1. Introduction
Cell senescence refers to the stable cell-cycle
arrest caused by telomere erosion, DNA damage, the abnormal
activation of oncogenes and other conditions, which is accompanied
by morphological, biochemical and epigenetic changes to the cells.
Cell senescence plays important roles in non-disease-related
physiological processes, including embryo development, wound
healing, tissue repair and aging (1). A large number of previously
published studies have demonstrated that cell senescence plays an
important role in the inhibition of potential cancer cell
proliferation pathways (2-4).
However, as the detailed scientific knowledge in this field
continues to expand, it has been demonstrated that senescent cells
also secrete a variety of cytokines, chemokines, matrix-remodeling
proteases and growth factors to form the senescence-associated
secretory phenotype (SASP), which is a 'double-edged sword' in that
it may either inhibit or promote tumor formation (5). Therefore, establishing an in-depth
understanding of the association between cell senescence and tumors
that focuses on the inhibitory function of cell senescence in
tumors, rather than on its promoting function, may provide more
options in terms of tumor therapies.
2. Characteristics of cell senescence
related with tumors
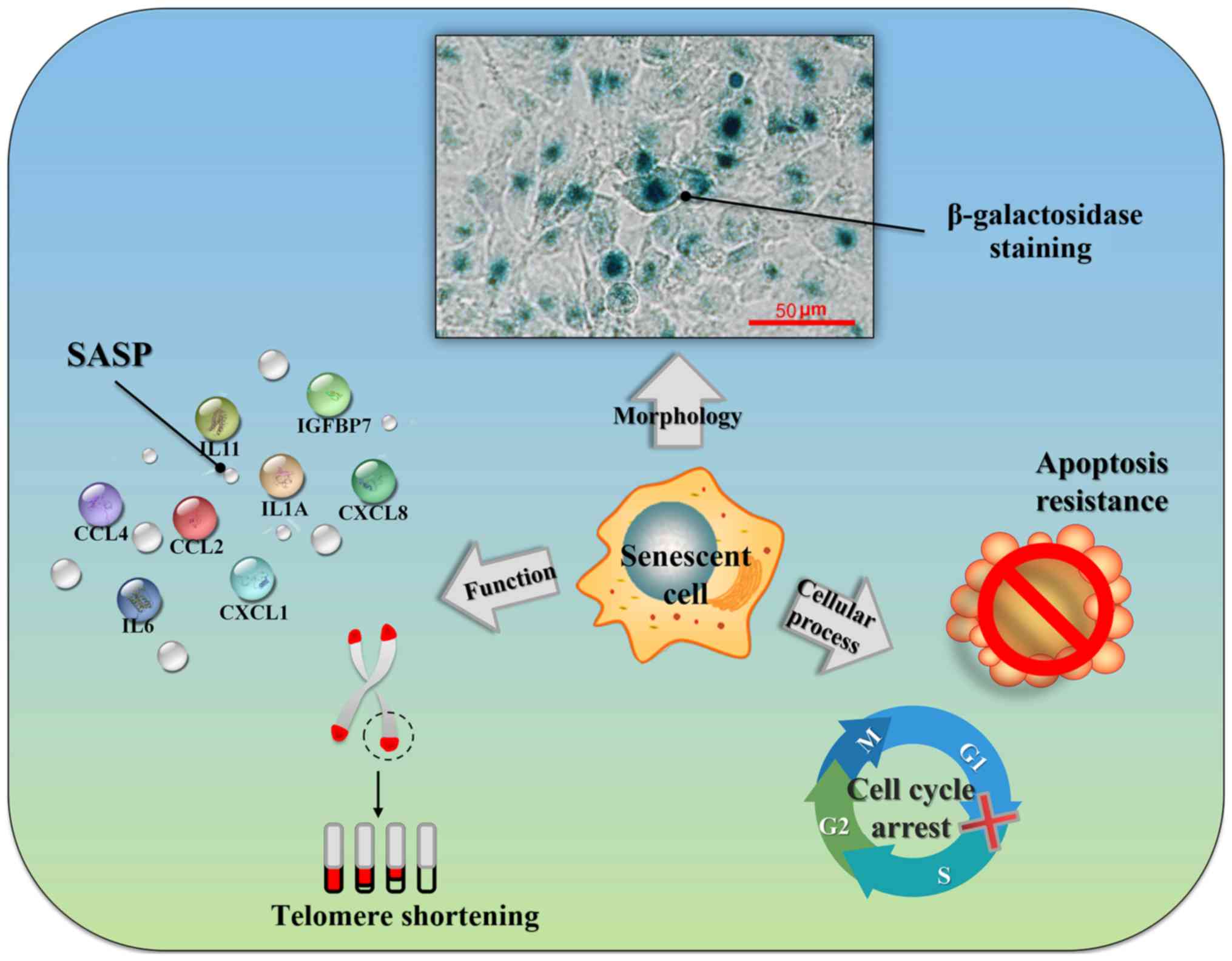
When cells become senescent, they undergo changes in
cellular morphology, function and cellular process (Fig. 1).
