Introduction
Psoriasis is one of the most chronic inflammatory
hypertrophic skin diseases, affecting 2-5% of the population
worldwide (1,2). Multiple factors may cause psoriasis,
including genetics, epigenetic modifications, environmental
triggers and immune dysfunction (3,4).
Although there are extensive studies available regarding the
etiologies and mechanisms underlying psoriasis, the precise
molecular mechanisms remain elusive (5). Therefore, crucial molecular markers
of psoriasis that are identifiable with more powerful technologies
are urgently required.
The methylation and hydroxymethylation of DNA are
the complementary processes of epigenetic regulation; these
epigenetic alterations are responsible for the modulation of gene
expression (6). Ten-eleven
translocation (TET)2 is a key methylation-regulating enzyme that
can alter the DNA methylation status by converting 5-methylcytosine
(5-mC) to 5-hydroxymethylcytosine (5-hmC) and can regulate the
expression of certain genes (7).
It has previously been reported that TET2 plays an important role
in biological processes, involving cell differentiation and
proliferation (8). In a recent
study, TET2 was also demonstrated to be involved in the progression
of psoriatic dermatitis in mice by regulating hydroxymethylation
(9). Although some methylation
microarrays have been performed to identify differentially
methylated genes (DMGs) in psoriasis (10), the comprehensive profile network
analysis remains largely elusive.
Gene microarray is a high-throughput gene detection
technique that has been widely used for the investigation of
differentially expressed genes (DEGs), and is an important method
for studying the pathogenesis of a variety of diseases (11). In recent years, a number of
microarray gene profile studies on psoriasis have been performed.
Using microarray data, the gene expression profiles in psoriasis
have been disclosed and described (12). Certain studies have integrated
analyses of gene expression microarray and methylation microarray
by bioinformatics analysis in different types of cancer (13,14). However, to the best of our
knowledge, there are currently no systematic studies available
focusing on mining expression profiles and methylated DEGs in
psoriasis. The integrated bioinformatics analysis will be more
reliable and provide valuable new molecular targets to foster the
development of accurate diagnosis and novel therapeutic strategies.
The present study downloaded original data from the Gene Expression
Omnibus (GEO; https://www.ncbi.nlm.nih.gov/geo/). The integrated
gene expression profile and DNA methylation profile microarrays of
psoriasis were analyzed using a series of packages in R in order to
identify the DEGs. Subsequently, The Database for Annotation,
Visualization and Integrated Discovery (DAVID, https://david.ncifcrf.gov/), the Kyoto Encyclopedia of
Genes and Genomes (KEGG, http://www.kegg.jp/), the Search Tool for the
Retrieval of Interacting Genes (STRING https://string-db.org/) and Cytoscape were used to
analyze the functional enrichment, protein-protein interaction
(PPI) network, cluster module and hub genes. For further
verification of the association of DMGs with epigenetic regulators
in vivo, the present study created an animal model of
imiquimod (IMQ)-induced psoriasiform dermatitis and knocked down
TET2 expression. The results of the present study provide
additional potential targets for the diagnosis and therapeutic
intervention in patients with psoriasis.
Materials and methods
Data collection
Data on gene expression profiles (GSE13355, GSE14905
and GSE78097) and gene methylation profiles (GSE31835) were
downloaded from the GEO database of The National Center for
Biotechnology Information (NCBI). The 3 selected gene expression
profile microarrays were based on the same platform (GPL570), which
can decrease batch differences in the integration analysis.
GSE31835 was based on the GPL8490 platform. GSE13355 contained the
data from 58 skin lesions of patients with psoriasis and 64 normal
healthy controls; GSE14905 contained 33 samples from patients with
psoriasis and 21 normal control samples; GSE78097 contained 13
samples from patients with psoriasis and 6 normal control samples;
19 samples from lesions of patients with psoriasis and 10 normal
skin samples were contained in GSE31835.
Data preprocessing and the identification
of DEGs
The new versions of affymetrix annotation files were
download from the official affymetrix website. The raw data of the
gene expression profile were pre-processed by the Robust
Multi-array Average (RMA) algorithm with the use of the affy
package in Bioconductor (https://www.bioconductor.org/) for background
correction and normalization (15). Linear models for the microarray
data and corrections for the P-values were calculated using the
limma package in Bioconductor (16). |log2FC|>1 and
P<0.05 were set as the cut-off criteria to indicate statistical
significance. The methylation profile microarray GSE31835 was
analyzed using GEO2R software. The DMGs were identified with a
P-value cut-off of <0.05 and |t| >2-fold. Subsequently, a
principal component analysis (PCA) and hierarchical cluster
analysis were performed with the scatterplot3d package and gplots
package in R software.
Functional enrichment analysis
The DAVID provides gene analysis tools that link
genomic information to biological annotations in order to help
users identify the most valuable and significant annotations and
links (17). DAVID was used for
the analysis of DEGs of the KEGG pathway enrichment and Gene
Ontology (GO) functional analysis, including cellular components,
molecular function and biological processes (18). P<0.05 and counts ≥2 were used
as the significance threshold.
PPI network construction and module
analysis
STRING is an online tool for searching for known
proteins and predicting the interaction between proteins (19). DEGs were mapped into STRING, and a
PPI network was established by calculating a combined score of
>0.4. The PPI network was constructed with Cytoscape (version
3.5.1) software. Significant modules were selected based on MCODE,
with scores >4 and the number of nodes >5 in Cytoscape. The
hub genes were identified according to the 12 topological analysis
methods of cytoHubba (20).
Animal procedures
A total of 72 female BALB/c mice (weighing 18-20 g)
were purchased from Beijing Vital River Laboratory Animal
Technology Co., Ltd. The animals were housed in a controlled
environment (23±2°C, 12-h light/dark cycle) with sufficient food
and water at The Animal Centre of Tongji Medical College.
Experimental mice were 8-10 weeks of age. All experimental
protocols with animals were conducted following the ARRIVE
guidelines. All mouse care and procedures were approved by the
Ethics Committee for experimental animals, Tongji Medical College
(no. SYXK HUBEI 2016-0057).
The BALB/c mice were randomly divided into 3 groups
as follows and the mice in all groups were shaved on their backs
(average area of 2×3 cm): i) The control group (n=8), mice were
topically treated with vaseline and received an intradermal
injection of lentivirus-delivered scrambled RNAs (shRNA-scr) on
days 1 and 4; ii) the IMQ group (n=8), skin inflammation was
induced by IMQ treatment, as previously reported (21,22). Briefly, mice in the IMQ group
received a daily topical dose of 62.5 mg of 5% IMQ cream on their
shaved backs for 7 consecutive days and received injections of the
same volume of shRNA-scr; the IMQ + sh-TET2 group (n=8), mice
received intradermal injections of lentivirus-delivered shRNA
against TET2 (sh-TET2) followed by the application of IMQ cream for
7 days. The lentivirus-delivered shRNAs were designed and obtained
from Shanghai Genechem Co., LTD. The sequences of the shRNAs were
5′-TTC TCC GAA CGT GTC ACG T-3′ (sh-sc), 5′-GAG CGT TCC TCA GTA TCA
TTT-3′ (sh-TET2). On day 8, following the application of 20%
urethane (1,500-2,000 mg/kg) (23), the dorsal skin lesions of the mice
were taken for use in further experiments. Anesthetic mice were
unresponsive to external stimuli, with the loss of righting and
palpebral reflexes, but had a constant heart rate and respiratory
rate (24). Subsequently, the
mice were euthanized by rapid decapitation. The experiments were
performed in triplicate.
