Introduction
The use of hyperthermia (HT) has been gradually
increasing over the past three decades. In general, temperatures
ranging from 40 to 45°C are used in locoregional treatment, whereas
temperatures of up to 42°C are used in cases requiring whole-body
HT. HT combined with chemotherapy, radiotherapy, or both, is
considered as a promising approach to cancer therapy (1-10).
However, the thermotolerance due to the upregulation of heat shock
proteins (HSPs) in some cancer cells is a limiting factor that
reduces the efficacy of HT in the clinical setting (11-15).
HSPs are highly conserved proteins occurring in
almost all organisms and they are classified by their molecular
weight; these include heat shock 27 kDa protein (HSPB), DnaJ
(Hsp40) homolog (DNAJ), heat shock 60 kDa protein (HSPD), heat
shock 70 kDa protein (HSPA), heat shock protein 90 (HSP90) and heat
shock 105/110 kDa protein (HSPH) in humans. HSPs act as molecular
chaperones, and their expression is induced by various stressors,
including heat (16-20). The induction of HSPs principally
occurs through the activation of heat shock transcription factor 1
(HSF1), which binds to conserved regulatory sequences, referred to
as heat shock elements, which are located in the promoter regions
of inducible HSP genes (21,22). It has been well established that
HSPs, particularly Hsp70, exert a cytoprotective effect against
stressors and play a key role in the development of thermotolerance
(19,23). Several studies demonstrated that
abrogation of HSF1 (24,25), Hsp70 (26) or Hsp105 (27) expression increased the sensitivity
of human cancer cells to heat stress. In response to heat stress,
Hsp70 rapidly translocates from the cytoplasm into the nucleus
(28,29). Kose et al (30) reported for the first time that the
nuclear import of Hsp70 is mediated by the heat shock protein
nuclear import factor hikeshi (HIKESHI), also referred to as
C11orf73, under conditions of heat-induced stress. Although
silencing of HIKESHI had no discernible effect under normal
conditions, it was found to significantly inhibit the nuclear
translocation of Hsp70 or to reduce cell viability after exposure
of cancer cells to heat stress (30-32). In human gastric cancer tissues,
HIKESHI expression was reported to be associated with the
progression of lymphatic invasion (32). It has also been demonstrated that
HIKESHI is abundantly expressed in human clear cell renal cancer
(33).
In our previous studies, we used human oral squamous
cell carcinoma (OSCC) HSC-3 cells as a model for evaluation of HT
sensitivity (25,34-36). The aim of the present study was to
evaluate the effects of HIKESHI knockdown (KD) on the sensitivity
of human OSCC HSC-3 cells to mild HT (MHT).
Materials and methods
Cell culture
Human HSC-3 OSCC cells (JCRB0623) were obtained from
the Human Science Research Resources Bank, Japan Health Sciences
Foundation (Tokyo, Japan). HSC-3 cells were cultured in Eagle's
minimum essential medium (E-MEM; Wako Pure Chemical Industries,
Ltd.) supplemented with 10% fetal bovine serum (FBS; Equitech-Bio,
Inc.) at 37°C in a humidified atmosphere with 5% CO2 and
95% air. U0126 (Cell Signaling Technology, Inc.), an inhibitor of
mitogen-activated protein kinase (MAPK)/extracellular
signal-regulated kinase (ERK), was dissolved in dimethyl sulfoxide
and added to the culture medium 1 h before MHT treatment (final
concentration of U0126: 10 µM).
MHT treatment
MHT treatments were performed by immersing plastic
culture dishes sealed with laboratory film (Parafilm® M;
Nippon Genetics, Co., Ltd.) in a water bath at 42°C for 60 or 90
min. After MHT treatment, the cells were recovered at 37°C for the
indicated periods.
Small interfering RNA (siRNA)
transfection
Based on the human HIKESHI nucleotide database
(GenBank accession no. NM_016401), two siRNAs for HIKESHI,
designated as siHIK-1 and siHIK-2, were synthesized by Nippon Gene
Co., Ltd. Firefly luciferase siRNA (siLuc) was used as a negative
control siRNA. The sequences of the siRNAs are listed in Table I. Cells were incubated in
Opti-MEM® I Reduced Serum Medium (Life Technologies
Japan; Thermo Fisher Scientific, Inc.) containing 20 nM siRNA and
Lipofectamine™ RNAiMAX (Life Technologies Japan; Thermo Fisher
Scientific, Inc.) at 37°C. At 6 h after transfection, the medium
was exchanged for E-MEM supplemented with 10% FBS, and then the
cells were maintained at 37°C for 2 days (36).
 | Table INucleotide sequences of siRNAs for
HIKESHI and luciferase. |
Table I
Nucleotide sequences of siRNAs for
HIKESHI and luciferase.
| Name | Sequence-TT | Position | GenBank accession
nos. |
|---|
| siHIK-1 |
AUUACCUACAGGAGUCUGC | 607 | NM_016401 |
| siHIK-2 |
AAGAAAAGUAGACAGAUCC | 392 | NM_016401 |
| siLuc |
CGUACGCGGAAUACUUCGA | 282 | MH759210 |
Cellular fractionation
Nuclear and cytoplasmic fractions were prepared as
described previously (37). In
brief, the cells were lysed in the fractionation buffer
[phosphate-buffered saline containing 0.1% Nonidet P-40 and
protease inhibitor cocktail (Nacalai Tesque, Inc.)] and centrifuged
at 15,000 × g for 10 sec at 4°C to obtain the cytosolic fraction
(supernatant). The insoluble pellets were resuspended in the
fractionation buffer and centrifuged at 15,000 × g for 10 sec at
4°C to obtain the nuclear fraction (pellets). Either GAPDH
(38) or fibrillarin (FBL)
(39) was used as the cytoplasmic
or nuclear marker protein, respectively.
Analysis of cell viability
A trypan blue dye exclusion test was performed to
assess cell viability. The number of cells excluding the dye was
counted by using a hematocytometer (Burker-Turk; ERMA Inc.). Cell
Count Reagent SF (Nacalai Tesque, Inc.), a water-soluble
tetrazolium salt (WST-8)-based assay, was also used to test the
cell viability. Cells were incubated with the WST-8 solution at
37°C. After 30 min, the concentration of formazan dye was
determined from the absorbance at 450 nm.
SDS-PAGE and western blotting
Cells were lysed with lysis buffer (50 mM NaCl, 1%
Nonidet P-40 and 50 mM Tris-HCl, pH 8.0) containing protease
inhibitor cocktail (Nacalai Tesque, Inc.) and treated at 94°C for 3
min. Proteins (10 µg/lane) were separated by 10 or 12.5%
SDS-PAGE and transferred onto a PVDF membrane. Membranes were
blocked by PVDF Blocking Reagent (Toyobo Co., Ltd.) for 3 h at 25°C
(40,41). The protein concentration was
determined by a standard BCA protein assay. Proteins were detected
using the following primary antibodies: Mouse monoclonal anti-Hsp70
antibody (1:2,000 dilution, cat. no. SR-B810; Medical &
Biological Laboratories Co., Ltd.), rabbit polyclonal anti-HIKESHI
antibody (1:1,000 dilution, cat. no. 14808-1-AP; ProteinTech Group,
Inc.), rabbit polyclonal anti-HSF1 antibody (1:2,000 dilution, cat.
no. 4356; Cell Signaling Technology, Inc.), mouse monoclonal
anti-total ERK1/2 antibody (1:3,000 dilution, cat. no. 9107, Cell
Signaling Technology, Inc.), anti-phospho-ERK1/2 (Thr202/Tyr204)
(pERK1/2) rabbit monoclonal antibody (1:3,000 dilution, cat. no.
