Introduction
Non-small cell lung cancer (NSCLC) is one of the
leading causes of cancer related death in the world, with its
5-year survival rate <20% after diagnosis (1). Although targeted molecular therapy
has achieved great success in treatment of NSCLC, it is usually
limited to a group of patients harboring drug-sensitive epidermal
growth factor receptor (EGFR) mutations (2). Platinum-based chemotherapy remains
the main treatment option for NSCLC with wild-type EGFR, but the
efficacy is still not satisfactory. Combination therapy has been
widely studied and used to increase the efficacy of tyrosine kinase
inhibitors (TKIs) or chemotherapeutics on EGFR wild-type lung
cancer cells (3-5). Nevertheless, new therapeutic targets
are urgently needed in order to improve the therapeutic outcome of
the current therapy for these NSCLC patients.
Cluster of differentiation 44 (CD44), a
transmembrane glycoprotein, serves as an oncogenic regulator as
well as a cancer stem cell marker in numerous kinds of malignancies
(6). CD44 is found to be
over-expressed in cancer tissues and was significantly associated
with progression, migration and multi-drug resistance of various
cancers such as colorectal cancer, breast cancer and lung cancer
(7,8). Previous studies have shown that the
expression of CD44 is correlated with EGFR level in a variety of
neoplasms (9-11). It has been indicated that the TKI
erlotinib treatment significantly downregulated the CD44 level and
inhibited breast cancer cell migration and invasion (9). Moreover, one study also has shown
that the EGFR ligand, EGF increased the expression of CD44 as well
as the phosphorylation of ERK, STAT3 and AKT in SKBR3 breast cancer
cells (12). On the other hand,
it was indicated that CD44 is a promoting modulator for EGFR
activation. For example, Perez et al (13) showed that CD44 augmented
tumorigenesis and progression in head and neck squamous cell
carcinoma through interaction with EGFR. This provides direct
evidence for the relationship between CD44 and EGFR signaling.
Recently, it has been shown that CD44s, a splicing isoform of CD44,
could stabilize protein level of receptor tyrosine kinases (RTKs)
through interaction with Rab7A and the absence of CD44 facilitated
Rab7A-mediated trafficking of EGFR to lysosomes in glioblastoma
cells, contributing to EGFR degradation (14). In breast cancer, specific CD44
subtypes are recruited as co-receptors in the EGFR signaling
pathway in a ligand-dependent manner and their specificity is
determined by the ligand rather than the receptor itself (15). Hyaluronan facilitates transforming
growth factor-β1 (TGF-β1)-dependent activation of MAPK/ERK by
promoting the interaction between CD44 and EGFR, thereby promoting
cellular proliferation of fibroblasts (16). CD44 appears to be both a
co-regulator of RTK signaling and a downstream target of EGFR
signaling. However, the relationship of CD44 and EGFR or the role
of CD44 in modulation of EGFR signaling in NSCLC cells has not been
well investigated.
The present study hypothesized that blocking CD44
may result in altered EGFR signaling and increase sensitivity of
wild-type EGFR NSCLC cells to chemotherapeutics such as cisplatin.
The present study thus focused on wild-type EGFR NSCLC cell line
H460 and investigated the role of CD44 in regulation of EGFR
signaling as well as its impact on platinum-based chemotherapy. The
present study will provide new perspectives for enhancing the
efficacy of chemotherapeutics in clinical treatment for EGFR
wild-type NSCLC patients.
Materials and methods
Cell culture
Human EGFR wild-type NSCLC cell line H460 was
obtained from the School of Biomedical Sciences, Chinese University
of Hong Kong. The cell line was cultured in RPMI-1640 medium
(Gibco; Thermo Fisher Scientific, Inc.), supplemented with 10%
fetal bovine serum (FBS; Thermo Fisher Scientific, Inc.) and 100
µg/ml streptomycin/100 U/ml penicillin (Gibco; Thermo Fisher
Scientific, Inc.). All cells were maintained at 37°C under a
humidified atmosphere with 5% CO2 in an incubator (SHEL
LAB, Inc.). Cells were passaged by trypsin/EDTA (Gibco; Thermo
Fisher Scientific, Inc.) and the culture medium was changed every
other day.
RNA interference
The expression of CD44 was downregulated using
pre-designed target-specific small interfering RNA (siRNAs). Cells
were transfected with siRNA by using the jetPRIME reagent
(Polyplus-Transfection SA). The control siRNA (sictr,
siN05815122147) and CD44 siRNA (siCD44, siG000000960B) were
purchased from Guangzhou RiboBio Co., Ltd. The target sequence of
CD44 siRNA was 5′-CCG CTT TGC AGG TGT ATT C-3′. By using the Basic
Local Alignment Search Tool (BLAST) at NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi), the
targeted sequence was found to match well and specifically with
both standard and variant forms (transcript variant 1-8, x1-x19) of
CD44, suggesting that the designed siCD44 can knock down CD44 and
its variant forms. The transfection efficiency was evaluated by
RT-qPCR and western blot analysis. A total of 50 nM siCD44 or sictr
mixed with jetPRIME reagent (Polyplus-Transfection SA) was added to
the cells for 24 h, following which the solution was replaced with
normal culture medium. Follow-up experiments were conducted 48 h
after transfection.
Cell viability
Cells were seeded at a density of 10,00-3,000
cells/well in 96-well plates (Guangzhou Jet Bio-Filtration Co.,
Ltd.). Then cells were cultured overnight in an incubator at 37°C.
Next, cells were cultured with or without cisplatin at different
concentrations (2.5, 5, 10 and 20 µM). Cell viability was
detected using a Cell Counting Kit (CCK)-8 (Dojindo Molecular
Technologies, Inc.) according to the manufacturer's protocol
whereby the CCK-8 was diluted with RPMI-1640 (1:10). The culture
medium was replaced before adding 100 µl diluted solution
per well. The plates were incubated for another 3 h in an
incubator. The optical density at 450 nm was then measured with a
microplate reader (CYTATION 3; Agilent Technologies, Inc.).
Cell cycle analysis
Cells were seeded into 6-well plates at
1.5×105 cells/well. They were later transfected with
siRNAs for 24 h and then treated with or without cisplatin (10
µM) for 24 h. The cells were harvest after digestion by
0.25% trypsin, washed with PBS and fixed with cold 70% ethanol for
at least 24 h at −20°C. Fixed cells were washed with PBS and then
allowed to incubate for 30 min at 37°C prior to analysis with a
propidium iodide solution: 50 µg/ml propidium iodide
(Invitrogen; Thermo Fisher Scientific, Inc.) and 10 mg/ml RNase A
(Roche Diagnostics). Fluorescence intensity of PI was measured by
flow cytometry (BD FACS Canto™; Becton, Dickinson and Company),
which reflects the individual nuclear DNA content. The proportion
of G0/G1, S and G2/M cells in cell
cycle distribution were analyzed by ModFit LT software (MFLT32;
Verity Software House, Inc.). The experiment was repeated three
times.
Apoptosis assay
Apoptosis was analyzed with an Annexin V-FITC/PI
detection kit (Invitrogen; Thermo Fisher Scientific, Inc.).
