Introduction
Obesity is a chronic non-communicable disease, the
rate of which has been rising exponentially on a global scale
(1). It is ranked among the top
10 risk factors associated with several other conditions, such as
type II diabetes mellitus, dyslipidemia and cardiovascular disease
(2). Furthermore, obesity
increases the incidence of psychological diseases, in particular
major depressive disorder (3).
Several neural mechanisms associated with obesity may occur in the
hypothalamus and nucleus tractus solitarii (NTS). The hypothalamus
and NTS are areas of the brain that are primarily concerned with
food intake and energy expenditures (4,5).
Food intake responses, including appetite and satiety senses, are
regulated through the release of hormones and molecules from the
stomach, adipose tissue, the intestines, the pancreas, the
pituitary gland, the hypothalamus and the NTS. The functions of
this pathway are relatively clear when observed in isolation. After
the central nervous system receives the relevant signals from
organs, it secretes orexigenic and anorexigenic peptides
responsible for the amount of food consumed (6,7).
Pathologically, the etiology of obesity is attributed to the total
number and size of adipocytes, which include the cells that make up
white adipose tissue (WAT) and brown adipose tissue (BAT). Previous
studies have identified that BAT is able to produce heat and
energy. This mechanism increases energy expenditures,
thermogenesis, and the oxidation of lipids and may influence the
development of obesity (8,9).
The transient receptor potential vanilloid 1 (TRPV1)
is one of 6 members of the cation channel subfamily. TRPV1 is the
first unit in this sequence to be specified and most keenly
characterized, due to its ability to act as a homotetrameric,
nonselective, calcium-permeable cation channel (10,11). TRPV1 is well known for its role in
nociception and inflammation, which may occur at lowered pH levels
(pH 6.0) and high temperatures (43°C) (12). Aside from the factors already
mentioned, there are other factors that are associated with TRPV1
function, such as pathological weight gain and certain
psychosomatic disorders (13,14). This response causes a signaling
cascade in neurons via the activation of protein kinases, including
mitogen-activated protein kinase (MAPK), and cyclic AMP-response
element binding protein (CREB) (15). These protein kinase families are
the secondary messengers that mediate intracellular signaling.
These kinases include protein kinase A (PKA) and protein kinase C
(PKC). These signaling cascades are important for maintaining
homeostasis and may be stimulated and/or inhibited by various
factors such as acupuncture. A number of pathological mechanisms
are associated with the dysregulation of one, many, or all of the
factors associated with these signaling pathways (16,17).
The theory of Traditional Chinese Medicine
introduced the concept of obesity as a result of dysfunction within
the gastrointestinal organs, primarily the spleen and the stomach
(18,19). Acupuncture has been used as a
traditional method for treating several diseases, including weight
control. The combination of acupuncture and lifestyle modifications
may significantly improve weight loss (20). Acupoint catgut embedding (ACE)
involves the implantation of absorbable catgut sutures at
traditional Chinese meridian acupoints. This technique is believed
to confer beneficial therapeutic effects: Notable decreases in
treatment frequency from approximately twice a week to twice a
month have been observed (21).
The ability to conserve time and costs makes ACE an attractive
treatment method. Although side effects such as local inflammation,
pain, callosity, and discomfort may occur for several days
following treatment, this method has been widely used for the
management of obesity (22).
Zusanli (ST36) is most commonly used for the treatment of pain,
lower limb issues and gastrointestinal disorders. In TCM, it is
believed that it may also be used to treat diseases that manifest
along the stomach meridian. In addition to the positive effects
associated with using ACE to treat obesity, ST36 is also frequently
combined with local abdominal acupoints to treat obesity and
maintain normal body weight (23). The purpose of the present study
was to investigate the effect of ACE on obesity and its associated
complications by identifying various neural mechanisms associated
with the disease. Therefore, we hypothesized that ACE treatment may
decrease the incidence of obesity through the control and
maintenance of body weight. In addition, we also hypothesized that
ACE treatment would also decrease the complications associated with
obesity via modulation of TRPV1 signaling.
Materials and methods
Experimental animals
A total of 28 female C57/BL6 wild type (WT) mice and
14 female TRPV1 knockout (KO) mice that were 7 weeks old and
weighing 19-22 g, were purchased from BioLASCO Taiwan Co., Ltd. The
animals were housed individually in cages under a 12:12 h
light-dark cycle (from 8.00 a.m. to 8.00 p.m.), at a temperature of
25±1°C, and humidity of 60±5% relative humidity, with ad
libitum access to water and a normal mice chow diet (ND) for 1
week prior to initiation of the experiment. The study was approved
by the Institute of Animal Care and USE Committee of China Medical
University (Permit no. 2016-061), following the Guide for the Care
and Use of Laboratory Animals (24). The first day of experiment, the
cages, animal bedding and water were changed, and the mice chow was
superseded by high-fat diet (HFD) (Research Diets Inc.; cat. no.
D12451) composed of 45 kcal% fat (1,598 kcal% of lard; 225 kcal%
soybean oil, USP), 35 kcal% carbohydrate and 20 kcal% protein (473
kcal/100 g). Subjects were randomly divided into six groups:
Control group (WT-ND); obesity group (WT-HFD); Acupoint catgut
embedding (WT-HFD-ACE) group; sham acupoint catgut embedding
(WT-HFD-SHAM) group; TRPV1 knockout mice with normal diet (KO-ND);
and TRPV1 knockout mice with HFD (KO-HFD), with 7 mice in each
group. All food of the subjects was weighed before being
replenished with either ND or HFD mice chow weekly. The subjects
were weighed once a week throughout the 8 week experimental period
to study weight changes. Food was collected, measured, refilled and
replaced to detect food consumption patterns on the same day
following body weight measurement. The cages were cleaned once a
week, with the animal bedding and water also changed. A preliminary
pilot study was conducted with 4 animals to analyze the absorbable
ability of catgut at various time intervals. All 4 subjects were
treated via insertion of absorbable catgut into the ST36 acupoint.
After 7 days, the implantation area of 2 mice was incised, and the
implantation area of the remaining 2 mice was incised at 14 days
after the embedding to establish the ideal absorbability duration.
The experiment was designed to sacrifice that the mice at the end
of the 8th week of the experiment. They were fasted with no access
to food but were fed water ad libitum 12 h prior to
sacrifice. Efforts were made to minimize the number of animals used
and their suffering. A total of 8 subjects were removed during the
experimental period due to excessive food gnawing behavior and
weight gain or loss at 25±1 g, as established in Week 4 prior to
initiation of the treatment session.
Acupoint catgut embedding treatment
Mice in the WT-HFD-ACE group received ACE treatment
at the bilateral side of ST36 once a week on the first day of the 5
to 8th week. In mice, as in humans, the ST36 point is located
longitudinally at 3 cun below the knee joint and intersects with
the middle of the tibialis anterior muscle (25). Sterile conventional syringe
needles 0.6×25 mm (Terumo Corporation), acupuncture needles 0.35×40
mm (Suzhou Medical Appliance) and brown catgut 0.2×4 mm (CP Medical
Inc.) were used for the implantation. Animals from the WT-ND,
WT-HFD, KO-ND and KO-HFD groups were placed into a fixation machine
under anesthesia with 5% isoflurane for induction, which was then
decreased to 1% for maintenance. Bilateral ST36 acupoint were
selected, sterilized with 70% alcohol and iodine solution and the
catgut was embedded at the 5 mm depth before the animals were
returned to individual observation home cages after the procedure.
The subjects in the WT-HFD-SHAM group received sham ACE treatment,
which consisted of using similar equipment to insert the empty
needle at the ST36 acupoint without catgut implantation. The needle
embedding causes a minute red mark at the area of needle insertion,
which was fully recovered within 48 h after the treatment. In
addition, there was no apparent difficulty of movement following
anesthesia.
Collection of samples
In the samples collection process, after subjects
were fasted for 12 h, 38 subjects were euthanized with 5%
isoflurane by inhalation. Blood samples were collected from the
Orbital sinus into 3 ml BD Vacutainer glass tubes with 5.4 mg K2
EDTA and 2 ml BD Vacutainer glass tubes with 3 mg sodium fluoride
and 6 mg Na2 EDTA. The samples were centrifuged at 1,500
× g for 15 min at 25°C, following which the separated plasma was
collected into 1.5 ml microcentrifuge tubes and stored at -80°C.
Following collection of the blood samples, the animals were
decapitated, and brains were excised for western blot analysis. The
adipose tissues were then collected from various regions of the
body; BAT from the interscapular area, subcutaneous WAT (sWAT) from
both sides of the flank area and visceral WAT (vWAT) from the
perigonadal area (26). Finally,
the liver and both kidneys were immediately dissected out and
weighed.
Western blot analysis
Western blot analysis is an analytical technique
that is widely used to study proteins. This method, first described
by Towbin et al (27),
uses an antibody that recognizes and binds to an epitope unique to
the protein of interest. In the present study, following sacrifice,
the hypothalamus, prefrontal cortex (PFC) and NTS were immediately
dissected out and frozen in ice prior to storage at -80°C. Total
proteins were prepared by abrasion and lysis in solution of 50 mM
Tris-HCl pH 7.4, 250 mM NaCl, 1% NP-40, 5 mM EDTA, 50 mM NaF, 1 mM
Na3VO4, 0.02% NaN3 and 1X protease
inhibitor cocktail (Amresco, LLC) prior to centrifugation at 10,000
× g for 10 min at 4°C. Proteins from each sample were loaded into
8% SDS-Tris glycine gel electrophoresis gels and transferred onto
PVDF membranes, which were then blocked with 5% non-fat milk in
TBS-T buffer (10 mM Tris pH 7.5, 100 mM NaCl, 0.1% Tween-20), and
incubated for 1 h at room temperature with the following primary
antibodies: Anti-TRPV1 (~95 kDa; Alomone; cat. no. ACC-030;
1:1,000), anti-p-PI3K (~110 kDa; Novus Biologicals, LLC; cat. no.
