Introduction
Mesenchymal stem cells (MSCs) are pluripotent stem
cells derived from the mesoderm, with strong proliferative
capacity, multi-directional differentiation potential and low
immunogenicity (1). MSCs can
differentiate into other cell types, such as osteoblasts, myocytes,
hepatocytes and stromal cells, under certain conditions (2,3).
In the research field of regenerative medicine, human bone
marrow-derived MSCs (HBMSCs) can be easily acquired and cultivated,
and are widely used as seed cells for repairing and reconstructing
damaged tissues or organs (4).
Complex regulatory pathways at both the transcriptional and
post-transcriptional levels are involved in the differentiation
process (5). However, the exact
molecular regulatory mechanism underlying osteogenic
differentiation remains unknown. Therefore, the current study aimed
to investigate the mechanism of osteogenic differentiation of
HBMSCs.
Long-chain non-coding RNAs (lncRNAs) are RNA
molecules that are >200 nucleotides in length (6). Moreover, the majority of lncRNAs
lack substantial open reading frames for protein translation, and
are primarily transcribed by RNA polymerase II (6). Previous studies have reported that
lncRNAs serve an important regulatory role in biological processes,
such as cell proliferation, metastasis and differentiation
(7,8). Long intergenic non-protein coding
RNA 00707 (LINC00707) is located on chromosome 10p14 (9,10),
and upregulation of LINC00707 is associated with clinical
manifestations, such as lymphatic metastasis and tumor size in lung
adenocarcinoma (11). In
addition, LINC00707 promotes cell proliferation and migration by
regulating the target gene cell division cycle 42 (11). A previous study examined the role
of LINC00707 in hepatocellular carcinoma, and revealed that it may
be involved in the regulation of hepatocellular carcinoma
progression by downregulating microRNA (miRNA/miR)-206 to increase
cyclin dependent kinase 14 expression (12). In addition, the expression of
LINC00707 is upregulated in melanoma, and it could affect the
prognosis of patients by regulating inflammation and
mitogen-activated protein kinase-related pathways (9). However, the effect of LINC00707 on
the osteogenic differentiation of HBMSCs is yet to be fully
elucidated.
miRNAs are endogenous non-coding small
single-stranded RNAs (13,14),
and are involved in the regulation of osteogenic differentiation of
HBMSCs (15-17). For instance, miR-33a-5p has been
shown to regulate osteogenic differentiation by targeting SATB
homeobox 2 expression in HBMSCs (18). Moreover, inhibition of miR-222-3p
expression promotes osteoblast differentiation by modulating the
Smad5/RUNX family transcription factor 2 (RUNX2) signaling axis
(19). It has also been reported
that miR-125b regulates osteogenic differentiation of HBMSCs by
targeting bone morphogenetic protein receptor type 1B (20). In addition, miR-103a-3p can serve
as a reference gene for evaluating osteoarthritis (21).
Dickkopf WNT signaling pathway inhibitor (DKK1) is a
potent LDL receptor related protein (LRP)5/6 antagonist that
inhibits Wnt signaling pathways (22), and it has been reported that
excess glucocorticoids impair osteoblastogenesis by inducing Wnt
antagonists, including DKK1, sclerostin and secreted
frizzled-related protein 1 (23).
Furthermore, silencing DKK1 expression rescues
dexamethasone-induced suppression of primary human osteoblast
differentiation (24). Li et
al (22) also reported that
miR-291a-3p promoted BMSCs osteogenic differentiation via targeting
DKK1 in dexamethasone-induced osteoporosis.
In the present study, small interfering
(si)LINC00707 was constructed in HBMSCs, and the function of
LINC00707, miR-103a-3p and target genes were predicted by
bioinformatics analysis and further confirmed by performing
functional assays. In addition, the effects of these genes on
osteogenic differentiation were analyzed.
Materials and methods
Cell culture
HBMSCs (cat. no. PCS-500-012) were purchased from
the American Type Culture Collection. The cells were cultured in
CTS™ StemPro™ MSC serum-free medium [SFM; proliferation medium
(PM); cat. no. A1033201; Thermo Fisher Scientific, Inc.) at 37°C
with 5% CO2. After 2 days of culture, the original
medium was removed, the cells were washed once or twice with PBS
and then 5 ml fresh StemPro™ MSC SFM was added to the cells, which
were returned to the CO2 incubator. Cells were
subcultured after they reached 60-80% confluence.
HBMSCs were cultured in StemPro™ MSC SFM, passaged
to the third generation and osteogenic differentiation was then
induced using a StemPro™ Osteogenesis Differentiation kit
[osteogenic medium (OM); cat. no. A1007201; Thermo Fisher
Scientific, Inc.] after the cells had reached ≥80% confluence. Day
0 of the culture served as a control, and osteoblasts in
differentiation groups and RNAs of the undifferentiated cell groups
were extracted using TRIzol® reagent (cat. no. 15596026;
Thermo Fisher Scientific, Inc.) on day 1, 3, 5, 7, 14 and 21 after
the induction.
Transfection
After culture in OM, cells were incubated in a
CO2 incubator for 12-16 h at 37°C to reach 60-70%
confluency. Then, the cells were transfected with 20 nM si-negative
control (siNC; siN0000001-1-5; 5′-UUC UCC GAA CGU GUC ACG UTT -3′),
20 nM siLINC00707 (siG141020094742-1-5, 5′-GCA GGA ACA UCA CCA UCU
UUU -3′), 50 nM miR-103a-3p mimic (M, miR10000101-1-5; 5′-AGC AGC
AUU GUA CAG GGC UAUGA-3′), 50 nM mimic NC (MC;
miRB160401025525-2-1; 5′-UCA CAA CCU CCU AGA AAG AGU AGA -3′), 50
nM miR-103a-3p inhibitor (I; miR20000101-1-5; 5′-TCA TAG CCC TGT
ACA ATG CTG CT-3′) and 50 nM inhibitor NC (IC; miR2N0000001-1-5;
5′-CAG TAC TTT TGT GTA GTA CAA -3′) using Lipofectamine®
3000 (cat. no. L3000015; Invitrogen; Thermo Fisher Scientific,
Inc.). si-NC, siLINC00707, M, I, MC and IC were purchased from
Guangzhou RiboBio Co., Ltd. (https://www.ribobio.com/). Cells were collected for
further experiments 48 h after transfection.
To investigate the effect of overexpression of DKK1,
cells were co-transfected with 0.5 µg pcDNA3.1 empty vector
(NC; Invitrogen; Thermo Fisher Scientific, Inc.) and M or MC, and
co-transfected with 0.5 µg pcDNA3.1-DKK1 vector (DKK1) and M
or MC. Cell transfection was performed using
Lipofectamine® 3000 transfection reagent Invitrogen kit
(cat. no. L3000015; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. Cells were prepared at a confluence of
70-90% before transfection, Lipofectamine® 3000 reagent
and RNAs (100 ng/well) were diluted in Opti-MEM® medium
(cat. no. 31985062; Invitrogen; Thermo Fisher Scientific, Inc.).
Then, Lipofectamine® 3000 reagent-mix and the diluted
RNAs were added to the diluted Lipofectamine® 3000
reagent. Cells were incubated at room temperature for 10 min. Then,
50 µl RNA-lipid complex was added to cells and incubated for
48 h at 37°C. The transfection efficiency of LINC00707 and
miR-103a-3p was analyzed using reverse transcription-quantitative
PCR (RT-qPCR).
Alizarin red s staining (ARS)
Cells were fixed with 70% ice ethanol at room
temperature for 1 h and washed five times with ddH2O.
Cells were then stained with 40 mM ARS (cat. no. ECM815; EMD
Millipore) for 10-15 min at room temperature while gently stirring.
After staining, cells were washed five times with ddH2O
and the orange-red spots were identified as calcified nodules under
a light microscope (magnification, ×200).
Bioinformatics prediction
The biological target gene for LINC00707 was
predicted using StarBase v2.0 (http://starbase.sysu.edu.cn/index.php). Gene
prediction of the miR-103a-3p biological target gene was performed
using TargetScan7.2 (http://www.targetscan.org/vert_72/).
Dual-luciferase activity assay
The binding sites of LINC00707-wild-type (WT),
LINC00707-mutant (MUT), DKK1-WT and DKK1-MUT DNA were cloned into
pmirGLO luciferase vectors (cat. no. E1330; Promega Corporation).
