Introduction
Osteosarcoma (OS) represents ~60% of malignant bone
tumors (1) and mostly occurs in
young adults. The high distant metastatasis rate of OS leads to
limited survival in OS patients, particularly terminal patients or
patients with multidrug resistance (2,3).
Therefore, the need to explore the potential molecular mechanism of
OS to not only develop complementary or alternative treatments but
also enhance the therapeutic effects of current treatments is
urgent.
Over the past few years, the medical community has
paid great attention to the potential of gene treatment and
targeted therapy in elucidating the mechanisms of disease and
altering diseases at the molecular level (4). The identification of specific genes
can be essential for success. TP53, which is commonly
regarded as one of the most critical tumor suppressor genes, serves
as a central regulatory factor in cell proliferation, cell
apoptosis and other biological processes (5-8).
The p53 protein prevents the proliferation of injured or mutated
cells by inducing cell cycle arrest, apoptosis or senescence;
therefore, the p53 dysfunction might promote the generation and
development of tumors (9-11). Interestingly, it was reported that
TP53 could act as a potential prognostic biomarker for
patients with OS (12,13) and was altered in ~50% of OS cases
(12). Zhao et al
(14) revealed that the
overexpression of p53 enhanced the chemical sensitivity of
multidrug-resistant OS cell lines, while Wu et al (15) demonstrated that the expression of
p53 has emerged as a valid prognostic biomarker for predicting the
survival of OS patients. Fully understanding its functional network
will be of great benefit to clinical treatment.
Located at the center of a complex molecular
regulatory network, TP53 can induce cell cycle arrest and
apoptosis via regulating the transcription of microRNAs
(miRNAs/miRs) and other different genes. miRNAs play a crucial role
as post-transcriptional regulatory factors that bind target mRNAs
at their 3′ untranslated region (UTR) to repress gene expression
(16-18). Previously, Jones et al
(19) regarded the miR-181 group
as an essential OS oncomiR; three of the four miRNAs from the
miR-181 group, including miR-181a, miR-181b and miR-181c, were
highly upregulated within OS samples (19). Furthermore, miR-181 can activate
the Wnt signaling pathway (20),
which is essential for the pathogenesis of OS (21). miR-181a (22,23) and miR-181b (24,25) both promote the capacity of cells
proliferation and invasion but inhibit OS cell apoptosis. More
notably, based on online predictive tools, p53 might bind miR-181b
at its promoter region, while miR-181b might target TP53 at
its 3′-UTR to negatively regulate one another. Therefore, the
present study hypothesized that the abnormal upregulation of
miR-181b in OS disturbs the negative feedback balance between p53
and miR-181b, causing excessive OS cell proliferation and
invasion.
In the present study, miR-181b and p53 expression
was determined in normal noncancerous and OS tissue samples; then
the correlation between miR-181b and p53 expression in tissues was
examined. Next, the predicted interactions between p53 and miR-181b
promoter and between miR-181b and the TP53 3′-UTR and the
negative regulation of miR-181b and p53 were verified. Moreover,
the full effects of miR-181b on p53 signaling and the
proliferation, and invasion of OS cells and the dynamic effects of
miR-181b and p53 on OS cells were evaluated in vitro and
in vivo. In summary, the present study provides a mechanism
by which the miR-181b-p53 negative feedback axis affects the
proliferation and invasion of OS cells.
Materials and methods
Clinical tissue samples
OS tissues and adjacent normal noncancerous tissues
(n=12, each) were collected during routine therapeutic surgery at
Xiangya Hospital from Feburay, 2018 to Feburay, 2019. (F/M is 3/9,
the average age is 24.33±7.10 years). OS patients who participated
in this study had been not received chemotherapy or radiotherapy.
Inclusion criteria and exclusion criteria were adopted from
previous studies (26,27). The adjacent normal noncancerous
tissues were 5 cm distance from OS tissues. All tissue specimens
were frozen in liquid nitrogen immediately and stored at −80°C
until use. The present study was reviewed and approved by the
Ethics Committee of Xiangya Hospital and each patient signed an
informed consent document.
Cell lines and cell transfection
Primary human osteoblasts (HOBs) were obtained from
Merck KGaA (cat. no. 406-05F) and cultured in a 1:1 mixture of
Ham's F12 medium and Dulbecco's modified Eagle's medium (both from
Gibco; Thermo Fisher Scientific, Inc.), with 2.5 mM L-glutamine
(without phenol red) and 10% fetal bovine serum (FBS) (Gibco;
Thermo Fisher Scientific, Inc.). A total of two OA cell lines, U2OS
(American Type Cututre Collection; ATCC HTB-96™) and MG63 (ATCC
CRL-1427™) cells were obtained from ATCC. U2OS cells were cultured
in McCoy's 5a medium (cat. no. 30-2007; ATCC) supplemented with 10%
FBS. MG63 cells were cultured in Eagle's Minimum essential medium
(cat. no. 30-2003; ATCC) supplemented with 10% FBS. Cells were
cultured at 37°C in 5% CO2.
For cell transfection, 20 nM negative control (NC)
mimics and miR-181b-5p mimics, 20 nM NC inhibitor and miR-181b-5p
inhibitor, 1 µg/ml control vector and p53-overexpression
vector (p53 OE), or 20 nM small intereferring (si)-NC and si-p53
that had been designed and purchased from Shanghai GenePharma Co.,
Ltd., were used. Cells in log phase were seeded into 12-well plates
at a density of 5×105 cells/ml one day before
transfection. When the cell confluence reached 70%, siRNAs, vectors
and miRNA mimics or inhibitors were diluted in Opti-MEM
(Invitrogen; Thermo Fisher Scientific, Inc.) and incubated with 5
µl Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific,
Inc.) for 5 min at room temperature. Next, the DNA-lipid complex
were added to cells. Then, cells were incubated at 37°C in a
CO2 incubator. Medium was then replaced with fresh
complete medium containing 10% FBS after 6 h of transfection. A
total of 48 h after transfection, cells were harvested for further
experiments. The sequence of miRNA mimics or inhibitor and primers
for plamid construction were listed in Table SI.
PCR-based analyses
Total RNA from tissues or target cells was extracted
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.),
following the manufacturer's procedures. A NanoDrop-1000 (Thermo
Fisher Scientific, Inc.) was used to determine concentrations and
for quality control purposes. Complementary DNA was synthesized
from extracted RNA 9 (5 µg) using the Prime
Script® RT Reagent kit with gDNA Eraser (Invitrogen;
Thermo Fisher Scientific, Inc.) in accordance with the recommended
protocol (samples and reagent mix were incuabated at 37°C for 15
min and 85°C for 5 sec, and finally stored at 4°C). Quantitative
(q)PCR was performed using SYBR®Premix Ex Taq™ II
(Invitrogen; Thermo Fisher Scientific, Inc.). ABI7500 real-time PCR
detection system (Applied Biosystems; Thermo Fisher Scientific,
Inc.) was used for detection. The qPCR thermocycling conditions
were as follows: The initial denaturation was first performed at
95°C for 2 min followed by denaturation at 95°C for 15 sec and
annealing and extension at 60°C for 30 sec. Repeat the
denaturation, annealing and extension for 40 circles. After
amplification, data were collected and processed by the comparative
cycle threshold method. U6 and GAPDH expression levels were used as
internal references for miRNA and mRNA expression detection,
respectively. Finally, the data were processed using the
2−ΔΔCq relative expression method (28). The primers were listed in Table SI.