Transfected cells were collected and were stained with 5 µl
Annexin V-FITC and 10 µl PI per sample in the dark for 10
min at room temperature. Finally, cell apoptosis was detected and
quantified by a flow cytometer (BD FACS Canto™; Becton, Dickinson
and Company). A total of ~10,000 cells were collected for data
analysis.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated using Trizol reagent (Thermo
Fisher Scientific, Inc.) based on the manufacturer's protocol. cDNA
was synthesized from mRNA using a FastKing RT reagent kit (Tiangen,
Inc.). The RT reaction was performed at 42°C for 15 min and
95°C for 3 min. QPCR was performed with a SYBR Green
Real Time PCR kit (Thermo Fisher Scientific, Inc.) on CFX96 Touch
Real Time PCR System (BioRad Laboratories, Inc.) under the
following conditions: 95°C for 1 min, then 40 cycles of 95°C for 5
sec and 60°C for 15 sec. The primers used for qPCR were as follows:
Human CD44, forward 5′-TGG AGA AAA ATG GTC GCT ACA G-3′, reverse
5′-GGG CAA GGT GCT ATT GAA AGC-3′; human GAPDH, forward 5′-CTG GGC
TAC ACT GAG CAC C-3′, reverse 5′-AAG TGG TCG TTG AGG GCA ATG-3′.
The formula 2−ΔΔCq was employed to analyze the fold
change of mRNA expression levels (17).
Western blotting
Western blotting was used to test the EGFR signaling
pathway-associated proteins, as well as the levels of proteins
associated with the cell cycle and apoptosis. RIPA lysis buffer
(Beijing Solarbio Science & Technology Co., Ltd.) containning
protease inhibitor PhosSTOP EASYpack (Roche Diagnostics) was used
for total protein extraction according to the manufacturer's
protocol. Equal amounts of protein samples (25 µg) were
separated by 10% SDS-PAGE gel and transferred to a polyvinylidene
fluroride hybridization transfer membrane (Immobilon-p; EMD
Millipore). After blocking with 5% bovine serum albumin (BSA,
Sigma-Aldrich; Merck KGaA) at room temperature for 1 h and the
membranes were probed with the corresponding primary antibodies
incubated at 4°C overnight. Afterwards, membranes were inclubated
with HRP-conjugated secondary antibodies at room temperature for 2
h. Chemiluminescent signals were finally reacted with ECL blotting
detection reagents (Clarity; Bio-Rad Laboratories, Inc.) and the
signals were detected by the ChemiDoc™ Imaging System (Bio-Rad
Laboratories, Inc.). Band density were evaluated by ImageJ 1.48v
(National Institutes of Health). Primary antibodies against
phosphorylated (p)-EGFR (Tyr1173; cat. no. 4407; 1:1,000), Src
(cat. no. 2123; 1:1,000), p-Src (Tyr416; cat. no. 6943; 1:1,000),
CD44 (cat. no. 5640s; 1:1,000), ERK (cat. no. 4695; 1:1,000), p-ERK
(cat. no. 4370; 1:1,000), CDK2 (cat. no. 2546s; 1:1,000), CDK4
(cat. no. 12790s; 1:1,000), CDK6 (cat. no. 13331s; 1:1,000), Bax
(cat. no. 2774; 1:1,000) and Bcl-2 (cat. no. 4223; 1:1,000) was
from Cell Signaling Technology, Inc. Primary antibodies against
EGFR (cat. no. ET1603-37; 1:1,000), p-EGFR (Y1068; cat. no.
ET1612-30; 1:1,000), GAPDH (cat. no. M1310-2; 1:1,000), AKT1/2/3
(cat. no. ET1609-51; 1:1,000), p-AKT1 (Ser473; cat. no. ET1607-73;
1:1,000), as well as the secondary antibodies of goat anti-rabbit
IgG HRP (cat. no. HA1006; 1:1,000) and goat anti-mouse IgG HRP
(cat. no. HA1001; 1:1,000) were purchased from HUABIO. Anti-c-Myc
(cat. no. 469301-22; 1:200) was purchased from Calbiochem, while
anti-CyclinD1 (cat. no. SC450; 1:200) was purchased from Santa Cruz
Biotechnology, Inc. Cycloheximide was purchased from
MedChemExpress.
Immunofluorescence assay
H460 cells were seeded in 8-well chamber (EMD
Milipore) at ~1×104 cells/well. Cells were transfected
with CD44 siRNA as well as control siRNA with JetPrime. After 48 h
incubation, the cells were washed three times with PBS, fixed for
15 min in 4% paraformaldehyde and permeabilized with 0.5%
TritonX-100 (Sigma-Aldrich; Merck KGaA) in PBS for 20 min at room
temperature. They were later washed three times with PBS and
blocked with 5% BSA for 30 min at room temperature. Cells were
incubated with rabbit anti-EGFR (cat. no. ET1603-37; 1:400; HUABIO)
and mouse anti-CD44 (cat. no. 5640s; 1:800; Cell Signaling
Technology, Inc.) for 1 h at room temperature. Subsequently, the
slide was washed with PBS three times and incubated with secondary
antibody conjugated to the fluorescent dye Alexa Fluor™ 488 goat
anti-rabbit (1966932; 1:800; Invitrogen; Thermo Fisher Scientific,
Inc.) and Alexa Fluor™ 568 goat anti-mouse (1906484; 1:1,600;
Invitrogen; Thermo Fisher Scientific, Inc.) for 1 h in dark at room
temperature, and mounted with DAPI (Abcam) mounting medium at room
temperature for 5 min. Finally, the images were captured using a
fluorescent microscope (Nikon Ts2R; Nikon Corporation).
The cancer genome atlas (TCGA) data
collection and bioinformatics analysis
The scatter diagram was based on Log2(RSEM+1)
analysis using data from 255 lung adenocar-cinoma samples in TCGA
(https://portal.gdc.cancer.gov/) and
tumor samples (n=224) with EGFR wild-type were selected. The
cBioPortal (http://www.cbioportal.org/) for Cancer Genomics
provides information on EGFR mutation.
Statistical analysis
Statistical analysis of the data was carried out
using GraphPad Prism 7.0 (GraphPad Software, Inc.). The data are
expressed as mean ± SD of three independent experiments. Unpaired
student t test, or one-way ANOVA with a post hoc Tukey test was
used for analysis of difference between two groups or among three
or more groups, respectively. The spearman correlation between EGFR
and CD44 was analyzed by GraphPad Prism 7.0 (GraphPad Software,
Inc.). P<0.05 was considered as significant difference.
