Introduction
Acute kidney injury (AKI) is a clinical syndrome
characterized by a rapid decrease in renal function accompanied by
accumulation of waste products such as urea (1,2).
The incidences of non-dialysis-requiring and dialysis-requiring AKI
in the U.S. are 522 and 29.5 per 100,000 people/years, respectively
(3). Drug-induced AKI is a common
occurrence in clinical practice because the kidney is a major
target of various toxic chemicals (4,5).
Although the incidence is difficult to estimate due to variable
patient characteristics, many newly developed drugs have distinct
mechanisms of nephrotoxicity, making it the second most common
cause of hospital-acquired AKI (4). Recent studies have revealed a close
relationship between the gut microbiome and various kidney
diseases; however, the association between intestinal bacteria and
nephrotoxic AKI remains unclear (6,7).
Cis-diammineplatinum (II) dichloride (cisplatin) is
an inorganic platinum-based chemotherapy drug that is widely used
in the treatment of solid tumors including testicular, ovarian,
cervical and head and neck cancers (5,8).
However, nephrotoxicity is major side effect of cisplatin (9,10),
which binds to DNA to form inter- and intra-stand crosslinks that
damage DNA templates or inhibit DNA synthesis and replication. The
potential for damage is even greater in rapidly dividing cells such
as cancer or epithelial cells (10); however, it can also disrupt the
intestinal epithelium, thereby increasing intestinal permeability
and causing systemic inflammation by permitting the egress of
microbes and their toxins (11,12).
Probiotics are living, non-pathogenic microbes
(bacteria or yeast) that provide beneficial nutrients to the host
(13,14). Probiotics can directly inhibit the
growth or promote death of pathogenic microorganisms by producing
anti-microbial substances such as short-chain fatty acids (SCFAs)
and have shown immunomodulatory effects in several chronic
inflammatory disease models. L. salivarius BP121 (BP121)
that was isolated from infant feces. L. salivarius has been
shown to have remarkable probiotic efficacy because it had high
survival rate in gastrointestinal tract conditions and produced
antimicrobial compounds such as SCFAs (15). Previous studies have shown that
L. salivarius produces abundant propionate/butyrate
(16,17). Additionally, in previous studies,
improved barrier function by SCFA was reported (18-20). SCFAs such as butyrate and acetate
are known to protect intestinal barrier integrity by regulating
through the activation of AMPK and TLR4. To date, there have been
no studies investigating the possible benefits of probiotics
including lactic acid bacteria (LAB) BP121 in the context of
AKI.
AST-120 is an oral spherical carbon absorbent
consisting of oral urea toxin remover and porous carbon particles.
AST-120 reduces the accumulation of indoxyl sulfate in patients by
adsorbing indole, a precursor of toxin sulfate. This function has
the effect of slowing the progress of kidney disease (21).
To address this issue, the effect of BP121 treatment
on a rat model of cisplatin-induced AKI was investigated, as well
as the mechanistic basis for the observed effects of BP121 using
Caco-2 human intestinal epithelial cells.
Materials and methods
Isolation and selection of strains from
infant feces
LAB were isolated from fresh feces of new born
infants (22). The samples were
blended with 50 ml 0.85% NaCl solution and serially diluted 10-fold
in 0.85% NaCl; each diluted solution was plated onto De Man,
Rogosa, and Sharpe (MRS) agar (BD Biosciences; Becton, Dickinson
and Company). The plates were incubated in an anaerobic jar with
Anaeropack (BD Biosciences; Becton, Dickinson and Company) at 37°C
for 72 h. White colonies on the plates were individually picked and
streaked on fresh MRS agar plates. Isolated strains were identified
based on the near-complete 16S rRNA gene sequence using the
EZ-taxon server (http://www.ezbiocloud.net).
LAB screen for indoxyl sulfate (IS)
inhibitory activity
LAB strains were screened for their ability to
reduce IS levels as previously described (23). The strains were incubated in MRS
broth with 60 µg/ml IS and the one with the highest
inhibitory activity was selected. After incubation for 72 h, IS
levels were measured with the Indican Assay kit (Sigma-Aldrich;
Merck KGaA).
Analysis of SCFA in the probiotics
conditioned medium and stool
L. salivarius BP121, L. acidophilus
BP105 and L. rhamnosus GG were used for analysis of SCFA in
conditioned medium. A total of three strains were incubated in MRS
broth at 37°C for 48 h. SCFA analyzed using a high-performance
liquid chromatography (HPLC; Hewlett-Packard HP1100 series; Agilent
Technologies, Inc.), equipped with a UV-vis detector and diode
array detector. (Colum; YMC-Pack FA 250/6 mm I.D., Mobile phase;
acetonitrile:water (85:15), pH 4.5 adjusted by 0.1% trifluoro
acetic acid; flow rate, 1.2 ml/min, Temperature 35 UV at 400 nm).
Culture medium was directly injected into the chromatograph after
filtering with 0.2 um syringe filter. Also, 200 mg stool was mixed
with 0.15 mM HCIO4 and then sonicated at 37°C and 70 KHz
for 15 min. The prepared samples were filtered with 0.2 µm
syringe filter. The filtered sample 20 µl was injected into
the HPLC. Acetate, propionate and butyrate mixture were used to
prepare a standard curve (range of 1-1,000 ppm).
Animals and cell line
A total of 48 male Sprague-Dawley rats
(SantacoBioKorea, Inc.) weighing 187-221 g (8 weeks old) were
housed two rats per cage in a room with controlled temperature
(20-24°C) and humidity (40-70%) on a 12-h light/dark cycle with
free access to food and sterile water. Experimental procedures
involving the rats were approved by the Institutional Animal Care
and Use Committee of Hoseo University [appr oval no.
HSIACUC-18-097(1)]. Caco-2 cells
were purchased from the Korean Cell Line Bank (cat. no. 30037.1)
and were maintained as a monolayer in minimal essential medium
(Thermo Fisher Scientific, Inc.) supplemented with 10% fetal bovine
serum (Thermo Fisher Scientific, Inc.), 25 mM HEPES, and 25 mM
NaHCO3 at 37°C in a humidified atmosphere of 5%
CO2.
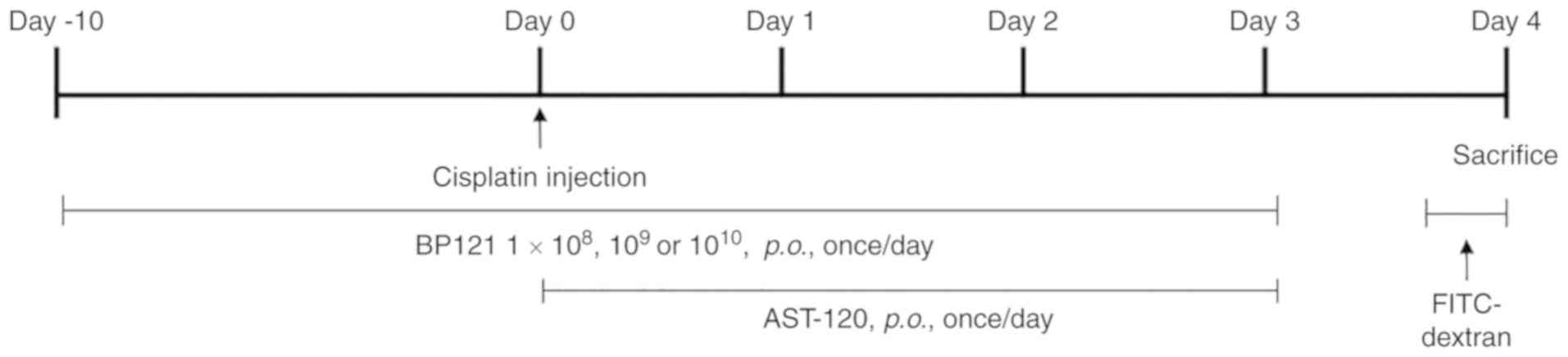
Experimental design
The 48 rats were randomly divided into six groups
(n=8 each). The AKI model was established by single intraperitoneal
(i.p.) injection of cisplatin (8 mg/kg body weight in 10 ml
saline). BP121 was administered daily by oral gavage [i.e., per os
(p.o.) administration] starting from 10 days prior to and
continuing until 4 days after cisplatin injection. Kremezin fine
granules (AST120; CJ Corp.) were orally administered at 3.6 g/kg
body weight for 4 days after cisplatin injection. The groups were
as follows: Normal control, saline (i.p.) + saline (p.o.);
cisplatin, cisplatin (i.p.) + saline (p.o.); BP121-L, cisplatin
(i.p.) + BP121 1×108 (p.o.); BP121-M, cisplatin (i.p.) +
BP121 1×109 (p.o.); BP121-H, cisplatin (i.p.) + BP121
1×1010 (p.o.), AST120, cisplatin (i.p.) + AST120 (p.o.;
Fig. 1). BP121 was obtained from.