Chromatin immunoprecipitation (ChIP)
analysis
Target cells were suspended in 1% formaldehyde in
PBS, cross-linked for 10 min at room temperature, washed twice with
PBS and lysed with SDS lysis buffer (Promega Corporation). At 4°C,
the lysates were centrifuged at 1,0000 × g for 10 min; the
supernatants were diluted with ChIP dilution buffer (EMD Millipore)
and then immunoprecipitated using an anti-p53 antibody (cat. no.
ab1101; 1:20) or mouse IgG (cat. no. ab190475; 1:20) (both from
Abcam). IgG served as a negative control. The immunoprecipitated
DNA was then analyzed by qPCR.
Luciferase reporter assay
To test the relationship between TP53 and miR-181b,
the online tool LncTar (http://www.cuilab.cn/lnctar) was used to predicate the
biding site between TP53 3′UTR and miR-181b-5p. Then,
dual-luciferase reporter assay was carried out. First, TP53 3′-UTR
was cloned into the downstream of the psiCheck2 vector (Promega
Corporation) to generate the wild-type TP53 3′-UTR luciferase
reporter vector; a mutant TP53 3′-UTR luciferase reporter vector
was generated by mutating the predicted miR-181b-binding site
within the TP53 3′-UTR. These two reporter vectors were
cotransfected with miR-181b mimics or miR-181b inhibitor into 293T
cells (ATCC). A total of 48 h later, the cells were harvested and
underwent a dual-luciferase reporter assay (Promega Corporation) to
evaluate firefly and Renilla the luciferase activities.
Renilla luciferase activity served as a normalization
control.
Immunoblotting
The cell lysates of OS cell lines were prepared
using RIPA lysis buffer (Beyotime Institute of Biotechnology), and
a bicinchoninic acid protein assay kit (Pierce; Thermo Fisher
Scientific, Inc.) was used to detect protein concentration. The
30-50 µg protein samples were then separated by 10-15%
SDS-PAGE and transferred onto PVDF membranes (Thermo Fisher
Scientific, Inc.). The membranes were blocked with 5% non-fat milk
in TBST (0.1% Tween-20) for 2 h at room temperature and incubated
with primary antibodies against p53 (1:1,000; cat. no. ab26;
Abcam), p21 (1:1,000; cat. no. ab109520; Abcam), Cyclin D1
(1:1,000; cat. no. ab16663; Abcam), epithelial (E)-cadherin
(1:1,000; cat. no. ab1416; Abcam) and GAPDH (1:2,000; cat. no.
ab8245; Abcam) overnight at 4°C. After washing three times with
TBST, the membranes were incubated for 1 h with goat-anti-rabbit or
goat-anti-mouse HRP antibody (both 1:5,000; cat. nos. ab205718 and
ab205719; both from Abcam) at room temperature. Intensity analyses
were performed using ImageJ software version 1.8.0 (National
Institute of Health).
Cell viability determined by MTT
assay
Cells were seeded in 96-well plates at a density of
5×103/well and transfected or treated. A total of 48 h
later, MTT solution (20 µl; 5 mg/ml) was added to the plates
at the 0, 12, 24 and 48 h and incubated at 37°C for 4 h. At the end
of the incubation, dimethyl sulfoxide was added to lyse the cells
and the supernatant was subsequently removed. Then the absorbance
value at 490 nm was measured.
DNA synthesis determined by
5-ethynyl-2′-deoxyuridine (EdU) assay
The ability to synthesize DNA was detected by EdU
assays using an EdU assay kit (Guangzhou RiboBio Co., Ltd.)
following a previously described method (29). Nuclei were stained with DAPI for
15 min at room temperature. Representative images were taken under
an IX71 fluorescence microscope (Olympus Corporation). A total of
five random fields were analyzed per sample.
Cell invasion determined by Transwell
assay
Treated OS cells (1×105 cells) were
suspended in serum-free medium in 24-well Transwell chambers
precoated with Matrigel; medium containing 10% FBS was added to the
bottom chamber. After cultured for 48 h at 37°C in a CO2
incubator, noninvasive cells in the upper chambers were discarded
whereas invasive cells were fixed with 5% paraformaldehyde for 20
min at room temperature and then stained with 0.1% crystal violet
for 20 min at room temperature. Invasive cells were in 4 random
fields in each group were counted under a light microscope
(magnification ×200) (Olympus).
Establishment of xenograft nude mice
Antagomir-miR-181b, antagomir-NC, p53 short hairpin
(sh)RNA plasmid (sh-p53) and NC shRNA plasmid (sh-NC) were obtained
from Shanghai GenePharma Co., Ltd. Antagomir-miR-181b, antagomir-NC
sh-p53 or sh-NC was transfected into MG63 cells with Lipofectamine
2000. A total of 48 h later, the transfected cells were harvested
for subcutaneous injection. To establish a human OS cell xenograft
nude mouse model, a total of 24 female nude mice (BALB/c-nu/nu) at
6-week-old and weighing 16-20 g were obtained from the SLAC
Laboratory Animal Center. The mice were housed in specific
pathogen-free room (22°C, ~50% humidity) and fed a standard
irradiated diet and had unrestricted access to purified water. Mice
were maintained on a 12-h light cycle. A total of 24 mice were
randomly divided into the following 4 groups: An antagomir-NC+sh-NC
group, antagomir-miR-181b+sh-NC group, antagomir-NC+sh-p53 group
and antagomir-miR-181b+sh-p53 group. A total of 1×106
transfected MG63 cells were suspended in 100 µl PBS and
subcutaneously injected into the right anterior armpits of the nude
mice. A total of 28 days later, mice were anesthetized via the
respiratory route by exposing them to ether for 2 min in a
transparent acrylic jar. The depth of anesthetization was
sufficient when the following vital criteria are reached: Regular
spontaneous breathing. No reflex after the setting of pain stimuli
between toes and no response to pain. Then, mice were sacrificed by
cervical dislocation. The tumor weight and volume were meausured.
The animal epxeriments were approved by the Ethic Committee of
Xiangya Hospital of Central South University.
Statistical analysis
All data from at least three independent experiments
were processed using GraphPad software (version 7; GraphPad Prism),
and are presented as the mean ± standard deviation. Differences
between two groups were determined using both unpaired or paired
Student's t-test where appropriate, and following one-way analysis
of variance, Tukey's test was used for comparison among three
groups or more. For the correlation analysis, the Pearson
correlation coefficient analysis was used. P<0.05 was considered
to indicate a statistically significant difference.
