Introduction
Allogenic bone marrow (BM) transplantation (BMT) is
the standard treatment for a number of hematopoietic disorders,
such as aplastic anemia and leukemia. However, several challenges
related to allogenic BMT exist, including cytotoxic conditioning
effects, infections, relapse and graft vs. host disease (GVHD). To
reduce the occurrence of such complications, approaches for
depleting graft T cells, partially depleting specific T cell
subsets, and ex vivo manipulation of donor T cells have been
developed (1,2). Although these methods have limited
BMT-associated toxicity and GVHD, T cell depletion also negatively
affects the effectiveness of adaptive immunity against viruses,
fungal pathogens, and cancer cells.
Following allogenic BMT, the recovery of myeloid
cells that participate in innate immunity occurs within weeks or
months, whereas lymphoid cells for adaptive immunity may require up
to 2 years for recovery based on quantitative and qualitative
reconstitution studies of functional T cell compartments following
BMT (3). T cell recovery
following BMT is accomplished through two pathways. In the
thymus-independent pathway, the initial recovery of T cells for
regaining immune competency following allogenic BMT primarily
involves the peripheral expansion of memory T cells transferred
from the donor T cell pool or host cells that survive pre-BMT
cytotoxic conditioning. The conditioning regimen is used to secure
the available space of donor graft following BM cell depletion and
reduce overall tumor mass in the recipient. Alternatively, the
thymus-dependent pathway leads to the eventual reconstitution of a
full repertoire of diverse, self-tolerant and naïve T cells from
the host thymus via the de novo production of T cells
(4). In the thymus-dependent
recovery of T cells, crosstalk between thymic stromal cells and
developing thymocytes must be regulated. However, this regulation
can be restricted by damaged or altered thymic niches due to
pre-conditioning regimens, infections, GVHD, or recipient age
(5,6). The function of thymic epithelial
cells (TECs) in T cell development is related to the development of
immature thymocytes into competent T cells that respond to foreign
antigens, but are self-tolerant. Essential extracellular factors
for TEC development include fibroblast growth factor (FGF)7
(7,8) and FGF10 from mesenchymal cells.
Thymus growth is attenuated in mice lacking FGF-R2IIIb, a receptor
for FGF7 and FGF10 (9). In
addition, FGF7 administration in GVHD mice has been shown to exert
a protective effect on the thymic epithelium (10), and bone morphogenic protein 4 from
thymic endothelial cells contributes to endogenous regeneration
following thymic damage (11).
Furthermore, medullary thymic epithelial cells can transfer host
antigens to CD8α dendritic cells via CD36 to induce tolerance
following allogenic BMT (12).
FMS-like tyrosine kinase 3 ligand (FLT3) is a receptor tyrosine
kinase homologous to c-Kit and c-fms and is expressed on
hematopoietic progenitor cells. The ligand of FLT3, FLT3L, is
important for hematopoietic stem cell generation and survival in
vitro (13) and for de
novo thymus-derived T cell development (14). FLT3L administration increases the
numbers of LSK cells, and early thymocyte progenitor precursors
leads to thymopoiesis following BMT (15). For naïve T cells, IL-7 is
essential for proliferation and maintenance in the periphery
(16). For memory CD4+
T cells, IL-7 and TCR stimulation is critical (17). By contrast, memory CD8+
T cell maintenance mainly depends on IL-15, although TCR
stimulation is dispensable (18).
Thus, IL-7 and IL-15 primarily affect thymus-independent or
peripheral reconstitution of T cells after BMT.
Delayed T cell recovery and restricted T cell
diversity following allogenic BMT are associated with an increased
risk of infection and cancer recurrence. To accelerate post-BMT,
the thymus-dependent recovery of T cells, thymic reconstitution
must be preceded by stimulation of the thymic niche and its output
(i.e., recent thymic emigrants) by expanding memory T cells through
cytokine action and through enhancing naïve T cell production by
the recipient's thymus.
Previously, several immune regulatory effects of
tonsil-derived mesenchymal stromal cells (T-MSCs) in BMT and
inflammatory disease mouse models were reported. For example, TSG-6
released from T-MSCs attenuates acute GVHD responses in mice,
represented by rapid reversal of weight loss and improved
histological scoring of damaged organs (19). PD-L1 expressed by T-MSCs prevents
Th17 differentiation and neutrophil-mediated inflammation (20,21), and T-MSCs also inhibit dendritic
cell maturation and CD4+ cell differentiation (22). As these effects of T-MSCs were
observed in the context of adaptive immune responses of mature T
cells, the possible effect of T-MSCs on the recovery of the thymus
during allogenic BMT was thus examined.
The present study focused primarily on the ability
of T-MSC co-transplantation to reconstitute the T cell repertoire
in a thymus-dependent manner in mice undergoing allogenic BMT. The
possible mechanism(s) of the effects T-MSCs on thymic regeneration
following allogenic BMT were also investigated by comparatively
analyzing the effects of MSCs from various origins, such as the BM
or adipose tissue.
Materials and methods
Animals
In the present study, 8-week-old female BALB/c
(n=40) and male C57BL/6 (n=14) mice (body weight, 18±2 g) were
purchased from OrientBio. All animals were housed at 21-23°C with
51-54% humidity in a pathogen-free environment on a 12-h light/dark
cycle and allowed free access to food and water. The experimental
procedures were approved by the Animal Care and Use Committee of
the College of Medicine, Ewha Womans University (Seoul, Korea;
approval no. ESM18-0403).
Cells and cell culture
BM-MSCs, adipose tissue-derived MSCs (AT-MSCs) and
T-MSCs were cultured in low-glucose Dulbecco's modified Eagle
medium (DMEM; Welgene) in 100-mm cell culture plates. The BM-MSCs
were purchased from the Severance Hospital Cell Therapy Center and
the AT-MSCs were purchased from the American Type Culture
Collection (ATCC). In the present study the T-MSCs were obtained
previously (IRB File no. EUMC 2018-01-011-002) and maintained from
the same patients as previously described (23). To prepare for cell injection,
cells were collected at 80% confluence with trypsin treatment
followed by 3 washes with PBS. Prior to injection with BM cells
(BMCs) into the mice, MSCs were resuspended with low-glucose DMEM
at a final volume of 200 µl for tail vein injection. To
generate conditioned medium (CM) to analyze cellular soluble
factors, cells at 80% confluence were washed four times with PBS,
and the medium was replaced with serum-free DMEM. The medium was
collected after 48 h of culture as previously described (19), centrifuged at 190 × g for 5 min at
25°C, passed through a 0.2-µm filter (Merck Millipore), and
concentrated 20-fold by high-speed centrifugal filtration (Sorvall
LYNX4000, Thermo Fisher Scientific, Inc.). The concentrated CM was
then frozen and stored at -80°C for future use. As a negative
control, serum-free culture medium was processed using the same
method.
Allogenic BMT
Female BALB/c recipient mice were injected with
busulfan (Bu; 20 mg/kg/day) for 4 days, followed by
cyclophosphamide (Cy; 100 mg/kg/day) as previously described
(23) for 2 days via
intraperitoneal injection. After 1 day of rest, BMT was performed.
To isolate BMCs, male C57BL/6 donor mice were sacrificed by
cervical dislocation. Their BM was dislodged from the medullary
cavities of both the femurs and tibias and prepared as a
single-cell suspension. The cells were filtered using a cell
strainer (70 µm, SPL Life Sciences) and centrifuged at 190 ×
g for 5 min at room temperature. The pellet was resuspended with
ACK lysis buffer (150 mM NH4Cl, 10 mM KHCO3
and 0.1 mM Na2EDTA) to lyse erythrocytes. Female BALB/c
recipient mice were then injected with 1×107 BMCs
(Bu-Cy+BMT group) or 1×107 BMCs combined with
2×106 T-MSCs (Bu-Cy+BMT+T-MSC group) in a total volume
of 200 µl via lateral tail vein injection. During the
experimental periods, humane endpoints were defined as the time
when the animal lost 20% of its starting weight, at which point the
animal was immediately sacrificed. Recipient mice were examined
daily for 40 days, and their survival and changes in weight were
recorded. Mice were sacrificed by cervical dislocation on days 3,
10 and 40 following allogenic BMT. Thymus size was estimated by
pixel calculations of the images using ImageJ software (version
1.51).
