Introduction
In recent years, screening cancer therapeutic drugs
from natural products has become a focus in cancer prevention and
treatment research. The efficacy of traditional Chinese medicines
and their extracts in the treatment of diseases has been
recognized, and more effective drugs for treating tumors are being
identified. Brucea javanica is a shrub that mainly grows in
Asia, particularly in southern China, and is used to treat a
variety of diseases, such as malaria (1), amoebic dysentery (2) and tumors (3). Brusatol (BRU), an important
component extracted from Brucea javanica (2), exerts a multitude of biological
effects, including inhibiting the growth of tumor cells, reducing
the reproduction of malaria parasites, reducing inflammation and
resisting virus invasion (4).
Clinical trials have demonstrated that BRU is a potential
anticancer drug with potent cytotoxicity towards several types of
cancer cells, including colorectal cancer (5), pancreatic cancer (6) and lung cancer (7). In addition, BRU can enhance the
sensitivity of cancer cells to chemotherapeutic drugs by
specifically blocking the expression of nuclear factor erythrocyte
2 related factor 2 (NRF2) (7,8).
These findings suggest that BRU may be an effective antineoplastic
drug and may be developed as a chemotherapeutic adjuvant for the
treatment of a variety of tumors (9). Unfortunately, BRU is associated with
several toxicities, including cardiac ischemia/reperfusion injury
(10). It also reverses the
therapeutic effects of other drugs, leading to aggravation of
neuroinflammation and nerve injury (11), septicemic kidney injury (12), liver injury (13) and intestinal epithelial cell
injury (14). These toxicities
are attributed to the inhibition of NRF2. Furthermore, other
studies have reported that BRU can affect the early development of
mouse embryos and exerts toxic effects on mouse oocytes (15,16). However, the response rate to most
chemotherapeutics in the treatment of human cancer remains low, and
there is an urgent need for developing new and safe therapeutic
agents.
Cancer is one of the most devastating diseases and
constitutes a major threat to global public health and quality of
life. Cancer is the second most fatal disease after cardiovascular
disease in developed and developing countries (17). There were a reported 9.6 million
deaths and 18.1 million new cancer cases worldwide in 2018
(18,19). Lung cancer is one of the most
common malignant tumors and a leading cause of cancer-related
mortality (20), whereas
endometrial and ovarian cancers are the most common malignant
tumors of the female reproductive system. The incidence of ovarian
cancer is slightly lower compared with that of endometrial cancer
(21). In the early stages of the
three cancers mentioned above, the symptoms are not obvious;
therefore, these cancers are often diagnosed after extensive
metastasis has occurred, and the treatment methods are ineffective,
resulting in poor prognosis (22-24). Therefore, with the rapid increase
of cancer cases worldwide, it is crucial to develop and screen
potential anticancer drugs (25).
However, the currently available anticancer drugs can cause serious
side effects and complications. Therefore, there is an urgent need
for effective and low-toxicity treatment methods, and for
innovative anticancer strategies in order to reduce the mortality
of patients with malignant tumors and improve their quality of life
(26).
Targeted drug therapy using nanoparticles (NPs) is a
new method of cancer treatment, which greatly improves the
therapeutic effect of several existing drugs (27,28). The effectiveness of NPs and
selective killing of cancer cells have been confirmed in several
studies (29-31). A major study reported that
glycosaminoglycan-placental chondroitin sulfate A (plCSA) is widely
expressed in human tumors, with absent-to-low expression in normal
tissues other than the placenta (32). In addition, our previous research
demonstrated that plCSA-binding peptide (plCSA-BP) lipid polymer
NPs could rapidly bind to choriocarcinoma cells and notably enhance
the anticancer activity of doxorubicin in vivo (33). Therefore, the use of
plCSA-targeting lipid polymer NPs to treat human cancers is a newly
recognized tool for delivering drugs to cancer cells, while
minimizing the risk of injury to normal cells.
In the present study, plCSA-targeted lipid polymer
NPs that encapsulated BRU were used to validate its efficiency and
targeting of various tumors in vitro, such as lung,
endometrial and ovarian cancer. The uptake of plCSAcoumarin 6 NPs
(CNPs) by tumor cells was compared to that of other types of NPs,
and it was investigated whether plCSA-brusatol-loaded NPs (BNPs)
effectively promoted the apoptosis of cancer cells and inhibited
their proliferation, invasion and migration through regulating the
B-cell CLL/lymphoma 2 (BCL2), BCL2-associated X protein (BAX),
cleaved caspase-3, matrix metalloproteinase (MMP)-2 and MMP-9
pathways.
Materials and methods
Experimental materials
Soybean lecithin and
1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N-[carboxy
(polyethylene glycol)-2000] (DSPEPEG-COOH) were purchased from
Avanti Polar Lipids, Inc. Poly(DL-lactic-co-glycolic acid) (50:50)
(PLGA), 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide
hydrochloride (EDC), and N-hydroxysuccinimide (NHS), were purchased
from Sigma-Aldrich; Merck KGaA. The plCSA-binding peptide (EDV KDI
NFD TKE KFL AGC LIV SFH EGKC) and the scrambled peptide (SCR; EVD
NDK KLG LVF EKD KIF TEF ACI SHC G) were synthesized by
ChinaPeptides Co., Ltd. and Shanghai GL Biochem Co. Ltd. BRU was
purchased from Absin. Transwell 24-well 8.0-µm pore
transparent plates were purchased from BD Biosciences. Matrigel was
purchased from Corning, Inc. The Cell Counting Kit-8 (CCK-8) was
purchased from Dojindo Molecular Technologies, Inc. The
anti-Bcl-2-associated X protein (BAX) (cat. no. 2774), anti-B-cell
lymphoma (BCL)-2 (cat. no. 2875), anti -cleaved caspase-3 (cat. no.
9661), anti-MMP-2 (cat. no. 40994), anti-MMP-9 (cat. no. 13667) and
anti-GAPDH (cat. no. 5174) were purchased from Cell Signaling
Technology, Inc. All other materials were obtained from
Sigma-Aldrich; Merck KGaA, unless otherwise specified.
BNPs or CNPs were synthesized using a single-step
sonication method using PLGA, soybean lecithin, BRU or coumarin 6,
and DSPE-PEG-COOH. PLGA was dissolved in acetonitrile, and soybean
lecithin and DSPE-PEG-COOH were dissolved in 4% ethanol solution.
BRU was dissolved in DMSO at a concentration of 10 mg/ml and
coumarin 6 was dissolved in DMSO at a concentration of 1 mg/ml. To
prepare BNPs, 750 µg of BRU, 90 µg of soybean
lecithin and 210 µg of DSPE-PEGCOOH were added into 3 ml of
4% ethanol aqueous solution, and then an ultrasonic processor (VCX
130; Sonics & Materials) was used to treat the solution. The
frequency of the ultrasonic processor was 20 kHz, the power was 30
W, and the time interval was 5 min. The BNPs were then centrifuged
(34,000 × g) for 30 min at 4°C (Optima™ MAX-XP; Beckman Coulter,
Inc.). After centrifugation, the supernatant was removed, and the
precipitate was resuspended in 1 ml of PBS (pH 7.4). The NPs were
purified by washing in PBS through an Amicon Ultra-4 centrifugal
filter (molecular weight cut-off, 10 kDa; EMD Millipore), and these
steps were repeated three times. CNPs were prepared using the same
method.
Conjugation of plCSA-BP or SCR to the
NPs
EDC/NHS technology was used to bind peptides to the
surface of the NPs. In brief, to bind plCSA-BP or SCR to the BNP
surface, activation buffer, EDC and NHS were added, and the molar
ratio of DSPE-PEG-COOH:NHS:EDC was 1:2:2 to activate the carboxyl
groups. The EDC/NHS solution and BNPs were mixed at room
temperature for 30 min, and then plCSA-BP or SCR were added. The
mixture was stirred at room temperature for 1-2 h, and then stored
overnight at 4°C. To remove the unbound peptide, the product was
reconstituted and purified as described above. PlCSA-BP and
SCR-conjugated CNPs were prepared using the same method.
Encapsulation efficiency (EE) and drug
loading efficiency (LE) of BRU in the NPs
The EE and LE of BRU in the NPs were determined as
described below. First, the following steps were used to produce a
standard curve: BRU was dissolved in DMSO at concentrations of 1,
5, 10, 20, 30, 50 and 80 µg/ml. The absorption of the BRU
solution at 280 nm was measured using an ultraviolet visible
(UV-vis) spectrophotometer (f900, Edinburgh Instruments Ltd.), and
then the standard curve was generated according to the
concentration of BRU. Subsequently, 25 µl of NPs were
diluted with 500 µl of ultra-pure water, and the absorption
at 280 nm was measured using the UV-vis spectrophotometer. The drug
concentration was calculated using the standard curve. The EE and
LE were calculated as follows: EE=(amount of drug in NPs/amount of
added drug) ×100%; and LE=(amount of drug in NPs/total weight of
materials) ×100%.