Results
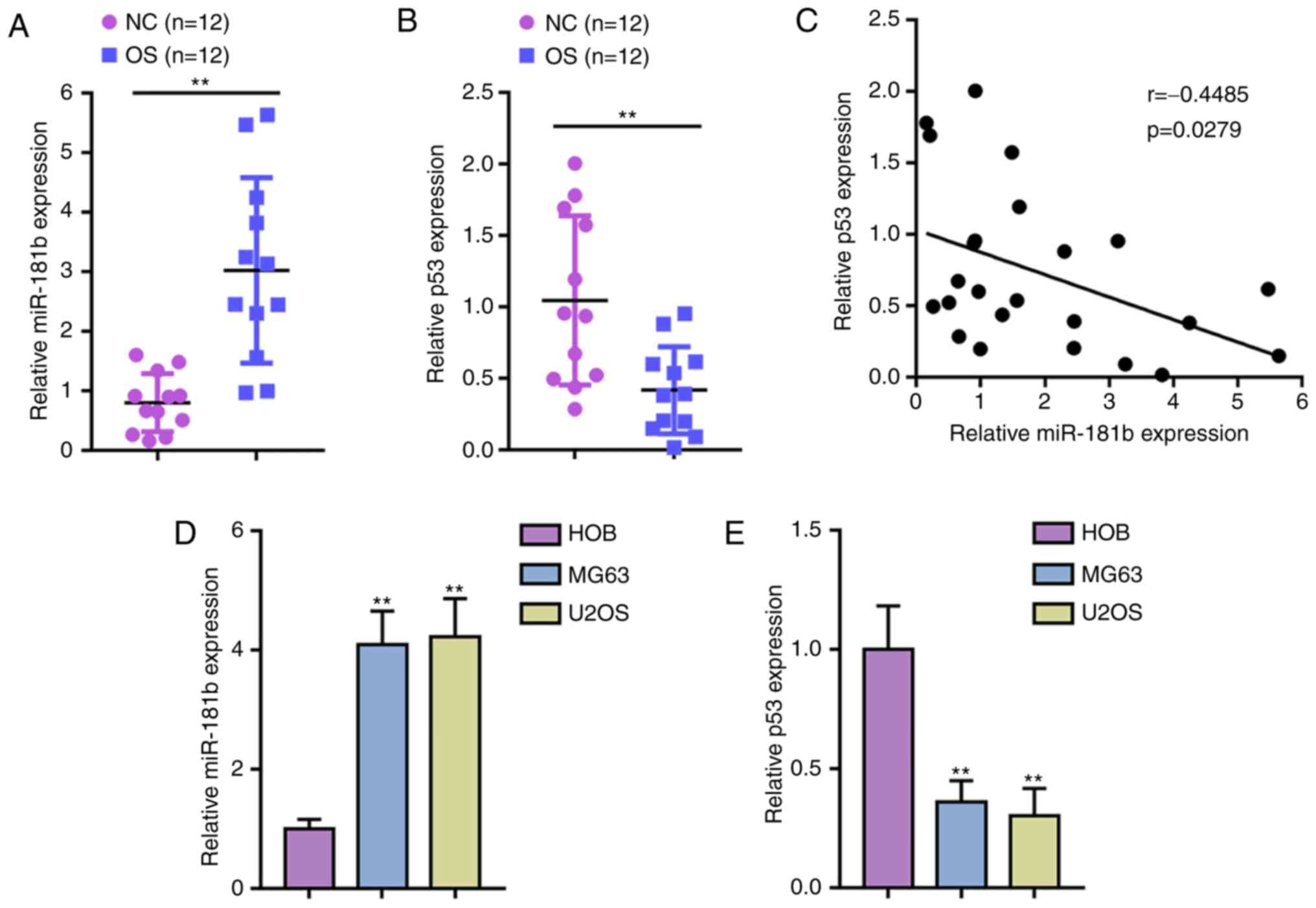
miR-181b and p53 expression within OS
tissue samples and cell lines
First, miR-181b and p53 expression was verified
within tissues and cell lines. Fig.
1A and B show that the expression of miR-181b was significantly
upregulated, while p53 expression was downregulated within OS
tissue samples compared with noncancerous tissue samples. Pearson
correlation coefficient analysis showed that miR-181b expression
was negatively correlated with p53 expression in OS tissue
(Fig. 1C). Consistently, miR-181b
expression was upregulated and p53 expression was downregulated in
two OS cell lines, MG63 and U2OS, compared with normal HOBs
(Fig. 1D and E).
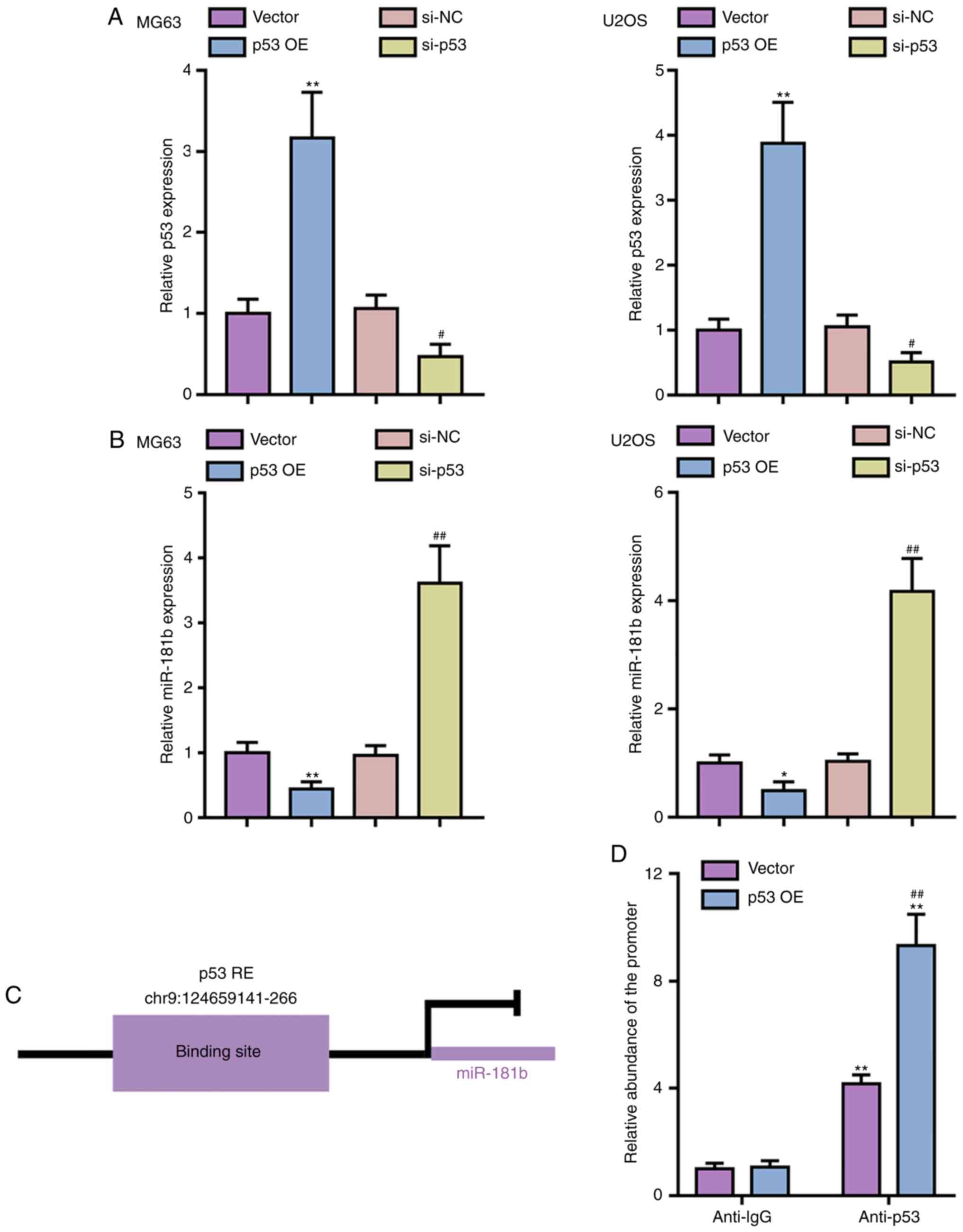
p53 inhibits the transcription of
miR-181b in OS cells
To verify the predicted binding between p53 and the
miR-181b promoter region, the MG63 and U2OS cell lines were
transfected with p53 OE or si-p53 vector for p53 OE or p53
knockdown, respectively, and qPCR was performed to verify the
transfection efficiency (Fig.