NBP2-15071; 1:1,000), anti-PI3K (~126 kDa; Abcam; cat. no.
ab154598; 1:1,000), anti-phosphorylated (p)-Akt (~65 kDa, Thermo
Fisher Scientific, Inc.; cat. no. 44-621G; 1:1,000), anti-Akt (~65
kDa; Merck KGaA; cat. no. 16-293; 1:1,000), anti-p-mTOR (~289 kDa;
Merck KGaA; cat. no. 09-213; 1:1,000), anti-mTOR (~238 kDa; Abcam;
cat. no. ab2732; 1:1,000), anti-p-extracellular signal-regulated
kinase (ERK) 1/2 (p-ERK)~42 kDa; Abcam; cat. no. ab138482;
1:1,000), anti-ERK (~42-44 kDa; Cell Signaling Technology, Inc.;
cat. no. 4695; 1:1,000), anti-p-c-Jun N-terminal kinase (p-JNK;
~45-55 kDa; Thermo Fisher Scientific, Inc.; cat. no. 44-682G;
1:1,000), anti-JNK (~46-54 kDa; Cell Signaling Technology, Inc.;
cat. no. 9252; 1:1,000), anti-p-p38 mitogen-activated protein
kinase (p-p38; ~41 kDa; Thermo Fisher Scientific, Inc.; cat. no.
44-684G; 1:1,000), anti-p38 (~43 kDa; Cell Signaling Technology,
Inc.; cat. no. 9212; 1:1,000), anti-p-NF-κB (~65 kDa; Merck KGaA;
cat. no. ABS403; 1:1,000), anti-NF-κB (~65 kDa; Cell Signaling
Technology, Inc.; cat. no. 8242; 1:1,000), anti-p-CREB (~43 kDa;
Merck KGaA; cat. no. 06-519; 1:1,000), anti-CREB (~43 kDa; Cell
Signaling Technology, Inc.; cat. no. 9197; 1:1,000), anti-p-protein
kinase C epsilon type (p-PKCε; ~82 kDa; Santa Cruz Biotechnology,
Inc.; cat. no. SC-12355; 1:1,000), anti-PKCε (~84 kDa; Abcam; cat.
no. ab63638; 1:1,000), anti-p-protein kinase AII α (p-PKAIIα; ~40
kDa; Santa Cruz Biotechnology, Inc.; cat. no. SC-12905; 1:1,000) or
anti-PKAIIα (~40 kDa; Santa Cruz Biotechnology, Inc.; cat. no.
SC-136262; 1:1,000) in TBST with 1% bovine serum albumin (BSA)
(Sigma-Aldrich; Merck KGaA). Horseradish peroxidase-conjugated
AffiniPure goat anti-mouse (Jackson ImmunoResearch Laboratory,
Inc.; cat. no. 115-035-003; 1:5,000), goat anti-rabbit (Jackson
ImmunoResearch Laboratory, Inc; cat. no. 111-035-003; 1:5,000) and
donkey anti-goat (Jackson ImmunoResearch Laboratory, Inc.; cat. no.
705-035-003; 1:5,000) secondary antibodies were incubated with the
membranes for 1 h incubation at room temperature. The protein bands
on the membranes were visualized using an enhanced chemiluminescent
substrate kit (Pierce; Thermo Fisher Scientific, Inc.) with
LAS-3000 Fujifilm (Fuji Photo Film Co. Ltd). The image densities of
the specific bands were quantified by National Institutes of Health
(NIH) ImageJ software (version 1.8.0).
Immunofluorescence
A total of 4 subjects, from the WT-HFD, WT-HFD-ACE,
WT-HFD-SHAM, and KO-HFD groups, were anesthetized using 1%
isoflurane by inhalation and intracardially perfused with saline
followed by 4% paraformaldehyde. The brain was immediately
dissected and post-fixed with 4% paraformaldehyde at 4°C overnight.
Post-fixed tissues were placed overnight in 30% sucrose for
cryoprotection at 4°C. The brain was embedded in OCT and
instantaneously frozen using liquid nitrogen prior to storage at
-80°C. Frozen segments were cut at a thickness of 16 µm
width on a cryostat then placed on glass slides. The samples were
incubated with blocking solution, which consisted of 3% BSA, 0.1%
Triton X-100 and 0.02% sodium azide, for 2 h at room temperature.
Following blocking, the brain samples were incubated overnight with
the primary antibody, TRPV1 (~95 kDa; Alomone; cat. no. ACC-030;
1:200), pPKAIIα (~40 kDa; Santa Cruz Biotechnology, Inc.; cat. no.
SC-12905; 1:200), p-PI3K (~110 kDa; Novus Biologicals, LLC; cat.
no. NBP2-15071; 1:200), and p-CREB (~43 kDa; Merck KGaA; cat. no.
06-519; 1:200), prepared in BSA solution at 4°C overnight. The
secondary antibodies, Alexa Fluor 488-conjugated AffiniPure donkey
anti-rabbit (Jackson ImmunoResearch Laboratory, Inc.; cat. no.
711-545-152; 1:500), donkey anti-mouse (Jackson ImmunoResearch
Laboratory, Inc., cat. no. 715-545-150; 1:500) and 594-conjugated
AffiniPure donkey anti-goat (Jackson ImmunoResearch Laboratory,
Inc.; cat. no. 705-585-003; 1:500) were used for incubation at room
temperature for 2 h prior to being fixed with cover slips for
immunofluorescence visualization. The samples were observed by an
epifluorescent microscope (BX-51; Olympus Corporation) with ×200
total magnification (×20 numerical aperture (NA=0.4) objective lens
and ×10 ocular lens). The images were analyzed by NIH ImageJ
software (version 1.8.0).
ELISA
The level of mouse leptin (cat. no. KMC2281) and
insulin (cat. no. EMINS) in the blood plasma were detected using
ELISA kits following the manufacturer's protocol (Thermo Fisher
Scientific, Inc.; catalog no. EIAGLUC). The ELISA procedures were
prepared at room temperature. The plasma samples were placed in
96-well plates with mouse leptin or insulin antibody coating prior
to incubation at 25°C for 2 h. They were then removed from the
plate and washed with wash buffer. The biotinylated antibody
reagent was added in the wells and incubated for 1 h before the
washing process. The wells were then incubated at 25°C for 30 min
with a streptavidin-HRP reagent and then washed. Next,
tetramethylbenzidine substrate was added into each well to incubate
for 30 min in darkness. Following the addition of the stop
solution, the absorbent substance at 450 nm was measured using a
microplate reader. The glucose level was measured with the glucose
colorimetric detection kit (Thermo Fisher Scientific, Inc.,
EIAGLUC); in this procedure, the principles of the glucose
oxidation reaction with glucose to produce hydrogen peroxide were
used; the resulting colored product was then read by the microplate
reader at 450 nm for leptin plasma level and 550 nm for insulin and
glucose plasma levels. Finally, the samples were measured and
analyzed using the kit-specific standard CurveExpert Basic (version
2.1.0; Hyams Development).
Statistical analysis
Statistical analysis was performed using the SPSS
statistical software package program v.22.0 (IBM Corp.). All data
are presented as the mean ± standard error of the mean. Statistical
comparisons were evaluated using a one-way analysis of variance
followed by Tukey's post hoc test to determine the significance of
differences. P<0.05 was considered to indicate a statistically
significant difference.
Results
Effect of ACE treatment on weight
control
The simple standard of obesity in the mouse model
was indicated by an increase in net body weight throughout the
8-week experimental period. All analyzed data are presented in
Fig. 1A. During this experiment,
a significant difference in weight gain between the WT and the
TRPV1 KO groups was identified. This was observed in the initial
increase in body weight observed in the WT-ND, KO-ND, and KO-HFD
groups compared with that in the WT-HFD, WT-HFD-ACE, and
WT-HFD-SHAM groups at Week 4. A significant increase in body weight
was observed in the WT groups, with the WT-HFD-ACE group
demonstrating a gradual decrease in Week 7 and significant
difference in Week 8. At Week 8, there was a slight decrease in
body weight observed in the WT-HFD-SHAM group. Although the mean
body weight of the WT-HFD-SHAM group was decreased compared with
that of the WT-HFD group, a significant difference in body weight
between the WT-HFD-ACE and WT-HFD-SHAM groups was observed. At Week
4, there was a significant decrease in the KO-ND group compared
with that in the WT-ND group. Concomitantly, no significant
difference between the WT-ND and KO-HFD groups was observed at this
time. Food intake among the groups was also examined. A significant
increase in food consumption was observed in the WT-ND and KO-ND
groups from the first week of the experiment compared with that of
the other groups. Furthermore, a significantly decreased level of
food consumption was observed in the KO-HFD group compared with
that in the other HFD-fed groups (Fig. 1B). The results demonstrated a
slight decrease in food intake in the WT-HFD-ACE group following
ACE treatment compared with that in the WT-HFD and WT-HFD-SHAM
groups. However, food consumption in the WT-HFD-ACE group
significantly decreased during the 8th week compared with that in
the WT-HFD group. A nonsignificant increase in food consumption
across all groups was observed in accordance with the growth
requirements of the mice.
