Introduction
As one of the most prevalent malignancies, breast
cancer is a primary cause of mortality among gynecological cancer
cases, and with increasing morbidity and mortality rates, it poses
a considerable threat to women's health worldwide (1,2).
In 2019, statistics from the American Cancer Society estimated
271,270 newly diagnosed cases and 42,260 deaths from breast cancer
in the United States (3). The
leading causes of the high death rate are distal metastasis and
resistance to the existing treatments (4). Despite improvements in early
diagnosis and systemic treatment, the incidence of breast cancer
and metastasis-related mortality is steadily increasing (5,6).
Therefore, there is an urgent need to elucidate the mechanisms
responsible for the disordered cellular metastasis and to enhance
our understanding of the tumorigenesis and development processes,
hence facilitating the identification of more efficient breast
cancer treatments.
Long noncoding RNAs (lncRNAs) are a group of RNAs
>200 nucleotides in length, which lack protein-coding capacity
(7). Numerous studies have
revealed that lncRNAs have versatile biological functions in
pathological and physiological processes, including tumorigenesis
(8-10). lncRNAs are considered to regulate
the development of various types of cancer, including breast cancer
(10). For instance, LINC01089 is
downregulated in breast cancer tissues and cell lines, and
LINC01089 overexpression increases tumor cell proliferation,
migration and invasiveness. As an oncogene that regulates breast
cancer cell proliferation and apoptosis, hepatocellular carcinoma
upregulated EZH2-associated lncRNA is closely associated with the
clinical progression of breast cancer (11). These results indicate the
indispensability of research on lncRNAs and breast cancer.
lncRNA forkhead box D2 adjacent apposite strand RNA
1 (FOXD2-AS1) is a novel non-coding RNA identified to be an
oncogene in human cancers. FOXD2-AS1 has been shown to be
upregulated in various types of cancer, including glioma,
osteosarcoma and papillary thyroid cancer, as well as breast cancer
(12-15). A previous study indicated that
FOXD2-AS1 participates in regulating the development of breast
cancer via the miR-150-5p/PFN2 axis, and that it may be a potential
biomarker for the diagnosis and prognosis of breast cancer
(15). However, to the best of
our knowledge, there are no additional data regarding the
investigation of FOXD2-AS1 in breast cancer, and its effects and
the underlying mechanisms on the regulation of breast cancer cell
invasion and metastasis. Thus, the aim of the present study was to
determine the role and potential mechanisms of action of FOXD2-AS1
in breast cancer, and to provide further support for its use in
clinical diagnosis and treatment.
Materials and methods
Datasets
The present study evaluated the expression level of
FOXD2-AS1 in breast cancer samples using The Cancer Genome Atlas
(TCGA) dataset, which was downloaded from the TCGA data portal
(https://tcga-data-nci-nih-gov.ez.xjtlu.edu.cn). The
TCGA data subset for breast cancer included 246 normal samples and
1,110 tumor samples. The Mann-Whitney test was used to determine
statistically significant differences between normal and tumor
samples. P<0.05 was considered to indicate a statistically
significant difference.
Cell culture
A human normal breast epithelial cell line (MCF-10A)
and human breast cancer cell lines (MCF-7, MDA-MB-468, MDA-MB-453
and BT-549) were purchased from the American Type Culture
Collection (ATCC). The cells were incubated in RPMI-1640 medium
supplemented with 10% fetal bovine serum (FBS) in a humidified
incubator at 37°C (5% CO2) (15,16).
Transfection
To overexpress FOXD2-AS1, an overexpression vector
(pcDNA FOXD2-AS1) and its corresponding negative control vector
(pcDNA-NC) were synthesized by Shanghai GenePharma Co., Ltd. Short
hairpin (sh)RNAs targeting FOXD2-AS1 (100 nM; shRNA-FOXD2-AS1-1 and
shRNA-FOXD2-AS1-2) and a negative scramble control shRNA (shRNA)
(also purchased from Shanghai GenePharma Co., Ltd.) were used to
knock down FOXD2-AS1 expression. In addition, pcDNA-LATS1,
shRNA-S100A1-1 and shRNA-S100A1-2 were obtained from Shanghai
GenePharma Co., Ltd. to overexpress LATS1 or to knock down S100A1,
respectively. The shRNA sequences were as follows:
shRNA-FOXD2-AS1-1 targets, GGA CTC CAC TCT TCG CTT A;
shRNA-FOXD2-AS1-2 targets, GCT TCC AGG TAT GTG GGA A;
shRNA-S100A1-1 targets, GAT CCG GAG ACC CTC ATC AAC GTG TTC TTC CTG
TCA GAA ACA CGT TGA TGA GGG TCT CCT TTT TG; shRNA-S100A1-2 targets,
GAT CCG TGG ACT TCC AGG AGT ATG TGC TTC CTG TCA GAC ACA TAC TCC TGG
AAG TCC ACT TTT TG. Cells were transfected with pcDNA FOXD2-AS1 (15
nM), pcDNA-LATS1 (15 nM), pcDNA-NC (15 nM), shRNA-FOXD2-AS1-1 (500
ng/µl), shRNA-FOXD2-AS1-2 (500 ng/µl), shRNA-S100A1-1
(500 ng/µl), shRNA-S100A1-2 (500 ng/µl), shRNA (500
ng/µl), or co-transfected with pcDNA FOXD2-AS1 and
pcDNA-LATS1, or co-transfected with pcDNA FOXD2-AS1 and
shRNA-S100A1 using Lipofectamine® 2000 transfection
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. Lipofectamine 2000 reagent was
first mixed with vectors to form a reagent-vector complex, followed
by incubation with cells at 37°C for 5 h. The transfection efficacy
was assessed by reverse transcription-quantitative PCR (RT-qPCR)
after 48 h of transfection.
RNA extraction and RT-qPCR
Total RNA was extracted from all cell lines using
TRIzol® reagent (Takara Bio, Inc.) according to the
manufacturer's instructions. Total RNA was then reverse transcribed
into cDNA using the PrimeScript™ RT Master kit, and the mRNA
expression levels were quantified using the SYBR Premix Ex Taq™ kit
(Both Takara Bio, Inc.) with a 7500 Fast Real-Time PCR System
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The sequences
of specific primers used for RT-qPCR were as follows: FOXDA-AS1
forward, 5′-TGG ACC TAG CTG CAG CTC CA-3′ and reverse, 5′-AGT TGA
AGG TGC ACA CAC TG-3′; S100A1 forward, 5′-GAG TAT GTG GTG CTT GTG
GC-3′ and reverse, 5′-CTT GGA CCG CTA CTC TTG CG-3′; large tumor
suppressor homolog 1 (LATS1) forward, 5′-ACC GCT TCA AAT GTG ACT
GTG ATG CCA C CT-3′ and reverse, 5′-CTT CCT TGG GCA AGC TTG GCT GAT
CCT CT-3′; and GAPDH forward, 5′-GCG AGA TCG CAC TCA TCA TCT -3′
and reverse, 5′-TCA GTG GTG GAC CTG ACC -3′. The data were
displayed as 2−ΔΔCq values with GAPDH as the
constitutive marker (17). The
PCR conditions were as follows: 95°C for 5 min, 40 cycles of 95°C
for 20 sec and 62°C for 30 sec, followed by 72°C for 3 min.
Cell Counting Kit-8 (CCK-8) assay
Cell proliferation was determined using the Cell
Counting Kit-8 assay (CCK-8; Dojindo Molecular Technologies, Inc).
Briefly, cells were seeded into a 96-well plate and incubated for
24 h at 37°C. Following culture for the indicated periods of time
(12, 24 and 48 h), 10 µl of the CCK-8 reagent were added to
each well and the plate was incubated at 37°C for a further 3 h.
The optical density values at 450 nm were then measured using a
micro-plate spectrophotometer (Thermo Fisher Scientific, Inc.).
Western blot analysis
The total protein was extracted from the cells using
RIPA lysis buffer (Beyotime Institute of Biotechnology) and
quantified using a BCA protein assay kit (Thermo Fisher Scientific,
Inc.). The same amount of each protein sample (20 µg) was
subjected to 10% SDS-PAGE; the proteins were then transferred onto
PVDF membranes (EMD Millipore) and was blocked in 5% non-fat milk
for 1 h at room temperature. The membranes were then incu-bated
with primary antibodies against cyclinE1 (1:1,000; cat. no.
ab33911; Abcam), cyclin-dependent kinase-2 (CDK2; 1:1,000; cat. no.
ab32147; Abcam), p21 (1:1,000; cat. no. ab109520; Abcam), matrix
metalloproteinases (MMP)2 (1:1,000; cat. no. ab92536; Abcam), MMP9
(1:1,000; cat. no. ab38898; Abcam), S100 calcium binding protein A1
(S100A1; 1:1,000; cat. no. 5066; Cell Signaling Technology, Inc.),
phosphorylated (p)-yes-associated protein (YAP; 1:1,000; cat. no.