Western blot analysis
The samples were lysed with cell lysis buffer (RIPA
Lysis Buffer; cat. no. P0013B; Beyotime), and the supernatant was
collected following centrifugation at 13,000 × g for 5 min at 4°C.
The concentrations of proteins were measured using a BCA protein
assay kit (cat. no. G2026; Servicebio). Equal quantities of protein
were separated via SDS-PAGE (stacking gel, 5%; separating gel,
8%-TET2, 12%-S100A9) and then transferred onto polyvinylidene
difluoride membranes. After blocking with 5% skim milk at room
temperature for 1 h, the membranes were incubated with primary
antibodies against GAPDH (1:1,000; cat. no. AF7021; Affinity
Biosciences), TET2 (1:1,000; cat. no. ab94580; Abcam) or S100A9
(1:1,000; cat. no. NB110-89726SS; Novus Biologicals) at 4°C
overnight, followed by the appropriate horseradish
peroxidase-conjugated secondary antibodies (anti-rabbit IgG,
HRP-linked antibody/anti-mouse IgG, HRP-linked antibody 1:2,000;
cat. no. 7074/7076; Cell Signaling Technology) for 1 h at room
temperature. Signals were detected using DNR bio-imaging systems
according to the manufacturer's protocol. The densitometric
quantification analysis of proteins was performed using ImageJ
software (Ver. 1.45s; National Institutes of Health).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the whole skin samples
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). cDNA was synthesized by a reverse-transcriptase
reaction with a PrimeScript RT Reagent kit (Takara Biotechnology).
The specific cDNA fragments were amplified using a Real-time PCR
Detection System and Power SYBR-Green (Takara Biotechnology) with
the relevant primers. The data are expressed as the relative
expression to the GAPDH gene. The relative expression level of each
target gene was analyzed using the 2−ΔΔCq method and
normalized to GAPDH (25). The
PCR primer sequences are presented in Table SI.
Hydroxymethylated DNA immunoprecipitation
(hmeDIP)
The extraction of genomic DNA was performed using
the SimpleDIP™ Hydroxymethylated DNA IP kit (Cell Signaling
Technology) according to the manufacturer's protocol. Genomic DNA
was fragmented to sub 100-500 bp using an Ultrasonic Homogenizer.
The 5-hmC mouse monoclonal antibody (1:1; cat. no. 51660, Cell
Signaling Technology) was added to the single stranded DNA samples.
The samples were incubated overnight at 4°C with rotation. After
washing and eluting, the immunoprecipitated DNA was purified using
spin columns. RT-qPCR was then utilized for the detection of
specific hydroxymethylated DNA levels. Quantitative analysis was
performed using the following formula: 10% × 2(C[T] 10% Input
Sample-C[T] IP Sample).
Statistical analysis
Comparisons between groups were performed using an
unpaired two-tailed Student's t-test. In some analyses, one-way
analysis of variance (ANOVA) was used, as indicated. Tukey's post
hoc test was used for multiple comparisons among diverse groups.
Data are expressed as the means ± standard deviation, and P<0.05
was considered to indicate a statistically significant difference.
GraphPad Prism software (version 6.0; GraphPad Software) was used
for statistical analysis.
Results
Identification of DEGs in psoriasis
Following quantification and normalization, DEGs
were identified based on a threshold of P<0.05 and fold change
>2. The present study screened 951, 1,530 and 2,836 DEGs in the
psoriasis samples compared with the normal samples in the GSE13355,
GSE14905 and GSE78097 datasets, respectively. Using GSE13355 as a
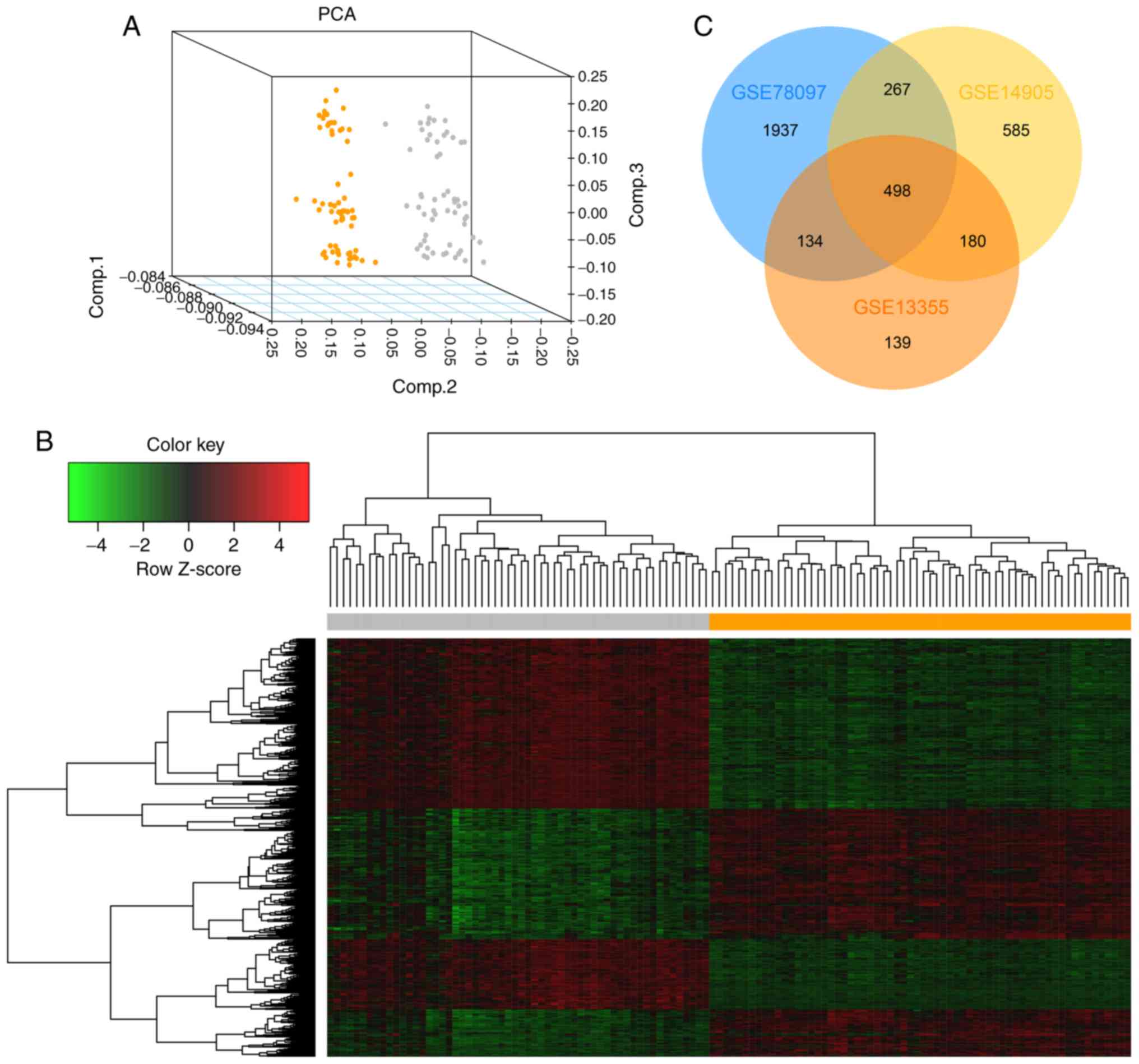
representative, the results of the PCA and hierarchical cluster
analysis of the DEGs are presented in Fig. 1A and B. The other two PCAs and
heatmaps are presented in Fig.