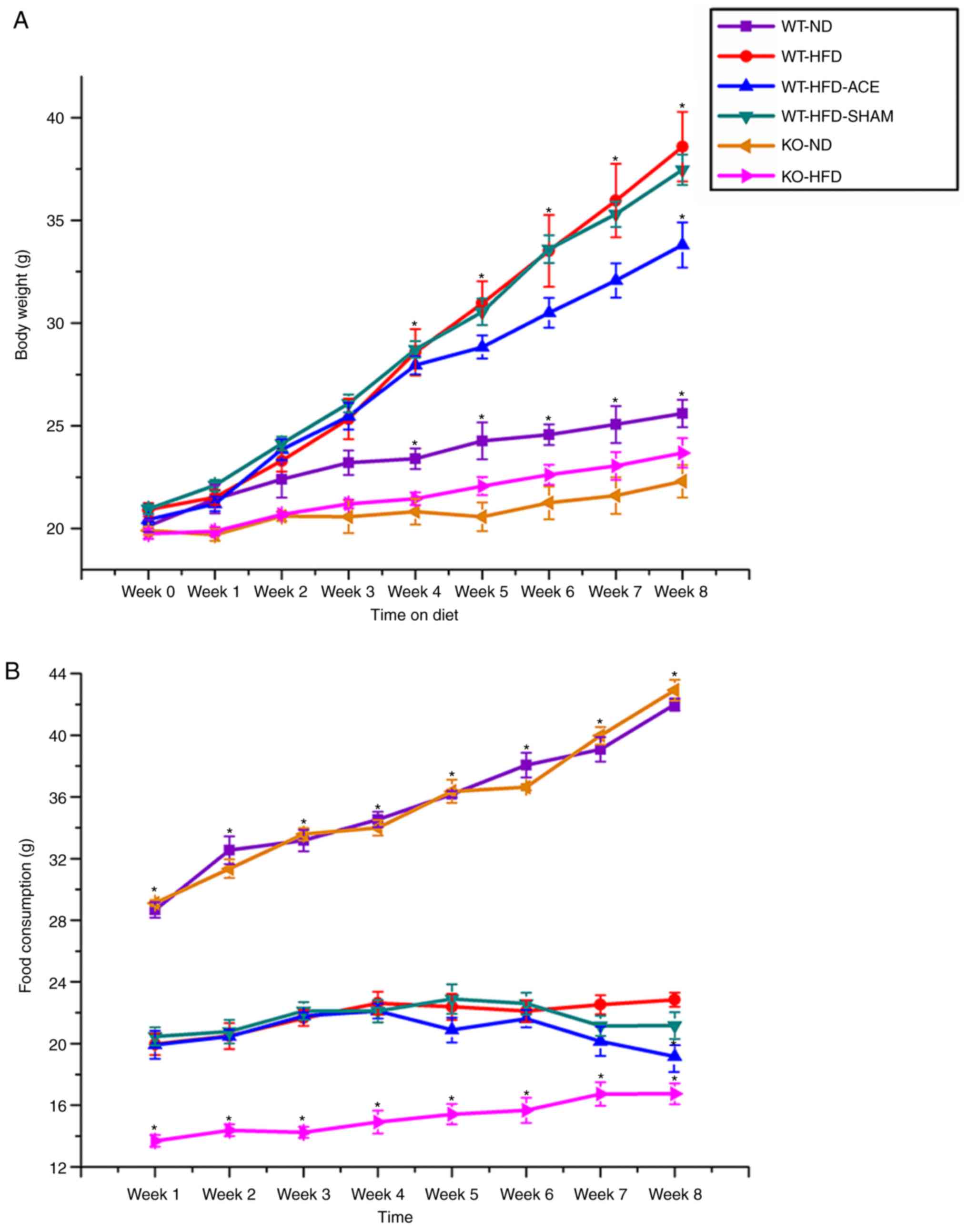 | Figure 1Weekly body weight alterations and
food consumption in the six subject groups. (A) The graph presents
comparisons of body weight in the WT-ND, WT-HFD, WT-HFD-ACE,
WT-HFD-SHAM, KO-ND and KO-HFD groups. Significant body weight
increases in the WT-HFD, WT-HFD-ACE and WT-HFD-SHAM groups compared
to the WT-ND group and both TRPV1 KO mouse groups were observed.
*P<0.05. There was significant body weight decease in
the WT-HFD-ACE group compared to the WT-HFD group and WT-HFD-SHAM
group. *P<0.05. (B) The graph presents comparisons in
food consumption in the WT-ND, WT-HFD, WT-HFD-ACE, WT-HFD-SHAM,
KO-ND and KO-HFD groups. Significant increases of food intake in
all normal mice chow, WT-ND and KO-ND groups compared to the
HFD-fed mouse groups were observed. *P<0.05. A
significant difference in food consumption was also observed for
the KO-HFD group when compared with the WT-HFD, WT-HFD-ACE and
WT-HFD-SHAM groups. There was significant decrease of food intake
in the WT-HFD-ACE group compared with the WT-HFD group in Week 8.
*P<0.05. WT, wild-type; ND, normal diet; HFD,
high-fat diet; ACE, acupoint catgut embedding; KO, knockout. |
Effect of ACE treatment on tissue
weight
The tissue weights, including the weights of adipose
tissue and associated organs, were measured and recorded
immediately following euthanasia. Statistically significant
differences in the weight of adipose tissue were consistent with
differences in net body weight. Compared with those in the ND-fed
groups, significant increases in the total recorded BAT, sWAT, and
vWAT weights were observed in the WT-HFD-fed groups (Fig. 2B-D). The data revealed significant
decreases in the weights of BAT and vWAT in the WT-HFD-ACE groups
compared with those in the WT-HFD group. Furthermore, there was
significant decrease in liver weight in the WT-ND, the TRPV1 KO and
both the ND and HFD groups when compared with that of the other
three WT groups. Conversely, there were significant decreases in
net kidney weight observed in the KO-ND and KO-HFD groups compared
with that in the WT-ND, WT-HFD, WT-HFD-ACE, and WT-HFD-SHAM groups.
No statistically significant differences were observed among the
four WT groups.
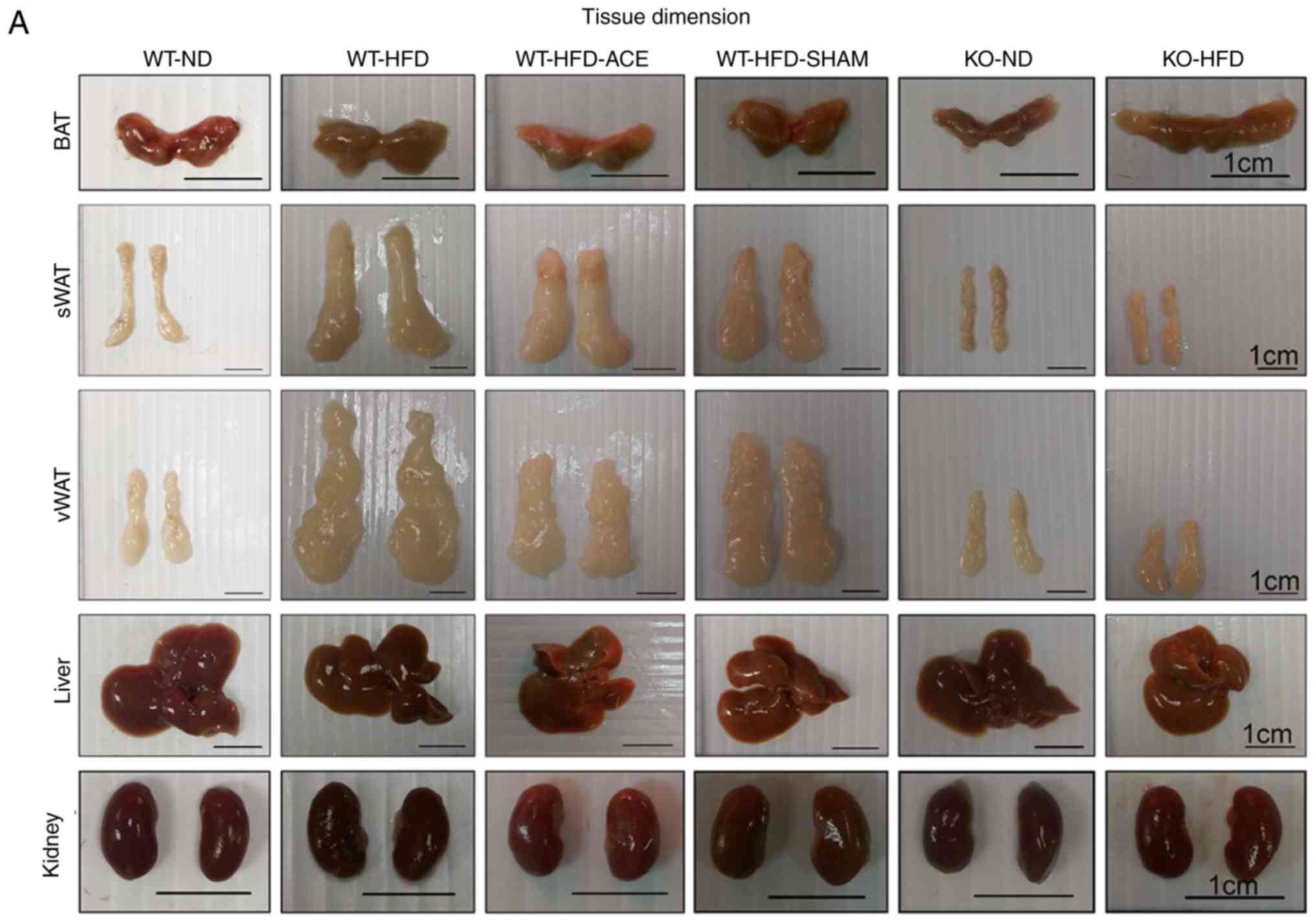 | Figure 2BAT, sWAT, vWAT, liver and kidney
weight and dimension in the six subject groups. The comparisons in
(A) tissue dimension and (B-F) tissue weight in the WT-ND, WT-HFD,
WT-HFD-ACE, WT-HFD-SHAM, KO-ND and KO-HFD groups. Significantly
increased BAT and vWAT levels were observed in the WT-HFD and
WT-HFD-SHAM groups compared with the WT-ND, WT-HFD-ACE, KO-ND and
KO-HFD groups, which exhibited significantly smaller values. The
sWAT and liver results presented significant decreases in the
WT-ND, KO-ND, and KO-HFD groups, whilst only a significant decrease
in kidney weight was observed in two KO groups.
*P<0.05. BAT, brown adipose tissue; sWAT,
subcutaneous white adipose tissue; vWAT, visceral white adipose
tissue; WT, wild-type; ND, normal diet; HFD, high-fat diet; ACE,
acupoint catgut embedding; KO, knockout. |
Glucose, insulin and leptin plasma level
changes following a 12 h fast
At 12 h, there was a significant difference in
fasting plasma glucose levels in the WT-HFD and WT-HFD-SHAM groups,
which were induced to become obese without any treatment
interventions. However, plasma glucose levels were significantly
diminished in the WT-HFD-ACE group following the receipt of
therapy, and in the KO-HFD group. Analysis of the insulin plasma
concentrations in the four groups revealed that the levels of this
hormone were significantly decreased in the WT-HFD-ACE group
compared with those in the WT-HFD and WT-HFD-SHAM groups. There was
also a significant decrease in insulin plasma levels in the KO-HFD
group compared with that in the other groups. Additionally, the
levels of leptin, which is a hormone released by adipocytes to
produce feelings of satiety, exhibited the same pattern to that
observed in the insulin plasma graph. The changes in leptin plasma
levels within the WT-HFD, WT-HFD-ACE, WT-HFD-SHAM, and KO-HFD
groups are presented in Fig.
