Introduction
Colorectal cancer (CRC), a type of malignant
gastrointestinal tumor, is the third leading cause of
tumor-associated mortality worldwide (1). Moreover, in China, the CRC incidence
in 2018 was 12.8% for men and 11.3% for women (2), and this rate is rapidly increases
along with the development of the Chinese economy (3). Currently, the primary curative
treatment for CRC is surgical resection; however, adjuvant
chemotherapy has been incorporated to reduce high rates of adjacent
tissue invasion and metastasis, thus decreasing the relapse rate
(4). Due to the invariable
incidence of drug resistance and serious side effects, including
diarrhea, nausea, swelling, vomiting, abdominal pain, tiredness,
low blood levels of albumin and other abnormalities, associated
with standard anti-cancer drugs, the outcomes of chemotherapy and
other effective measures are currently unsatisfactory for patients
with CRC (5). Therefore, the
investigation of novel treatment strategies with a safe profile
that act via different signaling pathways is urgently required to
develop improved targeted therapies.
Contrarily to traditional chemotherapeutic drugs,
certain natural products, including flavonoids and jatrorrhizine
(4), are considered to be
potential candidates for neoplastic therapy on account of their
substantial biological activities and relatively low adverse
effects (6). Moreover, ongoing
research for anti-cancer agents from medicinal plants has led to
the examination of Traditional Chinese Medicine (7). As a simple phenolic compound
extracted from the herbal root bark, paeonol
(2′-hydroxy-4′-methoxyacetophenone) has substantial biological and
pharmacological properties, including significant sedation,
analgesic action, antipyresis, anti-inflammation, anti-oxidation,
anti-hypertension, neuroprotection and immunomodulation (8,9).
In addition, paeonol has attracted increased attention in recent
years due to its desirable anti-tumor effect against various types
of cancer cell, both in vitro and in vivo, as
revealed by its ability to inhibit cell proliferation and induce
cell apoptosis (8,10).
Previous studies have reported that several
signaling path-ways have important roles in the progression of
cancer, including NF-κB, C-X-C motif chemokine ligand 4/C-X-C motif
chemokine receptor 3B and the PI3K/Akt/NF-κB pathway (10-12). It has been shown that the role of
the canonical Wnt signaling pathway is to regulate its downstream
genes responsible for the cell cycle and cell survival (13). In addition, it is crucial to
maintain homeostasis in multiple tissues throughout the body via
the Wnt/β-catenin pathway, and dysregulation of this pathway is
regarded as an important oncogenic event in numerous types of human
tumor, particularly in the initiation and progression of CRC
(14). As aberrant activation of
the Wnt/β-catenin pathway is involved in cell proliferation,
invasive features and development of chemoresistance resistance in
cancer cells, this pathway is considered as a potential novel
target of chemotherapeutic or chemopreventive agents for cancer
types (15). Currently, several
natural agents from drug discovery platforms, including aesculetin
(16), baicalin (17), hydnocarpin (15), isobavachalcone (18), jatrorrhizine (4), Sanguisorba officinalis
(19) and wogonin (20), have been verified to directly or
indirectly target β-catenin and its downstream signaling partners T
cell-specific transcription factor/lymphoid-enhancer binding factor
(TCF/LEF), consequently reducing the viability of CRC cells.
However, the underlying mechanisms responsible for the effects of
paeonol against CRC are yet to be fully elucidated. Therefore, the
present study aimed to identify the mechanisms of the anti-tumor
effect of paeonol on human CRC cells.
Materials and methods
Major reagents
Paeonol (purity, >98%) was obtained from
Sigma-Aldrich (Merck KGaA; cat. no. H35803) and the stock solution
of paeonol in alcohol was diluted to obtain the required
concentrations (7.8125, 15.625, 31.25, 62.5, 125, 250 and 500
µg/ml). RPMI-1640 medium and FBS were provided by Thermo
Fisher Scientific, Inc. A Cell Counting Kit-8 (CCK-8) was obtained
from Beyotime Institute of Biotechnology. The TRIzol®
total extraction kit was from Invitrogen (Thermo Fisher Scientific,
Inc.). Ribonuclease (RNase) and propidium iodide (PI) were
purchased from Sigma-Aldrich (Merck KGaA). The Annexin-V-FITC/PI
apoptosis detection kit was from BD Biosciences. Colorimetric
caspase assay kits were obtained from Abcam [cat. nos. ab39401
(caspase-3), ab39700 (caspase-8) and ab65608 (caspase-9)]. The
TCF/LEF reporter plasmid (cat. no. GM-021042) was purchased from
Jiman Biotechnology (Shanghai) Co., Ltd. Micropoly-transfecter
(cat. no. MT103) was obtained from Biosky Biotechnology Corporation
and D-Luciferin sodium (cat. no. 7902-100) was from BioVision, Inc.
RIPA buffer (cat. no. 6505729) and the bicinchoninic acid (BCA)
Protein Assay kit (cat. no. BL52A-1) were obtained from Biosharp
Life Sciences.
The primary rabbit antibodies against human Bax
(cat. no. ab32503), Bcl-2 (cat. no. ab59348), p21Cip1
(cat. no. ab145), cytochrome C (cat. no. ab13575), cyclin D1 (cat.
no. ab226823), cyclin-dependent kinase (CDK)4 (cat. no. ab137675),
c-Myc (cat. no. ab12213), survivin (cat. no. ab76424), glycogen
synthase kinase (GSK)-3β (cat. no. ab32391), β-catenin (cat. no.
ab32572) and β-actin (cat. no. ab8229) were obtained from Abcam. In
addition, horseradish peroxidase-conjugated goat anti-rabbit or
mouse IgG antibodies (cat. no. SA00001-1 or SA00001-2) were
obtained from ProteinTech Group, Inc. The other chemicals were of
analytical grade and obtained from local reagent suppliers.
Cell line and culture
The human CRC HCT116 cell line was provided by the
Cell Bank of the Chinese Academy of Sciences and was cultured in
RPMI-1640 medium containing 10% FBS and 1% penicillin/streptomycin
at 37°C in a humidi-fied atmosphere with 95% air and 5%
CO2. The cells used in the experiments were in the
logarithmic growth phase.
Cell proliferation assay
The CCK-8 assay was performed to determine the
number of viable cells according to the manufacturer's protocol. In
brief, 5×103 HCT116 cells per well in a 96-well plate
were incubated at 37°C with a series of concentrations of paeonol
(0, 7.8125, 15.625, 31.25, 62.5, 125, 250 and 500 µg/ml) for
12, 24, 48 and 72 h. Each condition was set up in 6-wells and the
assay was performed in duplicate. Then, 10 µl CCK-8 solution
was added to each well of the plates at 12, 24, 48 and 72 h. After
incubation at 37°C for another 4 h, the absorbance (A) at 550 nm
was detected to determine the number of viable cells using a
microplate reader (iMark680; Bio-Rad Laboratories, Inc.). The
inhibitory rate (IR) of HCT116 cells was calculated as follows: IR
(%)=[(mean Acontrol-mean Ablank)-(mean
Atest-mean Ablank)]/(mean Acontrol-mean
Ablank) ×100%, and the IC50 was obtained from
the cell growth curve using Bliss software (version 2.0; Bliss
Software Technologies Inc.).
Analysis of cell cycle
Based on the IC50 value, different doses
of paeonol (20, 40 and 80 µg/ml) were selected for the
study. After incubation at 37°C with paeonol in a 6-well plate
(1×105 cells per well) for 12, 24 and 48 h, the cells
were harvested, washed with 1X PBS and then incubated with 50
µg/ml PI solution containing 0.1 mg/ml RNase A in PBS (pH
7.4) for 30 min at room temperature in the dark. Subsequently, flow
cytometry (FCM) was performed using a FACSCalibur (BD Biosciences)
to analyze the fluorescence of the PI-DNA complex and further to
quantify cell-cycle fractions from ≥1×104 cells using
Cell Quest software (version 3.3; BD Biosciences).
Determination of apoptosis
The percentage of early apoptosis (Annexin
V+/PI−) and late apoptosis (Annexin
V+/PI+) was detected by FCM according to our
previous study (7). 20, 40 and 80
µg/ml paeonol at an earlier time point (24 h) and a moderate
dose of paeonol (40 µg/ml) at 12, 24 and 48 h were selected.