S1. In total, 498 common DEGs were screened out, including 352
upregulated, 137 downregulated and 9 non-specific genes (Fig. 1C; Table SII).
DEGs GO analysis and pathway enrichment
analysis
In order to investigate the biological functions of
the DEGs, the present study performed a functional pathway
enrichment analysis of the DEGs through GO analysis and KEGG
pathway enrichment in DAVID. A P<0.05 was considered to indicate
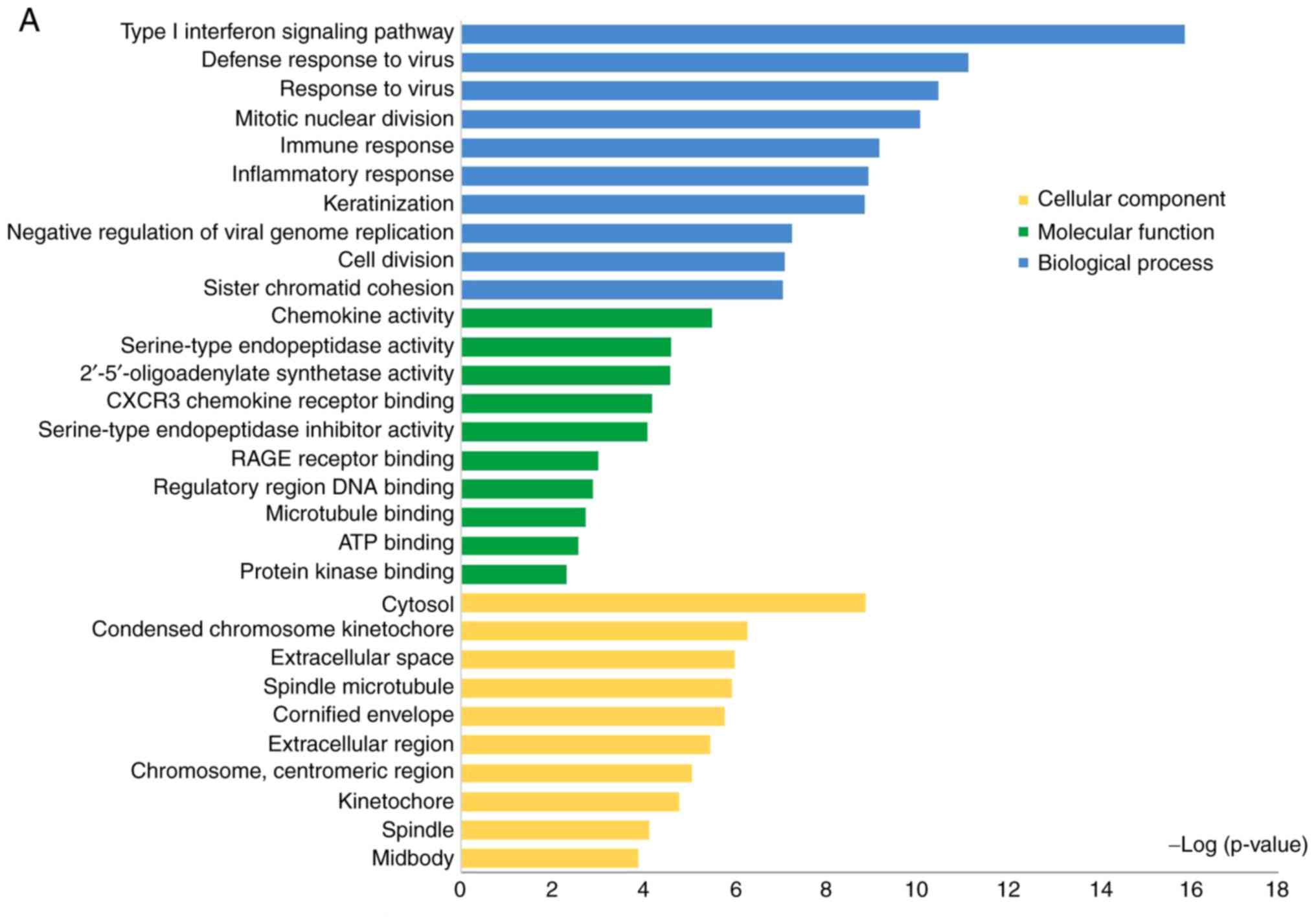
a statistically significant difference. As presented in Fig. 2A, in the biological processes
group, the upregulated DEGs were primarily involved in 'the type I
interferon signaling pathway', 'the defense response to virus',
'mitotic nuclear division', 'the immune response' and 'the
inflammatory response'; in the molecular function group, the
upregulated DEGs were significantly involved in 'chemokine
activity', 'serine-type endopeptidase activity', 'CXC motif
chemokine receptor (CXCR) 3 chemokine receptor binding', 'advanced
glycosylation end-product specific receptor (RAGE) receptor
binding' and 'regulatory region DNA binding'; in the cellular
component group, the upregulated DEGs were primarily involved in
'cytosol', 'condensed chromosome kinetochore', 'spindle
microtubule' and 'cornified envelope'. The downregulated DEGs were
primarily involved in 'cell adhesion', 'positive regulation of
protein kinase B signaling', 'RNA polymerase II transcription
factor binding', 'transcription factor binding' and 'extracellular
matrix' (Fig. 2B).
The significantly enriched KEGG pathways of
upregulated DEGs associated with psoriasis were the 'influenza A
pathway', 'chemokine signaling pathway', 'cell cycle pathway',
'RIG-I-like receptor signaling pathway' and 'p53 signaling
pathway'. The downregulated genes were primarily enriched in 'tight
junction pathways' (Fig. 2C).