Results
Downregulation of CD44 inhibits growth of
EGFR wild-type NSCLC cells and the effect is enhanced when combined
with cisplatin treatment
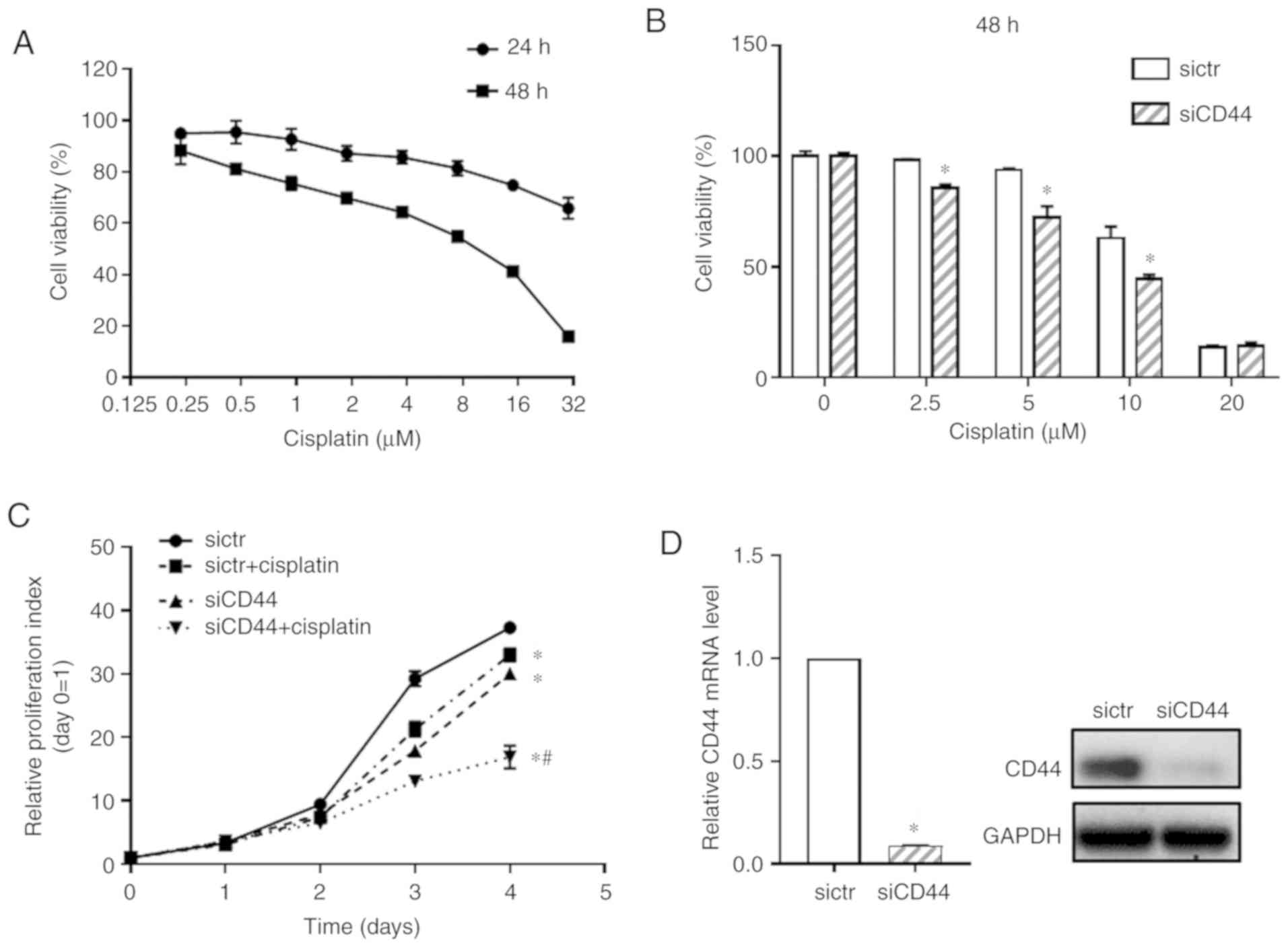
First, the cytotoxicity of cisplatin on EGFR
wild-type NSCLC cell H460 was detected (Fig. 1A). Cell viability of H460 cells
treated with cisplatin at concentrations from 0.2 to 30 µM
were tested by using the CCK-8 assay and IC50 of
cisplatin at 48 h was 6.852±0.405 µM. To examine the
functional relevance of CD44 expression in cisplatin sensitivity of
EGFR wild-type NSCLC cells, the CD44 level in the H460 cell line
was downregulated using specific RNA interference. It was confirmed
that, compared with blank cells, transfection with sictr did not
significantly affect CD44 expression and cell growth of H460 cells
(Fig. S1). Thus, the H460 cells
transfected with sictr were used for direct comparison with that
transfected with siCD44 in the following experiments. As shown in
Fig. 1B, after knockdown of CD44,
cisplatin significantly inhibited cell growth of H460 at
concentration of 2.5, 5 and 10 µM when compared with the
sictr group and 10 µM for cisplatin was selected following
in vitro studies. The proliferative curve of H460 was
further detected after silencing CD44 combined with cisplatin
treatment (10 µM). As shown in Fig. 1C, knockdown of CD44 significantly
inhibited cell growth of H460. Additionally, the present study
found that downregulation of CD44 followed by cisplatin treatment
inhibited the proliferation of H460 cell compared with sictr +
cisplatin group. Fig. 1D shows
that knockdown efficiency of siCD44 in H460 was >90% at 48 h
post transfection based on RT-qPCR assessment. Although there was
no synergistic effect of siCD44 and cisplatin, their combination
was effective in suppressing tumor cells. The results also
indicated cisplatin treatment might be more effective and could
achieve a better outcome for wild-type EGFR positive NSCLC patients
with low CD44 expression.
Downregulation of CD44 promotes
G0/G1 cell cycle arrest and apoptosis in H460
cells and the effects are significantly enhanced when in
combination with cisplatin treatment
It has been reported that heterodimeric complexes of
CDK4/6 with cyclin D and CDK3 with cyclin C regulated cell-cycle
transition from G0 to G1 and early phases of
G1 through the phosphorylation of retinoblastoma protein
(18) while S and G2/M
phase control were mediated by CDK2 and CDK1 (19). It was further observed that
knockdown of CD44 in H460 cells led to a significant
G0/G1 cell cycle arrest compared with the
control (Fig. 2A and B), which
was associated with downregulation of cell cycle related proteins
such as C-myc, CDK2, CDK4, and CDK6 (Fig. 2C). Cisplatin alone induced a
slight S phase arrest in H460 cells (Fig. 2B) with decreased expression of
CDK6 (Fig. 2C). Its combination
with CD44 silencing resulted in a significant increase of
G0/G1 populations in H460 cells (Fig. 2B) with a further repression of
corresponding proteins (Fig. 2C).
The cyclin D1 expression remain unchanged (Fig. 2C), suggesting that neither
cisplatin treatment nor siCD44 could influence cyclin D1 expression
in the NSCLC cell line H460. The cell cycle arrest is believed to
be mainly relying on the downregulation of CDKs as well as other
cyclins, such as cyclin C.
