Introduction
Osteoporosis is a systemic disease associated with
decreased bone mass, leading to reduced bone quality and an
increased risk of fracture (1)
due to the functional imbalance of osteoclasts and osteoblasts
(2). Osteoclasts and osteoblasts
are derived from hematopoietic stem cells (HSCs) and bone
marrow-derived mesenchymal stem cells (BMSCs) (3). In the bone marrow of patients with
osteoporosis, the adipogenic differentiation of BMSCs prevails over
osteogenic differentiation, leading to lipid accumulation (4,5).
An excessive amount of adipocytes in bone marrow may inhibit bone
formation and stimulate bone resorption (6). Adipocytes may produce several
adipokines such as leptin, adiponectin and chemerin (7) that alter bone matrix composition
(8). Hence, inhibiting the
adipogenesis of BMSCs may have the potential to increase
osteogenesis.
Statins are normally used to lower cholesterol in
patients with hyperlipidemia and cardiovascular diseases (9); however, they have also exhibited
efficacy in the treatment of bone loss-related diseases (10-12). The liposoluble simvastatin
exhibits anabolic effects on bones by stimulating osteogenesis,
inhibiting osteoblasts apoptosis (13), and by suppressing the
differentiation and activities of osteoclasts (13). Statins also increase the
expression of the bone morphogenic protein-2, enhancing bone
formation (14). An animal study
using ovariectomized rats suggested that the use of low-dose statin
increased bone resorption, while high-dose statin increased bone
formation (15). Simvastatin has
also been shown to nega-tively regulate the adipogenic
differentiation of BMSCs and preadipocytes (16) through the downregulation of the
peroxisome proliferator-activated receptor γ (PPARγ) (16,17).
PPARγ activation favors BMSC differentiation into
adipocytes, and the PPARγ blockade promotes osteogenesis (18). The overexpression of PPARγ and
lipid accumulation have been found in the bone marrow from
osteoporosis models (19). During
adipocyte differentiation, PPARγ directly stimulates the expression
and secretion of chemerin (20,21). Chemerin binds to at least 3
receptors: Chemokine-like receptor 1 (CMKLR1), G protein-coupled
receptor 1 (GPR1) and chemokine (C-C motif) receptor-like 2 (CCRL2)
(22-24). CCRL2 is an atypical receptor and
was not investigated in the present study. CMKLR1 and GPR1 are
similar in sequence and structure; thus, GPR1 may partially mediate
the function of chemerin (22-24). An increased level of chemerin or
the deficiency of CMKLR1 leads to bone loss in mice (25,26). Chemerin/CMKLR1 signaling is
essential for adipogenesis (20,27-29). On the other hand, the role of GPR1
in adipogenesis upon simvastatin treatment has been less
extensively studied, despite a link between GPR1 and bone
metabolism (30).
Nevertheless, the mechansims through which
simvastatin inhibit adipogenesis remain unclear. In the present
study, it was demonstrated that chemerin/CMKLR1 signaling drives
the adipogenesis of BMSCs. Simvastatin downregulated the expression
of chemerin, thereby inhibiting chemerin/CMKLR1 signaling and
adipogenesis. It was also founde that PPARγ induced the expression
of chemerin to activate downstream chemerin/CMKLR1 signaling.
Simvastatin thus exerts its inhibitory effects by inhibiting PPARγ
expression.
Materials and methods
Isolation, culture and identification of
BMSCs
All animal experiments were performed in compliance
with the research guidelines and were approved by the Ethics
Committee of the Shanxi Medical University (no. 2017005). The rats
were provided with free access to food and water. They were housed
in a facility at 25°C, 75% humidity, with a 12-12 h light/dark
cycle. BMSCs were isolated from one 4-week-old specific
pathogen-free (SPF) female Sprague-Dawley (SD) rat (Vital River
Laboratory), as previously described (30). The primary cells were cultured in
minimum essential medium, α-modification (α-MEM) containing 10%
fetal bovine serum (FBS) and 1% penicillin-streptomycin solution in
75-cm2 flasks. An atmosphere of 5% CO2 and
37°C was maintained. The medium was changed every 72 h until 80%
confluency was achieved. The BMSCs were trypsinized by treatment
with 0.05% trypsin-0.02% EDTA solution and passaged at a split
ratio of 1:3. Once the same level of confluence was obtained, the
cells at passage 3 were trypsinized and seeded at a density of
1.0×104 cells/cm2 in 6-cm dishes or 6-well
plates pre-coated with 0.1% gelatin solution.
According to the International Society for Cellular
Therapy position statement (31),
the identification of the cells was performed by examining the
morphological features and flow cytometry, as previously described
(30). The differentiation
capacity of the BMSCs was also detected using SD rat bone
mesenchymal stem cell osteogenic differentiation medium kit
(RASMX-90021) and adipogenic differentiation medium kit
(RASMX-90031), according to the manufacturer's instructions [Cyagen
Biosciences (Guangzhou) Co., Ltd.].
Flow cytometry
Flow cytometry was performed according to a
previously published method (30). BMSCs at passage 3 were trypsinized
according to the method described above, and were then centrifuged
at 179 × g for 4 min at room temperature. The cell concentration
was then adjusted to 1.0×107 cells/ml. Subsequently, the
cells were incubated with antibodies (BD Biosciences) against CD44
(FITC mouse anti-rat CD44, cat. no. 550974), CD45 (PE-Cy™5 mouse
anti-rat CD45, cat. no. 559135), CD90 (APC mouse anti-rat CD90,
cat. no. 561409) and CD11b (PE mouse anti-rat CD11b, cat. no.
562105) at a concentration of 10 µl of a 1/50 dilution for
106 cells in 100 µl. Following incubation for 30
min at 4°C protected from light, the samples were washed twice with
wash buffer and resuspended in flow cytometry buffer. Fluorescence
was measured using a FACSCalibur (BD Biosciences).
Proliferation analysis of BMSCs
The proliferation assay was performed using Cell
Counting kit-8 (CCK-8) according to the manufacturer's instructions
(AR1160, Wuhan Boster Biological Technology, Ltd.). Briefly, BMSCs
were seeded in 96-well plates (1,000 cells/well) and cultured in
the maintenance medium, which was changed every 3 days, with
simvastatin (HY-17502, MedChemExpress) at various concentrations
(10−5 M, 10−6 M, 10−7 M and
10−8 M). PBS was added to the control group. The CCK-8
solution (10 µl/well) was added at 1 h before absorbance
values at 450 nm were read using a SPECTRA max PLUS microplate
reader (Molecular Devices, LLC) to draw the growth curves.
Adipocyte differentiation
The conditioned media included basal media A and B.
Basal medium A contained α-MEM, 1% penicillin-streptomycin solution
(Wuhan Boster Biological Technology, Ltd.), 10% FBS (ScienCell
Research Laboratories, Inc.), 10 µM insulin, 200 µM
indomethacin, 500 µM 3-isobutyl-1-methylxanthine and 1
µM dexamethasone (Sigma-Aldrich; Merck KGaA). Basal medium B
contained α-MEM, FBS, penicillin-streptomycin solution and insulin
at the same concentrations as in basal medium A. The adipogenic
induction of BMSCs followed a protocol for 3 cycles involving
incubation in basal medium A for 3 days and then in basal medium B
for 1 day. Subsequently, the cells were incubated in basal medium B
until day 14 of differentiation. Moreover, to examine the role of
PPARγ, 0.5 µM rosiglitazone (HY-17386, MedChemExpress) was
used as an agonist; thus, it was not included in the adipogenic
induction medium.