After incubation at 37°C, the cells were collected and washed three
times with ice-cold PBS. Following staining with 5 µl
Annexin V-FITC and 5 µl PI in 100 µl 1X binding
buffer for 15 min at room temperature in the dark, the total
apoptotic rate was examined with a flow cytometer using Cell Quest
software (version 3.3; BD Biosciences).
Colorimetric caspase activity assay
Following the manufacturer's instructions,
colorimetric caspase assay kits were performed to measure the
caspase activity. In brief, 1×106 HCT116 cells/ml were
incubated at 37°C with 0, 20, 40 and 80 µg/ml paeonol for a
moderate period of time (48 h). After washing with ice-cold PBS,
ice-cold cell lysis buffer was used to lyse cells for 15 min and
then the supernatant was separated by centrifugation (12,000 × g at
4°C for 10 min). The cell lysate was added to assay plates
containing reaction buffer with 10 µl acetyl-Asp-Glu-Val-Asp
p-nitroanilide as a substrate for caspase-3, acetyl-Ile-Glu-Thr-Asp
p-nitroanilide for caspase-8 or acetyl-Leu-Glu-His-Asp
p-nitroanilide for caspase-9, followed by incubation at 37°C in the
dark for 1.5 h. Finally, the A at 405 nm was measured with a
micro-plate reader to quantify the formation of p-nitroanilide, and
the relative increases of caspase-3, -8 and -9 activity were
calculated by comparing the A of paeonol-treated HCT116 cells with
the control group.
TCF/LEF luciferase reporter assay
The TCF/LEF dual-luciferase reporter assay was
performed following the manufacturer's instructions with minor
modifications. In brief, 1×104 HCT116 cells/well were
seeded into 24-well microtiter plates and maintained in RPMI-1640
medium overnight at 37°C in an incubator with 5% CO2
prior to transfection. After incubation at room temperature for 10
min, the mixture of 2 µg TCF/LEF reporter plasmid and 2
µl micropolytransfecter was added to RPMI 1640 culture
medium with HCT116 cells. Following the anti-biotic screening for
24 h, the transfection efficiency of HCT116 cells was performed by
measuring the signals from TCF/LEF reporter (firefly luminescence).
Then, the transfected cells were incubated at 37°C with either 0
(control), 20 or 80 µg/ml paeonol for 48 h. Finally,
D-luciferin sodium at a final concentration of 15 mg/ml was added
to each well to quantify the luciferase activity and the
fluorescence images were acquired using the IVIS®
Spectrum system and Living Image® software (version 4.5;
IVIS® Spectrum; PerkinElmer, Inc.).
Western blot analysis
Following incubation with 20, 40 and 80 µg/ml
paeonol at 37°C for 48 h, RIPA buffer was used to extract proteins
from the harvested cells and then a BCA Protein Assay kit was used
to determine the protein concentration in the supernatant after
centrifugation at 12,000 x g and 4°C for 30 min. Aliquots
containing 10 µg protein per lane were subjected to 10%
SDS-PAGE and the separated proteins were transferred onto PVDF
membranes (EMD Millipore). After blocking with a mixture of 5%
skimmed milk/0.1% Tris-buffered saline containing 0.1% Tween-20
(TBST) at 25°C for ≥2 h, the membranes were probed with anti-CDK4,
anti-p21Cip1, anti-Bax, anti-Bcl-2, anti-cytochrome C,
anti-glycogen synthase kinase 3 β (GSK-3β), anti-c-Myc (all
1:1,000), anti-cyclin D1 (1:2,000), anti-survivin, anti-β-catenin
(both 1:5,000) and mouse anti-β-actin (1:1,000) on a shaker table
at 4°C for ≥12 h, followed by incubation with horseradish
peroxidase-conjugated goat anti-rabbit or anti-mouse IgG (1:2,000)
at room temperature for 2 h. After further rinsing with TBST three
times, the membranes were visualized using an enhanced
chemiluminescence substrate (Amersham; Cytiva). The intensity of
each band relative to β-actin was determined semi-quantitatively
using ImageQuant TL software (version 7.0; Cytiva) (21).
Statistical analysis
Data are presented as the mean ± standard deviation
of ≥3 independent experiments performed in triplicate. Comparisons
between two groups were analyzed with an unpaired Student's t-test,
and one-way ANOVA followed by Tukey's post hoc test was performed
to determine differences among >2 groups using SPSS 18.0 (SPSS,
Inc.). P<0.05 was considered to indicate a statistically
significant difference.
Results
Paeonol reduces the number of viable
HCT116 cells
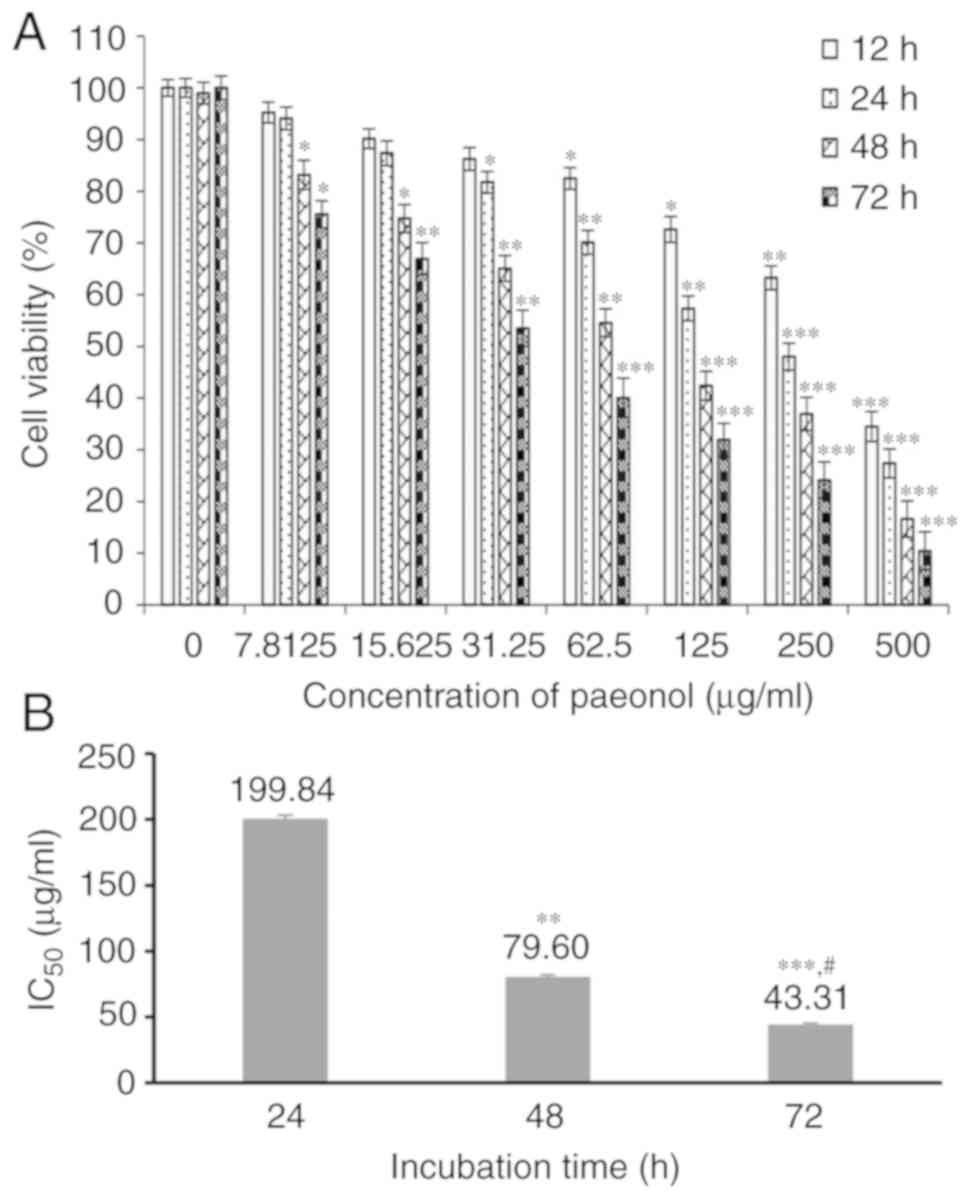
After incubation with paeonol for various intervals
(12-72 h), it was demonstrated that paeonol significantly
suppressed the proliferation of HCT116 cells, and the number of
viable cells was inhibited with increasing concentrations of
paeonol and the prolongation of incubation time (P<0.05;
Fig. 1A). Moreover, the
IC50 of paeonol was determined as 199.84 µg/ml at
24 h, 79.60 µg/ml at 48 h and 43.31 µg/ml at 72 h
(Fig. 1B). Collectively, these
results suggested that paeonol reduced the number of viable HCT116
cells in a dose- and time-dependent.