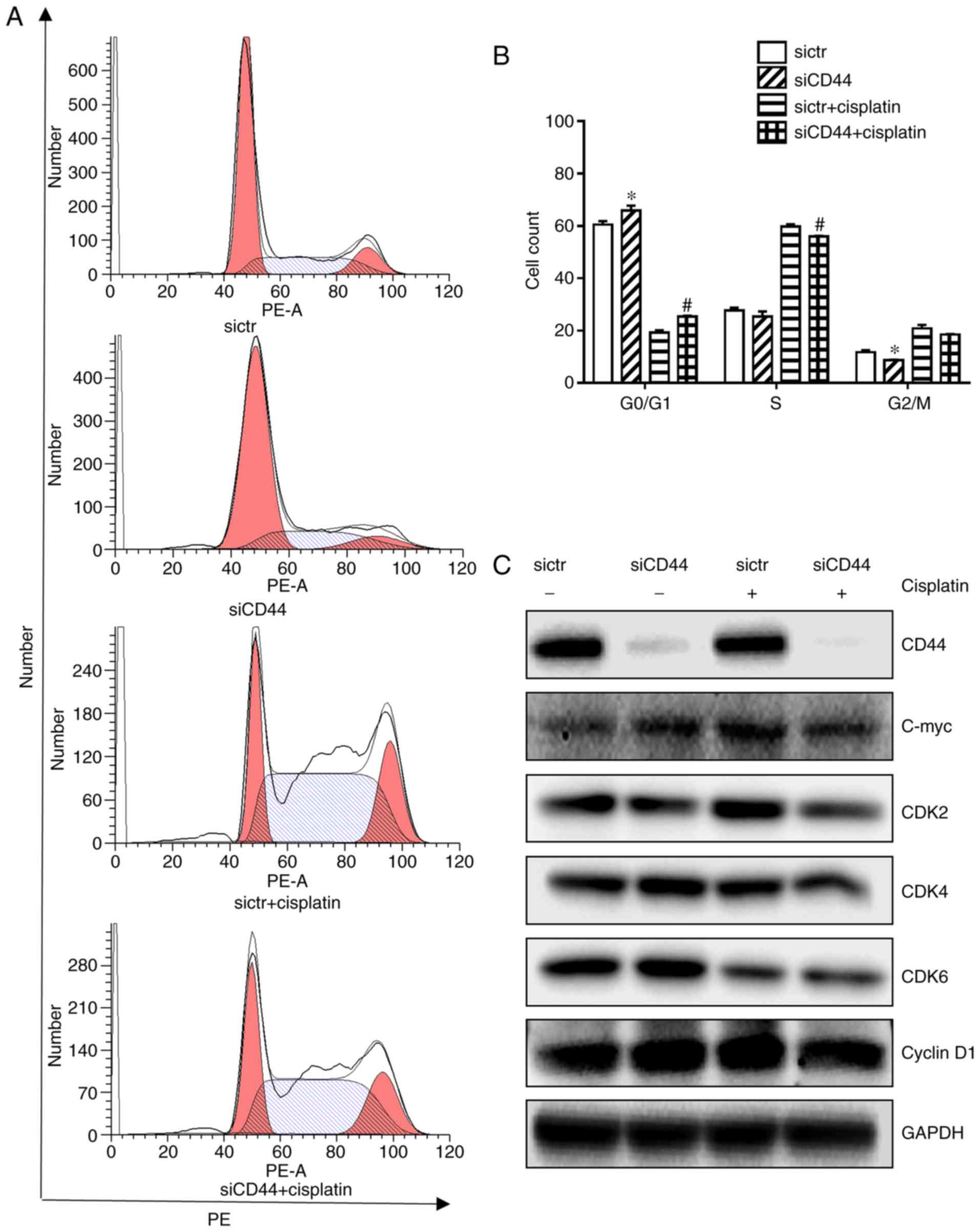 | Figure 2Effects of siCD44, cisplatin or their
combination on cell cycle distribution of H460 cells. H460 cells
were seeded into 6-well plate at 1.5×105 cells/well and
transfected with sictr and siCD44 respectively for 24 h, followed
by treatment of cisplatin at concentration of 10 µM for
another 24 h. (A) Transfected cells were stained with PI for cell
cycle analysis by flow cytometry. Cell distribution was analyzed by
MFLT32. (B) Corresponding statistical analysis of cell
distribution. (C) The protein expressions of CD44, c-Myc, CDK2,
CDK4, CDK6 and cyclin D1 in H460 were analyzed by western blotting,
with GAPDH as loading control. Data were presented as mean ± SD of
3 independent experiments. *P<0.05 vs. sictr;
#P<0.05 vs. sictr + cisplatin. sictr, control siRNA;
siCD44, CD44 siRNA; PI, propidium iodide. |
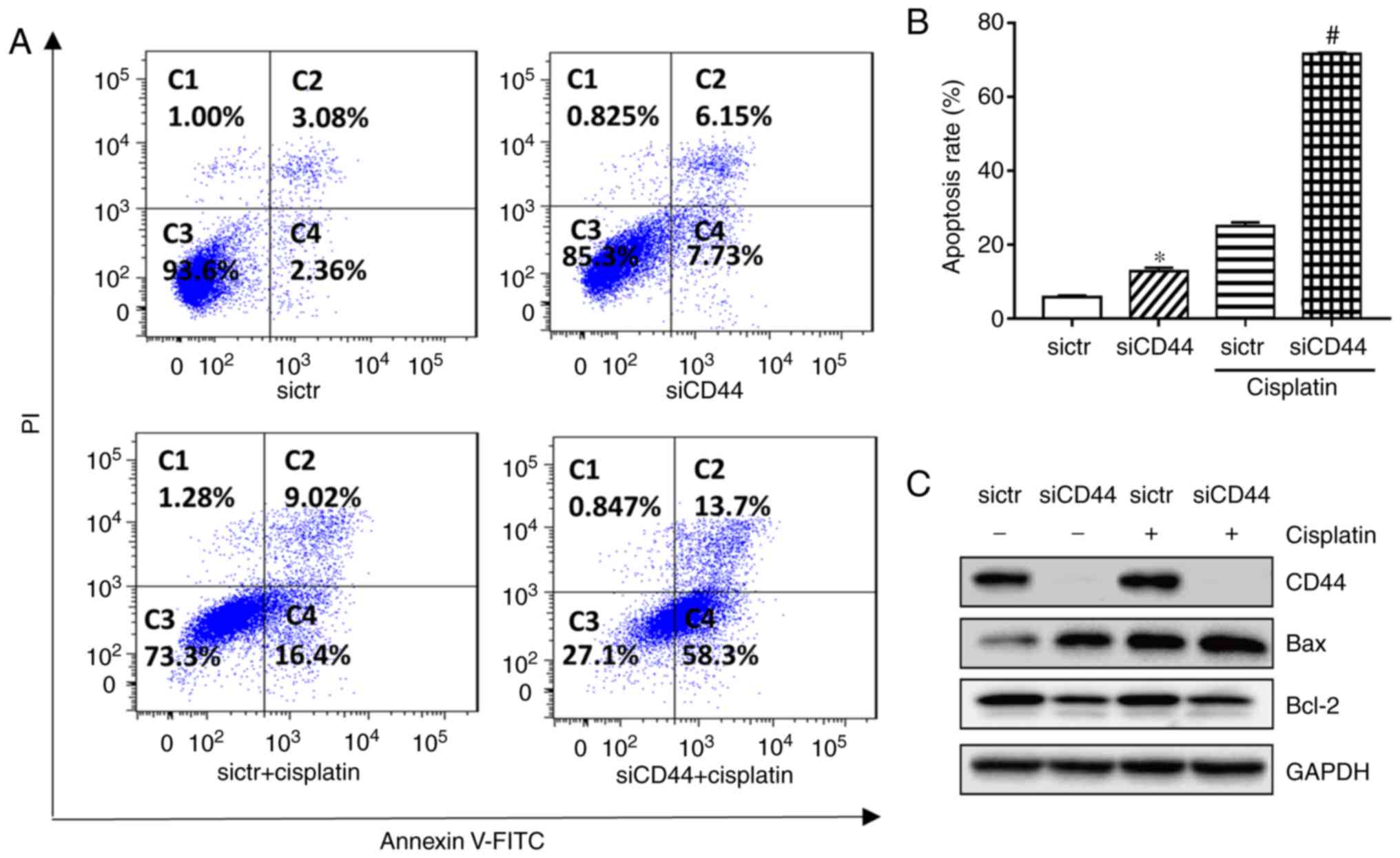
Flow cytometry results indicated that knockdown of
CD44 significantly increased the apoptotic population from 5.8 to
13.2% (Fig. 3A and B) in H460
cells. Western-blot analysis showed that expression of the
pro-apoptotic protein Bax was increased while that of
anti-apoptotic protein Bcl-2 was decreased after knockdown of CD44
in H460 cells (Fig. 3C). The
apoptotic rate of cisplatin alone was 25.4% in H460, while its
combination with CD44 knockdown resulted in an enhanced anticancer
effect. The apoptotic population in the combination group increased
to 72.0% in H460, while the corresponding protein Bax and Bcl-2
level was changed (Fig. 3). The
results demonstrated that suppression of CD44 promoted cell cycle
arrest and cell apoptosis of EGFR wild-type NSCLC cells and further
increased the antitumor effect of cisplatin.