3C.
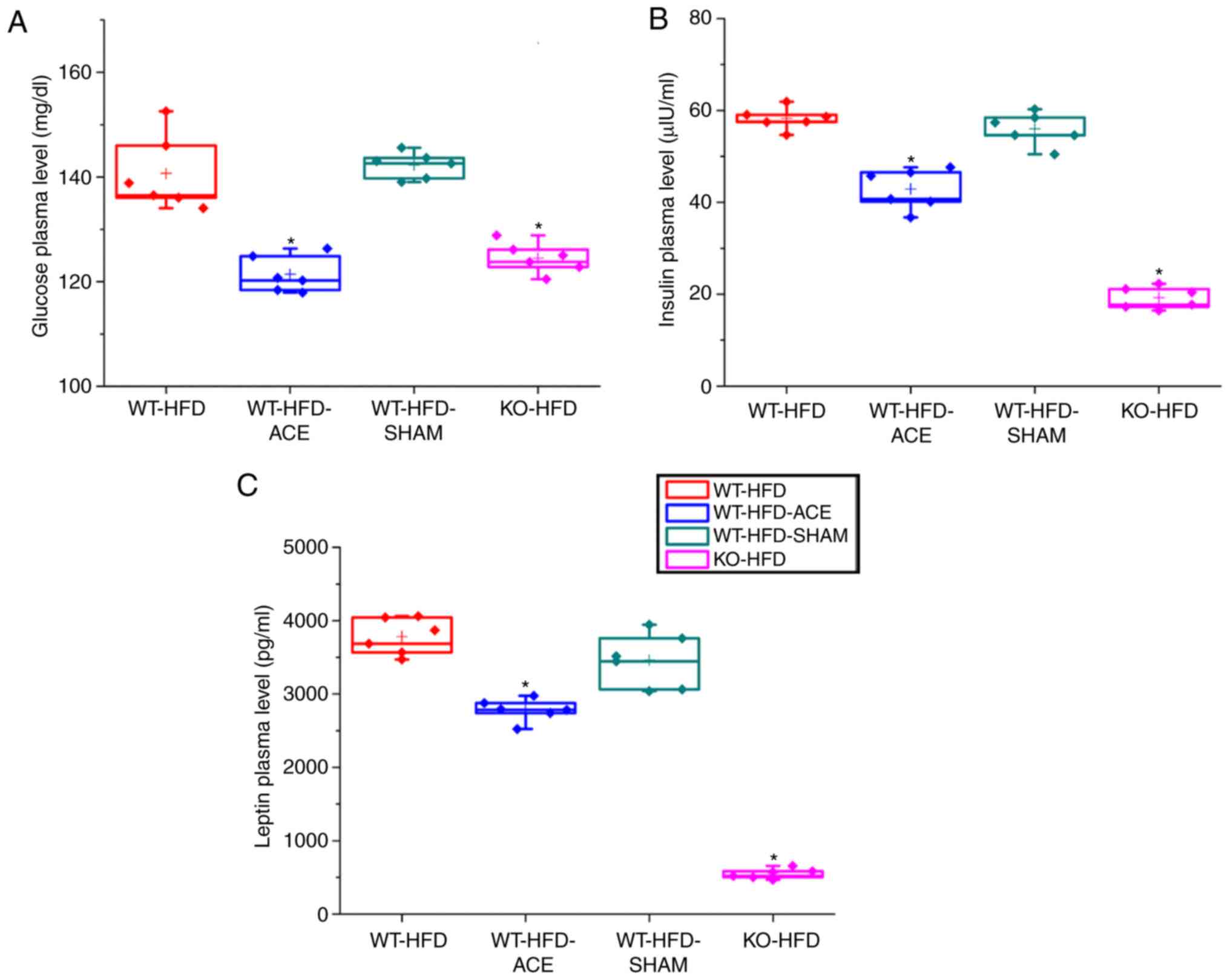 | Figure 3ELISA results for fasting glucose
insulin and leptin plasma level. A graphs present the comparisons
in fasting plasma levels of (A) glucose (B) insulin and (C) leptin
in the WT-HFD, WT-HFD-ACE, WT-HFD-SHAM and KO-HFD groups. There
were significant increases in fasting glucose plasma levels in the
WT-HFD and WT-HFD-SHAM groups. *P<0.05. Conversely,
significantly decreased plasma levels of glucose, insulin, and
leptin were observed in the WT-HFD-ACE and KO-HFD groups, which
also demonstrated decreased body weight. *P<0.05. WT,
wild-type; ND, normal diet; HFD, high-fat diet; ACE, acupoint
catgut embedding; KO, knockout. |
Effect of ACE treatment on proteins of
TRPV1 and associated molecules in the hypothalamus
Western blot analysis was used to determine the
effect of ACE treatment on the expression levels of the TRPV1
receptor and associated downstream molecules. TRPV1 protein levels
significantly decreased following the receipt of treatment. This
decrease was observed in the WT-HFD-ACE and KO-HFD groups. A
significant increase in TRPV1 expression was observed in the WT-HFD
and WT-HFD-SHAM treatment groups, which indicated increased amounts
of adipose tissue storage, and hence weight gain. The levels of
p-PI3K, p-Akt and p-mTOR were significantly increased in the WT-HFD
and WT-HFD-SHAM groups. However, the levels of these phosphorylated
proteins were significantly decreased in the WT-HFD-ACE and KO-HFD
groups. Similarly, a significant decrease in levels of
phosphorylated proteins from the MAPK pathway, specifically p-ERK,
pp38 and p-JNK, was observed. There were also significant decreases
in levels of p-PKCε and p-PKAIIα in the WT-HFD-ACE and KO-HFD
groups when compared to those in the WT-HFD groups (Fig. 4E and F). There was a significant
increase in p-PKCε protein expression in the WT-HFD-SHAM group
compared with that in the WT-HFD-ACE group. In the nucleus, there
was a correspondingly significant difference of p-NF-κB protein
levels between the obese groups; specifically, there were increased
levels of p-NF-κB in the WT-HFD and WT-HFD-SHAM groups compared
with those in the WT-HFD-ACE and KO-HFD groups. In addition, p-CREB
expression exhibited a similar pattern to that observed in the
protein expression levels of the WT-HFD, WT-HFD-ACE, WT-HFD-SHAM
and KO-HFD groups (Fig. 4K).
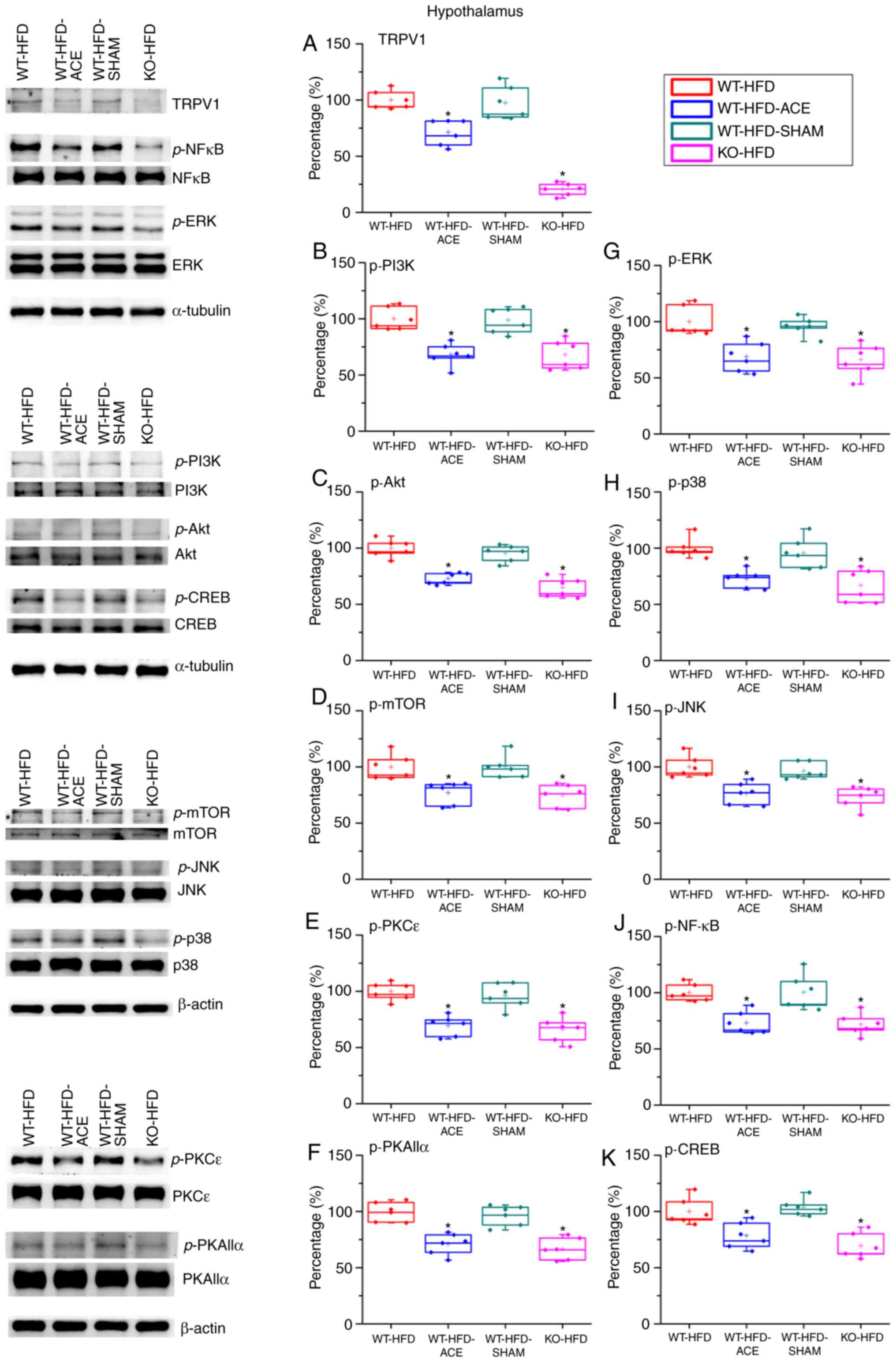 | Figure 4Expression levels of TRPV1 and
associated molecules in the hypothalamus. The expression pattern of
TRPV1 protein was detected in the following groups: WT-HFD;
WT-HFD-ACE; WT-HFD-SHAM; and KO-HFD. The results revealed
significant increases in (A) TRPV1, (B) p-PI3K, (C) p-Akt, (D)
p-mTOR, (E) p-PKCε, (F) p-PKAIIα, (G) p-ERK, (H) p-p38, (I) p-JNK,
(J) p-NF-κB and (K) p-CREB expression levels in the WT-HFD and
WT-HFD-SHAM groups compared with the other groups
(*P<0.05). These increases were statistically
decreased in the WT-HFD-ACE group that received ACE treatment and
in the KO-HFD group that lacked the TRPV1 receptor.