Cellular morphological changes
After entering the aging phase, the senescent cells
may be observed as polygonal-shaped flat cells occupying a large
volume, with increased nuclear and nucleolar volumes, as observed
under an optical microscope. Moreover, the numbers of Golgi
apparatus and lysosomes in senescent cells are increased, along
with a marked increase in intracytoplasmic particles (6). In 1995, Dimri et al (7) discovered that when the pH value was
6 in diploid fibroblasts cultured in vitro, the positive
rate of β-galactosidase staining gradually increased with a rise in
generation age. They defined this neutral galactosidase as
SA-β-gal. When cells become senescent, lysosomes expand and
increase, and galactosidase clearly accumulates within lysosomes
(8). Galactosidase is produced by
senescent cells and tissues, and is able to catalyze the hydrolysis
of its substrate, X-Gal, to produce an insoluble, intensely dark
blue product, which can be easily observed under an optical
microscope. The SA-β-gal is a type of biological marker that can be
simple and easy to use, it has been widely applied in the detection
of senescent cells.
Functional alterations
After the cells have entered the aging process,
their protein expression profiles are markedly altered. Most
importantly, senescent cells will secrete a large number of
cytokines and chemokines in this regard. The concept of SASP was
first proposed by Coppé et al (9) in 2008. These researchers discovered
that senescent cells are able to promote malignancy in adjacent
precancerous cells by secreting inflammatory and
oncogene-associated factors, and the SASP includes pro-inflammatory
cytokines [interleukin (IL)-6 and IL-8], chemokines [monocyte
chemoattractant proteins (MCP) and macrophage inflammatory proteins
(MIPs)], growth factors [transforming growth factors (TGFs) and
granulocyte-macrophage colony-stimulating factor (GM-CSF)] and
proteases (10). Different
biological activities induced by SASP components indicate that SASP
functions with other cells and regulates the local microenvironment
of tissues, which may either lead to or promote tumor
formation.
When telomeres decay to a critical extent, cells
begin a spontaneous aging process (11). Telomeres were first identified by
Muller in 1938 (12). He found
that the damaged and broken chromosome ends in Drosophila
were easily connected, forming various types of chromosome
aberrations. Telomeres, a special structure located at the
chromosome ends, maintain the stability of chromosome ends by
forming protective 'caps', thereby preventing them from being
exposed and fusing with adjacent chromosome ends. However, since
DNA polymerase may not completely replicate the lagging strands of
chromosomes, telomere DNA repeat sequences are continuously
shortened according to the increase in replication times. This
irreversible telomere DNA damage leads to a damage response that
may accelerate cell aging by activating tumor suppressor genes,
such as p53, and triggering the expression of age-associated
genes.
Cellular process alterations
Unlike resting cells, the growth stagnation of
senescent cells is relatively permanent. Even when subjected to
physiological stimulation, or after eliminating factors that induce
aging, senescent cells cannot regain the ability to divide
(13). Flow cytometric analysis
has been employed to demonstrated that senescent cells are mainly
held in a state of G1 phase arrest, although they may
also be found in G2/M phase arrest, or can be induced
into a state of G2/M phase arrest in certain cases
(14). In addition, in certain
types of cell, low-level DNA damage activates DNA damage repair,
triggers cell senescence, causes the expression of certain
anti-apoptosis proteins, and leads to the inhibition of apoptosis
(15). A number of studies have
confirmed that the occurrence of aging is closely related to
several oncogenes and tumor suppressor genes (16-18).
3. Cell senescence has important functions
in tumors
Senescence initiation in cancer is able to rely upon
genetic alterations, such as oncogene-induced senescence (OIS) and
tumor-suppressor gene (TSG) loss-induced senescence, or upon
therapeutic interventions [therapy-induced senescence (TIS)]. OIS
generally occurs early in tumorigenesis, and is able to limit cell
proliferation due to oncogene mutations, maintaining tumors in a
non-invasive precancerous state. By examining lesions associated
with OIS, new patterns of tumorigenesis have been proposed
(19,20). When cells have cancer gene
mutations, they are generally prevented from being able to
proliferate through the processes of apoptosis and senescence. When
the apoptotic or senescence program cannot be initiated, and other
genetic mutations occur at the same time, the cells experience
continuous growth and eventually enter into the state of malignancy
due to the lack of effective defense mechanisms. Senescent cells
remain senescent for decades, and are presented as benign lesions.