13008; Cell Signaling Technology, Inc.), YAP (1:1,000; cat. no.
15028; Cell Signaling Technology, Inc.), serine/threonine-protein
kinase LATS1 (1:1,000; cat. no. 3477; Cell Signaling Technology,
Inc.), p-LATS1 (1:1,000; cat. no. 8654; Cell Signaling Technology,
Inc.), mammalian STE20-like protein kinase (MST)1 (1:1,000; cat.
no. 3682; Cell Signaling Technology, Inc.), MST2 (1:1,000; cat. no.
3952; Cell Signaling Technology, Inc.), and GAPDH (1:2,000; cat.
no. ab8245; Abcam) at 4°C overnight. The membranes were washed with
Tris-buffered saline with Tween (TBST) and incubated with
horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG
secondary antibody (1:2,000; cat. no. sc-2005; Santa Cruz
Biotechnology, Inc.) at room temperature for 1.5 h. The protein
bands were visual-ized using an enhanced chemiluminescence kit
(Amersham Pharmacia Biotech) and quantified using ImageJ software
(version 1.46; National Institutes of Health).
Wound healing assay
The cellular migration rate was determined using a
wound healing assay. The cells were seeded into a 6-well plate and
cultured to 100% confluence. A wound was produced in each monolayer
using a 200-µl pipette tip, and the plate was washed 3 times
with PBS to remove detached cells. The cells were then cultured in
the fresh medium without FBS. Following incubation for 48 h, the
wound-healing ability was assessed under a light microscope
(magnification ×100; CKX41, Olympus Corporation), and the widths of
the wounds were measured at 0 and 48 h.
Transwell assay
The cell invasive rate was determined with a
Transwell assay. Cells (4×104/well) in serum-free medium
were placed in the upper chamber of each insert [which had been
precoated with 40 µl of Matrigel (BD biosciences) at 37°C
for 1 h], and complete medium containing 10% FBS was added to the
lower 24-well chamber. After 24 h, cells on the upper surface were
removed, and cells attached to the lower surface were stained with
0.05% crystal violet (Beijing Solarbio Science & Technology
Co., Ltd.) at room temperature for 10 min. The cells were viewed
under a light microscope (magnification ×100; CKX41, Olympus
Corporation), and the invasive ability of the cells was determined
using ImageJ software version 1.46 by counting the number of cells
attached to the lower surface.
Cell cycle analysis
The cell cycle distribution was determined by flow
cytometry. After being subjected to the indicated treatments, the
cells were collected and fixed in 70% ethanol at −20°C overnight.
The cells were then washed twice with PBS and incubated in the dark
with RNase A and PI staining solution (Roche Diagnostics) at 37°C
for 30 min. Finally, the cell samples were analyzed using a
FACSCalibur flow cytometer and CellQuest software version 5.1 (both
from BD Biosciences).
In vivo experiments
The present study was approved by the First
Affiliated Hospital of Zhengzhou University, and the animal
experiments were performed according to the National Institute of
Health Guidelines for the Care and Use of Laboratory Animals. A
total of 18 5-week-old male BALB/c nude mice were purchased from
the Experimental Animal Center of Shanghai Institute for Biological
Sciences, and housed in a standard environment (25°C; 50% humidity;
12 h light/dark cycle) with free access to food and water. Each
mouse was subcutaneously injected at the right axillary lymph nodes
with 1.0×107 MDA-MB-468 cells, which were stably
transfected with either the shRNA negative control shRNA or
shRNA-FOXD2-AS1. The weights and tumor volumes (tumor volume =1/2 ×
length × width2) of the mice were monitored every 5 days
until the mice were sacrificed. At 20 days after the injection, all
the 18 mice were sacrificed by cervical dislocation that caused a
sharp section of the spinal cord followed by an instantaneous
cardiac arrest. After the cessation of the heartbeat and
respiratory arrest of the mice was confirmed, the tumors were
excised, photographed and stored for the further investigation.
Statistical analysis
Data are presented as the means ± standard deviation
(SD) from ≥3 independent experiments, and each experiment was
conducted in triplicate. The data were analyzed using SPSS
statistical software version 20.0 (SPSS, Inc.) and the differences
among groups were analyzed using one-way analysis of variance
followed by Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
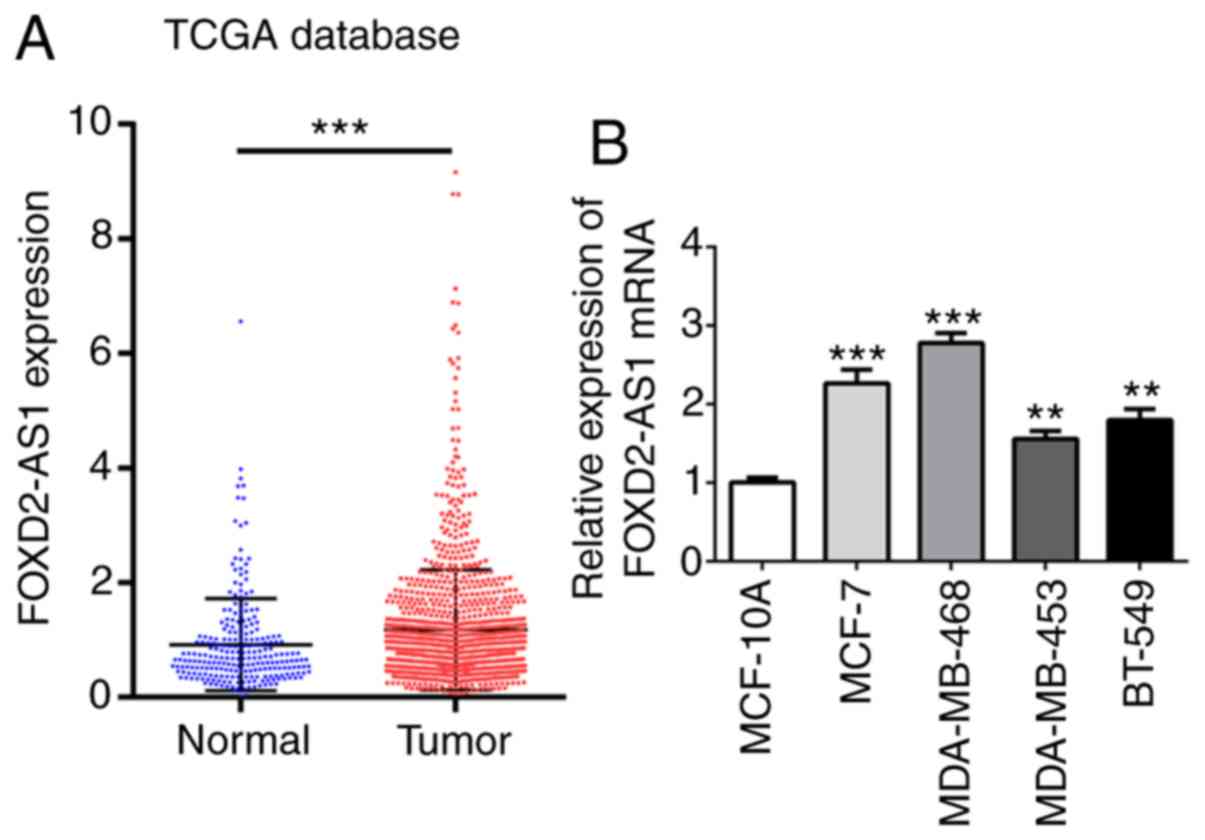
FOXD2-AS1 expression is upregulated in
breast cancer cells
The TCGA database (cancer.gov/tcga) was used to identify the association
between FOXD2-AS1 and breast cancer by evaluating the expression
profiles of FOXD2-AS1 in breast cancer tissues and normal tissues.
The results of TCGA analysis revealed a significantly higher
FOXD2-AS1 expression in breast cancer tissues than in normal
tissues (Fig. 1A). Human normal
breast epithelial cell line (MCF-10A) and human breast cancer cell
lines (MCF-7, MDA-MB-468, MDA-MB-453 and BT-549) were also obtained
to detect the mRNA levels of FOXD2-AS1. The results revealed that
FOXD2-AS1 expression was markedly upregulated in all breast cancer
cells, particularly in the MCF-7 (ER-positive breast cancer cell
line) and MDA-MB-468 cells (triple-negative breast cancer cell
line) (Fig. 1B), which were used
for further experiments, even though there may be some variability
in the results between the 2 cell lines. These findings indicate
that FOXD2-AS1 is upregulated in breast cancer.