Paeonol induces
G0/G1-phase arrest in HCT116 cells
As cell proliferation is closely regulated by the
cell cycle (22), FCM was used to
assess whether the effect of paeonol on inhibiting cell
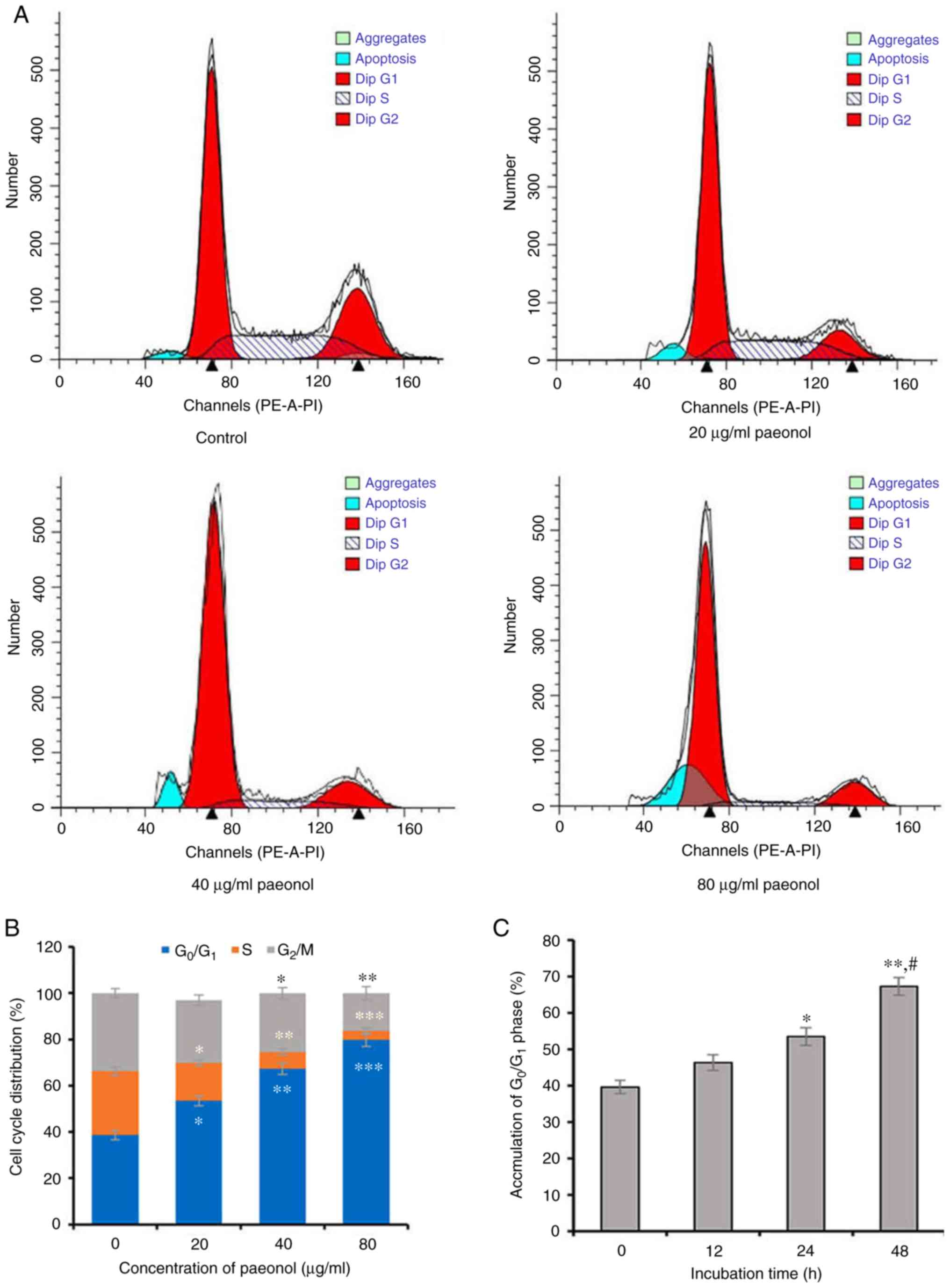
proliferation was due to induction of cell cycle arrest. Following
incubation with 0, 20, 40 and 80 µg/ml paeonol for 48 h, the
FCM results indicated that the proportion of HCT116 cells in
G0/G1 phase was 53.42±2.14, 67.37±2.43 and
79.78±2.86%, respectively, which was significantly higher compared
with the control group (38.68±1.96%; all P<0.05), demonstrating
that paeonol dose-dependently induced a significant accumulation of
HCT116 cells in G0/G1 phase. Furthermore, the
cell cycle profile of HCT116 cells exposed to different doses of
paeonol exhibited a distinctive broad sub-diploid DNA
(sub-G1) peak at 48 h, which was significantly different
compared with the control cells (Fig.
2A). The accumulation of HCT116 cells in
G0/G1 phase was also accompanied by
corresponding decreased percentages in the S and G2/M
phases (Fig. 2A and B). In
addition, the proportion of HCT116 cells in
G0/G1 phase was time-dependently increased in
the presence of paeonol (Fig.
2C), thus suggesting that paeonol dose- and time-dependently
inhibited the proliferation of HCT116 cells by causing
G0/G1-phase arrest.