Module analysis and hub gene selection
from the PPI network of DEGs
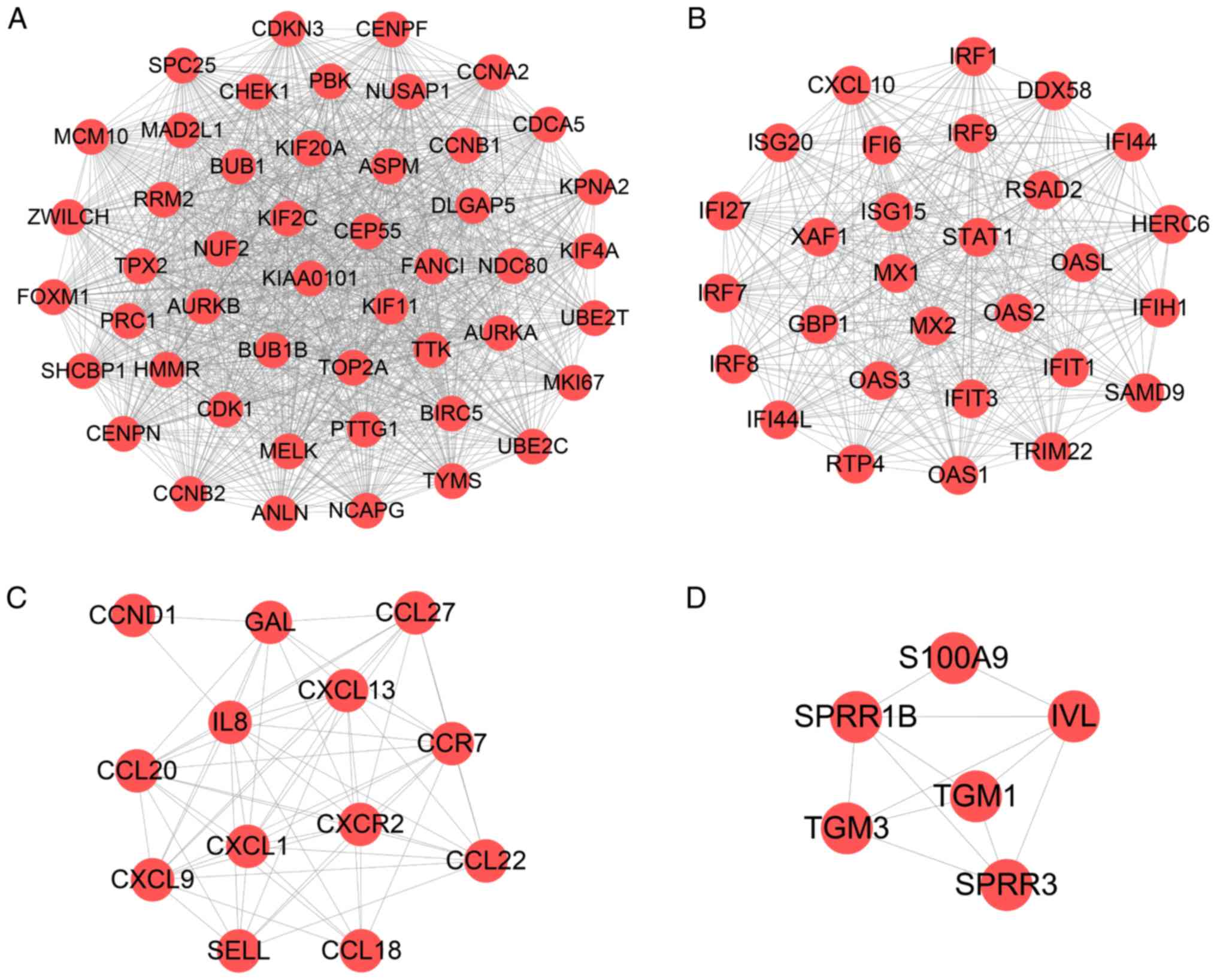
In order to screen out key modules and hub genes, a
PPI network of 475 DEGs was established with a combined score of
>0.4, including 475 nodes and 2,556 edges (Fig. S2), which was based on the STRING
database. A module analysis was performed using MCODE in Cytoscape.
MCODE scores >4 and nodes >5 were set as the cut-off
criteria. The 4 significant modules are presented in Fig. 3. Subsequently, the present study
analyzed the pathways involved in modules 1 and 2 using ClueGO, the
plugin of Cytoscape. The 'Cell cycle', 'NOD-like receptor signaling
pathway' and 'RIG-I-like receptor signaling pathway' were the
significantly enriched pathways (Fig. S3). The hub genes were screened by
cytoHubba in Cytoscape (data not shown) and it was revealed that
TOP2A, CDK1, IL8, KIAA0101, CCNB1, BIRC5, STAT3, TYMS, UBE2C,
ABCA12, TMPRSS11D and S100A9 may all play a pivotal role in the
molecular mechanisms underlying psoriasis.
Identification of abnormal
methylated-DEGs in psoriasis
The gene methylation microarray GSE31835 was
obtained from the public GEO database. Based on the cut-off
standards, a total of 2,730 DMGs was primitively identified between
the psoriasis samples and healthy controls. By the construction of
a Venn diagram (data not shown), the consistent differential genes
between DEGs and DMGs were obtained (Table I).
 | Table IAbnormal methylated-differentially
expressed genes in psoriasis. |
Table I
Abnormal methylated-differentially
expressed genes in psoriasis.
| Term | Gene name |
|---|
|
Methylated-differentially expressed
genes | TNIP3, XDH,
HSD17B2, HMGCS2, SELL, FOXM1, C10orf99, IFI27, PPP1R1B, ADRB1,
PI15, IRF7, CCND1, ZNF677, PI3, PDK4, CCR7, PDZK1IP1, IL19, ATP11B,
CRABP2, GALNT6, CHI3L2, FUT3, CLEC7A, GBP1, NFE2L3, OAS2, KRT16,
GZMB, FCGR3B, SMPD3, SLAMF7, LRRC8B, ZDHHC21, STS, ALDH1A3, LRG1,
CD47, RORC, S100A7, S100A12, PRELP, SERPINB3, IVL, APOC1, CYP4B1,
ARSF, TRIM22, SLC23A2, TIMP3, GJB6, APOL6, SLC2A13, FOXC1,
DNASE1L3, KLHDC7B, FADS2, ARNTL2, CCL27, ZBTB16, KYNU, CCL20,
CRISPLD1, TMEM40, CHAC1, LYN, BTC, TCN1, ID4, SLC7A5, GDPD3, GJB2,
GBP6, WNT2B, ISG20, CXCL1, CDK5R1, TMEM116, LYPD5, IL7R, PTAFR,
KLK10, ATP12A, SPRR3, CFB, AIM2, LCN2, KRT6B, UHRF1, MAMDC2,
KCNJ15, IFI44, S100A9, MMP9 |
Identification of key candidate genes and
pathways with the methylated-DEGs PPI network
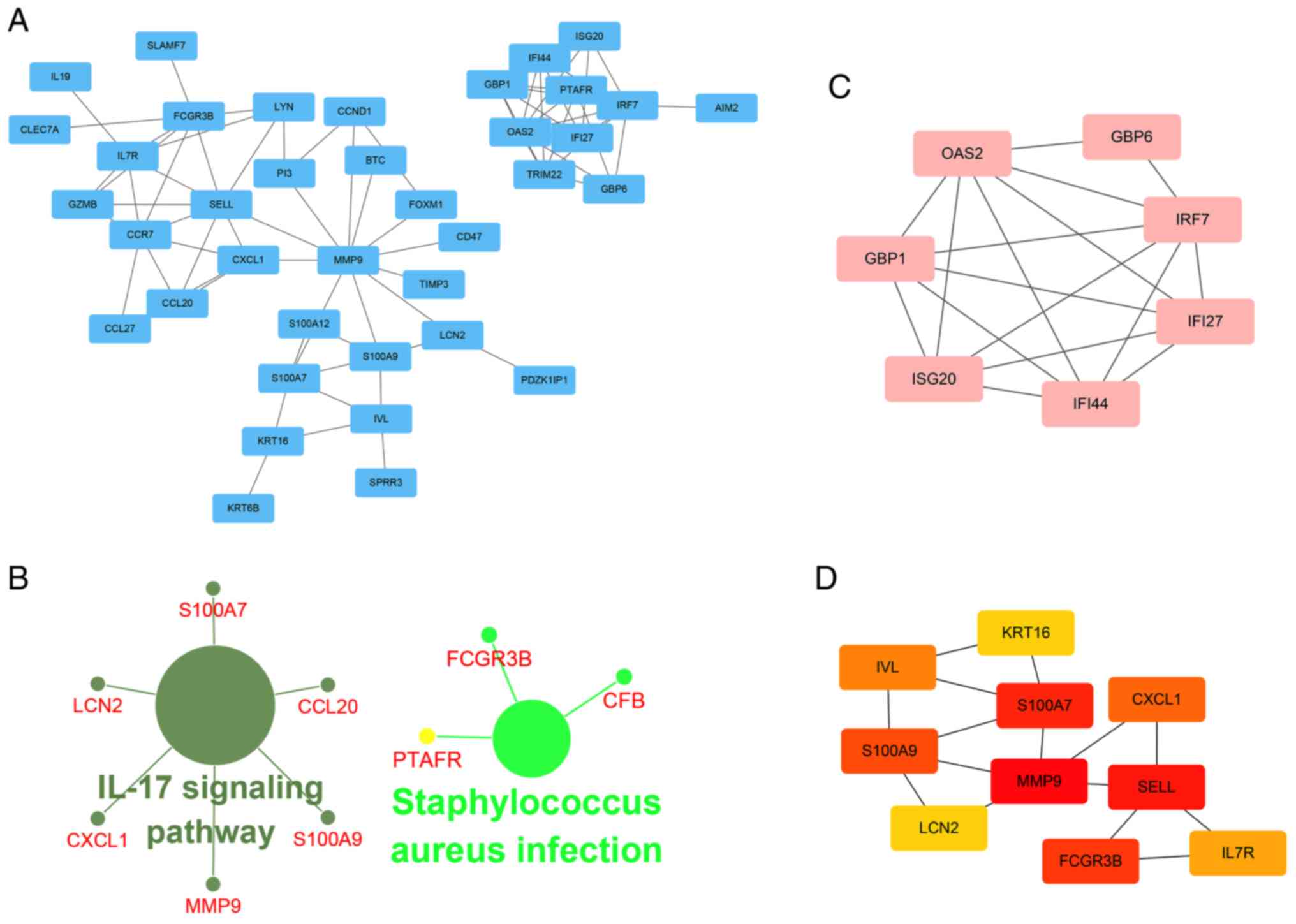
A PPI network of 95 methylated-DEGs was constructed
using Cytoscape software (Fig.
4A). According to the ClueGO analysis, these genes were
primarily involved in the 'interleukin (IL)-17 signaling pathway',