Green Cross Wellbeing Corp., (Fig.
1).
Intestinal permeability
Intestinal permeability was measured by
administering fluorescein isothiocyanate-labeled dextran (60 mg/kg
body weight, MW = 4,000 kDa; Sigma-Aldrich; Merck KGaA) to rats by
oral gavage on day 4 after cisplatin injection. At 4 h after
gavage, 0.6 ml of blood was drawn via the jugular vein and the
fluorescence intensity of the plasma was measured using a
GloMax-Multi+ fluorophotometer (excitation/emission, 490/525 nm;
Promega Corporation).
Histological analysis
Kidney and small intestine tissues were fixed at
room temperature for 48 h in 10% neutral buffered formalin,
embedded in paraffin, and sectioned at a thickness of 4 µm.
Kidney tissue sections were stained at room temperature with
periodic acid-Schiff (PAS) (periodic acid for 5 min and Schiff
reagent for 15 min) and observed under a light microscope. The
staining intensity (reflecting the area of tissue damage) was
assessed in five randomly selected areas at ×200 magnification as
follows: None (score: 0) = normal, weak (score: 1) = <10%, mild
(score: 2) = 10-25%, moderate (score: 3) = 26-75% and strong
(score: 4) = >75% (24).
Immunohistochemical detection of monocyte chemoattractant protein
(MCP)1, B cell lymphoma 2-associated X protein (Bax) in kidneys,
zona occludins (ZO)-1, and occludin in small intestine was
performed using a commercial kit (Abcam; cat. no. ab80436) and
antibodies [all from Abcam; cat. nos. ab9668 (1:500), ab32503
(1:250), ab96587 (1:500) and ab216327 (1:500), respectively].
Apoptotic cells were detected with the terminal deoxynucleotidyl
transferase dUTP nick end labeling (TUNEL) assay using the TACS 2
TdT DAB (diami-nobenzidine) kit (Trevigen, Inc.; cat. no.
4810-30-k) according to manufacturer's protocol. The average
positive cell number was calculated by observing 10 fields at ×200
magnification using a light microscope.
Biochemical parameters
Blood samples were collected from rats and serum
levels of blood urea nitrogen (BUN), creatinine, and C-reactive
protein (CRP) were measured with a Model 7020 automated
biochemistry analyzer (Hitachi, Ltd.). IS and p-cresol
sulfate (PCS) concentrations in serum were analyzed by liquid
chromatography (ACQuity™) followed by tandem mass spectrometry (TSQ
Quantum Ultra; Thermo Fisher Scientific, Inc.) (Data SI).
Evaluation of oxidative stress and
inflammation
A 10-ml volume of phosphate-buffered saline was
added to 0.5 g kidney tissue cut into 1-2 mm pieces and
homogenized. The homogenate was centrifuged at 5,000 × g for 10 min
at 4°C; the supernatant was collected and the levels of interleukin
(IL)-6 (cat. no. MBS824560) and tumor necrosis factor (TNF)-α (cat.
no. MBS175904) (both from MyBioSource); malondialdehyde (MDA;
Cayman Chemical; cat. no. 10009055); and reduced to oxidized
glutathione ratio (GSH/GSSG; Abcam; cat. no. ab138881) were
analyzed.
Analysis of gut microbiota abundance
Fecal samples for gut microbiome profiling were
collected at 4 days after cisplatin injection and stored at -70°C
until analysis. Species were identified by 16S rRNA pyrosequecing
using the EzTaxon database at Chunlab of Seoul National University
(Seoul, South Korea). The procedure included sorting and quality
pre-screening, trimming of primer sequences, schematic structure
assignment, and chimera testing.
Effect of BP121 on tight junction (TJ)
regulation in Caco-2 cells
Caco-2 cells were treated with 20 µM
H2O2 (Volatile Free Acid Mix; Sigma-Aldrich;
Merck KGaA; cat. no. CRM46975) to cause TJ protein damage; the
culture supernatant of BP121 was used as a control. The 5′
AMP-activated protein kinase (AMPK) inhibitor Compound C (cat. no.
P5499) and the Toll-like receptor (TLR)4 inhibitor TLR-IN-C34 (cat.
no. SML0832; both from Sigma-Aldrich; Merck KGaA) were used to
evaluate the mechanism of TJ protein regulation. Each inhibitor was
applied 30 min after H2O2 treatment. SCFAs
and culture medium were added simultaneously with
H2O2. ZO-1 and occludin were detected by
immunofluorescence after incubation at 37°C for 24 h; Alexa Fluor
488 (cat. no. ab150077; 1:400) and Alexa Fluor 647 (cat. no.
ab150079; 1:400) (both from Abcam) were used as secondary
antibodies and nuclei were stained with DAPI at 37°C for 15 min
(Sigma-Aldrich; Merck KGaA; cat. no. D9542).
Statistical analysis
Data are expressed as the mean ± standard error.
Data were analyzed using SPSS (version 21.0; IBM Corp.) software.
After homogeneity of variances using Levene's test, one way ANOVA
analysis was performed to evaluate the significant differences,
with Tukey's post-hoc test. P<0.05 was considered to indicate a
statistically significant difference. Experiments were repeated 5-6
times for the in vivo investigations, or 3 times for the
in vitro investigations.