Immunohistochemistry
Thymus tissues were isolated from healthy recipient
female BALB/c mice (normal controls), mice receiving cytotoxic
conditioning (Bu-Cy only group), Bu-Cy + BMT mice and Bu-Cy + BMT +
T-MSC mice and were embedded in paraffin. Sections of the thymus
were stained with hematoxylin (YD Diagnostics) for 2 min and eosin
(Sigma-Aldrich) for 1.5 min (H&E) at room temperature. For
immunohistochemistry, the thymus tissue sections were
deparaffinized by immersion in xylene 2 times for 5 min and then
hydrated. The slides were washed with PBS. The antigenic epitopes
were recovered with heat using citrate buffer (10 mM citric acid
and 0.05% Tween-20, pH 6.0) at 95°C for 20 min, and the slides were
held in cool citrate buffer for 20 min. Endogenous peroxide
blockade was performed using hydrogen peroxide (Dako North America,
Inc.) in a humidity chamber for 30 min. The slides were washed
again with PBS for 5 min. Non-specific binding blockade was
performed with protein blocking reagent (Dako North America, Inc.)
for 15 min. The slides were incubated overnight at 4°C with primary
anti-CD3E rabbit polyclonal antibody (bs-4815R, Bioss Antibodies)
diluted 1:200 with PBS. After returning to room temperature, the
slides were washed with PBS for 5 min and incubated with secondary
biotinylated anti-rabbit and anti-mouse antibodies in PBS
containing carrier protein and 0.015 mol/l sodium azide (K0609,
LSAB2 System-HRP, Dako North America, Inc.) in a humidity chamber
for 30 min. The slides were washed with PBS for 5 min and incubated
with streptavidin conjugated to horseradish peroxidase (HRP) in PBS
(Dako North America, Inc.) for 30 min. The slides were washed for 5
min with PBS, and the reaction was developed with
3,3′-diaminobenzidine solution (Dako North America, Inc.) for 10
min. The slides were washed in running water for 10 min and
counterstained with hematoxylin for 60 sec. The slides were washed
again in running water for 5 min, dipped 3 times in 1% acid
alcohol, washed in running water for 5 min, dehydrated and mounted.
Immunohistochemical analysis was performed with a slide scanner
(Aperio Scanscope FL).
Terminal deoxynucleotidyl transferase
dUTP nick-end labeling (TUNEL) assay
The TACS®2 TdT-Fluor In Situ Apoptosis
Detection kit (Trevigen, Inc.) was used to measure apoptosis in
accordance with the manufacturer's protocol. Briefly, the thymus
section slides were deparaffinized by warming to 57°C for 5 min,
immersing twice in xylene for 5 min, and immersing consecutively in
100, 95 and 70% ethanol for 5 min each. The tissue slides were
washed twice in 1X PBS for 5 min each. For labeling preparation,
the slides were carefully dried around the samples, which were
covered with 50 µl proteinase K solution, incubated for 15
min at room temperature, and washed twice in deionized water for 2
min each. The slides were immersed in 1X terminal deoxynucleotidyl
transferase (TdT) labeling buffer for 5 min, covered with 50
µl labeling reaction mix, and incubated at 37°C for 1 h in a
humidity chamber. The slides were immersed in 1X TdT stop buffer
for 5 min at room temperature to stop the labeling reaction and
then washed twice in deionized water at room temperature for 5 min
each time. The slides were covered with 50 µl strepfluor
solution, incubated for 20 min at room temperature in the dark, and
washed twice in 1X PBS for 2 min each time. After mounting (Vector
Laboratories, Inc.), the samples were viewed with a confocal
microscope (Carl Zeiss AG).
T cell receptor (TCR)β sequencing
Thymic RNA from experimental mice was extracted
using the RNeasy Plus Mini kit (Qiagen GmbH) in accordance with the
manufacturer's protocol. The RNA samples were analyzed for purity
and concentration using a BioPhotometer® D30
(Eppendorf). The RNA samples were sent to Macrogen for TCRβ chain
sequencing. The V-C primer was designed to cover V-D-J-C genes
averaging 380 bp. The resulting sequences were analyzed using the
iRepertoire TCR beta sequencing panel (https://www.irepertoire.com). To quantitatively
represent the degree of T cell diversity within the samples,
diversity 50 (D50) analysis was used. The D50 value is the percent
of dominant and unique T cell clones that cumulatively account for
50% of the total complementarity determining region 3 (CDR3)
sequences in each sample. The more diverse a library, the closer
the value will be to 50. The D50 was calculated as follows: (Number
of unique CDR3s that compose 50% of the reads of the top 10,000
unique CDR3s ×100)/10,000=D50. An illustrative tree map was used to
illustrate V-J-unique CDR3s. The entire plot area was divided into
sub-areas according to V usage that were further grouped according
to J usage. Each unique CDR3 within a given V-J-combination was
subsequently represented by a rounded rectangle. The unevenness of
squares reflects areas of thymus-dependent clonal expansion of the
thymocyte repertoire.
Reverse transcription-polymerase chain
reaction (RT-PCR)
To compare the expression levels of FGF7, FLT3L,
interleukin (IL)-7 and IL-15 in the T-MSCs compared with those from
BM-MSCs and AT-MSCs, total RNA was extracted from the cells
harvested at 80% confluence. Cells were collected with a cell
scraper and centrifuged at 190 × g for 5 min. Cell pellets were
resuspended completely with 1 ml TRIzol (Invitrogen; Thermo Fisher
Scientific, Inc.) and incubated for 5 min at room temperature. The
samples were added to 200 µl chloroform, inverted for 15
sec, and incubated at room temperature for 3 min. The samples were
centrifuged at 16,000 × g for 15 min at 4°C. The aqueous phase was
transferred into a new tube with 500 µl isopropyl alcohol
and incubated at room temperature for 10 min. The samples were
centrifuged at 16,000 × g for 10 min at 4°C. The supernatant was
removed, and the pellet was washed with 75% ethanol. The pellet was
inverted 5-6 times and centrifuged at 16,000 × g for 5 min at 4°C.
The RNA pellet was dried briefly and dissolved in RNase-free water.
RNA samples were analyzed for purity and concentration using a
BioPhotometer® D30 (Eppendorf) and adjusted to 1
µg/µl. Total RNA was transcribed into cDNA using a
reverse transcription regent (ELPIS-Biotech, Inc.) according to the
manufacturer's instructions. The samples were then mixed with their
respective primers and Maxime™ PCR PreMix (i-Taq, iNtRON
Biotechnology) according to the manufacturer's instructions.