4370, Cell Signaling Technology, Inc.), rabbit monoclonal anti-FBL
antibody (1:2,000 dilution, cat. no. 2639; Cell Signaling
Technology, Inc.) and mouse monoclonal anti-GAPDH antibody (as a
loading reference; 1:2,000 dilution, cat. no. MAB347; EMD
Millipore). Secondary fluorescent IRDye-conjugated anti-rabbit and
anti-mouse antibodies (1:10,000 dilution, LI-COR Biosciences) were
also used. Fluorescence images were acquired using an Odyssey
Infrared Imager (LI-COR Biosciences), and the band density was
quantified using Image Studio 5.1 software (LI-COR
Biosciences).
Immunocytochemistry
Cells were grown in a collagen type I-precoated
glass coverslip (AGC Techno Glass Co., Ltd.). The cells were fixed
in 4% paraformaldehyde phosphate-buffered solution (Nacalai Tesque,
Inc.) for 15 min at 25°C. The cells were incubated with a mouse
monoclonal anti-Hsp70 antibody (1:200 dilution, cat. no. SR-B810;
Medical & Biological Laboratories Co., Ltd.) for 18 h at 4°C,
followed by Chromeo™ 488-labeled secondary antibody (1:500
dilution, cat. no. 15031; Active Motif) for 1 h at 25°C. The
nucleus was stained for 5 min at 25°C with DAPI. Immunofluorescence
images were visualized by using a fluorescence microscope at a
magnification of ×20 (BX-50; Olympus Corporation). Fluorescence
intensity was measured using softWoRx Explorer software, version
1.3 (Applied Precision, Inc.).
Reverse transcription-quantitative PCR
(RT-qPCR) assay
Total RNA was extracted from cells using a
NucleoSpin® RNA isolation kit (Takara Bio, Inc.). RNA
quality was analyzed using a Bioanalyzer 2100 (Agilent
Technologies, Inc.). Complementary DNA (cDNA) was produced from the
reverse transcription of total RNA using a PrimeScript RT kit
(Takara Bio Inc.) with random 6-mers and an oligo dT primer. qPCR
was performed on a Mx3005P real-time PCR system (Agilent
Technologies, Inc.) using a SYBR® Premix Ex Taq™ II kit
(Takara Bio, Inc.). The thermocycling conditions for each primer
consisted of 10 min at 95°C followed by 40 cycles of 10 sec at 95°C
and 40 sec at 60°C. The specific primers are listed in Table II. GAPDH was used for
normalization (40).
| Table IINucleotide sequences of primers for
target genes. |
Table II
Nucleotide sequences of primers for
target genes.
| Genes | Orientation | Nucleotide sequence
(5'-3') | GenBank accession
nos. |
|---|
| DNAJB1 | Sense |
ACCCGGACAAGAACAAGGAG | NM_006145 |
| Antisense |
GCCACCGAAGAACTCAGCAA | |
| GAPDH | Sense |
AAGGCTGGGGCTCATTTGCA | NM_002046 |
| Antisense |
ATGACCTTGCCCACAGCCTT | |
| HIKESHI | Sense |
AGGGAATGGGAGGATCTGTC | NM_016401 |
| Antisense |
GATGTTGGCTTCCTTCTCCA | |
| HSPA1 | Sense |