FOXD2-AS1 knockdown suppresses breast
cancer cell proliferation, migration and invasiveness
To further elucidate the role of FOXD2-AS1 in breast
cancer, FOXD2-AS1 was knocked down in both the MCF-7 and MDA-MB-468
cells. Due to a higher transfection efficacy, shRNA-FOXD2-AS1-1
(referred to as shRNA-FOXD2-AS1) was subsequently used for breast
cancer cell experimentation (Fig. 2A
and B). The results of CCK-8 assay indicated that FOXD2-AS1
knockdown significantly inhibited the proliferative ability of the
MCF-7 and MDA-MB-468 cells (Fig. 2C
and D). The cell cycle was then analyzed by flow cytometry,
which revealed that FOXD2-AS1 knockdown increased the percentage of
cells in the G1 phase, whereas it decreased that in the S phase for
both the MCF-7 and MDA-MB-468 cells (Fig. 2E-G). Furthermore, FOXD2-AS1
knockdown decreased the protein expression levels of cyclin E1 and
CDK2, and increased the expression of p21 (Fig. 2H and I). These findings indicate
that FOXD2-AS1 knockdown suppresses cellular proliferation by
regulating the cell cycle, specifically by preventing G1 to S phase
progression. Moreover, FOXD2-AS1 knockdown significantly decreased
the migration rate (Fig. 2J-L)
and the invasiveness (Fig. 2M-O)
of the MCF-7 and MDA-MB-468 cells. The expression levels of MMP-2
and -9, which are critical to the migration, invasion and
metastasis of breast cancer cells (18), were both downregulated following
FOXD2-AS1 knockdown (Fig. 2P and
Q), indicating that FOXD2-AS1 may enhance cellular migration
and invasiveness by regulating MMP-2 and -9.
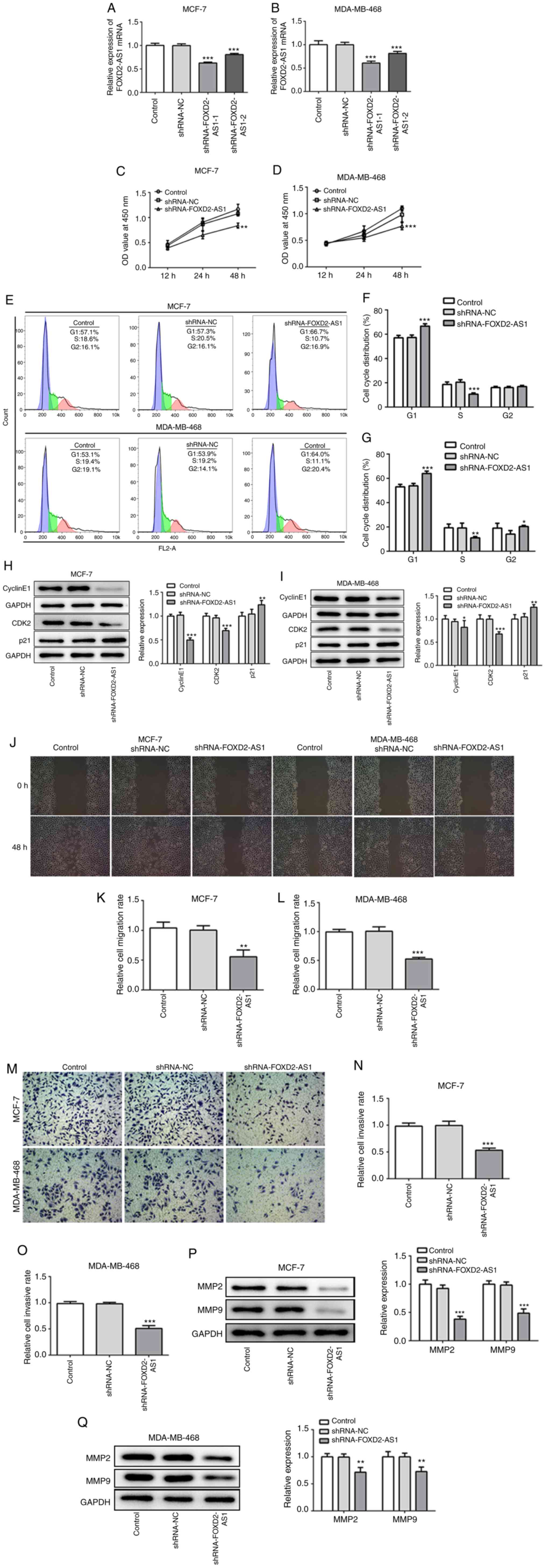 | Figure 2Knockdown of FOXD2-AS1 suppresses
cell proliferation, migration and invasion in breast cancer. (A and
B) Following transfection with shRNA-FOXD2-AS, the mRNA level of
FOXD2-AS1 was measured by RT-qPCR in MCF-7 and MDA-MB-468 cells. (C
and D) CCK-8 assay was performed to determine cell proliferation
following transfection. (E-G) Cell cycle distribution was
determined and analyzed by FACS. (H and I) The protein expression
of cyclin E1, CDK2 and p21 was determined by western blot analysis.
Knockdown of FOXD2-AS1 suppresses cell proliferation, migration and
invasion in breast cancer. (J) Wound healing assay was performed to
detect the migration of both MCF-7 and MDA-MB-468 cells. (K and L)
Relative migration rate of MCF-7 and MDA-MB-468 cells was
quantified, respectively. (M) Transwell assay was performed to
detect the invasion of both MCF-7 and MDA-MB-468 cells. (N and O)
Relative cell invasive rate of MCF-7 and MDA-MB-468 cells was
quantified, respectively. (P) MMP-2 and MMP-9 protein expression in
MCF-7 transfected with shRNA-FOXD2-AS1 was detected and quantified.
(Q) MMP-2 and MMP-9 protein expression in MDA-MB-468 transfected
with shRNA-FOXD2-AS1 was detected and quantified.
*P<0.05, **P<0.01 and ***P<0.001,
vs. shRNA-NC. CDK2, cyclin-dependent kinase-2; MMP, matrix
metalloproteinase. |
FOXD2-AS1 knockdown regulates the
Hippo/YAP signaling pathway
The Hippo/YAP signaling pathway is reportedly
involved in the progression of breast cancer (19). In the present study, the levels of
specific proteins involved in the YAP/Hippo signaling pathway were
assessed in the MCF-7 and MDA-MB-468 cells following FOXD2-AS1
knockdown. Western blot analysis revealed that the levels of p-YAP
and p-LATS1 were significantly upregulated, while those of MST1 and
2 were significantly downregulated by FOXD2-AS1 knockdown (Fig. 3A and B). Thus, the results
confirmed that FOXD2-AS1 regulates the Hippo/YAP signaling pathway
in breast cancer cells.