It is certainly possible that other genetic mutations may also be
acquired, and the cells are subsequently transformed into malignant
tumors. Other studies have found that certain aging markers are
only present in precancerous lesions, but are rarely expressed in
corresponding malignancies. Using a mouse prostate model in which
PTEN tumor suppressor was inactivated, Chen et al (21) observed similar results, i.e.,
precancerous lesions or non-lethal tumors were able to express
senescence markers, whereas malignant tumors were not. In cultured
cells, as well as in vivo, p53 restricts the growth and
malignant evolution of PTEN-deficient cells by inducing cellular
senescence. This is similar to the situation with human prostate
cancer, in that PTEN loss-induced cellular senescence (PICS)
appears during the early stage, but is not expressed in the
advanced malignant stages (22,23). Patients with cancer often receive
radiotherapy and chemotherapy as a means of treating the tumor. A
previous study demonstrated that the chemoradiotherapy of tumors
induced cellular senescence (24). Common chemoradiotherapeutic
treatments include cisplatin treatment, docetaxel treatment,
cyclophosphamide treatment, vincristine treatment, doxorubicin
treatment or gamma-ray irradiation. Notably, when lower drug
concentrations are used for chemotherapy or lower radiation
intensities are used for radiotherapy, such chemoradiotherapeutic
treatments can induce cell senescence. Conversely, if higher
chemotherapeutic drug concentrations or radiation intensities are
used, apoptosis can be induced (25). The main cause of cellular
senescence induced by chemoradiotherapy is the induction of
intracellular DNA damage; for example, in cultured cells and mouse
models of cancer, tumor cells expressing wild-type p53 gene are
more sensitive to chemotherapy than tumor cells with a p53 gene
mutation are, findings that are consistent with p53 playing a key
role in DNA damage-induced cellular senescence (26). Other explanations underlying
cellular senescence induced by chemoradiotherapy are that
chemotherapeutic drugs cause the production of intracellular
reactive oxygen species (ROS), or that the chemoradiotherapy leads
to the inhibition of telomerase activity in the cells; these
processes are also involved in the induction of cellular
senescence, and may accelerate the cellular senescence process
(17). After the body receives
chemoradiotherapy, even though the tumor cells undergo senescence,
exit the cell cycle, and lose the ability to proliferate, these
cells are potentially tumorigenic, and after the tumor cells enter
the state of senescence, the SASP phenomenon may occur.
The role of SASP in vivo is closely
associated with the microenvironment where it is located. The
secretion of SASP is a dynamic process, and SASP factors can
inhibit tumor development by altering the tissue microenvironment,
promoting embryonic development and tissue repair, and recruiting
and activating immune cells. On the other hand, as senescent cells
accumulate in organisms, SASP drives tissue senescence in a
cell-involuntary manner, thereby inducing body inflammation through
autocrine and paracrine pathways, and transmitting senescence
signals to adjacent cells, resulting in increased levels of
inflammatory factors. The inflammatory environment can promote the
development of tumors by inhibiting immune surveillance and/or
stimulating malignant phenotypes (27). SASP is secreted into the
surrounding environment, where it blocks the differentiation of
cells, prevents the renewal of damaged cells, interferes with the
rejuvenation of tissues, further accelerates aging and promotes
tumor migration, proliferation, invasion and angiogenesis,
ultimately leading to tumor metastasis. Tumor extracellular matrix
proteins are involved in the occurrence, development and immune
adaptation of tumors, and senescent fibroblasts can regulate the
secretion of integrins mediated by surrounding cells, thereby
affecting extracellular matrix proteins. Integrins can enhance both
contractility and the focal adhesions produced by ERK and Rho
kinase (ROCK) activation, which interfere with epithelial
morphology and promote the malignant transformation of tumors
(28). MMP members often exhibit
an increasing trend in SASP factors secreted by senescent
fibroblasts, whereas during tumor development, MMP mainly degrades
the extracellular matrix and destroys the basement membrane, which,
in turn, promotes the infiltration and metastasis of tumor cells
(29). Acosta and Gil (30) demonstrated that IL-8, as one of
the SASP factors secreted by senescent tumor cells, is an effective
agonist of chemokine (C-X-C motif) ligand 1 (CXCL1)/CXCL-2
receptors in granulocytes and monocytes, which can mediate the
migration of target cells to the tumor microenvironment and promote
tumor angiogenesis and tumor cell proliferation. Ortiz-Montero
et al (31) demonstrated
that IL-6 was a major regulator of the Janus kinase/signal
transducer and activator of transcription (JAK/STAT) signaling
pathway, which promoted tumor cell proliferation, invasion and
immunosuppression and induce fibroblast morphological changes in
MCF-7 breast cancer cells.