Paeonol induces apoptosis in HCT116
cells
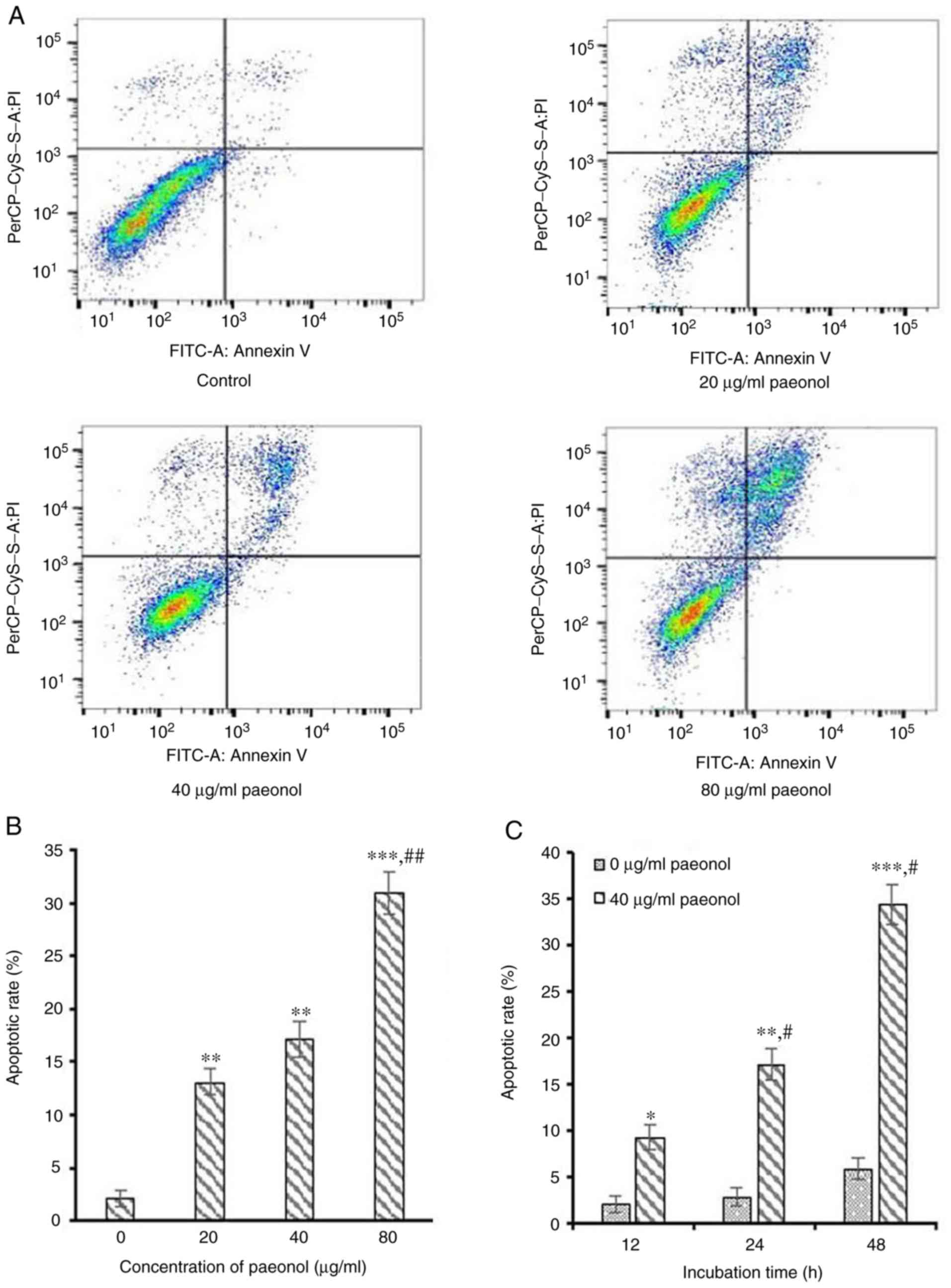
To assess whether cell apoptosis was responsible for
the reduction of viable cells following incubation with paeonol, an
Annexin V-FITC/PI double staining assay with FCM analysis was used
to monitor phosphatidylserine exposure. Following incubation with
20, 40 and 80 µg/ml paeonol for 24 h, the FCM results
indicated that the apoptotic rate of HCT116 cells was 13.07±1.23,
17.13±1.65 and 30.97±2.01%, respectively, which was significantly
higher compared with the control group 4.20±0.83% (all P<0.05;
Fig. 3A and B), indicating that
paeonol dose-dependently induced apoptosis of CRC cells. Moreover,
the apoptotic rates increased with longer exposure time of paeonol
(Fig. 3C). Therefore, the results
indicated that paeonol dose- and time-dependently promoted
apoptosis of CRC cells, which may be one of the underlying
mechanisms of the anti-cancer activity of paeonol.
Paeonol induces cell apoptosis via the
caspase-dependent pathway
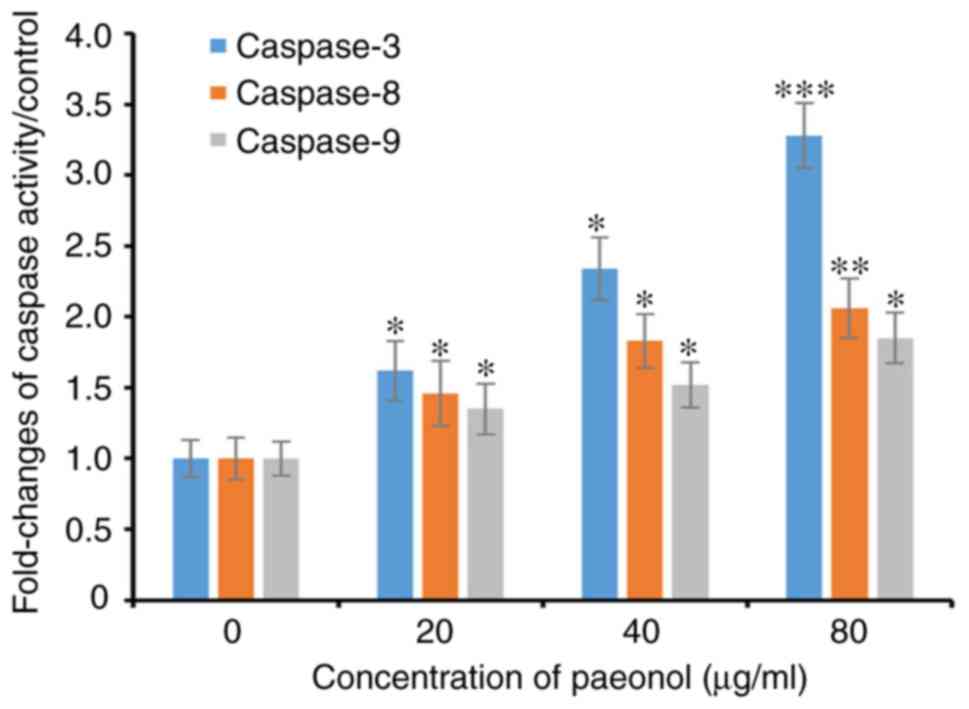
Following incubation with increasing concentrations
of paeonol for 48 h, caspase-3 activity gradually increased in
HCT116 cells and was higher compared with the control group
(P<0.05). In addition, similar trends for caspase-8 and
caspase-9 were observed in HCT116 cells exposed to paeonol for 48 h
(Fig. 4). Collectively, it was
demonstrated that paeonol dose-dependently increased the activity
of caspase-3, -8 and -9.
Paeonol represses the β-catenin-mediated
transcriptional activity of TCF/LEF
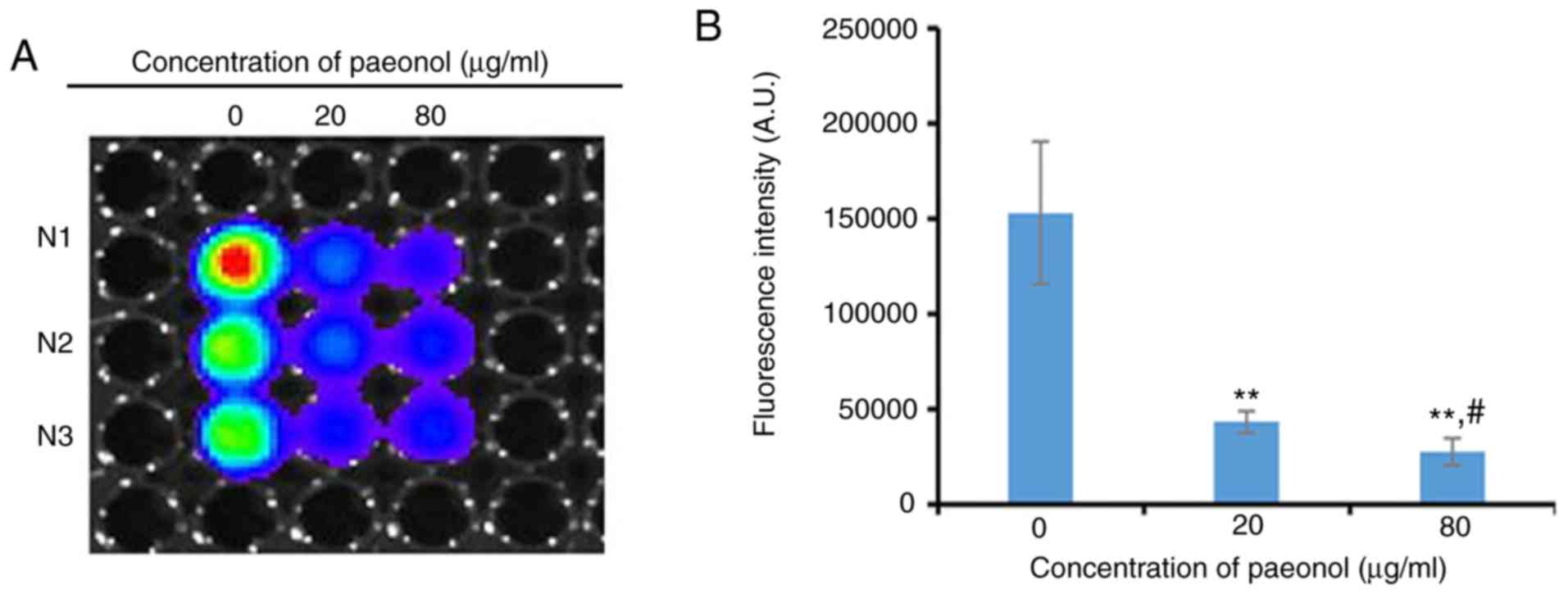
To examine the effect of paeonol on the activity of
TCF/LEF mediated by β-catenin, a TCF/LEF luciferase reporter assay
was performed. With increasing concentrations of paeonol, the
luciferase activity gradually decreased and there was a significant
difference between the 20 µg/ml paeonol-treated group and
the control group (P<0.05). Furthermore, the luciferase activity
following exposure to 80 µg/ml paeonol was significantly
weaker compared with the group treated with 20 µg/ml paeonol
(P<0.05; Fig. 5), suggesting
that a high concentration of paeonol significantly repressed the
transcriptional activity of TCF/LEF.