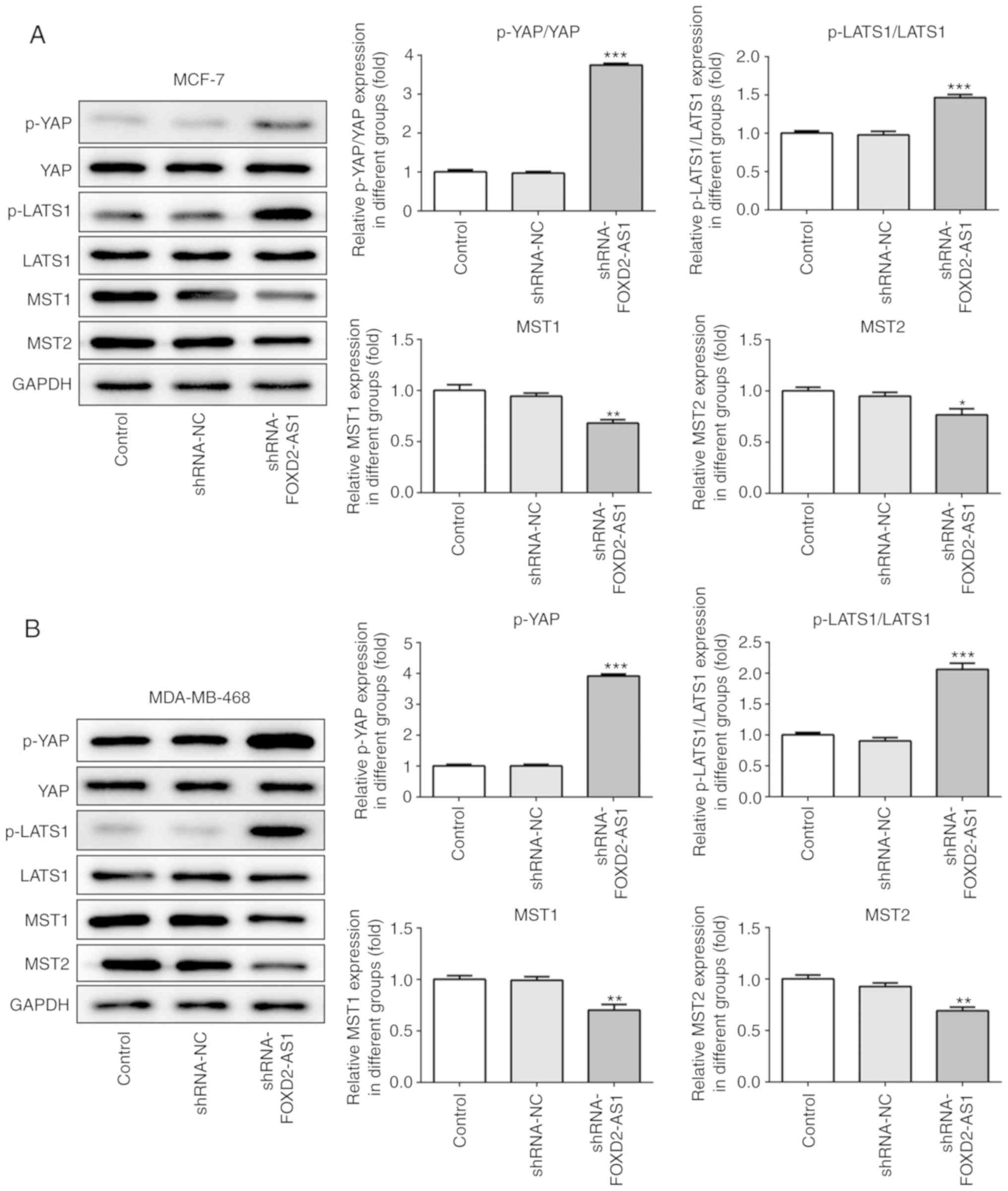 | Figure 3Knockdown of FOXD2-AS1 regulates the
Hippo/YAP signaling pathway. (A) The protein expression of
Hippo/YAP signaling pathway-related genes (p-YAP, YAP, p-LATS1,
LATS1, MST1 and MST2) was determined by western blot analysis in
MCF-7 cells. (B) The protein expression of Hippo/YAP signaling
pathway-related genes (p-YAP, YAP, p-LATS1, LATS1, MST1 and MST2)
was determined by western blot analysis in MDA-MB-468 cells.
*P<0.05, **P<0.01 and
***P<0.001, vs. shRNA-NC. LATS1, large tumor
suppressor homolog 1; MST, mammalian STE20-like protein kinase;
YAP, yes-associated protein. |
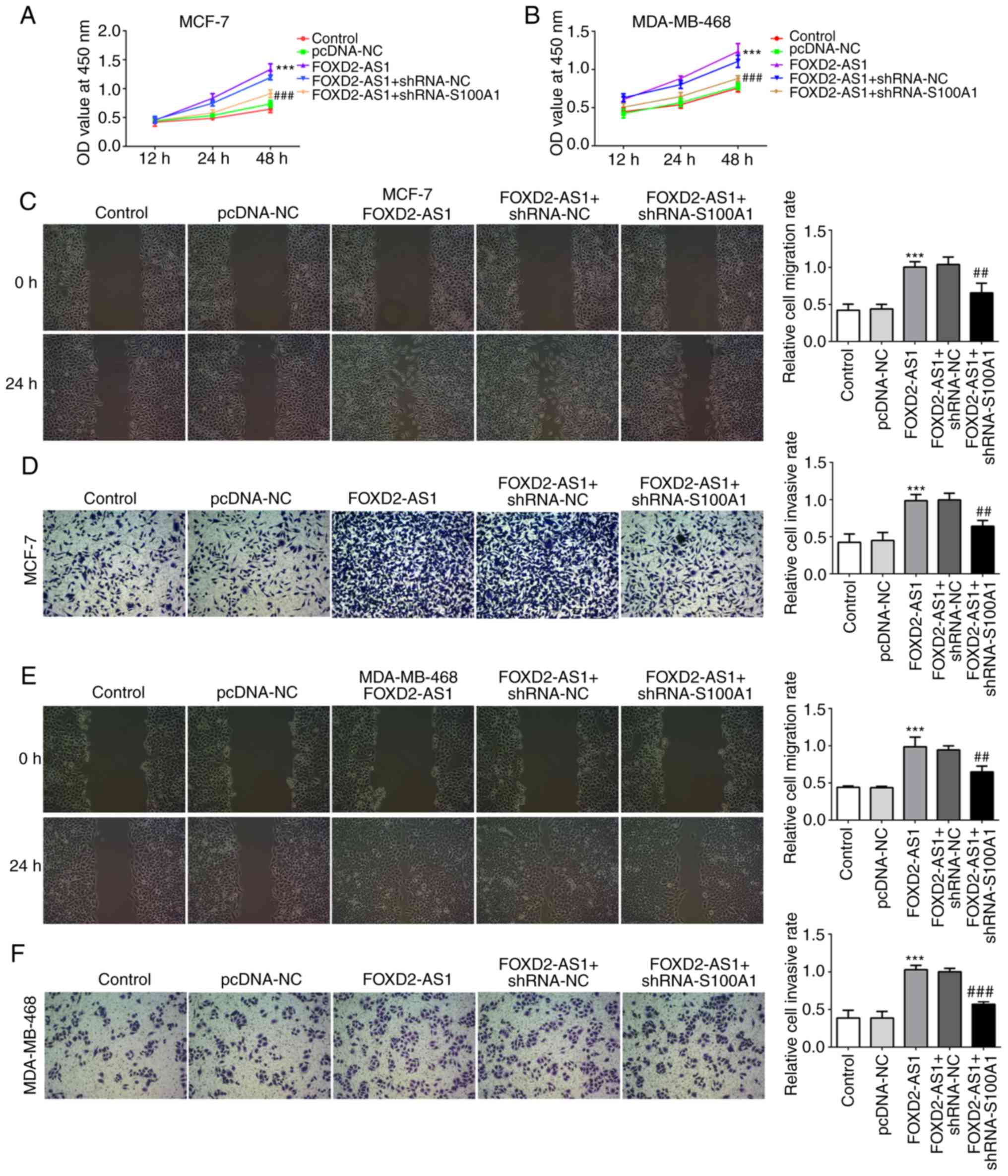
S100A1 mediates FOXD2-AS1-induced breast
cancer cell proliferation, migration and invasiveness
S100A1 is a calcium-binding protein of the S100
protein family, which is not only upregulated in, but is also
involved in the progression of ovarian cancer (20). In the present study, the
expression of S100A1 was evaluated in the breast cancer cell lines,
indicating that S100A1 was significantly upregulated in breast
cancer cells (particularly in the MCF-7 and MDA-MB-468 cells)
compared with the MCF-10A cells (Fig.
4A). In MCF-7 and MDA-MB-468 cells transfected with
shRNA-FOXD2-AS1, it was found that the protein expression level of
S100A1 was downregulated (Fig. 4B and
C). To further investigate the role of S100A1 in
FOXD2-AS1-mediated cellular proliferation, migration and
invasiveness, breast cancer cells were transfected with an
expression vector, pcDNA-FOXD2-AS1 (Fig. 4D and E), and shRNA-S100A1 to
inhibit S100A1 protein and mRNA expression (Fig. 4F-I). As shown in Fig. 5A and B, the overexpression of
FOXD2-AS1 significantly promoted the proliferation of MCF-7 and
MDA-MB-468 cells, which was subsequently reversed by the
downregulation of S100A1. Wound healing and Transwell assays
demonstrated that FOXD2-AS1 overexpression significantly increased
MCF-7 cell migration and invasiveness, respectively, which were
also reversed by the downregulation of S100A1 (Fig. 5C and D). A similar result was
observed in the MDA-MB-468 cells (Fig. 5E and F). These results suggest
that S100A1 regulates the FOXD2-AS1-mediated proliferation,
migration and invasiveness of breast cancer cells.