EGFR signaling is attenuated by CD44
silencing in EGFR wild-type NSCLC cells
In order to investigate the effect of silencing CD44
on EGFR signaling in H460 cells, western blotting was performed to
discover the expression level of EGFR, AKT and ERK as well as their
phosphorylated form. EGF was used to stimulate the EGFR signaling
pathway in H460 cells. As shown in Fig. 4A, EGF-mediated activation of EGFR
in H460 cells was reduced in the siCD44 group, as reflected by
protein expression of p-EGFR (Tyr1068 and Tyr1173) within 60 min.
Furthermore, protein expression of p-SRC (Tyr416), p-AKT (Ser473),
and p-ERK (Tyr204) was decreased, suggesting that the activation of
EGFR downstream signaling including SRC, ERK and AKT was
attenuated. All experiments were performed in triplicate and the
quantification results for the protein band intensity are shown in
Fig. 4B-F. The results indicated
that knockdown of CD44 attenuated EGF-mediated stimulation of EGFR
signaling in H460 cells. It is further confirmed that siCD44 led to
direct inhibition of EGFR signaling (Fig. 5A), which might contribute to the
increased sensitivity of EGFR wild-type NSCLC cells to
chemotherapy.
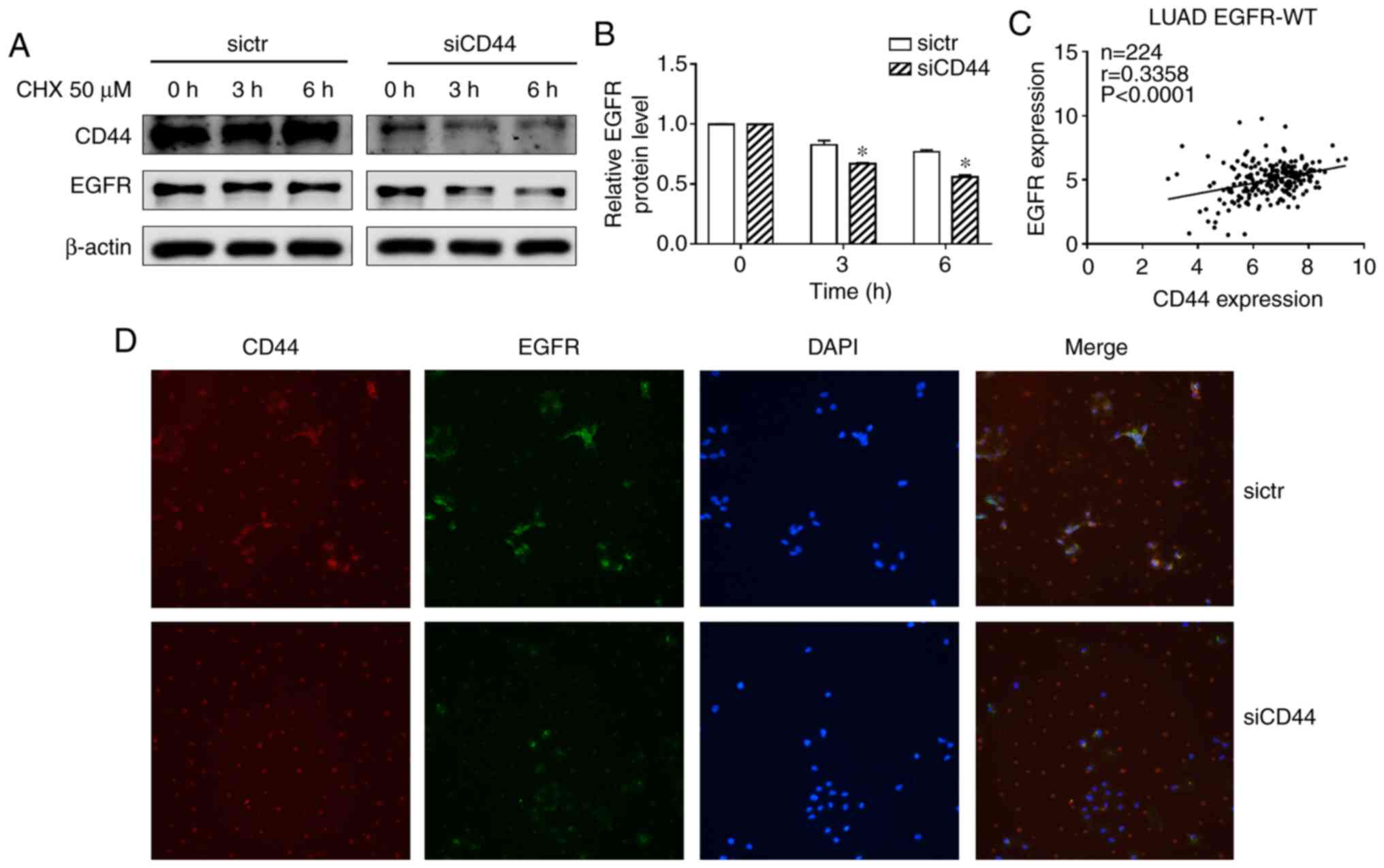
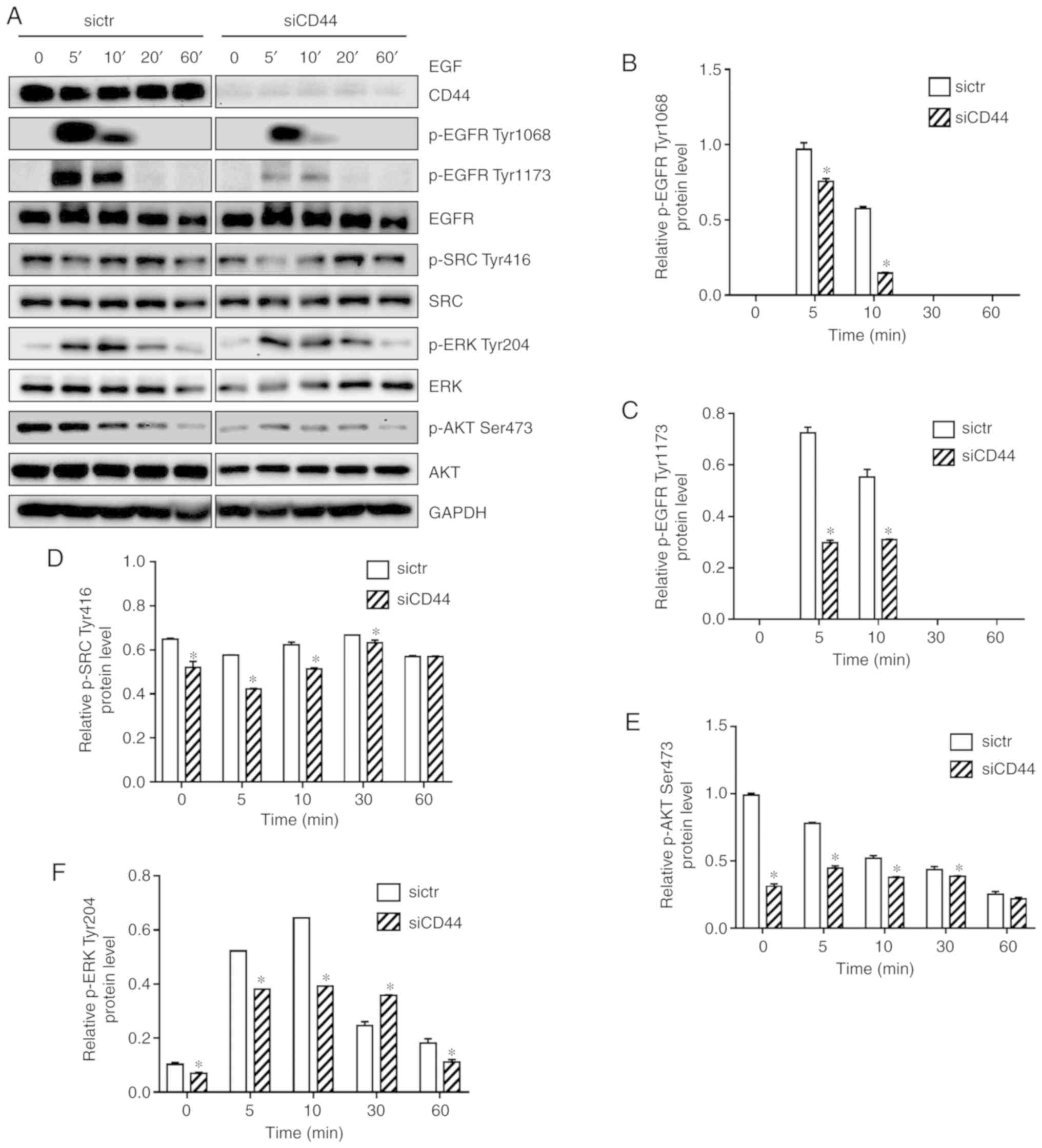 | Figure 4Downregulation of CD44 attenuates
EGF-mediated activation of EGFR signaling, potentially through
enhanced EGFR degradation. (A) Cells were seeded into 6-well plate
at 1.5×105 cells/well and transfected with sictr or
siCD44 for 24 h. After stimulation by EGF (50 ng/ml) for 0, 5, 10,
20 and 60 min, total protein were collected for western blotting.
The protein expression of EGFR, p-EGFR Tyr1068, p-EGFR Tyr1173,
SRC, p-SRC Tyr416, AKT, p-AKT Ser473, ERK and p-ERK Tyr204 were
assessed. Accordingly, the relative expressions of (B) p-EGFR
Tyr1068, (C) p-EGFR Tyr1173, (D) p-SRC Tyr416, (E) p-AKT Ser473,