Results
Isolation and selection of strains from
infant feces
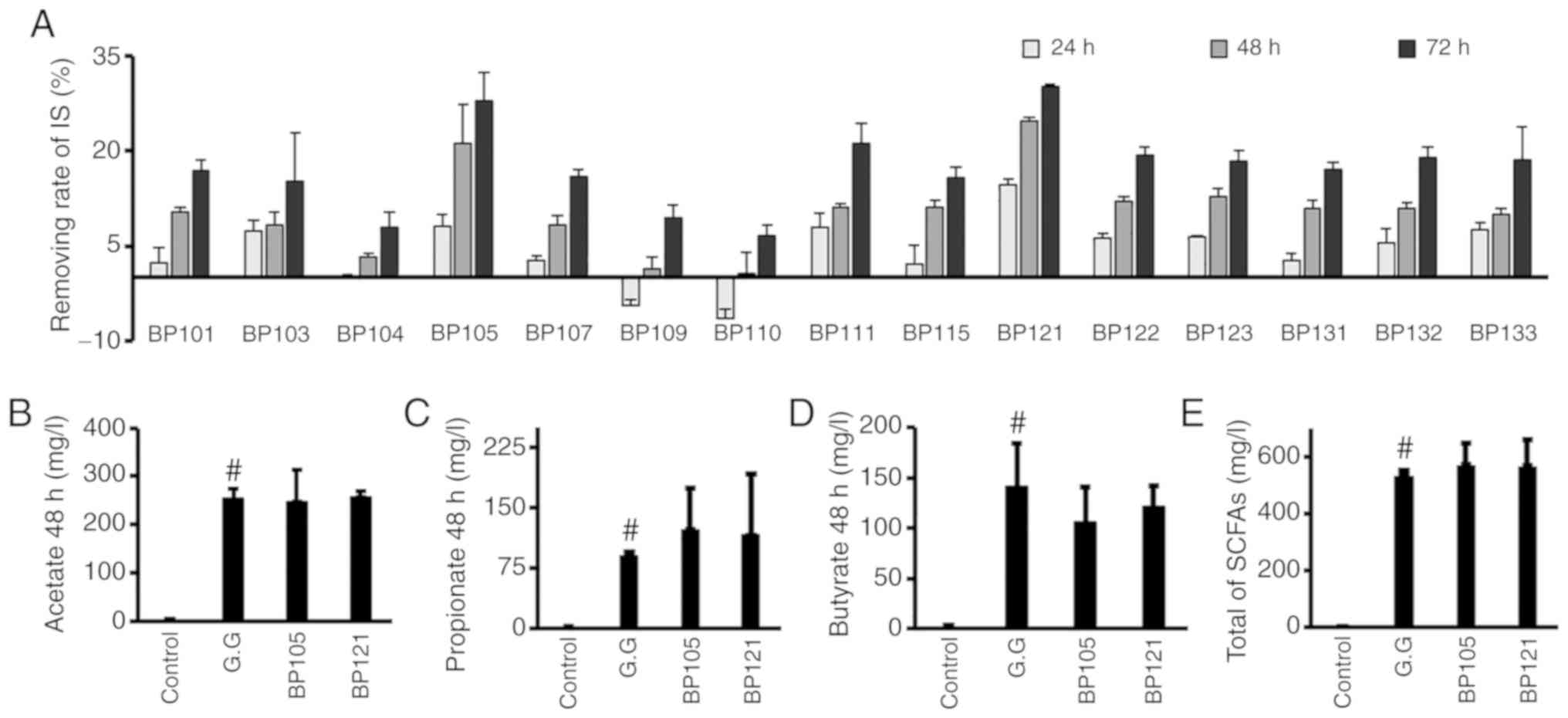
To screen for IS-inhibiting bacteria, bacteria
obtained from fresh feces of newborn infants were grown on MRS agar
plates. A total of 15 strains of LAB were selected and cultured in
MRS broth with IS for 72 h; the strain with the highest inhibitory
effect on IS was found to be BP121. This strain was able to reduce
the levels of IS by >30% after 72 h. To identify this strain,
analysis of the 16S rRNA gene sequence of BP121 (Table SI) revealed that it is closely
related to Lactobacillus salivarius JCM1231, with 99.93%
sequence similarity. After culturing BP121 and BP105 for 48 h, SCFA
production was analyzed. Both strains produced SCFA similar to
L. rhamnosus GG (Fig. 2
B-E). Based on these results, BP121 was selected as the test
strain, which showed the highest IS inhibitory effect and high SCFA
production (Fig. 2).
BP121 restores kidney weight and reduces
serum urinary toxin levels
Rats that were administered BP121 did not show any
difference in body weight compared with those that were treated
with cisplatin (Fig. 3A). On the
other hand, the increase of relative kidney weight in cisplatin
treated rats was partially decreased in rats who were administered
a high dose of BP121 (Fig. 3B).
Similar trends were observed for serum BUN and creatinine levels
(Fig. 3C and E). In addition,
BP121 caused a dose-dependent reduction in IS and PCS
concentrations (Fig. 3D and F).
AST-120 had no effect on body weight, relative kidney weight, or
blood BUN and creatinine levels, but significantly reduced IS and
PCS levels in the blood (Fig.
3).
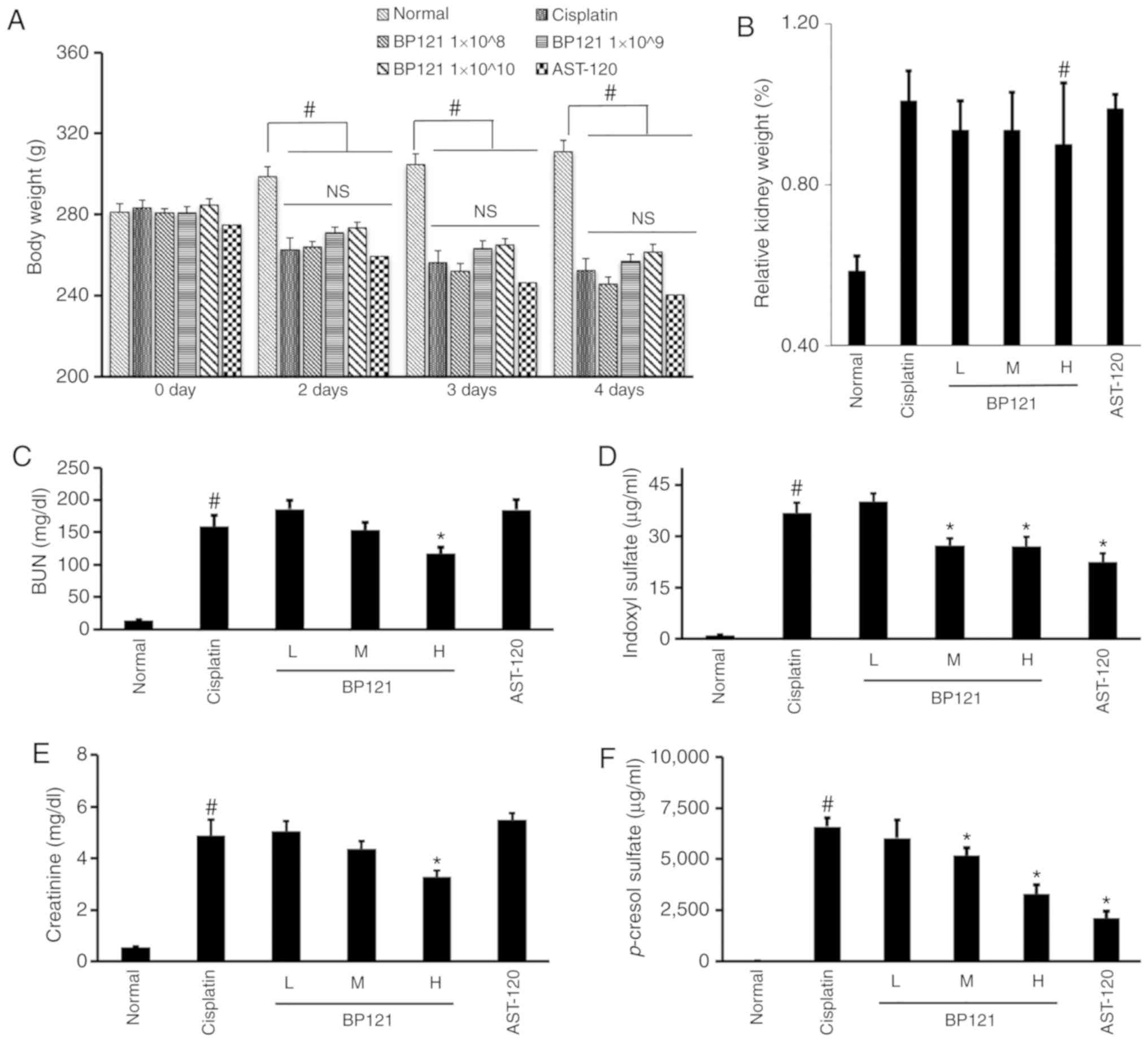 | Figure 3Physiological parameters in rats with
cisplatin-induced AKI. Changes in (A) body weight and (B) relative
kidney weight. Concentration of the urinary toxins (C) BUN, (D) IS,
(E) CREA and (F) p-cresol sulfate in rats with
cisplatin-induced AKI. L = BP121 1×108, M = BP121
1×109, H = BP121 1×1010.
#P<0.05, normal vs. cisplatin; *P<0.05,
cisplatin vs. treatment group. AKI, acute kidney injury; BUN, blood
urea nitrogen; IS, indoxyl sulfate; CREA, creatinine; NS, not
significant. |
BP121 suppresses inflammation and injury
in kidney
PAS stained kidney tissue revealed extensive renal
damage following cisplatin treatment; this was reduced by BP121-H
administration (Fig. 4A and B).