AGGTGCAGGTGAGCTACAAG | NM_005346 |
| Antisense |
ATGATCCGCAGCACGTTGAG | |
Statistical analysis
Data are shown as means ± standard deviation.
Differences between groups were analyzed by ANOVA, and correction
for multiple comparisons was made using Tukey's post hoc test.
Comparisons between two groups were made by using Student's t-test.
P<0.05 was considered to indicate statistically significant
differences.
Results
Effects of HIKESHI-KD on the viability in
HSC-3 cells
To evaluate the ability of siHIK-1 and siHIK-2 to
inhibit HIKESHI expression, RT-qPCR and western blot analyses were
carried out. The treatment of HSC-3 cells with siLuc did not alter
the mRNA and protein expression of HIKESHI (Fig. S1). Therefore, siLuc treatment was
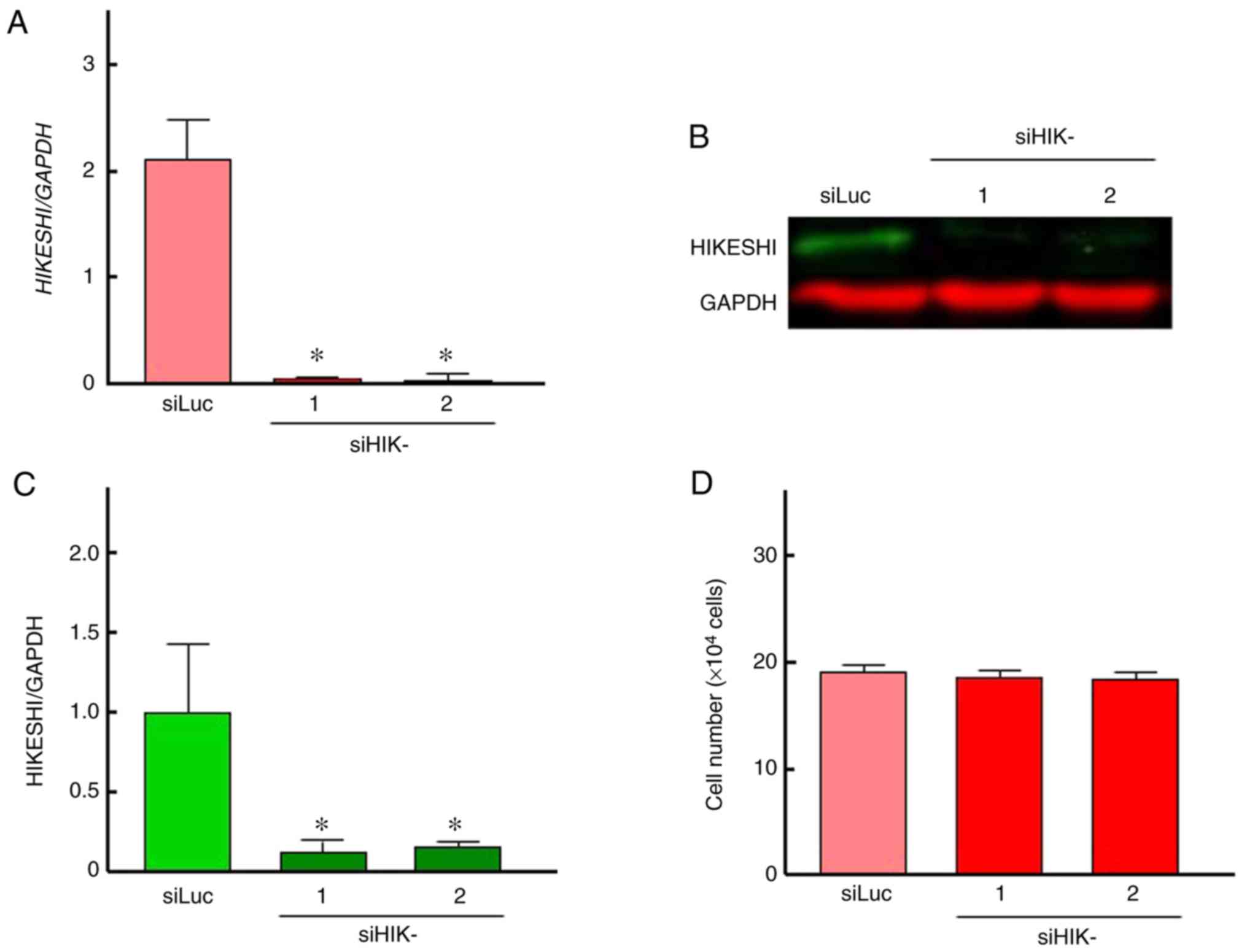
used as control in further experiments. As shown in Fig. 1A, the two siRNAs for HIKESHI
markedly reduced the mRNA expression level of HIKESHI in HSC-3
cells. This silencing was confirmed by western blotting, whereas
the inhibition percentage was ~85% (Fig. 1B and C). There was no difference
in the silencing efficiency between the two siHIKs. Next, the
effects of HIKESHI-KD on the cell growth under normal conditions
were monitored. Treatment of the cells with the siRNAs for HIKESHI
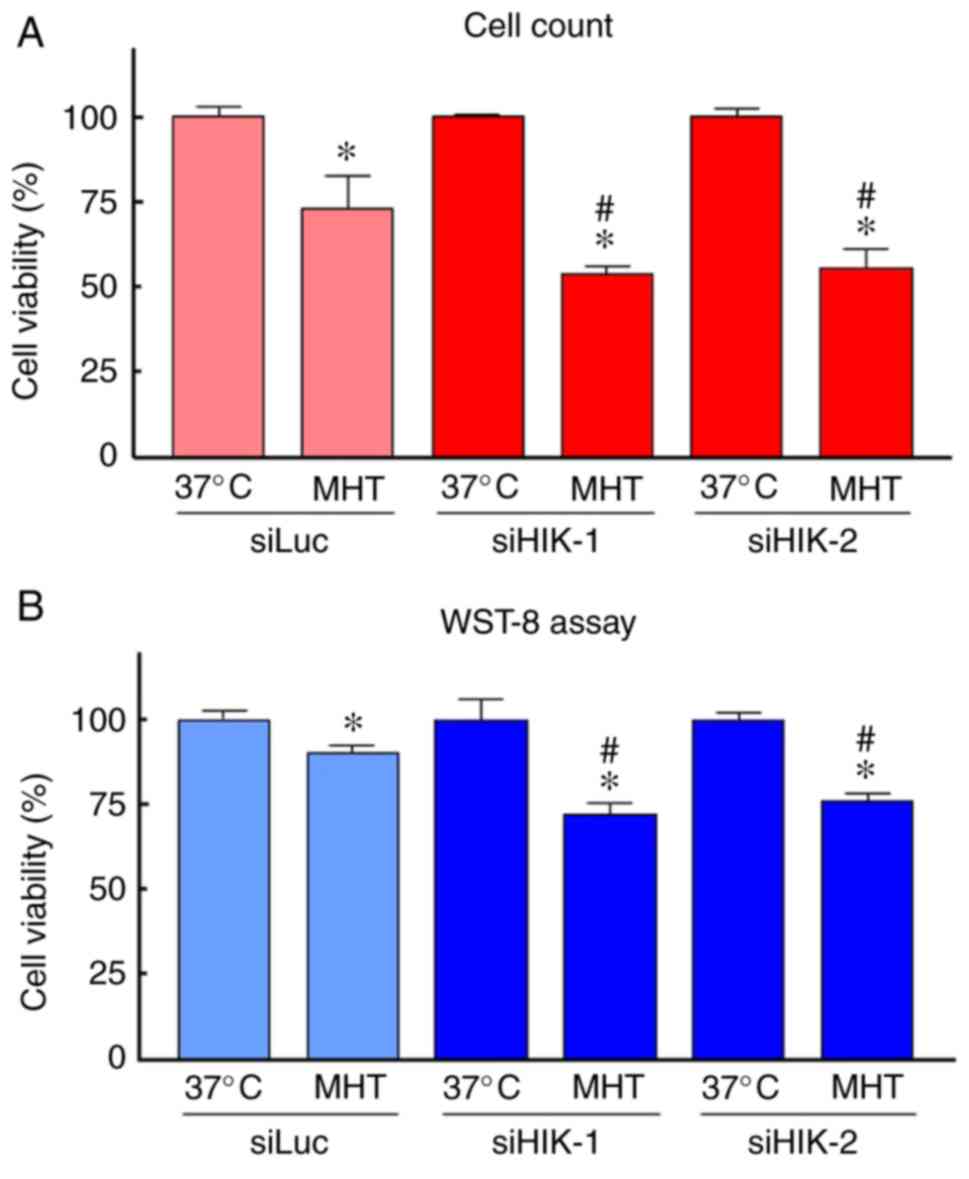
did not change the cell number at 37°C (Fig. 1D). With respect to MHT, the
viability of cells exposed to MHT (42°C for 90 min) alone was
slightly decreased compared with that of non-treated cells; the
mean values were 73 and 92% by trypan blue dye exclusion test and
WST-8 assay, respectively. HIKESHI-KD prior to MHT further
decreased the number of viable cells, with mean values of ~55 and
75% by trypan blue dye exclusion test and WST-8 assay, respectively
(Fig. 2).
Effects of HIKESHI-KD on the localization
of Hsp70 in HSC-3 cells
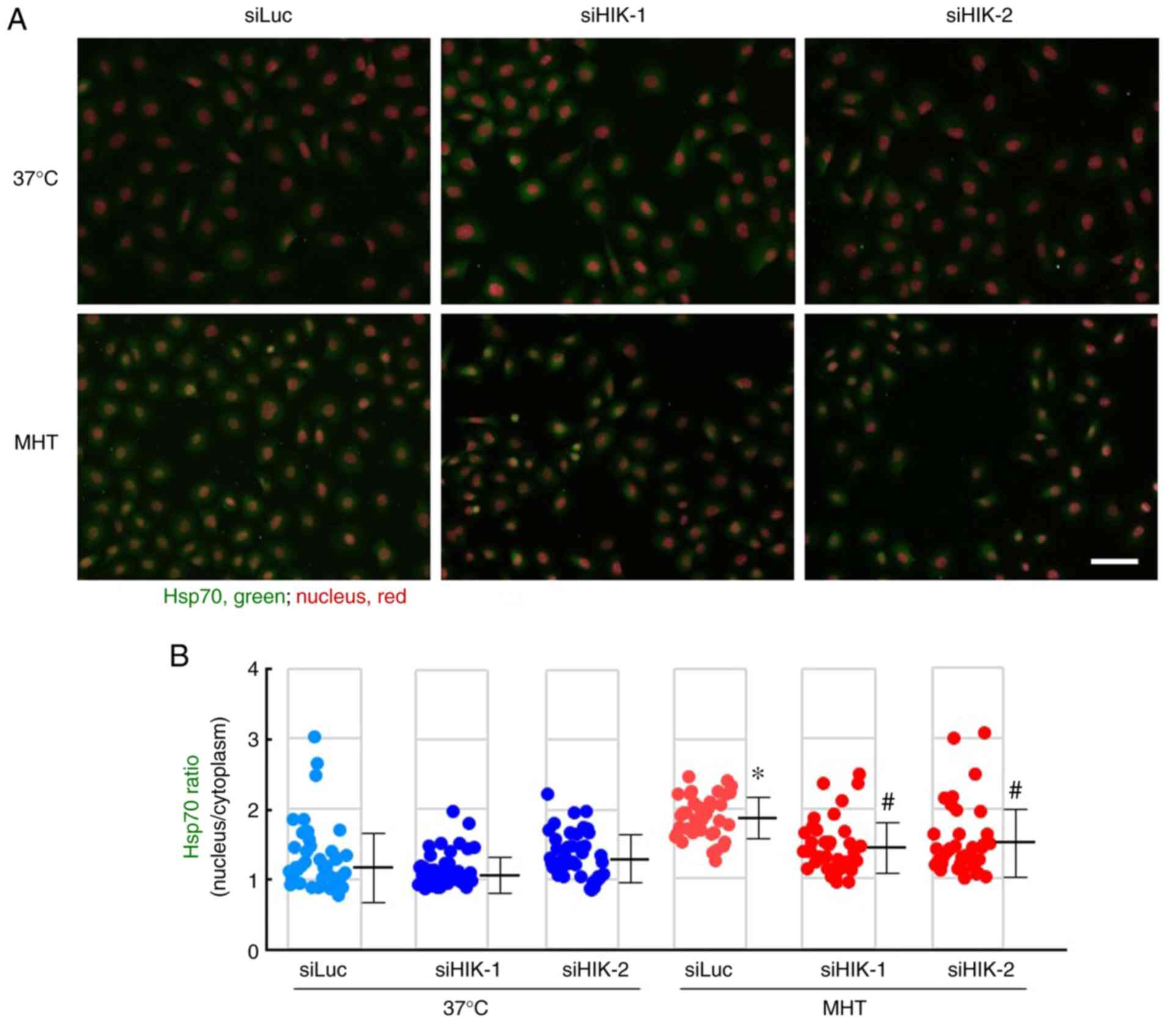
As demonstrated in Fig. 3A, Hsp70 was principally localized
in the cytosolic compartment, and neither siHIK-1 nor siHIK-2
affected the intracellular localization of this protein under
non-MHT conditions. Treatment of cells with MHT at 42°C for 60 min
markedly induced the nuclear localization of Hsp70, as reported
previously (28,29). Furthermore, the MHT-induced
nuclear localization was significantly inhibited in HIKESHI-KD
cells (Fig. 3A and B). Next,
cytoplasmic and nucleic fractions were prepared from the cells, and
the protein levels were verified by western blot analysis. As
expected, MHT at 42°C for 60 min induced the nuclear translocation
of Hsp70, and this induction of Hsp70 translocation by MHT was
significantly decreased in HIKESHI-KD cells (Fig. 4A and B).