Cell culture and treatment
The human ovarian adenocarcinoma cell line SKOV3 was
obtained from Wuhan Boster Biological Technology, Ltd. The human
lung epithelial cell adenocarcinoma cell line A549 was obtained
from the Shanghai Institute of Cell Biology, Chinese Academy of
Sciences. The human endometrial adenocarcinoma cell line HEC-1-A,
the normal human ovarian epithelial cell line IOSE80, human
endometrial stromal cells (hESCs), and the normal human lung
epithelial cells BEAS-2B, were purchased from BeNa Culture
Collection. The A549 cells were grown in DMEM; the SKOV3 and
HEC-1-A cell were maintained in McCoy's 5A medium; IOSE80 and
BEAS-2B cells were cultured in RPMI-1640 medium; and hESCs were
cultured in DMEM/F12 supplemented with 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.), 100 µg/ml streptomycin and 100
U/ml penicillin G. Apart from the different media, all the cell
culture conditions were the same as mentioned above. All cells were
placed in a cell culture chamber with 5% CO2 at 37°C
with sufficient saturated humidity.
BRU does not emit light; therefore, coumarin 6 was
used instead of BRU to treat cells, in order to observe the rate of
uptake of the NPs. First, cells were grown to 70-80% confluence and
treated with free coumarin 6, CNPs, SCR-CNPs, or the plCSA-CNPs, in
24-well plates for 1 h at 4°C. The cells were washed with PBS to
remove unconjugated coumarin 6 or NPs, and incubated for 30 min at
37°C. Only surface-bound NPs were allowed to internalize. The cells
in each group were then fixed at room temperature for 15 min with
4% paraformaldehyde, stained with
2-(4-amidinophenyl)-1H-indole-6-carboxamidine (DAPI) for 10 min at
room temperature, washed with PBS three times, and fixed at room
temperature for 15 min with 4% paraformaldehyde. Finally, the cells
of each group were observed under an inverted fluorescence
microscope (IX73; Olympus Corporation) at a magnification of ×200.
Four visual fields were randomly selected to calculate the
fluorescence intensity.
Detection of in vitro cell viability
using the CCK-8 assay
The CCK-8 method was used to detect the effect of
different treatments on the viability of tumor cells (SKOV3,
HEC-1-A and A549) and normal cells (IOSE80, hESCs and BEAS-2B).
Cells in the logarithmic growth phase were digested with trypsin.
Thereafter, the cell concentration was adjusted to 10,000 cells/ml
and the cells were inoculated into a 96-well plate at a density of
100 µl/well. Subsequently, the free BRU, BNPs, SCR BNPs, or
plCSA-BNPs at concentrations of 0, 0.2, 0.4, 0.6, 0.8, 1 and 2
µg/ml were added to the culture cells for drug intervention.
Each concentration was set with six replicate wells. The plates
were cultured at 37°C and 5% CO2. After 48 h of cell
culture, 20 µl of CCK-8 solution was added to each well. The
cells were incubated for 1 h before the reaction was terminated and
detected using an automatic enzyme marker.
Apoptosis assay using PE Annexin V
staining
The cellular apoptosis rate was determined using a
PE Annexin V Apoptosis Detection kit I (BD Biosciences) according
to the manufacturer's protocol. Briefly, tumor cells (SKOV3,
HEC-1-A and A549) and normal cells (IOSE80, hESCs and BEAS-2B) were
seeded into 6-well plates and treated with free BRU or plCSA-BNPs
for 24 h. On the following day, the cells were harvested, washed
twice with cold PBS, resuspended in 500 µl of binding
buffer, and stained with 5 µl 7-AAD and 5 µl PE
Annexin V for 15 min at room temperature (25°C) in the dark.
Fluorescence signals from at least 10,000 cells were analyzed
immediately using a FACSCalibur flow cytometer (BD Biosciences).
Dot plots and histograms were analyzed using FlowJo software,
v7.6.1 (FlowJo, LLC).
Wound healing and Transwell invasion
assays
For the cell invasion assay, pure Matrigel was
diluted with PBS at a ratio of 1:20, and then the diluted Matrigel
was added to the upper surface of the Transwell filter and placed
in the incubator at 37°C for 2 h. Subsequently, cells treated with
free BRU or plCSA-BNPs for 24 h were inoculated into the upper
chamber in 200 µl of culture medium without FBS, and 600
µl of culture medium with 10% FBS was added into the plate.
After 2 days of incubation, the cells attached to the upper surface
of the filter were removed with cotton swabs, and the cells
remaining on the lower surface were fixed with 4% paraformaldehyde
for 15 min at room temperature and stained with 0.5% crystal violet
solution for 10 min at room temperature. Subsequently, cells
invading the lower surface were counted in five random fields
(magnification, ×100) under an inverted fluorescence microscope
(IX73; Olympus Corporation). For the migration assay, cells were
seeded into 6-well plates until ~100% confluent, and the confluent
cell layer on an agar plate was scratched with a 200-µl
pipette tip. The cells were then washed with PBS and treated with
free BRU or plCSA-BNPs, then incubated in serum-free medium. The
degree of wound closure in each case was imaged and plotted at 0
and 24 h.
Western blotting
Free BRU or plCSA-BNPs were added to cells in the
logarithmic growth phase and incubated at 37°C for 24 h. The cells
were lysed by grinding on ice in RIPA buffer at 4°C for 30 min. The
lysate was centrifuged at 12,000 × g at 4°C for 10 min. One part of
the supernatant was used to detect the protein concentration using
a BCA Protein Assay kit (Beyotime Institute of Biotechnology). The
other part of the supernatant was added 4X sample buffer, boiled
for denaturation, and then subjected to SDS-PAGE with a 4%
concentrating gel and a 10% separation gel. Subsequently, the
separated proteins were transferred onto PVDF membranes (EMD
Millipore). The membranes were then blocked with 5% non-fat dry
milk in Tris-buffered saline with 0.1% Tween 20 (TBST) for 1.5 h
and incubated with the primary antibodies (BAX, 1:1,000 dilution;
BCL2 , 1:1,000 dilution; cleaved caspase-3, 1:1,000 dilution;
MMP-2, 1:1,000 dilution; MMP-9, 1:1,000 dilution; and GAPDH,
1:1,000 dilution) at 4°C overnight. After washing with TBST three
times, the membranes were further incubated with the secondary
antibody (cat. no. SA00001-2; 1:5,000 dilution; goat anti-rabbit;
ProteinTech Group, Inc.) for 1 h at room temperature, and the color
was developed using ECL chemiluminescent solution (Beyotime
Institute of Biotechnology). Finally, images were acquired and the
optical density value of the immunoreactive protein bands were
analyzed using Image J software, version 1.51j8 (Media Cybernetics,
Inc.). Loading was normalized using GAPDH.
Statistical analysis
All experimental data are expressed as the mean ±
standard deviation of at least three independent experiments.
GraphPad Prism 7.0 (GraphPad Software, Inc.) was used for the
statistical analysis. One-way ANOVA followed by Tukey's post hoc
test and two-way ANOVA followed by Bonferroni's post hoc test was
used to analyze the differences among groups. P<0.05 was
considered to indicate statistically significant differences.
Results
Characterization of NPs
The drug EE and LE of the NPs are crucial to ensure
a successful drug delivery system. The EE and LE of BRU in the NPs
were calculated by constructing standard curves. The EEs of BRU for
the BNPs, SCR-BNPs and plCSA-BNPs were 40.3±1.67, 38.8±1.83 and
39.5±1.94%, respectively. The LEs of BRU for the BNPs, SCR-BNPs and
plCSA-BNPs were 6.2±0.74, 5.3±0.56 and 5.1±0.42%, respectively.
These data indicate that the extent of drug loading and drug
encapsulation was maintained after plCSA-BP decoration of the
NPs.