'Staphylococcus aureus infection', 'response to interferon'
and 'granulocyte chemotaxis' (Figs.
4B and S4). GBP1, IRF7,
OAS2, IFI44, GBP6, IFI27 and ISG20 were selected as significant
module genes using MCODE (Fig.
4C). The top 10 genes, calculated by Betweenness algorithms,
are presented in Fig. 4D. S100A9,
SELL, FCGR3B, MMP9, S100A7, IL7R, IRF7, CCR7, IFI44, CXCL1 and LCN2
were the top outstanding key genes based on a comprehensive
evaluation of 12 algorithms in cytoHubba, which were selected when
the genes contained in five or more algorithms.
Evaluation of the hub DEGs in IMQ-induced
psoriasiform dermatitis
TET2 is a key DNA methylated regulatory enzyme that
converts methylation to hydroxymethylation, thereby regulating gene
expression. In order to verify the hub DMGs screened in the present
study, a mouse model of IMQ-induced psoriasis with or without the
knockdown of TET2 expression was established. Compared with the
control group, the IMQ group exhibited significant psoriasiform
dermatitis of the erythema, thickening and scaling, whereas the
skin lesions of lentivirus-delivered shTET2-injected mice were
significantly decreased (Fig.
5A). The mRNA levels and protein concentration of TET2 were
decreased in the sh-TET2-injected mice compared with the IMQ group
(Fig. 5B, D and E). Due to the
significantly high expression of S100A9 in the IMQ group, the
western blot bands were slightly conjoined, which may affect the
accurate quantification of the protein. The expression levels of
the hub genes were detected by RT-qPCR. The mRNA expression levels
of SELL, FCGR3B, MMP9, S100A7, IL7R, IRF7, CCR7, IFI44, CXCL1 and
LCN2 were increased in the IMQ group compared with the control
group. The expression levels of SELL, IL7R, MMP9, CXCL1 and LCN2
were suppressed by sh-TET2 treatment (Fig. S5). Furthermore, compared with the
control group, the IMQ group exhibited a significantly increased
expression of S100A9, which was evidently suppressed by sh-TET2
treatment at both the mRNA and protein levels (Fig. 5C, F and G). When investigating the
molecular mechanisms underlying the effects of TET2 on S100A9
expression, it was revealed that the hydroxymethylation level of
S100A9 was significantly decreased in the IMQ + sh-TET2 group
(Fig. 5H). These data thus
indicate that S100A9 may be regulated by epigenetic approaches and
may play an important role in the pathogenesis of psoriasis.
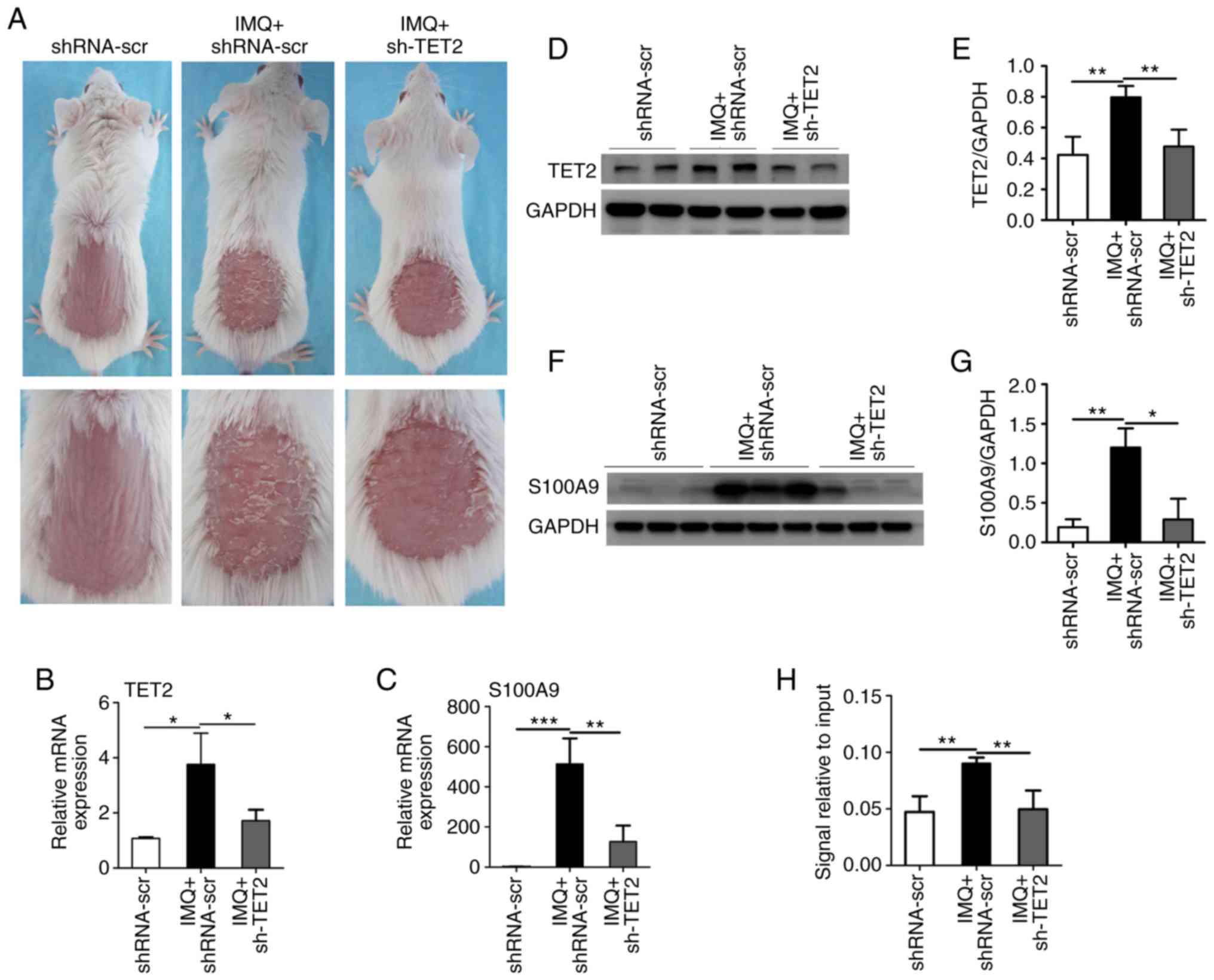 | Figure 5TET2 regulates the expression of
S100A9 in a model of psoriasiform dermatitis. (A) Representative
lesions of BALB/c mice following continuous treatment for 8 days.