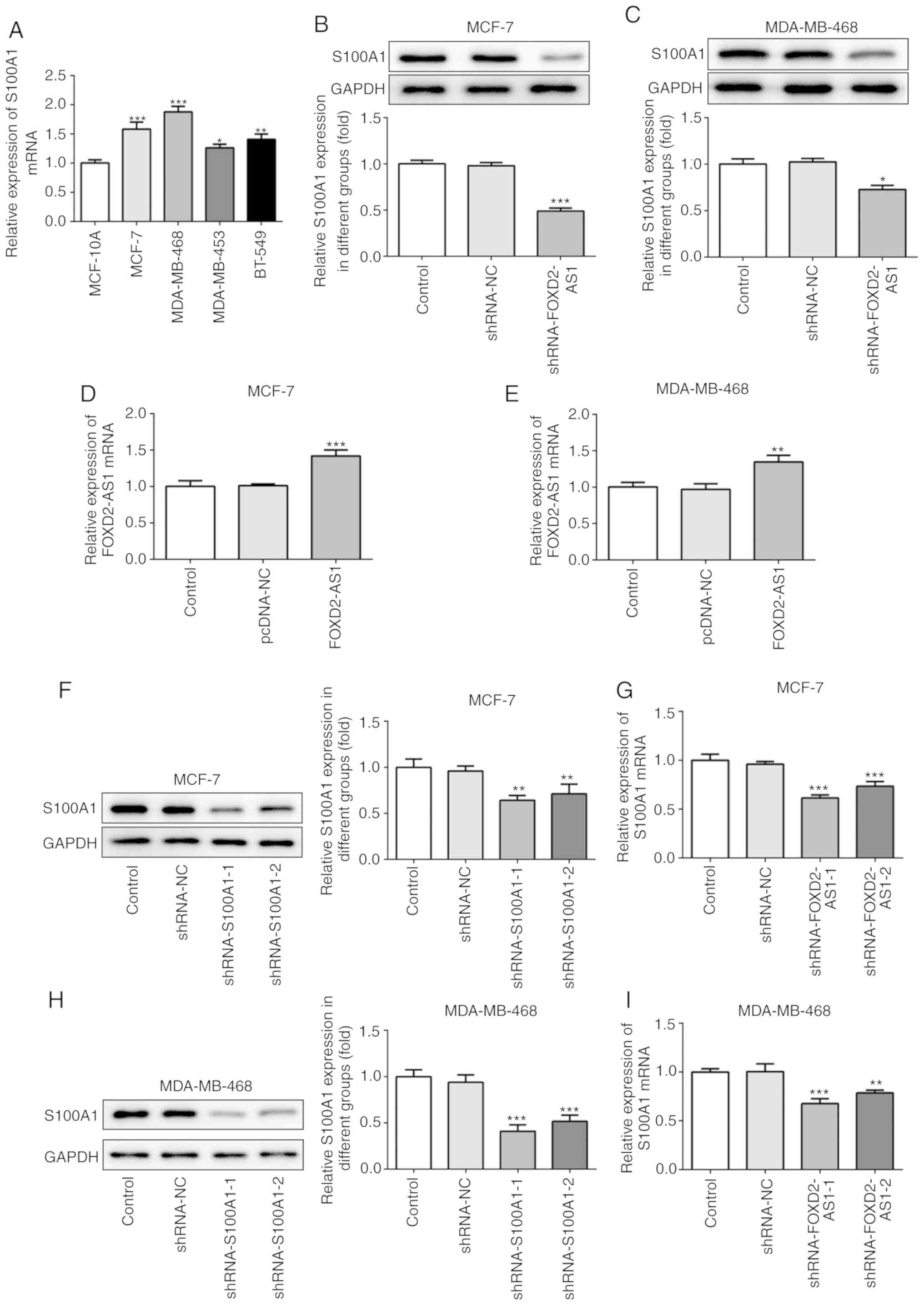 | Figure 4S100A1 is upregulated in breast
cancer cells. (A) The mRNA level of S100A1 in MCF-10A and human
breast cancer cell lines (MCF-7, MDA-MB-468, MDA-MB-453 and BT-549)
was determined by RT-qPCR. *P<0.05,
**P<0.01 and ***P<0.001, vs. MCF-10A
cells. (B and C) In shRNA-FOXD2-AS1-trans-fected MCF-7 and
MDA-MB-468 cells, the protein expression of Sl00A1 was detected by
western blot analysis. *P<0.05 and
***P<0.001, vs. shRNA-NC. (D and E) Following
transfection with pcDNA-FOXD2-AS1, the mRNA level of FOXD2-AS1 in
MCF-7 and MDA-MB-468 cells was detected by RT-qPCR.
**P<0.01 and ***P<0.001, vs. pcDNA-NC.
MCF-7 and MDA-MB-468 cells were transfected with shRNA-S100A1, and
the protein expression and mRNA level of S100A1 in (F and G) MCF-7
and (H and I) MDA-MB-468 cells were determined by western blot
analysis and RT-qPCR, respectively. **P<0.01 and
***P<0.001, vs. shRNA-NC. S100A1, S100 calcium
binding protein A1. |
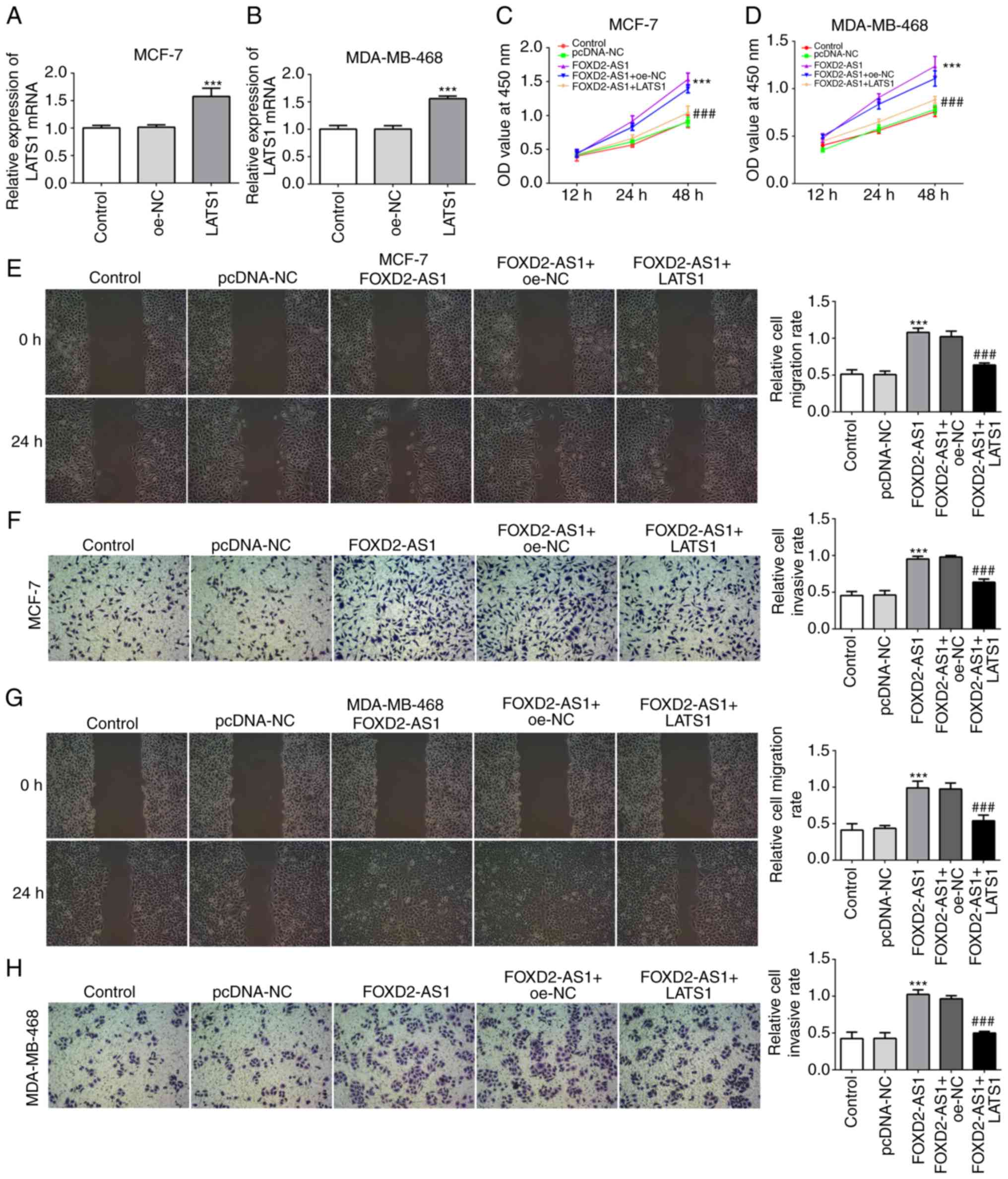
Overexpression of LATS1 inhibits
FOXD2-AS-induced cell proliferation, migration and
invasiveness
Since the Hippo/YAP signaling pathway is involved in
the FOXD2-AS1-mediated characteristics of breast cancer cells,
Hippo/YAP signaling was further investigated for its regulatory
role in breast cancer cell proliferation, migration and
invasiveness. For this purpose, LATS1 was overexpressed in the
MCF-7 and MDA-MB-468 cells (Fig. 6A
and B), and the results of CCK-8 assay revealed that LATS1
overexpression significantly inhibited FOXD2-AS1-induced cellular
proliferation (Fig. 6C and D).