Amplification was performed in duplicate with 35 cycles of a 30-sec
denaturation step at 94°C, 30-sec annealing step at 62°C, and
30-sec extension step at 72°C. The relative fold expression and
changes were observed from DNA gel electrophoresis (AE-9000
E-graph, ATTO-Gentaur). Primers for the gene expression by RT-PCR
were as follows: Human FGF7 (123 bp) forward, 5′-TCC TGC CAA
CTT TGC TCT ACA-3′ and reverse, 5′-CAG GGC TGG AAC AGT TCA CAT-3′;
human FLT3L (196 bp) forward, 5′-AAA ATC CGTGAG CTG TCT
GAC-3′ and reverse, 5′- TGA CAA AGT GTA TCT CCG TGT TC-3′; human
IL-7 (220 bp) forward, 5′-TTC CTC CCC TGA TCC TTG TTC-3′ and
reverse, 5′- CTT GCG AGC AGC ACG GAA TA-3′; human IL-15 (190
bp) forward, 5′-TTG GGA ACC ATA GAT TTG TGC AG-3′ and reverse,
5′-AGA GAA AGC ACT TCA TTG CTG TT-3′; and human GAPDH (192
bp) forward, 5′- GGT AAA GTG GAT ATT GTT GCC ATC AAT G-3′ and
reverse, 5′-GGA GGG ATC TCG CTC CTG GAA GAT GGT G-3′. The pixel
densities of the FGF7, FLT3L, IL-7 and IL-15 bands
were divided by the pixel densities of the corresponding
GAPDH bands for normalization of the expression levels using
UN-SCAN-IT-gel 6.1 software (Silk Scientific, Inc.).
Western blot analysis
Equal amounts of concentrated CM (20 µl for
2×105 cells) from BM-MSCs, AT-MSCs and T-MSCs were
loaded onto a 5% stacking/10% separating polyacrylamide gel,
separated by electrophoresis, transferred to polyvinylidene
difluoride (PVDF) membranes, blocked with 5% skim milk in TBST (50
mM Tris-HCl, pH 7.6, 150 mM NaCl, 0.1% Tween-20) and incubated with
primary antibodies overnight at 4°C. All primary antibodies were
prepared by diluting in 3% BSA (Bovogen Biologicals) and 0.02%
sodium azide (Sigma-Aldrich; Merck KGaA) in TBST. Anti-FGF7 mouse
monoclonal antibody (sc-365440, 1:200, F-9, IgG1, κ),
anti-FLT3L mouse monoclonal antibody (sc-365266, 1:200, F-6,
IgG1, κ) and anti-β-actin mouse monoclonal antibody
(sc-47778, 1:3,000, C4, IgG1, κ) were purchased from
Santa Cruz Biotechnology, Inc. The PVDF membranes were washed 3
times for 10 min in TBST and incubated with goat anti-mouse IgG (H
+ L)-HRP antibody (#1706516, Bio-Rad Laboratories, Inc.) and
diluted in TBST (1:3,000) for 1 h at room temperature. Following
incubation, the membranes were washed 3 times for 10 min in TBST
and developed using EZ-Western Lumi Femto (DoGenBio Co.). Images
were obtained using ImageQuant LAS 500 (GE Healthcare Life
Sciences). The pixel densities of the FGF7 and FLT3L bands were
divided by the pixel densities of the corresponding β-actin bands
for protein quantitation using UN-SCAN-IT-gel 6.1 software.
Statistical analysis
Data are presented as the means ± standard error of
the mean (SEM). Statistical significance was determined by mixed
analysis of variance (ANOVA) in conjunction with the Sidak test as
applied to the weight loss of Bu-Cy pre-conditioned mice. Survival
curves were plotted using Kaplan-Meier estimates and analyzed using
the log-rank test. One-way ANOVA in conjunction with Sidak's
multiple comparisons test were used for the results of
immunohisto-chemistry, TUNEL assay, and semi-quantitative PCR. All
analyses were performed using Prism 8 (GraphPad Software, Inc.).
P<0.05 was considered to indicate a statistically significant
difference for all comparisons.
Results
T-MSCs promote weight recovery and
survival in allogenic BMT following Bu-Cy conditioning
To identify the effects of T-MSCs on body weight
recovery and survival rate following allogenic BMT, mice were
injected with Bu-Cy for cytotoxic conditioning and/or transplanted
BMCs and BMCs with T-MSCs by tail vein injection and were observed
for 40 days after cell transfer (Fig.
1A). Rapid weight loss was observed within 7 days in mice after
the Bu-Cy injection, but not in the normal control (no treatment)
mice (Fig. 1B). Differences in
body weight recovery of the Bu-Cy + BMT + T-MSC group were noted
after experimental day 14. Mice co-transplanted with T-MSCs
significantly regained their body weight more rapidly than the
Bu-Cy only group and Bu-Cy + BMT mice. Although body weight
recovery of the Bu-Cy + BMT mice was less than that of the Bu-Cy
mice, their survival rate was much higher (Fig. 1B). There are 3 different levels of
intensity in the pre-conditioning regimen, such as myeloablative
(high dose), reduced intensity, or non-myeloablative (24). Due to severe pre-conditioning, the
Bu-Cy group exhibited high rates of fatality during the
experimental period without BM cells, due to severe weight loss
following cytotoxic preconditioning (2 mice died on their own from
weight loss (21 and 23% weight loss for each of the mice,
respectively) and 3 mice had reached the humane endpoint, which was
a ≥20% loss of body weight from the starting body weight). By
contrast, the Bu-Cy + BMT and Bu-Cy + BMT + T-MSC groups exhibited
extended survival times; specifically, Bu-Cy + BMT and Bu-Cy + BMT
+ T-MSC mice exhibited an 80 and 90% survival rate, respectively,
during the experimental period (Fig.
1C).
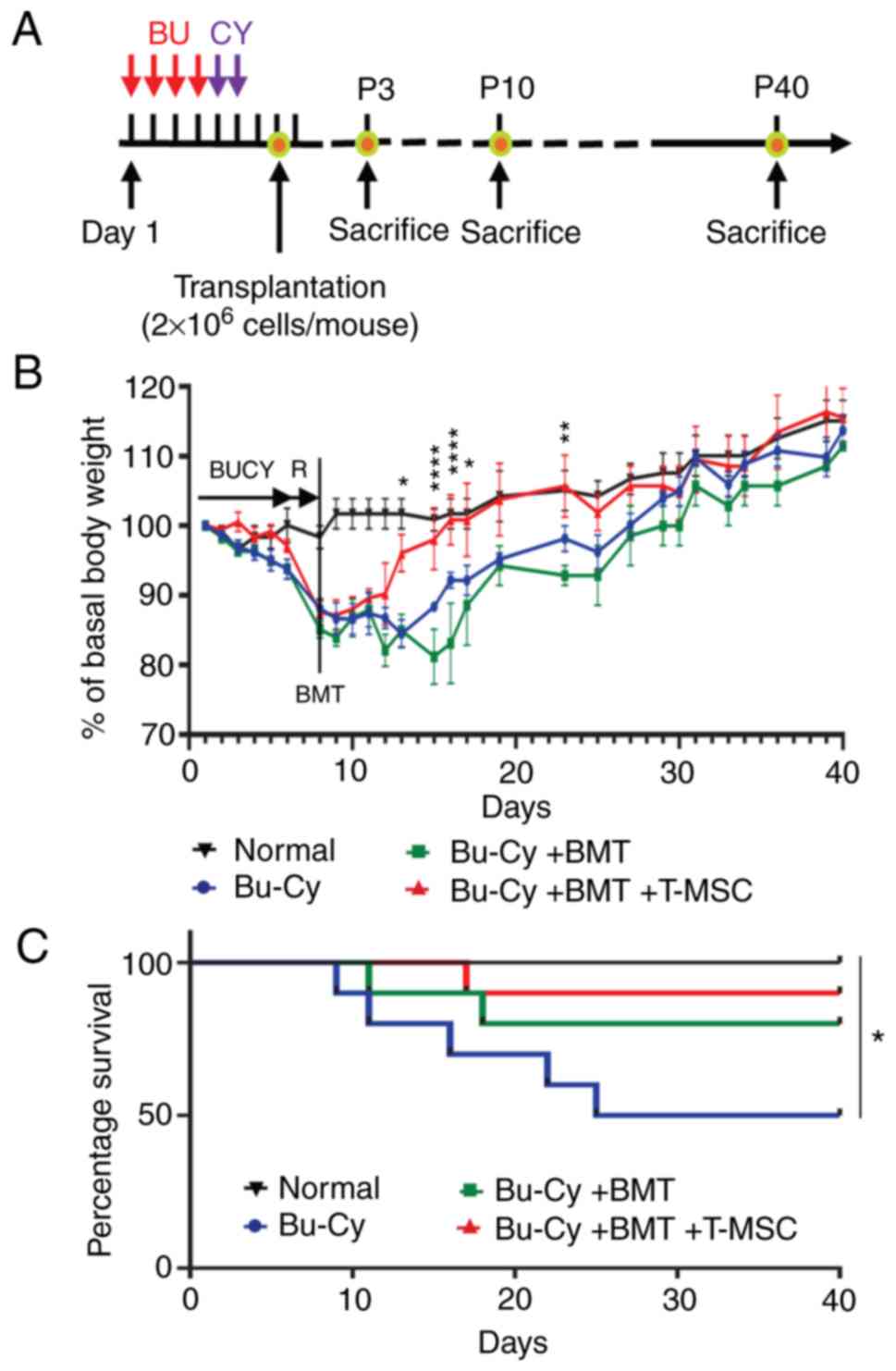 | Figure 1Weight change and survival course in
the experimental mice. (A) Experimental scheme for BMT and T-MSC
transfer time schedule. (B) BALB/c female mice were injected with
busulfan (Bu; 20 mg/kg/day) for 4 days, followed by
cyclophosphamide (Cy; 100 mg/kg/day) for 2 days. The mice were
classified into 4 groups as follows: Normal female BALB/c (normal
control), Bu and Cy treatment only (Bu-Cy), transplantation of BMCs
from C57BL/6 mice after Bu-Cy treatment (Bu-Cy + BMT), and
transplantation of BMCs plus T-MSCs after Bu-Cy treatment (Bu-Cy +
BMT + T-MSC). The graph presents body weight changes in
experimental mice (n=10 for each group) for 40 days after BMT.