(B and C) TET2 and S100A9 mRNA levels in the skin lesions were
evaluated by RT-qPCR. The protein expression levels of (D and E)
TET2 and (F and G) S100A9 in the skin lesions were measured by
western blot analysis. (H) The DNA hydroxymethylation level of
S100A9 was analyzed by hMeDIP-qPCR. The results are presented as
the means ± standard deviation. All experiments were performed 3
times, and the representative results are presented.
*P<0.05, **P<0.01 and
***P<0.001. S100A9, S100 calcium binding protein A9;
TET2, ten-eleven translocation 2; IMQ, imiquimod; hMeDIP-qPCR,
Hydroxymethylated DNA Immunoprecipitation-quantitative PCR;
shRNA-scr, lentivirus-delivered scrambled RNAs; shRNA-scr, control
group; IMQ+shRNA-scr, IMQ group; IMQ+sh-TET2, IMQ+sh-TET2
group. |
Discussion
The present study analyzed three gene expression
profiles and one gene methylation profile. A total of 352
upregulated DEGs and 137 downregulated DEGs were identified. In a
comparison between the DEGs and DMGs, 95 overlapping genes were
selected. The IL-17 signaling pathway and Staphylococcus
aureus infection pathway were the main enriched pathways. The
biological processes involved were the response to interferon and
granulocyte chemotaxis. S100A9, SELL, FCGR3B, MMP9, S100A7, IL7R,
IRF7, CCR7, IFI44, CXCL1 and LCN2 were the top outstanding genes.
Furthermore, S100A9 was regulated by epigenetic regulator TET2 in a
mouse model of psoriasis.
As the three raw gene expression microarrays had the
same normalization and quality control, the present study obtained
highly standardized and comparable data. Gene expression datasets
combined with comprehensive bioinformatics analysis can provide
reliable and new insights into potential crucial diagnostic and
therapeutic biomarkers. Researchers have previously focused on key
genes and pathways in different diseases (26). According to the analysis of
pathways and GO terms in the present study, DEGs were enriched in
the 'immune response', 'inflammatory response', 'cell adhesion',
'chemokine signaling pathway' and 'cell cycle', which is consistent
with extensive previous studies on psoriasis (5,27,28). Furthermore, all the top 3
significant terms of the biological process analysis and one of the
main pathways of the upregulated DEGs were associated with
responses to virus, suggesting that the pathogenesis of psoriasis
may be closely associated with viral infection. A recent study
revealed that activated RIG-I induced the production of IL-23 and
triggers psoriasiform dermatitis in mice, highlighting that the
dysregulation of antiviral immune responses may initiate the
inflammation of psoriasis (29).
Extracellular vesicles transport cytosolic proteins, lipids and
RNA, mediating intercellular communication between different cells
(30). Circulating endothelial-
and platelet-derived extracellular vesicles are increased in
psoriasis (31) and are decreased
following effective anti-TNF-α treatment (32). The present study demonstrated that
downregulated DEGs were evidently involved in extracellular
vesicles (OGN, APOE, EDIL3 and PRELP) in the GO cell component
analysis. Thus, these molecules may be potential biomarkers of
psoriasis.
Aberrant DNA methylation can influence the
expression of hub genes, and can induce a variety of
pathophysiological alterations (33). Therefore, the present study
identified 95 abnormally methylated DEGs by overlapping the DEGs
and DMGs. These genes were primarily involved in the 'IL-17
signaling pathway', 'response to interferon' and 'granulocyte
chemotaxis', which is reasonable, as these biological processes
have been demonstrated to increase the risk of developing psoriasis
(34). The top 11 hub genes were
selected by cyto-Hubba analysis. S100A7 is a calcium-binding
protein that participates in a number of biological functions, such
as proliferation and inflammation (35). S100A7 has been demonstrated to be
upregulated in psoriasis and is also named psoriasin, as it was
originally discovered in psoriasis (36). The knockdown of the histone
demethylase JMJD3 in keratinocytes has been shown to result in the
significant downregulation of the expression of S100A7 (37). MMP9 is an enzyme from the
gelatinase family that regulates cell matrix composition and
proliferation (38). Elevated
concentrations of MMP9 have been described in patients with
psoriasis (39). The aberrant
methylation of the promoter region of MMP9 has been shown to
decrease the levels of RNA and protein secretion in lymphoma cells
(40). FCGR3B, IL7R, IRF7, CCR7
and LCN2 have also been shown to be highly expressed in patients
with psoriasis (41-45). IRF7 is a key transcriptional
regulator of type I interferon (IFN)-dependent immune responses and
plays a vital role in the response against viruses (46). TET2 can maintain the
hypomethylation of IRF7 to regulate the TLR7/9-elicited IFN
response in plasmacytoid dendritic cells (pDCs) (47). Hence, these genes may be potential
candidate genes regulated by epigenetics that are involved in the
pathogenesis of psoriasis.
S100A9 is a calcium-binding protein of the S100
family, which plays a prominent role during the amplification of
the inflammatory response in autoimmunity, including chemotaxis,
adhesion and the degranulation of neutrophils (48). Through a comprehensive
bioinformatics analysis, S100A9 was one of the identified hub genes
of methylated DEGs. Furthermore, the research in the present study
demonstrated that the knockdown of epigenetic regulator, TET2, in
mice significantly decreased the expression of S100A9, both at the
mRNA and protein level. Compared with the IMQ group, the
hydroxymethylation level of S100A9 was also significantly decreased
in the IMQ + sh-TET2 group. These findings highlight the importance
of the regulation of S100A9 by an abnormal methylation status.