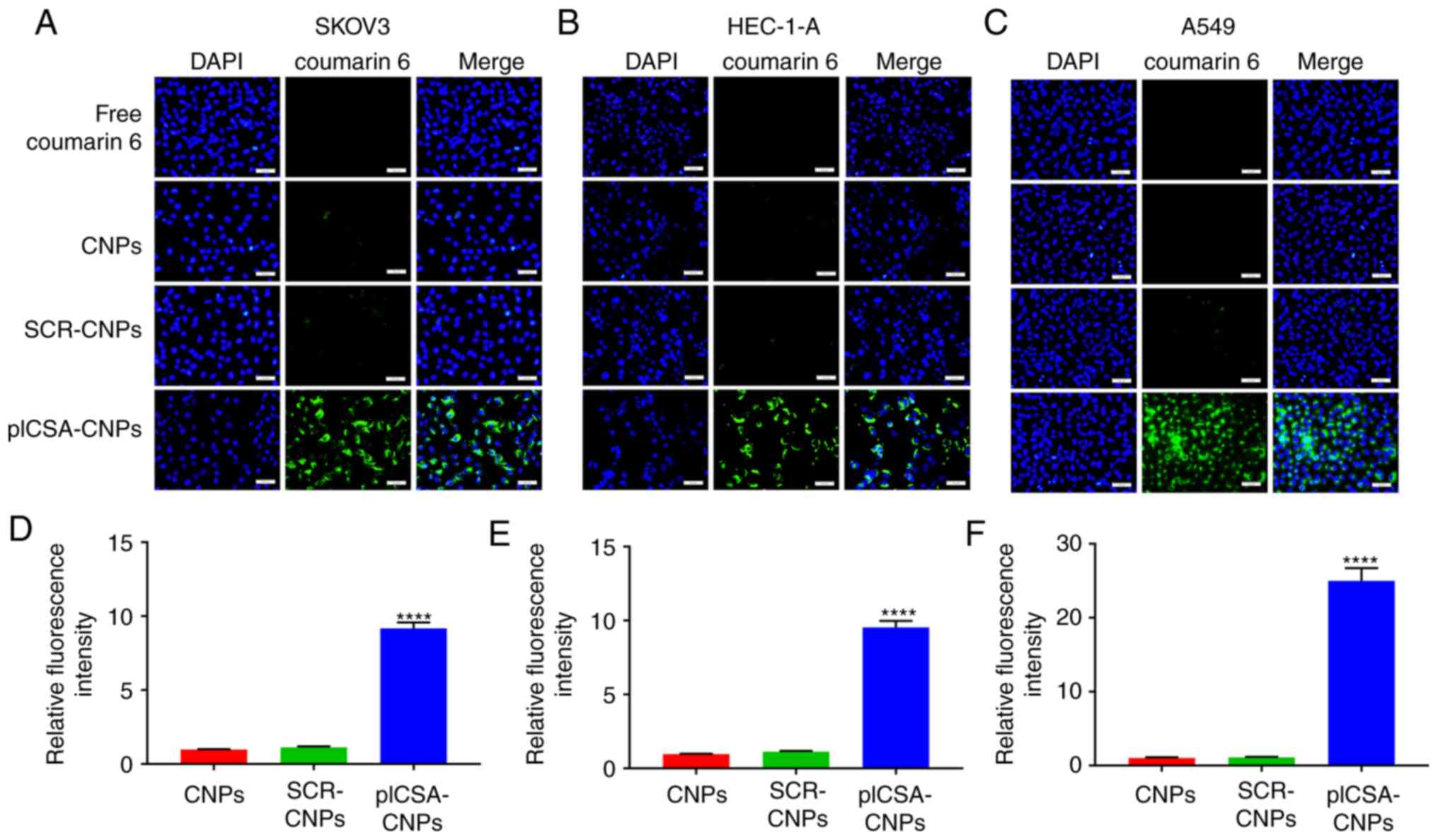
In vitro cellular uptake assay of the
NPs
In order to test whether the combination of plCSA-BP
with nanocarriers could increase the absorption of NPs by the
tumors, the SKOV3, HEC-1-A and A549 cell lines were cultured with
different NPs. Fluorescence microscopy demonstrated that cancer
cells treated with plCSA-CNPs had a higher fluorescence intensity
compared with that of the other treatments. Moreover, in SKOV3,
HEC-1-A and A549 cells, the NP uptake of the targeted preparation
into the cells was >9 times that of the non-targeted control
(P<0.0001; Fig. 1A-F). These
results suggest that the plCSA-targeting agent-absorbed NPs had a
better targeting efficiency compared with the non-targeted
controls. In addition, the combination with SCR did not lead to
enhanced uptake of NPs by the different cancer cells, indicating
the specific role of plCSA in the uptake by tumor cells.
Toxicity of BRU and plCSA-BNPs toward
normal cells
In order to evaluate the effect of BRU and
plCSA-BNPs on normal cells, the CCK-8 method was used to determine
the cytotoxicity of different NPs after 24 h. The results revealed
that 0-2 µg/ml of BRU could significantly inhibit the growth
of normal cells (IOSE80, hESCs and BEAS-2B), and the cytotoxicity
increased with increasing BRU concentration. However, the toxicity
of plCSA-BNPs toward normal cell lines was significantly lower
compared with that of free BRU (Fig.
2A). The P-values of IOSE80 cells treated with plCSA-BNPs at
0.2, 0.4, 0.6, 0.8, 1 and 2 µg/ml compared with that of
cells treated with free BRU were 0.0163, 0.0003, <0.0001,
<0.0001, <0.0001 and <0.0001, respectively. The P-values
of hESCs treated with plCSA-BNPs at 0.2, 0.4, 0.6, 0.8, 1 and 2
µg/ml compared with that of cells treated with free BRU were
0.0243, 0.0002, <0.0001, <0.0001, <0.0001 and <0.0001,
respectively. The P-values of BEAS-2B cells treated with plCSA-BNPs
at 0.2, 0.4, 0.6, 0.8, 1 and 2 µg/ml compared with that of
cells treated with free BRU were 0.0422, 0.0153, 0.0005,
<0.0001, <0.0001 and <0.0001, respectively. According to
the IC50 values of these NPs and free BRU, we analyzed
the advantages of the targeting NPs. The IC50 of
plCSA-BNPs (9.767 µg/ml) in IOSE80 cells was significantly
higher compared with that of free BRU (1.863 µg/ml). The
IC50 of plCSA-BNPs (5.492 µg/ml) in hESCs was
significantly higher compared with that of free BRU (1.8243
µg/ml). The IC50 of plCSA-BNPs (8.262
µg/ml) in BEAS-2B cells was significantly higher compared
with that of free BRU (1.938 µg/ml). These results
demonstrated that BNPs combined with plCSA-BP effectively reduced
the cytotoxicity compared with that of free BRU (P<0.05;
Fig. 2A). Moreover, to further
verify these findings, the ratios of apoptotic cells were detected
using flow cytometry. The apoptosis rates of IOSE80, hESCs and
BEAS-2B cells treated with plCSA-BNPs were 7.000±0.289, 6.033±0.145
and 6.133±0.240%, respectively, while those of IOSE80, hESCs and
BEAS-2B cells treated with free BRU were 11.500±0.252, 11.267±0.371
and 11.067±0.521%, respectively, which demonstrated that the
apoptosis rates in cells treated with BRU were 1.6 (P<0.0001),
1.8 (P<0.0001) and 1.8 (P=0.0002) times higher compared with
those in cells treated with free plCSA-BNPs (Fig. 2B). The results indicated that free
BRU increased the apoptosis of normal cells, whereas plCSA-BP could
reduce this effect. These in vitro results suggested that
plCSA-BNPs could reduce cytotoxicity compared with that of free
BRU.
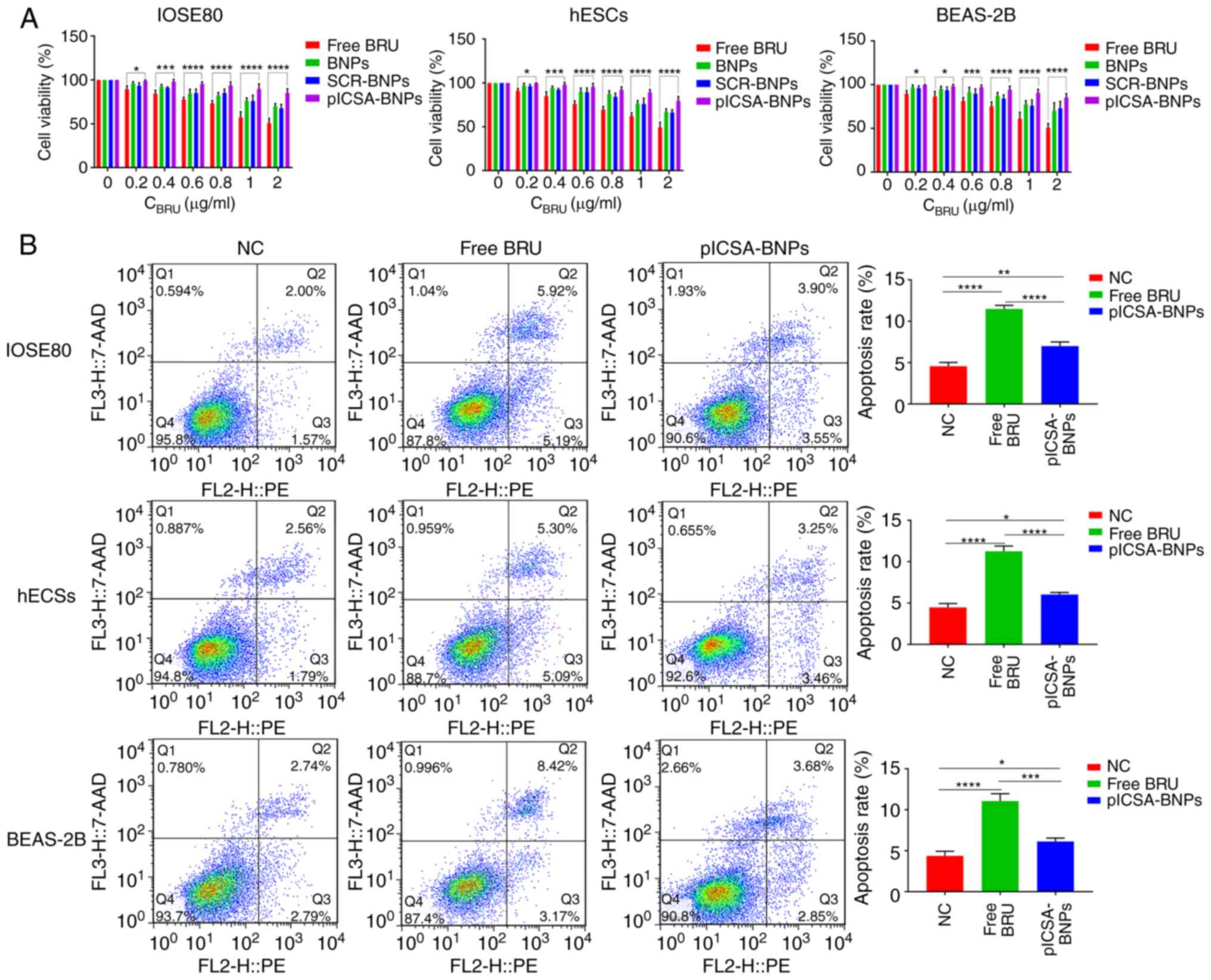 | Figure 2plCSA-BNPs reduce the cytotoxicity of
BRU. (A) Quantitative evaluation of the viability of different
normal cells treated with free BRU and the different nanoparticles
for 24 h using the Cell Counting Kit-8 assay. Two-way ANOVA was
used to analyze the significance of the differences among groups,
followed by Bonferroni's post hoc test. (B) After stimulation with
free BRU and the different nanoparticles for 24 h, the apoptosis
rate of different normal cells was detected using flow cytometry.