Basal body weight was determined as weight at BMT day (start).
Mixed ANOVA followed by the Sidak test; *P<0.05,
**P<0.01, and ****P<0.0001, Bu-Cy + BMT
group vs. Bu-Cy + BMT + T-MSC group. (C) Survival course of the 4
experimental groups measured using the Kaplan-Meier estimator and
compared using log-rank test (*P<0.05, P=0.0268).
BMT, one marrow transplantation; T-MSC, tonsil-derived mesenchymal
stromal cell; Bu-Cy, busulfan-cyclophosphamide. |
T-MSCs accelerate thymus regeneration
following cytotoxic conditioning
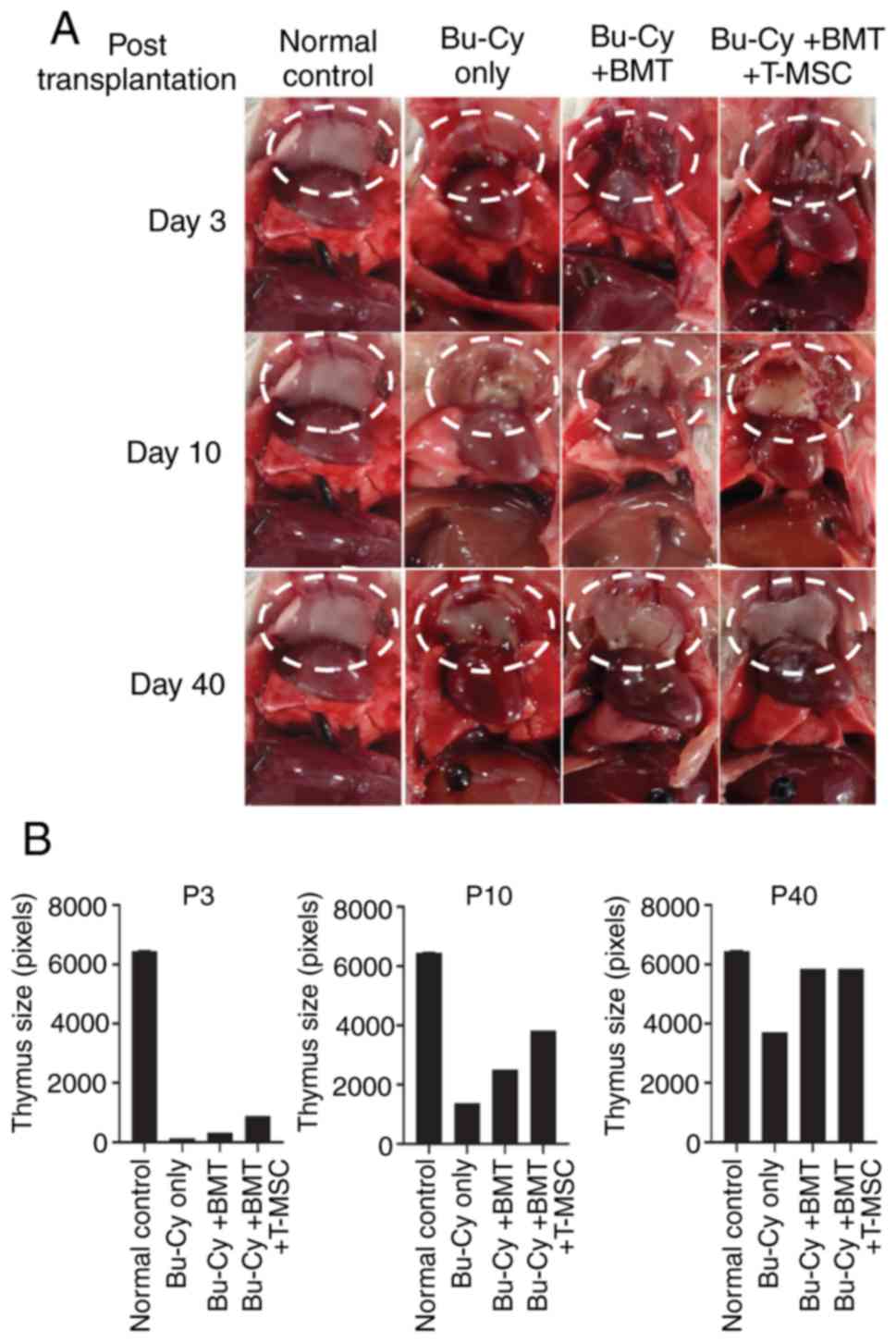
The thymus size of the experimental mice decreased
after pre-conditioning (Fig. 2A and
B). To observe the effects of T-MSCs on thymus size following
Bu-Cy treatment, mice from each group were sacrificed on days 3, 10
and 40 post-BMT. Gross observations revealed a shrunken thymus in
the mice on day 3. On day 10, however, the thymuses of the Bu-Cy +
BMT + T-MSC mice were larger than those of mice in all other
groups. Thus, BMT combined with T-MSCs rapidly reversed thymus
involution induced by the conditioning regimen. On day 40, the
thymus size of all groups recovered to a level similar to that of
the normal control (Fig. 2B).
To analyze cellularity and recovery of thymus
architecture, tissue sections were stained with H&E. Moreover,
immunohistochemical analysis of thymus tissue for the T cell
marker, CD3, revealed compositional changes of the thymus following
BMT and indicated its restoration. The medulla and cortex regions
in the thymus were distinct in the normal control groups, while
thymus organization was abolished and the medulla and cortex were
not distinguished in Bu-Cy only mice. The thymuses of the Bu-Cy
only group exhibited less cellularity, and most of the tissue was
fibrotic compared with the other groups (Fig. 2C). However, thymocyte cellularity
in the Bu-Cy + BMT + T-MSC mice was more prominent than that of the
other experimental groups. In addition, the thymic cortex and
medulla structures of Bu-Cy + BMT + T-MSC mice were defined as
clearly as those in the normal control (Fig. 2C). To further determine the
recovery and repopulation of thymocytes, the thymus tissues were
stained with anti-CD3 antibody and quantified areas of
CD3+ cells. The co-transplantation group (Bu-Cy + BMT +
T-MSCs) exhibited more repopulated CD3+ cells than those
of the Bu-Cy only or Bu-Cy + BMT groups following BMT (Fig. 2D and E).