*P<0.05. TRPV1, transient receptor vanilloid member
1; WT, wild-type; ND, normal diet; HFD, high-fat diet; ACE,
acupoint catgut embedding; KO, knockout; p, phosphorylated; PKCε,
protein kinase C epsilon type; PKAIIα, protein kinase AII α; ERK,
extracellular signal-regulated kinase; p38, p38 mitogen-activated
protein kinase; JNK, c-Jun N-terminal kinase; CREB, cyclic
AMP-response element binding protein. |
Effect of ACE treatment on proteins of
TRPV1 and associated molecules in the PFC
The expression of the proteins in the PFC was
examined to determine the association between obesity and
corresponding psychological problems, such as depression, mediated
through TRPV1-associated signaling molecules. There was a
significant increase in TRPV1 expression levels in the WT-HFD-ACE
treatment group compared with that in the WT-HFD and WT-HFD-SHAM
treatment groups. The KO-HFD group exhibited a significant decrease
in the protein level compared with that in the other three groups.
The data revealed contrasting results between the hypothalamus and
PFC. It was identified that the p-PI3K, p-Akt and p-mTOR expression
levels were significantly decreased in the WT-HFD and WT-HFD-SHAM
treatment group but were significantly increased in the WT-HFD-ACE
and KO-HFD groups. Additionally, the expression levels of p-ERK,
p-p38 and p-JNK exhibited similar trends to other proteins. The
levels of p-ERK, p-p38 and p-JNK were significantly increased in
the WT-HFD-ACE and KO-HFD groups. This increase was abolished in
the WT-HFD and WT-HFD-SHAM groups when compared with the other two
groups, WT-HFD-ACE and KO-HFD. There was a statistical decrease in
the expression of p-PKCε and p-PKAIIα between the WT-HFD and
WT-HFD-SHAM groups (Fig. 5F).
However, much larger significant increases in the WT-HFD-ACE and
KO-HFD groups were observed. Lastly, the levels of p-NF-κB and
p-CREB were investigated, and a significant variation between the
obese groups and groups with normal body weight was observed.
Increased values were observed in the WT-HFD-ACE and KO-HFD groups,
whereas decreased values were identified in the WT-HFD and
WT-HFD-SHAM groups.
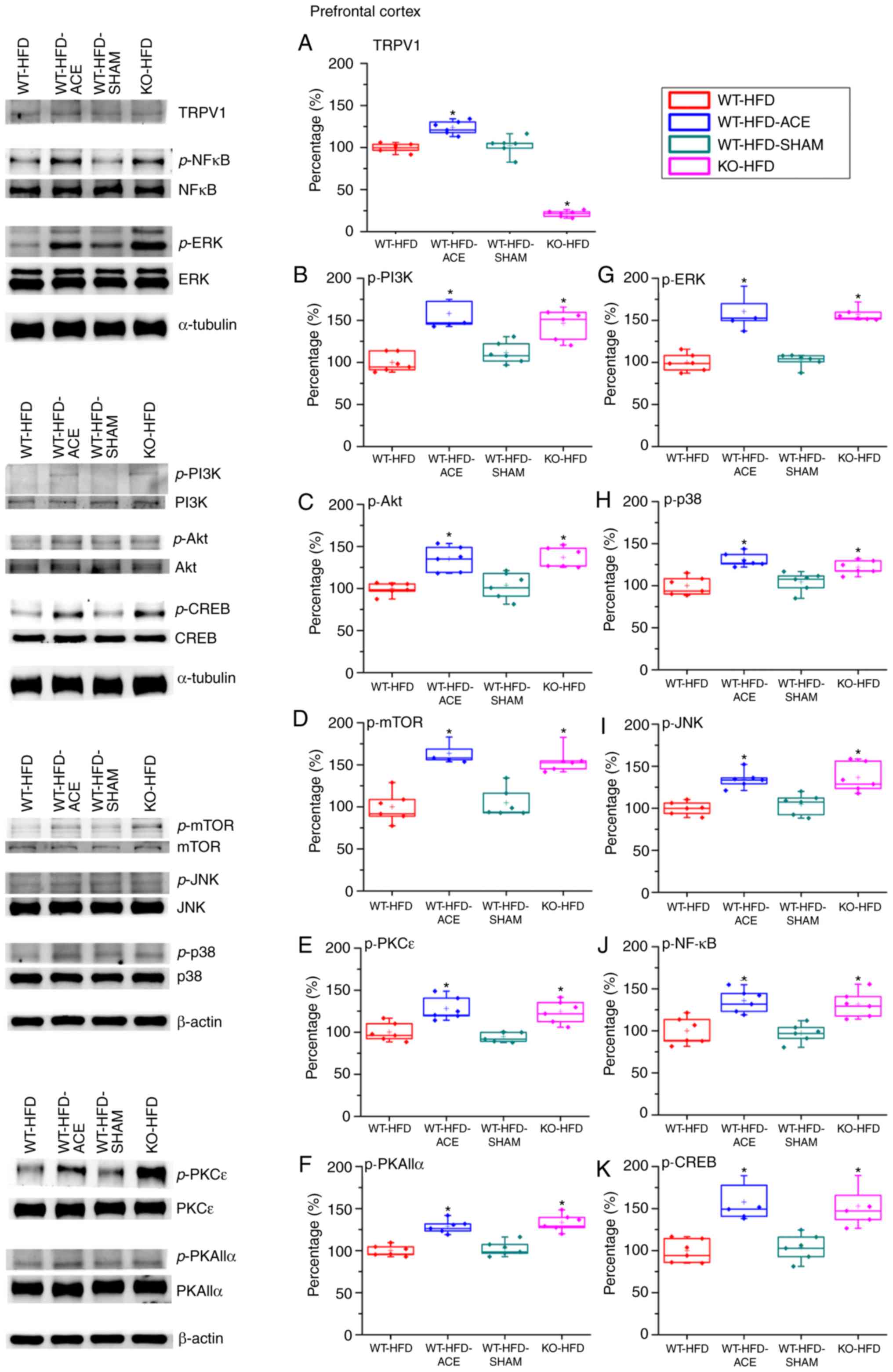 | Figure 5Expression levels of TRPV1 and
associated molecules in the PFC. The expression pattern of TRPV1
protein was detected in the following groups: WT-HFD; WT-HFD-ACE;
WT-HFD-SHAM; and KO-HFD. The results demonstrated significant
decreases in (A) TRPV1 expression in WT-HFD, WT-HFD-SHAM and KO-HFD
groups when compared with the WT-HFD-ACE group, which demonstrated
a significant increase following ACE treatment.
*P<0.05. The results also demonstrated significant
decreases in (B) p-PI3K (C) p-Akt (D) p-mTOR (E) p-PKCε (F)
p-PKAIIα (G) p-ERK (H) p-p38 (I) p-JNK (J) p-NF-κB and (K) p-CREB
expression levels in the WT-HFD and WT-HFD-SHAM groups compared
with the other groups. *P<0.05. The expression levels
of these proteins were statistically increased in the WT-HFD-ACE
and KO-HFD groups. *P<0.05. TRPV1, transient receptor
vanilloid member 1; WT, wild-type; ND, normal diet; HFD, high-fat
diet; ACE, acupoint catgut embedding; KO, knockout; PFC, prefrontal
cortex; p, phosphorylated; PKCε. protein kinase C epsilon type;
PKAIIα, protein kinase AII α; ERK, extracellular signal-regulated
kinase; p38, p38 mitogen-activated protein kinase; JNK, c-Jun
N-terminal kinase; CREB, cyclic AMP-response element binding
protein. |
Effect of weight control on TRPV1 and
associated molecules in the hypothalamus and NTS
Immunofluorescence was used to measure the
expression of TRPV1-associated downstream proteins in the
hypothalamus and NTS, which is the center that regulates appetite
and satiety in the brain. There was a significant increase in TRPV1
and p-PKAIIα immunoreactivity within the hypothalamic area of the
WT-HFD and WT-HFD-SHAM groups. However, these protein levels
significantly decreased in the WT-HFD-ACE and KO-HFD groups
(Fig. 6A). Additionally, levels
of p-PI3K and p-CREB (Fig. 6B)
exhibited similar patterns of expression in these treatment groups.
Decreased levels of TRPV1, p-PKAIIα, p-PI3K and p-CREB were also
detected in the NTS (Fig. 7A and
B).