and (F) p-ERK Tyr204 to GAPDH were calculated by ImageJ. Data were
presented as mean ± SD of 3 independent experiments.
*P<0.05 vs. sictr. sictr, control siRNA; siCD44, CD44
siRNA; p, phosphorylated; EGFR, epidermal growth factor
receptor. |
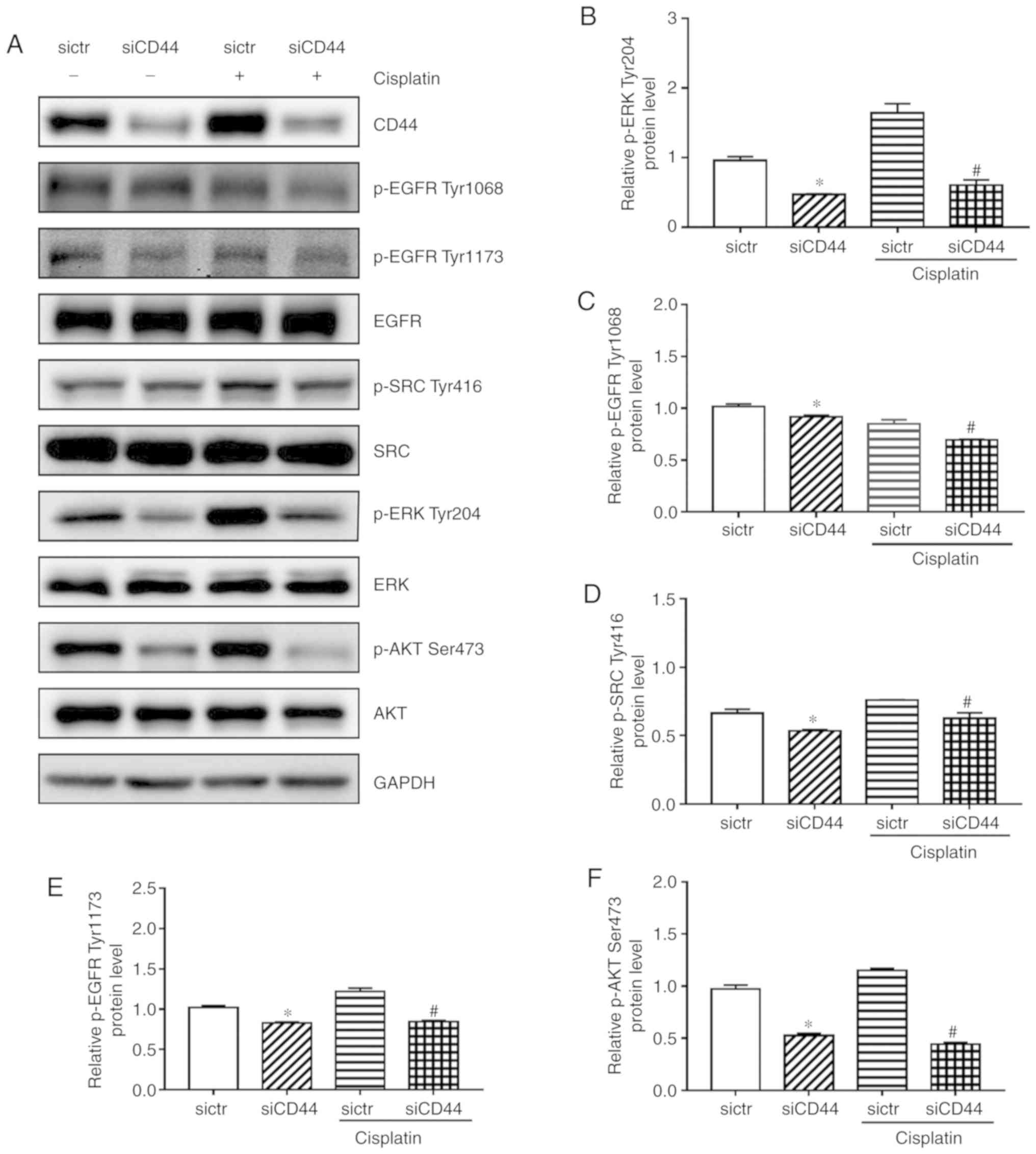 | Figure 5Influences of siCD44, cisplatin or
their combination on EGFR signaling in H460 cells. H460 cells were
seeded into 6-well plate at 1.5×105 cells/well and
transfected with sictr or siCD44 for 24 h. (A) The protein levels
of (B) p-ERK Tyr204 (C) p-EGFR Tyr1068, (D) p-SRC Tyr416, (E)
p-EGFR Tyr1173, (F) p-AKT Ser473, were assessed by western
blotting, with their relative expression to GAPDH calculated using
ImageJ. Data are presented as mean ± SD of 3 independent
experiments. *P<0.05 vs. sictr; #P<0.05
vs. sictr + cisplatin. sictr, control siRNA; siCD44, CD44 siRNA; p,
phosphorylated; EGFR, epidermal growth factor receptor. |
CD44 inhibition enhances cisplatin
sensitivity in H460 cells due to possible deactivation of EGFR
signaling
The current study demonstrated that EGFR signaling
pathway was attenuated after silencing of CD44 in H460 cells
(Fig. 4), which could be the
reason for siCD44-induced higher efficacy of cisplatin. In order to
confirm that, the EGFR signaling activation including the
phosphorylation of EGFR, SRC, AKT and ERK was observed after
knockdown of CD44 as well as in combination with cisplatin. While
cisplatin alone had a limited effect on EGFR signaling, as revealed
by the unchanged protein expression levels of p-EGFR (Tyr1068 and
Tyr1173), p-SRC (Tyr416), p-AKT (Ser473), and p-ERK (Tyr204),
cisplatin plus CD44 knockdown led to significantly repressed
phosphorylation of these proteins (Fig. 5). These results confirmed that
suppression of EGFR signaling by siCD44 contributed to the
increased efficacy of cisplatin on EGFR wild-type NSCLC cells.