Co-transplantation of T-MSCs reduces the
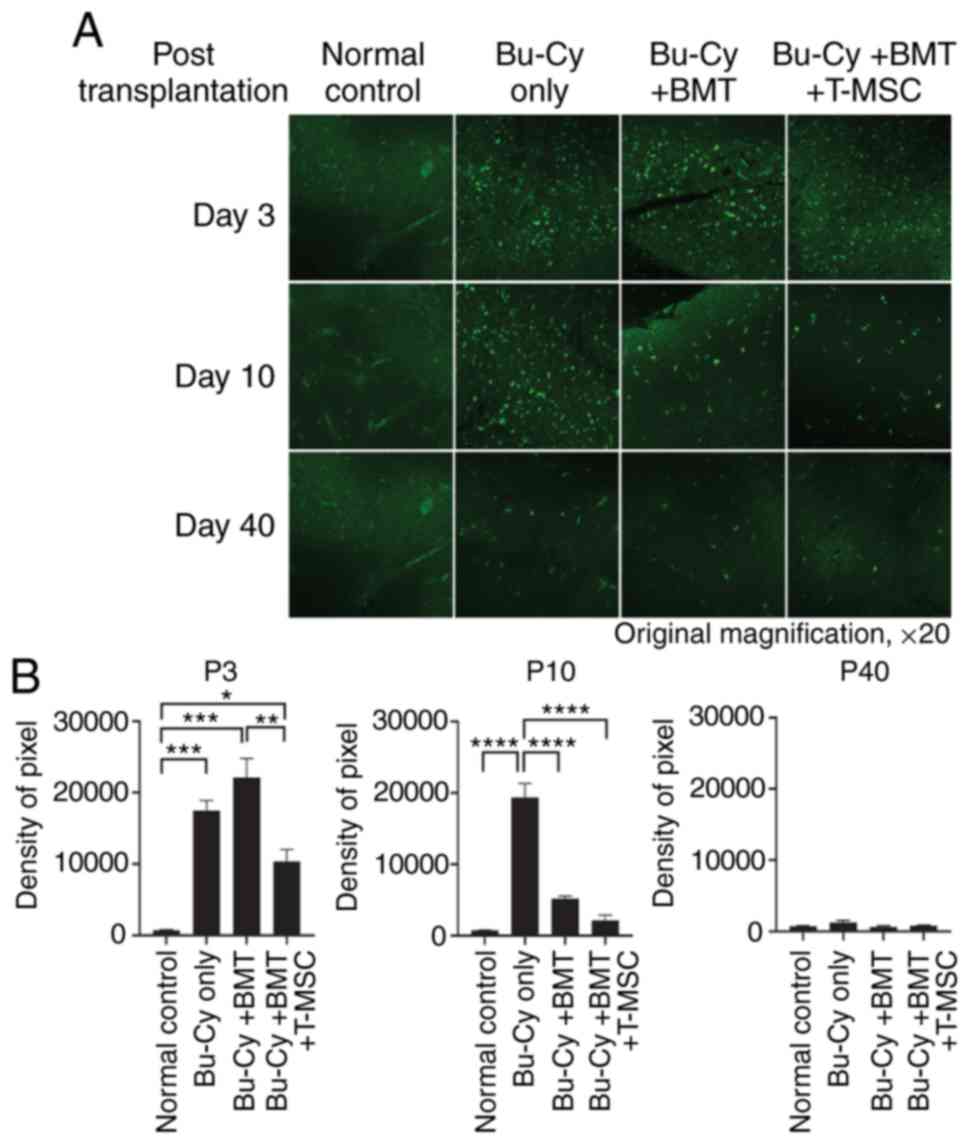
frequency of apoptosis
A TUNEL assay was performed to verify the effects of
T-MSC co-transplantation on thymus recovery in the context of
thymocyte apoptosis. Thymus sections from all experimental groups
exhibited high levels of apoptosis on day 3 post-BMT. On day 10,
the highest number of apoptotic cells in the thymus were observed
in the Bu-Cy only group, followed by the Bu-Cy + BMT and Bu-Cy +
BMT + T-MSC groups (Fig. 3A and
B). All thymic sections from the experimental groups exhibited
similarly low levels of fluorescence on day 40 and baseline
apoptosis frequency (Fig. 3B,
P40). Thus, results revealed that the co-transplantation of T-MSCs
distinctly decreased the appearance of apoptotic lymphocytes in the
thymus.
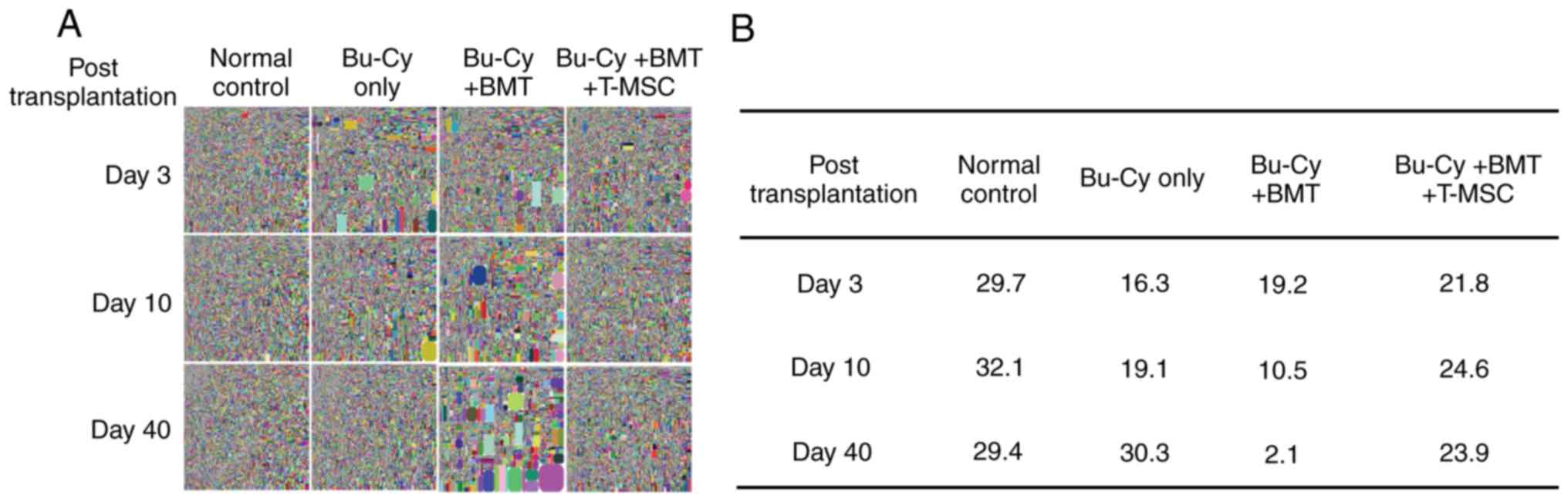
Co-transplantation of T-MSCs with BMCs
improves diversity of the TCR repertoire
The thymic TCR repertoires of the experimental
groups were compared by TCRβ chain sequencing. The squares
represent areas of clonal expansion within the sample (Fig. 4A). The Bu-Cy only group exhibited
the least amount of TCR repertoire diversity on day 3 post-BMT
among all groups. However, at 40 days post-BMT, the Bu-Cy + BMT
group exhibited the least TCR diversity.