To the best of our knowledge, there are limited
studies that have used integrated bioinformatics analyses to gather
valuable information and provide important clues for future
research. Using computational systems biology, Sevimoglu and Arga
reported that the JAK/STAT signaling pathway was significant for
psoriasis. They screened out several biomarkers and therapeutic
candidates by analyzing the raw data of gene expression datasets
(49). Certain feature algorithms
filtered gene expression profiles in order to acquire
classification models of psoriasis (50). Unlike previous studies, the
present study comprehensively analyzed DEGs and DMGs to identify
active modules and key epigenetically-associated DEGs.
The integrated bioinformatics analysis of both the
gene expression profile and the methylation profile is more
reliable and provide valuable, novel molecular targets to foster
the development of accurate diagnosis and novel therapeutic
strategies. However, one limitation of this study was the lack of
clinical human specimens. Additional studies with clinical
materials and experiment validation are required in order to verify
the key candidate genes. Furthermore, the present study analyzed
only a single methylation profile microarray; thus, more comparable
microarray data are required so that more valuable data can be
obtained. The present study selected S100A9 as a representative
gene, and preliminarily evaluated the expression and epigenetic
regulatory mechanism of S100A9 in psoriasis. Further experiments
are required to clarify the exact association between TET2 and
S100A9, and the molecular mechanisms of action.
In conclusion, the results from the present study
not only identify a series of DEGs and DEMGs, but also analyze the
significant modules and key candidate genes. In addition, in order
to further verify the bioinformatics analysis data, the present
study detected the expression levels of hub genes (S100A9, SELL,
FCGR3B, MMP9, S100A7, IL7R, IRF7, CCR7, IFI44, CXCL1 and LCN2) in a
mouse model of psoriasiform dermatitis by interfering with the
expression of methylation-regulating enzyme TET2. These key genes
may serve as potential biomarkers in psoriasis.
Supplementary Data
Acknowledgments
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81903212, 81673057 and
81502730).
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW, XL, NL and HC conceived and designed the study.
XW and NL performed the analysis procedures. XL, NL and HC analyzed
the results. XW performed the experiments. XL and HC contributed
analysis tools. XW, XL and NL wrote the manuscript. All authors
have read and approved the final manuscript.
Ethics approval and consent to
participate
All experimental protocols with animals were
conducted following the ARRIVE guidelines. All mouse care and
procedures were approved by the Ethics Committee for experimental
animals, Tongji Medical College (no. SYXK HUBEI 2016-0057).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Raychaudhuri SK, Maverakis E and
Raychaudhuri SP: Diagnosis and classification of psoriasis.
Autoimmun Rev. 13:490–495. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Duffin KC, Chandran V, Gladman DD, Krueger
GG, Elder JT and Rahman P: Genetics of psoriasis and psoriatic
arthritis: Update and future direction. J Rheumatol. 35:1449–1453.
2008.PubMed/NCBI
|
|
3
|
Fogel O, Richard-Miceli C and Tost J:
Epigenetic changes in chronic inflammatory diseases. Adv Protein
Chem Struct Biol. 106:139–189. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Griffiths CE: The immunological basis of
psoriasis. J Eur Acad Dermatol Venereol. 17(Suppl 2): S1–S5. 2003.
View Article : Google Scholar
|
|
5
|
Boehncke WH and Schön MP: Psoriasis.
Lancet. 386:983–994. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Turek-Plewa J and Jagodziński PP: The role
of mammalian DNA methyltransferases in the regulation of gene
expression. Cell Mol Biol Lett. 10:631–647. 2005.PubMed/NCBI
|
|
7
|
Ichiyama K, Chen T, Wang X, Yan X, Kim BS,
Tanaka S, Ndiaye-Lobry D, Deng Y, Zou Y, Zheng P, et al: The
methyl-cytosine dioxygenase Tet2 promotes DNA demethylation and
activation of cytokine gene expression in T cells. Immunity.
42:613–626. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Solary E, Bernard OA, Tefferi A, Fuks F
and Vainchenker W: The ten-eleven translocation-2 (TET2) gene in
hematopoiesis and hematopoietic diseases. Leukemia. 28:485–496.
2014. View Article : Google Scholar
|
|
9
|
Wang X, Liu X, Duan X, Zhu K, Zhang S, Gan
L, Liu N, Jaypaul H, Makamure JT, Ming Z and Chen H: Ten-eleven
translocation-2 regulates DNA hydroxymethylation status and
psoriasiform dermatitis progression in mice. Acta Derm Venereol.
98:585–593. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Roberson ED, Liu Y, Ryan C, Joyce CE, Duan
S, Cao L, Martin A, Liao W, Menter A and Bowcock AM: A subset of
methylated CpG sites differentiate psoriatic from normal skin. J
Invest Dermatol. 132:583–592. 2012. View Article : Google Scholar
|
|
11
|
Chen L, Sun F, Yang X, Jin Y, Shi M, Wang
L, Shi Y, Zhan C and Wang Q: Correlation between RNA-Seq and
microarrays results using TCGA data. Gene. 628:200–204. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li B, Tsoi LC, Swindell WR, Gudjonsson JE,
Tejasvi T, Johnston A, Ding J, Stuart PE, Xing X, Kochkodan JJ, et
al: Transcriptome analysis of psoriasis in a large case-control
sample: RNA-seq provides insights into disease mechanisms. J Invest
Dermatol. 134:1828–1838. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu J, Li H, Sun L, Wang Z, Xing C and
Yuan Y: Aberrantly methylated-differentially expressed genes and
pathways in colorectal cancer. Cancer Cell Int. 17:752017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huo X, Sun H, Cao D, Yang J, Peng P, Yu M
and Shen K: Identification of prognosis markers for endometrial
cancer by integrated analysis of DNA methylation and RNA-Seq data.
Sci Rep. 9:99242019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gautier L, Cope L, Bolstad BM and Irizarry
RA: Affy-analysis of affymetrix GeneChip data at the probe level.
Bioinformatics. 20:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: Limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dennis G Jr, Sherman BT, Hosack DA, Yang
J, Gao W, Lane HC and Lempicki RA: DAVID: Database for annotation,
visualization, and integrated discovery. Genome Biol. 4:P32003.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The gene
ontology consortium Nat Genet. 25:25–29. 2000.
|
|
19
|
Szklarczyk D, Franceschini A, Kuhn M,
Simonovic M, Roth A, Minguez P, Doerks T, Stark M, Muller J, Bork
P, et al: The STRING database in 2011: Functional interaction
networks of proteins, globally integrated and scored. Nucleic Acids
Res. 39:D561–D568. 2011. View Article : Google Scholar :
|
|
20
|
Chin CH, Chen SH, Wu HH, Ho CW, Ko MT and
Lin CY: cyto-Hubba: Identifying hub objects and sub-networks from
complex interactome. BMC Syst Biol. 8(Suppl 4): S112014. View Article : Google Scholar
|
|
21
|
van der Fits L, Mourits S, Voerman JS,
Kant M, Boon L, Laman JD, Cornelissen F, Mus AM, Florencia E, Prens
EP and Lubberts E: Imiquimod-induced psoriasis-like skin
inflammation in mice is mediated via the IL-23/IL-17 axis. J
Immunol. 182:5836–5845. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schukur L, Geering B, Charpin-El Hamri G
and Fussenegger M: Implantable synthetic cytokine converter cells
with AND-gate logic treat experimental psoriasis. Sci Transl Med.