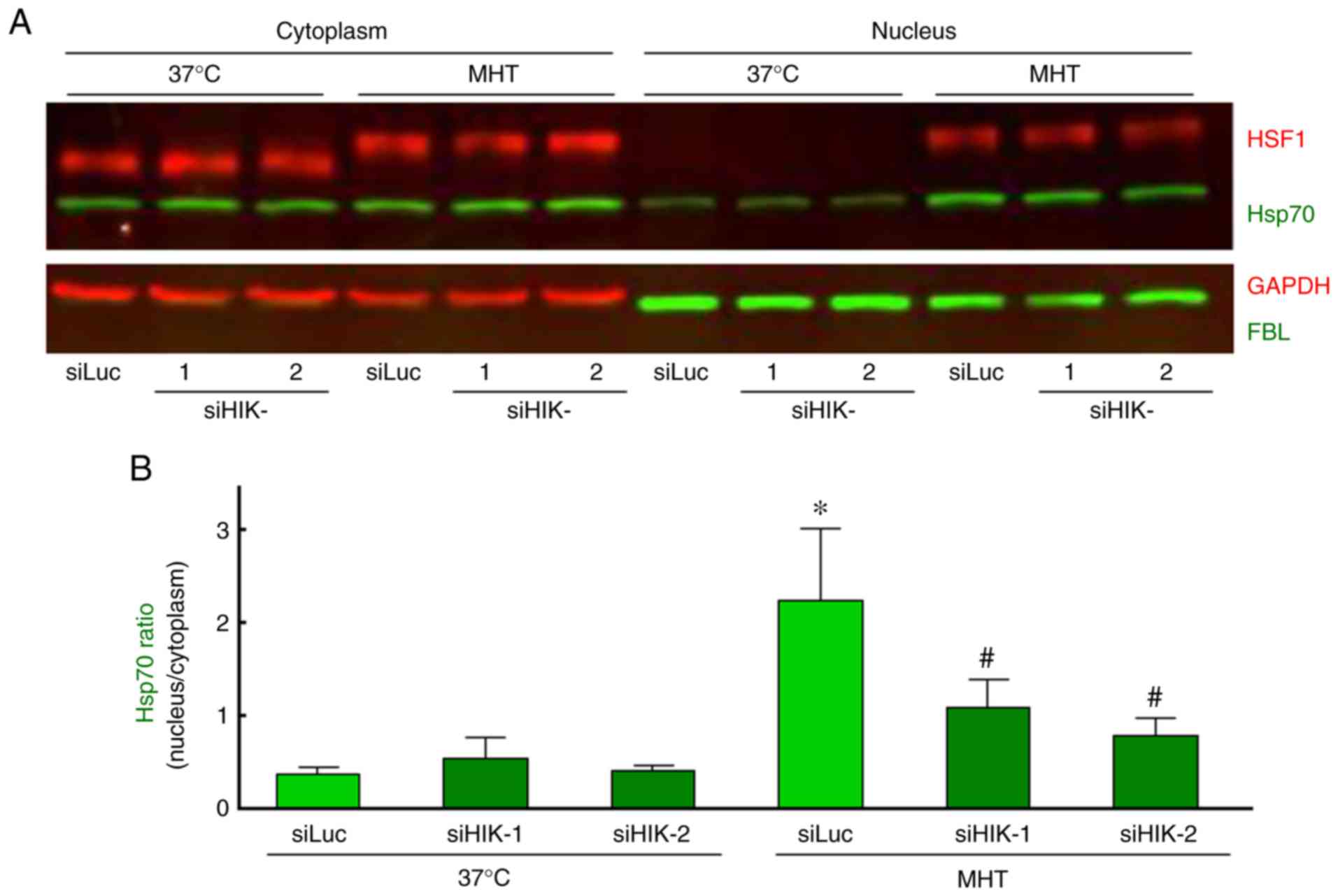 | Figure 4Western blot analysis of the
intracellular localization of Hsp70 and HSF1 in HIKESHI-knockdown
cells under mild hyperthermia (MHT) conditions. HIKESHI-knockdown
HSC-3 cells were incubated at 42°C for 60 min. Immediately after
heat exposure, the cells were harvested and either the cytoplasmic
or nuclear fraction was separated. (A) Western blotting was carried
out using specific primary antibodies against Hsp70, HSF1, GAPDH
and FBL. GAPDH and FBL served as marker proteins for the cytoplasm
and nucleus, respectively. (B) Each band density of Hsp70 was
quantified, and the ratio (nucleus to cytoplasm) was calculated.
Data are presented as means ± standard deviation (n=3).
*P<0.05 vs. the siLuc-treated group at 37°C;
#P<0.05 vs. the siLuc-treated group at 42°C. siLuc,
siRNA for luciferase; siHIKE, siRNA for HIKESHI; HIKESHI, heat
shock protein nuclear import factor hikeshi; Hsp70, heat shock
protein 70 kDa; HSF, heat shock transcription factor; FBL,
fibrillarin. |
Effects of HIKESHI-KD on HSF1 activation
in HSC-3 cells
It is well known that the mobility shift and nuclear
translocation of HSF1 due to its phosphorylation indicates
activation of the molecule (42).
In our experiments, either a mobility shift or nuclear
translocation of HSF1 was observed immediately after MHT (42°C for
60 min), whereas silencing of HIKESHI did not affect the activation
of HSF1 under MHT conditions (Fig.