One-way ANOVA was used to analyze the significance of the
differences among groups, followed by Tukey's post hoc test. Values
are expressed as means ± standard deviation. *P<0.05,
**P<0.01, ***P<0.001,
****P<0.0001. The experiments were repeated three
times. IOSE80, human normal ovarian epithelial cell line; hESCs,
human endometrial stromal cells; BEAS-2B, human normal lung
epithelial cells; BRU, brusatol; plCSA, placental chondroitin
sulfate A; BNPs, BRU-loaded nanoparticles; NC, negative
control. |
plCSA-BNPs strongly inhibits tumor cell
growth
To confirm the antitumor activity of BRU and
plCSA-BNPs, the CCK-8 assay was used to evaluate cell viability
after 24 h of treatment with different NPs. The results
demonstrated that BRU significantly inhibited the activity of the
three types of cancer cells, and both targeted (plCSA-BNPs) and
non-targeted (BNPs and SCR-BNPs) NPs exhibited dose-dependent
cytotoxicity. When the dosage was 0.2 -2 µg/ml, the killing
effect of plCSA-BNPs on SKOV3, HEC-1-A and A549 cells was stronger
compared with that of BNPs and SCR-BNPs (Fig. 3A). The P-values of SKOV3 cells
treated with plCSA-BNPs at 0.2, 0.4, 0.6, 0.8, 1 and 2 µg/ml
compared with that of cells treated with free BRU were 0.0173,
<0.0001, <0.0001, <0.0001, <0.0001 and <0.0001,
respectively. The P-values of HEC-1-A cells treated with plCSA-BNPs
at 0.2, 0.4, 0.6, 0.8, 1 and 2 µg/ml compared with that of
cells treated with free BRU were <0.0001, <0.0001,
<0.0001, 0.0004, <0.0001 and <0.0001, respectively. The
P-values of A549 cells treated with plCSA-BNPs at 0.2, 0.4, 0.6,
0.8, 1 and 2 µg/ml compared with that of cells treated with
free BRU were 0.0055, <0.0001, <0.0001, 0.002, <0.0001 and
<0.0001, respectively. According to the IC50 values
of these NPs and free BRU, the advantages of the targeting NPs were
analyzed. The IC50 of plCSA-BNPs (0.5734 µg/ml)
in SKOV3 cells was significantly lower compared with that of free
BRU (1.832 µg/ml). The IC50 of plCSA-BNPs (0.2378
µg/ml) in HEC-1-A cells was significantly lower compared
with that of free BRU (0.975 µg/ml). The IC50 of
plCSA-BNPs (0.7084 µg/ml) in A549 cells was significantly
lower compared with that of free BRU (2.364 µg/ml). These
results suggested that conjugating the BNPs with plCSA-BP
effectively improved the growth inhibition of tumor cells compared
with that of free BRU (P<0.05; Fig. 3A). Moreover, to further verify the
findings mentioned above, the ratios of apoptotic cells were
detected using flow cytometry. The apoptosis rates of SKOV3,
HEC-1-A and A549 cells treated with plCSA-BNPs were 18.867±0.593,
35.533±0.742 and 15.60±0.702%, respectively, while those of SKOV3,
HEC-1-A and A549 cells treated with free BRU were 13.933±0.581,
12.033±0.578 and 10.00±0.577%, respectively, which demonstrated
that the apoptosis rates in cells treated with plCSA-BNPs were 1.3
(P=0.0012), 2.9 (P<0.0001) and 1.5 (P=0.0009) times higher
compared with those treated with free BRU (Fig. 3B). These results demonstrated that
free BRU increased cell apoptosis, whereas plCSA-BP further
enhanced this effect. These in vitro results suggested that
plCSA-BNPs may be a suitable candidate for cancer therapy.
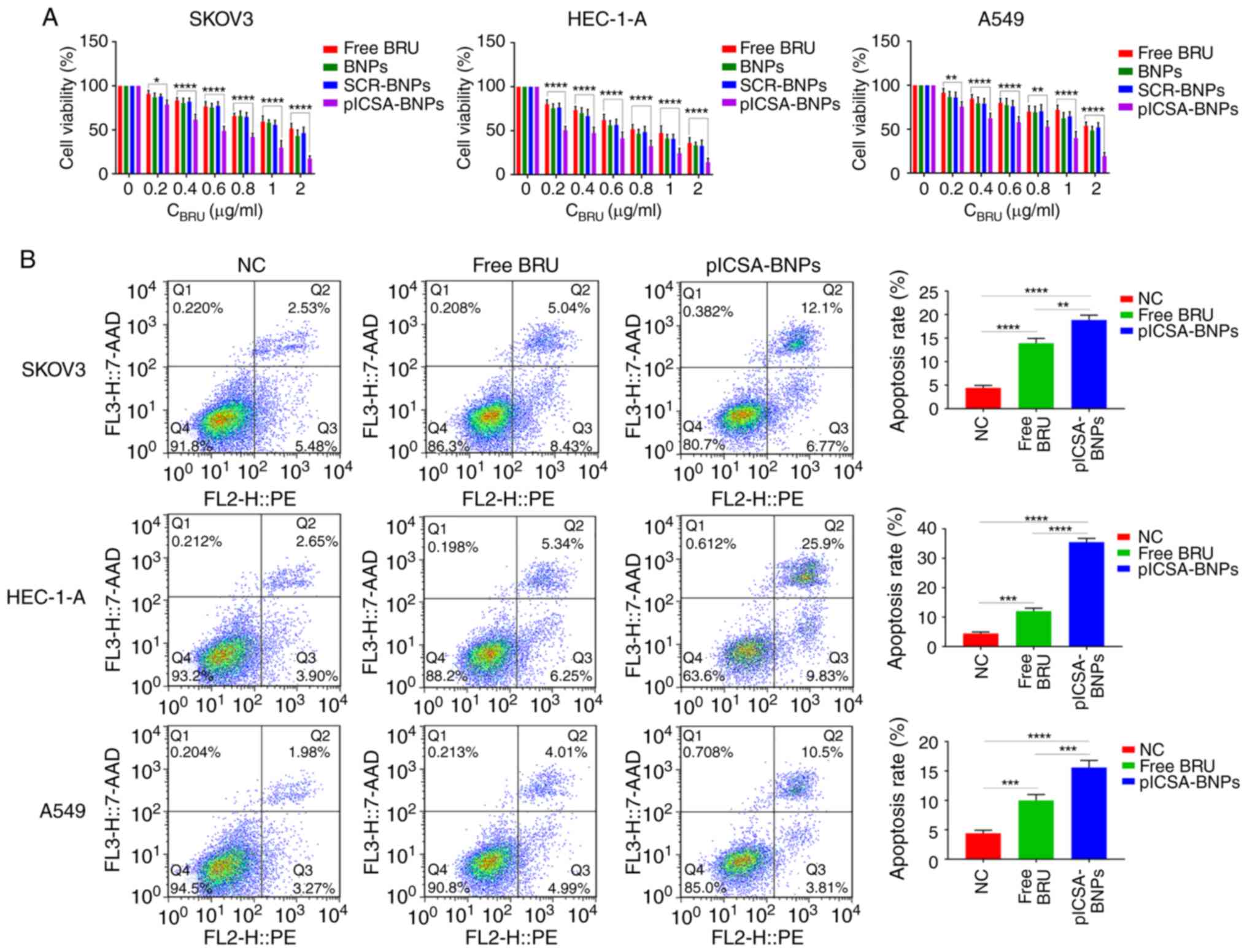 | Figure 3Growth inhibition of cell lines by
plCSA-BNPs. (A) Quantitative evaluation of the viability of
different cancer cells treated with free BRU and the different
nanoparticles for 24 h using the Cell Counting Kit-8 assay. Two-way
ANOVA was used to analyze the significance of the differences among
groups, followed by Bonferroni's post hoc test. (B) After
stimulation with free BRU and the different nanoparticles for 24 h,
the apoptosis rate of different cancer cells was detected using
flow cytometry. One-way ANOVA was used to analyze the significance
of the differences among groups, followed by Tukey's post hoc test.