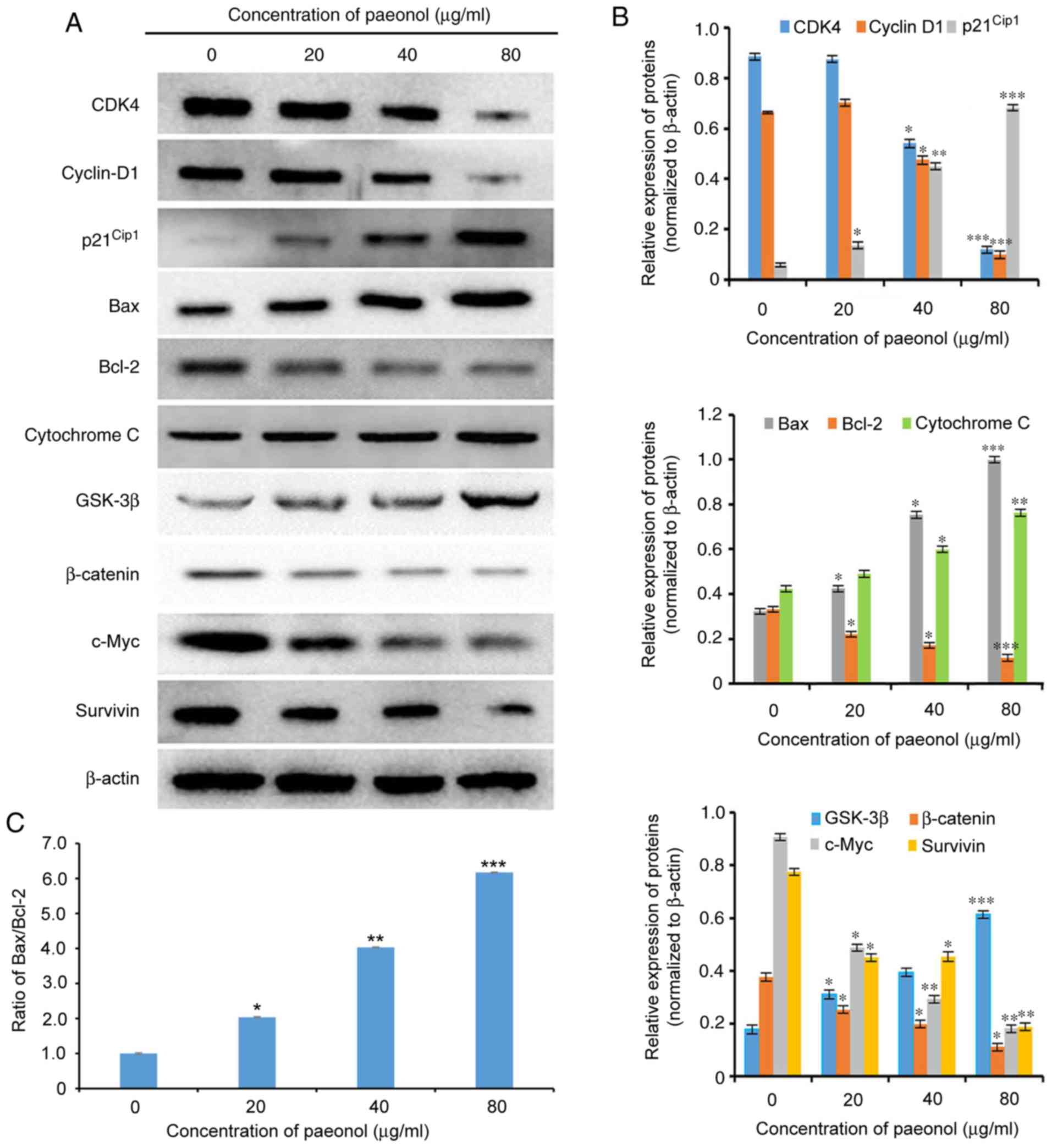
Paeonol inhibits proliferation via the
Wnt/β-catenin signaling pathway
To elucidate the possible mechanism responsible for
the inhibition of transition to the DNA synthesis phase, the cell
cycle-associated proteins, including cyclin D1, CDK4 and
p21Cip1, which are able to promote cell cycle
progression (23-24), were further investigated by
western blot analysis. The protein expression levels of cyclin D1
and CDK4 were significantly downregulated, while p21Cip1
expression was upregulated in HCT116 cells exposed to paeonol for
48 h (Fig. 6). Thus, paeonol may
be able to cause G0/G1 phase arrest at least
partly due to interference of the expression levels of the key
G1-regulatory proteins CDK4, cyclin D1 and
p21Cip1.
To investigate the mechanisms underlying the
anti-tumor effect of paeonol against HCT116 cells, which were via
inducing cell apoptosis, western blot analysis was used to detect
the expression levels of apoptosis-associated proteins. Following
incubation with 20, 40 and 80 µg/ml paeonol for 48 h, the
expression levels of Bax and cytochrome C were significantly
upregulated, while those of Bcl-2 were significantly downregulated
in HCT116 cells in a dose-dependent manner (Fig. 6). Moreover, the Bax/Bcl-2 ratio
was elevated compared with the control group and was
dose-dependently increased by paeonol in HCT116 cells (P<0.05).
Collectively, the results indicated that paeonol induced cell
apoptosis via increasing the Bax/Bcl-2 ratio in HCT116 cells.
To further elucidate whether paeonol exerts its
anti-tumor effect against HCT116 cells via the Wnt/β-catenin
pathway, the protein expression levels of β-catenin, as well as its
down-stream signaling molecules cyclin D1, c-Myc and survivin
proto-oncogene, were determined by western blot analysis. Following
incubation with 20, 40 and 80 µg/ml paeonol for 48 h, the
expression levels of β-catenin, c-Myc and survivin were
dose-dependently downregulated, while those of GSK-3β were
dose-dependently upregulated in HCT116 cells compared with the
control group (P<0.05; Fig.
