Introduction
Neuropathic pain is defined by the International
Association for the Study of Pain in 2011 as pain caused by a
lesion or disease of the somatosensory nervous system (1). Neuropathic pain is usually caused by
peripheral nerve injury and can lead to allodynia or hyperalgesia,
that is, the patient feels pain with weak or no stimulation
(2).
Studies have shown that dorsal root ganglion (DRG)
neurons are closely associated with neuropathic pain. Changes in
neurotrophic factors, inflammatory factors and expression of
various ion channels in DRG neurons result in increased
excitability of DRG neurons, which underlie the development of
neuropathic pain (3-7). Voltage-gated sodium channels (VGSCs
or Navs) have been classified into nine different
subtypes (Nav1.1-1.9), seven of which are expressed on
human DRG neurons (Nav1.1-1.3 and
Nav1.6-1.9). PCR analysis showed that Nav1.7
is primarily expressed in healthy human DRG neurons (8). Nav1.7 is closely
associated with neuropathic pain. Mutations in SCN9A, the gene
encoding Nav1.7, are associated with inherited primary
erythromelalgia, paroxysmal extreme pain disorder and idiopathic
small fiber neuropathies (9-11).
Hydrogen sulfide (H2S), has been reported
to be involved in numerous biological functions. Studies have shown
that H2S can enhance the sodium current density
(12,13), and inhibiting the endogenous
production of H2S mediated by the enzyme cystathionine
β-synthetase (CBS) with O-(carboxymethyl) hydroxylamine
hemihydrochloride (AOAA) can significantly reduce the sodium
current density carried by Nav1.7 and Nav1.8
in DRG neurons of rats (14). In
addition, mitogen-activated protein kinases (MAPKs) are transducers
of cell signaling that have been implicated in neuronal responses
to pathologies such as pain. Previous studies have shown that
expression of Nav1.7, Nav1.8 and
phosphorylated (p)-MAPKs, including p-extracellular
signal-regulated kinase (ERK) and p-p38, are significantly
increased in the blind-ending axon of the neuroma (15). Furthermore, p-ERK modulates the
gating kinetics of Nav1.7 through phosphorylation of
Nav1.7 at multiple sites (16).
The aim of the present study was to assess whether
endogenous H2S regulated Nav1.7 through the
MEK/ERK pathway in DRG neurons, and whether this may underlie the
pathogenesis of neuropathic pain.
However, there are few reports regarding the
relationship and role of endogenous H2S with
Nav1.7 (14). In the
present study, it was shown that endogenous H2S
activated the MEK/ERK pathway and upregulated the expression and
function of Nav1.7 in SNI rats, suggesting that
inhibiting endogenous H2S may provide a potentially
novel therapeutic method for treatment of neuropathic pain.
Materials and methods
Animals and groups
Male Sprague-Dawley rats, aged 8-12 weeks, and
weighing 180-200 g were used in the present study. Two or three
rats were housed in each cage and under a 12-h light/dark cycle,
and food and water were provided ad libitum. The present
study was approved by the Xinjiang Medical University Animal Center
(approval no. SCXK Xin2003-0001). Animal experiments were performed
in accordance with the Institutional Ethics Review Board at the
First Affiliated Hospital of the Shihezi University School of
Medicine. A total of 150 rats were randomly divided into five
groups: i) Sham group (n=30); ii) SNI+Vehicle group (n=30); iii)
SNI+AOAA group (n=30); iv) SNI+U0126 group (n=30); and iv)
SNI+PF-04856264 group (n=30). Rats were used for immunofluorescence
experiments (n=6 per group), western blot analysis (n=9 per group),
patch clamp experiments (n=3 per group) and nociceptive behavioral
studies (n=6 per group for thermal nociceptive tests and n=6 per
group for acetone experiments).
Reagents and instruments
The primary reagents used in the present study were
anti-Nav1.7 mouse monoclonal antibody (cat. no. ab85015;
Abcam), anti-Nav1.7 rabbit monoclonal antibody (cat. no.
orb447636; Biorbyt), anti-CBS rabbit polyclonal antibody (cat. no.
ab96252; Abcam), mouse anti-NF-200 (a marker for myelinated
A-fibers; cat. no. ab82259; Abcam), mouse anti-calcitonin gene
related peptide (CGRP; a marker of peptidergic C-type neurons;
ab81887; Abcam), rabbit anti-p-MEK (cat. no. 2338S; Cell Signaling
Technology, Inc.), rabbit anti-MEK (cat. no. 4694S; Cell Signaling
Technology, Inc.), rabbit anti-p-ERK (cat. no. 4370S; Cell
Signaling Technology, Inc.), rabbit anti-ERK (cat. no. 4695S; Cell
Signaling Technology, Inc.), mouse anti-GAPDH (cat. no. ab8245;
Abcam) and mouse anti-β-actin (cat. no. ab8226; Abcam). In
addition, the following secondary antibodies were used: Goat
anti-mouse antibody IgG H&L (HRP) (cat. no. 12-349;
Sigma-Aldrich; Merck KGaA), goat anti-rabbit antibody IgG H&L
(HRP) (cat. no. 12-348; Sigma-Aldrich; Merck KGaA), goat
anti-rabbit IgG H&L (TRITC) (cat. no. ab6718; Abcam), goat
anti-mouse IgG H&L (FITC) (cat. no. ab6785; Abcam), goat
anti-rabbit IgG H&L (FITC) (cat. no. ab6717; Abcam) and IB4
(FITC-conjugated; a marker for non-peptidergic C-type neurons; cat.
no. L2895; Sigma-Aldrich; Merck KGaA). During acute digestion of
DRG neurons, collagenase I (cat. no. C0130; Sigma-Aldrich; Merck
KGaA), trypsin (cat. no. T1426; cat. no. Sigma-Aldrich; Merck KGaA)
and trypsin inhibitor (cat. no. T6522; Sigma-Aldrich; Merck KGaA)
were used. PF-04856264 (Sigma-Aldrich; Merck KGaA), AOAA
(Sigma-Aldrich; Merck KGaA), dimethyl sulfoxide (DMSO;
Sigma-Aldrich; Merck KGaA), radio-immunoprecipitation assay buffer
(RIPA; Thermo Fisher Scientific, Inc.) and phenylmethylsulfonyl
fluoride (PMSF; Sigma-Aldrich; Merck KGaA) and DMEM (Gibco; Thermo
Fisher Scientific, Inc.) were also used in the present study.
The equipment used included an automatic thermal
radiation stimulator (37370 Plantar Test Apparatus; Ugo Basile
SRL), a BX51 fluorescence microscope (Carl Zeiss AG), a patch clamp
amplifier (MultiClamp 700B; Axon Instruments; Molecular Devices,
LLC), a microelectrode control instrument (P-2000; Sutter
Instrument Company), a biological data acquisition system (Digidata
1550A; Axon Instruments; Molecular Devices, LLC), a
micromanipulator (MP-225; Sutter Instrument Company) and an
inverted microscope (Olympus Corporation).
Establishment of neuropathic pain in
rats
Rats were anesthetized with 1% pentobarbital sodium
(50 mg/kg) by intraperitoneal (i.p.) injection, as described
previously (17). Following
anesthesia, three terminal branches of the sciatic nerve were
gently separated proximal to the popliteal fossa, and the tibial
and the common peroneal nerves were tightly ligated using a 4-0
silk and sectioned distal to the ligation, removing 3 mm of the
distal stumps at this level. Subsequently, the wound was closed in
layers. The rats were kept in a warm environment until complete
recovery from anesthesia. Following the operation, the rats were
subjected to single-cage feeding, enhanced animal nutrition,
infection prevention and a strict 12 h light/dark cycle. In the
Sham group, the sciatic nerve was isolated and exposed but not
injured.
Fluorescence probe for measuring
H2S
Rats were anesthetized with 1% pentobarbital sodium
(50 mg/kg, i.p.) and subsequently sacrificed by cervical
dislocation. The L4-L6 DRGs from the SNI
surgery side were then removed immediately. WSP-5 (cat. no. C3378;
APExBIO), a fluorescent probe for rapid detection of
H2S, was dissolved in D-Hanks balanced salt solution for
use. The samples were cut into pieces using eye scissors and
digestive enzymes (DMEM containing 0.24 mg/ml trypsin and 0.6 mg/ml
collagenase I) were used to acutely isolate DRG neurons at 37°C for
2-4 min, followed by centrifugation at 153 × g for 5 min at 4°C.
After removing the supernatant, the specimens were placed on a
96-well plate. Samples were incubated with WSP-5 (50 µm/l)
at 37°C for 30 min, and the specimens were cleaned with D-Hanks. A
fluorescence microscope (magnification, ×100) was used to record
the results. When the concentration of H2S in the cell
increased, the fluorescence intensity of WSP-5 increased
proportionally. Fluorescence intensity was analyzed using CellSens
Standard 2.1 Image Acquisition (Olympus Corporation) and Image Pro
Plus version 6.0 (Media Cybernetics).
Immunofluorescence experiments
As described previously (18), rats were anesthetized with 1%
pentobarbital sodium (50 mg/kg, i.p.) and perfused through the
aorta with normal saline followed by fresh 4% PFA in PBS for 10
min. L4, L5 and L6 DRGs were
removed rapidly and post-fixed in 4% PFA in PBS for 24 h at room
temperature, followed by dehydration in 20 or 30% sucrose in
phosphate buffer at 4°C. DRGs were embedded in paraffin and
sectioned (5-µm) using a freezing microtome. Sections were
washed and incubated with blocking buffer (4% BSA in PBS with 0.2%
Tween-20) at room temperature for 30 min. Subsequently, sections
were rinsed with 0.01 M PBS three times and incubated with mouse
anti-Nav1.7 antibody (1:100) or rabbit anti-CBS antibody
(1:100) in a wet box overnight at 4°C. Next, FITC-conjugated goat
anti-mouse secondary antibodies (1:200) and FITC-conjugated goat
anti-rabbit secondary antibodies (1:200) were incubated with the
sections at room temperature for 120 min.
In the co-expression experiments, sections were
incubated with a mixture of primary rabbit anti-Nav1.7
antibody (1:100) and mouse anti-NF-200 (1:100)/mouse anti-CGRP
(1:100) in a wet box overnight at 4°C. Next, the sections were
rinsed with 0.01 M PBS three times, and then incubated with a
mixture of the TRITC-conjugated goat anti-rabbit secondary antibody
(1:200) and FITC-conjugated goat anti-mouse secondary antibody
(1:200) in the dark for 120 min at room temperature. However, in
order to detect the co-expression of Nav1.7 and IB4 on
DRG neurons, the tissue sections were incubated only with the
primary antibody rabbit anti-Nav1.7 antibody (1:100) overnight at
4°C, followed by rinsing three times, and incubation with IB4
(1:500) mixed with TRITC-conjugated goat anti-rabbit secondary
antibody (1:200) at room temperature for 120 min in the dark.