Expression of factors related to thymus
regeneration
To explore T-MSC-associated factors related to
thymus regeneration, the expression of FGF7, FLT3L, IL-7 and IL-15
was measured, which are soluble factors that promote T cell
reconstitution in BM-MSCs, AT-MSCs and T-MSCs using RT-PCR and
western blot analysis. At the mRNA level, T-MSCs highly expressed
FGF7 and FLT3L compared with the BM-MSCs (P<0.05;
Fig. 5A and B). At the protein
level, T-MSCs highly expressed FGF7 and FLT3L compared to the
BM-MSCs and AT-MSCs (Fig. 5C).
Moreover, the T-MSCs produced higher levels of factors related to
thymus regeneration than BM-MSCs and AT-MSCs as shown by western
blot analysis (Fig. 5D). To
confirm whether the factors associated thymus regeneration were
increased, the expression of mouse Fgf7, Flt3l, human
FGF7 and FLT3L were compared in the thymuses on days
3 and 10 post-BMT. In the thymus on day 3 post-BMT, Fgf7 and
FGF7 was significantly more expressed in the Bu-Cy + BMT +
T-MSCs group than the other groups and Flt3l was more
expressed in both BMT groups than in Bu-Cy only group (Fig. S1A and C). In the thymus on day 10
post-BMT, Fgf7, Flt3l and FLT3L expression
levels in Bu-Cy only group were restored up to those of the normal
control (Fig. S1B and D).
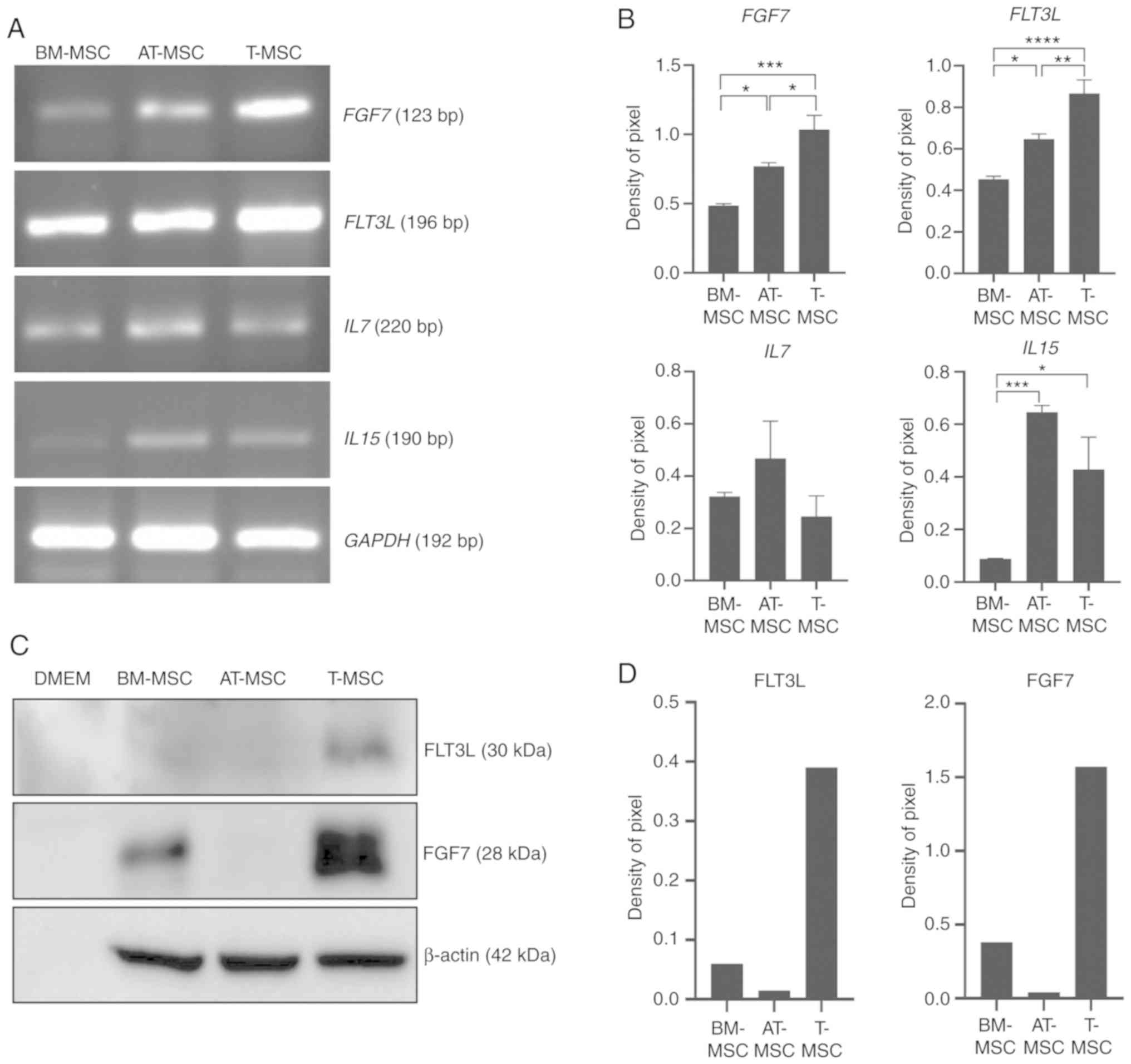 | Figure 5Expression of factors related to
thymus regeneration. (A) RNA was extracted from BM-MSCs, AT-MSCs,
and T-MSCs at 80% confluence and analyzed by RT-PCR. (B) The pixel
densities of the FGF7, FLT3L, IL7 and IL15 bands were
divided by the pixel densities of the corresponding GAPDH
bands. Data are presented as the means ± standard error (one-way
ANOVA followed by Sidak's multiple comparisons test;
*P<0.05, **P<0.01,
***P<0.001 and ****P<0.0001 as
indicated). (C) Cell culture medium was collected and concentrated
for immunoblotting to detect the expression of FLT3L and FGF7 in
BM-MSCs, AT-MSCs, and T-MSCs. DMEM was loaded as the negative
control. (D) The pixel densities of the FLT3L and FGF7 bands from
representative blots were divided by the pixel densities of the
corresponding β-actin bands. BM-MSCs, bone marrow-derived
mesenchymal stromal cells; T-MSCs, tonsil-derived mesenchymal
stromal cells; AT-MSCs, adipose tissue-derived MSCs. |
Discussion
In the present study, it was found that the
co-transplantation of T-MSCs with BMCs accelerates the recovery of
the involuted thymus after partial pre-BMT conditioning with Bu-Cy
treatment. The co-transplantation of T-MSCs with BMCs replenished
CD3+ cell populations by inhibiting the apoptosis of
thymocytes following cytotoxic pre-conditioning. In addition, T-MSC
co-transplantation improved TCR repertoire expansion and increased
the diversity of thymus-derived T cells.
Full T cell reconstitution following BMT needs de
novo production of naïve T cells in the thymus of the recipient
via thymus-dependent pathway. Such endogenous T cell regeneration
result in increasing TCR repertoire and antigen specificity as well
as long lasting immune recovery (6). Critically, the recovery of the
thymus is essential for the reconstitution of immune tolerance
after BMT. Patients who develop chronic GVHD exhibit reduced CD4
TCR diversity compared to those without GVHD; naïve T cell
reconstitution through the thymus-dependent pathway was also
observed in the non-GVHD patients (25,26). These findings suggest that
peripheral T cell expansion via the thymus-independent pathway may
be associated with development of GVHD. Clinically, GVHD is
associated with damage to the gut, liver and skin but thymus is
proven to be sensitive target of alloreactive T cells in murine
model (27). Paradoxically, the
most common treatment of GVHD is corticosteroids, which lead to
acute thymic involution (28),
eventually mediate thymic damage and lead to constriction of T cell
repertoire. Therefore, a different approach is required to repress
the thymus-independent pathway, but enhance thymus-dependent T cell
production, as shown in the present study. In this experimental
system, it was found that T-MSCs express more FGF7 and
FLT3L than BM-MSC or AT-MSC, but not for IL-7 or
IL-15, which are related to thymus-independent recovery
(Fig. 5). As these results are
relevant models of non-GVHD patients and focus on thymus-dependent
T cell production, the effects of T-MSCs on the thymus in a GVHD
model remain to be elucidated.