Furthermore, the results of wound healing and Transwell assays
revealed that LATS1 overexpression significantly inhibited the
FOXD2-AS1-induced migration and invasiveness of both the MCF-7
(Fig. 6E and F) and MDA-MB-468
cells (Fig. 6G and H).
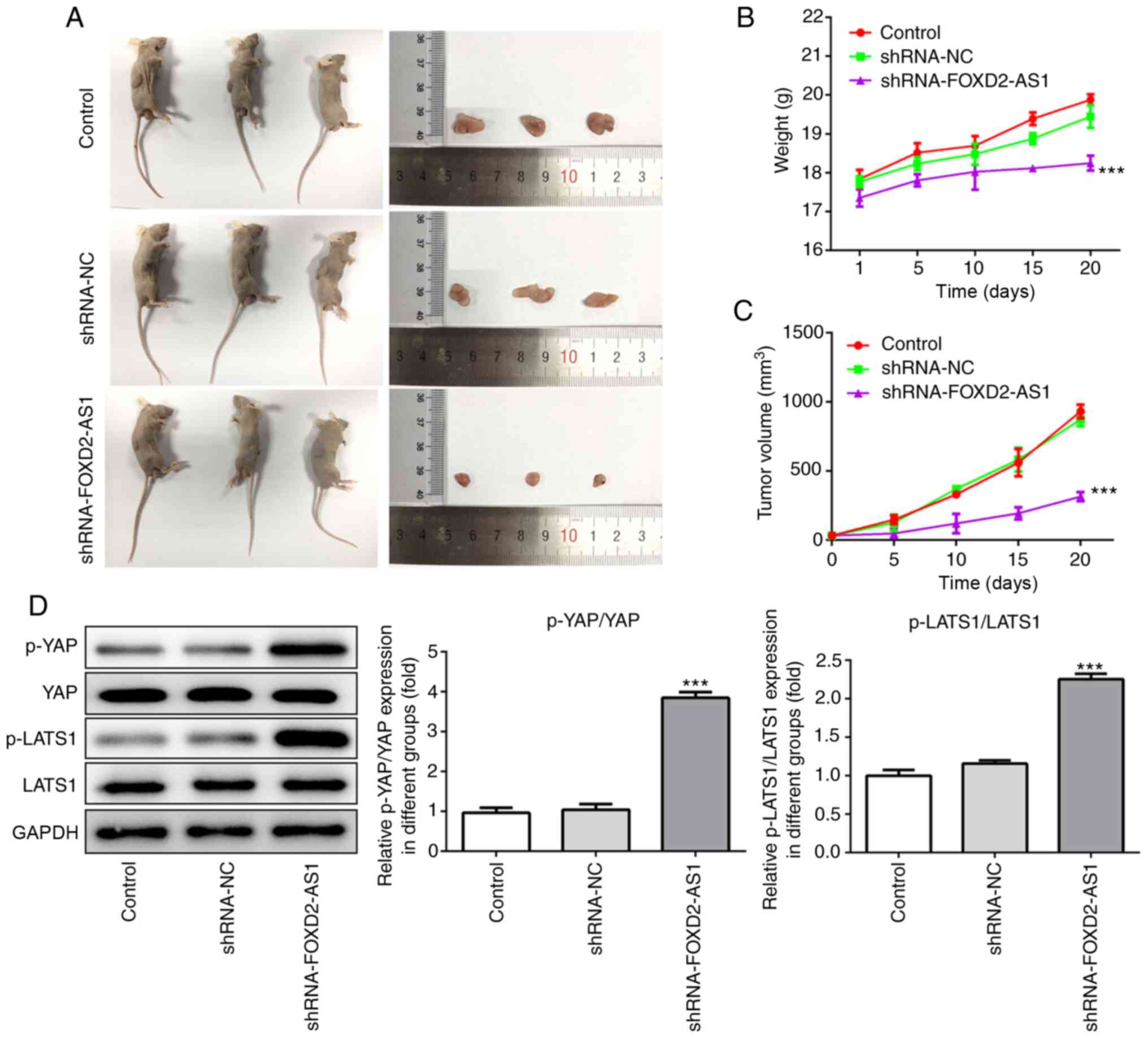
FOXD2-AS1 knockdown suppresses breast
cancer tumor progression in vivo
From the aforementioned results, the role of
FOXD2-AS1 in both the MCF-7 and MDA-MB-468 cells was confirmed. To
explore the role of FOXD2-AS1 in breast cancer in vivo, mice
were injected with MDA-MB-468 cells, which were stably transfected
with either an shRNA negative control or shRNA-FOXD2-AS1. Following
sacrifice, the tumors were excised and photographed (Fig. 7A). Tumors in the shRNA-FOXD2-AS1
group were the smallest in size, directly reflecting the
suppressive effect of FOXD2-AS1 knockdown on tumor growth. During
the experiment, body weights and tumor volumes were recorded every
5 days. Body weight increased at a slower rate in the
shRNA-FOXD2-AS1 group, and the tumor volumes of this group also
increased at a slower rate than those in the other groups (Fig. 7B and C). Additionally, western
blot analysis of the extracted tumor tissues indicated a
significant increase in p-YAP and p-LATS1 expression in the
shRNA-FOXD2-AS1 group (Fig. 7D),
which was consistent with the in vitro results. These
findings thus suggest that FOXD2-AS1 knockdown suppresses the
progression of breast cancer and regulates the Hippo/YAP signaling
pathway in vivo.
Discussion
Breast cancer is one of the most common types of
human tumors, particularly among females. The high rates of
metastasis and recurrence typically result in the deterioration and
death of patients with breast cancer (21). Increasing evidence suggests that
lncRNAs function as oncogenic or antitumor genes in various tumor
types, cells types and the microenvironment, and that they may be
used as effective and specific biomarkers for clinical diagnosis
and prognosis (22,23). Due to its oncogenic properties,
lncRNA FOXD2-AS1 has been investigated in several malignant tumors.
In the present study, FOXD2-AS1 expression was found to be
upregulated in breast cancer cell lines; thus, it was knocked down
in MCF-7 and MDA-MB-468 cell to investigate its role in breast
cancer. FOXD2-AS1 knockdown inhibited the proliferation, migration
and invasiveness of MCF-7 and MDA-MB-468 cells, and inhibited tumor
growth in vivo. Notably, S100A1 expression was also found to
be upregulated in breast cancer cells, and further investigation
revealed that S100A1 was inhibited following FOXD2-AS1 knockdown,
indicating that the expression of FOXD2-AS1 and S100A1 was
positively associated in breast cancer cells. Subsequent
experiments revealed that the overexpression of FOXD2-AS1
significantly accelerated tumorigenesis by promoting cellular
proliferation, migration and invasiveness. However, the effects of
FOXD2-AS1 were reversed by the downregulation of S100A1. These
results suggest that both FOXD2-AS1 and S100A1 knockdown exert
antitumor effects on the progression of breast cancer, and that
FOXD2-AS1 may exert its oncogenic functions by regulating
S100A1.
S100A1 is a calcium-binding protein belonging to the
S100 protein family, which exhibit a range of biological properties
surrounding cellular proliferation, metastasis, immune evasion and
angiogenesis, and are also involved in tumorigenesis (20,24). For example, S100A4 enhances
p53-dependent apoptosis and facilitates more aggressive tumor
progression (25). S100A6 has
been reported to be upregulated in human osteosarcoma, colorectal
carcinoma and hepatocellular carcinoma, which was mostly associated
with its suppressive properties towards cancer cell migration and
tumor metastasis (26,27). Therefore, S100 proteins play an
important role in the development and progression of tumors,
highlighting the necessity to further understand their roles and
potential underlying mechanisms. In the present study, S100A1
expression was found to be upregulated in breast cancer cell lines.