Immunofluorescence quantification for all target proteins was
performed by measuring the mean absorbance following laser confocal
microscopy (magnification, ×200; Carl Zeiss AG) and analyzed with
ZEN 2009 Light Edition (Carl Zeiss AG).
Western blot analysis
The L4-L6 DRGs from each group
were homogenized in RIPA buffer (1 µl PMSF to 100 µl
RIPA buffer) with freshly added protease inhibitor (10 mg tissue to
100 µl RIPA buffer), and centrifuged at 15,294 x g for 15
min at 4°C. The protein concentration was determined using the BCA
method. Protein aliquots (40 µg/lane) were separated using a
Tris-glycine denaturing gradient gel electrophoresis on an 8%
SDS-gel using SDS-PAGE. The proteins were then transferred to a
PVDF membrane (EMD Millipore), blocked with 5% non-fat milk in TBST
buffer (pH 8.0, 10 mmol/l Tris-HCl, 150 mmol/l NaCl and 0.2%
Tween-20) for 2 h at room temperature, and then incubated with the
following primary antibodies in a box overnight at 4°C: Rabbit
anti-Nav1.7 monoclonal antibody (1:1,000), rabbit
anti-CBS polyclonal antibody (1:1,000), rabbit anti-p-MEK
(1:1,000), rabbit anti-MEK (1:1,000), rabbit anti-p-ERK (1:1,000),
rabbit anti-ERK (1:1,000), mouse anti-GAPDH (1:1,000) or mouse
anti-β-actin (1:1,000). Following incubation with primary
antibodies, the PVDF membrane was washed three times with TBST for
5 min and incubated with horseradish peroxidase-conjugated goat
anti-mouse antibody IgG H&L (HRP) (1:10,000) or goat
anti-rabbit antibody IgG H&L (HRP) (1:10,000) for 2 h at room
temperature. Protein levels were normalized to β-actin or GAPDH
using Image J 1.43 (National Institutes of Health).
Acute separation of DRG neurons
Rats were anesthetized with 1% sodium pentobarbital
(50 mg/kg, i.p.) and then sacrificed by cervical dislocation.
Subsequently, the segmental L4-L6 DRG
organization were removed immediately. The redundant nerve fibers
and connective tissues were amputated. Next, the ganglia were cut
using eye scissors, and gently dissociated using a fire-polished
Pasteur pipette and treatment with digestive enzymes (DMEM
containing 0.24 mg/ml trypsin and 0.6 mg/ml collagenase I) at 37°C
for 2-4 min. Incubation with trypsin inhibitors was then performed
at room temperature for 15 min to terminate digestion.
Extracellular fluid contained: 150 mM NaCl, 5 mM KCl, 2.5 mM
CaCl2, 1 mM MgCl2·H2O, 10 mM HEPES
and 10 mM D-glucose, and was adjusted to pH 7.4 with NaOH.
Extracellular fluid was added to the neuron suspension. The
solution osmotic pressure of the extracellular fluid was 300-330
mOsm. The neurons suspension was placed in a 35 mm cell culture
dish and used for electrophysiological recording once the neurons
had adhered to the wall.
Patch clamp recording
A borosilicate glass blank with core produced by
Sutter Instrument Company was transformed into a microelectrode
using a P-2000 electrode drawing instrument. Microelectrodes with
resistance values between 2-8 MΩ were used for current-clamp
recording. The pipette solution contained the following: 130 mM
K-gluconate, 10 mM NaCl, 1.2 mM MgCl2·H2O, 2
mM CaCl2, 5 mM EGTA, 10 mM HEPES and 7.5 mM D-glucose,
adjusted to pH 7.35 with KOH, and 300-330 mOsm with sucrose. The
extracellular solution contained: 140 mM NaCl, 5 mM KCl, 2 mM
CaCl2, 2 mM MgCl2·H2O, 10 mM HEPES
and 11 mM glucose, and was adjusted to pH 7.4 with NaOH and 315
mOsm with sucrose. In the voltage clamp mode, the pipette solution
contained the following: 140 mM CsF, 10 mM NaCl, 1 mM EGTA and 10
mM HEPES, adjusted to pH 7.3 with CsOH and 315 mOsm with sucrose.
The extracellular bath solution contained the following: 70 mM
NaCl, 70 mM choline chloride, 3 mM KCl, 1 mM
MgCl2·H2O, 1 mM CaCl2, 10 mM
HEPES, 5 mM CsCl, 20 mM tetraethylammonium chloride and 0.1 mM,
CdCl2, was adjusted to pH 7.32 with NaOH and 327 mOsm
with sucrose. The inverted microscope was used to observe the state
of DRG neurons, and the micromanipulator was used to slowly bring
the microelectrode into contact with DRG neurons under the
microscope. A GΩ seal was formed between the cells and the
electrode. After the membrane was broken, cells with a resistance
above GΩ were used for experiments. In the voltage clamp mode, the
holding potential used was -60 mV; a step protocol (from -100 to 20
mV in 10 mV increments with a pulse duration of 100 msec) was
applied to determine the Nav1.7 current. In the current
clamp mode, a step protocol (from 0 to 600 pA in 50 pA increments
with a pulse duration of 150 msec) was applied to determine the
rheobase (the minimum current intensity required to excite the
first action potential). Subsequently, another step protocol
(duration, 500 msec; amplitude, double-strength of rheobase) was
performed to record the number of action potentials. The recorded
signal was amplified by a MultiClamp 700B amplifier, filtered at
2.5 kHz, and converted by an Axon DigiData 1550A
digital-to-analog/analog-to-digital converter with a sampling
frequency of 10-20 kHz. And the 'net' current is obtained by
subtracting the 'administration' current from the 'no
administration' current, which represents the current sensitive to
this drug.
Nociceptive behavioral study
According to our previous study (18), the threshold responses to painful
thermal stimuli were evaluated on the surgical side hind paw at day
1 pre-operation and at days 1, 3, 7, 14 and 21 post-operation in
each group. Each rat was tested three times at intervals of 5 min,
and the mean of the three tests was obtained. Data are expressed as
thermal withdrawal latency in sec. As described previously
(19), acetone experiments were
performed on the plantar surface of the surgical side hind paw with
20 µl acetone by measuring the duration of paw withdrawal in
the 30 sec immediately following acetone application. These
experiments were evaluated 1 day before operation and 1, 3, 7, 14
and 21 days post-operation in each group. Each day, cold allodynia
measurements were repeated three times, and the mean was
calculated. The next stimulus was administered once the animal was
quiet, and data were expressed as the time that the paw was lifted
in sec.
Drug administration
AOAA, an inhibitor of CBS, was dissolved in normal
saline and immediately injected intrathecally into the animals (10
µg/kg body weight) 30 min before the nociceptive behavioral
study. An equivalent volume of normal saline was injected
intrathecally in the SNI+Vehicle group. For western blotting and
immunofluorescence experiments, AOAA was administered for 21
consecutive days from the day of surgery (14,20). In the patch clamp experiments,
AOAA (final concentration of 1 µm) was incubated with
acutely dissociated DRG neurons (modeling for 21 days) for 1 h. An
equivalent volume of normal saline was added during the incubation
in the SNI+Vehicle group.
U0126, an inhibitor of MEK, was freshly dissolved in
DMSO. For the nociceptive behavioral study, U0126 (10 µg
dissolved in 10 µl 10% DMSO) was administered intrathecally
30 min before the test. For western blotting and immunofluorescence
experiments, U0126 (10 µg dissolved in 10 µl 10%
DMSO) was administered intrathecally daily for 21 consecutive days
from the day of SNI surgery (20). For the patch clamp experiments,
U0126 (final concentration of 1 µm) was incubated with
acutely dissociated DRG neurons for 20 min (16).
PF-04856264, a specific inhibitor of
Nav1.7, was freshly prepared in DMSO according to the
manufacturer's protocol. For the nociceptive behavioral study, an
i.p. injection of PF-04856264 (30 mg/kg body weight) was performed
10 min before the test (21). For
western blot analysis and immunofluorescence experiments,
PF-04856264 (30 mg/kg body weight) was administered intrathecally
daily for 21 consecutive days from the day of surgery. For the
patch clamp experiments, PF-04856264 (1 µm) was incubated
with acutely dissociated DRG neurons for 1 h (22).
Statistical analysis
Data are presented as the mean ± standard error of
the mean of three independent experiments. SPSS version 17.0 (SPSS,
Inc.) was used for statistical analysis. Firstly, the distribution
of the data was determined using a Shapiro-Wilk's or One-Sample
Kolmogorov-Smirnov test. Variables with normal distribution were
analyzed using ANOVA followed by Bonferroni's post hoc test.
Whereas variables not normally distributed were analyzed using a
Friedman test or Kruskal-Wallis test with a Tukey's post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
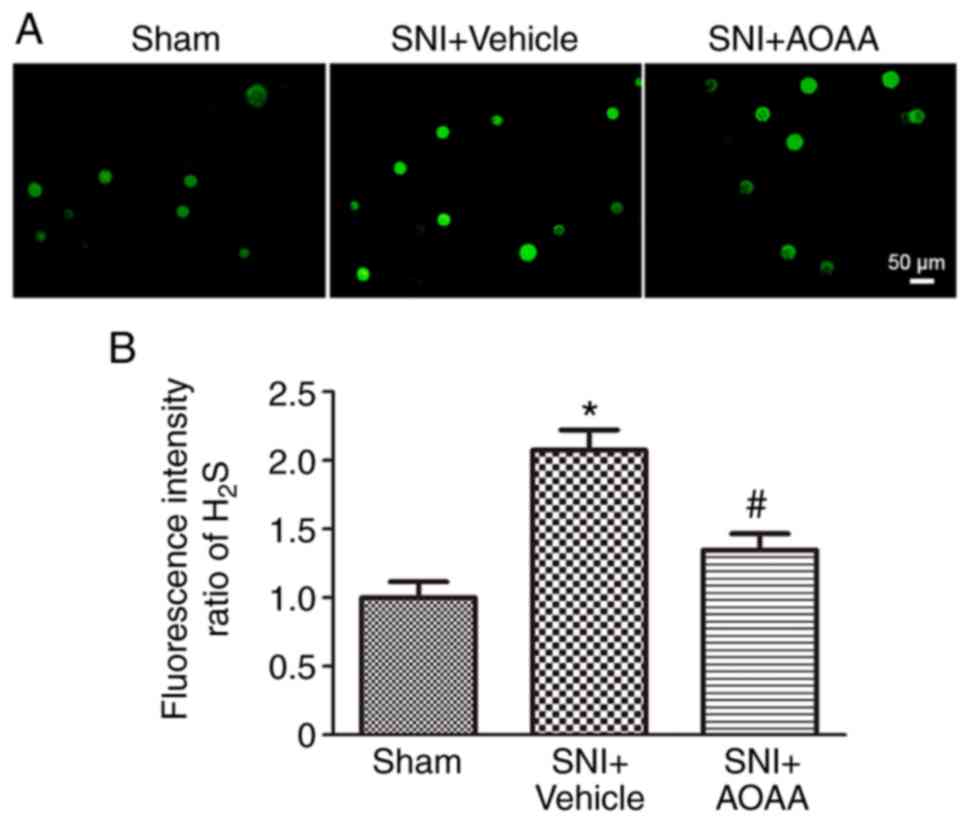
Detection of H2S concentration
changes in DRG neurons using a chlorine ion fluorescence probe
Firstly, DRG neurons were acutely dissociated and
WSP-5 was used to detect the concentration of H2S in DRG
neurons. The intensity of green fluorescence is directly
proportional to the concentration of H2S in DRG neurons.