In addition, the increase in the number of MCP-1-positive cells
induced by cisplatin was reversed by BP121 (Fig. 4C and D). Although there were no
differences in serum levels of IL-6, TNF-α, endotoxin and CRP
between the cisplatin- and BP121-treated groups (Fig. 4E, G and J), IL-6 and TNF-α
concentrations in kidney tissue homogenate were significantly
reduced by BP121 treatment (Fig. 4H
and I). Despite a substantial decrease of both IS and PC,
AST-120 failed to show a renal protective effect (Fig. 4A-I).
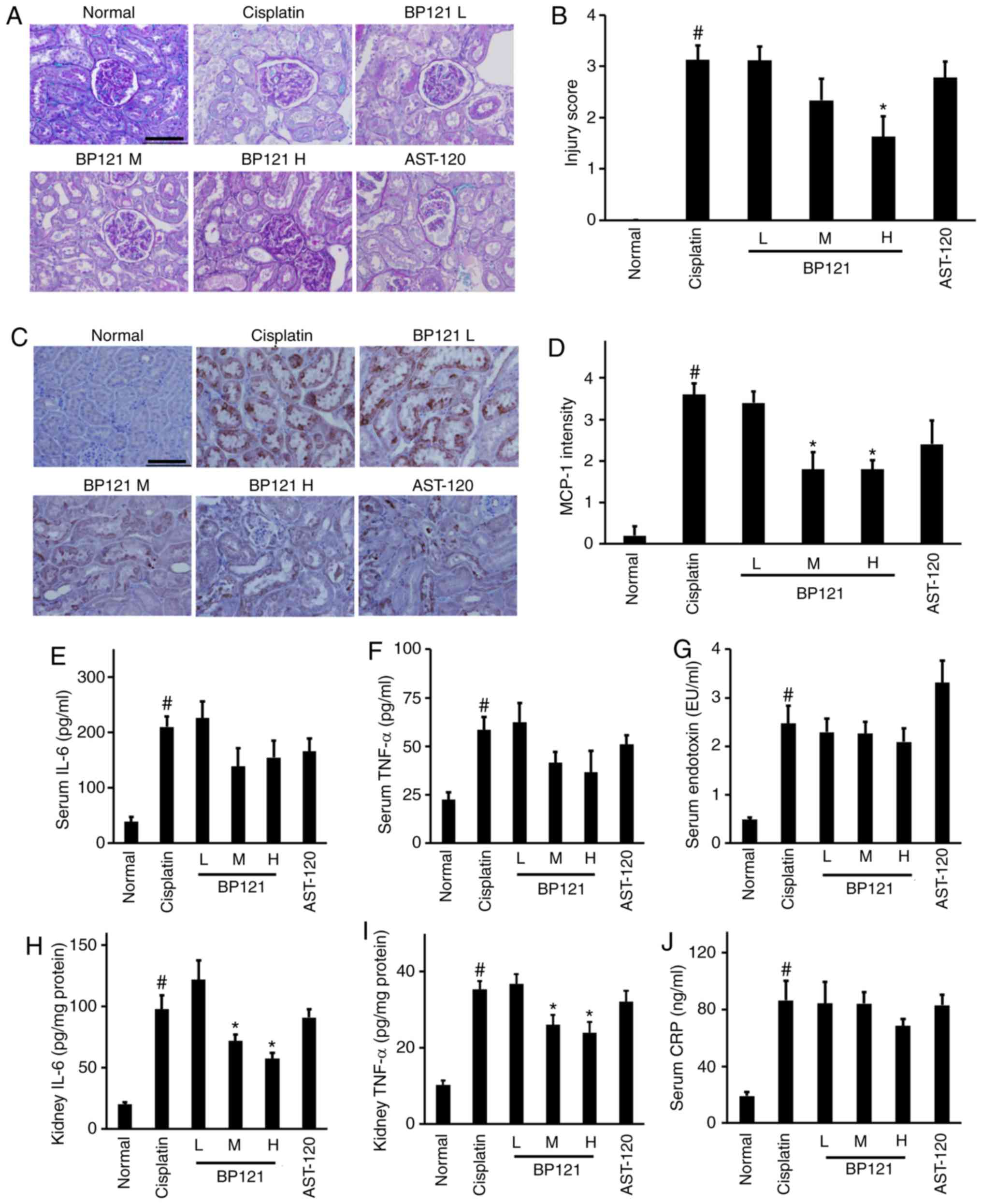 | Figure 4Histological analysis of the kidney
and IL-6, TNF-α, and CRP expression following cisplatin-induced
AKI. (A) PAS staining and (B) injury score. (C) MCP-1
immunoreactivity and (D) intensity. Concentrations of (E) serum
IL-6, (F) TNF-α, (G) serum endotoxin, (H) kidney IL-6, (I) kidney
TNF-α and (J) CRP levels were measured by ELISA. Scale bar=100
µm. #P<0.05, normal vs. cisplatin;
*P<0.05, cisplatin vs. treatment group. IL,
interleukin; TNF, tumor necrosis factor; CRP, c-reactive protein;
PAS, periodic acid Schiff; AKI, acute kidney injury. |
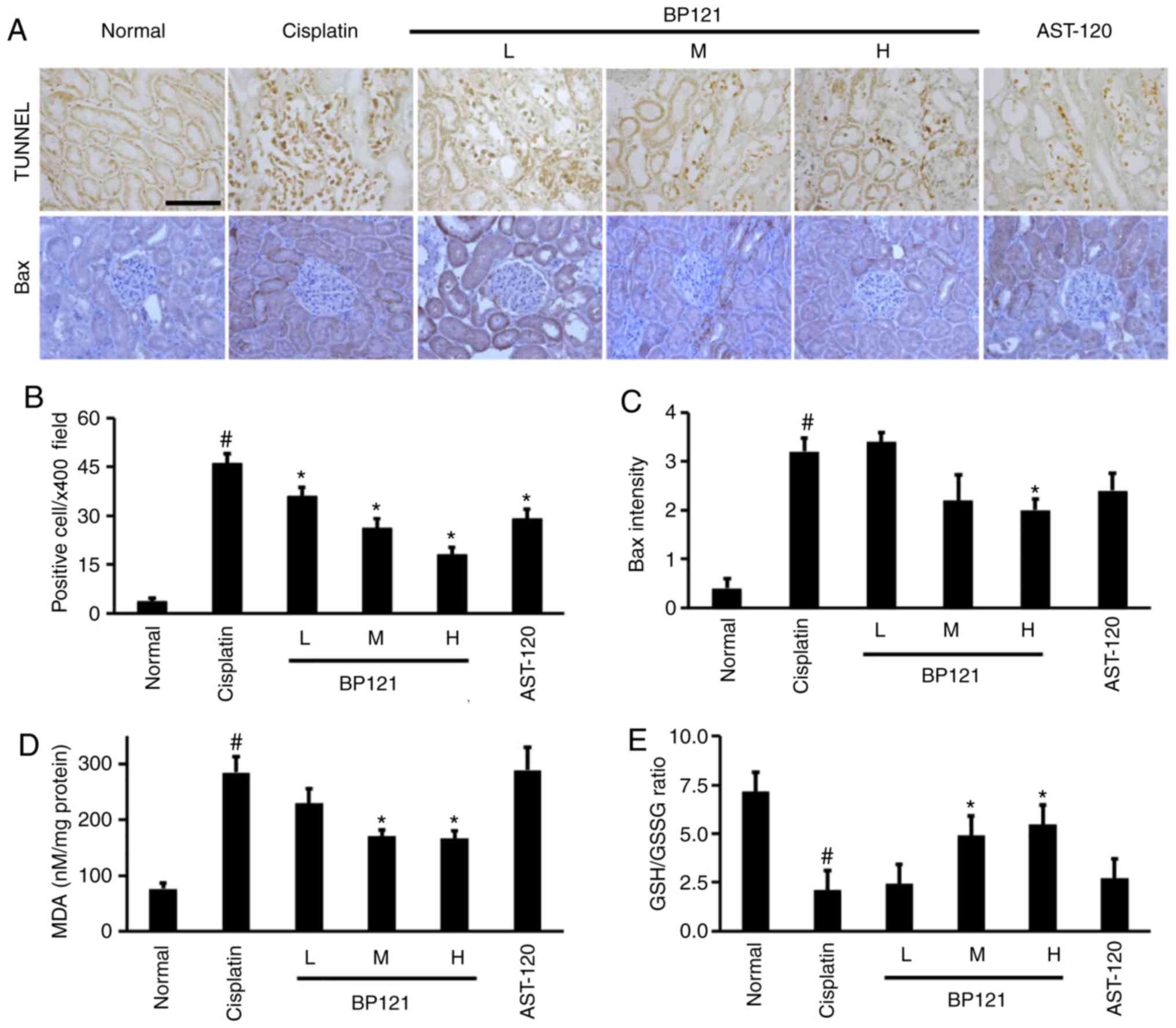
BP121 inhibits apoptosis and oxidative
stress in the kidney
The number of TUNEL positive apoptotic cells
significantly increased in cisplatin induced AKI and BP121
treatment dose dependently reduced apoptosis.