4A). Moreover, the effects of HIKESHI-KD on the gene expression
of HIKESHI, HSPA1 and DNAJB1 were evaluated
using RT-qPCR. The expression level of HIKESHI was slightly
but significantly increased 3 h after MHT, to a level 1.3-fold
higher compared with that of non-treated cells. As expected, the
expression of HIKESHI was almost completely eradicated in
HIKESHI-KD cells under MHT conditions (Fig. 5A). The expression levels of
HSPA1 and DNAJB1 were markedly increased in a
time-dependent-manner, by 66- and 40-fold, respectively, compared
with the levels in non-treated cells. However, the expressions of
these genes were not affected by HIKESHI-KD (Fig. 5B and C).
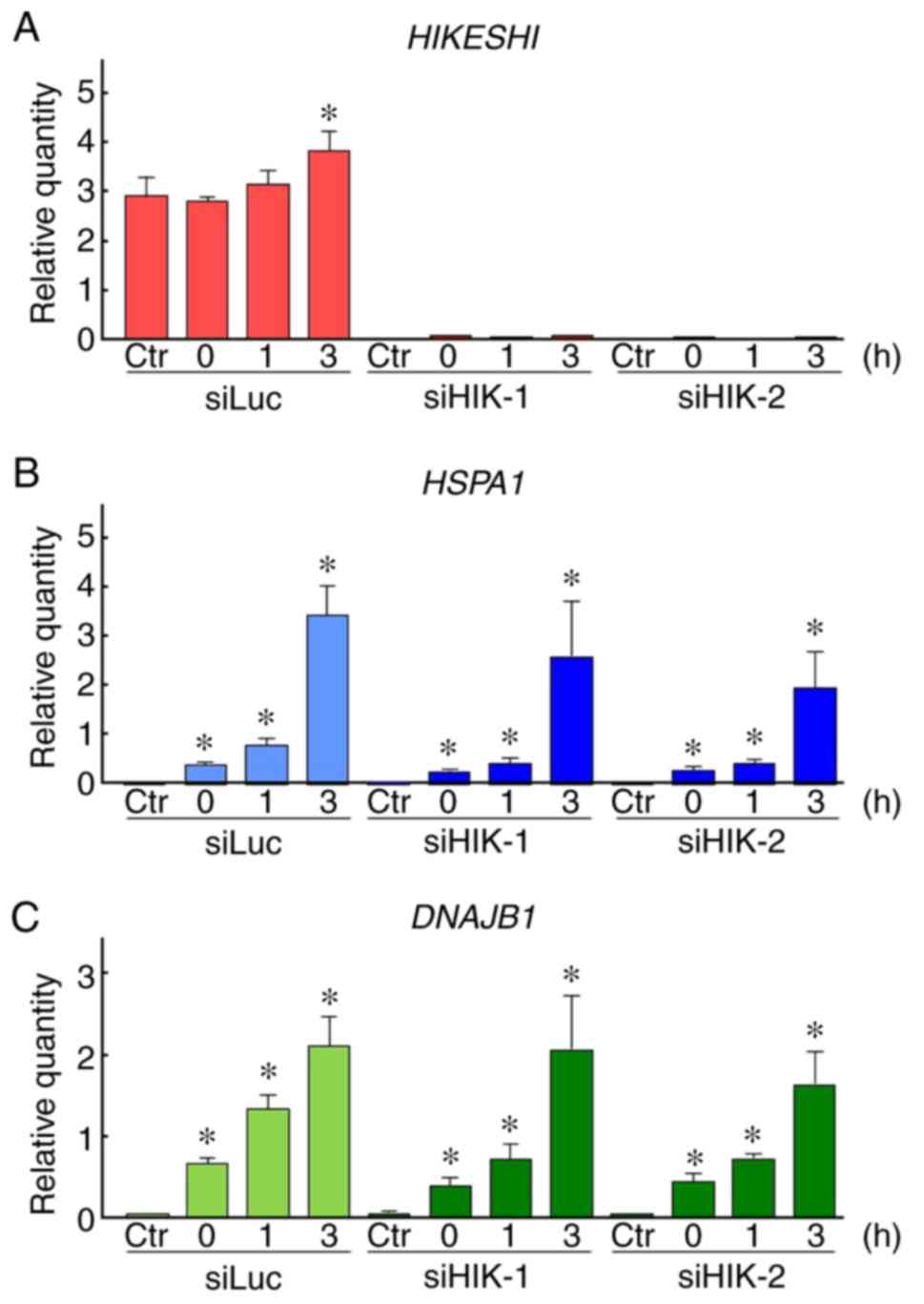 | Figure 5Effects of HIKESHI knockdown on the
gene expression in mild hyperthermia (MHT)-treated HSC-3 cells.
After treatment of HIKESHI-knockdown HSC-3 cells with mild
hyperthermia at 42°C for 90 min, the cells were cultured for 0, 1
or 3 h at 37°C. quantitative PCR was carried out with specific
primers for (A) HIKESHI, (B) HSPA1, (C) DNAJB1
and GAPDH. The expression level was normalized to that of
GAPDH. siLuc, Data are presented as means ± standard
deviation (n=4). Non-MHT-treated cells served as the control (Ctr).
*P<0.05 vs. each Ctr. siRNA for luciferase; siHIKE,
siRNA for HIKESHI; HIKESHI, heat shock protein nuclear import
factor hikeshi; HSPA1, heat shock 70 kDa protein 1; DNAJB1, DnaJ
(Hsp40) homolog, subfamily B, member 1. |
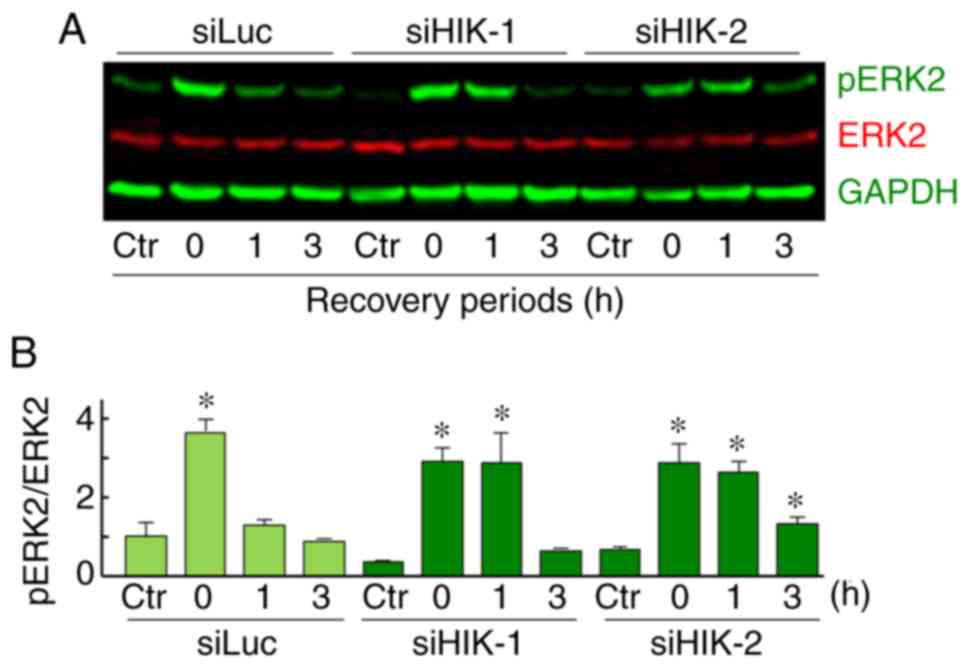
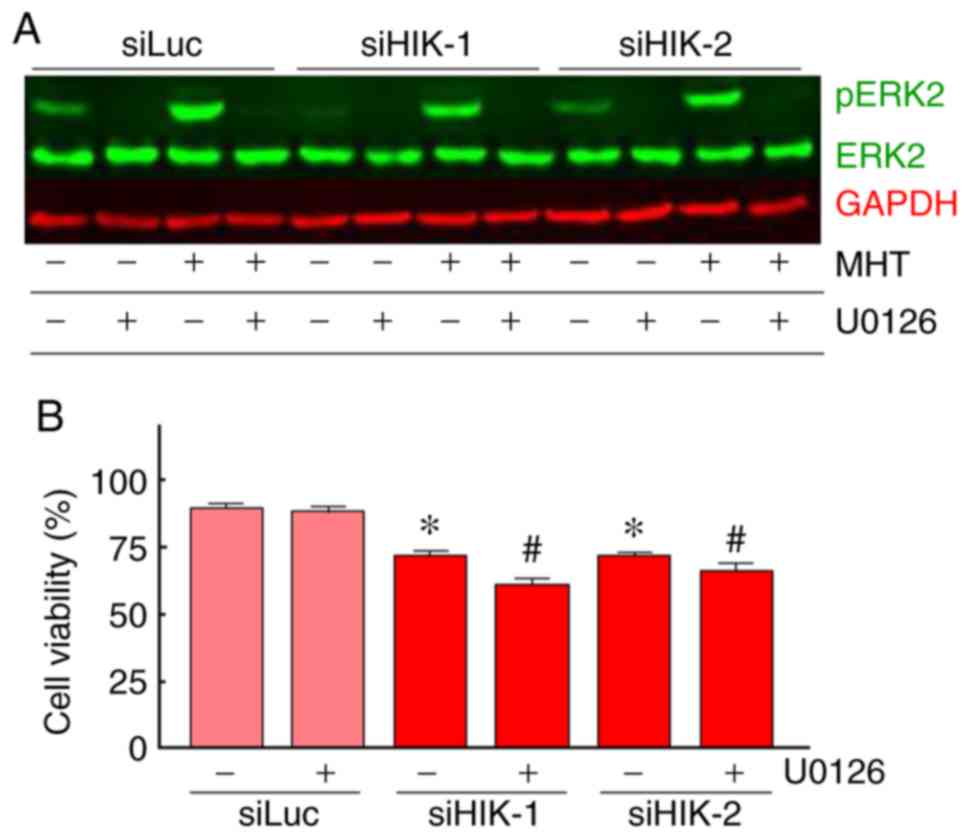
Effects of HIKESHI-KD on ERK2 activation
in HSC-3 cells and the role of the MAPK/ERK pathway in the
enhancement of MHT sensitivity by HIKESHI-KD
The expression levels of total and pERK1/2 proteins
were assessed using western blot analysis. In HSC-3 cells, a
relatively high expression level of total ERK2 (42 kDa) was
observed, while total ERK1 (44 kDa) was hardly detected. ERK2
expression levels were almost constant under all the treatments
tested. A significant and transient induction of pERK2 was observed
immediately after MHT treatment (0 h). In addition, an elevated
level of pERK2 was sustained for 0-1 or 0-3 h after treatment in
cells treated with siHIK-1 or siHIK-2, respectively (Fig. 6A and B). This increase in the
level of pERK2 was completely abolished by pretreatment with U0126,
a MAPK/ERK inhibitor, under both normal and MHT conditions
(Fig. 7A). Next, the effects of
U0126 on the viability of HIKESHI-KD cells were investigated and
the results are shown in Fig. 7B.
In control siLuc-treated cells, U0126 did not affect the cell
viability under MHT conditions. By contrast, the inhibitor
significantly decreased the viability of cells treated with the
combination of HIKESHI-KD and MHT.
Discussion
The present study investigated whether silencing of
HIKESHI by siRNA can sensitize human OSCC HSC-3 cells to MHT. The
results demonstrated that the downregulation of HIKESHI enhanced
MHT sensitivity, and inhibition of the MAPK/ERK pathway further
potentiated the synergistic effects of MHT and HIKESHI-KD.