The experiment evaluated changes in the intracellular
H2S concentrations of the L4-L6
DRG neurons on the surgical side of rats in the Sham group,
SNI+Vehicle group and SNI+AOAA group. The results demonstrated that
the concentration of H2S in L4-L6
DRG neurons in the SNI+Vehicle group was significantly increased
(P<0.05) compared with the Sham group. In addition,
compared with the SNI+Vehicle group, the concentration of
H2S in the SNI+AOAA group was significantly decreased
(P<0.05; Fig. 1). This
indicates that the change of H2S in DRG is closely
associated with pain. However, acute isolation may affect the
activity of DRG neurons and the concentration of H2S in
neurons, thus affecting the intensity of the H2S
fluorescence.
Expression of Nav1.7 and CBS
in DRG neurons of SNI rats
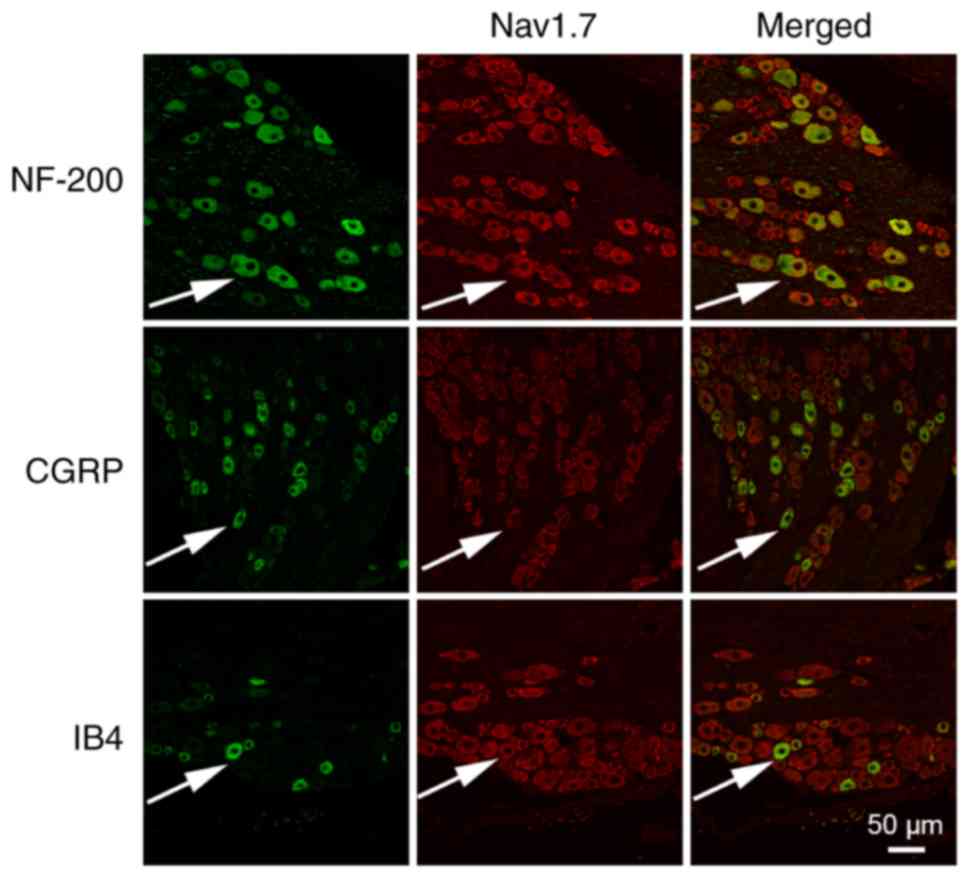
Immunofluorescence was performed to observe the
expression of Nav1.7 in the DRG neurons. The results
revealed that Nav1.7 was expressed in different types of
DRG neurons, including NF-200-marked DRG neurons, CGRP-marked DRG
neurons and IB4-marked DRG neurons (Fig. 2).
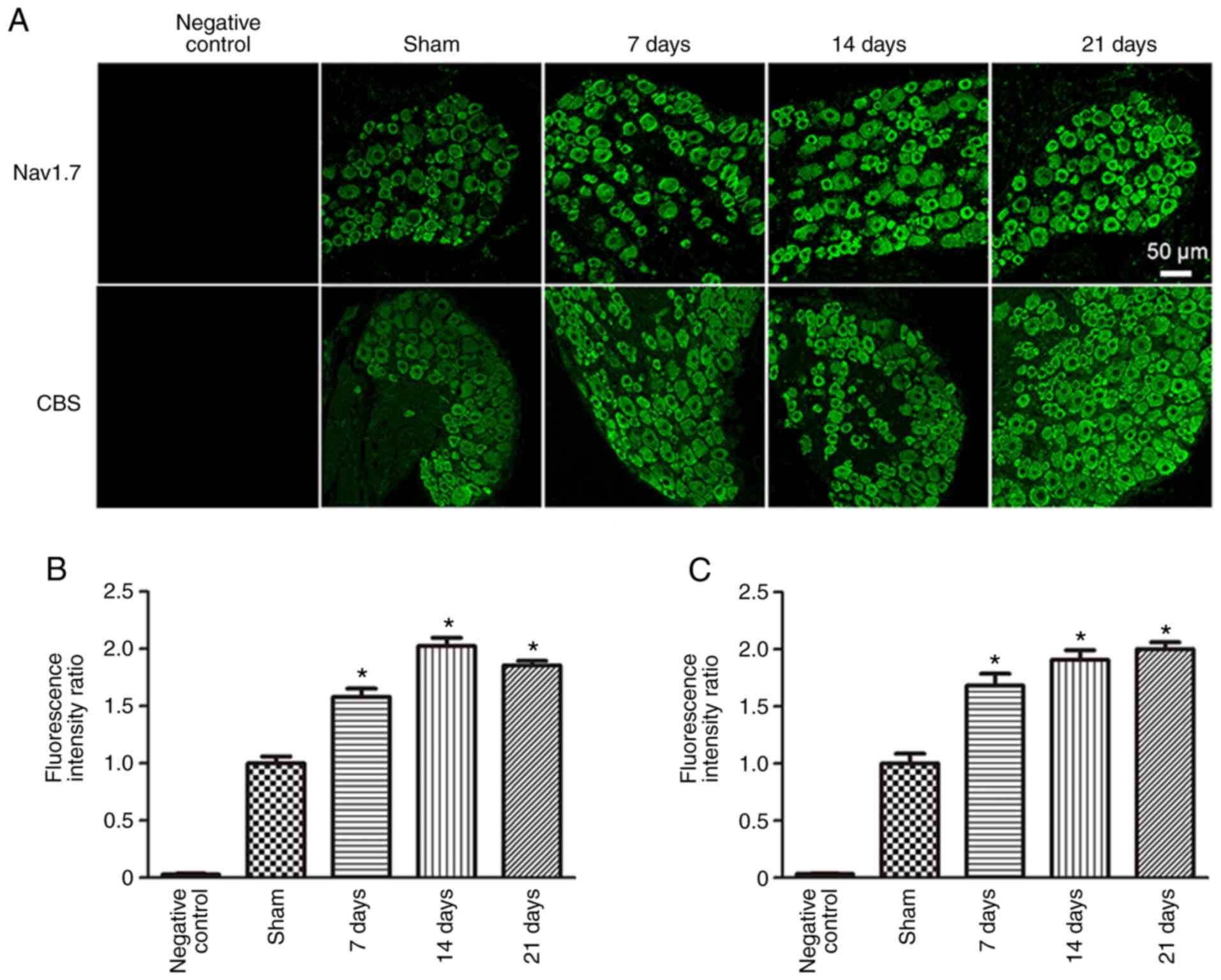
To observe changes in both Nav1.7 and CBS
protein expression levels in the DRG neurons of rats following SNI
surgery, immunofluorescence and western blotting were used. Both
Nav1.7 and CBS were expressed in DRG neurons. Compared
with the Sham group (1.00±0.06), the fluorescence intensity ratio
of Nav1.7 was significantly increased 7 (1.58±0.07;
P<0.05), 14 (2.03±0.07; P<0.05) and 21 days
(1.85±0.04; P<0.05) post-SNI surgery (Fig. 3). In addition, compared with the
Sham group (1.0±0.09), the fluorescence intensity ratio of CBS was
also significantly increased 7 (1.68±0.10; P<0.05), 14
(1.91±0.08; P<0.05) and 21 days (2.00±0.06;
P<0.05) post-SNI surgery.