Simvastatin was diluted beforehand with PBS and was
added to the medium at a final concentration of 10−7 M
throughout the differentiation process. The conditioning medium of
the control group contained the same amount of PBS. Oil Red O
staining was performed at 14 days after induction to verify the
effects of different intervention schemes of simvastatin
(simvastatin incubation for 14 days; for the first 3 days; after 3
days) on adipogenesis. The adipogenic differentiated cells were
washed twice with PBS, and then fixed with 2 ml of 4% neutral
formaldehyde (Boster Biological Technology) for 30 min. The cells
were washed again, and subsequently stained with filtered Oil Red O
solution (60% Oil Red O dye and 40% water) (Cyagen Biosciences,
Inc.) at room temperature for 30 min. The Oil Red O dye was further
extracted from the adipogenic cells with 1 ml isopropanol before
absorbance values at 490 nm were read.
Adenovirus generation and
transduction
CCX832, the only small-molecule antagonist for
CMKLR1, is not commercially available (24). Therefore, the present study sought
to knock down CMKLR1 (NM_008153) expression by RNA interference
(RNAi). Virus packaging requires two types of plasmids. One is the
packaging plasmid or auxiliary plasmid, which is used to express
the capsid elements of the virus and to form the virus itself. The
other is the shuttle vector, which is used to carry the shRNA
expression system and is wrapped in the virus shell to form
functional viral particles. An adenovirus backbone vector
containing the target sequence 5′-GCA ATG GCC TGG TGA TTG TCA-3′
was synthesized by GenePharma and co-transfected into 293A cells
(Cell Bank of the Chinese Academy of Sciences) along with a shuttle
vector, according to the manufacturer′s instructions. The negative
control carried a nonsense shRNA sequence (5′-GTT CTC CGA ACG TGT
CAC GT-3′). This sequence does not target any gene in human, rat,
or mouse. Adenovirus solution with a titer of 108 TU/ml
was prepared, and a multiplicity of infection of 100 was selected
for subsequent BMSC transfection. The BMSCs were seeded onto 6-cm
dishes (for western blot analysis) or 6-well plates [for reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)] 24
h prior to transfection. The adenovirus solution was then diluted
using the transfection medium, which was 10% FBS and 5 µg/ml
polybrene (Sigma-Aldrich; Merck KGaA) containing α-MEM, and added
into the culture dishes or plates. The transfection medium was
replaced with culture medium after 24 h. After a further 24 h, the
cell culture medium was replaced with the induction medium for
differentiation. The differentiated cells were transfected using
the adenovirus at 8 days post-differentiation with the same
protocol to maintain interference efficiency. Adenovirus
transfection efficiency was evaluated using an inverted
fluorescence microscope (LX70; Olympus Corporation) by quantifying
GFP expression. The interference efficiency was verified by the
analysis of the mRNA and protein expression levels of CMKLR1.
Western blot analysis
For western blot analysis, proteins from the 14-days
cultured cells of different groups were extracted using cell lysis
buffer containing 1% phenylmethanesulfonyl fluoride and were
centrifuged at 12,000 × g for 5 min at 4°C. A BCA Protein
Concentration Assay kit (AR0146, Wuhan Boster Biological
Technology, Ltd.) was used to determine the protein concentration,
according to the manufacturer's instructions. 10% SDS-PAGE gel was
used to separate the proteins, 50 µg/lane of which was
transferred onto a polyvinylidene difluoride membrane. The membrane
was then incubated with primary antibodies for adiponectin
(ab22554, mouse monoclonal antibody, 1:1,000 dilution, Abcam),
CMKLR1 (ab64881, rabbit polyclonal antibody, 1:500 dilution,
Abcam), β-actin (A5441, mouse monoclonal antibody, 1:10,000
dilution, Sigma-Aldrich; Merck KGaA) or β-tubulin (AF1216, rabbit
monoclonal antibody, 1:10,000 dilution, Beyotime Institute of
Biotechnology) overnight at 4°C. The membrane was incubated with
the appropriate secondary antibodies (adiponectin, A0216,
horseradish peroxidase-conjugated goat anti-mouse IgG, 1:1,000
dilution, Beyotime Institute of Biotechnology; β-tubulin, A0208,
horseradish peroxidase-conjugated goat anti-Rabbit IgG, 1:1,000
dilution, Beyotime Institute of Biotechnology; CMKLR1, 33101ES60,
horseradish peroxidase-conjugated goat anti-rabbit IgG, 1:2,000
dilution, Yeasen Biotechnology (Shanghai) Co., Ltd.; β-actin:
33201ES60, horseradish peroxidase-conjugated goat anti-mouse IgG,
1:2,000 dilution, Yeasen Biotechnology (Shanghai) Co., Ltd.) for 2
h at room temperature. The specific blotting bands were detected by
electrogenerated chemiluminescence and analyzed using a Gel-Pro
Analyzer (Media Cybernetics). Protein expression was normalized to
that of β-tubulin (Fig. 1) or
β-actin (Fig. S4). Densitometry
was performed using Gelpro32 4.0 software (Media Cybernetics).
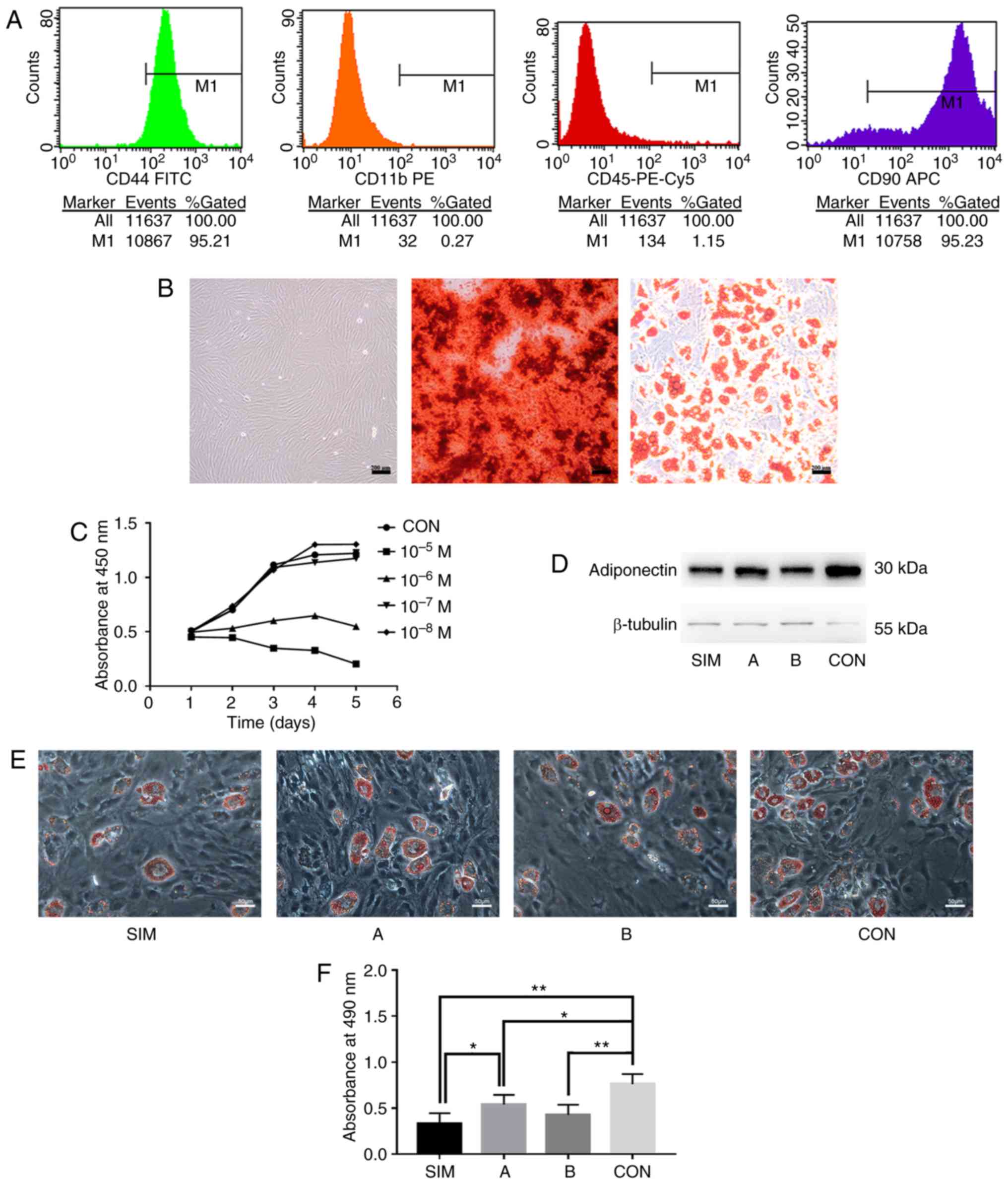 | Figure 1Determination of the concentration
and scheme of simvastatin stimulation. (A) Prevalence and number of
cells expressing the cell surface marker CD44, CD90, CD11b, or CD45
examined by flow cytometry. (B) Representative images of BMSCs from
passage 3 under a light microscope (left panel), Alizarin Red
staining after osteogenic induction (middle panel) and Oil Red O
staining following adipogenic induction (right panel)
(magnification, ×100). (C) Growth curves of BMSCs treated with
simvastatin at various concentrations or PBS (n=5). Data were
examined by analysis of variance followed by Dunnett's post hoc
test. (D) Western blot analysis of adiponectin levels. (E)
Representative images of Oil Red O staining on differentiated BMSCs