To investigate the binding of LINC00707 to miRNA, HBMSCs were
transfected with 50 nM I in OM, while the transfection of 50 nM M
into HBMSCs was used to determine the binding of miR-103a-3p to the
target gene DKK1. The cell transfection was performed with
Lipofectamine® 3000 (Thermo Fisher Scientific, Inc.)
Cells transfected with IC or MC served as blank control. Luciferase
activity was determined using a TransDetect®
Double-Luciferase Reporter Assay kit (cat. no. FR201-01; TransGen
Biotech Co., Ltd.) according to the manufacturer's instructions.
After removing the cell culture medium, cells were carefully rinsed
twice in PBS, 100 µl 1X cell lysis buffer was added, cells
were fully lysed at room temperature for 10 min and centrifuged
(12,000 × g; 4°C; 10 min) to obtain the supernatant. Then, 100
µl Luciferase Reaction reagent (at room temperature) was
added, 20 µl cell lysate was carefully pipetted into the
tube and mixed gently. The activity of the firefly luciferase
reporter gene was measured using a luminescence meter (Molecular
Devices, LLC; SpectraMaxL) after 48 h transfection. Then, 100
µl Luciferase Reaction Reagent II (at room temperature) was
added to the aforementioned reaction tube, vortexed and the
activity of the Renilla luciferase reporter gene was
detected using the luminescence meter.
RT-qPCR
The lysis of each group of cells, and the extraction
of total RNAs were all conducted at 4°C using TRIzol®
(cat. no. 15596-026; Invitrogen; Thermo Fisher Scientific, Inc.).
The quality and integrity of RNAs were detected using a NanoDrop
One instrument (Thermo Fisher Scientific, Inc.) and 1% modified
agarose gel electrophoresis. The reverse transcription reaction of
miRNA, lncRNA and mRNA was performed using TaqMan™ MicroRNA RT kit
(cat. no. 4366597; Thermo Fisher Scientific, Inc.) and TaqMan™ RT
reagents (cat. no. N8080234; Thermo Fisher Scientific, Inc.). The
reverse transcription reaction condition were set to 42°C for 30
min, and reverse transcriptase inactivation was conducted at 85°C
for 15 min. qPCR was performed using SYBR Premix Ex TaqTM II
(Takara Biotechnology Co., Ltd.) in the Opticon RT-PCR detection
system (ABI 7500; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The thermocycling conditions were as
follows: Initial denaturation at 95°C for 10 min, followed by 45
cycles at 95°C for 30 sec, 60°C for 1 min and 60°C for 1 min, and
preserved at 4°C. The relative expression levels were calculated
using the 2−ΔΔCq method (25). GAPDH served as an internal control
for mRNAs and lncRNAs, and U6 was an internal control for
miRNAs.
The sequences of primers were as follows: Alkaline
phosphatase (ALP)-forward (F), 5′-ACC ACC ACG AGA GTG AAC CA-3′ and
reverse (R), 5′-CGT TGT CTG AGT ACC AGT CCC-3′; osteocalcin
(OCN)-F, 5′-GGC GCT ACC TGT ATC AAT GG-3′ and R: 5′-GTG GTC AGC CAA
CTC GTC A-3′; osteopontin (OPN)-F, 5′-GGA GTT GAA TGG TGC ATA CAA
GG-3′ and R, 5′-CCA CGG CTG TCC CAA TCA G-3′; RUNX2-F, 5′-TGT CAT
GGC GGG TAA CGA T-3′ and R, 5′-AAG ACG GTT ATG GTC AAG GTG AA-3′;
miR-103a-3p-F, 5′-TGT ACA GGG CTA TGA GTC GT-3′ and R, 5′-TCG TAT
CCA GTG CGT G TC G-3′; LINC00707-F, 5′-TGG AAA GTA AGC CTA TTA CAT
ATA C-3′ and R, 5′-GGT ATC ACC AAC AAC CCT GA-3′; DKK1-F, 5′-CAG
TGC CAC CTT GAA CTC AGT -3′ and R, 5′-CCG CCC TCA TAG AGA ACT
CC-3′; GAPDH-F, 5′-GTC AGC CGC ATC TTC TTT TG-3′ and R, 5′-GCG CCC
AAT ACG ACC AAA TC-3′; and U6-F, 5′-CTC GCT TCG GCA GCA CA-3′ and
R, 5′-AACGCTTCACGAATTTGCGT-3′. Each sample was analyzed in
triplicate.
Western blotting (WB)
An RIPA lysate (cat. no. 89901; Thermo Fisher
Scientific, Inc.) containing protease inhibitor and phosphatase
inhibitor was used to extract total proteins from HBMSCs, and the
protein concentration was detected using a Pierce™ Rapid Gold
bicinchoninic acid protein assay kit (cat. no. A53227; Thermo
Fisher Scientific, Inc.). A total of 30 µg protein was
separated by 10% SDS-PAGE and transferred to PVDF membranes (cat.
no. HVLP04700; EMD Millipore), which were rinsed in TBS and blocked
with 5% non-fat milk powder solution at 37°C for 1 h. Then, the
membranes were incubated with antibodies against ALP (39 kDa;
Rabbit; 1:1,000; cat. no. ab83259; Abcam), OCN (12 kDa; Mouse;
1:1,000; cat. no. ab13420; Abcam), OPN (66 kDa; Rabbit; 1:1,000;
cat. no. ab8448; Abcam), RUNX2 (57 kDa; Mouse; 1:1,000; cat. no.
ab76956; Abcam) or GAPDH (36 kDa; Mouse; 1:1,000; cat. no. ab8245;
Abcam) overnight at 4°C. The target bands were then incubated with
a horseradish peroxidase-conjugated secondary antibody (1:1,000;
cat. nos. ab205718 and ab205719; Abcam) for 2 h at room
temperature. SignalFire™ ECL reagent (cat. no. 6883; Cell Signaling
Technology, Inc.) was used to detect the signals and the gray
values of the bands were analyzed using ImageJ (version 5.0;
National Institute of Health).
Statistical analysis
Data are presented as the mean ± SD. Comparisons
between the groups were performed using one-way ANOVA, followed by
Dunnett test or Tukey's test using GraphPad Prism 7.0 (Graph-Pad
Software, Inc.). P<0.05 was considered to indicate a
statistically significant difference. All experiments were
performed in triplicate.
Results
Expression levels of LINC00707,
miR-103a-3p and osteogenesis-related genes in HBMSCs cultured in PM
and OM
The expression levels of lncRNA LINC00707,
miR-103a-3p and osteogenesis-related genes in HBMSCs cultured in PM
and OM were detected by RT-qPCR. There was no changes in the
expression levels of LINC00707, miR-103a-3p, ALP, OCN, OPN and
RUNX2 in HBMSCs cultured in PM over time (P>0.05; Fig. 1). However, the expression of
LINC00707 was reduced, while those of miR-103a-3p, ALP, OCN, OPN
and RUNX2 were significantly increased in OM-cultured HBMSCs with
prolonged culture duration (P<0.001). It was also found that the
osteogenesis-related genes were all significantly increased during
osteogenic induction from day 3 compared with day 0 (Fig. 1); therefore, in the follow-up
experiment, the expression levels of osteogenic genes were detected
at 3 days of osteogenic induction.