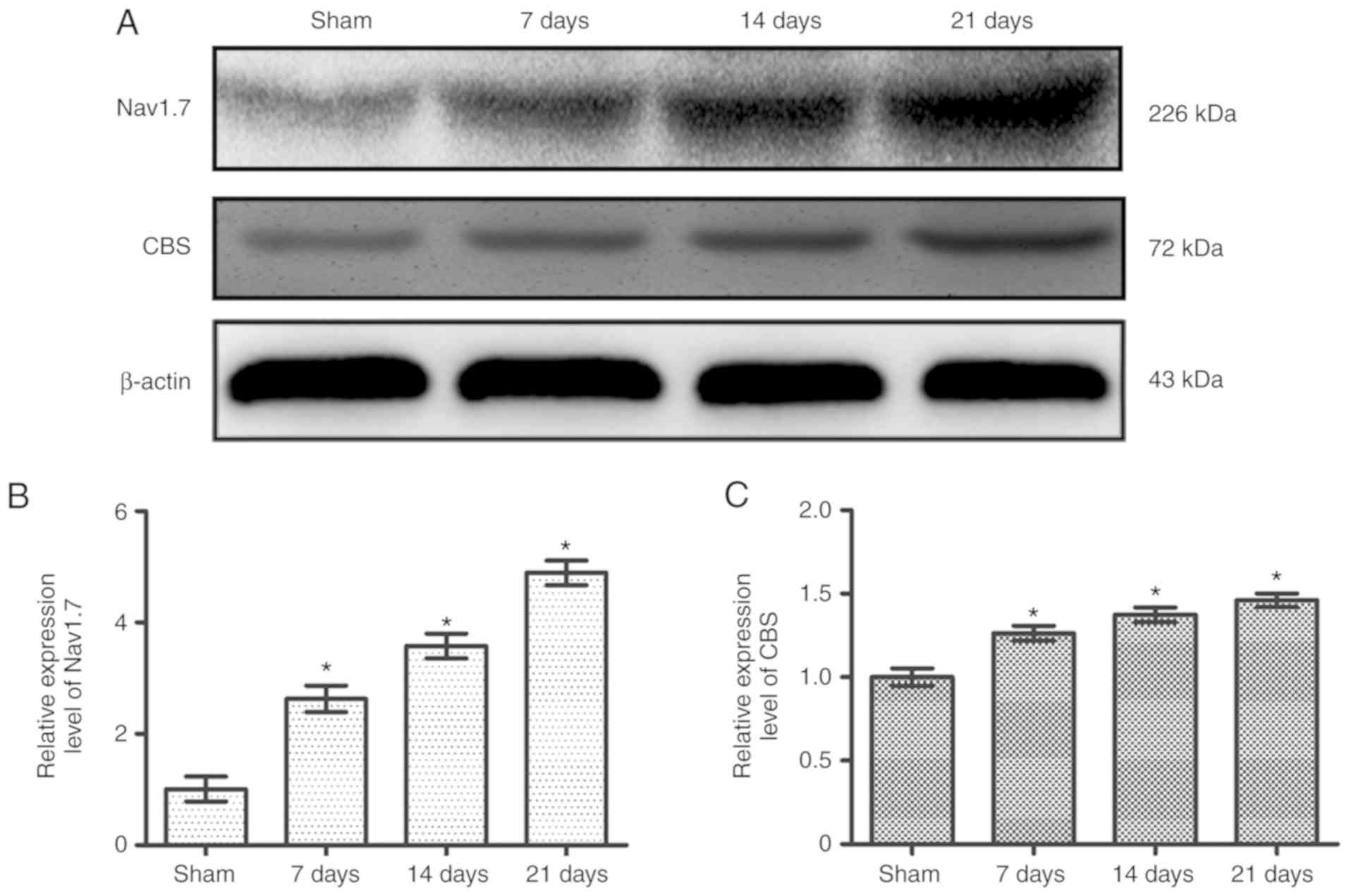
Compared with the Sham group (1.00±0.22), the
relative expression level of Nav1.7 was significantly
increased at 7 (2.63±0.24; P<0.05), 14 (3.58±0.22;
P<0.05) and 21 days (4.88±0.22; P<0.05)
post-SNI surgery (Fig. 4A and B).
Furthermore, compared with the Sham group (1.0±0.05), the relative
expression level of CBS was also significantly increased at 7
(1.26±0.04; P<0.05), 14 (1.37±0.04; P<0.05) and
21 days (1.46±0.04; P<0.05) post-SNI surgery (Fig. 4A and C). The changes observed were
most prominent after 21 days; therefore, in all subsequent
experiments, measurements were taken 21 days after surgery.
Endogenous H2S upregulates the
expression of Nav1.7 by activating the MEK/ERK
pathway
To investigate how H2S regulates the
expression of Nav1.7 in DRG neurons of rats following
SNI surgery, western blotting was performed following treatment
with AOAA, U0126 and PF-04856264 (Fig. 5A).
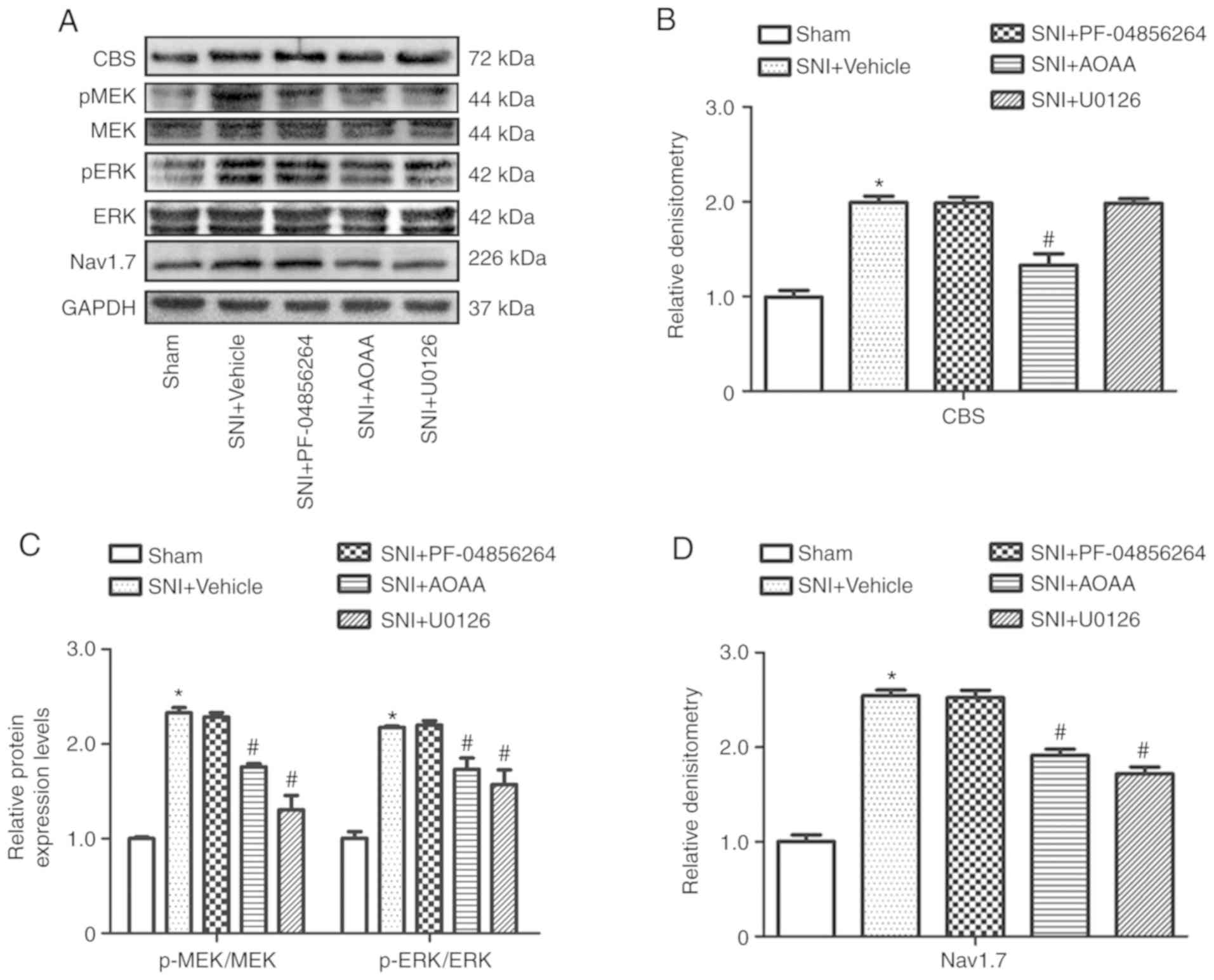 | Figure 5Changes in protein expression levels
following the administration of AOAA, U0126 or PF-04856264 in
dorsal root ganglions of SNI rats. (A) Expression levels of CBS,
MEK, p-MEK, ERK, p-ERK, Nav1.7 and GAPDH in each group.
Relative expression of (B) CBS, (C) p-MEK/MEK, p-ERK/ERK and (D)
Nav1.7 in each group. *P<0.05 vs. Sham
group. #P<0.05 vs. SNI+Vehicle group. n=6 per group.
AOAA, O-(carboxymethyl) hydroxylamine hemihydrochloride; SNI,
spared nerve injury; MEK, mitogen-activated protein kinase kinase;
ERK, extracellular signal-regulated kinase; p, phosphorylated;
cystathionine β-synthetase; Nav1.7, voltage-gated sodium channel
1.7. |
Compared with the Sham group (1.0±0.04), the
relative expression level of CBS (1.99±0.04; P<0.05) was
increased significantly in the SNI+Vehicle group. Compared with the
SNI+Vehicle group (1.99±0.04), the relative expression of CBS was
reduced significantly in the SNI+AOAA group (1.33±0.07;
P<0.05). However, there was no significant difference in
the relative expression of CBS following administration of U0126
(1.98±0.03; P>0.05) and PF-04856264 (1.99±0.04; P>0.05;
Fig. 5B).
Compared with the Sham group, the relative ratios of
p-MEK/MEK and p-ERK/ERK were significantly increased in DRG neurons
after SNI surgery (P<0.05). In addition, compared with the
SNI+Vehicle group, the relative ratios of p-MEK/MEK and p-ERK/ERK
were reduced significantly in the SNI+AOAA and SNI+U0126 groups
(P<0.05). However, there were no significant differences
in relative ratios of p-MEK/MEK and p-ERK/ERK following
administration of PF-04856264 (P>0.05; Fig. 5C).
Compared with the Sham group (1.0±0.04), the
relative expression of Nav1.7 was increased
significantly in the SNI+Vehicle group (2.54±0.03;
P<0.05). Compared with the SNI+Vehicle group (2.54±0.03),
the relative expression level of Nav1.7 was reduced
significantly in the SNI+AOAA group (1.91±0.04; P<0.05).
In addition, the relative expression of Nav1.7 was also
reduced significantly in the SNI+U0126 group (1.72±0.04;
P<0.05; Fig. 5D).