from different groups (magnification, ×200). (F) Absorbance values
(n=3) at 490 nm of cells as in (E). The data are expressed as the
means ± SD. Data were examined by analysis of variance followed by
the LSD post hoc test. *P<0.05 and
**P<0.01. BMSCs, bone marrow-derived mesenchymal stem
cells; SIM, BMSCs treated with simvastatin for 14 days; A, for the
first 3 days; B, after 3 days; and CON, with PBS for 14 days. |
Gene expression analysis
Gene expression analysis was performed using
RT-qPCR. According to the manufacturer's instructions, RNAiso Plus
(9108, Takara Bio, Inc.) was used to extract total RNA from the
cultured cells of the different groups. The absorbance values at
260 nm were analyzed using a biospectrometer (Eppendorf AG) to
determine the concentrations of extracted RNA, 0.4 µg of
which were subsequently mixed with 5X PrimeScript RT Master Mix
(RR036A, Takara Bio, Inc.) and RNase-free dH2O. The
mixed solution (10 µl) was used in reverse transcription
reactions in a Veriti 96-Well Fast Thermal Cycler (Applied
Biosystems) to generate cDNA. The cDNA solution was then mixed with
SYBR Premix Ex Taq II (2X) (RR820A, Takara Bio, Inc.), primers (0.4
µM), ROX Reference Dye, and RNase Free dH2O prior
to quantitative polymerase chain reaction. mRNA expression analysis
was performed using the StepOnePlus Real-Time PCR System (Applied
Biosystems) in a two-step protocol according to the following
conditions: Initial denaturation for 30 sec at 95°C, followed by 40
cycles with denaturation for 5 sec at 95°C, and annealing and
extension for 30 sec at 60°C. All primer sequences are provided in
Table I. The expression levels of
PPARγ, chemerin, CMKLR1, GPR1 and adiponectin were calculated
relative to those of GAPDH by the 2−ΔΔCq method
(32) for the 3, 7 and
14-day-differentiated cell samples. The melting curves obtained
were examined for the specificity of the products prior to
analysis.
 | Table IPrimer sequences used for
RT-qPCR. |
Table I
Primer sequences used for
RT-qPCR.
| Gene | Primer sequence
(5′→3′) | Product size |
|---|
| GAPDH | Fw:
GATGCTGGTGCTGAGTATGT | 104 bp |
| Rv:
GCGGAGATGATGACCCTTT | |
| PPARG | Fw:
TCCCGTTCACAAGAGCTGAC | 107 bp |
| Rv:
ATAATAAGGCGGGGACGCAG | |
| RARRES2 | Fw:
GGTGTGGACAGTGCTGATGA | 169 bp |
| Rv:
TGGGGTCCAGTTTGATGCAG | |
| CMKLR1 | Fw:
AGTGACTGATCAGCCGAGGA | 141 bp |
| Rv:
GATGTAGTCCGAGCCGTCAG | |
| GPR1 | Fw:
CCGGACCCTGAAGAACTCAC | 86 bp |
| Rv:
CCCGGAAGTACAGGGTAGGA | |
| ADIPOQ | Fw:
CAGCATTCAGCGTAGGGC | 191 bp |
| Rv:
GAAGAGGCTCACTTTCACATCC | |
Statistical analysis
Statistical analyses were performed using SPSS 22.0
(IBM Corp.). The results are expressed as the means ± SD. A
two-tailed Student's t-test was used to compare the mean values
between the 2 groups. One-way analysis of variance (ANOVA) followed
by the Dunnett or LSD post hoc tests was performed in the case of
multiple comparisons. P<0.05 was considered to indicate a
statistically significant difference.
Results
Identification of BMSCs
To identify the BMSCs of SD rats, flow cytometry was
performed and the presence of the BMSC marker proteins, CD44 and
CD90, and the absence of the monocyte marker, CD11b, or
hematopoietic cell marker, CD45, were observed on the cell surface
(Fig. 1A). The morphological
features and differentiation capacity of the cultured cells were
also examined and it was found that cultured BMSCs from passage 3
maintained a fibroblast-like morphology (Fig. 1B). The cells were incubated in a
specific induction medium to induce their differentiation into
osteoblasts or adipocytes. At 28 days postosteogenic
differentiation, calcium deposition was observed (Fig. 1B). Oil Red O staining at 21 days
postadipogenic differentiation revealed lipid accumulation within
the cells (Fig. 1B). Taken
together, these lines of evidence suggest that these cells were
BMSCs and could be used for further experiments.
Determination of the concentration and
intervention scheme of simvastatin
To determine whether various concentrations of
simvastatin exert differential effects on cell growth, cell growth
was monitored following simvastatin stimulation for 5 days. The
results revealed that simvastatin slightly enhanced the
proliferation of BMSCs at a low concentration (10−8 M),
but inhibited cell growth at higher concentrations (10−5
M and 10−6 M) (Fig.
1C). Cells treated with 10−7 M simvastatin exhibited
a similar growth pattern compared to the control group (Fig. 1C).
To verify the intervention scheme of simvastatin on
adipo-genesis, Oil Red O staining of adipogenic BMSCs treated with
simvastatin for 14 days (SIM group) was performed. In addition,
BMSCs were examined during the first 3 days (group A, early stage
of adipogenesis) and after 3 days (group B) of adipogenesis; BMSCs
treated with PBS (CON group) were also examined. The protein
expression of adiponectin in the simvastatin-treated group was
markedly decreased compared to that of the control group, and the
lipid accumulation within the cells exhibited a similar
simvastatin-dependent lipid loss (Fig. 1D and E), indicating that
simvastatin inhibited the adipogenic differentiation of BMSCs
during the whole process of adipogenesis. Moreover, the
simvastatin-treated group (SIM group) exhibited a lower absorbance
at 490 nm compared with the control group or group A (Fig. 1F). The SIM group exhibited a
comparable level of absorbance with group B (Fig. 1F). In addition, the fact that the
inhibitory effect was more potent in group B and the SIM group than
in group A (Fig. 1F) indicated
that longer stimulation with simvastatin enhanced its inhibitory
effects. The P-values obtained for the data in Fig. 1 are provided in detail in Tables SI and SII.
