Rep. 6:389792016. View Article : Google Scholar
|
|
40
|
Hwang YP, Yun HJ, Kim HG, Han EH, Lee GW
and Jeong HG: Suppression of PMA-induced tumor cell invasion by
dihydroartemisinin via inhibition of PKCalpha/Raf/MAPKs and
NF-kappaB/AP-1-dependent mechanisms. Biochem Pharmacol.
79:1714–1726. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Huang X, Xiu H, Zhang S and Zhang G: The
Role of macrophages in the pathogenesis of ALI/ARDS. Mediators
Inflamm. 2018:12649132018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Koay MA, Gao X, Washington MK, Parman KS,
Sadikot RT, Blackwell TS and Christman JW: Macrophages are
necessary for maximal nuclear factor-kappa B activation in response
to endotoxin. Am J Respir Cell Mol Biol. 26:572–578. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Morales-Nebreda L, Misharin AV, Perlman H
and Budinger GR: The heterogeneity of lung macrophages in the
susceptibility to disease. Eur Respir Rev. 24:505–509. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chen L, Jin Y, Chen H, Sun C, Fu W, Zheng
L, Lu M, Chen P, Chen G, Zhang Y, et al: Discovery of caffeic acid
phenethyl ester derivatives as novel myeloid differentiation
protein 2 inhibitors for treatment of acute lung injury. Eur J Med
Chem. 143:361–375. 2018. View Article : Google Scholar
|
|
45
|
Duan JX, Zhou Y, Zhou AY, Guan XX, Liu T,
Yang HH, Xie H and Chen P: Calcitonin gene-related peptide exerts
anti-inflammatory property through regulating murine macrophages
polarization in vitro. Mol Immunol. 91:105–113. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Qi Z, Qi S, Ling L, Lv J and Feng Z:
Salidroside attenuates inflammatory response via suppressing
JAK2-STAT3 pathway activation and preventing STAT3 transfer into
nucleus. Int Immunopharmacol. 35:265–271. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hosseinian N, Cho Y, Lockey RF and
Kolliputi N: The role of the NLRP3 inflammasome in pulmonary
diseases. Ther Adv Respir Dis. 9:188–197. 2015. View Article : Google Scholar : PubMed/NCBI
|