A similar trend was observed in the expression of
the pro-apoptotic protein Bax, which was upregulated in the kidney
of cisplatin-treated rats and downregulated upon administration of
a high dose BP121 (Fig. 5A-C).
The MDA level and GSH/GSSG ratio were also measured as indices of
oxidative damage and found that lipid peroxidation (MDA level) was
increased whereas GSH/GSSG ratio was decreased by cisplatin
injection; these changes were partially reversed by pretreatment
with medium and high concentrations of BP121 (Fig. 5D and E). No changes in oxidative
stress relative to the cisplatin-treated group was observed upon
AST-120 administration.
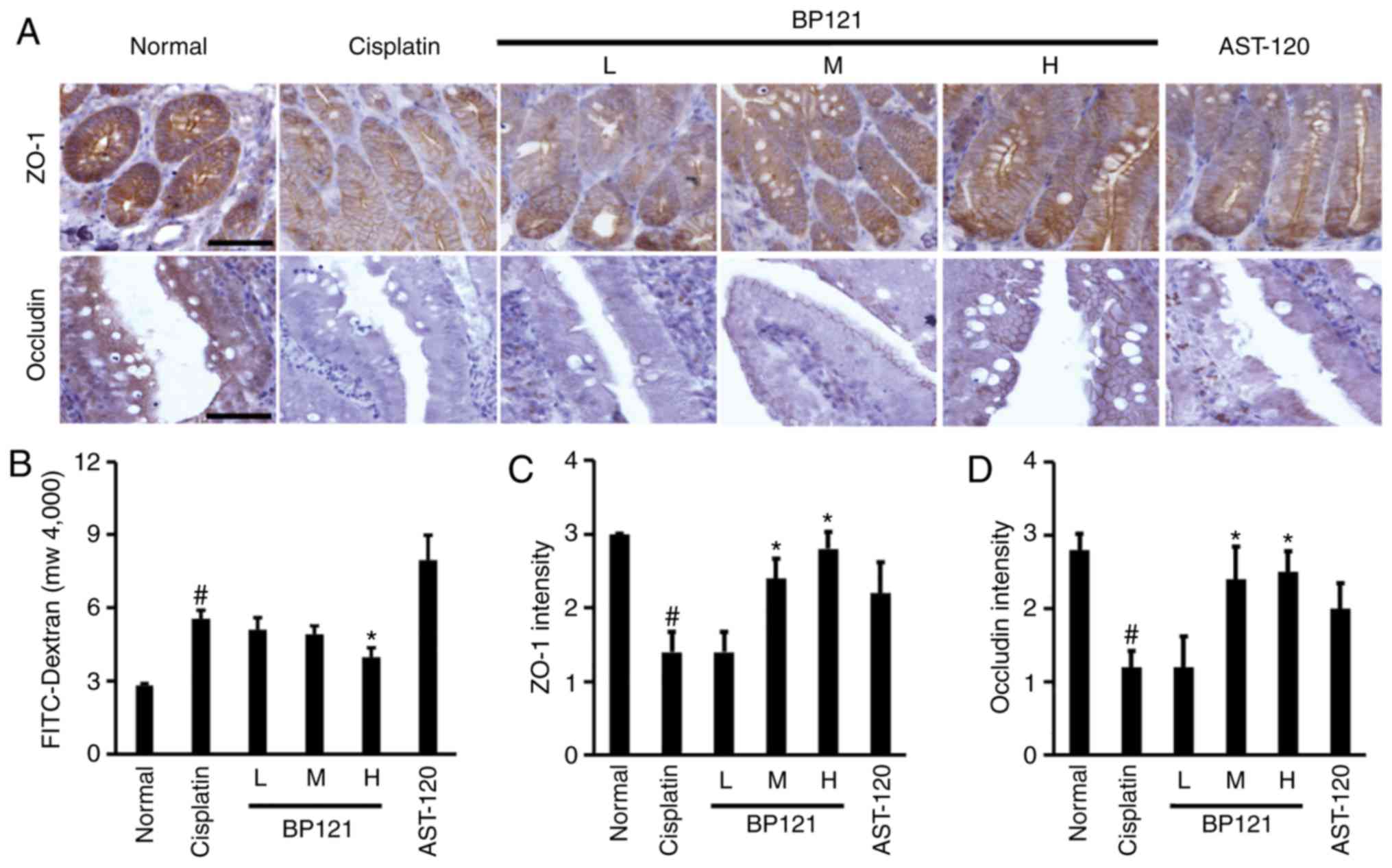
BP121 reverses cisplatin-induced increase
in intestinal permeability
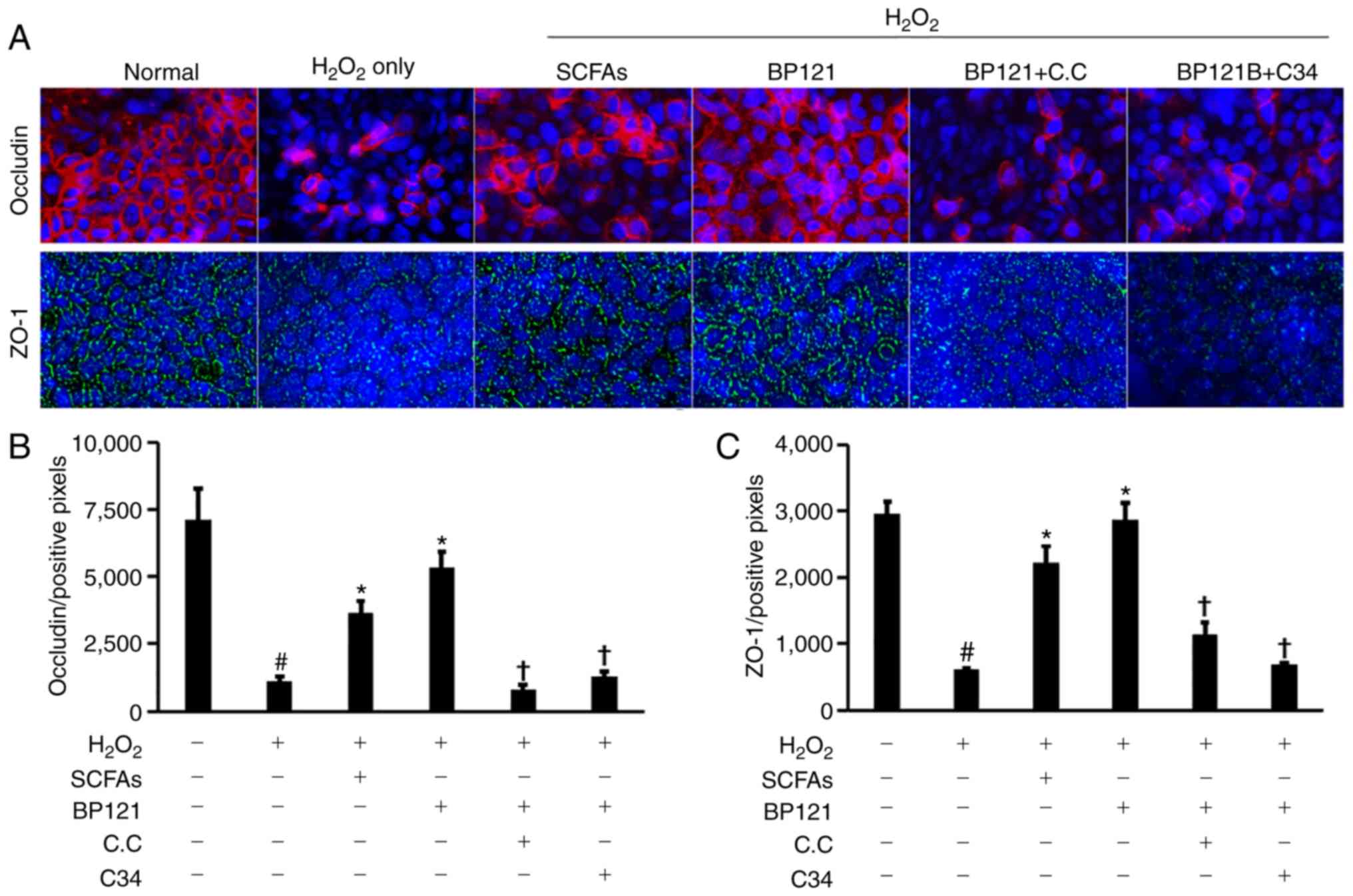
Intestinal permeability was increased by cisplatin;
this was accompanied by downregulation of ZO-1 and occludin
(Fig. 6A-D). However, medium and
high doses of BP121 restored the levels of these proteins and
reduced intestinal permeability. Treatment with AST 120 did not
affect TJ protein expression or intestinal permeability.