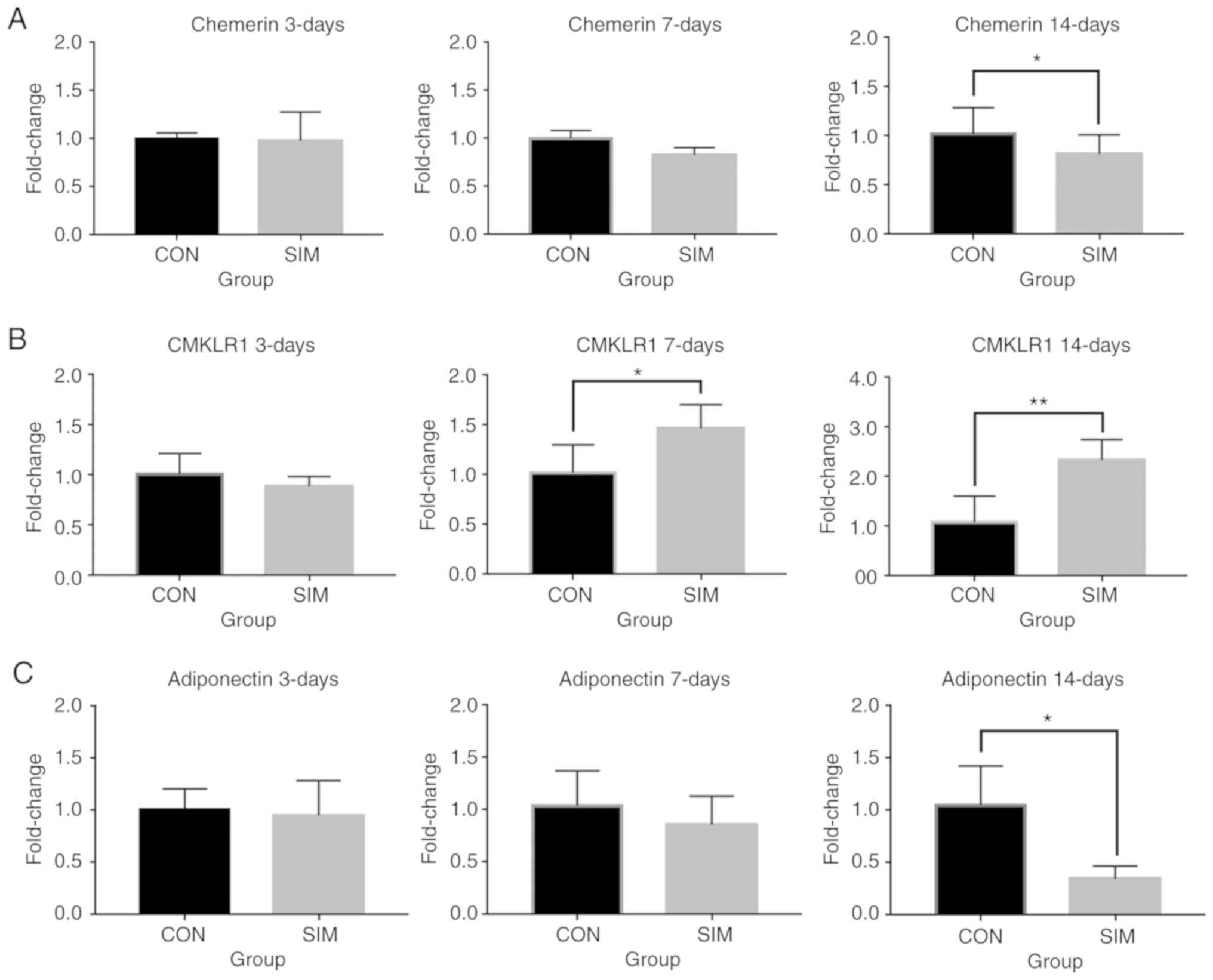
Effects of simvastatin on PPARγ,
chemerin/CMKLR1 signaling and adiponectin expression
In agreement with previous studies, the expression
of chemerin increased after 3, 7 and 14 days of adipogenesis, while
one of the receptors for chemerin, CMKLR1, exhibited a trend
towards a decreased expression (data not shown).
Simvastatin treatment did not affect the overall
kinetics of the expression patterns and exerted no marked effects
on the expression of chemerin and CMKLR, compared to the control
group on day 3 (Fig. 2A and B).
On days 7 and 14, the simvastatin-treated cells exhibited a
downregulated expression of chemerin, whereas they exhibited an
upregulated expression of its receptor, CMKLR1 (Fig. 2A and B). Notably, the expression
of the adipocyte marker, adiponectin, was decreased compared with
the control at day 14 (Fig. 2C),
suggesting that simvastatin exerted an inhibitory effect on
adipogenesis. PPARγ expression increased during adipogenesis while
the expression of the other chemerin receptor, GPR1, exhibited a
decreasing trend (data not shown). Simvastatin treatment decreased
PPARγ expression on day 3 (Fig.
S1). Simvastatin treatment did not affect the expression of
GPR1 on days 3 and 7, but induced GPR1 expression on day 14
(Fig. S2A). The P-values for the
data in Fig. 2 are provided in
detail Tables SIII-SV.
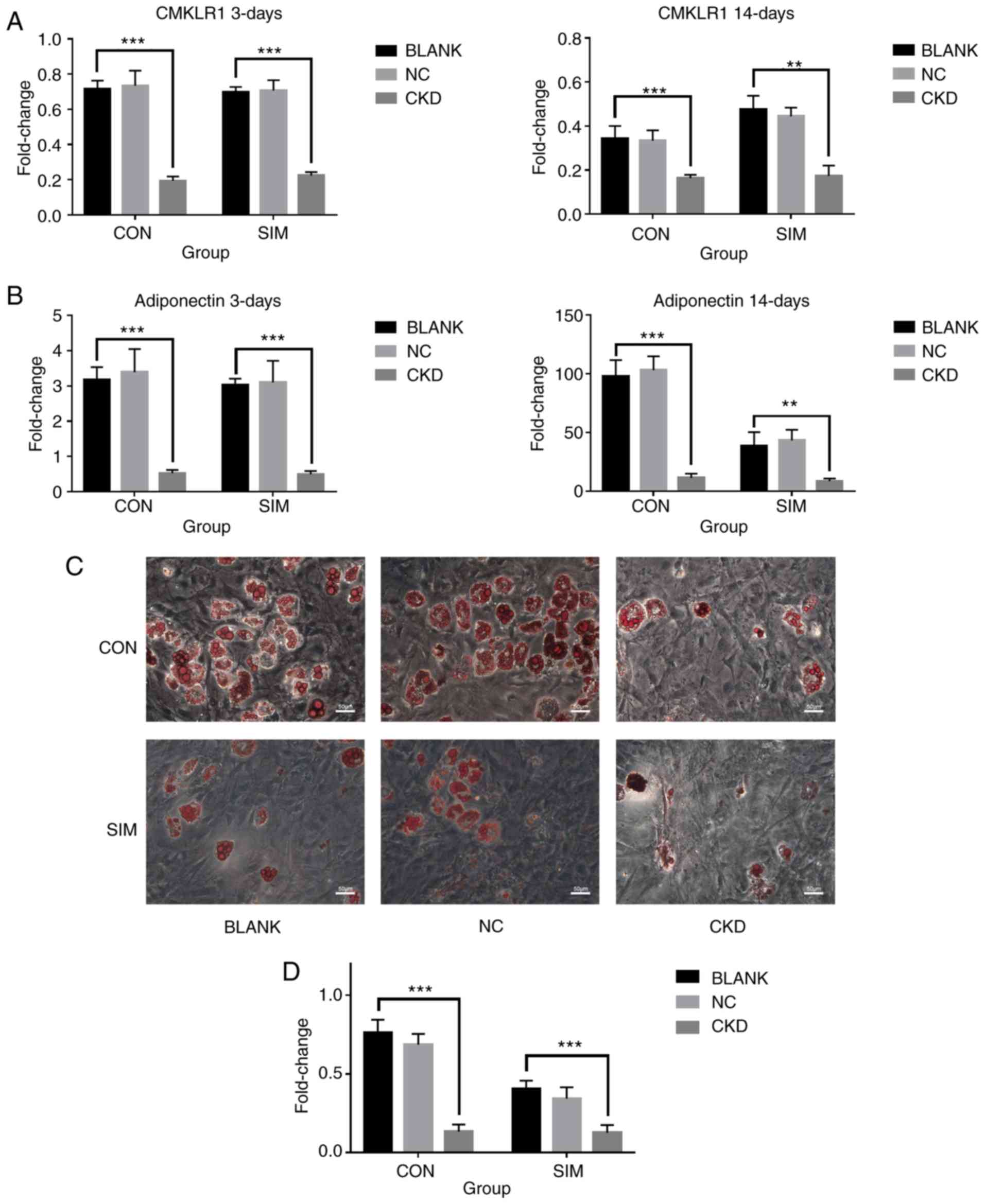
CMKLR1 is required for adipogenesis and
the simvastatin-mediated inhibitory effect on adipogenesis
Since simvastatin was found to differentially
regulate the expression of chemerin and its receptor CMKLR1 in
BMSCs, the present study directly examined the role of
chemerin/CMKLR1 signaling during the simvastatin-mediated
inhibition of adipogenesis by knocking down CMKLR1 in BMSCs using
RNAi. BMSCs were transfected with adenovirus at 48 h prior to
adipogenic differentiation, and the immunofluorescence of each
group was confirmed (Fig. S3).