FOXD2-AS1 overexpression was shown to accelerate breast cancer
progression by promoting cellular proliferation, migration and
invasiveness, and functional experiments demonstrated that the
knockdown of S100A1 reversed the effects induced by FOXD2-AS1.
Furthermore, S100A1 knock-down suppressed breast cancer progression
by inhibiting the proliferation, migration and invasiveness of
MCF-7 and MDA-MB-468 cells. In agreement with these findings,
S100A1 has been reported to be overexpressed in ovarian cancer, and
to be associated with lymph mode metastasis; the overexpression of
S100A1 was shown to enhance cellular proliferation and migration,
whilst its inhibition exerted an opposite effect on ovarian cancer
cells (20). Moreover, high tumor
expression levels of S100A1 have been shown to be positively
associated with decreased relapse-free survival time in an
endometrioid subtype of ovarian and endometrial cancers (28). It was thus hypothesized that
S100A1 functions as an important regulator in breast cancer, and
may therefore be a promising therapeutic target for this, as well
as other types of gynecological cancer.
The Hippo pathway is an important signaling pathway
that regulates cellular proliferation and apoptosis, the activation
of which is triggered by the phosphorylation of the large tumor
suppressor kinases, LATS1 and LATS2. The Hippo pathway is very
complex as a number of kinases relay upstream signals to LATS to
regulate this pathway. The STE20 protein kinases (MST1/2), as the
core components of the Hippo pathway, are considered responsible
for the phosphorylation and activation of LATS1/2 (29). YAP, a downstream effector of the
Hippo pathway, is highly activated in various types of cancer, and
targeting YAP may effectively suppress tumorigenesis (30,31). Both dysregulated Hippo signaling
and aberrant YAP activation contribute to cancer progression
(32,33). In the present study, FOXD2-AS1
knockdown significantly increased the phosphorylation of YAP and
LATS1, indicating that the Hippo signaling pathway was activated by
FOXD2-AS1 downregulation. Notably, it was found that MST1/2
expression levels were downregulated by FOXD2-AS1 downregulation,
which was contradictory with the activated Hippo pathway. There is
evidence to indicate that although MST1/2 are firmly established as
the initiating kinases of the Hippo kinase cascade in mammals, it
has been observed that MST1/2 are not absolutely required for LATS
and YAP regulation by numerous upstream signals. For example,
MST1/2 is not involved in YAP regulation in response to other
signals such as cAMP (34).
Moreover, the knockdown of MST1/2 was previously show to not affect
basal YAP phosphorylation in HeLa cells (35). Furthermore, MST1/2 has found to be
largely dispensable for YAP regulation, whereas MAPK4s, also
considered as Hippo pathway components, exert direct effects on
LATS1/2 and YAP phosphorylation and activation (36). Therefore, there is no simple
one-to-one linear association in MST1/2 and LATS activation, which
may provide a reasonable explanation for the results of the present
study. The present results revealed that Hippo/YAP may be involved
in the promoting effects of FOXD2-AS1 on cell proliferation,
migration and invasiveness. The Hippo/YAP signaling pathway is also
associated with the regulation of tumor growth in vivo, and
FOXD2-AS1 downregulation was demonstrated to suppress tumor growth
in breast cancer-bearing mice by regulating Hippo/YAP signaling
pathway.
Furthermore, existing evidence suggests that S100A1
exerts its oncogenic effects by interacting with LATS1 and
activating YAP. In view of the positive association between p-LATS1
and S100A1 in clinical samples of hepatocellular carcinoma, LATS1
was considered to be responsible for S100A1-induced cancer
progression (37). In the present
study, a positive association was observed between FOXD2-AS1 and
S100A1 expression, and the knockdown of S100A1 significantly
inhibited FOXD2-AS1-induced cellular proliferation, migration and
invasion. The overexpression of LATS1 also inhibited
FOXD2-AS1-induced cellular activity, suggesting that FOXD2-AS1 is
likely to exert its effects by interacting with S100A1 and the
LATS1-induced Hippo signaling pathway. Given that MST1/2 was not
involved in the activation of LATS1 in the present study, LATS may
be activated by other signals; thus, it was hypothesized that
S100A1 may be the upstream protein involved in directly activating
LATS1. Considering all of the above, the p-LATS1-induced activation
of Hippo/YAP signaling is partly dependent on the level of S100A1,
which is regulated by FOXD2-AS1.
However, some limitations exist in the present
study. First, although it was hypothesized that S100A1 was
responsible for the activation of Hippo-YAP signaling, whether
Sl00A1 was indispensable for LATS activation or whether its
mutation directly affected Hippo-YAP signaling in breast cancer was
not elucidated. Secondly, the mechanisms through which FOXD2-AS1
regulates S100A1 were not investigated in the present study.
Increasing evidence has revealed that lncRNAs contribute to
tumorigenesis by silencing tumor suppressors or activating
oncogenes by acting as competing endogenous RNAs (ceRNAs) to sponge
miRNAs. In previous studies, FOXD2-AS1 has been reported to
regulate cancer progression by sponging various miRNAs, including
miR-143, miR-7-5p and miR-185, thus modulating the suppression of
mRNAs (38-40). In particular, Chen et al
demonstrated that the oncogenic role of FOXD2-AS1 in nasopharyngeal
carcinoma was mediated largely in part by sponging miR-363-5p, and
subsequently activating S100A1, as S100A1 was confirmed to be a
direct target of miR-363-5p in its 3′-UTR mRNA (41). Therefore, it is possible that
FOXD2-AS1 may regulate S100A1 by sponging miR-363-5p in breast
cancer, thus activating S100A1-induced LATS1 and YAP activation.
FOXD2-AS1 may exert its oncogenic effects on breast cancer through
the miR-363-5p/S100A1/Hippo pathway; this hypothesis warrants
further investigations in the future.
In conclusion, the findings of the present study
demonstrate that FOXD2-AS1 is crucial for cellular proliferation,
migration and invasiveness, as well as tumor growth in breast
cancer. FOXD2-AS1 regulates malignancy by regulating the Hippo/YAP
signaling pathway, which is further mediated by the interaction
between S100A1 and p-LATS1. The present study suggests that the
FOXD2-AS1/S100A1/Hippo axis is involved in the tumorigenesis and
progression of breast cancer, which may contribute to the future
development of more effective treatments for breast cancer.
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PH made substantial contributions to the conception
and design of the study. PH and JX performed data acquisition, data
analysis and interpretation. PH drafted the manuscript and
critically revised it for important intellectual content. Both
authors gave final approval for the published version of the study
and agree to be accountable for all aspects of the work in ensuring
that questions related to the accuracy or integrity of the work are
appropriately investigated and resolved. Both authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the First
Affiliated Hospital of Zhengzhou University, and the animal
experiments were performed according to the National Institute of
Health Guidelines for the Care and Use of Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
DeSantis CE, Ma J, Goding Sauer A, Newman
LA and Jemal A: Breast cancer statistics, 2017 racial disparity in
mortality by state. CA Cancer J Clin. 67:439–448. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Global Burden of Disease Cancer
Collaboration; Fitzmaurice C, Allen C, Barber RM, Barregard L,
Bhutta ZA, Brenner H, Dicker DJ, Chimed-Orchir O, Dandona R, et al:
Global, regional, and national cancer incidence, mortality, years
of life lost, years Lived with disability, and disability-adjusted
life-years for 32 cancer groups, 1990 to 2015: A systematic
analysis for the global burden of disease study. JAMA Oncol.
3:524–548. 2017. View Article : Google Scholar
|
|
3
|
DeSantis CE, Miller KD, Goding Sauer A,
Jemal A and Siegel RL: Cancer statistics for African Americans,
2019. CA Cancer J Clin. 69:211–233. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gonzalez-Angulo AM, Morales-Vasquez F and
Hortobagyi GN: Overview of resistance to systemic therapy in
patients with breast cancer. Adv Exp Med Biol. 608:1–22. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schwartz RS and Erban JK: Timing of
metastasis in breast cancer. N Engl J Med. 376:2486–2488. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Beermann J, Piccoli MT, Viereck J and Thum
T: Non-coding RNAs in development and disease: Background,
mechanisms, and therapeutic approaches. Physiol Rev. 96:1297–1325.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yuan H, Qin Y, Zeng B, Feng Y, Li Y, Xiang
T and Ren G: Long noncoding RNA LINC01089 predicts clinical
prognosis and inhibits cell proliferation and invasion through the
Wnt/β-catenin signaling pathway in breast cancer. Onco Targets
Ther. 12:4883–4895. 2019. View Article : Google Scholar :
|
|
9
|
Hua X, Li G, Liu Z and Niu Z: LINK-A
lncRNA participates in the pathogenesis of glioma by interacting
with survivin. Exp Ther Med. 18:1581–1586. 2019.PubMed/NCBI
|
|
10
|
Liu Y, Sharma S and Watabe K: Roles of
lncRNA in breast cancer. Front Biosci (Schol Ed). 7:94–108. 2015.