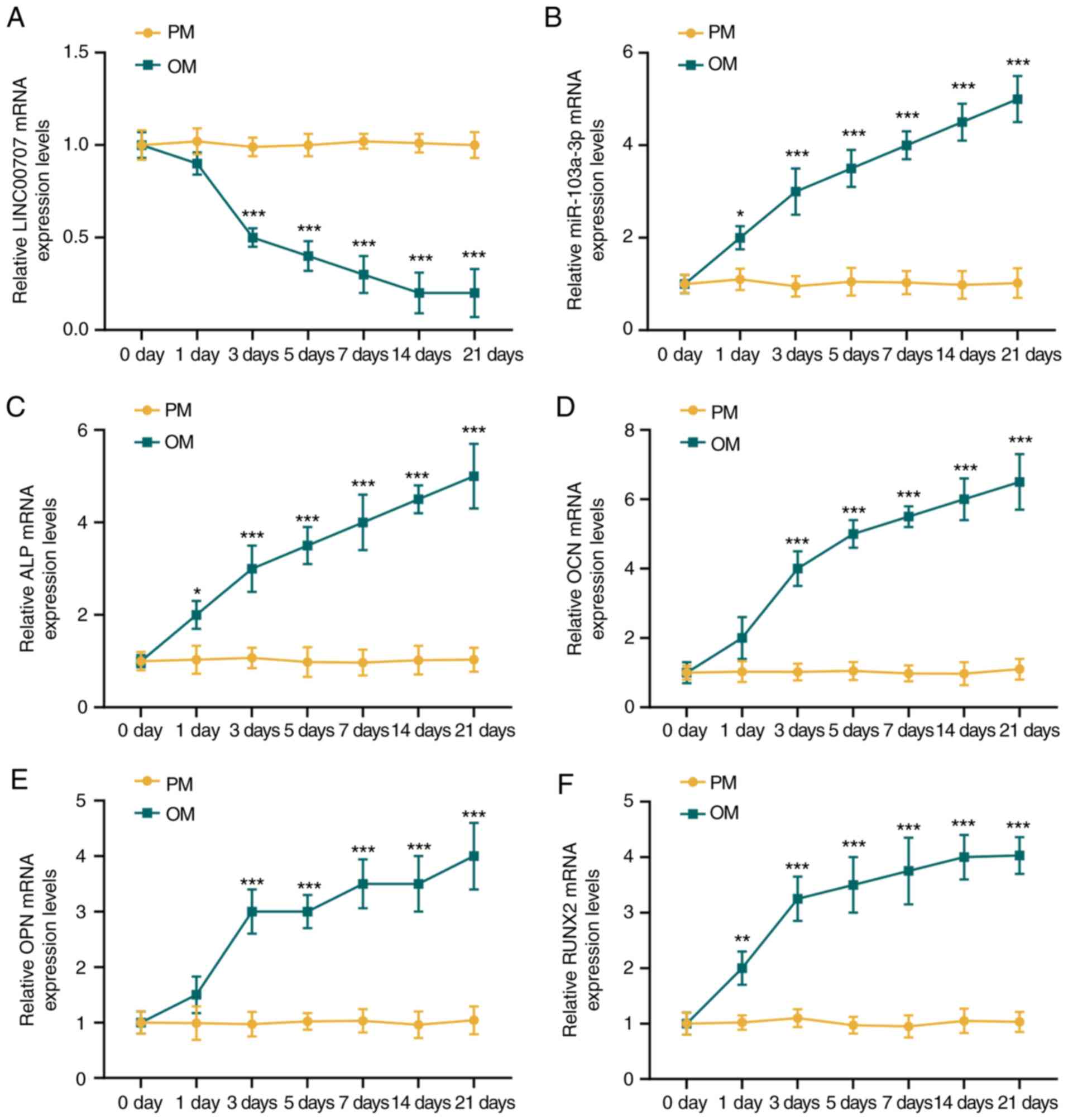 | Figure 1Expression levels of LINC00707,
miR-103a-3p and osteogenesis-related genes in HBMSCs cultured in PM
and OM. (A) LINC00707 and (B) miR-103a-3p expression levels in
HBMSCs cultured in PM and OM were measured by RT-qPCR. Expression
levels of the osteogenesis-related genes (C) ALP, (D) OCN, (E) OPN
and (F) RUNX2 in HBMSCs cultured in PM and OM were measured by
RT-qPCR. Each experiment was repeated three times, GAPDH served as
an internal control for mRNA and long non-coding RNA, and U6 served
as an internal control for miRNAs. *P<0.05,
**P<0.01, ***P<0.001 vs. 0 days.
HBMSCs, human bone mesenchymal stem cells; PM, proliferation
medium; OM, osteogenic medium; RT-qPCR, reverse
transcription-quantitative PCR; miR/miRNA, microRNA; LINC00707,
Long intergenic non-protein coding RNA 00707; ALP, Alkaline
phosphatase; OCN, osteocalcin; OPN, osteo-pontin; RUNX2, RUNX
family transcription factor 2. |
To further investigate the effects of PM and OM on
osteogenic differentiation, ARS was performed to detect mineralized
matrix deposition in PM and OM, and it was observed that
mineralized matrix deposition in OM was notably increased compared
with PM (Fig. 2A).
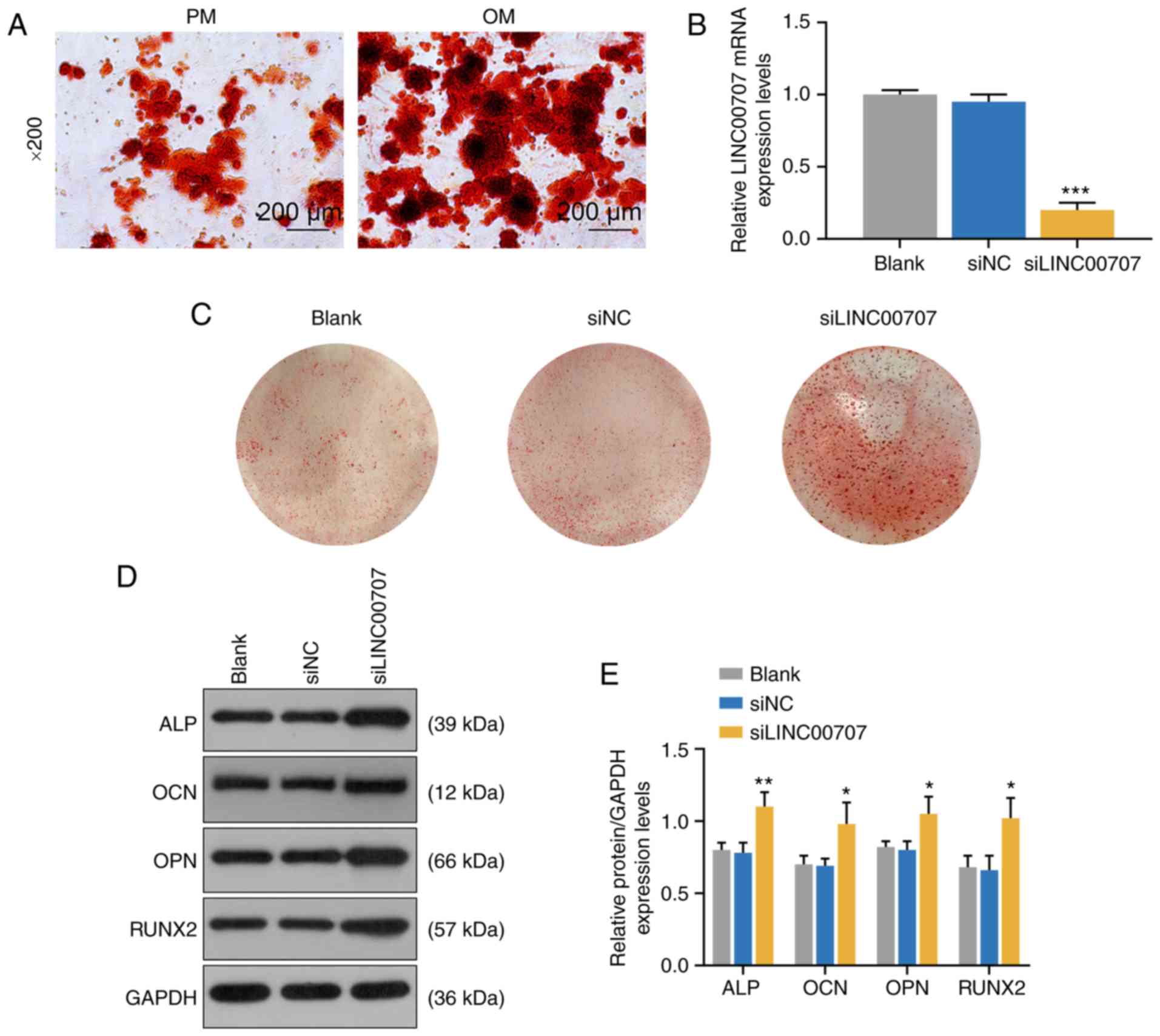 | Figure 2Effects of PM, OM and LINC00707
silencing on osteogenic differentiation of HBMSCs. (A) Alizarin Red
Staining of PM and OM were performed on day 21. Scale bar, 200
µm. Magnification, ×200. (B) LINC00707 expression after
transfection with blank, siNC or siLINC00707 in HBMSCs cultured in
OM was analyzed by reverse transcription-quantitative PCR. Each
experiment was repeated three times, GAPDH served as an internal
control. (C) Alizarin Red Staining of blank, siNC and siLINC00707
in HBMSCs cultured in OM was performed on day 21. (D) Western
blotting results of the (E) protein expression levels of
osteogenesis-related genes (ALP, OCN, OPN and RUNX2) in blank, siNC
and siLINC00707 group. Each experiment was repeated three times,
GAPDH served as an internal control. *P<0.05,
**P<0.01, ***P<0.001 vs. siNC. HBMSCs,
human bone mesenchymal stem cells; PM, proliferation medium; OM,
osteogenic medium; NC, negative control; LINC00707, Long intergenic
non-protein coding RNA 00707; ALP, Alkaline phosphatase; OCN,
osteocalcin; OPN, osteopontin; RUNX2, RUNX family transcription
factor 2; siRNA, small interfering RNA. |
Effects of LINC00707 silencing on
osteogenic differentiation of HBMSCs
To investigate the effect of LINC00707 on osteogenic
differentiation of HBMSCs, blank, siNC and siLINC00707 groups of
OM-cultured HBMSCs were established (P<0.001; Fig. 2B), and the mineralization matrix
deposition of the three groups were examined by ARS. The results
indicated that siLINC00707 markedly increased mineralized matrix
deposition (Fig. 2C). In
addition, WB was performed to measure the expression levels of
osteogenesis-related genes of OM-cultured cells 3 days after the
incubation, and it was found that siLINC00707 significantly
increased the expression levels of ALP, OCN, OPN and RUNX2 compared
with siNC group (P<0.05; Fig. 2D
and E).