Values are expressed as means ± standard deviation.
*P<0.05, **P<0.01,
***P<0.001, ****P<0.0001. The
experiments were repeated three times. BRU, brusatol; SCR,
scramble; plCSA, placental chondroitin sulfate A; BNPs, BRU-loaded
nanoparticles; NC, negative control. |
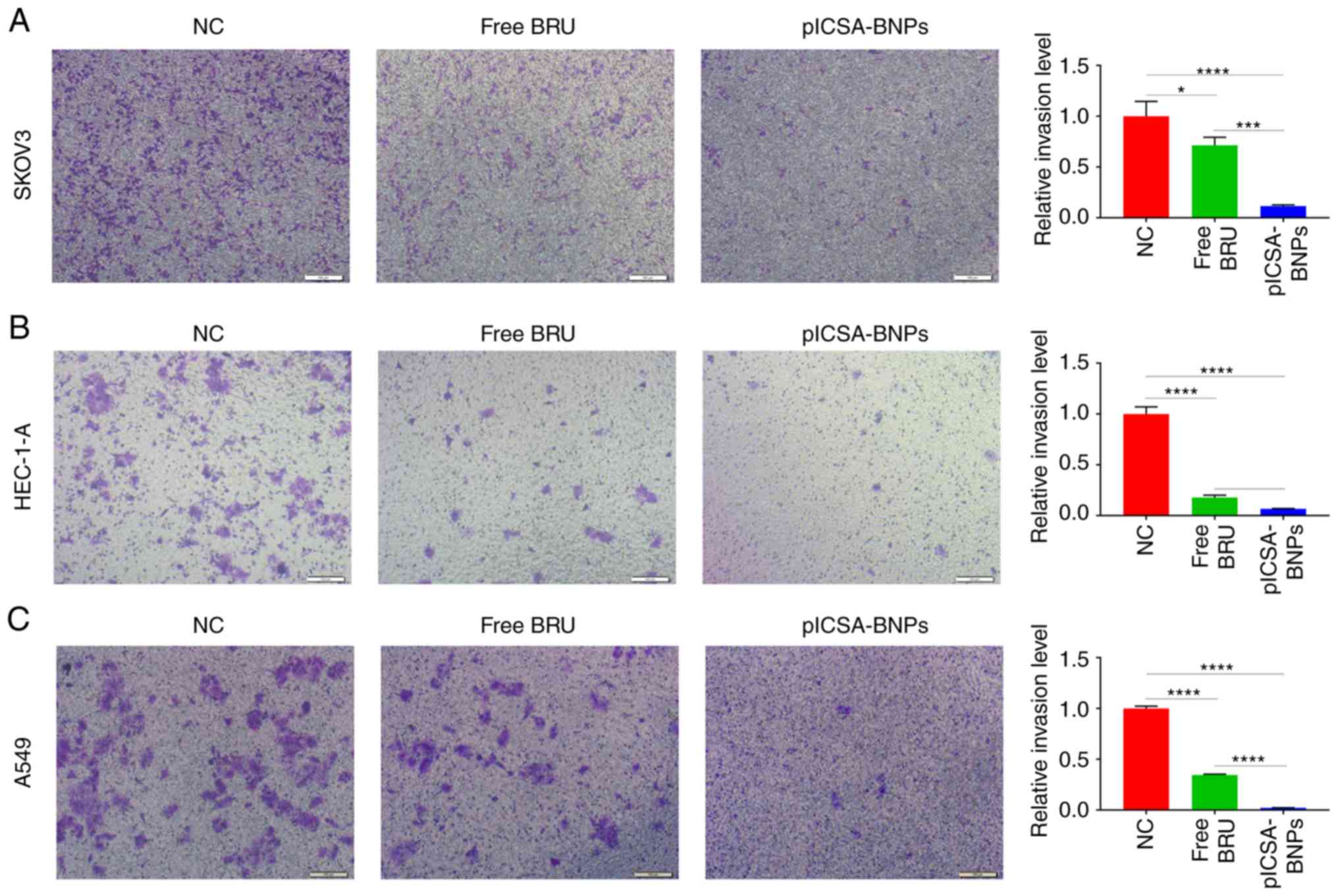
plCSA-BNPs strongly inhibit tumor cell
invasion
Transwell experiments were used to compare the
effects of free BRU and plCSA-BNPs on cell invasion. In SKOV3,
HEC-1-A and A549 cells, the cell invasion inhibition rates of the
plCSA-BNPs group were 6.2 (P=0.0007), 2.6 (P=0.0429) and 9.6
(P<0.0001) times higher compared with those of the BRU group,
respectively. These results demonstrated that plCSA-BNPs
significantly inhibited cell invasion (Fig. 4A-C).
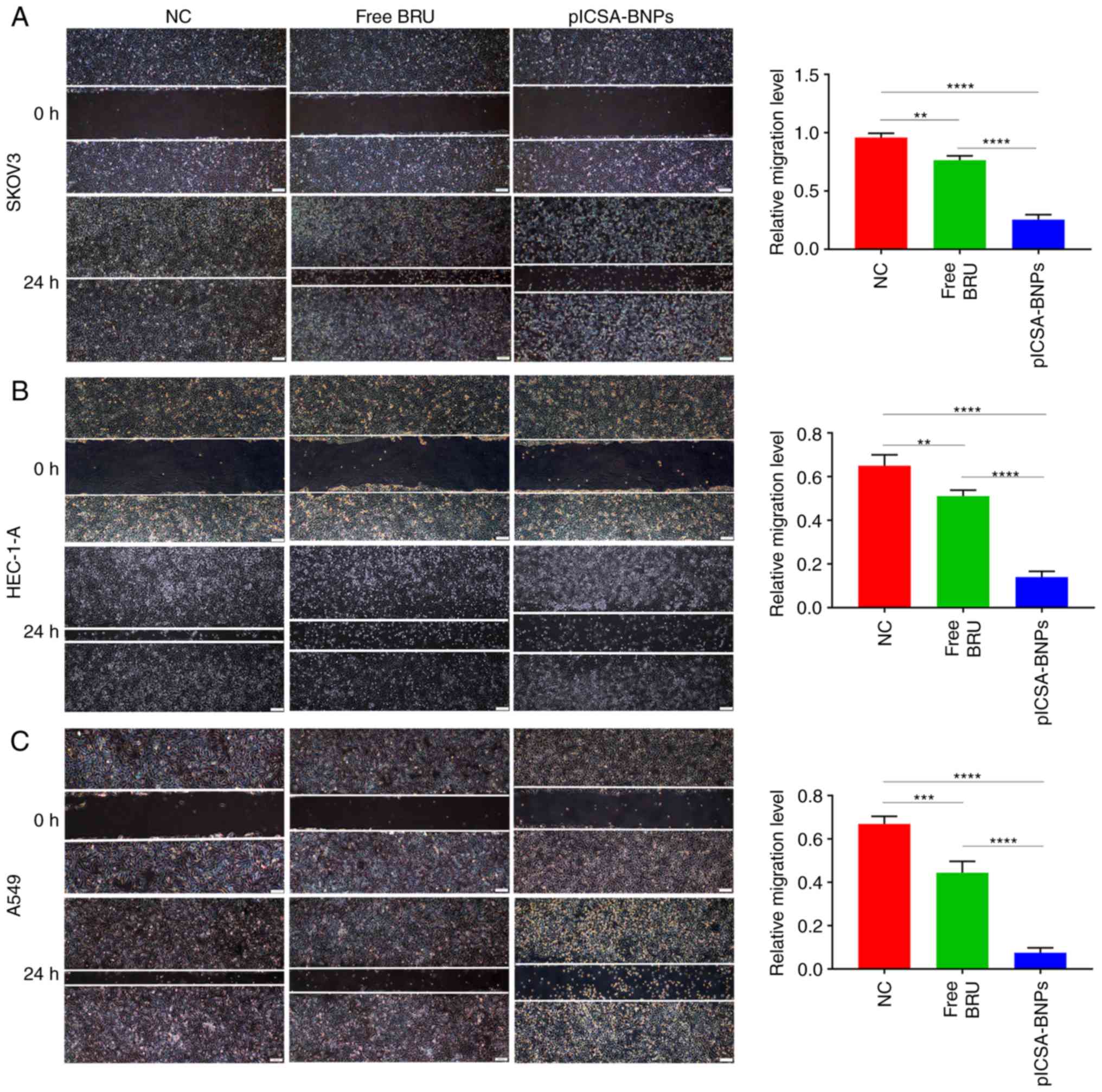
plCSA-BNPs strongly inhibit tumor cell
migration
Wound healing assays were performed to compare the
effects of plCSA-BNPs and free BRU on cell migration. In SKOV3,
HEC-1-A and A549 cells, the cell migration inhibition rates of the
plCSA-BNPs group were 3 (P<0.0001), 3.6 (P<0.0001) and 5.9
(P<0.0001) times higher compared with those of the BRU group,
respectively. These results demonstrated that plCSA-BNPs
significantly inhibited cell migration compared with the control
group (Fig. 5A-C).