2A). Next, miR-181b expression was determined in
p53-overexpressing or p53-silenced OS cells; p53 OE significantly
inhibited miR-181b expression, whereas p53 knockdown significantly
promoted miR-181b expression in both OS cell lines (Fig. 2B). To provide further evidence of
the predicted binding of p53 to the miR-181b promoter (Fig. 2C), ChIP assays were performed with
anti-p53 and anti-IgG antibodies. Fig. 2D shows that the relative abundance
of the miR-181b promoter precipitated by anti-p53 was significantly
increased compared with precipitated with anti-IgG, especially in
p53-overexpressing 293T cells, indicating that p53 can inhibit the
expression of miR-181b via targeting its promoter region.
miR-181b suppresses the expression of p53
via targeting its 3′-UTR
The online tool LncTar predicted that miR-181b might
target p53 3′-UTR. To verify this prediction, miR-181b mimic or
inhibitor were transfected to induce miR-181b OE or inhibition,
respectively, within the MG63 and U2OS cell lines, and performed
qPCR to confirm the transfection efficiency (Fig. 3A). Consistent to the prediction,
p53 expression was inhibited via miR-181b OE while expression was
promoted via miR-181b inhibitor transfection (Fig. 3B). To verify this interaction, two
luciferase reporter vectors were constructed, wild-type and mutant
TP53 3′-UTR luciferase reporter vectors, that contained the
wild-type or mutated miR-181b binding site, respectively (Fig. 3C). These reporter vectors were
transfected into 293T cells with the miR-181b mimic/inhibitor and
the luciferase activity was examined. Fig. 3D shows that the luciferase
activity of cells transfected with the wild-type TP53 3′-UTR vector
was reduced via the OE of miR-181b but increased via the inhibition
of miR-181b; mutation of the putative miR-181b binding site within
TP53 3′-UTR could abolish the alterations in the luciferase
activity. In summary, miR-181b can suppress the expression of TP53
via direct binding to its 3′-UTR.
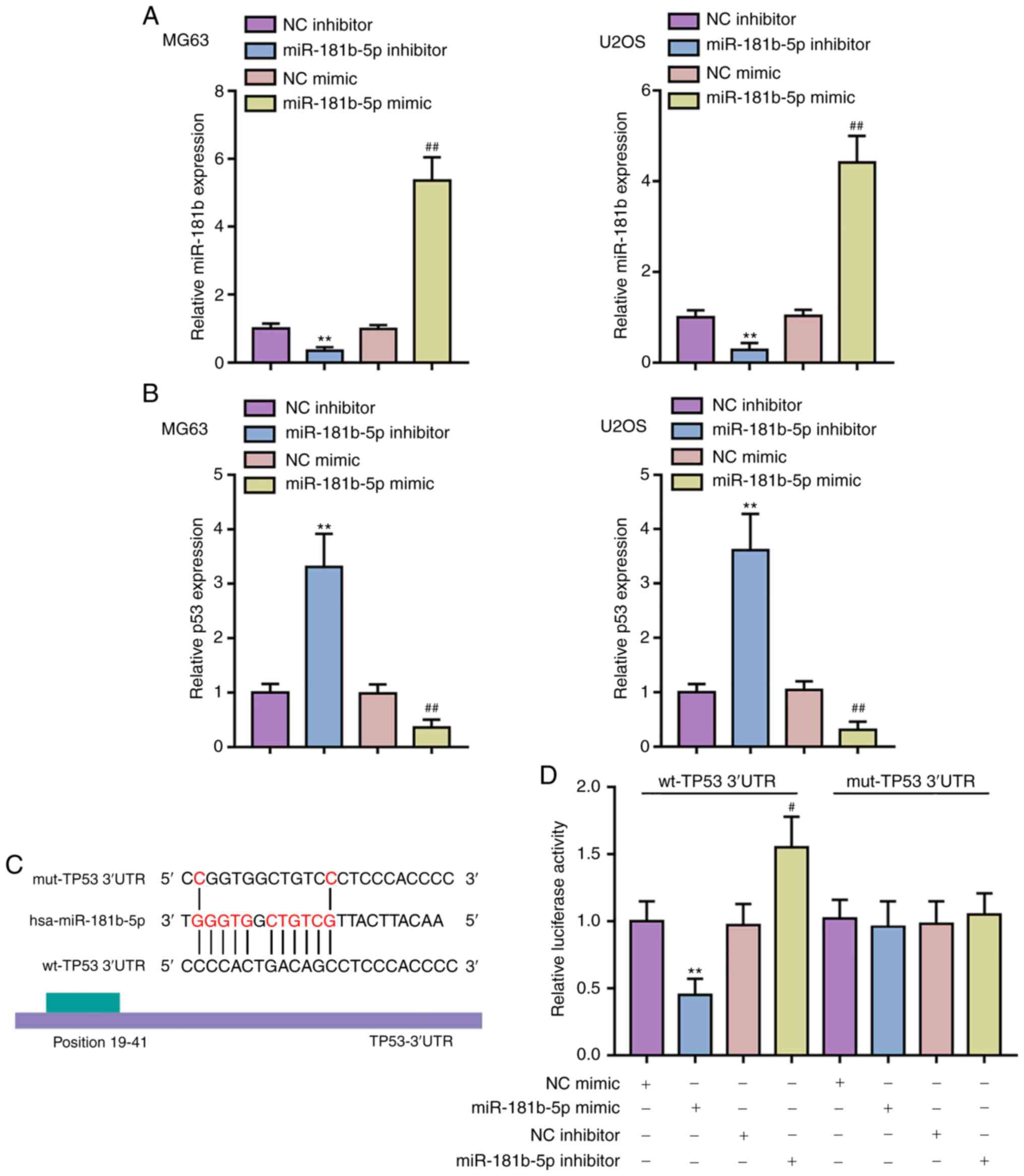 | Figure 3miR-181b binds the 3′-UTR of p53 to
inhibit its expression. (A) miR-181b OE and inhibition was carried
out in MG63 and U2OS cells by the transfection of miR-181b mimic or
miR-181b inhibitor, respectively, as confirmed by qPCR. (B) p53
expression in response to miR-181b OE or miR-181b inhibition was
determined in MG63 and U2OS cells by qPCR. **P<0.01,
vs. the NC inhibitor group; #P<0.05 and
##P<0.01 vs. NC mimic group. (C) A schematic diagram
showing the predicted binding site of miR-181b in TP53 3′-UTR. Wt