Relationship between LINC00707 and
miR-103a-3p in HBMSCs
The mechanism of LINC00707 on osteogenic
differentiation of HBMSCs was further investigated. StarBase
predicted that miR-103a-3p was the target miRNA of LINC00707
(Fig. 3A), and the prediction was
further demonstrated by the dual-luciferase activity assay results
(Fig. 3B). The results indicated
that cells in the LINC00707-WT and miR-103a-3p I group exhibited
significant enhanced luciferase activity, while no significant
difference was identified in luciferase activity in cells
co-transfected with LINC00707-MUT and miR-103a-3p I compared with
the blank group (P<0.05; Fig.
3B). Furthermore, the RT-qPCR assay results suggested that the
expression of miR-103a-3p was significantly higher in the
siLINC00707 group of HBMSCs compared with the siNC groups,
indicating that knocking down LINC00707 upregulated the expression
of miR-103a-3p (P<0.01; Fig.
3C).
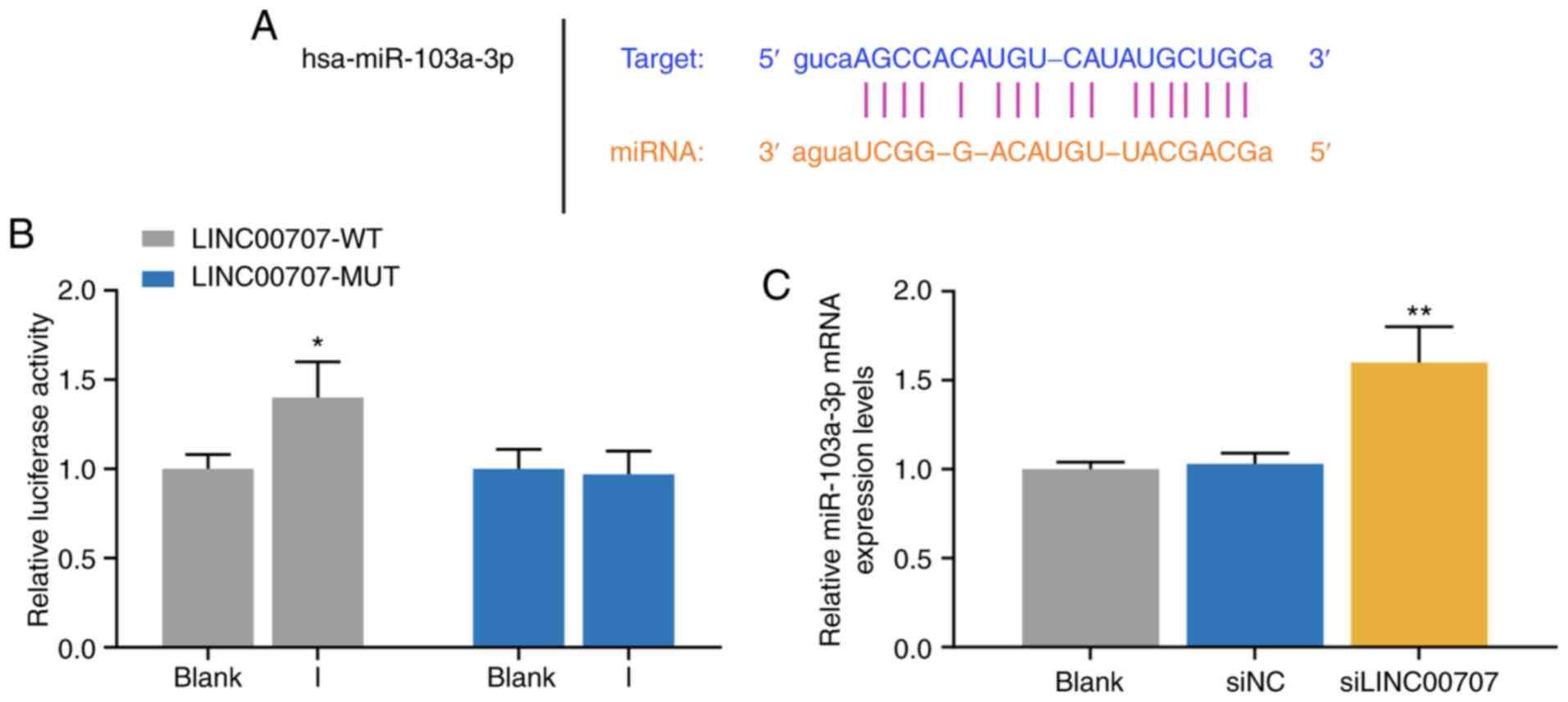 | Figure 3Relationship between the expression
levels of LINC00707 and miR-103a-3p in HBMSCs. (A) StarBase and (B)
dual-luciferase activity were used to identify the LINC00707
binding site in miR-103a-3p. *P<0.05 vs. blank. (C)
Expression of miR-103a-3p in blank, siNC and siLINC00707 group was
analyzed by reverse transcription-quantitative PCR. Each experiment
was repeated three times, and U6 served as an internal control.
**P<0.01 vs. siNC. HBMSCs, human bone mesenchymal
stem cells; PM, proliferation medium; OM, osteogenic medium; NC,
negative control; LINC00707, Long intergenic non-protein coding RNA
00707; siRNA, small interfering RNA; WT, wild-type; MUT, mutant;
miR/miRNA, microRNA. |
Effects of miR-103a-3p and LINC00707 on
expression levels of osteogenesis-related genes of HBMSCs
To further investigate the effect of miR-103a-3p on
the expression levels of osteogenesis-related genes, MC, M, IC, I
and siLINC00707 + I groups of HBMSCs were established. The RT-qPCR
assay results demonstrated that the mimic significantly increased
the expression of miR-103a-3p compared with MC group, while the
inhibitor downregulated the miR-103a-3p expression compared with IC
group, and miR-103a-3p expression was higher in the siLINC00707 + I
group compared with the I group (P<0.001; Fig. 4A).