In comparison with the control group, a markedly lower expression
of CMKLR1 at the protein (Fig.
S4) and mRNA (Fig. 3A) level
was detected on day 3 post-differentiation. The expression of GPR1,
which shares sequence and structural similarities with CMKLR1, was
also markedly decreased on days 3 and 14 post-differentiation
(Fig. S2B), while a compensatory
increase was not observed. Of note, the knockdown of CMKLR1
markedly inhibited adipogenesis, as evidenced by the decreased
expression of adiponectin at 14 days post-differentiation (Fig. 3B). Consistently, decreased Oil Red
O staining was observed in the cells in which CMKLR1 was knocked
down (Fig. 3C), which was further
confirmed by quantitative analysis (Fig. 3D), suggesting that the inhibition
of chemerin/CMKLR1 signaling prevented the adipogenesis of BMSCs.
Notably, simvastatin stimulation did not further decrease the
expression of adiponectin, or Oil red O staining in the cells in
which CMKLR1 was knocked down (Fig.
3C and D), suggesting that the inhibitory effects of
simvastatin on adipogenesis of BMSCs are dependent on
chemerin/CMKLR1 signaling. This is consistent with the negative
regulation of chemerin expression by simvastatin stimulation
observed above (Fig. 2), further
supporting the notion that simvastatin negatively regulates
adipogenesis by modulating chemerin/CMKLR1 signaling. The P-values
for the data presented in Fig. 3
are provided in detail in Tables
SVI-SVIII.
PPARγ agonist, rosiglitazone, partially
reverses the effects of simvastatin on the expression of specific
genes
To further determine whether PPARγ targets
chemerin/CMKLR1 signaling during the simvastatin-mediated
inhibition, the cells were treated with the PPARγ agonist,
rosiglitazone. On days 3 and 14, rosiglitazone significantly
increased the expression of PPARγ (Fig. 4A) and chemerin (Fig. 4B), suggesting that PPARγ indeed
activates chemerin/CMKLR1 signaling downstream. Moreover,
rosiglitazone also partially reversed the downregulation of PPARγ
and chemerin induced by simvastatin on days 3 and 14 (Fig. 4A and B). The expression of CMKLR1
and the other receptor GPR1 were decreased by treatment with
rosiglitazone (Figs. 4C and
S2C). Consistent with an
increase in the expression of chemerin, an increase in adipogenesis
was observed in the rosiglitazone-treated BMSCs following
simvastatin stimulation, as evidenced by the higher expression of
adiponectin at 14 days compared to the control group (Fig. 4D). The P-values for the data
presented in Fig. 4 are provided
in detail in Tables SIX-SX.
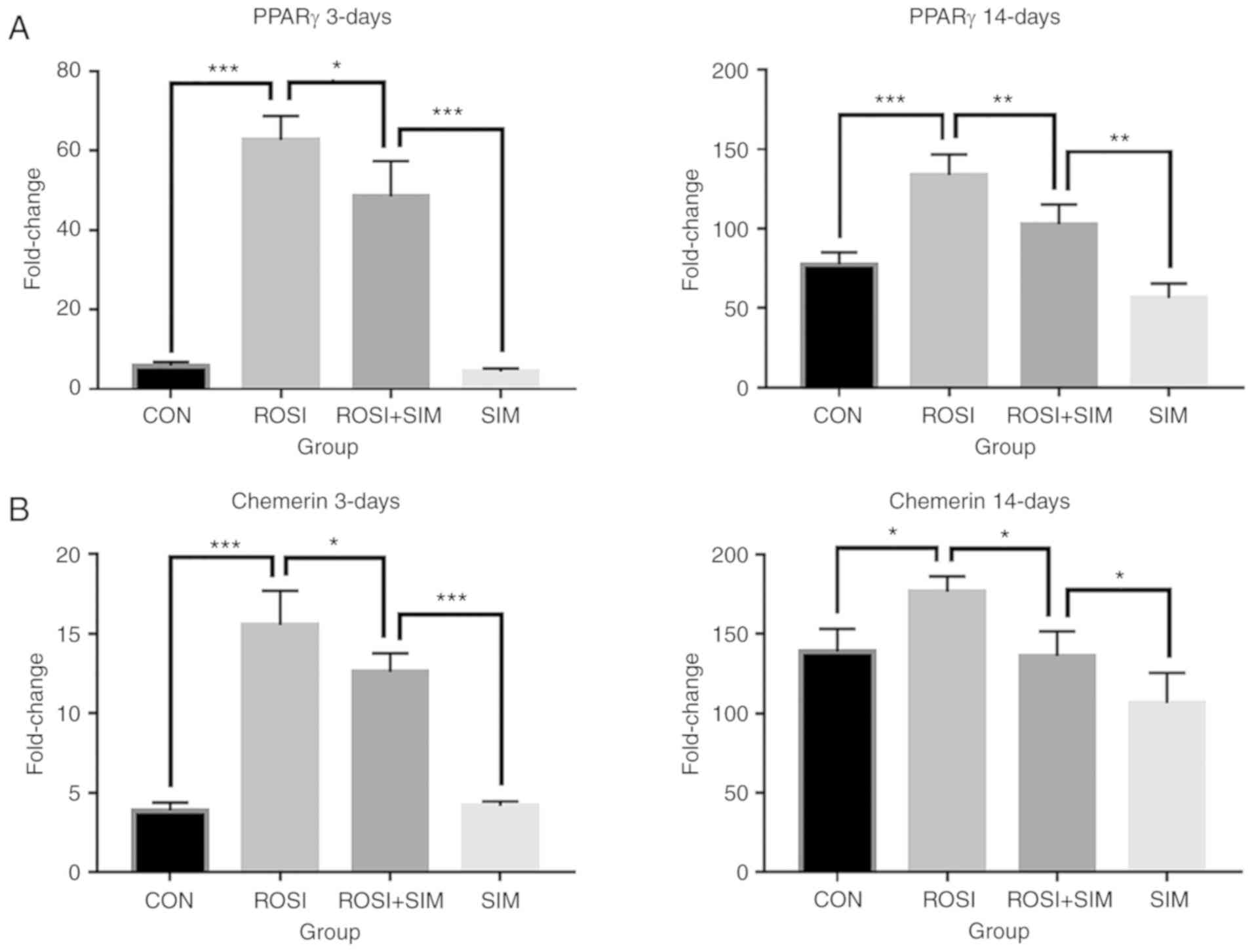 | Figure 4PPARγ agonist rosiglitazone partly
reverses the effects of simvastatin on the expression of specific
genes. At days 3 and 14 post-differentiation, cells that exhibited
adipogenic differentiation with PBS (CON), rosiglitazone (ROSI),
rosiglitazone and simvastatin (ROSI + SIM) and simvastatin (SIM)
treatment were analyzed by RT-qPCR for expression of (A) PPARγ, (B)
chemerin, (C) CMKLR1, and (D) adiponectin (n=3). Fold change was
expressed relative to undifferentiated BMSCs, and the data are
expressed as the means ± SD. Data were examined by analysis of
variance followed by the LSD post hoc test. *P<0.05,
**P<0.01 and ***P<0.001. BMSCs, bone
marrow-derived mesenchymal stem cells; CMKLR1, chemokine-like
receptor 1; PPARγ, peroxisome proliferator-activated receptor
γ. |
Taken together, these data suggest that simvastatin
inhibits PPARγ-mediated chemerin signaling to prevent adipogenesis
in BMSCs and that the activation of PPARγ signaling by
rosiglitazone is able to reverse, at least partially, such an
inhibitory effect.