plCSA-BNPs inhibit cancer cell
proliferation, invasion and migration by regulating the BCL2/BAX,
cleaved caspase-3 and MMP pathways
To explore the molecular mechanisms underlying the
effects of BRU on cancer cells, the SKOV3 cell line was selected,
treated with BRU and the different NPs, and then western blotting
was used to study the levels of apoptosis-related proteins, such as
BCL2, BAX and cleaved caspase-3, and migration- and
invasion-related proteins, such as MMP-2 and MMP-9. The levels of
the apoptosis-related proteins BCL2, BAX and cleaved caspase-3, as
well as those of the migration- and invasion-related proteins MMP-2
and MMP-9 are shown in Fig. 6B.
Treatment with free BRU resulted in significant downregulation of
BCL2, MMP-2 and MMP-9 levels, and upregulation of BAX and cleaved
caspase-3 levels, compared with those in the control group. In
addition, the same expression trends were observed following
plCSA-BNPs treatment for 24 h; however, the protein levels were
altered to a greater degree compared with those after free BRU
treatment. For BCL2, BAX, cleaved caspase-3, MMP-2 and MMP-9, the
changes the protein levels in the plCSA-BNPs group were 2
(P=0.0028), 2.2 (P=0.0192), 2 (P=0.0185), 3.5 (P<0.0001), and 2
(P<0.0001) times greater compared with those in the BRU group,
respectively. These results indicated that BRU promoted cancer cell
death by inhibiting cancer cell proliferation, invasion and
migration, whereas encapsulating BRU as plCSA-BNPs enhanced these
effects.
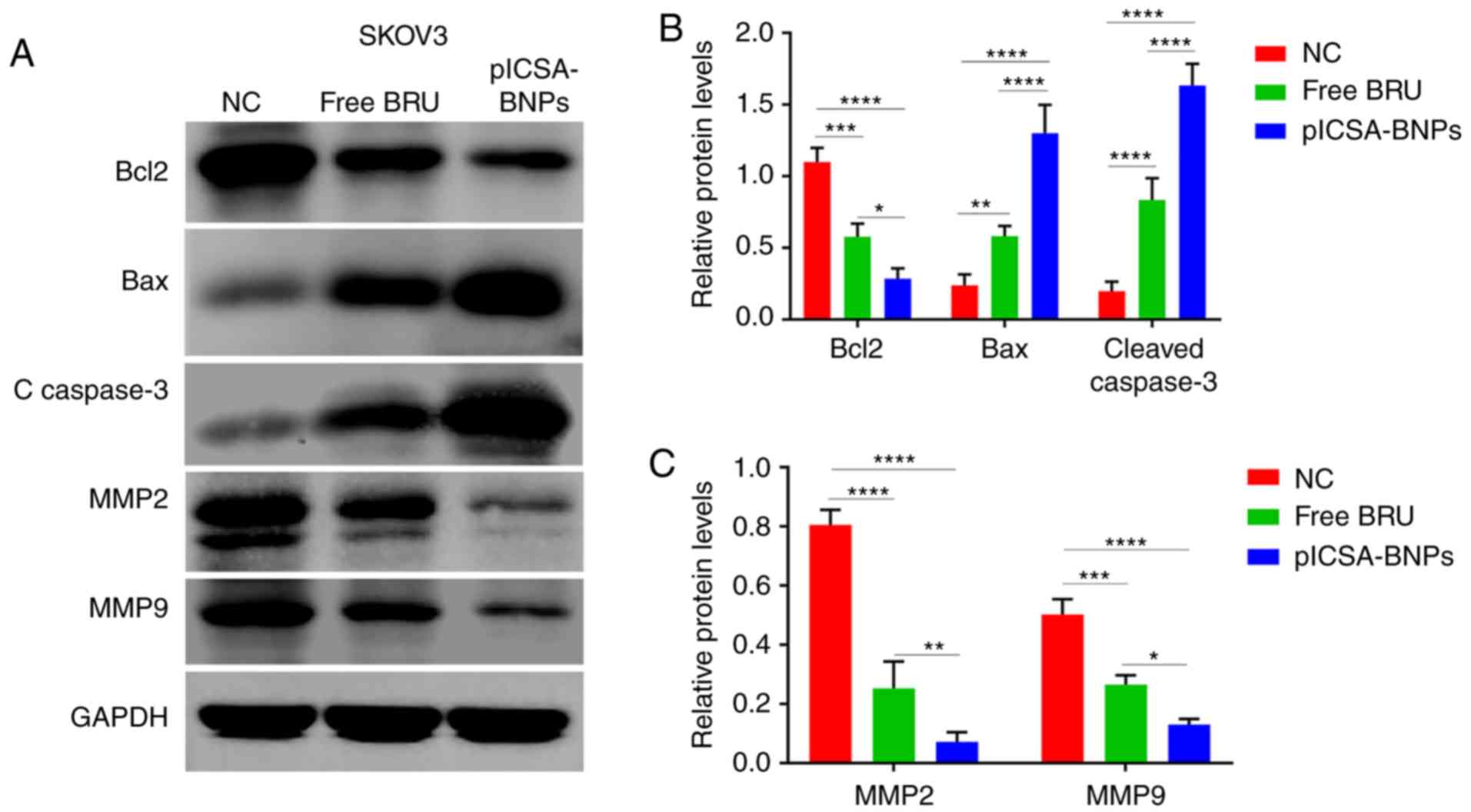 | Figure 6plCSA-BNPs promote cancer cell death
by regulating the BCL2/BAX, cleaved caspase-3 and MMP pathways.
Western blotting demonstrated that cancer cells treated with the
plCSA-BNPs exhibited significant downregulation of BCL2, MMP2 and
MMP 9 levels and upregulation of BAX and cleaved caspase-3 levels.
(A) Representative immunoblots of BCL2, BAX, cleaved caspase-3,
MMP-2 and MMP-9. GAPDH was used as the internal control. (B)
Densitometry results of BCL2, BAX and cleaved caspase-3 as analyzed
using ImageJ software. (C) Densitometry of MMP-2 and MMP-9 results
as analyzed using ImageJ software. One-way ANOVA was used to
analyze the significance of the differences among groups, followed
by Tukey's post hoc test. Values are expressed as means ± standard
deviation. *P<0.05, **P<0.01,
***P<0.001, ****P<0.0001. The
experiments were repeated three times. BRU, brusatol; plCSA,
placental chondroitin sulfate A; BNPs, BRU-loaded nanoparticles;
NC, negative control; BCL2, B-cell lymphoma 2; BAX, BCL2-associated
X protein; MMP, matrix metallopeptidase. |
Discussion
BRU has notable anticancer activity and may improve
disease outcomes in various cancers (34,35). However, BRU is associated with a
number of toxic adverse effects (10-16), which may limit its application in
the clinical setting. Nanocarriers are promising systems for the
effective treatment of cancer (36,37). As mentioned above,
glycosaminoglycan plCSA is specifically expressed in most cancer
cells and placental trophoblasts. Therefore, plCSA-NPs may be used
in targeted therapy of human tumor cells. Thus, it was hypothesized
that encapsulating BRU in plCSA-modified NPs would prevent its
inherent side effects (36). In
the present study, plCSA-BP-conjugated lipid-polymer NPs loaded
with BRU (plCSA-BNPs) were synthesized. We observed that the uptake
of plCSA-BP modified NPs by cancer cells was markedly higher
compared with that of other types of NPs. Three different cancer
cell lines were treated with free BRU, BNPs, SCR-BNPs or
plCSA-BNPs. The results demonstrated that plCSA-BNPs promoted the
apoptosis of cancer cells more effectively, and inhibited the
proliferation, invasion and migration of cancer cells by regulating
the BCL2, BAX, cleaved caspase-3, MMP-2 and MMP-9 pathways. The
results of the present study will hopefully improve our
understanding of the successful application of plCSA-BP modified
NPs and provide an effective tool for targeted drug delivery to
cancer cells. These plCSA targeting lipid polymer NPs may be used
to deliver drugs that target most human cancers, thereby
representing a breakthrough in the targeted therapy of cancer and
supporting the development of new drug targeting methods for
traditional Chinese medicines.