and mut TP53 3′-UTR luciferase reporter vectors were constructed as
described in the Materials and methods section. (D) These reporter
vectors were co-transfected in 293T cells with miR-181b mimic or
miR-181b inhibitor and luciferase activities were determined. UTR,
untranslated; miR, microRNA; NC, negative control; q, quantitative;
wt, wild-type; mut, mutant; OE, overexpression. |
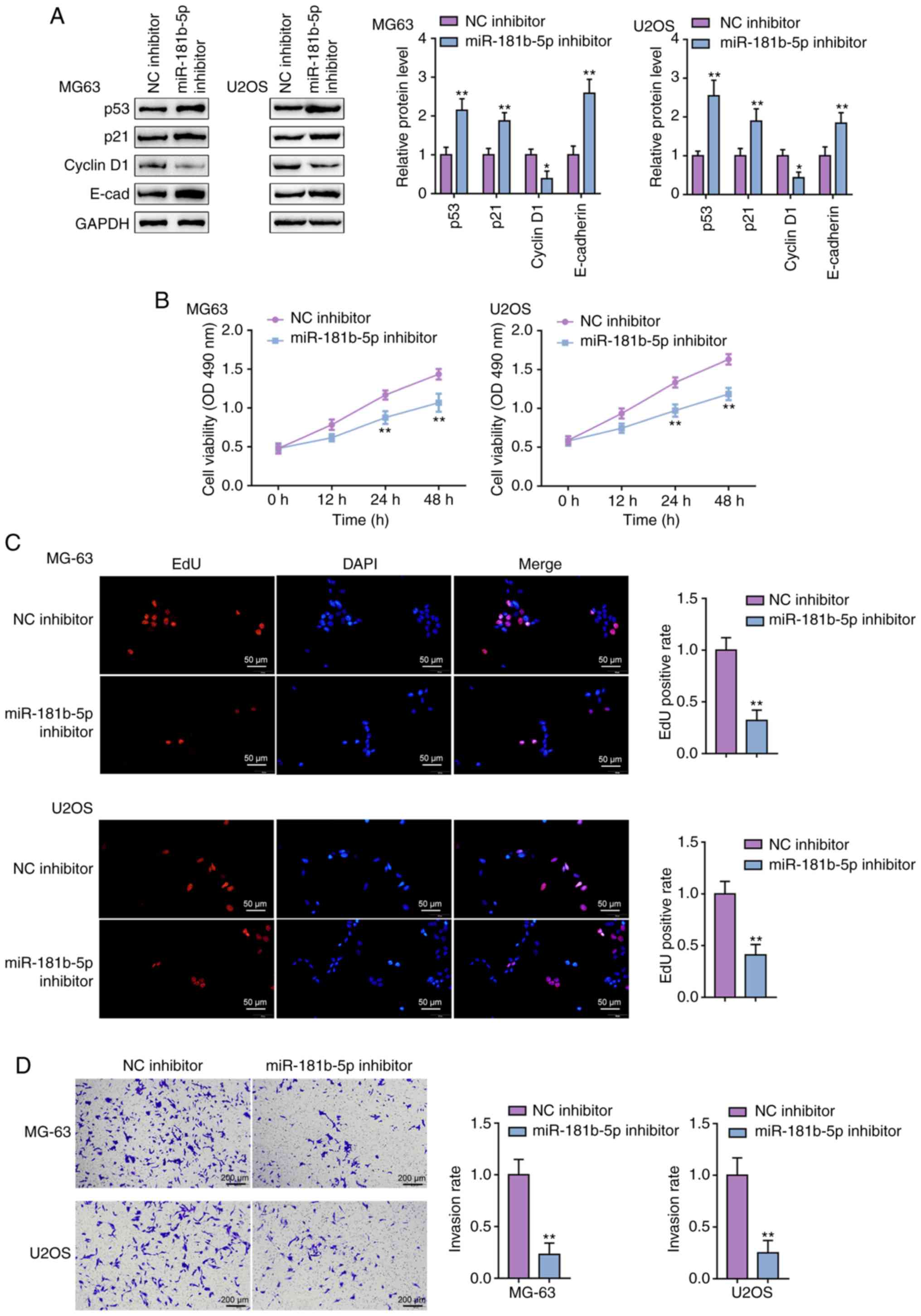
Detailed effects of miR-181b on p53
signaling and OS cell proliferation and invasion
After confirming the binding between p53 and the
miR-181b promoter and miR-181b and the TP53 3′-UTR, the
specific effects of miR-181b on p53 signaling and OS cells were
determined. The MG63 and U2OS cell lines were transfected with
miR-181b inhibitor and then examined p53, p21, Cyclin D1, and
E-cadherin protein levels. In both cell lines, miR-181b inhibition
dramatically increased p53, p21 and E-cadherin protein levels, but
decreased the protein levels of Cyclin D1 (Fig. 4A). The MTT assay showed that the
miR-181b inhibitor decreased the proliferative potential of the
MG63 and U2OS cell lines (Fig.
4B). Similarly, miR-181b inhibition also decreased the ability
of OS cells to synthesize DNA (Fig.
4C). Finally, the Transwell assay indicated that metastatic
ability of OS cells was significantly restricted by the miR-181b
inhibitor (Fig. 4D). In summary,
miR-181b inhibition enhanced p53 signaling activation.
Consistently, miR-181b inhibition reduced cell viability, DNA
synthesis and the invasive ability of both OS cell lines.
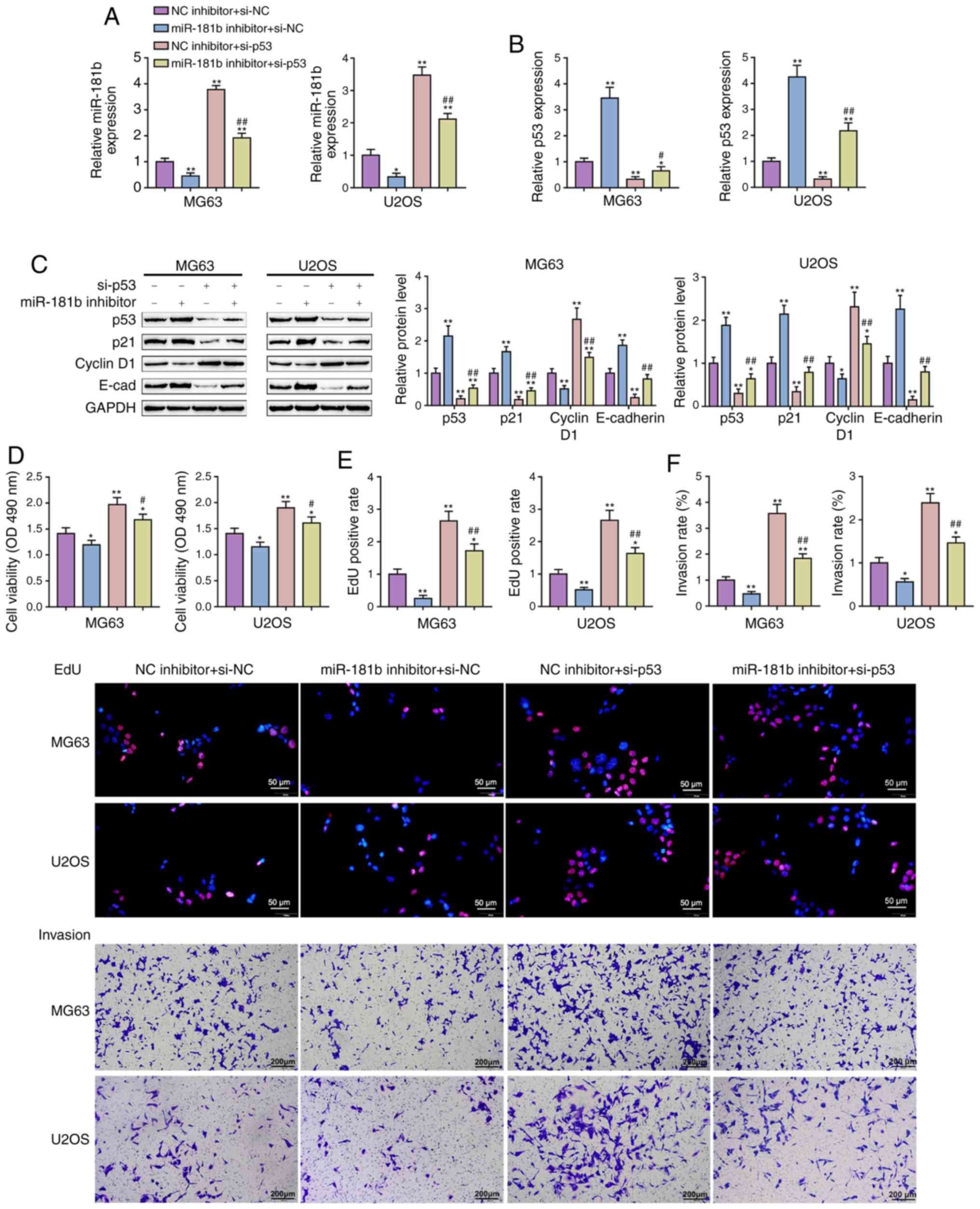
Dynamic effects of miR-181b and p53 on
the proliferation and invasion of OS cells
After confirming the full impact of miR-181b on p53
signaling and OS cell lines, whether miR-181b brings about these
effects via targeting p53 was determined. The MG63 and U2OS cell
lines were cotransfected with miR-181b inhibitor and si-p53 and
then miR-181b and p53 expression was evaluated. miR-181b expression
was dramatically reduced via miR-181b inhibitor transfection but
increased via si-p53 transfection; miR-181b inhibitor transfection
significantly attenuated the effect of si-p53 (Fig. 5A). Conversely, p53 expression was
considerably increased via miR-181b inhibition but reduced via
si-p53 transfection; the effects of the miR-181b inhibitor were
significantly attenuated via si-p53 transfection (Fig. 5B). These data further confirm that
miR-181b and p53 are negatively regulated by each other.