Discussion
In addition to clinical applications for the
treatment of hyperlipidemia and cardiovascular diseases, statins
reduce the production of mevalproic acid and subsequent downstream
products involved in various physiological and pathological
processes (33). These are the
so-called statin pleiotropic effects, one of which is on
adipogenesis through the chemerin signaling (29,34). Other examples are that statins may
activate Wnt/β-catenin signaling (35,36) and inhibit Rho (37,38) in different biological processes,
and coincidentally, both CMKLR1 and GPR1 mediate chemerin signaling
through the RhoA/ROCK pathway (39), and the former has been proven to
be a Wnt target gene (27). Thus,
the regulation of chemerin signaling via multiple mechanisms may,
at least partly, account for the pleiotropic effects of statins.
Nevertheless, although this coincidence indicates a possible link
between chemerin signaling and statins, no direct evidence has been
found to date. To the best of our knowledge, the present study is
the first to bridge the inhibitory effect of simvastatin on
adipogenesis of BMSCs and chemerin signaling through the regulation
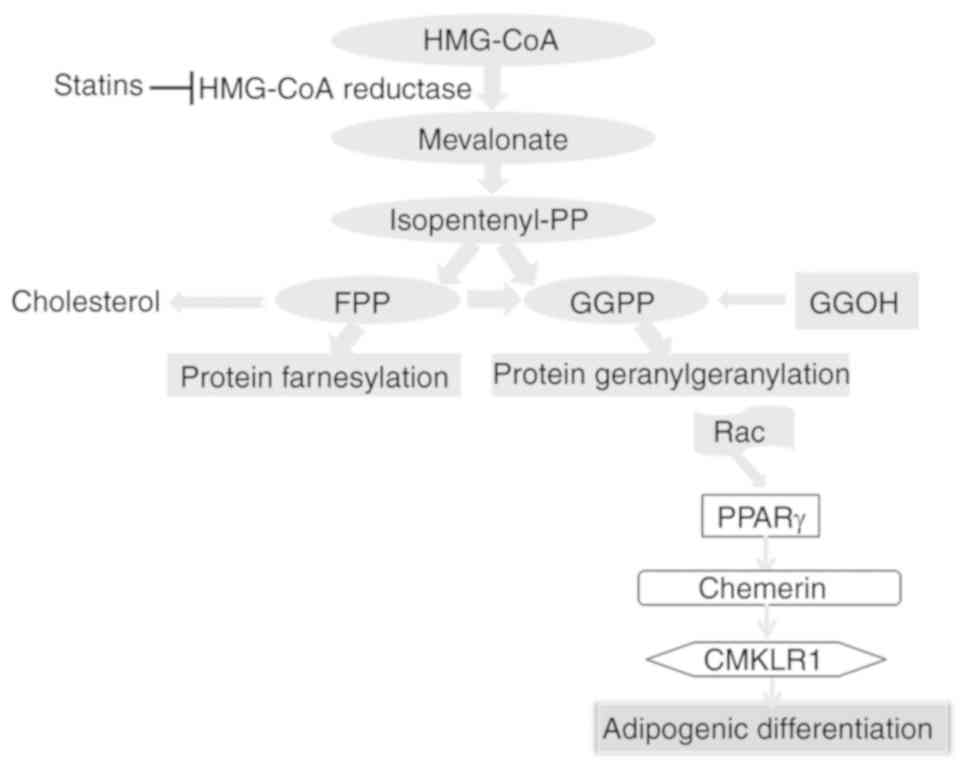
of PPARγ (Fig. 5).
Owing to evidence that the post-confluence clone
expansion of BMSCs may influence adipogenesis (40), and that simvastatin had effects,
though with controversy, on BMSC proliferation (41,42), the present study selected the
concentration of simvastatin which minimally influenced
proliferation to minimize its effect on proliferation-related
adipogenesis. Furthermore, although the early stage is of vital
importance for adipogenesis (28), simvastatin intervention was
performed in the whole adipogenic differentiation process of BMSCs
to maintain its continuous role in adipogenesis and its possible
role on the expression of PPARγ and chemerin signaling. It has been
reported that the decreased expression of PPARγ leads to the
simvastatin-mediated inhibition of adipogenesis (43,44), and chemerin, which plays vital
roles in adipogenesis, was a target gene of PPARγ in adipogenesis
of BMSCs (20). The results of
the present study demonstrated that the expression of both genes
increased significantly during the adipogenic differentiation
process, and simvastatin intervention inhibited such increase,
demonstrating that the inhibitory effect of simvastatin may be due
to the downregulation of PPARγ and chemerin signals.
It is noteworthy that while the chemerin receptor
CMKLR1 expression was upregulated by simvastatin, chemerin-mediated
adipogenesis was still inhibited by simvastatin. This may be due to
that the decreased expression of chemerin by simvastatin
stimulation overrides the elevated expression of CMKLR1, thereby
leading to the inhibition of chemerin-CMKLR1 signaling and reduced
adipogenesis. Similarly, the PPARγ agonist rosiglitazone partially
rescued the inhibitory effect of simvastatin but reduced the levels
of CMKLR1. In this case, the upregulated expression of chemerin by
PPARγ activation may override CMKLR1 down-regulation leading to
increased chemerin-CMKLR1 signaling and adipogenesis. Indeed, PPARγ
has been shown to decrease b-catenin-dependent CMKLR1 expression by
promoting the degradation of b-catenin (29). Moreover, consistent with the
findings of the present study, the downregulation of CMKLR1 is
associated with an enhancement of chemerin/CMKLR1 signaling leading
to bone resorption activity in mature osteoclasts (28).
Apart from the expression of PPARγ, an increase in
chemerin expression was also detected by rosiglitazone intervention
in the present study, and a possible role of PPARγ in the
simvastatin-mediated inhibition of adipogenesis was demonstrated.
Moreover, the decreased expression of both chemerin receptors,
CMKLR1 and GPR1, along with adipogenesis, was further reduced in
the context of rosiglitazone, and such a decrease was partially
reversed by simvastatin intervention. To further validate this
hypothesis, RNAi was used to knock down the expression of CMKLR1.