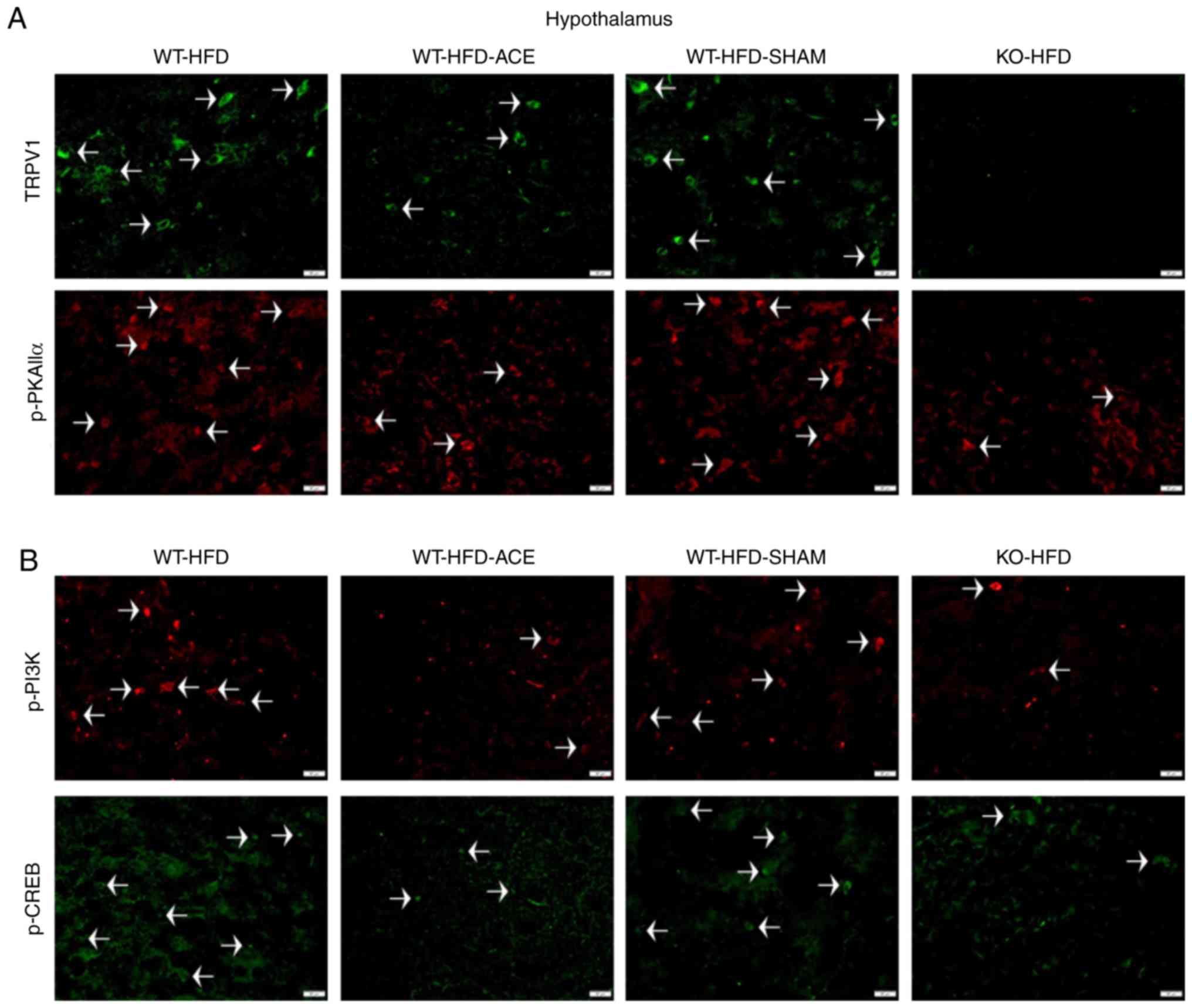 | Figure 6Expression levels of TRPV1, p-PI3K,
p-CREB and p-PKAIIα in the hypothalamus. (A) Representative
immunofluorescence staining of TRPV1 (green) and p-PKAIIα (red) and
(B) representative immunofluorescence staining of p-PI3K (red) and
p-CREB (green) were performed in the hypothalamus of subjects in
the WT-HFD, WT-HFD-ACE, WT-HFD-SHAM and KO-HFD groups. White
arrowheads indicate immunopositive cells. TRPV1, transient receptor
vanilloid member 1; WT, wild-type; ND, normal diet; HFD, high-fat
diet; ACE, acupoint catgut embedding; KO, knockout; p,
phosphorylated; PKAIIα, protein kinase AII α; CREB, cyclic
AMP-response element binding protein. |
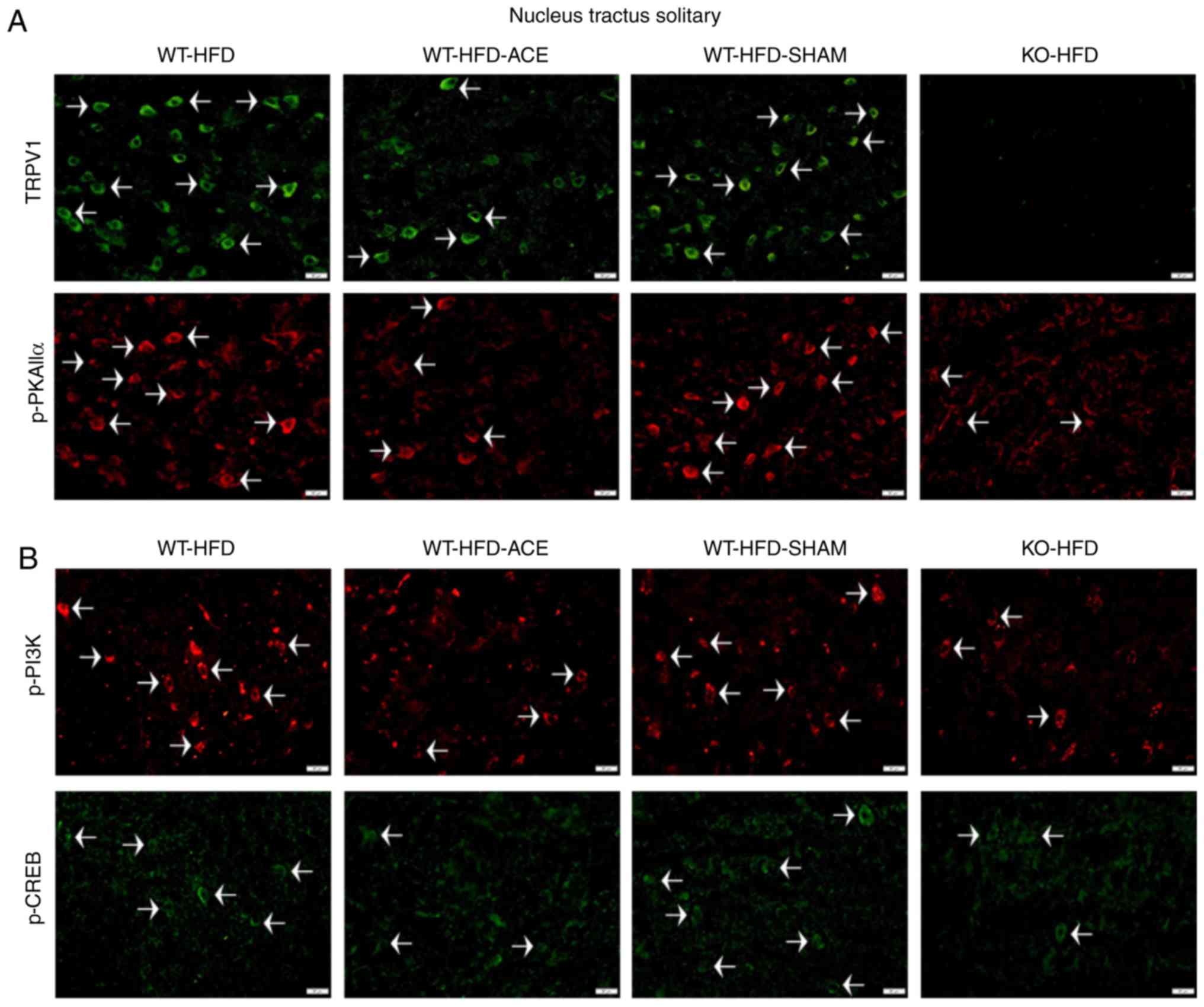 | Figure 7Expression levels of TRPV1, p-PI3K,
p-CREB and p-PKAIIα in the NTS. (A) Representative
immunofluorescence staining of TRPV1 (green) and p-PKAIIα (red) and
(B) representative immunofluorescence staining of p-PI3K (red) and
p-CREB (green) were performed in the NTS of subjects in the WT-HFD,
WT-HFD-ACE, WT-HFD-SHAM and KO-HFD groups. White arrowheads
indicate immunopositive cells. TRPV1, transient receptor vanilloid
member 1; WT, wild-type; ND, normal diet; HFD, high-fat diet; ACE,
acupoint catgut embedding; KO, knockout; p, phosphorylated; PKAIIα,
protein kinase AII α; CREB, cyclic AMP-response element binding
protein; NTS, nucleus tractus solitarii. |
Discussion
Obesity, which has a very high incidence rate, is
largely attributed to sedentary lifestyles among the global
population. Furthermore, high obesity rates have also been
attributed to poor nutrition and decreased physical activity among
individuals. Obesity can lead to various complications that not
only affect public health but also create an economic burden in the
form of increased healthcare costs. ACE treatment produces a
lasting and potent stimulatory effect, thereby decreasing the
treatment frequency when compared with other techniques of
acupuncture. In clinical therapy, 14 days between each treatment is
the interval commonly used in weight control (21). Therefore, in the present study,
the catgut absorption ability in a mouse model was determined prior
to initiation of the experimental model. In the present study, a
time period of 7 days was established to be the optimal interval
between treatments, as it is the shortest time in which the catgut
can be completely absorbed. TRPV1, a calcium ion channel, is
sensitively activated by capsaicin and heat, and it is distributed
in various areas of the brain, including the hypothalamus, NTS and
PFC. TRPV1 is heavily involved in regulating the MAPK signaling
pathway. Previous studies aimed to identify the involvement of the
TRPV1 receptor in obesity. Capsaicin and associated substances
increased the activation of BAT, amplified energy expenditures, and
induced the browning of WAT (28,29). Dietary capsaicin also possesses
the ability to prevent adipogenesis, particularly vWAT, and also
promote lipolysis (30). These
results from previous studies suggest that TRPV1 promotes weight
reduction by elevating energy expenditures and decreasing food
consumption (9,31).