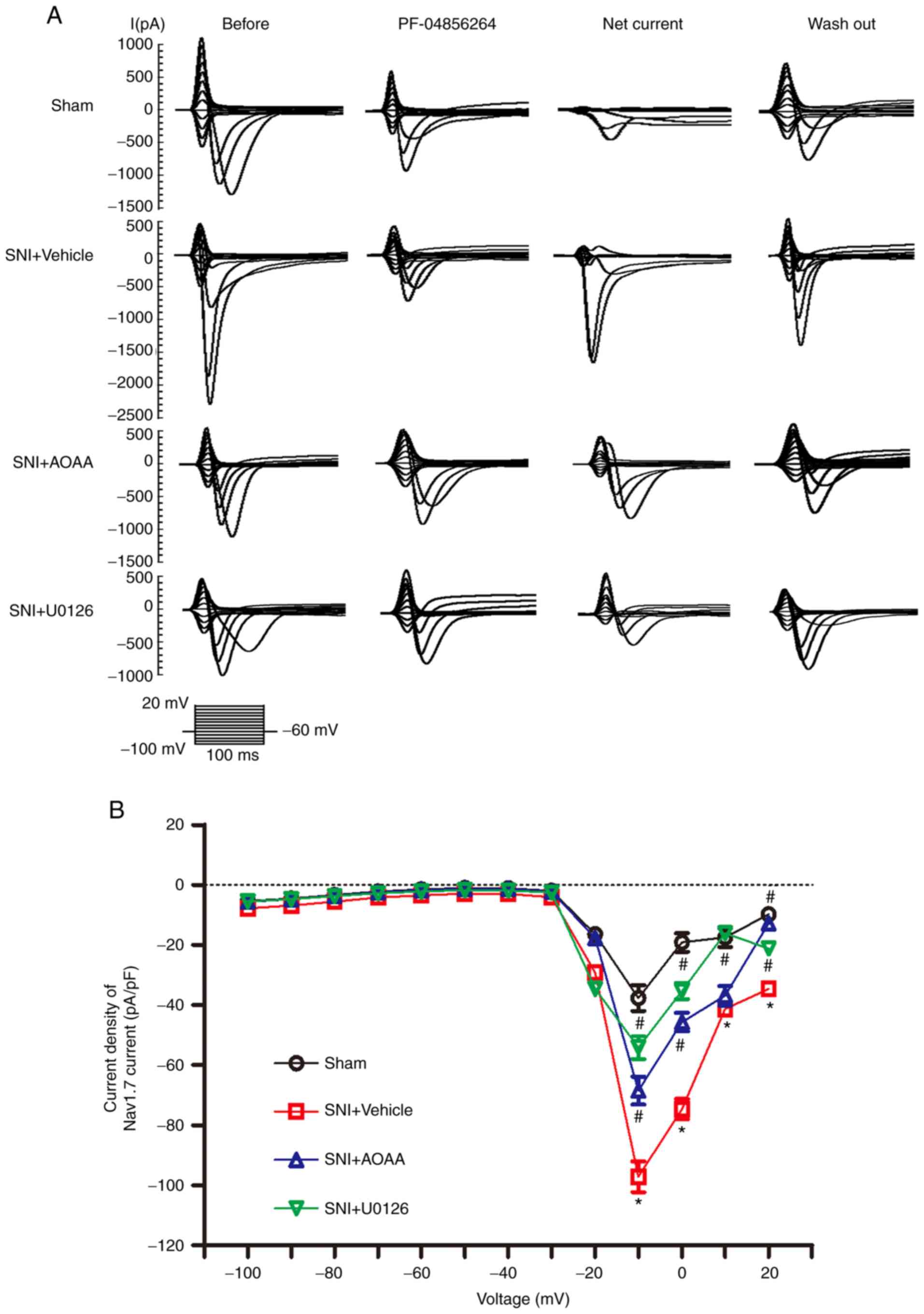
Endogenous H2S upregulates
Nav1.7 expression by activating the MEK/ERK pathway and
increasing the Nav1.7-carried current density
To investigate whether endogenous H2S
upregulates Nav1.7 expression via the MEK/ERK signaling
pathway and increases the sodium current carried by
Nav1.7 in DRG neurons, patch-clamp experiments were
performed. The 'Net' current was obtained by subtracting the
PF-04856264 from the 'Before' current, which represents the current
sensitive to PF-04856264, and the Net current was taken to
represent the current carried by Nav1.7 (Fig. 6A). For a more accurate measure of
the current 'felt' by the cells, the current density was
calculated.
At the voltage of -10 mV, compared with the Sham
group (-37.69±4.31), the current density of Nav1.7 was
significantly increased in the SNI+Vehicle group (-97.19±5.14;
P<0.05). Compared with the SNI+Vehicle group,
administration of AOAA (-68.53±4.66; P<0.05) and U0126
(-54.18±3.78; P<0.05) significantly decreased the current
density of Nav1.7 (Fig.
6B). At the voltage of 0 mV, the current density of
Nav1.7 was significantly increased in the SNI+Vehicle
group (-74.71±3.38) compared with Sham group (-19.18±3.06;
P<0.05). Compared with the SNI+Vehicle group,
administration of AOAA (-45.69±3.07; P<0.05) and U0126
(-35.40±2.69; P<0.05) significantly decreased the current
density of Nav1.7. Furthermore, at the voltage of 20 mV,
the current density of Nav1.7 was significantly
increased in the SNI+Vehicle group (-34.71±1.45) compared with the
Sham group (-9.79±1.58; P<0.05). Compared with the
SNI+Vehicle group, administration of AOAA (-12.84±1.32;
P<0.05) and U0126 (-21.25±1.39; P<0.05)
significantly decreased the current density of Nav1.7.
At the voltage of 10 mV, the current density of Nav1.7
was significantly increased in the SNI+Vehicle group (-41.35±1.6)
compared with the Sham group (-17.47±3.16; P<0.05).
Compared with the SNI+Vehicle group, administration of U0126
(-16.10±1.74; P<0.05) decreased the current density of
Nav1.7. However, the current density of
Nav1.7 was not changed after administration of AAOA
(-36.93±3.22; P>0.05).
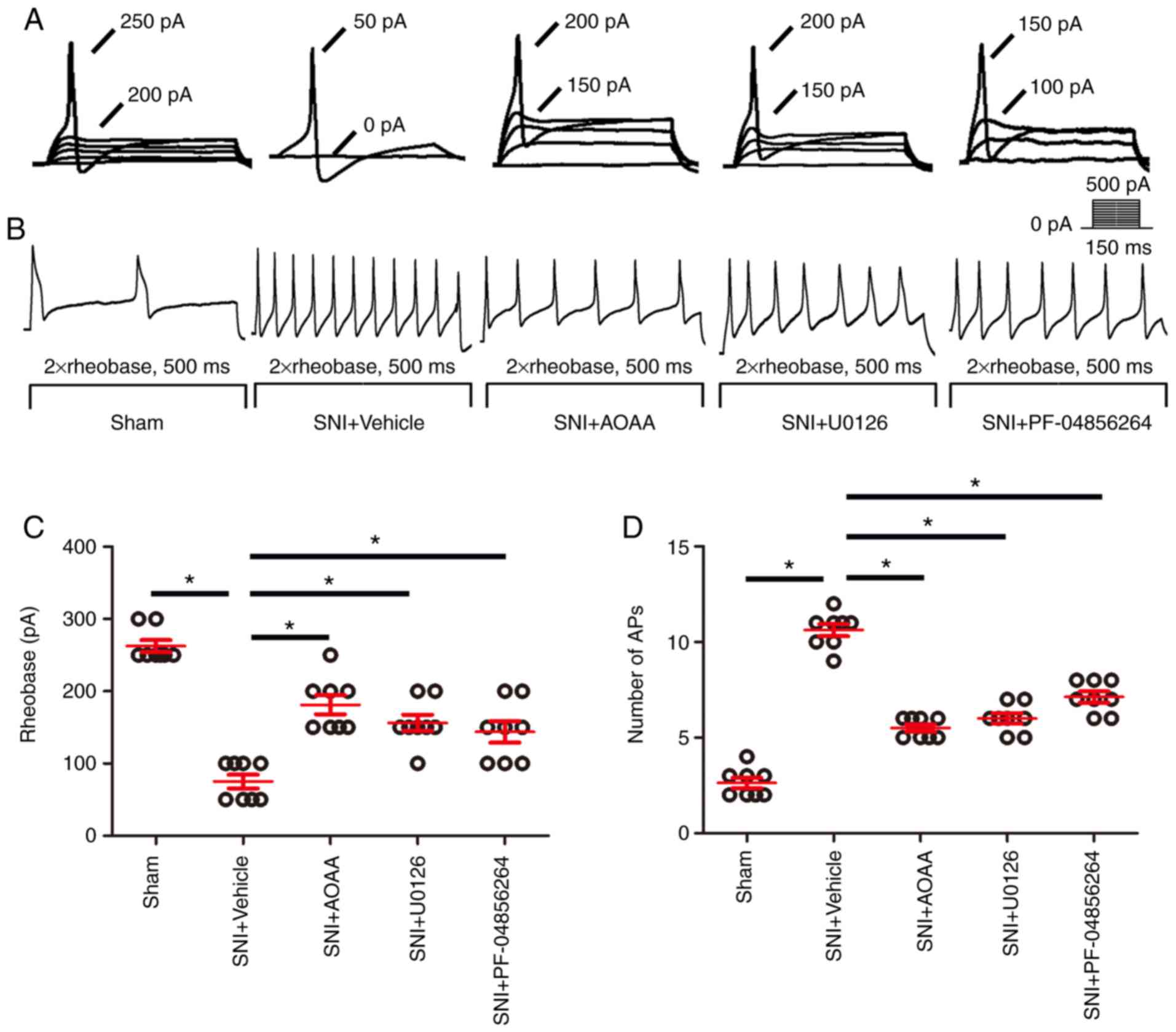
Endogenous H2S upregulates
Nav1.7 by activating the MEK/ERK signaling pathway,
resulting in increased excitability of DRG neurons
To investigate whether endogenous H2S
results in upregulated expression of Nav1.7 by the
MEK/ERK pathway and enhances the excitability of DRG neurons,
patch-clamp experiments were performed (Fig. 7). The rheobase (the minimum
current intensity required to excite the first action potential)
and the number of the action potentials were used as indicators of
the excitability of DRG neurons.
Compared with the Sham group (262.5±8.18), the
rheo-base of action potentials was significantly decreased in the
SNI+Vehicle group (75.00±9.45; P<0.05). Compared with the
SNI+Vehicle group, administration of AOAA (181.13±16.33), U0126
(156.25±11.33) and PF-04856264 (143.75±14.75) significantly
increased the rheobase of action potentials (P<0.05;
Fig. 7C).