6). In addition, paeonol significantly inhibited the expression
of cyclin D1 compared with the control group (P<0.05). Moreover,
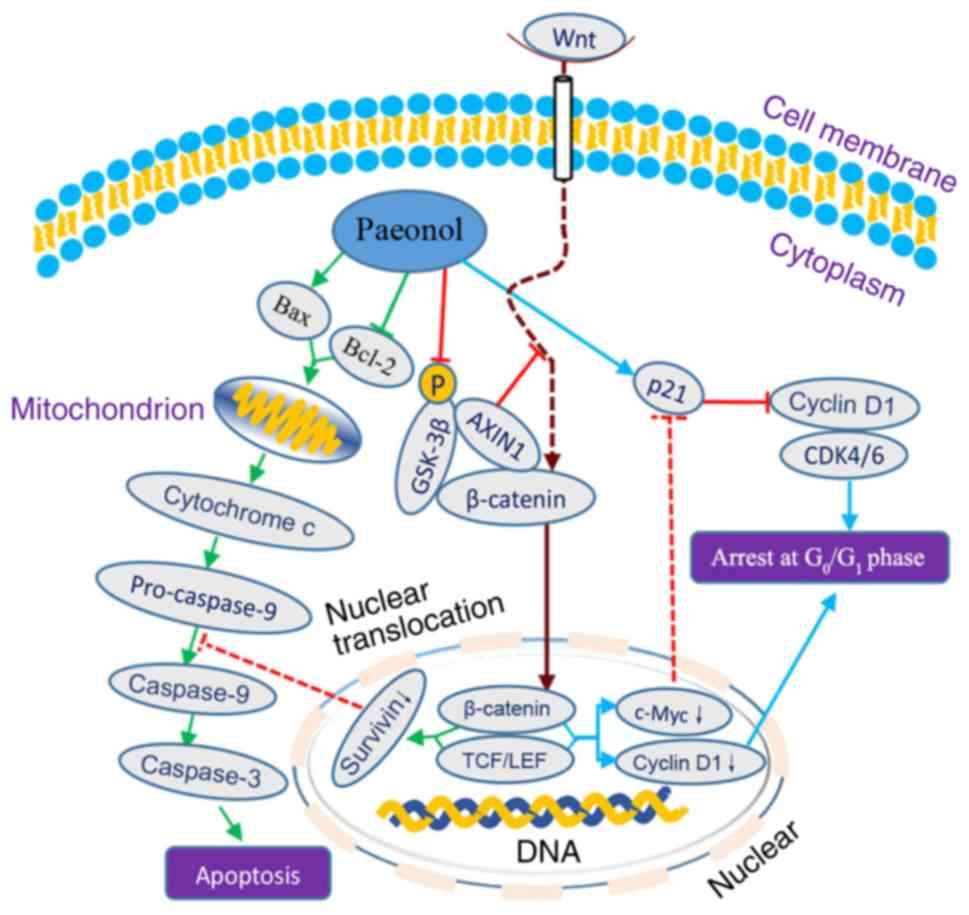
a possible mechanism responsible for the effects of paeonol against
human CRC cells is schematically presented in Fig. 7.
Discussion
Despite progress in systemic anti-cancer therapy,
the effective treatment of CRC remains a major clinical challenge
due to its high mortality and metastasis potential (25). Active components, including
hydnocarpin (15), aesculetin
(16), baicalin (17), isobavachalcone (18), wogonin (20) and lycorine (26), of Traditional Chinese Medicine
formulations have attracted increased attention worldwide due to
their unique advantages over western drugs in cancer treatment
(26). For instance, experimental
data have revealed that paeonol has mild anti-tumor activities
(10,27,28). In the present study, paeonol time-
and dose-dependently suppressed the viability of HCT116 cells, with
an IC50 of 79.60 µg/ml at 48 h; these results are
consistent with other CRC cell lines (LoVo and SW620) (29). Moreover, paeonol has been shown to
be a relatively safe medicine in mice with a median lethal dose of
3,430 mg/kg (30). Collectively,
these experimental data suggest that paeonol may be a novel
candidate for anti-cancer therapy.
Cell proliferation is closely regulated by the cell
cycle (22), and disruption of
the cell cycle may inhibit cell proliferation and suppress tumor
growth. The present results indicated that paeonol induced an
accumulation in the G0/G1 phase, accompanied
by a concomitant decrease in S and G2/M phases in HCT116
cells, which is consistent with findings in other cancer cell lines
(11,31). Cell cycle control is regulated by
CDKs, cyclins and CDK inhibitors, such as p21Cip1 and
p27Kip1 (23), and the
activity of cyclin-CDK complexes may be affected by multiple
signaling pathways (32).
Furthermore, binding of cyclin D1 to CDK4 or CDK6 causes the
formation of the cyclin D1-CDK4/6 complex, eventually driving cell
transition from G0/G1 to S phase (33). In the present study, the
expression levels of cyclin D1 and CDK4 were downregulated and that
of p21Cip1 was upregulated, subsequently blocking the
cell cycle procession. Thus, it was hypothesized that paeonol
exerted anti-proliferative effects by blocking cell cycle
transition from G1 phase to S phase.
Induction of apoptosis is one of the most important
and direct pathways that controls and eliminates cancer
proliferation (11,34). The FCM results of the present
study demonstrated that paeonol dose- and time-dependently induced
cell apoptosis, which was in line with previous studies of CRC
cells (29) and other cancer
cells (11,31). In addition, HCT116 cells exposed
to paeonol exhibited a distinctive broad sub-G1 peak at
48 h, the appearance of which is usually regarded as a result of
the degradation of nuclear DNA in the early stages of cell
apoptosis (35). Although the
mechanisms of apoptosis are complex, it has been reported that
mitochondrial (intrinsic) and cell-surface death receptor-mediated
(extrinsic) apoptosis are the two principal pathways (36). Bcl-2 family proteins have a
significant role in the intrinsic apoptotic pathway, during which
the imbalance between pro- and anti-apoptotic proteins determines
the ultimate fate of cancer cells (37). Bax causes mitochondrial
disruption, release of cytochrome c, activation of the downstream
caspase-9 and, ultimately, activation of caspase-3 (38). The present results demonstrated an
increase in the Bax/Bcl-2 ratio, via which the apoptotic effect of
paeonol on HCT116 cells was exerted. To investigate which pathway
was responsible for cell apoptosis, the activities of caspase-3, -8
and -9 were detected using a colorimetric caspase assay. Following
incubation for 48 h, caspase-3, -8 and -9 activities in HCT116
cells were enhanced with increasing doses of paeonol in comparison
with those in the control group. All of these results are
consistent to a previous study, which showed that paeonol induces
cell apoptosis via suppressing the expression of Bcl-2 and
increasing the expression levels of Bax, caspase-8 and caspase-3
(39). Furthermore, the
aforementioned results suggested that the suppressive effect of
paeonol on the viability of CRC cells was associated with the
induction of apoptosis via the caspase pathway.
In most healthy cells, the Wnt pathway is commonly
inactive and β-catenin is sequestered in the cytoplasm by a
multi-protein complex containing axin 1 (AXIN1), APC regulator of
Wnt signaling pathway, casein kinase 1 α and GSK-3β (40), resulting in a low level of
β-catenin in the nucleus. Activated Wnt may cause the translocation
of β-catenin from the cell cytoplasm to the nucleus, where it
activates the β-catenin-mediated LEF/TCF transcriptional machinery,
thus inducing the transcription of TCF/LEF-responsive genes, such
as c-Myc (41) and cyclin D1
(42). Moreover, abnormal
activation of the Wnt signaling pathway, a known hallmark of CRC,
has been reported to be associated with cell proliferation, cell
cycle and cell apoptosis, as a result of an activated canonical
β-catenin and LEF/TCF pathway (43), in which suppressing the expression
of β-catenin has a beneficial anti-tumor effect (44). In the present study, the
expression level of β-catenin was significantly downregulated,
while that of GSK-3β was upregulated by paeonol in a dose-dependent
manner.
c-Myc, a downstream effector of the β-catenin
pathway, has been revealed to be upregulated in ~30% of cancer
cells and is associated with cancer progression (45). With the disruption of
β-catenin/TCF activity, decreased c-Myc may cause the transcription
of p21Cip1/Waf1, which in turn promotes cell cycle
arrest at the G0/G1 phase and cell
differentiation (46).
Furthermore, cyclin D1, another downstream effector of the
β-catenin pathway, exerts a vital role in regulating the cell cycle
progression in different types of cells (47,48). The activated β-catenin signaling
pathway may also stimulate the transcription of cyclin D1, which is
an important marker for cells undergoing mitosis (49). The present findings identified the
roles of c-Myc and cyclin D1 in cell cycle regulation, as
demonstrated by the decreased TCF/LEF activity and concomitant
downregulation of c-Myc and cyclin D1 protein. Apart from c-Myc and
cyclin D1, survivin, an inhibitor of apoptosis, was identified as
another target gene that is implicated in suppressing cell
proliferation and regulating the cell life span (50). A previous study reported that
downregulation of c-Myc, cyclin D1 and survivin may be an effective
treatment strategy for CRC (51).