In the present study, body mass, tissue weight and
food consumption were examined to analyze feeding behavior. The
results of the WT-HFD and WT-HFD-SHAM groups revealed similar
increases in body mass and tissue weight, including increased
weights for BAT, sWAT, vWAT, and in the liver, but not in the
kidneys. There were significant decreases in BAT and vWAT in the
WT-HFD-ACE treatment group compared with those in the WT-HFD and
WT-HFD-SHAM groups. There was a significant decrease in food
consumption in the WT-HFD-ACE group during the last week of
treatment when compared with that in the WT-HFD-fed groups that did
not undergo catgut insertion. Furthermore, the results revealed
that the KO-ND and KO-HFD groups exhibited significant decreases in
weight when compared with those in the other four WT groups. Even
though the KO groups did not receive any medication, the results
from these groups were similar to those of the WT-HFD-ACE group.
These results are supported by the results of previous studies
suggesting that TRPV1 deletion protects against obesity-inducing
diets (32-35). Marshall et al (36) demonstrated that although there was
similarity between the body weights of WT and KO groups on an HFD,
KO mice were protected against obesity-inducing hypertension,
low-grade inflammation and glucose tolerance. Other previous
studies described similar results in terms of body weight for WT
and KO mice fed an HFD (8,37).
These results indicated that the KO-HFD-fed mice gained
significantly more body mass compared with that in the HFD-fed WT
mice (38). Accordingly, the
western blot analysis results of the present study demonstrated
similarities in the protein expression intensities in the
hypothalamus and NTS, with greater protein density in the WT-HFD
and WT-HFD SHAM treatment groups and significantly decreased
density in the WT-HFD-ACE and KO-HFD groups. These results revealed
the role of TRPV1 signaling in both the hypothalamus and the NTS on
obesity. The fasting levels of blood, glucose, insulin and leptin
revealed a proportionately correlated interaction with body mass.
These data corroborate previous data, suggesting a close
association between TRPV1 and glucose tolerance during obesity
(36). Previous studies have
identified the role of TRPV1 antagonists in promoting insulin
secretion and improving insulin resistance (39-41). Additionally, a previous study
examining supplementary outcomes demonstrated that TRPV1 actively
participates in the regulation of leptin signaling (42). On the basis of these conflicting
results concerning the effect of TRPV1 on the regulation of
obesity, the results of the present study suggested that TRPV1
expression serves an integral role in the regulation and
maintenance of body weight. Given the stark differences in the data
from various previous studies, the association between TRPV1 and
obesity remains an interesting avenue of study.
The association between obesity and depression, an
important psychological problem, has also been investigated.
Previous data suggest that depression is a comorbidity of obesity,
and these conditions may increase the risk of developing the other
condition (43). In addition,
adipose tissue accumulation, which is increased in females, serves
an important role in this association (24,40,44). The present study exposed the
different patterns of protein expression in the PFC, which is
commonly associated with depression. There were lower protein
levels in the WT-HFD and WT-HFD-SHAM groups compared with those in
the WT-HFD-ACE group, which underwent catgut insertion at the ST36
acupoints. However, in the KO-HFD group, only TRPV1 levels were
significantly decreased compared with those in the other WT-HFD-fed
groups. By contrast, the protein expression of the MAPK molecules
was significantly increased compared with that in the WT-HFD and
WT-HFD-SHAM groups. Similar tendencies were exhibited among groups
with decreased body weights, specifically the WT-HFD-ACE and KO-HFD
groups, which demonstrated significantly increased expression
levels of the MAPK proteins compared with those in the WT-HFD and
WT-HFD-SHAM groups. This evidence suggests that depression serves
an integral role in obesity, as indicated through the expression of
MAPK proteins.
Accordingly, obesity affects multiple organ systems,
even if there are only two causes, namely, excessive food intake
and/or the absence of physical activity. Western and alternative
medicines, such as acupuncture, are utilized to prevent the onset
of obesity. Traditional Chinese acupuncture involves the
stimulation of qi in meridians, which are also referred to
as channels. These theories have been described as the stimulation
of energy through the autonomic and central nervous systems in
terms of Western medical science. In more detail, the energy (or
qi) has be described as the release of endogenous
neurotransmitters between at least 2 cells (45-47). Various studies have comparatively
analyzed the efficacy of acupuncture for the treatment of obesity.
The results indicated that ACE, electroacupuncture and manual
acupuncture treatment significantly decreased overall body weight
and the body mass index with no distinguishable difference between
these therapeutic outcomes. By contrast, placebo or sham methods
was considerably less effective in decreasing body weight (48,49). Although behavior modification is
the ultimate solution for controlling weight, remedies that promote
and maintain the performance of this treatment are in high demand.
Generally, weight reduction requires more time in order to yield
the desired results. Therefore, a fast-acting medication with less
side effects can be combined with diet control, and exercise is
optimal for the treatment of obesity.
In summary, the results of the present study
identified significant associations between ACE treatment and
obesity that involved TRPV1 receptors and their associated
downstream signaling cascades. Additionally, the psychological
problems that may arise from obesity are mediated through
downstream MAPK signaling pathways. These changes were verified
through the measurement of body mass and the mass of adipose
tissue, TRPV1 protein expression, and the expression of proteins in
the MAPK signaling pathway. The similar tendencies of the TRPV1
receptor and the downstream pathways were demonstrated through the
significant decrease in protein levels in the hypothalamus and NTS.
This decrease in protein expression would affect food consumption,
whereas increased protein expression in the PFC would affect mental
health. Nonetheless, the present study aimed to observe the effect
of ACE treatment and protein expression changes in specific brain
areas. Therefore, the glucose, insulin and leptin plasma level
modification were studied using ELISA, in order to detect their
general involvement among body and tissue weight, food consumption
and protein expression. The present study also had limitations: The
comparison of plasma levels and protein expression were only
observed in HFD-fed subject groups, as the specified aim of the
present study was to only observe the treatment effect without diet
modification. For future studies, the comparison of diet
modification should be considered and analyzed as a secondary
independent variable, to investigate the treatment efficacy in more
detail and corroborate the results of the present study.
Therefore, the present study concluded that levels
of TRPV1, p-PI3K, p-mTOR, p-Akt, p-ERK, p-p38, p-JNK, p-PKCε,
p-PKAIIα, p-CREB and p-NF-κB are involved with the development of
obesity, and they may be regulated by ACE treatment. In addition,
the deletion of the TRPV1 gene decreased the risk of
developing obesity regardless of food intake, as demonstrated by
the comparison between ND and HFD feeding regimes. Lastly, the ACE
treatment at the bilateral ST36 acupoint promotes weight control by
decreasing food intake.
Funding
The present study was financially supported by MOST
107-2320-B-039-033.
Availability of data and materials
The data used to support the findings of this study
are available from the corresponding author upon request.
Authors' contributions
CI designed the protocol, performed the experiments
and analyzed and wrote the manuscript. YL and YH provided technical
guidance and advice on the study. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The use of these animals was approved by Institute
of Animal Care and USE Committee of China Medical University
(Permit no. 2016-061), Taiwan, following the Guide for the use of
Laboratory Animals (National Academy Press).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
ACE
|
acupoint catgut embedding
|
|
ST36
|
Zusanli acupoint
|
|
NTS
|
nucleus tractus solitarii
|
|
PFC
|
prefrontal cortex
|
|
sWAT
|
subcutaneous white adipose tissue
|
|
vWAT
|
visceral white adipose tissue
|
|
BAT
|
brown adipose tissue
|
|
WT
|
wild type mouse
|
|
KO
|
TRPV1 knockout mouse
|
|
ND
|
normal diet
|
|
HFD
|
high fat diet
|
Acknowledgments
The authors would like to thank Bernice Lottering,
Pei-Hsuan Chen and Hsin-Ping Ku (all China Medical University,
Taichung, Taiwan) for their support with the present study. In
addition, the authors would also like to thank Panupong Ngamwong
(Chao Phya Abhaibhubejhr Hospital Foundation, Prachin Buri,
Thailand), for their expertise with the image editing.
References
|
1
|
GBD 2015 Obesity Collaborators; Afshin A,
Forouzanfar MH, Reitsma MB, Sur P, Estep K, Lee A, Marczak L,
Mokdad AH, Moradi-Lakeh M, et al: Health effects of overweight and
obesity in 195 countries over 25 years. N Engl J Med. 377:13–27.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cho SH, Lee JS, Thabane L and Lee J:
Acupuncture for obesity: A systematic review and meta-analysis. Int
J Obes (Lond). 33:183–196. 2009. View Article : Google Scholar
|
|
3
|
Thaler JP, Guyenet SJ, Dorfman MD, Wisse
BE and Schwartz MW: Hypothalamic inflammation: Marker or mechanism
of obesity pathogenesis? Diabetes. 62:2629–2634. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Coveleskie K, Kilpatrick LA, Gupta A,
Stains J, Connolly L, Labus JS, Sanmiguel C and Mayer EA: The
effect of the GLP-1 analogue Exenatide on functional connectivity
within an NTS-based network in women with and without obesity. Obes
Sci Pract. 3:434–445. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lockie SH: Glucagon-like peptide-1
receptor in the brain: Role in neuroendocrine control of energy
metabolism and treatment target for obesity. J Neuroendocrinol.
25:597–604. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sobrino Crespo C, Perianes Cachero A,
Puebla Jiménez L, Barrios V and Arilla Ferreiro E: Peptides and
food intake. Front Endocrinol (Lausanne). 5:582014. View Article : Google Scholar
|
|
7
|
Valassi E, Scacchi M and Cavagnini F:
Neuroendocrine control of food intake. Nutr Metab Cardiovasc Dis.
18:158–168. 2008. View Article : Google Scholar
|
|
8
|
Baskaran P, Krishnan V, Fettel K, Gao P,
Zhu Z, Ren J and Thyagarajan B: TRPV1 activation counters
diet-induced obesity through sirtuin-1 activation and PRDM-16
deacetylation in brown adipose tissue. Int J Obes (Lond).