HT and the combination of HT with radiotherapy,
chemotherapy, or both, have been recognized as effective treatments
for malignant tumors (1-10). However, the acquisition of
thermotolerance constitutes a limitation of HT therapy (11-15). Although the detailed mechanisms
are not well known, the induction of HSPs, particularly Hsp70,
plays a central role in the acquisition of thermotolerance
(19,23). Therefore, Hsp70 has been
considered as a valuable target in HT therapy (23). Furthermore, previous findings
suggest the utilization of Hsp70 tumor antigens for cancer
immunotherapy based on HT (43,44). When the cells are exposed to heat
stress, Hsp70 rapidly trans-locates from the cytoplasm into the
nucleus (28,29). Recently, HIKESHI was reported to
be a nuclear import carrier of Hsp70 under heat stress conditions
(30). In addition, previous
reports demonstrated that HIKESHI expression was induced by HT in
cancer cells (30,32). These findings prompted us to
investigate a unique strategy, namely MHT in combination with
targeting of HIKESHI, which prevents only the nuclear translocation
of Hsp70 under heat stress conditions. In the present study, it was
confirmed that HIKESHI was induced at the mRNA level under
MHT conditions. However, its induction ratio was markedly lower
compared with those of HSPA1 and DNAJB1, the
expression levels of which are principally regulated by HSF1
(21,22). These results indicated that heat
may promote the expression of HIKESHI via HSF1-independent
transcriptional mechanisms.
In line with previous reports (30-32), our experiments demonstrated that
HIKESHI-KD did not affect the number of viable cells under normal
conditions at 37°C, suggesting that HIKESHI may not be required for
the normal growth of OSCC HSC-3 cells. Interestingly, HIKESHI
silencing significantly enhanced MHT sensitivity of HSC-3 cells, as
demonstrated by the cell viability assay. These results were
comparable to those of previous studies (30-32). Immunocytochemical analysis clearly
demonstrated that HIKESHI-KD effectively prevented the nuclear
translocation of Hsp70. This was confirmed by western blotting with
the cellular fractionation assay. Of note, neither HSF1 activation
nor HSP expression were affected by downregulation of HIKESHI under
MHT conditions. Thus, disruption of the nuclear translocation of
Hsp70 may play a major role in the enhancement of thermosensitivity
by HIKESHI-KD. However, over half of HSC-3 cells were viable even
after combined treatment with MHT (42°C for 90 min) and siRNA for
HIKESHI. We previously reported that, under comparable experimental
conditions, HSF1 silencing markedly enhanced MHT sensitivity, with
damage occurring in ~75% of HSC-3 cells treated with MHT (42°C for
90 min) and siRNA for HSF1 (25).
Under these HSF1-silencing conditions, the expressions of
HSF1-regulated proteins, such as Hsp70, Hsp40 and Hsp27, were
maintained at a low level. It appears likely that the potential of
HIKESHI-KD for enhancement of MHT is weaker compared with that of
HSF1-KD.