To assess thymic output and T cell reconstitution
following allogenic BMT or cancer chemotherapy alone, the detection
of excisional DNA circles generated from TCR chain rearrangement
(29) and TCRβ gene sequencing
have been used (30,31). In the present study, TCRβ chain
sequencing was performed and D50 values were compared to determine
repertoire reconstitution following BMT under various experimental
conditions. To compare the effect of T-MSCs on thymic recovery, the
BMT and BMT + T-MSC groups were primarily compared and it was found
that T-MSC co-transplantation accelerated thymocyte rescue and
expanded TCR repertoire following allogenic BMT. As shown in
Fig. 4B, the diversity index of T
cells from T-MSC co-transplanted mice was increased compared with
those of the BMT only group. Notably, the recipient mice in the
Bu-Cy + BMT and Bu-Cy + BMT + T-MSC groups were BALB/c female mice,
and the donor BMCs originated from C57BL/6 male mice. Therefore,
the thymic recovery of Bu-Cy + BMT and Bu-Cy + BMT + T-MSC mice
involved two distinct cellular origins (i.e., BALB/c and C57BL/6)
consisting of thymic chimerism. By contrast, the thymic recovery of
the normal control and Bu-Cy only groups involved only the BALB/c
recipient mouse cells. Interestingly, the D50 of the Bu-Cy only
mice on day 40 post-BMT was nearly the same as that of the control
mice. Although the overall survival rate of the Bu-Cy only group
was 50% (n=5 out of 10), the mice that survived until the 40-day
endpoint spontaneously recovered from the cytotoxic conditioning
and regained thymic structure and function, represented as a
decreased number of apoptotic cells on day 40 in Bu-Cy only group
(Fig. 3B). However, as presented
in Fig. 2E, the re-populated
CD3+ cells in the thymuses were the lowest in Bu-Cy only
group and it might lead to a decreased survival compared to those
of Bu-Cy + BMT group (Fig. 1C).
Therefore, the interpretation of D50 must be accompanied with total
T cell number and apoptotic cell fraction in the thymuses as an
index of thymus recovery, especially representing expansion of
selected TCRβ clones.
In order to evaluate immune reconstitution following
BMT, γδ T cell reconstitution is associated with fewer infection
following HSC transplantation (32). γδ T cells are innate-like effector
cells and exert GVL without GVHD (33,34). Such γδ TCR repertoire analyses
from peripheral blood are possibly as much effective as thymus
sequencing for αβ TCR to evaluate T cell-mediated immune
reconstitution. γδ TCR reconstitution following the
co-transplantation of T-MSC also needs to be analyzed and compared
in future studies.
A marked loss of body weight and depletion of fat
and muscle storage occur following chemotherapy (35). The possible mechanism driving the
increased body weight in the Bu-Cy + BMT + T-MSC group compared
with the Bu-Cy only or Bu-Cy + BMT groups can be explained by the
effect of MSCs on muscle loss due to Bu-Cy treatment. These muscle
wasting was associated with the upregulation of ERK1/2 and p38
MAPKs (35). In airway
inflammation model, MSCs down-regulate cyclooxygenase-2 via p38 and
ERK/MAPK pathways (36). It has
also been reported that human MSCs from Wharton's jelly may
attenuate sarcopenia, which is loss of muscle mass and function,
via activation of muscle satellite cells, reduction of apoptosis
and reduced inflammation (37).
In conclusion, the approach used in the present
study to explore the effects of T-MSCs on thymic regeneration
following allogenic BMT revealed that the co-transplantation of
T-MSCs with BMCs improved mouse survival, restored thymic
structure, reduced apoptosis and expanded TCR diversity compared
with the BMT-only group, possibly by inducing FLT3L and FGF7
expression in T-MSCs. Further studies are required to examine the
mechanisms of T cell repertoire diversity expansion and to analyze
peripheral T cell subsets following BMT. Additional analysis of the
effects of co-transplantation on thymic and peripheral immune
reconstitution may also contribute to the development of clinical
approaches for preventing infection or GVHD following BMT.
Supplementary Data
Acknowledgments
Not applicable.
Funding
The present study was supported by a grant of the
Korea Health Technology R&D Project through the Korea Health
Industry Development Institute (KHIDI), which was funded by the
Ministry of Health & Welfare, Republic of Korea (grant no.
HI18C2392).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DWC and KAC performed the experiments and wrote the
manuscript. HJL, YHK and JWP analyzed the data and assisted in the
writing of the manuscript. KJW and KHR interpreted the data and
assisted in the writing of the manuscript. SYW designed the
experiments and wrote the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The experimental procedures were approved by the
Animal Care and Use Committee of the College of Medicine, Ewha
Womans University (approval no. ESM18-0403).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Klein OR, Buddenbaum J, Tucker N, Chen AR,
Gamper CJ, Loeb D, Zambidis E, Llosa NJ, Huo JS, Robey N, et al:
Nonmyeloablative haploidentical bone marrow transplantation with
post-transplantation cyclophosphamide for pediatric and young adult
patients with high-risk hematologic malignancies. Biol Blood Marrow
Transplant. 23:325–332. 2017. View Article : Google Scholar
|
|
2
|
Sengsayadeth S, Savani BN, Blaise D and
Mohty M: Haploidentical transplantation: Selecting optimal
conditioning regimen and stem cell source. Semin Hematol.
53:111–114. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Storek J, Geddes M, Khan F, Huard B, Helg
C, Chalandon Y, Passweg J, Roosnek E, et al: Reconstitution of the
immune system after hematopoietic stem cell transplantation in
humans. Semin Immunopathol. 30:425–437. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Krenger W, Blazar BR and Hollander GA:
Thymic T-cell development in allogeneic stem cell transplantation.
Blood. 117:6768–6776. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Simons L, Cavazzana M and Andre I: Concise
Review: Boosting T-cell reconstitution following allogeneic
transplantation-current concepts and future perspectives. Stem
Cells Transl Med. 8:650–657. 2019.PubMed/NCBI
|
|
6
|
Chaudhry MS, Velardi E, Malard F and van
den Brink MR: Immune reconstitution after allogeneic hematopoietic
stem cell transplantation: Time to T up the thymus. J Immunol.
198:40–46. 2017. View Article : Google Scholar
|
|
7
|
Rossi SW, Jeker LT, Ueno T, Kuse S, Keller
MP, Zuklys S, Gudkov AV, Takahama Y, Krenger W, Blazar BR and
Holländer GA: Keratinocyte growth factor (KGF) enhances postnatal
T-cell development via enhancements in proliferation and function
of thymic epithelial cells. Blood. 109:3803–3811. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Alpdogan O, Hubbard VM, Smith OM, Patel N,
Lu S, Goldberg GL, Gray DH, Feinman J, Kochman AA, Eng JM, et al:
Keratinocyte growth factor (KGF) is required for postnatal thymic
regeneration. Blood. 107:2453–2460. 2006. View Article : Google Scholar
|
|
9
|
Sun L, Li H, Luo H and Zhao Y: Thymic
epithelial cell development and its dysfunction in human diseases.
Biomed Res Int. 2014:2069292014.PubMed/NCBI
|
|
10
|
Jenkinson WE, Jenkinson EJ and Anderson G:
Differential requirement for mesenchyme in the proliferation and
maturation of thymic epithelial progenitors. J Exp Med.