Compared with the Sham group (2.63±0.26), the
number of action potentials was significantly increased in the
SNI+Vehicle group (10.63±0.32; P<0.05). Compared with the
SNI+Vehicle group, administration of AOAA (5.50±0.19), U0126
(6.0±0.27) and PF-04856264 (7.13±0.30) significantly decreased the
number of action potentials (P<0.05; Fig. 7D).
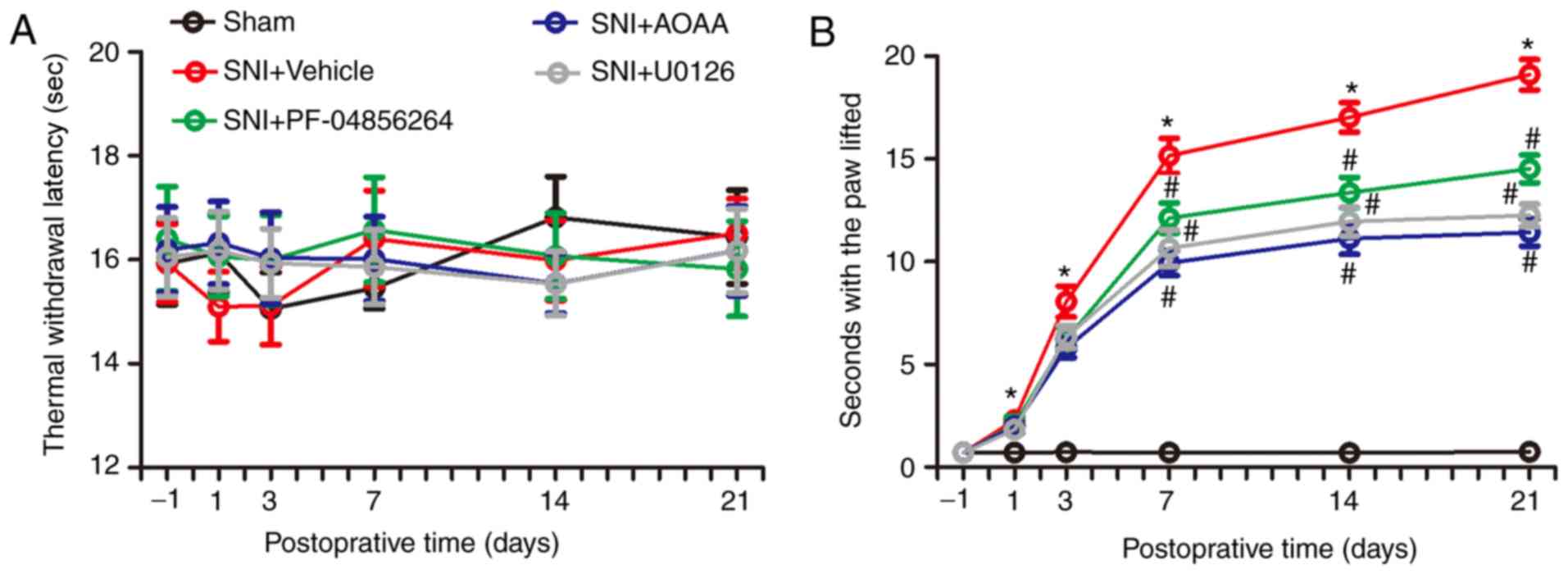
Endogenous H2S alters the
behavior of rats by upregulating expression of
Nav1.7
In order to investigate the changes in nociceptive
behavior of rats following SNI surgery, and to detect changes in
nociceptive behavior in SNI rats after administration of AOAA,
U0126 or PF-04856264, a Hargreaves test and acetone experiments
were performed. There were no significant differences between each
group in terms of thermal withdrawal latency (TWL) values (F=0.130;
P=0.9416; Fig. 8A). However, the
acetone experiments demonstrated that, compared with the Sham
group, the time animals spent with the paw lifted in the
SNI+Vehicle group increased significantly between days 1 and 21
(P<0.05). Compared with the SNI+Vehicle group, administration of
AOAA or U0126 significantly decreased the time with the paw lifted
between days 7 and 21 (P<0.05). In addition, compared with the
SNI+Vehicle group, administration of PF-04856264 significantly
decreased the time with the paw lifted between days 14 and 21
(P<0.05; Fig. 8).
Discussion
The primary findings of the present study were: i)
The concentration of H2S in DRG neurons was increased
after SNI modeling; ii) the expression levels of Nav1.7
and CBS were increased in DRG neurons of SNI rats; iii) endogenous
H2S upregulates the expression of Nav1.7 by
activating the MEK/ERK signaling pathway; iv) endogenous
H2S upregulates Nav1.7 by activating the
MEK/ERK pathway, resulting in increased Nav1.7 carried
current, and thus an increase in the excitability of DRG neurons;
and v) endogenous H2S alters the behavior of rats by
upregulating Nav1.7. Together, these results suggest
that endogenous H2S and Nav1.7 serve an
important role in neuropathic pain.
Previous studies have shown that Nav1.7,
one of the nine subtypes of VGSCs, serves an important role in
human pain disorders, including idiopathic small fiber
neuropathies, paroxysmal extreme pain disorder, inherited
erythromelalgia and congenital indifference to pain (23-26). Upon plantar incision surgery, the
expression of Nav1.7 in L4-L6 DRG
neurons of rats is increased, as well as the cumulative pain score.
Those changes are inhibited when rats are pretreated with SCN9A-RNA
interference lentivirus delivered via an intrathecal tube (4). He et al (25) confirmed that R1488*, a
variant of SCN9A, results in a complete loss-of-function of
Nav1.7, which is consistent with variants in this gene
in subjects with congenital insensitivity to pain. Geha et
al (27) demonstrated that
pharmacotherapy guided by genomic analysis, molecular modeling and
functional profiling attenuated neuropathic pain in patients
carrying an S241T Nav1.7 mutant channel. In the present
study, it was shown that Nav1.7 is expressed in
different types of DRG neurons (NF-200, CGRP and IB4) and the
expression of Nav1.7 was increased in
L4-L6 DRG neurons of SNI rats. Previous
studies have reported that there is a relationship between the
diameter of DRG neurons in rats and their excitatory typing, and
the excitability of small and medium-sized cells was higher
compared with that of small and medium-sized cells, indicating that
small and medium-sized cells play a more important role in the
generation of neuropathic pain (28,29). In addition, our previous study
(30) demonstrated that the
changes in excitatory typing of DRG neurons with different sizes
potentially explains the mechanisms of neuropathic pain, and after
SNI surgery the excitatory type of DRG neurons in rats changed,
with the proportion of type 1 and type 2 cells increased, but the
proportion of type 3 cells decreased. Therefore, neurons with
excitatory changes were selected to be recorded in the patch clamp
experiment. This result is consistent with the findings of a
previous study (30). In
addition, in the present study, the excitability of rat DRG neurons
increased, and rats developed cold allodynia following SNI surgery,
which was inhibited by the Nav1.7 specific blocker
PF-04856264.
An increasing number of studies have shown that
endogenous H2S has a variety of physiological functions,
including considerable support for a role of H2S as a
neuromodulator (31-33) or an endogenous gaseous transmitter
(34). Under physiological
conditions, H2S has been shown to regulate key neuronal
functions, including modulation of inward or outward currents on
dorsal raphe serotonergic neurons in vitro (35), or regulating the release of
corticotrophin-releasing hormone from the hypothalamus (36). H2S is an important
endogenous vasoactive factor and is a gaseous opener of
K+-ATP channels in vascular smooth muscle cells
(34). CBS and systathionine
γ-lyase (CSE) are two important enzymes involved in the generation
of endogenous H2S (37-41), which are expressed in the spinal
cord and colon, and detectable quantities of H2S are
produced by these tissues in the presence of L-cysteine, a CSE/CBS
substrate (42). CBS and CSE
expression have been observed in several mammalian tissues,
including liver, kidney, brain, ileum and blood lymphocytes
(34). In the cardiovascular
system, H2S is predominantly derived from CSE, and
modulates endothelium-dependent and endothelium-independent
vasodilatation (43), whereas
CBS-derived H2S is a physiologically relevant
neuromodulator in the central nervous system (CNS) (44). Consistent with this view, it has
been shown that H2S is present at relatively high levels
in the mammalian brain, and that in the CNS, the activity of CBS is
>30-fold greater than that of CSE (45). Xu et al (46) reported that CBS, but not CSE, is
expressed by colon-specific sensory neurons. Similarly, the
expression of CBS in L4-L6 DRG neurons was
also shown in the present immunofluorescence experiments. These
results suggest that the CBS-H2S pathway may serve an
important role in the nervous system. Previous studies have shown
that sodium channels, T-type calcium channels, transient receptor
potential cation channel subfamily V member 1 receptors, transient
receptor potential cation channel subfamily A member 1 receptors
and N-methyl-D-aspartic acid receptors are targets of
H2S-induced pain hypersensitivity (47-53). Yan et al (14) demonstrated that the expression of
CBS increased in DRG neurons in a lumbar disc herniation pain model
in rats, and that the administration of AOAA partially reversed
this change. In the present study, the expression of CBS increased
in DRG neurons of SNI rats. Furthermore, it has previously been
shown that the activity of sodium channels is enhanced by
upregulation of CBS expression in DRG neurons (54), and that treatment with AOAA
significantly suppresses the expression of Nav1.7 and
Nav1.8 in DRG neurons (14). Wang et al (55) reported that AOAA treatment
significantly hyperpolarized the resting potential of
colon-specific DRG neurons and enhanced the rheobase. The results
of the present study demonstrated that the concentration of
H2S in DRG neurons increased following SNI modeling and
the administration of AOAA partially reversed this change.
Furthermore, the increased expression of Nav1.7 and
enhanced excitability of DRG neurons induced by SNI surgery was
partially reversed by inhibiting CBS with AOAA. The reduced number
of action potentials and the increased rheobase may explain the
reversal in neuronal excitability following application of
AOAA.
H2S can regulate various physiological
and pathophysiological processes by regulating MAPK. Lan et
al (56) demonstrated that
H2S protects PC12 cells against chemical hypoxia-induced
injuries by inhibition of reactive oxygen species-mediated
activation of the ERK1/2 and p38-MAPK signaling pathways. Gobbi
et al (57) reported that
H2S impairs the growth and adhesion of keratinocyte
cells by inhibiting the Raf-MAPK-ERK signaling pathway.