The ERK1/2 cascade is a central signaling pathway
activated by a wide variety of stressors, including heat (45-47). In the present study, although a
significant increase in pERK2 was observed in HSC-3 cells treated
with MHT, cells pretreated with U0126, an inhibitor of MAPK/ERK,
exhibited little change in viability, which was consistent with the
findings of previous studies using heat-treated cancer cells
(46,47). Chen et al (46) reported that pretreatment of human
HT-29 colon cancer cells with U0126 resulted in enhancement of HT
(43°C for 60 min) sensitivity in cells treated with a combination
of HT and MG132, a proteasome inhibitor, suggesting that activated
ERK is a prosurvival mechanism under conditions of combination of
HT with proteasome inhibition. By contrast, ERK activity was
reported to be a proapoptotic mechanism in human Y79
retino-blastoma cells treated with HT (44°C for 60 min) combined
with silencing of BAG cochaperone 3, a cochaperone for Hsp70
(47). In the present study,
inhibition of the sustained activation of ERK by U0126 induced
enhancement of MHT sensitivity in HSC-3 cells treated with a
combination of MHT and siRNA for HIKESHI. These data suggest that
the addition of HIKESHI silencing to MHT triggers ERK activation as
a prosurvival mechanism, as reported previously (46). However, the molecular mechanism
through which HIKESHI silencing induces the sustained activation of
ERK under MHT conditions is unclear. Moreover, it has been
demonstrated that nucleocytoplasmic transport is affected by the
MAPK/ERK pathway under stress conditions (48,49). At present, few details are known
on the role of the MAPK/ERK pathway in the enhancement of MHT
sensitivity by HIKESHI silencing.
In conclusion, the findings of the present study
clearly demonstrated that downregulation of HIKESHI enhances the
sensitivity to MHT in human OSCC HSC-3 cells; therefore, this
molecule may be considered as a potential target in HT therapy of
cancer. However, further studies are needed to investigate the
detailed mechanisms underlying the effects of HIKESHI in HT therapy
of cancer in animals and humans.
Supplementary Data
Funding
The present study was supported in part by JSPS
KAKENHI (grant nos. JP26560205 and JP17K01353).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YT designed all the experiments and wrote the
manuscript. YT, KM, MT and SM performed the experiments. YT, KM and
YF analyzed the data. All authors (YT, KM, MT, YF, TH, SM, TY and
AH) have discussed the data and commented on the manuscript. All
the authors have read and approved the final version of this
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Abbreviations:
|
DNAJ
|
DnaJ (Hsp40) homolog
|
|
FBL
|
fibrillarin
|
|
FBS
|
fetal bovine serum
|
|
GAPDH
|
glyceraldehyde 3-phosphate
dehydrogenase
|
|
HIKESHI
|
heat shock protein nuclear import
factor hikeshi
|
|
HSF1
|
heat shock transcription factor 1
|
|
HSP90
|
heat shock protein 90
|
|
HSPA
|
heat shock 70 kDa protein
|
|
HSPB
|
heat shock 27 kDa protein
|
|
HSPD
|
heat shock 60 kDa protein
|
|
HSPH
|
heat shock 105kDa/110kDa protein
|
|
HSPs
|
heat shock proteins
|
|
HT
|
hyperthermia
|
|
KD
|
knockdown
|
|
MAPK/ERK
|
mitogen-activated protein
kinase/extracellular signal-regulated kinase
|
|
MHT
|
mild HT
|
|
OSCC
|
oral squamous cell carcinoma
|
|
pERK1/2
|
phosphorylated ERK1/2
|
|
siRNA
|
small interfering RNA
|
|
siHIKE
|
siRNA for HIKESHI
|
|
siLuc
|
siRNA for luciferase
|
References
|
1
|
van der Zee J, González González D, van
Rhoon GC, van Dijk JD, van Putten WL and Hart AA: Comparison of
radiotherapy alone with radiotherapy plus hyperthermia in locally
advanced pelvic tumours: A prospective, randomised, multicentre
trial. Dutch Deep Hyperthermia Group. Lancet. 355:1119–1125. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wust P, Hildebrandt B, Sreenivasa G, Rau
B, Gellermann J, Riess H, Felix R and Schlag PM: Hyperthermia in
combined treatment of cancer. Lancet Oncol. 3:487–497. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Issels RD: Hyperthermia adds to
chemotherapy. Eur J Cancer. 44:2546–2554. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zagar TM, Oleson JR, Vujaskovic Z,
Dewhirst MW, Craciunescu OI, Blackwell KL, Prosnitz LR and Jones
EL: Hyperthermia combined with radiation therapy for superficial
breast cancer and chest wall recurrence: A review of the randomised
data. Int J Hyperthermia. 26:612–617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Westermann A, Mella O, Van Der Zee J,
Jones EL, Van Der Steen-Banasik E, Koper P, Uitterhoeve AL, De Wit
R, Van Der Velden J, Burger C, et al: Long-term survival data of
triple modality treatment of stage IIB-III-IVA cervical cancer with
the combination of radiotherapy, chemotherapy and hyper-thermia-an
update. Int J Hyperthermia. 28:549–553. 2012. View Article : Google Scholar
|
|
6
|
Ahmed K, Tabuchi Y and Kondo T:
Hyperthermia: An effective strategy to induce apoptosis in cancer
cells. Apoptosis. 20:1411–1419. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ohguri T, Harima Y, Imada H, Sakurai H,
Ohno T, Hiraki Y, Tuji K, Tanaka M and Terashima H: Relationships
between thermal dose parameters and the efficacy of definitive
chemoradiotherapy plus regional hyperthermia in the treatment of
locally advanced cervical cancer: Data from a multicentre
randomised clinical trial. Int J Hyperthermia. 34:461–468. 2018.
View Article : Google Scholar
|
|
8
|
Bakker A, van der Zee J, van Tienhoven G,
Kok HP, Rasch CRN and Crezee H: Temperature and thermal dose during
radiotherapy and hyperthermia for recurrent breast cancer are
related to clinical outcome and thermal toxicity: A systematic
review. Int J Hyperthermia. 36:1024–1039. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tohnai I, Hayashi Y, Mitsudo K, Shigetomi
T, Ueda M and Ishigaki T: Prognostic evaluation of preoperative
thermochemo-radiotherapy for N(3) cervical lymph node metastases of
oral cancer. Oncology. 62:234–240. 2002. View Article : Google Scholar
|
|
10
|
Nozato T, Koizumi T, Hayashi Y, Iida M,
Iwai T, Oguri S, Hirota M, Kioi M, Koike I, Hata M, et al:
Thermochemora-diotherapy using superselective intra-arterial
infusion for patients with oral cancer with cervical lymph node
metastases. Anticancer Res. 39:1365–1373. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Urano M: Kinetics of thermotolerance in
normal and tumor tissues: A review. Cancer Res. 46:474–482.
1986.PubMed/NCBI
|
|
12
|
Sapareto SA: Thermal isoeffect dose:
Addressing the problem of thermotolerance. Int J Hyperthermia.
3:297–305. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Overgaard J: The current and potential
role of hyperthermia in radiotherapy. Int J Radiat Oncol Biol Phys.
16:535–549. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li GC, Mivechi NF and Weitzel G: Heat
shock proteins, thermotolerance, and their relevance to clinical
hyperthermia. Int J Hyperthermia. 11:459–488. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li GC and Calderwood SK: Hyperthermia
classic article commentary: 'Re-induction of hsp70 synthesis: An
assay for thermotolerance' by Gloria C. Li and Johnson Y. Mak,
International Journal of Hyperthermia 1989;5:389-403. Int J
Hyperthermia. 25:258–261. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lindquist S and Craig EA: The heat-shock
proteins. Annu Rev Genet. 22:631–677. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ohtsuka K and Hata M: Molecular chaperone
function of mammalian Hsp70 and Hsp40-a review. Int J Hyperthermia.
16:231–245. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hartl FU and Hayer-Hartl M: Molecular
chaperones in the cytosol: From nascent chain to folded protein.
Science. 295:1852–1858. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kregel KC: Heat shock proteins: Modifying
factors in physiological stress responses and acquired
thermotolerance. J Appl Physiol 1985. 92:2177–2186. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kampinga HH, Hageman J, Vos MJ, Kubota H,
Tanguay RM, Bruford EA, Cheetham ME, Chen B and Hightower LE:
Guidelines for the nomenclature of the human heat shock proteins.