The EE and LE of drugs in NPs are crucial for their
success as drug delivery systems (33). It was reported that the particles
were more stable when the drug loading levels were lower (38). Furthermore, the drug loading
amount is a key factor for the drug EE (38). Therefore, in the present study,
loading of small drug amounts was performed to ensure that the
prepared NPs were more stable.
Previous studies demonstrated that tumor cells
display good uptake of liposome polymer NPs (38,39). For example, Chu et al
proved that PEGPE/PLGA NPs exhibited higher drug loading, less
sudden release, good serum stability and higher cell uptake
compared with traditional PLGA NPs (40). The results of the present study
demonstrated that tumor cells were also capable of taking up
plCSA-CNPs, which is consistent with previous research results
(39-41). Our data suggested that plCSA-BNPs
efficiently delivered drugs into tumor cells to eliminate them.
BRU has been shown to modulate the apoptosis,
migration and invasion of human cancer cells (42), indicating its potential role in
cancer metastasis. Therefore, CCK-8 and cell apoptosis assays were
used to study the effects of BRU and different NPs on tumor cells
and normal cells. The effects of BRU and different NPs on tumor
cells were studied using migration and Transwell invasion assays.
The cytotoxicity of targeted (plCSA-BNPs) and non-targeted (BNPs
and SCR-BNPs) NPs against normal cells was less prominent compared
with that of free BRU, while BRU exerted a significant inhibitory
effect on the tumor cells, and both targeted (plCSA-BNPs) and
non-targeted (BNPs and SCR-BNPs) NPs exhibited dose-dependent
cytotoxicity. When the drug concentration was 0.2-2 µg/ml,
the killing effect of plCSA-BNPs on SKOV3, HEC-1-A and A549 cells
was stronger compared with that of free BRU, BNPs and SCR-BNPs,
suggesting that the combination of BNPs and plCSA-BP significantly
enhanced the growth inhibitory and pro-apoptotic effect of BRU on
tumor cells. Moreover, plCSA-BNPs could significantly reduce the
killing effect of BRU on normal cells. Thus, the NPs maintained the
antitumor activity of BRU and reduced its toxicity toward non-tumor
cells. The process of tumorigenesis and development is extremely
complex. In the present study, one cell line was selected, treated
with BRU and the different NPs, and western blotting was further
employed to examine the levels of apoptosis-related proteins and
migration- and invasion-related proteins. Apoptosis may occur
through two pathways: Extrinsic and intrinsic. However, there is
now evidence that these two pathways are interrelated, with
molecules in one pathway affecting the other (43). There is an additional pathway that
involves T-cell mediated cytotoxicity and
perforingranzyme-dependent killing of cells. This perforin/granzyme
pathway can induce apoptosis via either granzyme B or granzyme A.
The extrinsic, intrinsic and granzyme B pathways converge on the
same terminal, or execution, pathway. This pathway is initiated by
the cleavage of caspase-3 (44),
and the direct activation of caspase-3 is essential for granzyme
B-induced cell killing (45). It
has been reported that members of the BCL2 family regulate the
process of apoptosis, including anti-apoptotic and pro-apoptotic
transcription factors, such as BCL2 and BAX, respectively. The
activity of BAX leads to cytochrome c release to promote
apoptosis. By contrast, as an anti-apoptotic protein, BCL2 can
prevent apoptosis-related death (46). BRU may also inhibit invasion,
migration and epithelial-to-mesenchymal transition (42). The interaction between apoptotic
and anti-apoptotic proteins determines cell survival. Members of
the MMP family are associated with cell migration and invasion.
MMP-2 and MMP-9 are the two most extensively investigated members
of the MMP family (47). In
accordance with previous studies, we found that among the three
groups, the levels of cleaved caspase-3 were significantly higher
in the plCSA-BNPs group. Therefore, plCSA-BNPs significantly
increased the transformation of the inactive procaspase-3 to active
cleaved caspase-3, which is the main inducer of apoptosis.
plCSA-BNPs promoted the levels of pro-apoptotic factors by
significantly increasing the expression of pro-apoptotic BAX. A
slight reduction in the levels of the anti-apoptotic BCL2 protein
was also observed. In addition, the plCSA-BNPs strongly inhibited
the expression of migration- and invasion-related proteins, such as
MMP-2 and MMP-9. These results demonstrated that BRU, delivered via
plCSA-modified NPs, may play a key role in promoting cancer cell
apoptosis and inhibiting invasion and migration via regulation of
important related proteins.
In conclusion, as a drug delivery system,
plCSA-BP-conjugated lipid polymer NPs loaded with BRU exerted a
notable antitumor effect in vitro, which could provide the
basis for the development of new anticancer drug delivery
systems.
Funding
This study was supported by the National Key
Research and Development Program of China (grant nos.
2018YFC1004103, 2018YFC1002804 and 2016YFC1000600), the National
Natural Science Foundation of China (grant nos. 81571513, 81771662
and 81771618), the National Key Natural Projects (grant no.
81830041), the National Natural Sciences Foundation (grant nos.
81771617 and 81771611), and the Shenzhen Basic Research Fund (grant
no. JCYJ20170412140326739).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
JY and XF were involved in the study design and
revision of the manuscript. XC, TY and BZ performed all the
experiments and drafted the manuscript. XC, TY, BZ, JC, TX, BW, ML
and BS performed data acquisition. XC, TY and BZ performed data
analysis. All the authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
All the authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Kitagawa I, Mahmud T, Simanjuntak P, Hori
K, Uji T and Shibuya H: Indonesian medicinal plants. VIII. Chemical
structures of three new triterpenoids, bruceajavanin A,
dihy-drobruceajavanin A, and bruceajavanin B, and a new alkaloidal
glycoside, bruceacanthinoside, from the stems of Brucea javanica
(Simaroubaceae). Chem Pharm Bull (Tokyo). 42:1416–1421. 1994.
View Article : Google Scholar
|
|
2
|
Liu JH, Qin JJ, Jin HZ, Hu XJ, Chen M,
Shen YH, Yan SK and Zhang WD: A new triterpenoid from. Arch Pharm
Res. 32:661–666. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cuendet M and Pezzuto JM: Antitumor
activity of bruceantin: An old drug with new promise. J Nat Prod.
67:269–272. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tang W, Xie J, Xu S, Lv H, Lin M, Yuan S,
Bai J, Hou Q and Yu S: Novel nitric oxide-releasing derivatives of
brusatol as anti-inflammatory agents: Design, synthesis, biological
evaluation, and nitric oxide release studies. J Med Chem.
57:7600–7612. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Oh ET, Kim CW, Kim HG, Lee JS and Park HJ:
Brusatol-mediated inhibition of c-myc increases HIF-1α degradation
and causes cell death in colorectal cancer under hypoxia.
Theranostics. 7:3415–3431. 2017. View Article : Google Scholar :
|
|
6
|
Lu Z, Lai ZQ, Leung AWN, Leung PS, Li ZS
and Lin ZX: Exploring brusatol as a new anti-pancreatic cancer
adjuvant: Biological evaluation and mechanistic studies.
Oncotarget. 8:84974–84985. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ren D, Villeneuve NF, Jiang T, Wu T, Lau
A, Toppin HA and Zhang DD: Brusatol enhances the efficacy of
chemotherapy by inhibiting the Nrf2-mediated defense mechanism.
Proc Natl Acad Sci USA. 108:1433–1438. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Olayanju A, Copple IM, Bryan HK, Edge GT,
Sison RL, Wong MW, Lai ZQ, Lin ZX, Dunn K, Sanderson CM, et al:
Brusatol provokes a rapid and transient inhibition of Nrf2
signaling and sensitizes mammalian cells to chemical
toxicity-implications for therapeutic targeting of Nrf2. Free Radic
Biol Med. 78:202–212. 2015. View Article : Google Scholar :
|
|
9
|
Zhang L, Feng X, Ma D, Yang J, Jiang H,
Zhang Y and He W: Brusatol isolated from Brucea javanica (L.) Merr.
induces apoptotic death of insect cell lines. Pestic Biochem
Physiol. 107:18–24. 2013. View Article : Google Scholar
|
|
10
|
Xiao C, Xia ML, Wang J, Zhou XR, Lou YY,
Tang LH, Zhang FJ, Yang JT and Qian LB: Luteolin attenuates cardiac
ischemia/reperfusion injury in diabetic rats by modulating nrf2
antioxidative function. Oxid Med Cell Longev. 2019:27192522019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang X, Lai W, Ying X, Xu L, Chu K, Brown
J, Chen L and Hong G: Salidroside reduces inflammation and brain
injury after permanent middle cerebral artery occlusion in rats by
regulating PI3K/PKB/Nrf2/NFKB signaling rather than complement C3
activity. Inflammation. 42:1830–1842. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang Y, Zhou F, Shen C, Wang H and Xiao
Y: LBP reduces theinflammatory injuryof kidney in septic rat and
regulates the Keap1-Nrf2ARE signaling pathway1. Acta Cir Bras.