Induction of TCF target gene transcription via activating the
β-catenin pathway in CRC constitutes the primary transforming
event, while TCF transcriptional activity may be reduced via
suppressing the expression of β-catenin, subsequently followed by
cell apoptosis via caspase-3 activation (52). The present results are in line
with a previous study, reporting that blocking the interaction
between β-catenin and TCF induced pancreatic cancer cell apoptosis
via decreasing the expression levels of c-Myc and cyclin D1
(52). In addition, antagonism of
Wnt/β-catenin signaling occurs at 20, 40 and 80 µg/ml
paeonol to those required to suppress cell proliferation, block the
cell cycle at G0/G1 phase and induce
apoptosis in HCT116 cells. A schematic illustration of the possible
mechanism under-lying the anti-tumor activity of paeonol against
CRC cells, including the induction of G0/G1
phase arrest and apoptosis via suppressing the Wnt/β-catenin
signaling pathway is presented in Fig. 7. However, the lack of multiple
cell lines to assess the present findings is a limitation of the
current study. In addition, the suppressive effect of AXIN1 on Wnt
signaling pathways and the intervention of Wnt/β-catenin signaling
pathways on the cell cycle and apoptosis require further
investigation in future studies.
In conclusion, the present results indicated that
paeonol exerted an anti-tumor effect against CRC cells, which, at
least partly, involved the blockage of the Wnt/β-catenin signaling
pathway. Therefore, the current findings support the use of paeonol
as a novel treatment for CRC, acting via distinct mechanisms.
However, in subsequent studies, the results of the present study
should be verified using multiple cell lines and the long-term
effects of paeonol are required to be assessed in vivo.
Acknowledgements
Not applicable.
Funding
This study was financially supported by Jiangsu
Provincial Administration of Traditional Chinese Medicine (grant
no. YB2017099).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article or are available from
the corresponding author on reasonable request.
Authors' contributions
LHL and RJS contributed to the conception and design
of the study. LHL and ZCC performed the experiments and contributed
to data analysis. LHL drafted the manuscript and RJS revised the
paper. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Nguyen MN, Choi TG, Nguyen DT, Kim JH, Jo
YH, Shahid M, Akter S, Aryal SN, Yoo JY, Ahn YJ, et al: CRC-113
gene expression signature for predicting prognosis in patients with
colorectal cancer. Oncotarget. 6:31674–31692. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Feng RM, Zong YN, Cao SM and Xu RH:
Current cancer situation in China: Good or bad news from the 2018
global cancer statistics? Cancer Commun (Lond). 39:222019.
View Article : Google Scholar
|
|
3
|
Li L and Ma BB: Colorectal cancer in
Chinese patients: Current and emerging treatment options. Onco
Targets Ther. 7:1817–1828. 2014.PubMed/NCBI
|
|
4
|
Wang P, Gao XY, Yang SQ, Sun ZX, Dian LL,
Qasim M, Phyo AT, Liang ZS and Sun YF: Jatrorrhizine inhibits
colorectal carcinoma proliferation and metastasis through
Wnt/β-catenin signaling pathway and epithelial-mesenchymal
transition. Drug Des Dev Ther. 13:2235–2247. 2019. View Article : Google Scholar
|
|
5
|
Stintzing S: Management of colorectal
cancer. F1000Prime Rep. 6:1082014. View
Article : Google Scholar
|
|
6
|
Koosha S, Alshawsh MA, Looi CY, Seyedan A
and Mohamed Z: An association map on the effect of flavonoids on
the signaling pathways in colorectal cancer. Int J Med Sci.
13:374–385. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen Z, Zhang B, Gao F and Shi R:
Modulation of G2/M cell cycle arrest and apoptosis by
luteolin in human colon cancer cells and xenografts. Oncol Lett.
15:1559–1565. 2018.PubMed/NCBI
|
|
8
|
Lou Y, Wang C, Tang Q, Zheng W, Feng Z, Yu
X, Guo X and Wang J: Paeonol inhibits IL-1β-induced inflammation
via PI3K/Akt/NF-κB pathways: In vivo and vitro studies.
Inflammation. 40:1698–1706. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen B, Ning M and Yang G: Effect of
paeonol on antioxidant and immune regulatory activity in
hepatocellular carcinoma rats. Molecules. 17:4672–4683. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fu J, Yu LH, Luo J, Huo R and Zhu B:
Paeonol induces the apoptosis of the SGC-7901 gastric cancer cell
line by downregulating ERBB2 and inhibiting the NF-κB signaling
pathway. Inter J Mol Mede. 42:1473–1483. 2018.
|
|
11
|
Saahene RO, Wang J, Wang ML, Agbo E and
Pang D: The anti-tumor mechanism of paeonol on CXCL4/CXCR3-B
signals in breast cancer through induction of tumor cell apoptosis.
Cancer Biother Radiopharm. 33:233–240. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu J, Xue X, Zhang B, Jiang W, Cao H, Wang
R, Sun D and Guo R: The protective effects of paeonol against
epirubicin-induced hepatotoxicity in 4T1-tumor bearing mice via
inhibition of the PI3K/Akt/NF-kB pathway. Chem Biol Interact.
244:1–8. 2016. View Article : Google Scholar
|
|
13
|
He X, Han W, Hu SX, Zhang MZ, Hua JL and
Peng S: Canonical Wnt signaling pathway contributes to the
proliferation and survival in porcine pancreatic stem cells (PSCs).
Cell Tissue Res. 362:379–388. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu N, Jiang F, Han XY, Li M, Chen WJ, Liu
QC, Liao CX and Lv YF: MiRNA-155 promotes the invasion of
colorectal cancer SW-480 cells through regulating the
Wnt/β-catenin. Eur Rev Med Pharmacol Sci. 22:101–109.
2018.PubMed/NCBI
|
|
15
|
Lee MA, Kim WK, Park HJ, Kang SS and Lee
SK: Anti-proliferative activity of hydnocarpin, a natural lignan,
is associated with the suppression of Wnt/β-catenin signaling
pathway in colon cancer cells. Bioorg Med Chem Lett. 23:5511–5514.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li T, Zhang L and Huo X: Inhibitory
effects of aesculetin on the proliferation of colon cancer cells by
the Wnt/β-catenin signaling pathway. Oncol Lett. 15:7118–7122.
2018.PubMed/NCBI
|
|
17
|
Jia Y, Chen L, Guo S and Li Y: Baicalin
induced colon cancer cells apoptosis through miR-217/DKK1-mediated
inhibition of Wnt signaling pathway. Mol Biol Rep. 46:1693–1700.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li Y, Qin X, Li P, Zhang H, Lin T, Miao Z
and Ma S: Isobavachalcone isolated from Psoralea corylifolia
inhibits cell proliferation and induces apoptosis via inhibiting
the AKT/GSK-3β/β-catenin pathway in colorectal cancer cells. Drug
Des Dev Ther. 13:1449–1460. 2019. View Article : Google Scholar
|
|
19
|
Liu MP, Li W, Dai C, Lam CWK, Li Z, Chen
JF, Chen ZG, Zhang W and Yao MC: Aqueous extract of Sanguisorba
officinalis blocks the Wnt/β-catenin signaling pathway in
colorectal cancer cells. RSC Adv. 8:10197–10206. 2018. View Article : Google Scholar
|
|
20
|
He L, Lu N, Dai Q, Zhao Y, Zhao L, Wang H,
Li Z, You Q and Guo Q: Wogonin induced G1 cell cycle arrest by
regulating Wnt/β-catenin signaling pathway and inactivating CDK8 in
human colorectal cancer carcinoma cells. Toxicology. 312:36–47.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lu H, Gao F, Shu G, Xia G, Shao Z, Lu H
and Chen K: Wogonin inhibits the proliferation of myelodysplastic
syndrome cells through the induction of cell cycle arrest and
apoptosis. Mol Med Rep. 12:7285–7292. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu W and McArthur G: Cell cycle regulation
and melanoma. Curr Oncol Rep. 18:342016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tane S, Ikenishi A, Okayama H, Iwamoto N,
Nakayama KI and Takeuchi T: CDK inhibitors, p21(Cip1) and
p27(Kip1), participate in cell cycle exit of mammalian
cardiomyocytes. Biochem and Bioph Res Co. 443:1105–1109. 2014.