198:325–332. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wertheimer T, Velardi E, Tsai J, Cooper K,
Xiao S, Kloss CC, Ottmüller KJ, Mokhtari Z, Brede C, deRoos P, et
al: Production of BMP4 by endothelial cells is crucial for
endogenous thymic regeneration. Sci Immunol. 3:eaal27362018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Perry JSA, Russler-Germain EV, Zhou YW,
Purtha W, Cooper ML, Choi J, Schroeder MA, Salazar V, Egawa T, Lee
BC, et al: Transfer of cell-surface antigens by scavenger receptor
CD36 promotes thymic regulatory T cell receptor repertoire
development and allotolerance. Immunity. 48:12712018. View Article : Google Scholar
|
|
13
|
Rasko JE, Metcalf D, Rossner MT, Begley CG
and Nicola NA: The flt3/flk-2 ligand: Receptor distribution and
action on murine haemopoietic cell survival and proliferation.
Leukemia. 9:2058–2066. 1995.PubMed/NCBI
|
|
14
|
Sitnicka E, Buza-Vidas N, Ahlenius H,
Cilio CM, Gekas C, Nygren JM, Månsson R, Cheng M, Jensen CT,
Svensson M, et al: Critical role of FLT3 ligand in IL-7 receptor
independent T lymphopoiesis and regulation of lymphoid-primed
multipotent progenitors. Blood. 110:2955–2964. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Williams KM, Moore AR, Lucas PJ, Wang J,
Bare CV and Gress RE: FLT3 ligand regulates thymic precursor cells
and hematopoietic stem cells through interactions with CXCR4 and
the marrow niche. Exp Hematol. 52:40–49. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tan JT, Dudl E, LeRoy E, Murray R, Sprent
J, Weinberg KI and Surh CD: IL-7 is critical for homeostatic
proliferation and survival of naive T cells. Proc Natl Acad Sci
USA. 98:8732–8737. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kondrack RM, Harbertson J, Tan JT, McBreen
ME, Surh CD and Bradley LM: Interleukin 7 regulates the survival
and generation of memory CD4 cells. J Exp Med. 198:1797–1806. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Murali-Krishna K, Lau LL, Sambhara S,
Lemonnier F, Altman J and Ahmed R: Persistence of memory CD8 T
cells in MHC class I-deficient mice. Science. 286:1377–1381. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cho KA, Kim YH, Park M, Kim HJ, Woo SY,
Park JW and Ryu KH: Conditioned medium from human palatine tonsil
mesenchymal stem cells attenuates acute graft-vs.-host disease in
mice. Mol Med Rep. 19:609–616. 2019.
|
|
20
|
Kim JY, Park M, Kim YH, Ryu KH, Lee KH,
Cho KA and Woo SY: Tonsil-derived mesenchymal stem cells (T-MSCs)
prevent Th17-mediated autoimmune response via regulation of the
programmed death-1/programmed death ligand-1 (PD-1/PD-L1) pathway.
J Tissue Eng Regen Med. 12:e1022–e1033. 2018. View Article : Google Scholar
|
|
21
|
Cho KA, Park M, Kim YH, Ryu KH and Woo SY:
Poly I:C primes the suppressive function of human palatine
tonsil-derived MSCs against Th17 differentiation by increasing
PD-L1 expression. Immunobiology. 222:394–398. 2017. View Article : Google Scholar
|
|
22
|
Park M, Kim YH, Ryu JH, Woo SY and Ryu KH:
Immune suppressive effects of tonsil-derived mesenchymal stem cells
on mouse bone-marrow-derived dendritic cells. Stem Cells Int.
2015:1065402015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim YH, Cho KA, Lee HJ, Park M, Shin SJ,
Park JW, Woo SY and Ryu KH: Conditioned medium from human
tonsil-derived mesenchymal stem cells enhances bone marrow
engraftment via endothelial cell restoration by pleiotrophin.
Cells. 9:2212020. View Article : Google Scholar :
|
|
24
|
Gyurkocza B and Sandmaier BM: Conditioning
regimens for hematopoietic cell transplantation: One size does not
fit all. Blood. 124:344–353. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Soares MV, Azevedo RI, Ferreira IA, Bucar
S, Ribeiro AC, Vieira A, Pereira PNG, Ribeiro RM, Ligeiro D, Alho
AC, et al: Naive and stem cell memory T cell subset recovery
reveals opposing reconstitution patterns in CD4 and CD8 T cells in
chronic graft vs. host disease Front Immunol. 10:3342019.
View Article : Google Scholar
|
|
26
|
Alachkar H and Nakamura Y: Deep-sequencing
of the T-cell receptor repertoire in patients with haplo-cord and
matched-donor transplants. Chimerism. 6:47–49. 2015. View Article : Google Scholar
|
|
27
|
Krenger W, Rossi S and Hollander GA:
Apoptosis of thymocytes during acute graft-versus-host disease is
independent of gluco-corticoids. Transplantation. 69:2190–2193.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ashwell JD, Lu FW and Vacchio MS:
Glucocorticoids in T cell development and function. Annu Rev
Immunol. 18:309–345. 2000. View Article : Google Scholar
|
|
29
|
Douek DC, Vescio RA, Betts MR, Brenchley
JM, Hill BJ, Zhang L, Berenson JR, Collins RH and Koup RA:
Assessment of thymic output in adults after haematopoietic
stem-cell transplantation and prediction of T-cell reconstitution.
Lancet. 355:1875–1881. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Seo YD, Jiang X, Sullivan KM, Jalikis FG,
Smythe KS, Abbasi A, Vignali M, Park JO, Daniel SK, Pollack SM, et
al: Mobilization of CD8+ T Cells via CXCR4 blockade
facilitates PD-1 checkpoint therapy in human pancreatic cancer.
Clin Cancer Res. 25:3934–3945. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Toivonen R, Arstila TP and Hanninen A:
Islet-associated T-cell receptor-β CDR sequence repertoire in
prediabetic NOD mice reveals antigen-driven T-cell expansion and
shared usage of VβJβ TCR chains. Mol Immunol. 64:127–135. 2015.
View Article : Google Scholar
|
|
32
|
Perko R, Kang G, Sunkara A, Leung W,
Thomas PG and Dallas MH: Gamma delta T cell reconstitution is
associated with fewer infections and improved event-free survival
after hema-topoietic stem cell transplantation for pediatric
leukemia. Biol Blood Marrow Transplant. 21:130–136. 2015.
View Article : Google Scholar
|
|
33
|
Handgretinger R and Schilbach K: The
potential role of gammadelta T cells after allogeneic HCT for
leukemia. Blood. 131:1063–1072. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Minculescu L, Marquart HV, Ryder LP,
Andersen NS, Schjoedt I, Friis LS, Kornblit BT, Petersen SL,
Haastrup E, Fischer-Nielsen A, et al: Improved overall survival,
relapse-free-survival, and less Graft-vs.-host-disease in patients
with high immune reconstitution of TCR gamma delta cells 2 months
after allogeneic stem Cell Transplantation. Front Immunol.
10:19972019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Barreto R, Waning DL, Gao H, Liu Y,
Zimmers TA and Bonetto A: Chemotherapy-related cachexia is
associated with mitochondrial depletion and the activation of
ERK1/2 and p38 MAPKs. Oncotarget. 7:43442–43460. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gu W, Song L, Li XM, Wang D, Guo XJ and Xu
WG: Mesenchymal stem cells alleviate airway inflammation and
emphysema in COPD through down-regulation of cyclooxygenase-2 via
p38 and ERK MAPK pathways. Sci Rep. 5:87332015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang QQ, Jing XM, Bi YZ, Cao XF, Wang YZ,
Li YX, Qiao BJ, Chen Y, Hao YL and Hu J: Human Umbilical Cord
Wharton's Jelly Derived mesenchymal stromal cells may attenuate
sarco-penia in aged mice induced by hindlimb suspension. Med Sci
Monit. 24:9272–9281. 2018. View Article : Google Scholar : PubMed/NCBI
|