View Article : Google Scholar
|
|
11
|
Li P, Zhou B, Lv Y and Qian Q: LncRNA HEIH
regulates cell proliferation and apoptosis through miR-4458/SOCS1
axis in triple-negative breast cancer. Hum Cell. 32:522–528. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang J, Li B, Wang C, Luo Y, Zhao M and
Chen P: Long non-coding RNA FOXD2-AS1 promotes glioma cell cycle
progression and proliferation through the FOXD2-AS1/miR-31/CDK1
pathway. J Cell Biochem. 120:19784–19795. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ren Z, Hu Y, Li G, Kang Y, Liu Y and Zhao
H: HIF-1α induced long noncoding RNA FOXD2-AS1 promotes the
osteosarcoma through repressing p21. Biomed Pharmacother.
117:1091042019. View Article : Google Scholar
|
|
14
|
Zhang Y, Hu J, Zhou W and Gao H: LncRNA
FOXD2-AS1 accelerates the papillary thyroid cancer progression
through regulating the miR-485-5p/KLK7 axis. J Cell Biochem. Nov
19–2018. View Article : Google Scholar : Online ahead of
print.
|
|
15
|
Jiang M, Qiu N, Xia H, Liang H, Li H and
Ao X: Long noncoding RNA FOXD2AS1/miR1505p/PFN2 axis regulates
breast cancer malignancy and tumorigenesis. Int J Oncol.
54:1043–1052. 2019.PubMed/NCBI
|
|
16
|
Tseng LM, Chiu JH, Liu CY, Tsai YF, Wang
YL, Yang CW and Shyr YM: A comparison of the molecular subtypes of
triple-negative breast cancer among non-Asian and Taiwanese women.
Breast Cancer Res Treat. 163:241–254. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
18
|
Zhou R, Xu L, Ye M, Liao M, Du H and Chen
H: Formononetin inhibits migration and invasion of MDA-MB-231 and
4T1 breast cancer cells by suppressing MMP-2 and MMP-9 through
PI3K/AKT signaling pathways. Horm Metab Res. 46:753–760. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang X, Liu X, Luo J, Xiao W, Ye X, Chen
M, Li Y and Zhang GJ: Notch3 inhibits epithelial-mesenchymal
transition by activating Kibra-mediated Hippo/YAP signaling in
breast cancer epithelial cells. Oncogenesis. 5:e2692016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tian T, Li X, Hua Z, Ma J, Liu Z, Chen H
and Cui Z: S100A1 promotes cell proliferation and migration and is
associated with lymph node metastasis in ovarian cancer. Discov
Med. 23:235–245. 2017.PubMed/NCBI
|
|
21
|
Wang T: Association between HIF1α 1772 C/T
polymorphism and breast cancer susceptibility: A systematic review.
Crit Rev Eukaryot Gene Expr. 27:297–304. 2017. View Article : Google Scholar
|
|
22
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou S, Wang J and Zhang Z: An emerging
understanding of long noncoding RNAs in kidney cancer. J Cancer Res
Clin Oncol. 140:1989–1995. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang T, Huo X, Chong Z, Khan H, Liu R and
Wang T: A review of S100 protein family in lung cancer. Clin Chim
Acta. 476:54–59. 2018. View Article : Google Scholar
|
|
25
|
Grigorian M, Andresen S, Tulchinsky E,
Kriajevska M, Carlberg C, Kruse C, Cohn M, Ambartsumian N,
Christensen A, Selivanova G and Lukanidin E: Tumor suppressor p53
protein is a new target for the metastasis-associated Mts1/S100A4
protein: Functional consequences of their interaction. J Biol Chem.
276:22699–22708. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Luu HH, Zhou L, Haydon RC, Deyrup AT,
Montag AG, Huo D, Heck R, Heizmann CW, Peabody TD, Simon MA and He
TC: Increased expression of S100A6 is associated with decreased
metastasis and inhibition of cell migration and anchorage
independent growth in human osteosarcoma. Cancer Lett. 229:135–148.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Melle C, Ernst G, Schimmel B, Bleul A and
von Eggeling F: Colon-derived liver metastasis, colorectal
carcinoma, and hepatocellular carcinoma can be discriminated by the
Ca(2+)-binding proteins S100A6 and S100A11. PLoS One. 3:e37672008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
DeRycke MS, Andersen JD, Harrington KM,
Pambuccian SE, Kalloger SE, Boylan KL, Argenta PA and Skubitz AP:
S100A1 expression in ovarian and endometrial endometrioid
carcinomas is a prognostic indicator of relapse-free survival. Am J
Clin Pathol. 132:846–856. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chan EH, Nousiainen M, Chalamalasetty RB,
Schafer A, Nigg EA and Sillje HH: The Ste20-like kinase Mst2
activates the human large tumor suppressor kinase Lats1. Oncogene.
24:2076–2086. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kim W, Khan SK, Liu Y, Xu R, Park O, He Y,
Cha B, Gao B and Yang Y: Hepatic Hippo signaling inhibits
protumoural microenvironment to suppress hepatocellular carcinoma.
Gut. 67:1692–1703. 2018. View Article : Google Scholar
|
|
31
|
Zhou Y, Jin Q, Xiao W and Sun C:
Tankyrase1 antisense oligodeoxynucleotides suppress the
proliferation, migration and invasion through Hippo/YAP pathway in
human osteosarcoma cells. Pathol Res Pract. 215:1523812019.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ehmer U and Sage J: Control of
Proliferation and Cancer Growth by the Hippo Signaling Pathway. Mol
Cancer Res. 14:127–140. 2016. View Article : Google Scholar :
|
|
33
|
Han H, Yang B, Nakaoka HJ, Yang J, Zhao Y,
Le Nguyen K, Bishara AT, Mandalia TK and Wang W: Hippo signaling
dysfunction induces cancer cell addiction to YAP. Oncogene.
37:6414–6424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yu FX, Zhang Y, Park HW, Jewell JL, Chen
Q, Deng Y, Pan D, Taylor SS, Lai ZC and Guan KL: Protein kinase A
activates the Hippo pathway to modulate cell proliferation and
differentiation. Genes Dev. 27:1223–1232. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao B, Li L, Wang L, Wang CY, Yu J and
Guan KL: Cell detachment activates the Hippo pathway via
cytoskeleton reorganization to induce anoikis. Genes Dev. 26:54–68.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Meng Z, Moroishi T, Mottier-Pavie V,
Plouffe SW, Hansen CG, Hong AW, Park HW, Mo JS, Lu W, Lu S, et al:
MAP4K family kinases act in parallel to MST1/2 to activate LATS1/2
in the Hippo pathway. Nat Commun. 6:83572015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Guo Q, Wang J, Cao Z, Tang Y, Feng C and
Huang F: Interaction of S100A1 with LATS1 promotes cell growth
through regulation of the Hippo pathway in hepatocellular
carcinoma. Int J Oncol. 53:592–602. 2018.PubMed/NCBI
|
|
38
|
Dong H, Cao W and Xue J: Long noncoding
FOXD2-AS1 is activated by CREB1 and promotes cell proliferation and
metastasis in glioma by sponging miR-185 through targeting AKT1.
Biochem Biophys Res Commun. 508:1074–1081. 2019. View Article : Google Scholar
|
|
39
|
Liu X, Fu Q, Li S, Liang N, Li F, Li C,
Sui C, Dionigi G and Sun H: LncRNA FOXD2-AS1 functions as a
competing endogenous RNA to regulate TERT expression by sponging
miR-7-5p in thyroid cancer. Front Endocrinol (Lausanne).
10:2072019. View Article : Google Scholar
|
|
40
|
An Q, Zhou L and Xu N: Long noncoding RNA
FOXD2-AS1 accelerates the gemcitabine-resistance of bladder cancer
by sponging miR-143. Biomed Pharmacother. 103:415–420. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen G, Sun W, Hua X, Zeng W and Yang L:
Long non-coding RNA FOXD2-AS1 aggravates nasopharyngeal carcinoma
carcinogenesis by modulating miR-363-5p/S100A1 pathway. Gene.
645:76–84. 2018. View Article : Google Scholar
|