Downregulation of CD44 enhances EGFR
degradation in H460 and EGFR level is positively correlated with
CD44 expression in EGFR wild-type NSCLC tissues
A previous study suggested that CD44 may interfere
with EGFR degradation (14). In
the present experiments, a protein synthesis inhibitor
cycloheximide (CHX) was used to block protein synthesis in NSCLC
cells. The EGFR level decreased after treatment with CHX in the
siCD44 group suggesting that CD44 stabilized the EGFR level while
CD44 silencing accelerated EGFR degradation (Fig. 6A and B). This indicated that
suppression of CD44 expression contributed to a reduction in EGFR
activation via increased degradation of EGFR. An immunofluorescence
(IF) assay was also performed to stain CD44 and EGFR to confirm
this result. As shown in Fig. 6D,
after knockdown of CD44, EGFR IF staining was significantly
decreased, which suggested that siCD44 promoted EGFR degradation,
thereby resulting in deactivation of EGFR downstream signaling. On
the other hand, in order to elucidate the correlation between CD44
and EGFR in lung cancer, the CD44 and EGFR expression data of lung
adenocarcinoma patient tissues with wild-type EGFR was obtained
from TCGA database. The correlation between CD44 and EGFR mRNA
level was analyzed and the result showed that CD44 was positively
associated with EGFR level in EGFR wild-type lung cancer tissues
(Fig. 6C). This further confirmed
the findings and pointed to the importance of screening CD44
expression in lung cancer tissue to achieve a better outcome for
lung cancer patients undergoing chemotherapy.
Discussion
EGFR mutations or hyperactivation is common among
lung cancer patients. While EGFR TKIs achieved a favorable outcome
in EGFR-mutant patients and were recommended as first-line
treatment for patients with positive EGFR mutation (20), chemotherapeutics including
cisplatin, carboplatin, etoposide and docetaxel were used as the
first line of treatment for EGFR-wild-type patients (21). However, the outcome of
chemotherapy in EGFR wild-type NSCLC tumors remains poor (22). Cisplatin, a commonly used
chemotherapeutic, is widely used for treatment of NSCLC patients,
while both its cytotoxicity and effective dose are relatively high
(23). It is important to find
novel therapeutic strategies to increase the efficacy of cisplatin
while reducing its side effects. The current study pointed to the
functional role of CD44 in regulating wild-type EGFR activation,
which provided a way to increase the effectiveness of cisplatin in
treatment of EGFR wild-type NSCLC cells.
CD44 is commonly considered to be a cancer stem cell
marker and it is found overexpressed in lung cancer tissues
especially in lung cancer stem cells. High expression of CD44 in
lung cancer tissue was significantly correlated with poor prognosis
of NSCLC patients (24). A
previous finding showed that the absence of CD44 in lung cancer
cells could result in suppression of cancer stem cell properties
including downregulation of tumor occurrence and reversing
multi-drug resistance (25).
Knockdown of CD44 might benefit numerous chemotherapeutic
treatments in wild-type EGFR NSCLC. The present results showed that
suppression of CD44 expression increased the sensitivity of H460 to
cisplatin. Additionally, cell cycle arrest and cell apoptosis were
induced by knockdown of CD44 and the effects were enhanced when in
combination with cisplatin. These results suggested EGFR wild-type
NSCLC patients with low CD44 expression might achieve a favorable
outcome from chemotherapeutic treatment.
Previous studies have indicated that expression of
CD44 and EGFR was significantly correlated in a variety of cancer
types; both were overexpressed in cancer tissues (14,24,26,27), and they can be considered as
prognostic factors for cancer patients. It was also reported that a
CD44 isoform could attenuate Rab7-induced EGFR degradation by
lysosomes in glioblastoma (14),
which provides a novel mechanism for CD44 activated the EGFR
signaling pathway. The current study further investigated the
related mechanism and found that EGF-mediated activation of EGFR
signaling was restrained by CD44 silencing. In addition, knockdown
of CD44 augments EGFR degradation in CHX in the treatment group,
indicating the mechanism underlying the relationship between CD44
and EGFR was consistent with previous findings (14). The modulation of EGFR expression
as well as its downstream signaling pathway by siCD44 was further
supported by the results of the IF assay and bioinformatics data
analysis. The clinical sample data from TCGA database indicated the
positive association between CD44 and EGFR mRNA level in EGFR
wild-type lung cancer tissue samples. CD44 might be involved in
EGFR degradation through interaction with wild-type EGFR in NSCLC
cells, thereby affecting its downstream AKT and ERK. Downregulation
of CD44 may benefit lung cancer patients undergoing chemotherapy
such as cisplatin treatment.
The current results demonstrated that CD44
inhibition enhanced cisplatin sensitivity in H460 cells, which
might be due to the downregulating of EGFR signaling activation.
Knockdown of CD44 decreased the EGFR protein level and
phosphorylation and affected the stimulation of the down-stream
signaling, thereby enhancing sensitivity of lung cancer cells to
cisplatin. The present results suggested that down-regulation of
CD44 could be considered as a useful strategy for cisplatin
treatment in wild-type EGFR NSCLC and the expression level of CD44
in NSCLC tissue might be viewed as an effective prognostic factor
for NSCLC patients.
Supplementary Data
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81803237 and 81602578) and
the Joint Funds of the Southwest Medical University & Luzhou,
China (grant no. 2018LZXNYD-ZK34).
Availability of data and materials
The analyzed data sets generated during the present
study are available from corresponding author on reasonable
request.