Regarding the p53 signaling, the protein levels of
p53, p21 and E-cadherin were significantly increased by miR-181b
inhibitor transfection and decreased by si-p53 transfection; on the
contrary, Cyclin D1 was reduced by miR-181b inhibition and raised
by p53 knockdown (Fig. 5C). p53
knockdown significantly attenuated the effects of miR-181b
inhibition (Fig. 5C).
Consistently, miR-181b inhibition significantly suppressed, whereas
p53 knockdown promoted cell viability, DNA synthesis and the
invasive abilities of both OS cell lines (Fig. 5D-F). These data indicate that
miR-181b functions in OS cells via targeting p53 and thus affecting
p53 signaling.
Dynamic effects of miR-181b and p53 on OS
cell xenograft nude mice
To further validate the effects of miR-181b and p53
in vivo, an OS cell xenograft model was established in nude
mice using antagomir-181b transfected and/or p53 knockdown MG63
cells. As shown in Fig. 6A and B,
antagomir-miR-181b transfection significantly decreased the tumor
weight and volume, and these changes were reversed by p53
knockdown. The protein levels of p53, p21 and E-cadherin in tumors
were significantly increased by antagomir-miR-181b tranfection and
decreased by sh-p53 transfection; on the contrary, the Cyclin D1
protein level was reduced by antagomir miR-181b and increased by
sh-p53 transfection (Fig. 6C).
sh-p53 transfection significantly attenuated the effects of
antagomir-miR-181b transfection (Fig.
6C). These results suggested that the miR-181b/p53 axis was
involved in OS tumor growth in nude mouse model.
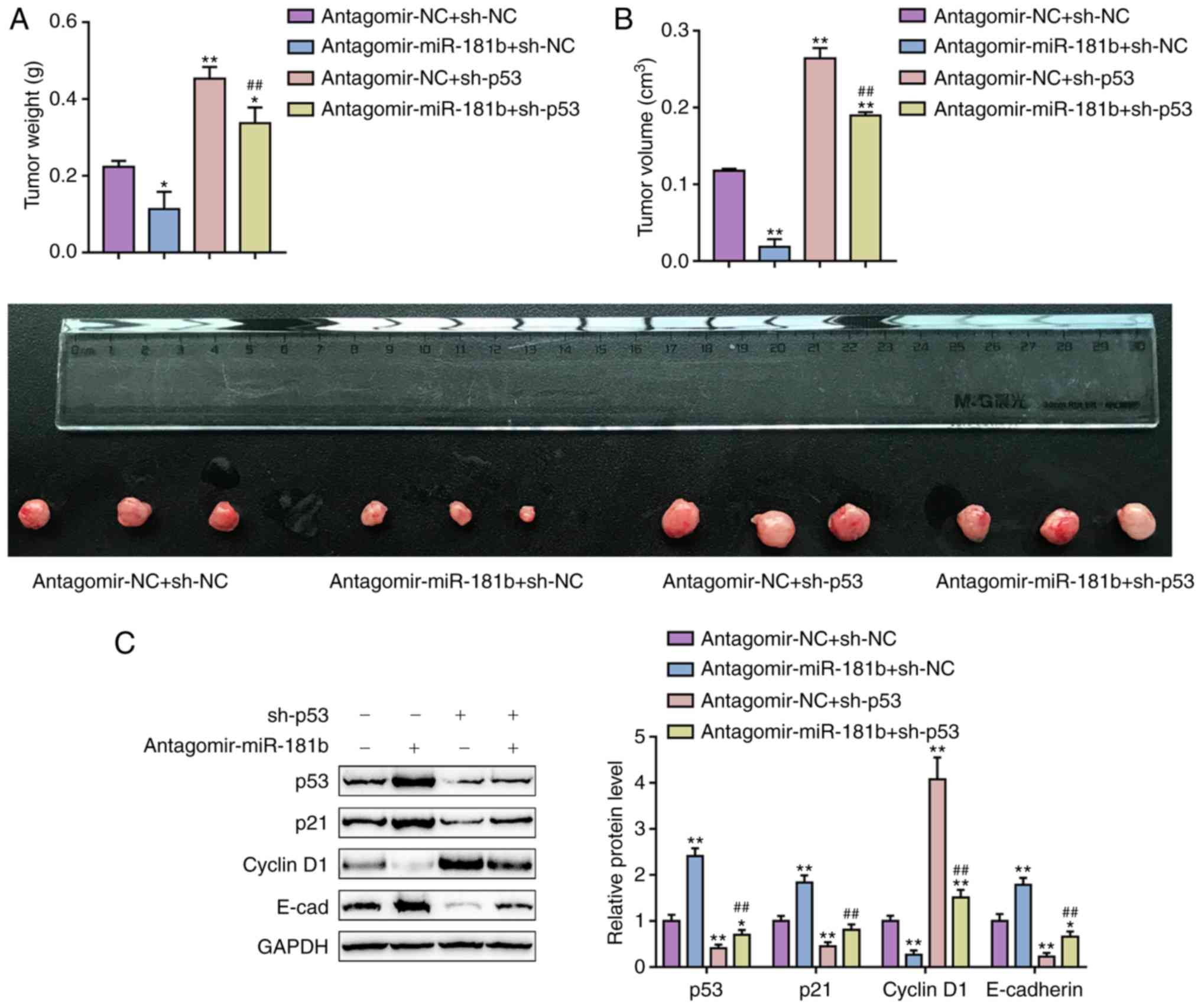 | Figure 6Dynamic effects of miR-181b and p53
on OS cell xenograft nude mice. OS cell xenograft (A) tumor weight
and (B) tumor volume, and (C) the protein levels of p53, p21,
cyclin D1, and E-cad in tumors from the antagomir-NC + sh-NC group,
antagomir-miR-181b + sh-NC group, antagomir-NC + sh-p53 group and
antagomir-miR-181b + sh-p53 group. The data are presented as the
mean ± standard deviation of three independent experiments.