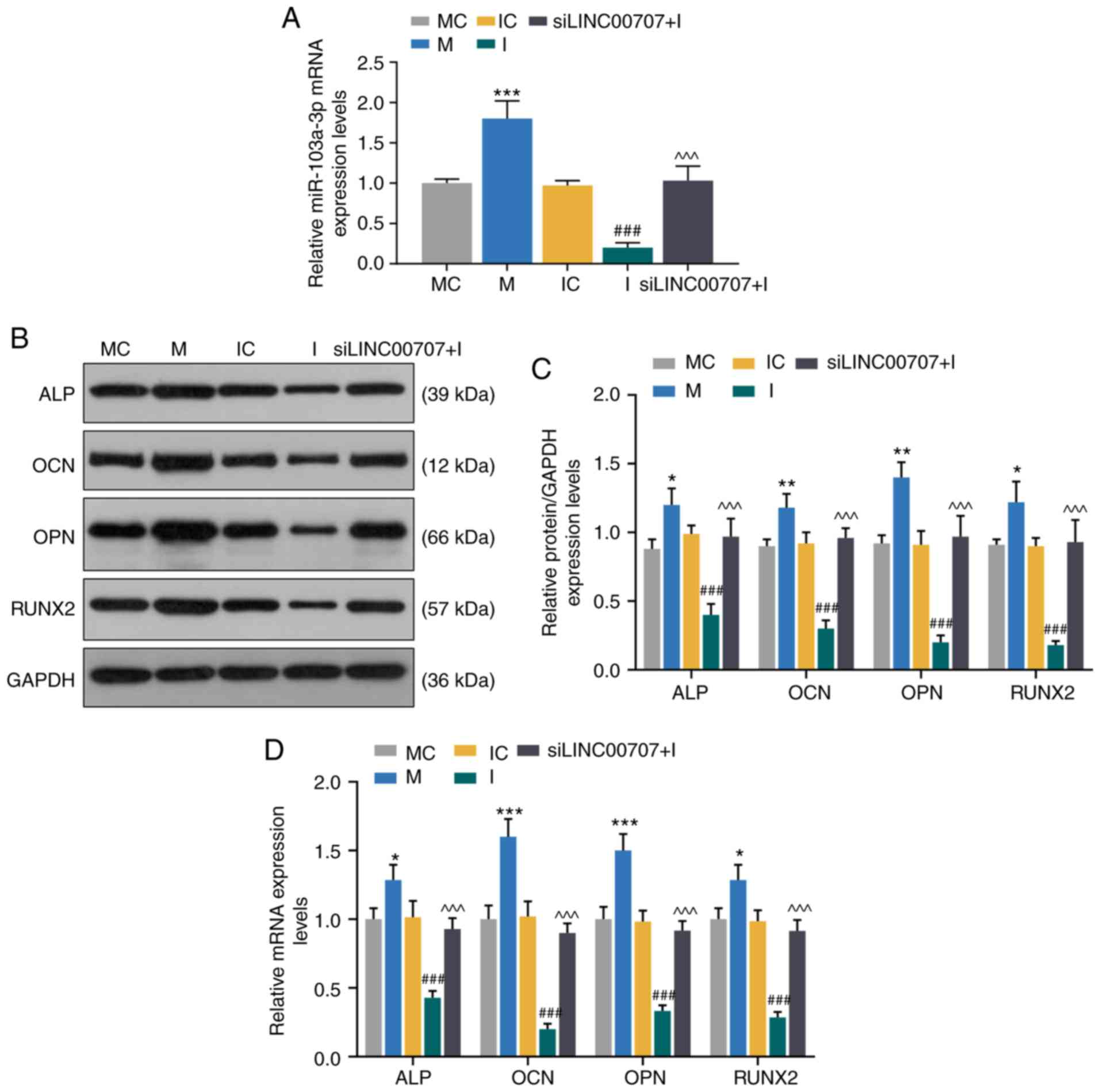 | Figure 4Effects of miR-103a-3p and LINC00707
on expression levels of osteogenesis-related genes of HBMSCs. (A)
Expression of miR-103a-3p after trans-fection with MC, M, IC, I or
siLINC00707 + I in HBMSCs was analyzed by RT-qPCR. Each experiment
was repeated three times, and U6 served as an internal control. (B)
Western blotting results of the (C) protein expression levels of
osteogenesis-related genes (ALP, OCN, OPN and RUNX2) in MC, M, IC,
I and siLINC00707 + I HBMSCs. (D) mRNA expression levels of
osteogenesis-related genes (ALP, OCN, OPN and RUNX2) in MC, M, IC,
I and siLINC00707 + I HBMSCs were detected by RT-qPCR. Each
experiment was repeated three times, and GAPDH served as an
internal control. *P<0.05, **P<0.01,
***P<0.001 vs. MC; ###P<0.001 vs. IC;
^^^P<0.001 vs. I. HBMSCs, human bone mesenchymal stem
cells; RT-qPCR, reverse transcription-quantitative PCR; MC, mimic
control; IC, inhibitor control; I, inhibitor; M, mimic; LINC00707,
Long intergenic non-protein coding RNA 00707; siRNA, small
interfering RNA; ALP, Alkaline phosphatase; OCN, osteocalcin; OPN,
osteopontin; RUNX2, RUNX family transcription factor 2. |
The expression levels of osteogenesis-related genes
of the OM-cultured cells 3 days after the incubation were detected
by WB and RT-qPCR. The results identified that the mimic
upregulated the mRNA and protein expression levels of ALP, OCN, OPN
and RUNX2 compared with MC group, while the inhibitor downregulated
the mRNA and protein expression levels of ALP, OCN, OPN and RUNX2
compared with IC group (P<0.001; Fig. 4B-D). In addition, the mRNA and
protein expression levels of ALP, OCN, OPN and RUNX2 were
significantly higher in siLINC00707 + I group compared with the I
group, indicating that upregulation of miR-103a-3p increased the
mRNA and protein expression levels of osteogenesis-related genes
(P<0.001; Fig. 4B-D).
miR-103a-3p increases the expression
levels of osteogenesis-related genes of HBMSCs by regulating DKK1
expression
TargetScan7.2 predicted that DKK1 was a target gene
for miR-103a-3p in HBMSCs (Fig.
5A), and this was further demonstrated by dual-luciferase
activity assay, as it was found that the mimic significantly
reduced the luciferase activity of DKK1-WT (P<0.001; Fig. 5B).
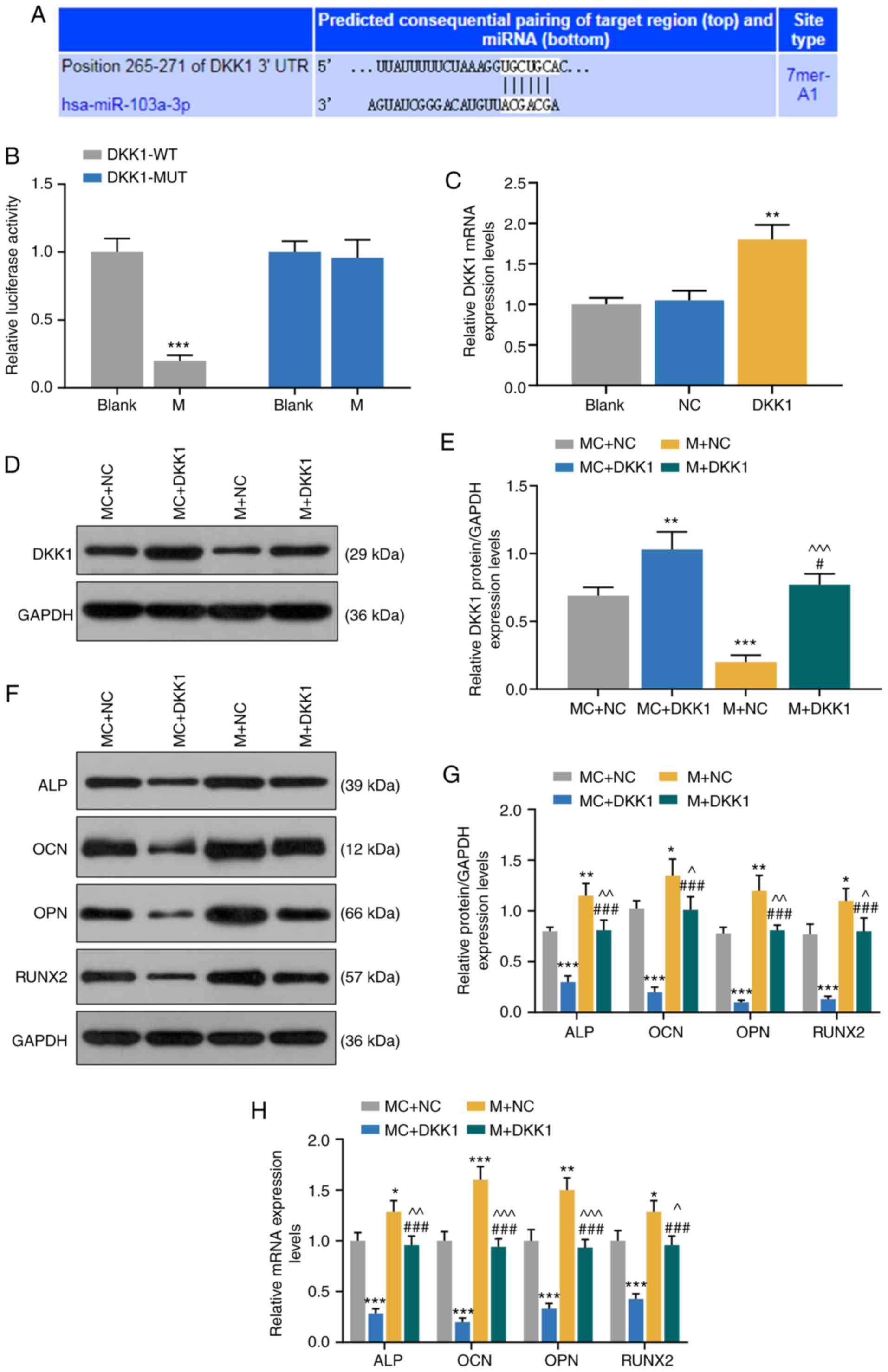 | Figure 5miR-103a-3p promotes expression
levels of osteogenesis-related genes in HBMSCs by regulating DKK1.