View Article : Google Scholar
|
|
24
|
Gao N, Flynn DC, Zhang Z, Zhong XS, Walker
V, Liu KJ, Shi X and Jiang BH: G1 cell cycle progression and the
expression of G1 cyclins are regulated by PI3K/AKT/mTOR/p70S6K1
signaling in human ovarian cancer cells. Am J Physiol Cell Physiol.
287:C281–C291. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Akbari A, Amanpour S, Muhammadnejad S,
Ghahremani MH, Ghaffari SH, Dehpour AR, Mobini GR, Shidfar F,
Abastabar M, Khoshzaban A, et al: Evaluation of antitumor activity
of a TGF-beta receptor I inhibitor (SD-208) on human colon
adeno-carcinoma. Daru. 22:472014. View Article : Google Scholar
|
|
26
|
Hu H, Wang S, Shi D, Zhong B, Huang X, Shi
C and Shao Z: Lycorine exerts antitumor activity against
osteosarcoma cells in vitro and in vivo xenograft model through the
JAK2/STAT3 pathway. OncoTargets Ther. 12:5377–5388. 2019.
View Article : Google Scholar
|
|
27
|
Zhang L, Tao L, Shi T, Zhang F, Sheng X,
Cao Y, Zheng S, Wang A, Qian W, Jiang L and Lu Y: Paeonol inhibits
B16F10 melanoma metastasis in vitro and in vivo via disrupting
proinflammatory cytokines-mediated NF-κB and STAT3 pathways. IUBMB
Life. 67:778–788. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu Y, Zhu JY, Lei ZM, Wan LJ, Zhu XW, Ye F
and Tong YY: Anti-proliferative effects of paeonol on human
prostate cancer cell lines DU145 and PC-3. J Physiol Biochem.
73:157–165. 2017. View Article : Google Scholar
|
|
29
|
Li M, Tan SY and Wang XF: Paeonol exerts
an anticancer effect on human colorectal cancer cells through
inhibition of PGE2 synthesis and COX-2 expression. Oncol
Rep. 32:2845–2853. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang LH, Xiao PG and Huang Y: Recent
progresses in pharmacological and clinical studies of paeonol.
Zhongguo Zhong Xi Yi Jie He Za Zhi. 16:187–190. 1996.In Chinese.
PubMed/NCBI
|
|
31
|
Yang S, Wang X and Zhong G: Paeonol
inhibits the growth of gastric cancer cells via suppressing HULC
expression. Int J Clin Exp Med. 9:13900–13908. 2016.
|
|
32
|
Wang S, Wang X, Gao Y, Peng Y, Dong N, Xie
Q, Zhang X, Wu Y, Li M and Li J: RN181 is a tumour suppressor in
gastric cancer by regulation of the ERK/MAPK-cyclin D1/CDK4
pathway. J Pathol. 248:204–216. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Obaya AJ, Mateyak MK and Sedivy JM:
Mysterious liaisons: The relationship between c-Myc and the cell
cycle. Oncogene. 18:2934–2941. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li Q, Zhang Y, Sun J and Bo Q:
Paeonol-mediated apoptosis of hepatocellular carcinoma cells by
NF-κB pathway. Oncol Lett. 17:1761–1767. 2019.PubMed/NCBI
|
|
35
|
Qian L, Murakami T, Kimura Y, Takahashi M
and Okita K: Saikosaponin A-induced cell death of a human hepatoma
cell line (HuH-7): The significance of the 'sub-G1 peak' in a DNA
histogram. Pathol Int. 45:207–214. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ghobrial IM, Witzig TE and Adjei AA:
Targeting apoptosis pathways in cancer therapy. CA Cancer J Clin.
55:178–194. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Boise LH, González-García M, Postema CE,
Ding L, Lindsten T, Turka LA, Mao X, Nuñez G and Thompson CB:
bcl-x, a bcl-2-related gene that functions as a dominant regulator
of apoptotic cell death. Cell. 74:597–608. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jia L, Macey MG, Yin Y, Newland AC and
Kelsey SM: Subcellular distribution and redistribution of Bcl-2
family proteins in human leukemia cells undergoing apoptosis.
Blood. 93:2353–2359. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ou Y, Li Q, Wang J, Li K and Zhou S:
Antitumor and apoptosis induction effects of paeonol on mice
bearing EMT6 breast carcinoma. Biomol Ther (Seoul). 22:341–346.
2014. View Article : Google Scholar
|
|
40
|
Li DW, Liu ZQ, Chen W, Yao M and Li GR:
Association of glycogen synthase kinase-3β with Parkinson's disease
(Review). Mol Med Rep. 9:2043–2050. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shi L, Wu YX, Yu JH, Chen X, Luo XJ and
Yin YR: Research of the relationship between β-catenin and
c-myc-mediated Wnt pathway and laterally spreading tumors
occurrence. Eur Rev Med Pharmacol Sci. 21:252–257. 2017.PubMed/NCBI
|
|
42
|
Chen Y, Jiang T, Shi L and He K: hcrcn81
promotes cell proliferation through Wnt signaling pathway in
colorectal cancer. Med Oncol. 33:32016. View Article : Google Scholar
|
|
43
|
Krishnamurthy N and Kurzrock R: Targeting
the Wnt/beta-catenin pathway in cancer: Update on effectors and
inhibitors. Cancer Treat Rev. 62:50–60. 2018. View Article : Google Scholar
|
|
44
|
Shen Y, Wang Q and Tian Y: Reversal effect
of ouabain on multi-drug resistance in esophageal carcinoma
EC109/CDDP cells by inhibiting the translocation of Wnt/β-catenin
into the nucleus. Tumor Biol. 37:15937–15947. 2016. View Article : Google Scholar
|
|
45
|
Xiao ZD, Han L, Lee H, Zhuang L, Zhang Y,
Baddour J, Nagrath D, Wood CG, Gu J, Wu X, et al: Energy
stress-induced lncRNA FILNC1 represses c-Myc-mediated energy
metabolism and inhibits renal tumor development. Nat Commun.
8:7832017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
van de Wetering M, Sancho E, Verweij C, de
Lau W, Oving I, Hurlstone A, van der Horn K, Batlle E, Coudreuse D,
Haramis AP, et al: The beta-catenin/TCF-4 complex imposes a crypt
progenitor phenotype on colorectal cancer cells. Cell. 111:241–250.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cai F, Chen P, Chen L, Biskup E, Liu Y,
Chen PC, Chang JF, Jiang W, Jing Y, Chen Y, et al: Human RAD6
promotes G1-S transition and cell proliferation through
upregulation of cyclin D1 expression. PLoS One. 9:e1137272014.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang H, Zhang X, Ji S, Hao C, Mu Y, Sun J
and Hao J: Sohlh2 inhibits ovarian cancer cell proliferation by
upregulation of p21 and downregulation of cyclin D1.
Carcinogenesis. 35:1863–1871. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tetsu O and McCormick F: Beta-catenin
regulates expression of cyclin D1 in colon carcinoma cells. Nature.
398:422–426. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ho YS, Tsai WH, Lin FC, Huang WP, Lin LC,
Wu SM, Liu YR and Chen WP: Cardioprotective Actions of TGFβRI
Inhibition Through Stimulating Autocrine/Paracrine of Survivin and
Inhibiting Wnt in Cardiac progenitors. Stem Cells. 34:445–455.
2016. View Article : Google Scholar
|
|
51
|
Yeh CT, Yao CJ, Yan JL, Chuang SE and Lai
GM, Chen CM, Yeh CF, Li CH and Lai GM: Apoptotic cell death and
inhibition of Wnt/β-catenin signaling pathway in human colon cancer
cells by an active fraction (HS7) from Taiwanofungus camphoratus.
Evid Based Complement Alternat Med. 2011:7502302011. View Article : Google Scholar
|
|
52
|
Garg B, Giri B, Majumder K, Dudeja V,
Banerjee S and Saluja A: Modulation of post-translational
modifications in β-catenin and LRP6 inhibits Wnt signaling pathway
in pancreatic cancer. Cancer Lett. 388:64–72. 2017. View Article : Google Scholar
|