*P<0.05 and **P<0.01 vs. the
antagomir-NC + sh-NC group; ##P<0.01 vs. the
antagomir-NC + sh-p53 group. miR, microRNA; OS, osteosarcoma;
E-cad, epithelial-cadherin; si, small interfering; NC, negative
control; sh, short hairpin. |
Discussion
In the present study, miR-181b expression was
demonstrated to be upregulated whereas p53 expression was
downregulated, within OS tissues and cells; in tissue samples,
miR-181b and p53 were negatively correlated. p53 might inhibit the
transcription of miR-181b via targeting its promoter region,
whereas miR-181b bound the TP53 3′-UTR to inhibit p53
expression. The inhibition of miR-181b increased p53, p21 and
E-cadherin protein levels but decreased Cyclin D1 protein levels in
OS cells and OS cell xenografts; in addition, miR-181b inhibition
reduced the capacity for OS cell proliferation and invasion both
in vitro and in vivo. In contrast, p53 knockdown had
opposite effects on these proteins and OS cell proliferation and
invasion. Above all, p53 knockdown attenuated the effects of
miR-181b inhibition.
miR-181b was reported to be abnormally upregulated
within various cancer types. The expression of miR-181b was
increased within patient serum and breast cancer cells compared
with those of healthy controls (30). Within OS cells, miR-181b OE
induced, while miR-181b inhibition decreased the ability of OS
cells to migrate and invade via directly binding NDRG2 (24). In the present study, consistent
with previous studies, miR-181b expression was shown to be
significantly enhanced, while the expression of p53, a
well-established tumor suppressor (31,32), was decreased within OS tissue
samples and cell lines.
TP53 can induce cell cycle arrest and
apoptosis by regulating the transcription of miRNAs and other
genes. p53 is regarded as mostly a transcriptional activator, but
p53 has been revealed to inhibit specific gene expression in some
reports (33,34). As demonstrated by studies on
p53-mediated suppression, both genes that regulate apoptotic
responses and genes that promote cell cycle progression can be
inhibited via p53 (35,36). In the present study, both an
online tool and experimental analysis indicated that p53 could
directly bind miR-181b. p53 inhibited the transcription of miR-181b
via targeting its promoter region. Interestingly, miR-181b also
targeted TP53 at its 3′-UTR to inhibit p53 expression, thereby
forming a negative feedback regulatory axis with p53. In OS cell
lines, p53 and miR-181b were negatively regulated by one another.
miR-181b was abnormally upregulated within OS cells, indicating
that the overexpression of miR-181b might disturb the balance of
the regulatory feedback axis to promote OS development.
To verify this speculation, the cellular effects of
miR-181b and dynamic effects of miR-181b and p53 were further
evaluated on OS cells. The oncogenic role of miR-181b has been
widely reported. miR-181b downregulation within prostate cancer
PC-3 cells induced apoptosis and suppressed PC-3 cell growth and
invasion in vitro (37).
miR-181b suppressed adenylyl cyclase 9 expression within cervical
cancer cell lines to enhance cell proliferation and to inhibit
apoptosis (38). Within
colorectal cancer, miR-181b also bound PDCD4 to act as an oncomiR,
promoting cancer cell proliferation and migratory capacity,
suppressing apoptosis within CRC cells, and accelerating tumor
growth within xenograft mice (39). In the present study, miR-181b
inhibition enhanced p53, p21 and E-cadherin protein levels while
decreasing Cyclin D1 protein levels. Apoptosis and growth arrest
are the as two main endpoints of p53 activation. Induction of the
cyclin dependent kinase suppressor p21 may be essential for
p53-mediated growth arrest. Consistently, miR-181b inhibition leads
to a significant suppression upon the proliferation and invasion of
OS cells, suggesting that the inhibition of miR-181b carries out
its effects on OS cells via targeting p53 to modulate p53-mediated
cancer cell growth arrest. As further confirmation, p53 knockdown
had effects opposite to those of miR-181b inhibition on p53
signaling and OS cells. Moreover, the effects of miR-181b
inhibition were reversed by p53 knockdown, indicating that abnormal
miR-181b upregulation inhibited p53 and promoted OS cells
proliferation and invasion. OS cell xenograft assays further
confirmed the roles of miR-181b/p53 axis in OS growth.
In conclusion, miR-181b and p53 are negatively
regulated by one another and therefore form a negative feedback
axis that regulates OS cells proliferation and invasion abilites.
Targeting miR-181b to inhibit its abnormal upregulation might be a
potent strategy for OS treatment.
Supplementary Data
Acknowledgements
The authors would like to thank Mr Xin Liu from
Central South University (Changsha, China) for his help with the
xenograft nude mice experiments.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81301671).
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author upon reasonable
request.
Authors' contributions
JW, YL made substantial contribution to the
conception and design of the study. FL, CZ analyzed and interpreted
the data. JW, CZ drafted the manuscript; CZ, YL revised the work
critically for important intellectual content. YL collected grants.
All authors approved the final published version of this
manuscript.
Ethics approval and consent to
participate
All procedures performed in studies involving human
participants were in accordance with the Ethics Committee of the
Xiangya Hospital and with the 1964 Helsinki declaration. Informed
consent to participate in the study was obtained from participants.
The animal epxeriments were approved by the Ethic Committee of
Xiangya Hospital of Central South University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ma O, Cai WW, Zender L, Dayaram T, Shen J,
Herron AJ, Lowe SW, Man TK, Lau CC and Donehower LA: MMP13, Birc2
(cIAP1), and Birc3 (cIAP2), amplified on chromosome 9, collaborate
with p53 deficiency in mouse osteosarcoma progression. Cancer Res.
69:2559–2567. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Anderson ME: Update on survival in
osteosarcoma. Orthop Clin North Am. 47:283–292. 2016. View Article : Google Scholar
|
|
3
|
Duchman KR, Gao Y and Miller BJ:
Prognostic factors for survival in patients with high-grade
osteosarcoma using the surveillance, epidemiology, and end results
(SEER) program database. Cancer Epidemiol. 39:593–599. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kedmi R, Veiga N, Ramishetti S, Goldsmith
M, Rosenblum D, Dammes N, Hazan-Halevy I, Nahary L,
Leviatan-Ben-Arye S, Harlev M, et al: A modular platform for
targeted RNAi therapeutics. Nat Nanotechnol. 13:214–219. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shaikh AB, Li F, Li M, He B, He X, Chen G,
Guo B, Li D, Jiang F, Dang L, et al: Present advances and future
perspectives of molecular targeted therapy for osteosarcoma. Int J
Mol Sci. 17:5062016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Papiol S, Arias B, Barrantes-Vidal N,
Guitart M, Salgado P, Catalán R and Fañanás L: Analysis of
polymorphisms at the tumor suppressor gene p53 (TP53) in
contributing to the risk for schizophrenia and its associated
neurocognitive deficits. Neurosci Lett. 363:78–80. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li J, Yang L, Gaur S, Zhang K, Wu X, Yuan
YC, Li H, Hu S, Weng Y and Yen Y: Mutants TP53 p.R273H and p.R273C
but not p.R273G enhance cancer cell malignancy. Hum Mutat.
35:575–584. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xie Y, Zhu S, Song X, Sun X, Fan Y, Liu J,
Zhong M, Yuan H, Zhang L, Billiar TR, et al: The tumor suppressor
p53 limits ferroptosis by blocking DPP4 activity. Cell Rep.
20:1692–1704. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bedi A and Mookerjee B: Biological
significance and molecular mechanisms of p53-induced apoptosis.
Apoptosis. 3:237–244. 1998. View Article : Google Scholar
|
|
10
|
Xie C, Wu B, Chen B, Shi Q, Guo J, Fan Z
and Huang Y: Histone deacetylase inhibitor sodium butyrate
suppresses proliferation and promotes apoptosis in osteosarcoma
cells by regulation of the MDM2-p53 signaling. Onco Targets Ther.
9:4005–4013. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou Y, Niu W, Luo Y, Li H, Xie Y, Wang H,
Liu Y, Fan S, Li Z, Xiong W, et al: p53/Lactate dehydrogenase A
axis negatively regulates aerobic glycolysis and tumor progression
in breast cancer expressing wild-type p53. Cancer Sci. 110:939–949.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen Z, Guo J, Zhang K and Guo Y: TP53
mutations and survival in osteosarcoma patients: A meta-analysis of
published data. Dis Markers. 2016:46395752016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Luo Y, Deng Z and Chen J: Pivotal
regulatory network and genes in osteosarcoma. Arch Med Sci.