34:e201900100000032019. View Article : Google Scholar
|
|
13
|
Lu Y, Wu S, Xiang B, Li L and Lin Y:
Curcumin attenuates oxaliplatin-induced liver injury and oxidative
stress by activating the Nrf2 pathway. Drug Des Devel Ther.
14:73–85. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Han X, Yao W, Liu Z, Li H, Zhang ZJ, Hei Z
and Xia Z: Lipoxin A4 preconditioning attenuates intestinal
ischemia reperfusion injury through Keap1/Nrf2 pathway in a lipoxin
A4 receptor independent manner. Oxid Med Cell Longev.
2016:93036062016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin Y, Sui LC, Wu RH, Ma RJ, Fu HY, Xu JJ,
Qiu XH and Chen L: Nrf2 inhibition affects cell cycle progression
during early mouse embryo development. J Reprod Dev. 64:49–55.
2018. View Article : Google Scholar :
|
|
16
|
Ma R, Li H, Zhang Y, Lin Y, Qiu X, Xie M
and Yao B: The toxic effects and possible mechanisms of Brusatol on
mouse oocytes. PLoS One. 12:e01778442017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kumar A, Sharma PR and Mondhe DM:
Potential anticancer role of colchicine-based derivatives: An
overview. Anticancer Drugs. 28:250–262. 2017. View Article : Google Scholar
|
|
18
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guo L, Liu Z and Tang X: Overexpression of
SLFN5 induced the epithelial-mesenchymal transition in human lung
cancer cell line A549 through β-catenin/Snail/E-cadherin pathway.
Eur J Pharmacol. 862:1726302019. View Article : Google Scholar
|
|
21
|
Gao S, Bian T, Zhang Y, Su M and Liu Y:
TCF12 overexpression as a poor prognostic factor in ovarian cancer.
Pathol Res Pract. 215:1525272019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhou X, Shi H, Jiang G, Zhou Y and Xu J:
Antitumor activities of ginseng polysaccharide in C57BL/6 mice with
Lewis lung carcinoma. Tumour Biol. 35:12561–12566. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou ML, Chen FS and Mao H: Clinical
significance and role of up-regulation of SERPINA3 expression in
endometrial cancer. World J Clin Cases. 7:1996–2002. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang T, Zhou R, Yu S, Yu S, Cui Z, Hu P,
Liu J, Qiao Q and Zhang J: Cytoplasmic SIRT1 inhibits cell
migration and invasion by impeding epithelial-mesenchymal
transition in ovarian carcinoma. Mol Cell Biochem. 459:157–169.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zeng L, Gupta P, Chen Y, Wang E, Ji L,
Chao H and Chen ZS: The development of anticancer ruthenium(ii)
complexes: From single molecule compounds to nanomaterials. Chem
Soc Rev. 46:5771–5804. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Geiger S, Lange V, Suhl P, Heinemann V and
Stemmler HJ: Anticancer therapy induced cardiotoxicity: Review of
the literature. Anticancer Drugs. 21:578–590. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
McDaid WJ, Greene MK, Johnston MC,
Pollheimer E, Smyth P, McLaughlin K, Van Schaeybroeck S,
Straubinger RM, Longley DB and Scott CJ: Repurposing of cetuximab
in antibody-directed chemotherapy-loaded nanoparticles in EGFR
therapy-resistant pancreatic tumours. Nanoscale. 11:20261–20273.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zheng W, Wang C, Ding R, Huang Y, Li Y and
Lu Y: Triptolide-loaded nanoparticles targeting breast cancer in
vivo with reduced toxicity. Int J Pharm. 572:1187212019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Benguigui M, Weitz IS, Timaner M, Kan T,
Shechter D, Perlman O, Sivan S, Raviv Z, Azhari H and Shaked Y:
Copper oxide nanoparticles inhibit pancreatic tumor growth
primarily by targeting tumor initiating cells. Sci Rep.
9:126132019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Alavi SE, Muflih Al, Harthi S, Ebrahimi
Shahmabadi H and Akbarzadeh A: Cisplatin-loaded
polybutylcyanoacrylate nanoparticles with improved properties as an
anticancer agent. Int J Mol Sci. 20:15312019. View Article : Google Scholar :
|
|
31
|
Yang B, Liu H, Yang H, Chen W, Wu J, Feng
X, Tong R, Yu H, Chen Y, Lv Z, et al: Combinatorial
photochemotherapy on liver cancer stem cells with
organoplatinum(ii) metallacage-based nanoparticles. J Mater Chem B.
7:6476–6487. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Salanti A, Clausen TM, Agerbaek MO, Al
Nakouzi N, Dahlback M, Oo HZ, Lee S, Gustavsson T, Rich JR, Hedberg
BJ, et al: Targeting human cancer by a glycosaminoglycan binding
malaria protein. Cancer Cell. 28:500–514. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang B, Cheng G, Zheng M, Han J, Wang B,
Li M, Chen J, Xiao T, Zhang J, Cai L, et al: Targeted delivery of
doxorubicin by CSA-binding nanoparticles for choriocarcinoma
treatment. Drug Deliv. 25:461–471. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee JH, Rangappa S, Mohan CD, Basappa,
Sethi G, Lin ZX, Rangappa KS and Ahn KS: Brusatol, a Nrf2 inhibitor
targets STAT3 signaling cascade in head and neck squamous cell
carcinoma. Biomolecules. 9:5502019. View Article : Google Scholar :
|
|
35
|
Zhang J, Fang X, Li Z, Chan HF, Lin Z,
Wang Y and Chen M: Redox-sensitive micelles composed of
disulfide-linked Pluronic-linoleic acid for enhanced anticancer
efficiency of brusatol. Int J Nanomedicine. 13:939–956. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Beg S, Kawish SM, Panda SK, Tarique M,
Malik A, Afaq S, Al-Samghan AS, Iqbal J, Alam K and Rahman M:
Nanomedicinal strategies as efficient therapeutic interventions for
delivery of cancer vaccines. Semin Cancer Biol. Oct 13–2019.Epub
ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Farokhzad OC, Cheng J, Teply BA, Sherifi
I, Jon S, Kantoff PW, Richie JP and Langer R: Targeted
nanoparticle-aptamer bioconjugates for cancer chemotherapy in vivo.
Proc Natl Acad Sci USA. 103:6315–6320. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lin WJ, Juang LW and Lin CC: Stability and
release performance of a series of pegylated copolymeric micelles.
Pharm Res. 20:668–673. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang G, Chen Y, Wang P, Wang Y, Hong H, Li
Y, Qian J, Yuan Y, Yu B and Liu C: Preferential tumor accumulation
and desirable interstitial penetration of poly(lactic-co-glycolic
acid) nanoparticles with dual coating of chitosan oligosaccharide
and polyethylene glycol-poly(D, L-lactic acid). Acta Biomater.
29:248–260. 2016. View Article : Google Scholar
|
|
40
|
Chu CH, Wang YC, Huang HY, Wu LC and Yang
CS: Ultrafine PEG-coated poly(lactic-co-glycolic acid)
nanoparticles formulated by hydrophobic surfactant-assisted one-pot
synthesis for biomedical applications. Nanotechnology.
22:1856012011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang B, Zheng M, Cai L and Fan X:
Synthesis and characterization of placental chondroitin sulfate A
(plCSA)-targeting lipid-polymer nanoparticles. J Vis Exp.
18:582092018.
|
|
42
|
Ye R, Dai N, He Q, Guo P, Xiang Y and
Zhang Q, Hong Z and Zhang Q: Comprehensive anti-tumor effect of
Brusatol through inhibition of cell viability and promotion of
apoptosis caused by autophagy via the PI3K/Akt/mTOR pathway in
hepatocellular carcinoma. Biomed Pharmacother. 105:962–973. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Igney FH and Krammer PH: Death and
anti-death: Tumour resistance to apoptosis. Nat Rev Cancer.
2:277–288. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
44
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Goping IS, Barry M, Liston P, Sawchuk T,
Constantinescu G, Michalak KM, Shostak I, Roberts DL, Hunter AM,
Korneluk R and Bleackley RC: Granzyme B-induced apoptosis requires
both direct caspase activation and relief of caspase inhibition.
Immunity. 18:355–365. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Korsmeyer SJ: BCL-2 gene family and the
regulation of programmed cell death. Cancer Res. 59(7 Suppl):
1693S–1700S. 1999.PubMed/NCBI
|
|
47
|
Jia YL, Shi L, Zhou JN, Fu CJ, Chen L,
Yuan HF, Wang YF, Yan XL, Xu YC, Zeng Q, et al: Epimorphin promotes
human hepatocellular carcinoma invasion and metastasis through
activation of focal adhesion kinase/extracellular signal-regulated
kinase/matrix metalloproteinase-9 axis. Hepatology. 54:1808–1818.
2011. View Article : Google Scholar : PubMed/NCBI
|