Simvastatin treatment did not exert a further inhibition of
adipogenesis with CMKLR1 knockdown; in other words, simvastatin
intervention did not affect adipogenesis without functional CMKLR1.
Hence, CMKLR1 mediates the downregulated signals of PPARγ and
chemerin in the process of simvastatin-mediated inhibition of
adipogenesis (Fig. 5).
Considering the controversies in functional studies
of CMKLR1, researchers have proposed that the functional
differences of CMKLR1 may partlly be due to the activation of GPR1
by chemerin (28). The results of
the present study demonstrated a similar decrease in the gene
expression levels of CMKLR1 and GPR1 during adipogenesis, and the
decreased expression of both receptors was inhibited by
simvastatin, thus suggesting that GPR1 may function similarly as
CMKLR1 during this process. Nevertheless, although both receptors
can modulate the osteogenic differentiation of BMSCs, only CMKLR1
was proven to be effective in adipogenesis. Surprisingly, it was
found that GPR1 expression markedly decreased after CMKLR1
knockdown, and this decrease was not promoted by simvastatin. These
results indicate that in the absence of CMKLR1, GPR1 may not
significantly affect the inhibitory effect of simvastatin, and the
increase in the expression of GPR1 with simvastatin may simply be
due to the inhibitory effect of simvastatin on adipogenesis.
Moreover, GPR1 showed lower expression than CMKLR1, demonstrating
that the function of GPR1 may not be as important as that of CMKLR1
during adipogenesis. This result was in accordance with the
findings of the study by Rourke et al, that GPR1 did not
contribute to adipogenesis (45).
Despite the expression data on GPR1 from the present study, the
role of this receptor in the adipogenesis of BMSCs remains to be
further revealed.
In the present study, the effects of simvastatin on
chemerin and CMKLR1 expression were not verified at the protein
level, and translation, post-translation processing and
modification, and different detection times may lead to
inconsistent mRNA and protein levels. It is not known whether the
lipid-inhibiting effect of simvastatin is directly achieved through
the chemerin and CMKLR1 proteins, or whether it affects proteins
downstream of the chemerin signaling pathway. This is a limitation
of the present study. Therefore, the expression of chemerin and
CMKLR1 at the protein level may or may not be consistent with the
changes at the RNA level. To solve the problem of gene expression
differences at different levels, well-designed experiments are
required to eliminate the interference of various factors in gene
translation.
In conclusion, for the first time, to the best of
our knowledge, the present study demonstrates that PPARγ bridges
the inhibitory effect of statins on adipogenesis and chemerin
signaling. Simvastatin inhibits the adipogenesis of BMSCs through
the downregulation of PPARγ and subsequently prevents
PPARγ-mediated induction of chemerin/CMKLR1 signaling. This
partially explains how simvastatin inhibits the adipogenesis of
BMSCs and provides new ideas for the clinical application of
statins on osteoporosis treatment. Considering that there may be
more possible connections between chemerin signaling and the
pleiotropic effects of statins, more information about the relevant
downstream events must be obtained to have a better understanding
of their interplay.
Supplementary Data
Funding
The present study was supported by the Shanxi
Province Science and Technology Research Project (grant no.
20150313012-1); and Key Research and Development (R&D) Projects
of Shanxi Province (grant no. 201803D31134).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YG was the main contributor to the conception and
design of the present study and in the drafting of the manuscript.
JH and DW played an important role in data analysis and contributed
to the writing of the manuscript. HH and XJ performed the
experiments of cell culture, cell identification and RNAi. EZ and
BN performed RT-qPCR and western blot analysis, and analyzed the
collected data. QL played an important role in the conception and
design of the study, and revised the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were performed in compliance
with the research guidelines and approved by the Ethics Committee
of the Shanxi Medical University (no. 2017005).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Abbreviations:
|
BMSCs
|
bone marrow-derived mesenchymal stem
cells;
|
|
HSCs
|
hematopoietic stem cells;
|
|
PPAR γ
|
peroxisome proliferator-activated
receptor γ;
|
|
CMKLR1
|
chemokine-like receptor 1;
|
|
GPR1
|
G protein-coupled receptor 1;
|
|
HMG-CoA reductase
|
3-hydroxy-3-methylglutaryl-coenzyme A
reductase;
|
|
FPP
|
farnesyl pyrophosphate;
|
|
GGPP
|
geranylgeranyl diphosphate;
|
|
GGOH
|
geranylgeraniol
|
Acknowledgments
Not applicable.
References
|
1
|
Kanis JA, McCloskey EV, Johansson H, Oden
A, Melton LJ III and Khaltaev N: A reference standard for the
description of osteoporosis. Bone. 42:467–475. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Teitelbaum SL: Bone resorption by
osteoclasts. Science. 289:1504–1508. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Marks SC Jr: The origin of osteoclasts:
Evidence, clinical implications and investigative challenges of an
extra-skeletal source. J Oral Pathol. 12:226–256. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chamberlain G, Fox J, Ashton B and
Middleton J: Concise review: Mesenchymal stem cells: Their
phenotype, differentiation capacity, immunological features, and
potential for homing. Stem Cells. 25:2739–2749. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bianco P, Robey PG, Saggio I and Riminucci
M: 'Mesenchymal' stem cells in human bone marrow (skeletal stem
cells): A critical discussion of their nature, identity, and
significance in incurable skeletal disease. Hum Gene Ther.
21:1057–1066. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Muruganandan S and Sinal CJ: The impact of
bone marrow adipocytes on osteoblast and osteoclast
differentiation. IUBMB Life. 66:147–155. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
de Paula FJA and Rosen CJ: Structure and
function of bone marrow adipocytes. Compr Physiol. 8:315–349. 2017.
View Article : Google Scholar
|
|
8
|
Mauney J and Volloch V: Progression of
human bone marrow stromal cells into both osteogenic and adipogenic
lineages is differentially regulated by structural conformation of
collagen I matrix via distinct signaling pathways. Matrix Biol.
28:239–250. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Anderson TJ, Grégoire J, Pearson GJ, Barry
AR, Couture P, Dawes M, Francis GA, Genest J Jr, Grover S, Gupta M,
et al: 2016 Canadian cardiovascular society guidelines for the
management of dyslipidemia for the prevention of cardiovascular
disease in the adult. Can J Cardiol. 32:1263–1282. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shah SR, Werlang CA, Kasper FK and Mikos
AG: Novel applications of statins for bone regeneration. Natl Sci
Rev. 2:85–99. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin TK, Chou P, Lin CH, Hung YJ and Jong
GP: Long-term effect of statins on the risk of new-onset
osteoporosis: A nationwide population-based cohort study. PLoS One.
13:e01967132018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin TK, Liou YS, Lin CH, Chou P and Jong
GP: High-potency statins but not all statins decrease the risk of
new-onset osteoporotic fractures: A nationwide population-based
longitudinal cohort study. Clin Epidemiol. 10:159–165. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Moshiri A, Sharifi AM and Oryan A: Role of
Simvastatin on fracture healing and osteoporosis: A systematic
review on in vivo investigations. Clin Exp Pharmacol Physiol.
43:659–684. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jadhav SB and Jain GK: Statins and
osteoporosis: New role for old drugs. J Pharm Pharmacol. 58:3–18.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Maritz FJ, Conradie MM, Hulley PA, Gopal R
and Hough S: Effect of statins on bone mineral density and bone
histomorphometry in rodents. Arterioscler Thromb Vasc Biol.
21:1636–1641. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bajaj MS, Kulkarni RS, Ghode SS, Limaye LS
and Kale VP: Irradiation-induced secretion of BMP4 by marrow cells
causes marrow adipogenesis post-myelosuppression. Stem Cell Res.