BP121 causes changes in intestinal flora
and stimulates SCFA production
The 16S rRNA analysis showed that the proportion of
Lactobacillales, a representative order of beneficial
bacteria, was decreased by cisplatin; however, the abundance was
restored upon BP121 administration. On the other hand, there were
no changes in the proportion of Clostridiales, an order of
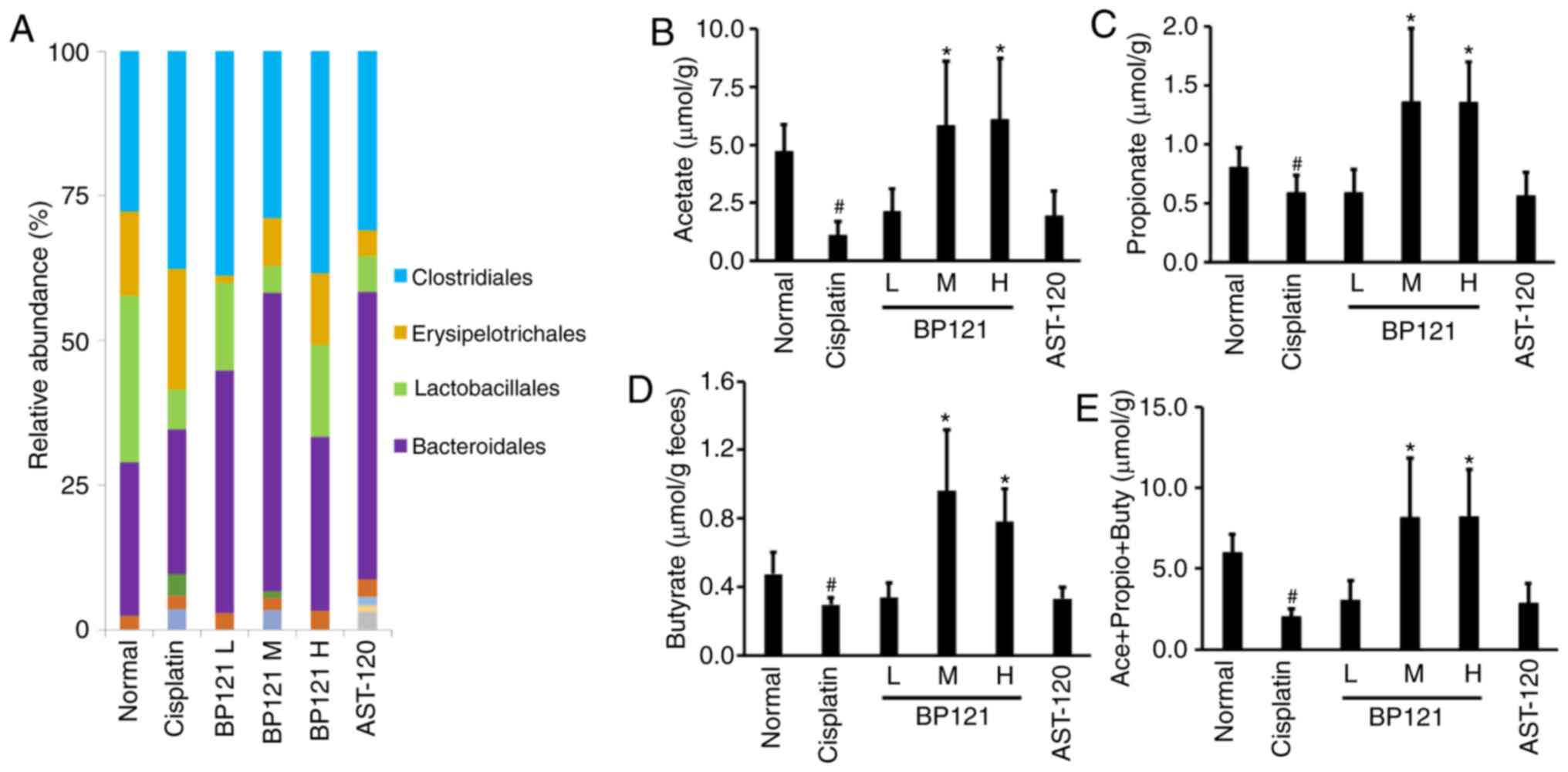
harmful bacteria, across treatment groups (Fig. 7A). The concentrations of the
representative SCFAs acetate, propionate and butyrate, which are
known to regulate the expression of TJ proteins-in feces were
examined and it was observed that they were decreased in
cisplatin-treated rats. However, medium and high concentrations of
BP121 increased the levels of all three SCFAs. AST-120
administration had no effect on SCFA levels (Fig. 7B-E).
BP121 increases TJ protein expression in
Caco-2 cells
In vitro experiments were carried out using
Caco-2 cells to clarify the mechanism by which BP121 improves
intestinal permeability. H2O2 treated Caco-2
cells showed a marked decrease of both occludin and ZO-1 expression
and pretreatment with SCFA or BP121 partially restored their
expression. However, cotreatment with inhibitors of AMPK or TLR4
blocked the beneficial effect of BP121, suggesting that BP121
mediated barrier modulation is dependent on AMPK and TLR4 signaling
(Fig. 8A-C).
Discussion
The human intestine contains over 1,000 species of
bacteria that interact dynamically with the host, which plays an
important role in health and disease (25). The microbiota is important for
maintaining the integrity of the intestinal barrier as well as for
immunological tolerance and metabolic activities that have not
evolved in the host (26,27). Intestinal dysbiosis is implicated
in various pathological processes including inflammatory bowel
disease, diabetes, obesity and AKI (27-29). Supplementing the gut microbiome
with beneficial bacteria such as LAB can promote host health.
The present study investigated the effects of L.
salivarius BP121, one of the 15 intestinal microorganisms
isolated from the feces of newborns-on AKI. Previous studies have
shown that the two-peptide bacteriocin ABP-118 produced by L.
salivarius inhibits the growth of pathogenic bacteria but not
most Lactobacillus species (30,31). The current study speculated that
L. salivarius could contribute to the treatment of diseases
caused by dysbiosis. Indeed, the present results showed that BP121
increased the abundance of Lactobacillales in rats with
cisplatin-induced dysbiosis.