Additionally, it has been shown that ERK serves an active role in
mediating H2S-induced apoptosis in human aorta smooth
muscle cells by activating caspase-3 (58). Stamboulian et al (16) reported that p-ERK phosphorylates
Nav1.7 and induces hyperpolarized activation and fast
inactivation of Nav1.7. U0126 inhibits the expression of
p-ERK (16,59). The ERK signaling cascade regulates
a variety of important cellular processes including growth,
proliferation and pain (60,61). Activated cAMP response
element-binding protein (CREB) is known to exhibit neuroprotective
and growth promoting effects (62). Sabbir and Fernyhough (63) reported that 1 h of treatment with
muscarinic toxin 7 and pirenzepine activates ERK through muscarinic
acetylcholine type 1 receptor and induces a significant increase in
levels of p-CREB in cultured sensory neurons. However, in the
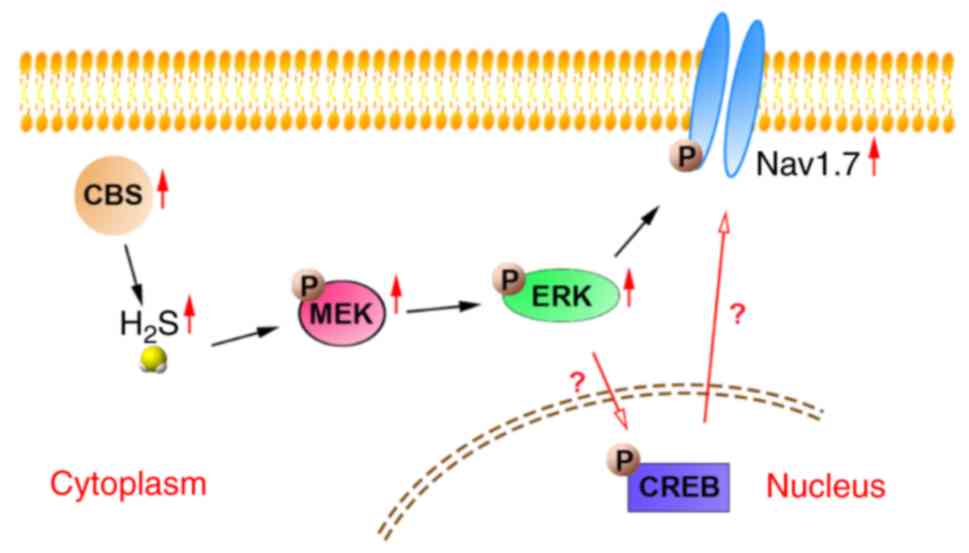
present study, it was demonstrated that p-MEK and p-ERK levels were
increased in DRG neurons of SNI rats, and these changes were
reversed by administration of AOAA, but not by administration of
PF-04856264. In addition, administration of U0126 inhibited the
increase in Nav1.7 expression, suggesting that
p-MEK/p-ERK regulated the expression of Nav1.7 in DRG
neurons of SNI rats (Fig. 9). The
process of p-ERK regulating p-CREB needs to be further evaluated in
future experiments. Furthermore, there may be a certain
relationship between p-CREB and Nav1.7 (Fig. 9), which needs to be further
researched in subsequent experiments.
In conclusion, endogenous H2S serves an
important role in the development and maintenance of neuropathic
pain by activating the MEK/ERK pathway to upregulate the
Nav1.7 channel. These results may assist in the
identification of novel drug targets for the treatment of
neuropathic pain. One limitation of the present study was that the
mechanisms underlying the increase in Nav1.7 were not
determined, and thus require further study.
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81560081).
Availability of data and materials
The datasets used and/or analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
JQS, LL and KTM conceived and designed the study,
implementation of the study and writing of the manuscript: JJT, CYT
and QYC performed the experiments and wrote the manuscript. WYS,
YZ, ZWQ and MZ analyzed and described the data. WYS, LL and KTM
were involved in critically revising the manuscript for important
intellectual content. All authors read and approved the final
manuscript and agree to be responsible for all aspects of the
study.
Ethics approval and consent to
participate
Animal experiments were approved by the
Institutional Ethics Review Board at the First Affiliated Hospital
of the Shihezi University School of Medicine (approval no.
A2017-169-01) and were performed in accordance with the Ethical
Guidelines for Investigations of Experimental Pain in Conscious
Animals of the International Association for the Study of Pain.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
This study was performed at the Department of
Physiology and the Key Laboratory of Xinjiang Endemic and Ethnic
Diseases of Xinjiang Provincial, Shihezi University School of
Medicine (Shihezi, China).
References
|
1
|
Jensen TS, Baron R, Haanpää M, Kalso E,
Loeser JD, Rice AS and Treede RD: A new definition of neuropathic
pain. Pain. 152:2204–2205. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bouhassira D, Lantéri-Minet M, Attal N,
Laurent B and Touboul C: Prevalence of chronic pain with
neuropathic characteristics in the general population. Pain.
136:380–387. 2008. View Article : Google Scholar
|
|
3
|
Zhang Y, Wang K, Lin M, Li Q and Hong Y:
Inhibition of morphine tolerance by MrgC receptor via modulation of
interleukin-1β and matrix metalloproteinase 9 in dorsal root
ganglia in rats. Eur J Pharmacol. 815:10–17. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sun J, Li N, Duan G, Liu Y, Guo S, Wang C,
Zhu C and Zhang X: Increased Na1.7 expression in the dorsal root
ganglion contributes to pain hypersensitivity after plantar
incision in rats. Mol Pain. 14:17448069187823232018. View Article : Google Scholar
|
|
5
|
Kawai H, Asaoka N, Miyake T, Nagayasu K,
Nakagawa T, Shirakawa H and Kaneko S: Neurotropin inhibits neuronal
activity through potentiation of sustained K currents in primary
cultured DRG neurons. J Pharmacol Sci. 137:313–316. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pachuau J and Martin-Caraballo M:
Expression pattern of T-type Ca (2+) channels in embryonic chick
nodose ganglion neurons. Dev Neurobiol. 67:1901–1914. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kitamura N, Nagami E, Matsushita Y, Kayano
T and Shibuya I: Constitutive activity of transient receptor
potential vanilloid type 1 triggers spontaneous firing in nerve
growth factor-treated dorsal root ganglion neurons of rats. IBRO
Rep. 5:33–42. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chang W, Berta T, Kim YH, Lee S, Lee SY
and Ji RR: Expression and role of voltage-gated sodium channels in
human dorsal root ganglion neurons with special focus on
Nav1.7, species differences, and regulation by
paclitaxel. Neurosci Bulletin. 34:4–12. 2018. View Article : Google Scholar
|
|
9
|
Koenig J, Werdehausen R, Linley JE, Habib
AM, Vernon J, Lolignier S, Eijkelkamp N, Zhao J, Okorokov AL, Woods
CG, et al: Regulation of Nav1.7: A conserved SCN9A
natural anti-sense transcript expressed in dorsal root ganglia.
PLoS One. 10:e01288302015. View Article : Google Scholar
|
|
10
|
Kim DT, Rossignol E, Najem K and Ospina
LH: Bilateral congenital corneal anesthesia in a patient with SCN9A
mutation, confirmed primary erythromelalgia, and paroxysmal extreme
pain disorder. J AAPOS. 19:478–479. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Faber CG, Hoeijmakers JG, Ahn HS, Cheng X,
Han C, Choi JS, Estacion M, Lauria G, Vanhoutte EK, Gerrits MM, et
al: Gain of function Naν1.7 mutations in idiopathic small fiber
neuropathy. Ann Neurol. 71:26–39. 2012. View Article : Google Scholar
|
|
12
|
Wang Y, Qu R, Hu S, Xiao Y, Jiang X and Xu
GY: Upregulation of cystathionine β-synthetase expression
contributes to visceral hyperalgesia induced by heterotypic
intermittent stress in rats. PLoS One. 7:e531652012. View Article : Google Scholar
|
|
13
|
Hu S, Xiao Y, Zhu L, Li L, Hu CY, Jiang X
and Xu GY: Neonatal maternal deprivation sensitizes voltage-gated
sodium channel currents in colon-specific dorsal root ganglion
neurons in rats. Am J Physiol Gastrointest Liver Physiol.
304:G311–G321. 2013. View Article : Google Scholar
|
|
14
|
Yan J, Hu S, Zou K, Xu M, Wang Q, Miao X,
Yu SP and Xu GY: Inhibition of cystathionine β-synthetase
suppresses sodium channel activities of dorsal root ganglion
neurons of rats with lumbar disc herniation. Sci Rep. 6:381882016.
View Article : Google Scholar
|
|
15
|
Ji RR and Strichartz G: Cell signaling and
the genesis of neuropathic pain. Sci STKE.
2004:reE142004.PubMed/NCBI
|
|
16
|
Stamboulian S, Choi JS, Ahn HS, Chang YW,
Tyrrell L, Black JA, Waxman SG and Dib-Hajj SD: ERK1/2
mitogen-activated protein kinase phosphorylates sodium channel
Na(v)1.7 and alters its gating properties. J Neurosci.
30:1637–1647. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Casals-Díaz L, Casas C and Navarro X:
Changes of voltage-gated sodium channels in sensory nerve
regeneration and neuropathic pain models. Restor Neurol Neurosci.
33:321–334. 2015.PubMed/NCBI
|
|
18
|
Zhang M, Gao CX, Wang YP, Ma KT, Li L, Yin
JW, Dai ZG, Wang S and Si JQ: The association between the
expression of PAR2 and TMEM16A and neuropathic pain. Mol Med Rep.
17:3744–3750. 2018.
|
|
19
|
Norcini M, Sideris A, Martin Hernandez LA,
Zhang J, Blanck TJ and Recio-Pinto E: An approach to identify
microRNAs involved in neuropathic pain following a peripheral nerve
injury. Front Neurosci. 8:2662014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu D, Wu X, Grabauskas G and Owyang C:
Butyrate-induced colonic hypersensitivity is mediated by
mitogen-activated protein kinase activation in rat dorsal root
ganglia. Gut. 62:1466–1474. 2013. View Article : Google Scholar
|
|
21
|
Deuis JR, Wingerd JS, Winter Z, Durek T,
Dekan Z, Sousa SR, Zimmermann K, Hoffmann T, Weidner C, Nassar MA,
et al: Analgesic effects of GpTx-1, PF-04856264 and CNV1014802 in a
mouse model of Nav17-mediated pain. Toxins (Basel). 8. pp. pii:
E782016, View Article : Google Scholar
|
|
22
|
Zhang H, Reichert E and Cohen AE: Optical
electrophysiology for probing function and pharmacology of
voltage-gated ion channels. ELife. 5:pii: e152022016. View Article : Google Scholar
|
|
23
|
Choi JS, Boralevi F, Brissaud O,
Sánchez-Martín J, Te Morsche RH, Dib-Hajj SD, Drenth JP and Waxman
SG: Paroxysmal extreme pain disorder: A molecular lesion of
peripheral neurons. Nat Rev Neurol. 7:51–55. 2011. View Article : Google Scholar
|
|
24
|
Cox JJ, Reimann F, Nicholas AK, Thornton
G, Roberts E, Springell K, Karbani G, Jafri H, Mannan J, Raashid Y,
et al: An SCN9A channelopathy causes congenital inability to
experience pain. Nature. 444:894–898. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
He W, Young GT, Zhang B, Cox PJ, Cho LT,
John S, Paciga SA, Wood LS, Danziger N, Scollen S and Vangjeli C:
Functional confirmation that the R1488* variant in SCN9A results in
complete loss-of-function of Na1.7. BMC Med Genet. 19:1242018.