(A) TargetScan7.2 and (B) dual-luciferase activity were performed
to identify the miR-103a-3p binding site in DKK1.
***P<0.001 vs. blank. (C) Expression of DKK1 in
blank, NC and DKK1 group was detected by RT-qPCR. (D) Western
blotting results of (E) DKK1 transfection with MC + NC, MC + DKK1,
M + NC or M + DKK1 in HBMSCs. (F) Western blotting results of (G)
protein expression levels of osteogenesis-related genes (ALP, OCN,
OPN and RUNX2) in MC + NC, MC + DKK1, M + NC and M + DKK1 group.
(H) mRNA expression levels of osteogenesis-related genes (ALP, OCN,
OPN and RUNX2) in MC, M, IC, I and siLINC00707 + I group were
detected by RT-qPCR. Each experiment was repeated three times, and
GAPDH served as an internal control. *P<0.05,
**P<0.01, ***P<0.001 vs. MC + NC;
#P<0.05, ###P<0.001 vs. MC + DKK1;
^P<0.05, ^^P<0.01,
^^^P<0.001 vs. M + NC. HBMSCs, human bone mesenchymal
stem cells; RT-qPCR, reverse transcription-quantitative PCR; MC,
mimic control; IC, inhibitor control; I, inhibitor; M, mimic; NC,
negative control; LINC00707, Long intergenic non-protein coding RNA
00707; siRNA, small interfering RNA; ALP, Alkaline phosphatase;
OCN, osteocalcin; OPN, osteopontin; RUNX2, RUNX family
transcription factor 2; miR/miRNA, microRNA; WT, wild-type; MUT,
mutant; DKK1, dickkopf WNT signaling pathway inhibitor 1. |
To further investigate the effects of miR-103a-3p
and DKK1 on osteogenic differentiation, MC + NC, MC + DKK1, M + NC
and M + DKK1 groups were established, and the transfection
efficiency of DKK1 was detected by RT-qPCR. The results identified
that DKK1 expression was significantly increased in DKK1 group
compared with the NC group (P<0.001; Fig. 5C). Moreover, the expression of
DKK1 was higher in MC + DKK1 group but lower in M + NC group,
compared with the MC + NC group (P<0.001, Fig. 5D and E). It was also found that
DKK1 expression in M + DKK1 group was lower compared with the MC +
DKK1 group, but higher compared with the M + NC group (P<0.001;
Fig. 5D and E).
The expression levels of osteogenesis-related genes
of OM-cultured HBMSCs 3 days after the incubation were detected by
WB and RT-qPCR, and it was identified that the mRNA and protein
expression levels of ALP, OCN, OPN and RUNX2 were lower in MC +
DKK1 group compared with the MC + NC group. Furthermore, the mRNA
and protein expression levels of ALP, OCN, OPN and RUNX2 in M + NC
group were higher compared with the MC+NC group, while these were
reduced in the M + DKK1 group compared with the M + NC group
(P<0.001; Fig. 5F-H).
Discussion
lncRNAs serve important roles in a variety of
biological processes, such as embryonic development, angiogenesis
and cell differentiation, and participate in a several of cellular
regulation activities, such as transcriptional activation,
transcriptional interference and intranuclear transport (6,26).
For example, Kikuchi et al (27) showed that adipogenesis
up-regulated transcript 1 may be involved in the differentiation of
MSCs into adipocytes. MSCs are important sources of multifunctional
cells for tissue repair, and lncRNAs function in the regulation of
HBMSCs (28). Jia et al
(29) reported that LINC00707
sponges miR-370-3p to promote osteogenesis of HBMSCs by
upregulating Wnt family member 2B. In addition, Cai et al
(30) revealed that LINC00707
promoted osteogenic differentiation of HBMSCs via the Wnt/β-catenin
pathway activated by the LINC00707/miR-145/LDL receptor related
protein 5 axis. The current study investigated LINC00707 and
osteogenesis-related genes (31,32) in HBMSCs cultured in PM and OM.
lncRNAs are significantly correlated with MSCs in
osteogenic differentiation (33-36). Zhu and Xu (37) reported that Angelman syndrome
chromosome region (ANCR) inhibited the expression of RUNX2 by
interacting with enhancer of zeste 2 polycomb repressive complex 2
subunit, thus inhibiting the osteogenic differentiation of MSCs.
Moreover, Zuo et al (38)
studied lncRNA expression profiles in the early osteogenic
differentiation of murine MSCs, and found that 116 lncRNAs were
differentially expressed in the bone morphoge-netic protein
2-treated group compared with the control group. lncRNA LINC00707
is abnormally expressed in lung adeno-carcinoma and melanoma
(9,11), but its effect on osteogenic
differentiation of HBMSCs are not fully understood. ARS can be used
to measure mineralized matrix deposition (39,40). The current results demonstrated
that LINC00707 expression of OM-cultured HBMSCs was significantly
reduced, while there was no significant change in LINC00707
expression in PM-cultured HBMSCs, indicating that LINC00707 may be
involved in the regulation of osteogenic differentiation. In
addition, the mineralized matrix deposition in OM was markedly
increased compared with PM. It was also found that knocking down
LINC00707 promoted the expression levels of osteogenesis-related
genes ALP, OCN, OPN and RUNX2, suggesting that LINC00707 may have
similar function to ANCR in inhibiting osteogenic differentiation
(37).
Osteoblasts are generally derived from BM-derived
stromal stem cells (1). Moreover,
significant changes in miRNA profiles during BM-derived
morphogenetic protein-2-induced differentiation of BM stromal stem
cells into osteoblasts have been observed, and miR-133 and miR-135
are closely related to osteogenic differentiation (41). It has been previously reported
that the negative regulation of miR-138 on osteoblasts is caused by
mediating the focal adhesion kinase/ERK1/2 signaling pathway
(42). Moreover, the
miR-196a/homeobox C8 pathway regulates the differentiation and
proliferation of adipose stem cells into osteoblasts (43), while miR-103a-3p may play a key
role in osteogenic differentiation (21). The role of miR-103a-3p in
pancreatic cancer (44), bladder
cancer (45) and gastric cancer
cells (46) has also been
previously revealed, but the specific regulatory mechanism of
miR-103a-3p in osteogenic differentiation of HBMSCs remains to be
investigated (47). The present
results suggested that the expression of miR-103a-3p in OM-cultured
HBMSCs was significantly increased, and that miR-103a-3p was a
targeted gene for LINC00707, indicating that miR-103a-3p had
opposite effects on osteogenic differentiation compared with
miR-133 and miR-135 (41).
Different miRNAs play diverse roles in osteogenic differentiation
(48). Previous studies have
shown that miR-210 inhibits the transforming growth
factor-β/activin signaling pathway by downregulating the expression
of activin A receptor type 1B protein, thus promoting cell
osteogenic differentiation (49).
In the present study, overexpression of miR-103a-3p promoted the
expression levels of osteogenesis-related genes (ALP, OCN, OPN and
RUNX2), which could be reversed by inhibition of LINC00707.