7:318ra2012015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Taves S, Berta T, Liu DL, Gan S, Chen G,
Kim YH, Van de Ven T, Laufer S and Ji RR: Spinal inhibition of p38
MAP kinase reduces inflammatory and neuropathic pain in male but
not female mice: Sex-dependent microglial signaling in the spinal
cord. Brain Behav Immun. 55:70–81. 2016. View Article : Google Scholar
|
|
24
|
Gargiulo S, Greco A, Gramanzini M,
Esposito S, Affuso A, Brunetti A and Vesce G: Mice anesthesia,
analgesia, and care, part I: Anesthetic considerations in
preclinical research. ILAR J. 53:E55–E69. 2012. View Article : Google Scholar
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
26
|
Dong B, Wang G, Yao J, Yuan P, Kang W, Zhi
L and He X: Predicting novel genes and pathways associated with
osteosar-coma by using bioinformatics analysis. Gene. 628:32–37.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mabuchi T and Hwang ST: ACKR2: Nature's
decoy receptor lures unsuspecting chemokines in psoriasis. J Invest
Dermatol. 137:7–11. 2017. View Article : Google Scholar
|
|
28
|
Reich K: The concept of psoriasis as a
systemic inflammation: Implications for disease management. J Eur
Acad Dermatol Venereol. 26(Suppl 2): S3–S11. 2012. View Article : Google Scholar
|
|
29
|
Zhu H, Lou F, Yin Q, Gao Y, Sun Y, Bai J,
Xu Z, Liu Z, Cai W, Ke F, et al: RIG-I antiviral signaling drives
interleukin-23 production and psoriasis-like skin disease. EMBO Mol
Med. 9:589–604. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Colombo M, Raposo G and Théry C:
Biogenesis, secretion, and intercellular interactions of exosomes
and other extracellular vesicles. Annu Rev Cell Dev Biol.
30:255–289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pelletier F, Garnache-Ottou F, Angelot F,
Biichlé S, Vidal C, Humbert P, Saas P, Seillès E and Aubin F:
Increased levels of circulating endothelial-derived microparticles
and small-size platelet-derived microparticles in psoriasis. J
Invest Dermatol. 131:1573–1576. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pelletier F, Garnache-Ottou F, Biichlé S,
Vivot A, Humbert P, Saas P, Seillès E and Aubin F: Effects of
anti-TNF-α agents on circulating endothelial-derived and
platelet-derived micropar-ticles in psoriasis. Exp Dermatol.
23:924–925. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li J, Chen C, Bi X, Zhou C, Huang T, Ni C,
Yang P, Chen S, Ye M and Duan S: DNA methylation of CMTM3, SSTR2,
and MDFI genes in colorectal cancer. Gene. 630:1–7. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lowes MA, Russell CB, Martin DA, Towne JE
and Krueger JG: The IL-23/T17 pathogenic axis in psoriasis is
amplified by keratinocyte responses. Trends Immunol. 34:174–181.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Donato R, Cannon BR, Sorci G, Riuzzi F,
Hsu K, Weber DJ and Geczy CL: Functions of S100 proteins. Curr Mol
Med. 13:24–57. 2013. View Article : Google Scholar :
|
|
36
|
Madsen P, Rasmussen HH, Leffers H, Honoré
B, Dejgaard K, Olsen E, Kiil J, Walbum E, Andersen AH, Basse B, et
al: Molecular cloning, occurrence, and expression of a novel
partially secreted protein 'psoriasin' that is highly up-regulated
in psoriatic skin. J Invest Dermatol. 97:701–712. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gschwandtner M, Zhong S, Tschachler A,
Mlitz V, Karner S, Elbe-Bürger A and Mildner M: Fetal human
keratinocytes produce large amounts of antimicrobial peptides:
Involvement of histone-methylation processes. J Invest Dermatol.
134:2192–2201. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Monhian N, Jewett BS, Baker SR and Varani
J: Matrix metal-loproteinase expression in normal skin associated
with basal cell carcinoma and in distal skin from the same
patients. Arch Facial Plast Surg. 7:238–243. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Głażewska EK, Niczyporuk M, Ławicki S,
Szmitkowski M, Donejko M, Zajkowska M, Będkowska GE and Przylipiak
A: Narrowband ultraviolet B light treatment changes plasma
concentrations of MMP-3, MMP-9 and TIMP-3 in psoriatic patients.
Ther Clin Risk Manag. 13:575–582. 2017. View Article : Google Scholar :
|
|
40
|
Chicoine E, Estève PO, Robledo O, Van
Themsche C, Potworowski EF and St-Pierre Y: Evidence for the role
of promoter methylation in the regulation of MMP-9 gene expression.
Biochem Biophys Res Commun. 297:765–772. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wu Y, Zhang Z, Tao L, Chen G, Liu F, Wang
T, Xue F, Chen Y, He L, Zheng J and Liu Y: A high copy number of
FCGR3B is associated with psoriasis vulgaris in Han Chinese.
Dermatology. 229:70–75. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lee SK, Jeon EK, Kim YJ, Seo SH, Kim CD,
Lim JS and Lee JH: A global gene expression analysis of the
peripheral blood mono-nuclear cells reveals the gene expression
signature in psoriasis. Ann Dermatol. 21:237–242. 2009. View Article : Google Scholar
|
|
43
|
Raposo RA, Gupta R, Abdel-Mohsen M, Dimon
M, Debbaneh M, Jiang W, York VA, Leadabrand KS, Brown G, Malakouti
M, et al: Antiviral gene expression in psoriasis. J Eur Acad
Dermatol Venereol. 29:1951–1957. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rittié L and Elder JT: Capturing the finer
points of gene expression in psoriasis: Beaming in on the
CCL19/CCR7 axis. J Invest Dermatol. 132:1535–1538. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Shao S, Cao T, Jin L, Li B, Fang H, Zhang
J, Zhang Y, Hu J and Wang G: Increased lipocalin-2 contributes to
the pathogenesis of psoriasis by modulating neutrophil chemotaxis
and cytokine secretion. J Invest Dermatol. 136:1418–1428. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Honda K, Yanai H, Negishi H, Asagiri M,
Sato M, Mizutani T, Shimada N, Ohba Y, Takaoka A, Yoshida N and
Taniguchi T: IRF-7 is the master regulator of type-I
interferon-dependent immune responses. Nature. 434:772–777. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ma S, Wan X, Deng Z, Shi L, Hao C, Zhou Z,
Zhou C, Fang Y, Liu J, Yang J, et al: Epigenetic regulator CXXC5
recruits DNA demethylase Tet2 to regulate TLR7/9-elicited IFN
response in pDCs. J Exp Med. 214:1471–1491. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ehrchen JM, Sunderkötter C, Foell D, Vogl
T and Roth J: The endogenous Toll-like receptor 4 agonist
S100A8/S100A9 (calprotectin) as innate amplifier of infection,
autoimmunity, and cancer. J Leukoc Biol. 86:557–566. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Sevimoglu T and Arga KY: Computational
systems biology of psoriasis: Are we ready for the age of Omics and
systems biomarkers? OMICS. 19:669–687. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Guo P, Luo Y, Mai G, Zhang M, Wang G, Zhao
M, Gao L, Li F and Zhou F: Gene expression profile based
classification models of psoriasis. Genomics. 103:48–55. 2014.
View Article : Google Scholar
|