Cell Stress Chaperones. 14:105–111. 2009. View Article : Google Scholar :
|
|
21
|
Akerfelt M, Morimoto RI and Sistonen L:
Heat shock factors: Integrators of cell stress, development and
lifespan. Nat Rev Mol Cell Biol. 11:545–555. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fujimoto M and Nakai A: The heat shock
factor family and adaptation to proteotoxic stress. FEBS J.
277:4112–4125. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Calderwood SK and Asea A: Targeting
HSP70-induced thermo-tolerance for design of thermal sensitizers.
Int J Hyperthermia. 18:597–608. 2002. View Article : Google Scholar
|
|
24
|
McMillan DR, Xiao X, Shao L, Graves K and
Benjamin IJ: Targeted disruption of heat shock transcription factor
1 abolishes thermotolerance and protection against heat-inducible
apoptosis. J Biol Chem. 273:7523–7528. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tabuchi Y, Furusawa Y, Wada S, Ohtsuka K
and Kondo T: Silencing heat shock transcription factor 1 using
small interfering RNA enhances mild hyperthermia and hyperthermia
sensitivity in human oral squamous cell carcinoma cells. Thermal
Med. 27:99–108. 2011. View Article : Google Scholar
|
|
26
|
Bettaieb A and Averill-Bates DA:
Thermotolerance induced at a mild temperature of 40°C alleviates
heat shock-induced ER stress and apoptosis in HeLa cells. Biochim
Biophys Acta. 1853:52–62. 2015. View Article : Google Scholar
|
|
27
|
Matozaki M, Saito Y, Yasutake R, Munira S,
Kaibori Y, Yukawa A, Tada M and Nakayama Y: Involvement of Stat3
phosphorylation in mild heat shock-induced thermotolerance. Exp
Cell Res. 377:67–74. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pelham HR: Hsp70 accelerates the recovery
of nucleolar morphology after heat shock. EMBO J. 3:3095–3100.
1984. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Welch WJ and Feramisco JR: Nuclear and
nucleolar localization of the 72,000-dalton heat shock protein in
heat-shocked mammalian cells. J Biol Chem. 259:4501–4513.
1984.PubMed/NCBI
|
|
30
|
Kose S, Furuta M and Imamoto N: Hikeshi, a
nuclear import carrier for Hsp70s, protects cells from heat
shock-induced nuclear damage. Cell. 149:578–589. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rahman KMZ, Mamada H, Takagi M, Kose S and
Imamoto N: Hikeshi modulates the proteotoxic stress response in
human cells: Implication for the importance of the nuclear function
of HSP70s. Genes Cells. 22:968–976. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yanoma T, Ogata K, Yokobori T, Ide M,
Mochiki E, Toyomasu Y, Yanai M, Kogure N, Kimura A, Suzuki M, et
al: Heat shock-induced HIKESHI protects cell viability via nuclear
translocation of heat shock protein 70. Oncol Rep. 38:1500–1506.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bhalla S, Chaudhary K, Kumar R, Sehgal M,
Kaur H, Sharma S and Raghava GP: Gene expression-based biomarkers
for discriminating early and late stage of clear cell renal cancer.
Sci Rep. 7:449972017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tabuchi Y, Wada S, Furusawa Y, Ohtsuka K
and Kondo T: Gene networks related to the cell death elicited by
hyperthermia in human oral squamous cell carcinoma HSC-3 cells. Int
J Mol Med. 29:380–386. 2012.
|
|
35
|
Yunoki T, Kariya A, Kondo T, Hayashi A and
Tabuchi Y: The combination of silencing BAG3 and inhibition of the
JNK pathway enhances hyperthermia sensitivity in human oral
squamous cell carcinoma cells. Cancer Lett. 335:52–57. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yunoki T, Tabuchi Y, Hayashi A and Kondo
T: Network analysis of genes involved in the enhancement of
hyperthermia sensitivity by the knockdown of BAG3 in human oral
squamous cell carcinoma cells. Int J Mol Med. 38:236–242. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Suzuki K, Bose P, Leong-Quong RY, Fujita
DJ and Riabowol K: REAP: A two minute cell fractionation method.
BMC Res Notes. 3:2942010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chang KL, Wong LR, Pee HN, Yang S and Ho
PC: Reverting metabolic dysfunction in cortex and cerebellum of
APP/PS1 mice, a model for Alzheimer's disease by pioglitazone, a
peroxi-some proliferator-activated receptor gamma (PPARγ) agonist.
Mol Neurobiol. 56:7267–7283. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yap K, Mukhina S, Zhang G, Tan JSC, Ong HS
and Makeyev EV: A short tandem repeat-enriched RNA assembles a
nuclear compartment to control alternative splicing and promote
cell survival. Mol Cell. 72:525–540.e13. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Furusawa Y, Yunoki T, Hirano T, Minagawa
S, Izumi H, Mori H, Hayashi A and Tabuchi Y: Identification of
genes and genetic networks associated with BAG3-dependent cell
proliferation and cell survival in human cervical cancer HeLa
cells. Mol Med Rep. 18:4138–4146. 2018.PubMed/NCBI
|
|
41
|
Hirano T, Minagawa S, Furusawa Y, Yunoki
T, Ikenaka Y, Yokoyama T, Hoshi N and Tabuchi Y: Growth and neurite
stimulating effects of the neonicotinoid pesticide clothianidin on
human neuroblastoma SH-SY5Y cells. Toxicol Appl Pharmacol.
383:1147772019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sarge KD, Murphy SP and Morimoto RI:
Activation of heat shock gene transcription by heat shock factor 1
involves oligomerization, acquisition of DNA-binding activity, and
nuclear localization and can occur in the absence of stress. Mol
Cell Biol. 13:1392–1407. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Todryk SM, Gough MJ and Pockley AG: Facets
of heat shock protein 70 show immunotherapeutic potential.
Immunology. 110:1–9. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ito A, Honda H and Kobayashi T: Cancer
immunotherapy based on intracellular hyperthermia using magnetite
nanoparticles: A novel concept of 'heat-controlled necrosis' with
heat shock protein expression. Cancer Immunol Immunother.
55:320–328. 2006. View Article : Google Scholar
|
|
45
|
Deschênes-Simard X, Kottakis F, Meloche S
and Ferbeyre G: ERKs in cancer: Friends or foes? Cancer Res.
74:412–419. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chen F, Rezavi R, Wang CC and Harrison LE:
Proteasome inhibition potentiates the cytotoxic effects of
hyperthermia in HT-29 colon cancer cells through inhibition of heat
shock protein 27. Oncology. 73:98–103. 2007. View Article : Google Scholar
|
|
47
|
Yunoki T, Tabuchi Y, Hayashi A and Kondo
T: BAG3 protects against hyperthermic stress by modulating NF-κB
and ERK activities in human retinoblastoma cells. Graefes Arch Clin
Exp Ophthalmol. 253:399–407. 2015. View Article : Google Scholar
|
|
48
|
Czubryt MP, Austria JA and Pierce GN:
Hydrogen peroxide inhibition of nuclear protein import is mediated
by the mitogen-activated protein kinase, ERK2. J Cell Biol.
148:7–16. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kosako H, Yamaguchi N, Aranami C, Ushiyama
M, Kose S, Imamoto N, Taniguchi H, Nishida E and Hattori S:
Phosphoproteomics reveals new ERK MAP kinase targets and links ERK
to nucleoporin-mediated nuclear transport. Nat Struct Mol Biol.
16:1026–1035. 2009. View Article : Google Scholar : PubMed/NCBI
|