Authors' contributions
ML, XW and DY designed and conceived the study. JY,
YZhang, HZ and YW performed the experiments. JY, XW and YZhao
analyzed the data. JL, ZX, JS, YZhao, ZZ, FD, PJK, CHC, LL and CH
provided the resources and experiments for the study. XW wrote the
first draft. ML, HZ and XW reviewed and edited the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Noone AM, Howlader N, Krapcho M, Miller D,
Brest A, Yu M, Ruhl J, Tatalovich Z, Mariotto A, Lewis DR, et al:
SEER cancer statistics review, 1975-2015. National Cancer
Institute; Bethesda, MD: https://seer.cancer.gov/csr/1975_2015/,
based on November 2017, SEER data submission, posted to the SEER
web site. Accessed September 10, 2018.
|
|
2
|
Jiang W, Cai G, Hu PC and Wang Y:
Personalized medicine in non-small cell lung cancer: A review from
a pharmacogenomics perspective. Acta Pharm Sin B. 8:530–538. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tang JC, Ren YG, Zhao J, Long F, Chen JY
and Jiang Z: Shikonin enhances sensitization of gefitinib against
wild-type EGFR non-small cell lung cancer via inhibition
PKM2/stat3/cyclinD1 signal pathway. Life Sci. 204:71–77. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kobayashi T, Koizumi T, Agatsuma T, Yasuo
M, Tsushima K, Kubo K, Eda S, Kuraishi H, Koyama S, Hachiya T and
Ohura N: A phase II trial of erlotinib in patients with EGFR
wild-type advanced non-small-cell lung cancer. Cancer Chemother
Pharmacol. 69:1241–1246. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Su SF, Li M, Geng YC, Yang WG, Ma Z, Li
QS, Hu YX, Ou Yang WW, Liu LF and Lu B: Randomized phase II study
of pemetrexed-cisplatin or docetaxel-cisplatin plus thoracic
intensity-modulated radiation therapy in patients with stage IV
lung adenocarcinoma. Am J Cancer Res. 9:1235–1245. 2019.PubMed/NCBI
|
|
6
|
Toole BP and Slomiany MG: Hyaluronan: A
constitutive regulator of chemoresistance and malignancy in cancer
cells. Semin Cancer Biol. 18:244–250. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen C, Zhao S, Karnad A and Freeman JW:
The biology and role of CD44 in cancer progression: Therapeutic
implications. J Hematol Oncol. 11:642018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Park NR, Cha JH, Jang JW, Bae SH, Jang B,
Kim JH, Hur W, Choi JY and Yoon SK: Synergistic effects of CD44 and
TGF-β1 through AKT/GSK-3β/β-catenin signaling during
epithelial-mesenchymal transition in liver cancer cells. Biochem
Biophys Res Commun. 477:568–574. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu H and Wu K, Tian Y, Liu Q, Han N, Yuan
X, Zhang L, Wu GS and Wu K: CD44 correlates with
clinicopathological characteristics and is upregulated by EGFR in
breast cancer. Int J Oncol. 49:1343–1350. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Grass GD, Tolliver LB, Bratoeva M and
Toole BP: CD147, CD44, and the epidermal growth factor receptor
(EGFR) signaling pathway cooperate to regulate breast epithelial
cell invasiveness. J Biol Chem. 288:26089–26104. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wobus M, Rangwala R, Sheyn I, Hennigan R,
Coila B, Lower EE, Yassin RS and Sherman LS: CD44 associates with
EGFR and erbB2 in metastasizing mammary carcinoma cells. Appl
Immunohistochem Mol Morphol. 10:34–39. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim S, Kil WH, Lee J, Oh SJ, Han J, Jeon
M, Jung T, Lee SK, Bae SY, Lee HC, et al: Zerumbone suppresses
EGF-induced CD44 expression through the inhibition of STAT3 in
breast cancer cells. Oncol Rep. 32:2666–2672. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Perez A, Neskey DM, Wen J, Pereira L,
Reategui EP, Goodwin WJ, Carraway KL and Franzmann EJ: CD44
interacts with EGFR and promotes head and neck squamous cell
carcinoma initiation and progression. Oral Oncol. 49:306–313. 2013.
View Article : Google Scholar :
|
|
14
|
Wang W, Zhang H, Liu S, Kim CK, Xu Y,
Hurley LA, Nishikawa R, Nagane M, Hu B, Stegh AH, et al:
Internalized CD44s splice isoform attenuates EGFR degradation by
targeting Rab7A. Proc Natl Acad Sci USA. 114:8366–8371. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Morath I, Jung C, Lévêque R, Linfeng C,
Toillon RA, Warth A and Orian-Rousseau V: Differential recruitment
of CD44 isoforms by ErbB ligands reveals an involvement of CD44 in
breast cancer. Oncogene. 37:1472–1484. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Meran S, Luo DD, Simpson R, Martin J,
Wells A, Steadman R and Phillips AO: Hyaluronan facilitates
transforming growth factor-β1-dependent proliferation via CD44 and
epidermal growth factor receptor interaction. J Biol Chem.
286:17618–17630. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
18
|
Chohan TA, Qayyum A, Rehman K, Tariq M and
Akash MSH: An insight into the emerging role of cyclin-dependent
kinase inhibitors as potential therapeutic agents for the treatment
of advanced cancers. Biomed Pharmacother. 107:1326–1341. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Asghar U, Witkiewicz AK, Turner NC and
Knudsen ES: The history and future of targeting cyclin-dependent
kinases in cancer therapy. Nat Rev Drug Discov. 14:130–146. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu SG and Shih JY: Management of acquired
resistance to EGFR TKI-targeted therapy in advanced non-small cell
lung cancer. Mol Cancer. 17:382018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park K, Vansteenkiste J, Lee KH,
Pentheroudakis G, Zhou C, Prabhash K, Seto T, Voon PJ, Tan DSW,
Yang JCH, et al: Pan-Asian adapted ESMO clinical practice
guidelines for the management of patients with locally-advanced
unresectable non-small-cell lung cancer: A KSMO-ESMO initiative
endorsed by CSCO, ISMPO, JSMO, MOS, SSO and TOS. Ann Oncol.
31:191–201. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Su C, Zhou F, Shen J, Zhao J and O'Brien
M: Treatment of elderly patients or patients who are performance
status 2 (PS2) with advanced non-small cell lung cancer without
epidermal growth factor receptor (EGFR) mutations and anaplastic
lymphoma kinase (ALK) translocations-Still a daily challenge. Eur J
Cancer. 83:266–278. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang TH, Wu TH, Guo YH, Li TL, Chan YL
and Wu CJ: The concurrent treatment of Scutellaria baicalensis
Georgi enhances the therapeutic efficacy of cisplatin but also
attenuates chemotherapy-induced cachexia and acute kidney injury. J
Ethnopharmacol. 243:1120752019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Roudi R, Madjd Z, Korourian A, Mehrazma M,
Molanae S, Sabet MN and Shariftabrizi A: Clinical significance of
putative cancer stem cell marker CD44 in different histological
subtypes of lung cancer. Cancer Biomark. 14:457–467. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Quan YH, Lim JY, Choi BH, Choi Y, Choi YH,
Park JH and Kim HK: Self-targeted knockdown of CD44 improves
cisplatin sensitivity of chemoresistant non-small cell lung cancer
cells. Cancer Chemother Pharmacol. 83:399–410. 2019. View Article : Google Scholar
|
|
26
|
Zheng Z, Shao N, Weng H, Li W, Zhang J,
Zhang L, Yang L and Ye S: Correlation between epidermal growth
factor receptor and tumor stem cell markers CD44/CD24 and their
relationship with prognosis in breast invasive ductal carcinoma.
Med Oncol. 32:2752015. View Article : Google Scholar
|
|
27
|
Rho JH, Ladd JJ, Li CI, Potter JD, Zhang
Y, Shelley D, Shibata D, Coppola D, Yamada H, Toyoda H, et al:
Protein and glycomic plasma markers for early detection of adenoma
and colon cancer. Gut. 67:473–484. 2018. View Article : Google Scholar
|