DDK1 is an inhibitor of Wnt signaling (50,51). The Wnt pathway induces bone cell
differentiation, prevents bone aging and inflammation and is an
important pathway in bone metabolism (52). DDK1 binds to low-density
lipoprotein receptor-related proteins 5 or 6 and blocks
heterodimerization with frizzled, thus affecting osteogenic
differentiation (53).
Furthermore, Xia et al (54) demonstrated that miR-203 could
promote the differentiation of rat MSCs into osteoblast-like cells
by regulating the expression of DKK1. In the present study, DDK1
was identified as a target gene of miR-103a-3p, and it inhibited
the expression levels of ALP, OCN, OPN and RUNX2, while
overexpression of miR-103a-3p reversed the inhibitory effects of
DDK1 on the expression levels of osteogenesis-related genes.
lncRNAs expressed in cytoplasm can act as a molecular sponge of
miRNA and function as competing endogenous RNA (ceRNA) (55). In the current study, LINC00707
acted as a molecular decoy for miR-103a-3p, and LINC00707 also
served as a ceRNA to regulate miR-103a-3p and DKK1. However, this
study also has some limitations. For example, the effect of
LINC00707/miR-103a-3p on the expression levels of genes in the Wnt
pathway has yet to be detected, such as JNK and ERK1/2. In
addition, LINC00707 knockout mice should be generated to measure
the bone mineral density, which will provide evidence for the
function of LINC00707 in HBMSCs osteogenic differentiation.
In conclusion, during the osteogenic differentiation
of HBMSCs, the expression of LINC00707 was reduced, while that of
miR-103a-3p was increased. Moreover, inhibition of LINC00707
promoted osteogenic differentiation gene expression of HBMSCs by
targeting miR-103a-3p to regulate the DKK1 signaling pathway. Thus,
the current findings provide evidence for further study on the
regulatory mechanisms of osteogenic differentiation of HBMSCs.
Acknowledgments
Not applicable.
Funding
This study was supported by the Science and
Technology Development Plan Project of Jilin Province (grant no.
20180520128JH).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
Substantial contributions to conception and design:
JL. Data acquisition, data analysis and interpretation: MW, GF, RL,
YW and JJ. Drafting the article or critically revising it for
important intellectual content: JL. Final approval of the version
to be published: All authors. Agreement to be accountable for all
aspects of the work in ensuring that questions related to the
accuracy or integrity of the work are appropriately investigated
and resolved: All authors.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Liu H, Xia X and Li B: Mesenchymal stem
cell aging: Mechanisms and influences on skeletal and non-skeletal
tissues. Exp Biol Med (Maywood). 240:1099–1106. 2015. View Article : Google Scholar
|
|
2
|
Ji N, Yu JW, Ni XC, Wu JG, Wang SL and
Jiang BJ: Bone marrow-derived mesenchymal stem cells increase drug
resistance in CD133-expressing gastric cancer cells by regulating
the PI3K/AKT pathway. Tumour Biol. 37:14637–14651. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Polymeri A, Giannobile WV and Kaigler D:
Bone marrow stromal stem cells in tissue engineering and
regenerative medicine. Horm Metab Res. 48:700–713. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Joshi R, Goihberg E, Ren W, Pilichowska M
and Mathew P: Proteolytic fragments of fibronectin function as
matrikines driving the chemotactic affinity of prostate cancer
cells to human bone marrow mesenchymal stromal cells via the α5β1
integrin. Cell Adh Migr. 11:305–315. 2017. View Article : Google Scholar
|
|
5
|
Takizawa N, Okubo N, Kamo M, Chosa N,
Mikami T, Suzuki K, Yokota S, Ibi M, Ohtsuka M, Taira M, et al:
Bone marrow-derived mesenchymal stem cells propagate
immu-nosuppressive/anti-inflammatory macrophages in cell-to-cell
contact-independent and -dependent manners under hypoxic culture.
Exp Cell Res. 358:411–420. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Han P and Chang CP: Long non-coding RNA
and chromatin remodeling. RNA Biol. 12:1094–1098. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jain S, Thakkar N, Chhatai J, Pal Bhadra M
and Bhadra U: Long non-coding RNA: Functional agent for disease
traits. RNA Biol. 14:522–535. 2017. View Article : Google Scholar :
|
|
8
|
Kondo Y, Shinjo K and Katsushima K: Long
non-coding RNAs as an epigenetic regulator in human cancers. Cancer
Sci. 108:1927–1933. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang S, Xu J and Zeng X: A six-long
non-coding RNA signature predicts prognosis in melanoma patients.
Int J Oncol. 52:1178–1188. 2018.PubMed/NCBI
|
|
10
|
Li T, Li Y and Sun H: MicroRNA-876 is
sponged by long noncoding RNA LINC00707 and directly targets
metadherin to inhibit breast cancer malignancy. Cancer Manag Res.
11:5255–5269. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ma T, Ma H, Zou Z, He X, Liu Y, Shuai Y,
Xie M and Zhang Z: The long intergenic noncoding RNA 00707 promotes
lung adenocarcinoma cell proliferation and migration by regulating
Cdc42. Cell Physiol Biochem. 45:1566–1580. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tu J, Zhao Z, Xu M, Chen M, Weng Q, Wang J
and Ji J: LINC00707 contributes to hepatocellular carcinoma
progression via sponging miR-206 to increase CDK14. J Cell Physiol.
234:10615–10624. 2019. View Article : Google Scholar
|
|
13
|
Tüfekci KU, Oner MG, Meuwissen RL and Genç
S: The role of microRNAs in human diseases. Methods Mol Biol.
1107:33–50. 2014. View Article : Google Scholar
|
|
14
|
Horsburgh S, Fullard N, Roger M, Degnan A,
Todryk S, Przyborski S and O'Reilly S: MicroRNAs in the skin: Role
in development, homoeostasis and regeneration. Clin Sci (Lond).
131:1923–1940. 2017. View Article : Google Scholar
|
|
15
|
Wen D, Peng Y, Liu D, Weizmann Y and
Mahato RI: Mesenchymal stem cell and derived exosome as small RNA
carrier and Immunomodulator to improve islet transplantation. J
Control Release. 238:166–175. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ren G, Sun J, Li MM, Zhang YD, Li RH and
Li YM: MicroRNA-23a-5p regulates osteogenic differentiation of
human bone marrow-derived mesenchymal stem cells by targeting
mitogen-activated protein kinase-13. Mol Med Rep. 17:4554–4560.
2018.PubMed/NCBI
|
|
17
|
Jiang C and Gong F: MiR-148a promotes
myocardial differentiation of human bone mesenchymal stromal cells
via DNA methyltransferase 1 (DNMT1). Cell Biol Int. 42:913–922.
2018. View Article : Google Scholar
|
|
18
|
Mi W, Shi Q, Chen X, Wu T and Huang H:
miR-33a-5p modulates TNF-α-inhibited osteogenic differentiation by
targeting SATB2 expression in hBMSCs. FEBS Lett. 590:396–407. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yan J, Guo D, Yang S, Sun H, Wu B and Zhou
D: Inhibition of miR-222-3p activity promoted osteogenic
differentiation of hBMSCs by regulating Smad5-RUNX2 signal axis.
Biochem Biophys Res Commun. 470:498–503. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang H, Xie Z, Hou T, Li Z, Huang K, Gong
J, Zhou W, Tang K, Xu J and Dong S: MiR-125b regulates the
osteogenic differentiation of human mesenchymal stem cells by
targeting BMPR1b. Cell Physiol Biochem. 41:530–542. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ragni E, Perucca Orfei C, De Luca P,
Colombini A, Viganò M, Lugano G, Bollati V and de Girolamo L:
Identification of miRNA reference genes in extracellular vesicles
from adipose derived mesenchymal stem cells for studying
osteoarthritis. Int J Mol Sci. 20:11082019. View Article : Google Scholar :
|
|
22
|
Li ZH, Hu H, Zhang XY, Liu GD, Ran B,
Zhang PG, Liao MM and Wu YC: MiR-291a-3p regulates the BMSCs
differentiation via targeting DKK1 in dexamethasone-induced
osteoporosis. Kaohsiung J Med Sci. 36:35–42. 2020. View Article : Google Scholar
|
|
23
|
Komori T: Glucocorticoid signaling and
bone biology. Horm Metab Res. 48:755–763. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Butler JS, Queally JM, Devitt BM, Murray
DW, Doran PP and O'Byrne JM: Silencing Dkk1 expression rescues
dexamethasone-induced suppression of primary human osteoblast
differentiation. BMC Musculoskelet Disord. 11:2102010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
26
|
Ransohoff JD, Wei Y and Khavari PA: The
functions and unique features of long intergenic non-coding RNA.