17:646–653. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu M, Wang K, Tang T, Dai K and Zhu Z:
The effect of simvastatin on the differentiation of marrow stromal
cells from aging rats. Pharmazie. 64:43–48. 2009.PubMed/NCBI
|
|
18
|
Yuan Z, Li Q, Luo S, Liu Z, Luo D, Zhang
B, Zhang D, Rao P and Xiao J: PPARγ and Wnt signaling in adipogenic
and osteogenic differentiation of mesenchymal stem cells. Curr Stem
Cell Res Ther. 11:216–225. 2016. View Article : Google Scholar
|
|
19
|
Harsløf T, Tofteng CL, Husted LB, Nyegaard
M, Børglum A, Carstens M, Stenkjær L, Brixen K, Eiken P, Jensen JE,
et al: Polymorphisms of the peroxisome proliferator-activated
receptor γ (PPARγ) gene are associated with osteoporosis.
Osteoporos Int. 22:2655–2666. 2011. View Article : Google Scholar
|
|
20
|
Muruganandan S, Parlee SD, Rourke JL,
Ernst MC, Goralski KB and Sinal CJ: Chemerin, a novel peroxisome
proliferator-activated receptor gamma (PPARgamma) target gene that
promotes mesenchymal stem cell adipogenesis. J Biol Chem.
286:23982–23995. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mattern A, Zellmann T and Beck-Sickinger
AG: Processing, signaling, and physiological function of chemerin.
IUBMB Life. 66:19–26. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
De Henau O, Degroot GN, Imbault V, Robert
V, De Poorter C, Mcheik S, Galés C, Parmentier M and Springael JY:
Signaling properties of chemerin receptors CMKLR1, GPR1 and CCRL2.
PLoS One. 11:e01641792016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Helfer G and Wu QF: Chemerin: A
multifaceted adipokine involved in metabolic disorders. J
Endocrinol. 238:R79–R94. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kennedy AJ and Davenport AP: International
union of basic and clinical pharmacology CIII: Chemerin receptors
CMKLR1 (Chemerin1) and GPR1 (Chemerin2) nomenclature, pharmacology,
and function. Pharmacol Rev. 70:174–196. 2018. View Article : Google Scholar :
|
|
25
|
Zhao H, Yan D, Xiang L, Huang C, Li J, Yu
X, Huang B, Wang B, Chen J, Xiao T, et al: Chemokine-like receptor
1 deficiency leads to lower bone mass in male mice. Cell Mol Life
Sci. 76:355–367. 2019. View Article : Google Scholar
|
|
26
|
Ramos-Junior ES, Leite GA, Carmo-Silva CC,
Taira TM, Neves KB, Colón DF, da Silva LA, Salvador SL, Tostes RC,
Cunha FQ and Fukada SY: Adipokine chemerin bridges metabolic
dyslipidemia and alveolar bone loss in mice. J Bone Miner Res.
32:974–984. 2017. View Article : Google Scholar
|
|
27
|
Muruganandan S, Govindarajan R, McMullen
NM and Sinal CJ: Chemokine-like receptor 1 is a novel Wnt target
gene that regulates mesenchymal stem cell differentiation. Stem
Cells. 35:711–724. 2017. View Article : Google Scholar
|
|
28
|
Muruganandan S, Roman AA and Sinal CJ:
Role of chemerin/CMKLR1 signaling in adipogenesis and
osteoblastogenesis of bone marrow stem cells. J Bone Miner Res.
25:222–234. 2010. View Article : Google Scholar
|
|
29
|
Goralski KB, McCarthy TC, Hanniman EA,
Zabel BA, Butcher EC, Parlee SD, Muruganandan S and Sinal CJ:
Chemerin, a novel adipokine that regulates adipogenesis and
adipocyte metabolism. J Biol Chem. 282:28175–28188. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
An Q, Wu D, Ma Y, Zhou B and Liu Q:
Suppression of Evi 1 promotes the osteogenic differentiation and
inhibits the adipogenic differentiation of bone marrow-derived
mesenchymal stem cells in vitro. Int J Mol Med. 36:1615–1622. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dominici M, Le Blanc K, Mueller I,
Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A,
Prockop DJ and Horwitz E: Minimal criteria for defining multipotent
mesenchymal stromal cells. The international society for cellular
therapy position statement. Cytotherapy. 8:315–317. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Schmittgen TD: Real-time quantitative PCR.
Methods. 25:383–385. 2001. View Article : Google Scholar
|
|
33
|
Bedi O, Dhawan V, Sharma PL and Kumar P:
Pleiotropic effects of statins: New therapeutic targets in drug
design. Naunyn Schmiedebergs Arch Pharmacol. 389:695–712. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Roh SG, Song SH, Choi KC, Katoh K,
Wittamer V, Parmentier M and Sasaki S: Chemerin-a new adipokine
that modulates adipogenesis via its own receptor. Biochem Biophys
Res Commun. 362:1013–1018. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gao K, Shen Z, Yuan Y, Han D, Song C, Guo
Y and Mei X: Simvastatin inhibits neural cell apoptosis and
promotes locomotor recovery via activation of Wnt/β-catenin
signaling pathway after spinal cord injury. J Neurochem.
138:139–149. 2016. View Article : Google Scholar :
|
|
36
|
Robin NC, Agoston Z, Biechele TL, James
RG, Berndt JD and Moon RT: Simvastatin promotes adult hippocampal
neurogenesis by enhancing Wnt/β-catenin signaling. Stem Cell
Reports. 2:9–17. 2013. View Article : Google Scholar
|
|
37
|
Sawada N and Liao JK: Rho/Rho-associated
coiled-coil forming kinase pathway as therapeutic targets for
statins in atherosclerosis. Antioxid Redox Signal. 20:1251–1267.
2014. View Article : Google Scholar :
|
|
38
|
Rikitake Y and Liao JK: Rho GTPases,
statins, and nitric oxide. Circ Res. 97:1232–1235. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rourke JL, Dranse HJ and Sinal CJ: CMKLR1
and GPR1 mediate chemerin signaling through the RhoA/ROCK pathway.
Mol Cell Endocrinol. 417:36–51. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tang QQ, Otto TC and Lane MD: Mitotic
clonal expansion: A synchronous process required for adipogenesis.
Proc Natl Acad Sci USA. 100:44–49. 2003. View Article : Google Scholar
|
|
41
|
Zanette DL, Lorenzi JC, Panepucci RA,
Palma PV, Dos Santos DF, Prata KL and Silva WA Jr: Simvastatin
modulates mesenchymal stromal cell proliferation and gene
expression. PLoS One. 10:e01201372015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li Y, Müller AL, Ngo MA, Sran K, Bellan D,
Arora RC, Kirshenbaum LA and Freed DH: Statins impair survival of
primary human mesenchymal progenitor cells via mevalonate
depletion, NF-κB signaling, and Bnip3. J Cardiovasc Transl Res.
8:96–105. 2015. View Article : Google Scholar
|
|
43
|
Weivoda MM and Hohl RJ: Geranylgeranyl
pyrophosphate stimulates PPARγ expression and adipogenesis through
the inhibition of osteoblast differentiation. Bone. 50:467–476.
2012. View Article : Google Scholar
|
|
44
|
Nicholson AC, Hajjar DP, Zhou X, He W,
Gotto AM Jr and Han J: Anti-adipogenic action of pitavastatin
occurs through the coordinate regulation of PPARgamma and Pref-1
expression. Br J Pharmacol. 151:807–815. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Rourke JL, Muruganandan S, Dranse HJ,
McMullen NM and Sinal CJ: Gpr1 is an active chemerin receptor
influencing glucose homeostasis in obese mice. J Endocrinol.
222:201–215. 2014. View Article : Google Scholar : PubMed/NCBI
|