The intercellular junctional complex is an important
component of the intestinal barrier. TJ proteins such as ZO-1 and
occludin seal the paracellular space between epithelial cells
(32), thus preventing the
leakage of solutes such as microorganisms and antigens as they are
transported across the epithelium (33). In this study, BP121 increased the
expression of TJ proteins that were downregulated by
H2O2 and cisplatin treatments. This is
consistent with a previous study in which Lactobacillus
rhamnosus GG increased the expression of human epithelial TJ
proteins in vivo, which conferred protection against
alcohol-induced epithelial barrier disruption (34). Moreover, L. plantarum
administration increased ZO-1 and occludin levels in an animal
model of alcoholic liver disease (34), and prevented the loss of TJ
proteins and abolished the increase in epithelial permeability
induced by phorbol ester in Caco-2 cells (35). Probiotics prevent damage to TJs by
stimulating lipoxygenase activity via SCFAs (36). The maintenance of TJ proteins also
involves activation of AMPK signaling (34,35). In the present study, a 48-h
treatment with BP121 culture supernatant containing SCFAs
alleviated TJ protein damage caused by H2O2
in Caco-2 cells; however, this effect was blocked by AMPK
inhibitor. These results are in agreement with previous reports
that SCFAs such as butyrate and acetate can preserve intestinal
barrier integrity by SCFAs (37-39).
SCFAs are straight-chain saturated FAs with fewer
than six carbon atoms; acetate, propionate and butyrate are the
most abundant SCFAs in the human intestinal tract (11). SCFAs are the end products of
fermentation of complex polysaccharides (40) including non-digestible dietary
fibers such as inulin (41) and
endogenous substrates such as epithelium-derived mucus (42). SCFAs are not only present in the
gut, but can also be absorbed in the bloodstream (13) and protect against kidney injury by
suppressing pro-inflammatory chemokines and cytokines such as IL-6,
TNF-α, and nitric oxide along with the production of reactive
oxygen species (ROS) and apoptosis (43,44), and/or by stimulating the
production of the anti-inflammatory cytokine IL-10 (45) via G protein-coupled receptor
activation (46) or histone
deacetylase inhibition (47).
The antioxidant properties of probiotics have been
confirmed and demonstrated by multiple studies (48,49). ROS were shown to mediate
cisplatin-induced AKI (5,8) and it was demonstrated that
H2O2 and OH radicals are involved in
cisplatin-induced necrosis and apoptosis, respectively (49). In this study, BP121 decreased
lipid peroxidation and increased the GSH/GSSG ratio; this was
accompanied by reductions in the number of TUNEL- and Bax-positive
cells and decreases in the levels of MCP-1 and the pro-inflammatory
cytokines IL-6 and TNF-α in kidney tissue.
IS and PCS are uremic toxins that are not
efficiently removed by hemodialysis (50). They are generated from bacterial
fermentation of tryptophan, aromatic amino acids and tyrosine
followed by sulfation in the liver or colonic mucosa (50). In the plasma, IS and PCS are
protein-bound and accumulate upon kidney failure (51). The concentrations of both toxins
are correlated with progression of chronic kidney disease and
higher risk of mortality (52).
The observed increases in serum IS and PCS levels could result from
enhanced production of these toxins, presumably due to an increase
in the abundance of specific microbiota and disruption of the
intestinal barrier by IS itself or by a decline in the
Lactobacillales population (53). In the present study, treatment of
AKI rats with BP121 stabilized intestinal barrier function and
reduced serum levels of IS and PCS, which suppressed in renal
inflammation.
In conclusion, the current results demonstrate that
the newly isolated L. salivarius strain BP121 protects
against cisplatin-induced AKI by suppressing IS and PCS production
through modulation of the gut environment and regulation of TJ
protein assembly via AMPK activation. These findings suggest that
probiotics including BP121 are an effective treatment for dysbiosis
and can prevent AKI.
Supplementary Data
Funding
The present study was supported by Green Cross
Wellbeing Corp (grant no. GCWB108).
Availability of data and materials
All data generated or analyzed in the present study
are included in this published article.
Authors' contributions
THL, DP and YJK contributed to the study conception
and design, as well as the data acquisition and analysis,
interpretation. THL, DP drafted the manuscript. IL performed the
immunohistochemical staining and in vitro studies. SK, CTO
and JYK contributed to data interpretation. JY and SKJ contributed
in study conception and reviewing the draft. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Animal Care and Use Committee of Hoseo University [approval no.
HSIACUC-18-097(1)].
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Zuk A and Bonventre JV: Acute kidney
injury. Annu Rev Med. 67:293–307. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bellomo R, Kellum JA and Ronco C: Acute
kidney injury. Lancet. 380:756–766. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hsu CY, McCulloch CE, Fan D, Ordonez JD,
Chertow GM and Go AS: Community-based incidence of acute renal
failure. Kidney Int. 72:208–212. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rahman M, Shad F and Smith MC: Acute
kidney injury: A guide to diagnosis and management. Am Fam
Physician. 86:631–639. 2012.PubMed/NCBI
|
|
5
|
Ozkok A and Edelstein CL: Pathophysiology
of cisplatin-induced acute kidney injury. Bio Med Res Int.
2014:9678262014.
|
|
6
|
Chen YY, Chen DQ, Chen L, Liu JR, Vaziri
ND, Guo Y and Zhao YY: Microbiome-metabolome reveals the
contribution of gut-kidney axis on kidney disease. J Transl Med.
17:52019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gong J, Noel S, Pluznick JL, Hamad ARA and
Rabb H: Gut microbiota-kidney cross-talk in acute kidney injury.
Semin Nephrol. 39:107–116. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pabla N and Dong Z: Cisplatin
nephrotoxicity: Mechanisms and renoprotective strategies. Kidney
Int. 73:994–1007. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang D and Lippard SJ: Cellular processing
of platinum anticancer drugs. Nat Rev Drug Discov. 4:307–320. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cohen SM and Lippard SJ: Cisplatin: From
DNA damage to cancer chemotherapy. Prog Nucleic Acid Res Mol Biol.
67:93–130. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Joyce K, Saxena S, Williams A, Damurjian
C, Auricchio N, Aluotto S, Tynan H and Demain AL: Antimicrobial
spectrum of the antitumor agent, cisplatin. J Antibiot (Tokyo).
63:530–532. 2010. View Article : Google Scholar
|
|
12
|
Taur Y and Pamer EG: Microbiome mediation
of infections in the cancer setting. Genome Med. 8:402016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Andrade-Oliveira V, Amano MT, Correa-Costa
M, Castoldi A, Felizardo RJ, de Almeida DC, Bassi EJ, Moraes-Vieira
PM, Hiyane MI, Rodas AC, et al: Gut bacteria products prevent AKI
induced by ischemia-reperfusion. J Am Soc Nephrol. 26:1877–1888.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Barrows IR, Ramezani A and Raj DS: Gut
feeling in AKI: The long arm of short-chain fatty acids. J Am Soc
Nephrol. 26:1755–1757. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
LeBlanc JG, Chain F, Martín R,
Bermúdez-Humarán LG, Courau S and Langella P: Beneficial effects on
host energy metabolism of short-chain fatty acids and vitamins
produced by commensal and probiotic bacteria. Microb Cell Fact.
16:792017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shimizu J, Kubota T, Takada E, Takai K,
Fujiwara N, Arimitsu N, Murayama MA, Ueda Y, Wakisaka S, Suzuki T
and Suzuki N: Propionate-producing bacteria in the intestine may
associate with skewed responses of IL10-producing regulatory T
cells in patients with relapsing polychondritis. PLoS One.
13:e02036572018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Parada Venegas D, De la Fuente MK,
Landskron G, González MJ, Quera R, Dijkstra G, Harmsen HJM, Faber
KN and Hermoso MA: Short chain fatty acids (SCFAs)-mediated gut
epithelial and immune regulation and its relevance for inflammatory
bowel diseases. Front Immunol. 10:2772019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen T, Kim CY, Kaur A, Lamothe L, Shaikh
M, Keshavarzian A and Hamaker BR: Dietary fibre-based SCFA mixtures
promote both protection and repair of intestinal epithelial barrier
function in a caco-2 cell model. Food Funct. 8:1166–1173. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Diao H, Jiao AR, Yu B, Mao XB and Chen DW:
Gastric infusion of short-chain fatty acids can improve intestinal
barrier function in weaned piglets. Genes Nutr. 14:42019.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Meimandipour A, Hair-Bejo M, Shuhaimi M,
Azhar K, Soleimani AF, Rasti B and Yazid AM: Gastrointestinal tract
morphological alteration by unpleasant physical treatment and
modulating role of lactobacillus in broilers. Br Poult Sci.