View Article : Google Scholar
|
|
26
|
Cummins TR, Dib-Hajj SD and Waxman SG:
Electrophysiological properties of mutant Nav1.7 sodium
channels in a painful inherited neuropathy. J Neurosci.
24:8232–8236. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Geha P, Yang Y, Estacion M, Schulman BR,
Tokuno H, Apkarian AV, Dib-Hajj DS and Waxman SG: Pharmacotherapy
for pain in a family with inherited erythromelalgia guided by
genomic analysis and functional profiling. JAMA Neurol. 73:659–667.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fang X, Djouhri L, McMullan S, Berry C,
Waxman SG, Okuse K and Lawson SN: Intense isolectin-B4 binding in
rat dorsal root ganglion neurons distinguishes C-fiber nociceptors
with broad action potentials and high Nav1.9 expression.
J Neurosci. 26:7281–7292. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang J, Cavanaugh DJ, Nemenov MI and
Basbaum AI: The modality-specific contribution of peptidergic and
non-peptidergic nociceptors is manifest at the level of dorsal horn
nociresponsive neurons. J Physiol. 591:1097–1110. 2013. View Article : Google Scholar :
|
|
30
|
Tan CY, Ma KT, Si JQ, Zhou Y, Qu ZW, Zhang
M, Chen QY, Tian JJ, Xu ZZ and Deng SY: Type of excitability of DRG
neurons in rats with neuropathic pain. Chin J Mod Med. 29:1–7.
2019.
|
|
31
|
Abe K and Kimura H: The possible role of
hydrogen sulfide as an endogenous neuromodulator. J Neurosci.
16:1066–1071. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kimura H: Hydrogen sulfide as a
neuromodulator. Mol Neurobiol. 26:13–19. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Moore PK, Bhatia M and Moochhala S:
Hydrogen sulfide: From the smell of the past to the mediator of the
future? Trends Pharmacol Sci. 24:609–611. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang R: Two's company, three's a crowd:
Can H2S be the third endogenous gaseous transmitter?
FASEB J. 16:1792–1798. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kombian SB, Reiffenstein RJ and Colmers
WF: The actions of hydrogen sulfide on dorsal raphe serotonergic
neurons in vitro. J Neurophysiol. 70:81–96. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dello Russo C, Tringali G, Ragazzoni E,
Maggiano N, Menini E, Vairano M, Preziosi P and Navarra P: Evidence
that hydrogen sulphide can modulate hypothalamopituitary-adrenal
axis function: In vitro and in vivo studies in the rat. J
Neuroendocrinol. 12:225–233. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Eto K and Kimura H: A novel enhancing
mechanism for hydrogen sulfide-producing activity of cystathionine
beta-synthase. J Biol Chem. 277:42680–42685. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Julian D, Statile JL, Wohlgemuth SE and
Arp AJ: Enzymatic hydrogen sulfide production in marine
invertebrate tissues. Comp Biochem Physiol A Part, Mol Integr
Physiol. 133:105–115. 2002. View Article : Google Scholar
|
|
39
|
Chen X, Jhee KH and Kruger WD: Production
of the neuromodulator H2S by cystathionine beta-synthase
via the condensation of cysteine and homocysteine. J Biol Chem.
279:52082–52086. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yusuf M, Kwong Huat BT, Hsu A, Whiteman M,
Bhatia M and Moore PK: Streptozotocin-induced diabetes in the rat
is associated with enhanced tissue hydrogen sulfide biosynthesis.
Biochem Biophys Res Commun. 333:1146–1152. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Levonen AL, Lapatto R, Saksela M and
Raivio KO: Human cystathionine gamma-lyase: Developmental and in
vitro expression of two isoforms. Biochem J. 347:291–295. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Smith HS: Hydrogen sulfide's involvement
in modulating nociception. Pain Physician. 12:901–910.
2009.PubMed/NCBI
|
|
43
|
Zhao W, Zhang J, Lu Y and Wang R: The
vasorelaxant effect of H(2)S as a novel endogenous gaseous K(ATP)
channel opener. EMBO J. 20:6008–6016. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Boehning D and Snyder SH: Novel neural
modulators. Ann Rev Neurosci. 26:105–131. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Awata S, Nakayama K, Suzuki I, Sugahara K
and Kodama H: Changes in cystathionine gamma-lyase in various
regions of rat brain during development. Biochem Mol Biol Int.
35:1331–1338. 1995.PubMed/NCBI
|
|
46
|
Xu GY, Winston JH, Shenoy M, Zhou S, Chen
JD and Pasricha PJ: The endogenous hydrogen sulfide producing
enzyme cystathionine-beta synthase contributes to visceral
hypersensitivity in a rat model of irritable bowel syndrome. Mol
Pain. 5:442009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hatakeyama Y, Takahashi K, Tominaga M,
Kimura H and Ohta T: Polysulfide evokes acute pain through the
activation of nociceptive TRPA1 in mouse sensory neurons. Mol Pain.
11:242015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Maeda Y, Aoki Y, Sekiguchi F, Matsunami M,
Takahashi T, Nishikawa H and Kawabata A: Hyperalgesia induced by
spinal and peripheral hydrogen sulfide: Evidence for involvement of
Cav3.2 T-type calcium channels. Pain. 142:127–132. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kawabata AT, Ishiki K, Nagasawa K, Yoshida
S, Maeda Y, Takahashi T, Sekiguchi F, Wada T, Ichida S and
Nishikawa H: Hydrogen sulfide as a novel nociceptive messenger.
Pain. 132:74–81. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Schicho R, Krueger D, Zeller F, Von
Weyhern CW, Frieling T, Kimura H, Ishii I, De Giorgio R, Campi B
and Schemann M: Hydrogen sulfide is a novel prosecretory
neuromodulator in the Guinea-pig and human colon. Gastroenterology.
131:1542–1552. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Krueger D, Foerster M, Mueller K, Zeller
F, Slotta-Huspenina J, Donovan J, Grundy D and Schemann M:
Signaling mechanisms involved in the intestinal pro-secretory
actions of hydrogen sulfide. Neurogastroenterol Motil.
22:1224–1231. e319–320. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhao S, Liu FF, Wu YM, Jiang YQ, Guo YX
and Wang XL: Upregulation of spinal NMDA receptors mediates
hydrogen sulfide-induced hyperalgesia. J Neurol Sci. 363:176–181.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Hu S, Xu W, Miao X, Gao Y, Zhu L, Zhou Y,
Xiao Y and Xu GY: Sensitization of sodium channels by cystathionine
β-synthetase activation in colon sensory neurons in adult rats with
neonatal maternal deprivation. Exp Neurol. 248:275–285. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Qu R, Tao J, Wang Y, Zhou Y, Wu G, Xiao Y,
Hu CY, Jiang X and Xu GY: Neonatal colonic inflammation sensitizes
voltage-gated Na (+) channels via upregulation of cystathionine
β-synthetase expression in rat primary sensory neurons. Am J
Physiol Gastrointest Liver Physiol. 304:G763–G772. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wang HJ, Xu X, Xie RH, Rui YY, Zhang PA,
Zhu XJ and Xu GY: Prenatal maternal stress induces visceral
hypersensitivity of adult rat offspring through activation of
cystathionine- β-synthase signaling in primary sensory neurons. Mol
Pain. 14:17448069187774062018. View Article : Google Scholar
|
|
56
|
Lan A, Liao X, Mo L, Yang C, Yang Z, Wang
X, Hu F, Chen P, Feng J, Zheng D and Xiao L: Hydrogen sulfide
protects against chemical hypoxia-induced injury by inhibiting
ROS-activated ERK1/2 and p38MAPK signaling pathways in PC12 cells.
PLoS One. 6:e259212011. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Gobbi G, Ricci F, Malinverno C, Carubbi C,
Pambianco M, Panfilis GD, Vitale M and Mirandola P: Hydrogen
sulfide impairs keratinocyte cell growth and adhesion inhibiting
mitogen-activated protein kinase signaling. Lab Invest.
89:994–1006. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yang G, Sun X and Wang R: Hydrogen
sulfide-induced apoptosis of human aorta smooth muscle cells via
the activation of mitogen-activated protein kinases and caspase-3.
FASEB J. 18:1782–1784. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yang F, Sun W, Yang Y, Wang Y, Li CL, Fu
H, Wang XL, Yang F, He T and Chen J: SDF1-CXCR4 signaling
contributes to persistent pain and hypersensitivity via regulating
excitability of primary nociceptive neurons: Involvement of
ERK-dependent Nav1.8 up-regulation. J Neuroinflammation.
12:2192015. View Article : Google Scholar
|
|
60
|
Chen Y, Zhang Y, Huo Y, Wang D and Hong Y:
Adrenomedullin mediates tumor necrosis factor-α-induced responses
in dorsal root ganglia in rats. Brain Res. 1644:183–191. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ma W, Zheng WH, Powell K, Jhamandas K and
Quirion R: Chronic morphine exposure increases the phosphorylation
of MAP kinases and the transcription factor CREB in dorsal root
ganglion neurons: An in vitro and in vivo study. Eur J Neurosci.
14:1091–1104. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang WX, Wu Q, Liang SS, Zhang XK, Hu Q,
Chen QH, Huang HJ, Xu L and Lou FQ: Dexmedetomidine promotes the
recovery of neurogenesis in aged mouse with postoperative cognitive
dysfunction. Neurosci Lett. 677:110–116. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Sabbir MG and Fernyhough P: Muscarinic
receptor antagonists activate ERK-CREB signaling to augment neurite
outgrowth of adult sensory neurons. Neuropharmacology. 143:268–81.
2018. View Article : Google Scholar : PubMed/NCBI
|