Nat Rev Mol Cell Biol. 19:143–157. 2018. View Article : Google Scholar :
|
|
27
|
Kikuchi K, Fukuda M, Ito T, Inoue M, Yokoi
T, Chiku S, Mitsuyama T, Asai K, Hirose T and Aizawa Y: Transcripts
of unknown function in multiple-signaling pathways involved in
human stem cell differentiation. Nucleic Acids Res. 37:4987–5000.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sun X, Jia B, Qiu XL, Chu HX, Zhang ZQ,
Wang ZP and Zhao JJ: Potential functions of long non-coding RNAs in
the osteogenic differentiation of human bone marrow mesenchymal
stem cells. Mol Med Rep. 19:103–114. 2019.
|
|
29
|
Jia B, Wang Z, Sun X, Chen J, Zhao J and
Qiu X: Long noncoding RNA LINC00707 sponges miR-370-3p to promote
osteogenesis of human bone marrow-derived mesenchymal stem cells
through upregulating WNT2B. Stem Cell Res Ther. 10:672019.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cai WL, Zeng W, Liu HH, Zhu BY, Liu JL and
Liu Y: LncRNA LINC00707 promotes osteogenic differentiation of
hBMSCs through the Wnt/β-catenin pathway activated by
LINC00707/miR-145/LRP5 axis. Eur Rev Med Pharmacol Sci. 24:18–28.
2020.PubMed/NCBI
|
|
31
|
An J, Yang H, Zhang Q, Liu C, Zhao J,
Zhang L and Chen B: Natural products for treatment of osteoporosis:
The effects and mechanisms on promoting osteoblast-mediated bone
formation. Life Sci. 147:46–58. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang Q, Li Y and Zhang Y, Ma L, Lin L,
Meng J, Jiang L, Wang L, Zhou P and Zhang Y: LncRNA MEG3 inhibited
osteogenic differentiation of bone marrow mesenchymal stem cells
from postmenopausal osteoporosis by targeting miR-133a-3p. Biomed
Pharmacother. 89:1178–1186. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang J, Tao Z and Wang Y: Long non-coding
RNA DANCR regulates the proliferation and osteogenic
differentiation of human bone-derived marrow mesenchymal stem cells
via the p38 MAPK pathway. Int J Mol Med. 41:213–219. 2018.
|
|
34
|
Cui Y, Lu S, Tan H, Li J, Zhu M and Xu Y:
Silencing of long non-coding RNA NONHSAT009968 ameliorates the
staphylococcal protein A-inhibited osteogenic differentiation in
human bone mesenchymal stem cells. Cell Physiol Biochem.
39:1347–1359. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gao Y, Xiao F, Wang C, Wang C, Cui P,
Zhang X and Chen X: Long noncoding RNA MALAT1 promotes osterix
expression to regulate osteogenic differentiation by targeting
miRNA-143 in human bone marrow-derived mesenchymal stem cells. J
Cell Biochem. 119:6986–6996. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang J, Miao J, Meng X, Chen N and Wang Y:
Expression of long non-coding RNAs in human bone marrow mesenchymal
stem cells co-cultured with human amnion-derived mesenchymal stem
cells. Mol Med Rep. 16:6683–6689. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhu L and Xu PC: Downregulated LncRNA-ANCR
promotes osteoblast differentiation by targeting EZH2 and
regulating Runx2 expression. Biochem Biophys Res Commun.
432:612–617. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zuo C, Wang Z, Lu H, Dai Z, Liu X and Cui
L: Expression profiling of lncRNAs in C3H10T1/2 mesenchymal stem
cells undergoing early osteoblast differentiation. Mol Med Rep.
8:463–467. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Puchtler H, Meloan SN and Terry MS: On the
history and mechanism of alizarin and alizarin red S stains for
calcium. J Histochem Cytochem. 17:110–124. 1969. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Paul H, Reginato AJ and Schumacher HR:
Alizarin red S staining as a screening test to detect calcium
compounds in synovial fluid. Arthritis Rheum. 26:191–200. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li Z, Hassan MQ, Volinia S, van Wijnen AJ,
Stein JL, Croce CM, Lian JB and Stein GS: A microRNA signature for
a BMP2-induced osteoblast lineage commitment program. Proc Natl
Acad Sci USA. 105:13906–13911. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Eskildsen T, Taipaleenmäki H, Stenvang J,
Abdallah BM, Ditzel N, Nossent AY, Bak M, Kauppinen S and Kassem M:
MicroRNA-138 regulates osteogenic differentiation of human stromal
(mesenchymal) stem cells in vivo. Proc Natl Acad Sci USA.
108:6139–6144. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kim YJ, Bae SW, Yu SS, Bae YC and Jung JS:
miR-196a regulates proliferation and osteogenic differentiation in
mesenchymal stem cells derived from human adipose tissue. J Bone
Miner Res. 24:816–825. 2009. View Article : Google Scholar
|
|
44
|
Zhou H and Rigoutsos I: MiR-103a-3p
targets the 5′ UTR of GPRC5A in pancreatic cells. RNA.
20:1431–1439. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhong Z, Lv M and Chen J: Screening
differential circular RNA expression profiles reveals the
regulatory role of circTCF25-miR-103a-3p/miR-107-CDK6 pathway in
bladder carcinoma. Sci Rep. 6:309192016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hu X, Miao J, Zhang M, Wang X, Wang Z, Han
J, Tong D and Huang C: miRNA-103a-3p promotes human gastric cancer
cell proliferation by targeting and suppressing ATF7 in vitro. Mol
Cells. 41:390–400. 2018.PubMed/NCBI
|
|
47
|
Kim DS, Lee SY, Lee JH, Bae YC and Jung
JS: MicroRNA-103a-3p controls proliferation and osteogenic
differentiation of human adipose tissue-derived stromal cells. Exp
Mol Med. 47:e1722015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sun H, Hu S and Zhang Z, Lun J, Liao W and
Zhang Z: Expression of exosomal microRNAs during chondrogenic
differentiation of human bone mesenchymal stem cells. J Cell
Biochem. 120:171–181. 2019. View Article : Google Scholar
|
|
49
|
Mizuno Y, Tokuzawa Y, Ninomiya Y, Yagi K,
Yatsuka-Kanesaki Y, Suda T, Fukuda T, Katagiri T, Kondoh Y, Amemiya
T, et al: miR-210 promotes osteoblastic differentiation through
inhibition of AcvR1b. FEBS Lett. 583:2263–2268. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Schmid A, Sailland J, Novak L, Baumlin N,
Fregien N and Salathe M: Modulation of Wnt signaling is essential
for the differentiation of ciliated epithelial cells in human
airways. FEBS Lett. 591:3493–3506. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhang CY, Chen R, Wang F, Ren C, Zhang P,
Li Q, Li HH, Guo KT, Geng DQ and Liu CF: EGb-761 attenuates the
anti-proliferative activity of fluoride via DDK1 in PC-12 cells.
Neurochem Res. 42:606–614. 2017. View Article : Google Scholar
|
|
52
|
Westendorf JJ, Kahler RA and Schroeder TM:
Wnt signaling in osteoblasts and bone diseases. Gene. 341:19–39.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Li J, Sarosi I, Cattley RC, Pretorius J,
Asuncion F, Grisanti M, Morony S, Adamu S, Geng Z, Qiu W, et al:
Dkk1-mediated inhibition of Wnt signaling in bone results in
osteopenia. Bone. 39:754–766. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Xia ZL, Wang Y, Sun QD and Du XF: MiR-203
is involved in osteoporosis by regulating DKK1 and inhibiting
osteogenic differentiation of MSCs. Eur Rev Med Pharmacol Sci.
22:5098–5105. 2018.PubMed/NCBI
|
|
55
|
Paraskevopoulou MD and Hatzigeorgiou AG:
Analyzing MiRNA-LncRNA interactions. Methods Mol Biol.
1402:271–286. 2016. View Article : Google Scholar : PubMed/NCBI
|