51:52–59. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fujii H, Nishijima F, Goto S, Sugano M,
Yamato H, Kitazawa R, Kitazawa S and Fukagawa M: Oral charcoal
adsorbent (AST-120) prevents progression of cardiac damage in
chronic kidney disease through suppression of oxidative stress.
Nephrol Dial Transplant. 24:2089–2095. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang CY, Lin PR, Ng CC and Shyu YT:
Probiotics properties of lactobacillus strains isolated from the
feces of breast-fed infants and Taiwanese pickled cabbage.
Anaerobe. 16:578–585. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fang CY, Lu JR, Chen BJ, Wu C, Chen YP and
Chen MJ: Selection of uremic toxin-reducing probiotics in vitro and
in vivo. J Funct Foods. 7:407–415. 2014. View Article : Google Scholar
|
|
24
|
McDonald JW and Pilgram TK: Nuclear
expression of p53, p21 and cyclin D1 is increased in
bronchioloalveolar carcinoma. Histopathology. 34:439–446. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pagliari D, Piccirillo CA, Larbi A and
Cianci R: The interactions between innate immunity and microbiota
in gastrointestinal diseases. J Immunol Res. 2015:8982972015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ley RE, Turnbaugh PJ, Klein S and Gordon
JI: Microbial ecology: Human gut microbes associated with obesity.
Nature. 444:1022–1023. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Turnbaugh PJ and Gordon JI: The core gut
microbiome, energy balance and obesity. J Physiol. 587:4153–4158.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Y, Hoenig JD, Malin KJ, Qamar S,
Petrof EO, Sun J, Antonopoulos DA, Chang EB and Claud EC: 16S rRNA
gene-based analysis of fecal microbiota from preterm infants with
and without necrotizing enterocolitis. ISME J. 3:944–954. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jeffery IB, O'Toole PW, Öhman L, Clasesson
MJ, Deane J, Quigley EM and Simre'n M: An irritable bowel syndrome
subtype defined by species-specific alterations in faecal
microbiota. Gut. 61:997–1006. 2012. View Article : Google Scholar
|
|
30
|
Barrett E, Hayes M, O'Connor P, Gardiner
G, Fitzgerald GF, Stanton C and Hill C: Salivaricin P: One of a
family of two-component antilisterial bacteriocins produced by
intestinal isolates of Lactobacillus salivarius. Appl Environ
Microbiol. 73:3719–3723. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Giongo A, Gano KA, Crabb DB, Mukherjee N,
Novelo LL, Casella G, Drew JC, Ilonen J, Knip M, Hyöty H, et al:
Toward defining the autoimmune microbiome for type 1 diabetes. ISME
J. 5:82–91. 2011. View Article : Google Scholar :
|
|
32
|
Ley RE, Peterson DA and Gordon JI:
Ecological and evolutionary forces shaping microbial diversity in
the human intestine. Cell. 124:837–848. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wells JM, Rossi O, Meijerink M and van
Baarlen P: Epithelial crosstalk at the microbiota-mucosal
interface. Proc Natl Acad Sci USA. 108(Suppl 1): 4607–4614. 2011.
View Article : Google Scholar
|
|
34
|
Bryniarski MA, Hamarneh F and Yacoub R:
The role of chronic kidney disease-associated dysbiosis in
cardiovascular disease. Exp Biol Med (Maywood). 244:514–525. 2019.
View Article : Google Scholar
|
|
35
|
Karczewski J, Troost FJ, Konings I, Dekker
J, Kleerebezem M, Brummer RJM and Wells JM: Regulation of human
epithelial tight junction proteins by Lactobacillus plantarum in
vivo and protective effects on the epithelial barrier. Am J Physiol
Gastrointest Liver Physiol. 298:G851–G859. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vaziri ND: CKD impairs barrier function
and alters microbial flora of the intestine: A major link to
inflammation and uremic toxicity. Curr Opin Nephrol Hypertens.
21:587–592. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mariadason JM, Barkla DH and Gibson PR:
Effect of short-chain fatty acids on paracellular permeability in
Caco-2 intestinal epithelium model. Am J Physiol. 272:G705–G712.
1997.PubMed/NCBI
|
|
38
|
Kendle M and Maslowski: The role of GPR43
in the immune system: A novel connection between diet, gut
microbiota and immune function. University of New South Wales;
Sydney: 2013
|
|
39
|
Nyman M: Fermentation and bulking capacity
of indigestible carbohydrates: The case of inulin and
oligofructose. Br J Nutr. 87(Suppl 2): 163–168. 2002. View Article : Google Scholar
|
|
40
|
Kotzampassi K, Giamarellos-Bourboulis EJ
and Stavrou G: Obesity as a consequence of gut bacteria and diet
interactions. ISRN Obes. 2014:6518952014.PubMed/NCBI
|
|
41
|
Wong JM, de Souza R, Kendall CW, Emam A
and Jenkins DJ: Colonic health: Fermentation and short chain fatty
acids. J Clin Gastroenterol. 40:235–243. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pomare EW, Branch WJ and Cummings JH:
Carbohydrate fermentation in the human colon and its relation to
acetate concentrations in venous blood. J Clin Invest.
75:1448–1454. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kim MH, Kang SG, Park JH, Yanagisawa M and
Kim CH: Short-chain fatty acids activate GPR41 and GPR43 on
intestinal epithelial cells to promote inflammatory responses in
mice. Gastroenterology. 145:396–406. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tedelind S, Westberg F, Kjerrulf M and
Vidal A: Anti-inflammatory properties of the short-chain fatty
acids acetate and propionate: A study with relevance to
inflammatory bowel disease. World J Gastroenterol. 13:2826–2832.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu T, Li J, Liu Y, Xiao N, Suo H, Xie K,
Yang C and Wu C: Short-chain fatty acids suppress
lipopolysaccharide-induced production of nitric oxide and
proinflammatory cytokines through inhibition of NF-κB pathway in
RAW264.7 cells. Inflammation. 35:1676–1684. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tazoe H, Otomo Y, Karaki S, Kato I, Fukami
Y, Terasaki M and Kuwahara A: Expression of short-chain fatty acid
receptor GPR41 in the human colon. Biomed Res. 30:149–156. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wang Y, Wu Y, Wang Y, Xu H, Mei X, Yu D
and Li W: Antioxidant properties of probiotic bacteria. Nutrients.
9:521–535. 2017. View Article : Google Scholar :
|
|
48
|
Mishra V, Shah C, Mokashe N, Chavan R,
Yadav H and Prajapati J: Probiotics as potential antioxidants: A
systematic review. J Agric Food Chem. 63:3615–3626. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Baek SM, Kwon CH, Kim JH, Woo JS, Jung JS
and Kim YK: Differential roles of hydrogen peroxide and hydroxyl
radical in cisplatin-induced cell death in renal proximal tubular
epithelial cells. J Lab Clin Med. 142:178–186. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Meijers BK and Evenepoel P: The gut-kidney
axis: Indoxyl sulfate, p-cresyl sulfate and CKD progression.
Nephrol Dial Transplant. 26:759–761. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jourde-Chiche N, Dou L, Cerini C,
Dignat-George F, Vanholder R and Brunet P: Protein-bound
toxins-update 2009. Semin Dial. 22:334–339. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ramezani A and Raj DS: The gut microbiome,
kidney disease, and targeted interventions. J Am Soc Nephrol.
25:657–670. 2014. View Article : Google Scholar :
|
|
53
|
Yoshifuji A, Wakino S, Irie J, Tajima T,
Hasegawa K, Kanda T, Tokuyama H, Hayashi K and Itoh H: Gut
Lactobacillus protects against the progression of renal damage by
modulating the gut environment in rats. Nephrol Dial Transplant.
31:401–412. 2015. View Article : Google Scholar : PubMed/NCBI
|