41:739–749. 2017. View Article : Google Scholar
|
|
9
|
Yoneshiro T, Aita S, Kawai Y, Iwanaga T
and Saito M: Nonpungent capsaicin analogs (capsinoids) increase
energy expenditure through the activation of brown adipose tissue
in humans. Am J Clin Nutr. 95:845–850. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Abdelhamid RE, Kovács KJ, Nunez MG and
Larson AA: Depressive behavior in the forced swim test can be
induced by TRPV1 receptor activity and is dependent on NMDA
receptors. Pharmacol Res. 79:21–27. 2014. View Article : Google Scholar :
|
|
11
|
Li HB, Mao RR, Zhang JC, Yang Y, Cao J and
Xu L: Antistress effect of TRPV1 channel on synaptic plasticity and
spatial memory. Biol Psychiatry. 64:286–292. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ramírez-Barrantes R, Cordova C, Poblete H,
Muñoz P, Marchant I, Wianny F and Olivero P: Perspectives of TRPV1
function on the neurogenesis and neural plasticity. Neural Plast.
2016:15681452016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu D, Zhu Z and Tepel M: The role of
transient receptor potential channels in metabolic syndrome.
Hypertens Res. 31:1989–1995. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Menigoz A and Boudes M: The expression
pattern of TRPV1 in brain. J Neurosci. 31:13025–13027. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang GR, Zhao H, Choi EM, Svestka M, Wang
X, Cook RG and Geller AI: CaMKII, MAPK, and CREB are coactivated in
identified neurons in a neocortical circuit required for performing
visual shape discriminations. Hippocampus. 22:2276–2289. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Brown TE, Chirila AM, Schrank BR and Kauer
JA: Loss of interneuron LTD and attenuated pyramidal cell LTP in
Trpv1 and Trpv3 KO mice. Hippocampus. 23:662–671. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fernandes ES, Fernandes MA and Keeble JE:
The functions of TRPA1 and TRPV1: Moving away from sensory nerves.
Br J Pharmacol. 166:510–521. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nam MH, Lee SW, Na HY, Yoo JH, Paik SH,
Ahn KS, Ahn YM, Ahn SY, Choi SH and Lee BC: Herbal acupuncture for
the treatment of obesity. J Acupunct Meridian Stud. 9:49–57. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yeh ML, Chu NF, Hsu MY, Hsu CC and Chung
YC: Acupoint stimulation on weight reduction for obesity: A
randomized sham-controlled study. West J Nur Res. 37:1517–1530.
2015. View Article : Google Scholar
|
|
20
|
Kim SY, Shin IS and Park YJ: Effect of
acupuncture and intervention types on weight loss: A systematic
review and meta-analysis. Obes Res. 19:1585–1596. 2018. View Article : Google Scholar
|
|
21
|
Guo T, Ren Y, Kou J, Shi J, Tianxiao S and
Liang F: Acupoint catgut embedding for obesity: Systematic review
and meta-analysis. Evid Based Complement Alternat Med.
2015:4019142015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu X, Mo Q, He T, Zhi N, Huang Y and Yang
S: Acupoint catgut embedding for the treatment of obesity in
adults: A systematic review protocol. Medicine (Baltimore).
98:e146102019. View Article : Google Scholar
|
|
23
|
Garcia-Vivas JM, Galaviz-Hernandez C,
Fernandez-Retana J, Pedroza-Torres A, Perez-Plasencia C,
Lopez-Camarillo C and Marchat LA: Transcriptomic profiling of
adipose tissue in obese women in response to acupuncture catgut
embedding therapy with moxibustion. J Altern Complement Med.
22:658–668. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the Care and Use of Laboratory Animals. 8th
edition. National Academies Press (US); Washington, DC: 2011
|
|
25
|
Choowanthanapakorn M, Lu KW, Yang J, Hsieh
CL and Lin YW: Targeting TRPV1 for body weight control using
TRPV1(-/-) mice and electroacupuncture. Sci Rep. 5:173662015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ibrahim MM: Subcutaneous and visceral
adipose tissue: Structural and functional differences. Obes Rev.
11:11–18. 2010. View Article : Google Scholar
|
|
27
|
Towbin H, Staehelin T and Gordon J:
Electrophoretic transfer of proteins from polyacrylamide gels to
nitrocellulose sheets: Procedure and some applications. Proc Natl
Acad Sci USA. 76:4350–4354. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Baskaran P, Krishnan V, Ren J and
Thyagarajan B: Capsaicin induces browning of white adipose tissue
and counters obesity by activating TRPV1 channel-dependent
mechanisms. Br J Pharmacol. 173:2369–2389. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Saito M, Yoneshiro T and Matsushita M:
Activation and recruitment of brown adipose tissue by cold exposure
and food ingredients in humans. Best Pract Res Clin Endocrinol
Metab. 30:537–547. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang LL, Yan Liu D, Ma LQ, Luo ZD, Cao
TB, Zhong J, Yan ZC, Wang LJ, Zhao ZG, Zhu SJ, et al: Activation of
transient receptor potential vanilloid type-1 channel prevents
adipogenesis and obesity. Circ Res. 100:1063–1070. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang X, Miyares RL and Ahern GP:
Oleoylethanolamide excites vagal sensory neurones, induces visceral
pain and reduces short-term food intake in mice via capsaicin
receptor TRPV1. J Physiol. 564:541–547. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cui J and Himms-Hagen J: Long-term
decrease in body fat and in brown adipose tissue in
capsaicin-desensitized rats. Am J Physiol. 262:R568–R573.
1992.PubMed/NCBI
|
|
33
|
Melnyk A and Himms-Hagen J: Resistance to
aging-associated obesity in capsaicin-desensitized rats one year
after treatment. Obes Res. 3:337–344. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Motter AL and Ahern GP: TRPV1-null mice
are protected from diet-induced obesity. FEBS Lett. 582:2257–2262.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen J, Li L, Li Y, Liang X, Sun Q, Yu H,
Zhong J, Ni Y, Chen J, Zhao Z, et al: Activation of TRPV1 channel
by dietary capsaicin improves visceral fat remodeling through
connexin43-mediated Ca2+ influx. Cardiovasc Diabetol.
14:222015. View Article : Google Scholar
|
|
36
|
Marshall NJ, Liang L, Bodkin J,
Dessapt-Baradez C, Nandi M, Collot-Teixeira S, Smillie SJ, Lalgi K,
Fernandes ES, Gnudi L and Brain SD: A role for TRPV1 in influencing
the onset of cardiovascular disease in obesity. Hypertension.
61:246–252. 2013. View Article : Google Scholar
|
|
37
|
Kentish SJ, Frisby CL, Kritas S, Li H,
Hatzinikolas G, O'Donnell TA, Wittert GA and Page AJ: TRPV1
channels and gastric vagal afferent signalling in lean and high fat
diet induced obese mice. PLoS One. 10:e01358922015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lee E, Jung DY, Kim JH, Patel PR, Hu X,
Lee Y, Azuma Y, Wang HF, Tsitsilianos N, Shafiq U, et al: Transient
receptor potential vanilloid type-1 channel regulates diet-induced
obesity, insulin resistance, and leptin resistance. FASEB J.
29:3182–3192. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tanaka H, Shimaya A, Kiso T, Kuramochi T,
Shimokawa T and Shibasaki M: Enhanced insulin secretion and
sensitization in diabetic mice on chronic treatment with a
transient receptor potential vanilloid 1 antagonist. Life Sci.
88:559–563. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang S, Ma X, Zhang L, Sun H and Liu X:
Capsaicin reduces blood glucose by increasing insulin levels and
glycogen content better than capsiate in streptozotocin-induced
diabetic rats. J Agric Food Chem. 65:2323–2330. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zsombok A and Derbenev AV: TRP channels as
therapeutic targets in diabetes and obesity. Pharmaceuticals
(Basel). 9:E502016. View Article : Google Scholar
|
|
42
|
Zsombok A, Jiang Y, Gao H, Anwar IJ,
Rezai-Zadeh K, Enix CL, Münzberg H and Derbenev AV: Regulation of
leptin receptor-expressing neurons in the brainstem by TRPV1.
Physiol Rep. 2:e121602014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pratt LA and Brody DJ: Depression and
obesity in the U.S. adult household population, 2005-2010. NCHS
Data Brief. 1–8. 2014.
|
|
44
|
Derry HM, Padin AC, Kuo JL, Hughes S and
Kiecolt-Glaser JK: Sex differences in depression: Does inflammation
play a role? Curr Psychiatry Rep. 17:782015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Biella G, Sotgiu ML, Pellegata G, Paulesu
E, Castiglioni I and Fazio F: Acupuncture produces central
activations in pain regions. Neuroimage. 14:60–66. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Omura Y: Connections found between each
meridian (heart, stomach, triple burner, etc.) & organ
representation area of corresponding internal organs in each side
of the cerebral cortex; release of common neurotransmitters and
hormones unique to each meridian and corresponding acupuncture
point & internal organ after acupuncture, electrical
stimulation, mechanical stimulation (including Shiatsu), soft laser
stimulation or Qi Gong. Acupunct Electrother Res. 14:155–186. 1989.
View Article : Google Scholar
|
|
47
|
Zhang ZJ, Wang XM and McAlonan GM: Neural
acupuncture unit: A new concept for interpreting effects and
mechanisms of acupuncture. Evid Based Complement Alternat Med.
2012:4294122012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sheng J, Jin X, Zhu J, Chen Y and Liu X:
The effectiveness of acupoint catgut embedding therapy for
abdominal obesity: A systematic review and meta-analysis. Evid
Based Complement Alternat Med. 2019:97143132019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang Y, Li J, Mo G, Liu J, Yang H, Chen
X, Liu H, Cai T, Zhang X, Tian X, et al: Acupuncture and related
therapies for obesity: A network meta-analysis. Evid Based
Complement Alternat Med. 2018:95696852018. View Article : Google Scholar : PubMed/NCBI
|