9:569–575. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao YX, Wang YS, Cai QQ, Wang JQ and Yao
WT: Up-regulation of HDAC9 promotes cell proliferation through
suppressing p53 transcription in osteosarcoma. Int J Clin Exp Med.
8:11818–11823. 2015.PubMed/NCBI
|
|
15
|
Wu J, Guo A, Li Q and Wang D:
Meta-analysis of clinical significance of p53 protein expression in
patients with osteosarcoma. Future Oncol. 13:1883–1891. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shukla GC, Singh J and Barik S: MicroRNAs:
Processing, maturation, target recognition and regulatory
functions. Mol Cell Pharmacol. 3:83–92. 2011.PubMed/NCBI
|
|
17
|
Farh KK, Grimson A, Jan C, Lewis BP,
Johnston WK, Lim LP, Burge CB and Bartel DP: The widespread impact
of mammalian MicroRNAs on mRNA repression and evolution. Science.
310:1817–1821. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pasquinelli AE, Hunter S and Bracht J:
MicroRNAs: A developing story. Curr Opin Genet Dev. 15:200–205.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jones KB, Salah Z, Del Mare S, Galasso M,
Gaudio E, Nuovo GJ, Lovat F, LeBlanc K, Palatini J, Randall RL, et
al: miRNA signatures associate with pathogenesis and progression of
osteosarcoma. Cancer Res. 72:1865–1877. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qin L, Chen Y, Niu Y, Chen W, Wang Q, Xiao
S, Li A, Xie Y, Li J, Zhao X, et al: A deep investigation into the
adipogenesis mechanism: Profile of microRNAs regulating
adipogenesis by modulating the canonical Wnt/beta-catenin signaling
pathway. BMC Genomics. 11:3202010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guo Y, Zi X, Koontz Z, Kim A, Xie J,
Gorlick R, Holcombe RF and Hoang BH: Blocking Wnt/LRP5 signaling by
a soluble receptor modulates the epithelial to mesenchymal
transition and suppresses met and metalloproteinases in
osteosarcoma Saos-2 cells. J Orthop Res. 25:964–971. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zang X, Li Q, Wang W, Zhou Y, Chen S and
Xiao T: miR-181a promotes the proliferation and metastasis of
osteosarcoma cells by targeting RASSF1A. Zhong Nan Da Xue Xue Bao
Yi Xue Ban. 41:789–795. 2016.in Chinese; Abstract available in
Chinese from the publisher. PubMed/NCBI
|
|
23
|
Zhu ZJ, Huang P, Chong YX, Kang LX, Huang
X, Zhu ZX and Nie L: MicroRNA-181a promotes proliferation and
inhibits apoptosis by suppressing CFIm25 in osteosarcoma. Mol Med
Rep. 14:4271–4278. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shao JL, Li ZZ, Wang L, Jiao GL, Zhou ZG
and Sun GD: microRNA-181b promotes migration and invasion of
osteosarcoma cells by targeting N-myc downstream regulated gene 2.
Nan Fang Yi Ke Da Xue Xue Bao. 36:321–326. 2016.In Chinese.
PubMed/NCBI
|
|
25
|
Wang W, Wang Z, Chen S, Zang X and Miao J:
Interleukin-1β/nuclear factor-κB signaling promotes osteosarcoma
cell growth through the microRNA-181b/phosphatase and tensin
homolog axis. J Cell Biochem. 120:1763–1772. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kaste SC, Pratt CB, Cain AM, Jones-Wallace
DJ and Rao BN: Metastases detected at the time of diagnosis of
primary pediatric extremity osteosarcoma at diagnosis: Imaging
features. Cancer. 86:1602–1608. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
McCarville MB, Kaste SC, Cain AM,
Goloubeva O, Rao BN and Pratt CB: Prognostic factors and imaging
patterns of recurrent pulmonary nodules after thoracotomy in
children with osteosarcoma. Cancer. 91:1170–1176. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nolan T, Hands RE and Bustin SA:
Quantification of mRNA using real-time RT-PCR. Nat Protoc.
1:1559–1582. 2006. View Article : Google Scholar
|
|
29
|
Liu ZB, Wang JA and Lv RQ: Downregulation
of long non-coding RNA DBH-AS1 inhibits osteosarcoma progression by
PI3K-AKT signaling pathways and indicates good prognosis. Eur Rev
Med Pharmacol Sci. 23:1418–1427. 2019.PubMed/NCBI
|
|
30
|
Zheng Y, Lv X, Wang X, Wang B, Shao X,
Huang Y, Shi L, Chen Z, Huang J and Huang P: MiR-181b promotes
chemoresistance in breast cancer by regulating Bim expression.
Oncol Rep. 35:683–690. 2016. View Article : Google Scholar
|
|
31
|
Hong B, van den Heuvel AP, Prabhu VV,
Zhang S and El-Deiry WS: Targeting tumor suppressor p53 for cancer
therapy: Strategies, challenges and opportunities. Curr Drug
Targets. 15:80–89. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou J, Wu S, Chen Y, Zhao J, Zhang K,
Wang J and Chen S: microRNA-143 is associated with the survival of
ALDH1+CD133+ osteosarcoma cells and the
chemoresistance of osteosarcoma. Exp Biol Med (Maywood).
240:867–875. 2015. View Article : Google Scholar
|
|
33
|
Mack DH, Vartikar J, Pipas JM and Laimins
LA: Specific repression of TATA-mediated but not initiator-mediated
transcription by wild-type p53. Nature. 363:281–283. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dickins RA, Hemann MT, Zilfou JT, Simpson
DR, Ibarra I, Hannon GJ and Lowe SW: Probing tumor phenotypes using
stable and regulated synthetic microRNA precursors. Nat Genet.
37:1289–1295. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mirza A, Wu Q, Wang L, McClanahan T,
Bishop WR, Gheyas F, Ding W, Hutchins B, Hockenberry T, Kirschmeier
P, et al: Global transcriptional program of p53 target genes during
the process of apoptosis and cell cycle progression. Oncogene.
22:3645–3654. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Spurgers KB, Gold DL, Coombes KR,
Bohnenstiehl NL, Mullins B, Meyn RE, Logothetis CJ and McDonnell
TJ: Identification of cell cycle regulatory genes as principal
targets of p53-mediated transcriptional repression. J Biol Chem.
281:25134–25142. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
He L, Yao H, Fan LH, Liu L, Qiu S, Li X,
Gao JP and Hao CQ: MicroRNA-181b expression in prostate cancer
tissues and its influence on the biological behavior of the
prostate cancer cell line PC-3. Genet Mol Res. 12:1012–1021. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang L, Wang YL, Liu S, Zhang PP, Chen Z,
Liu M and Tang H: miR-181b promotes cell proliferation and reduces
apoptosis by repressing the expression of adenylyl cyclase 9 (AC9)
in cervical cancer cells. FEBS Lett. 588:124–130. 2014. View Article : Google Scholar
|
|
39
|
Liu Y, Uzair-Ur-Rehman, Guo Y, Liang H,
Cheng R, Yang F, Hong Y, Zhao C, Liu M, Yu M, et al: miR-181b
functions as an oncomiR in colorectal cancer by targeting PDCD4.
Protein Cell. 7:722–734. 2016. View Article : Google Scholar : PubMed/NCBI
|