Recently, it has been reported that
senescence-inducing factors can promote tumor formation. Carrière
et al (32) demonstrated
that the oncogene Kras was able to induce an aging response,
upregulating proinflammatory cytokines in the pancreas and
increasing the incidence of cystic tumors, while accelerating the
process of pancreatic ductal adenocarcinoma. Malaquin et al
(33) suggested that skin
carcinoma may be activated through a paracrine axis in which matrix
metalloproteinases (MMPs) are secured via the oversynthesis of the
MMP-active receptor, protease-activated receptor-1. Chen et
al (34) demonstrated that
stromal cell populations, after having suffered DNA damage via
radiotherapy and chemotherapy, entered the aging stage and
presented SASP, leading to the formation of an inflammatory
microenvironment. Their clinical investigations have indicated that
the high expression of serine proteinase inhibitor Kazal Type 1
(SPINK1) in the tumor microenvironment is associated with a worse
clinical prognosis and a shorter the survival time of patients.
Zhang et al (35) observed
that certain drugs, such as vinblastine, taxanes, nitrogen mustard,
nucleoside analogues and platinum compounds, which directly or
indirectly cause DNA damage, may stimulate the SASP in cells at a
high frequency, while triggering cell senescence. They also found
that Zscan4 which is highly expressed in plasma cells that suffer
from DNA damage, plays a key role in the cascade amplification of
SASP over time. Nacarelli et al (36) found that an increased metabolism
of NAD+ promoted the proliferation and progression of
tumor cells in pancreatic cancer and ovarian cancer in a mouse
model by manipulating the expression levels of high-mobility group
AT-hook 1 (HMGA1) protein and nicotinamide
phosphoribosyltransferase (NAMPT). Correspondingly, this
demonstrated that the increase in NAD+ metabolism caused
by the increase in HMGA1 and NAMPT expression promoted high levels
of SASP. Simultaneously, it strengthened the inflammatory
environment around the tumor, and stimulated tumor growth. Xu et
al (37) demonstrated that
human stromal cells passively entered the aging state during
chemotherapy, where they developed the typical SASP. Stromal cells
generated and released an exocrine factor termed AREG
(amphiregulin, bidirectional modulin), which continuously enters
damaged microenvironments. AREG promotes the malignant phenotype of
residual cancer cells in lesions, thereby conferring drug
resistance and causing resistance in clinical care. However, it
also induces cancer cells to upregulate the expression of
programmed death-ligand 1, forming an immunosuppressive
microenvironment, which enables cancer cells to escape from immune
monitoring. A recent study stated that SASP counteracted
atorvastatin-induced senescence in hepatocellular carcinoma cells,
and also found that the poor overall survival of patients with
hepatocellular carcinoma was associated with an increased hTERT
expression (38). Miyazoe et
al (39) reported that
senescent human hepatic stellate cells released increased
quantities of extracellular vesicles particles compared with normal
human hepatic stellate cells. Extracellular vesicles resulted in
increased EGF expression levels and may create a more conducive
tumor microenvironment for proliferation of hepatoma cells
(Table I).
 | Table ISenescence-inducing factors as
targets for the promotion of tumor formation. |
Table I
Senescence-inducing factors as
targets for the promotion of tumor formation.
| Category | Inducement | Target gene | Tumor
histology | Year, (Refs.) |
|---|
| Oncogenes
activated | Kras | p53 | Pancreatic
cancer | 2011, (32) |
| Telomere
shortening | 50-60 population
doublings | PAR-1 | Skin cancer | 2013, (33) |
| DNA damage | Radiotherapy,
chemotherapy | SPINK1 | Prostate
cancer | 2018, (34) |
| DNA damage | Chemotherapy | Zscan4 | Breast cancer,
prostate cancer | 2018, (35) |
| Oncogenes
activated | Ras |
NAD+ | Pancreatic cancer,
ovarian cancer | 2019, (36) |
| DNA damage | Chemotherapy | AREG | Prostate
cancer | 2019, (37) |
| DNA damage | Chemotherapy | hTERT | Hepatocellular
carcinoma | 2020, (38) |
| DNA damage | Extracellular
vesicles | EGF | Hepatocellular
carcinoma | 2020, (39) |
4. Targeting senescent cells to inhibit
tumor formation
Aging is a double-edged sword. When senescent cells
are retained, they promote chronic inflammation, which in turn
leads to aging-associated diseases, such as atherosclerosis, cancer
and fibrosis. Therefore, the targeted elimination of senescent
cells has become a promising new avenue for therapeutic
interventions, and researchers have confirmed the feasibility of
this approach. Currently, regulating SASP, activating apoptosis and
immune-based elimination of senescent cells are the three most
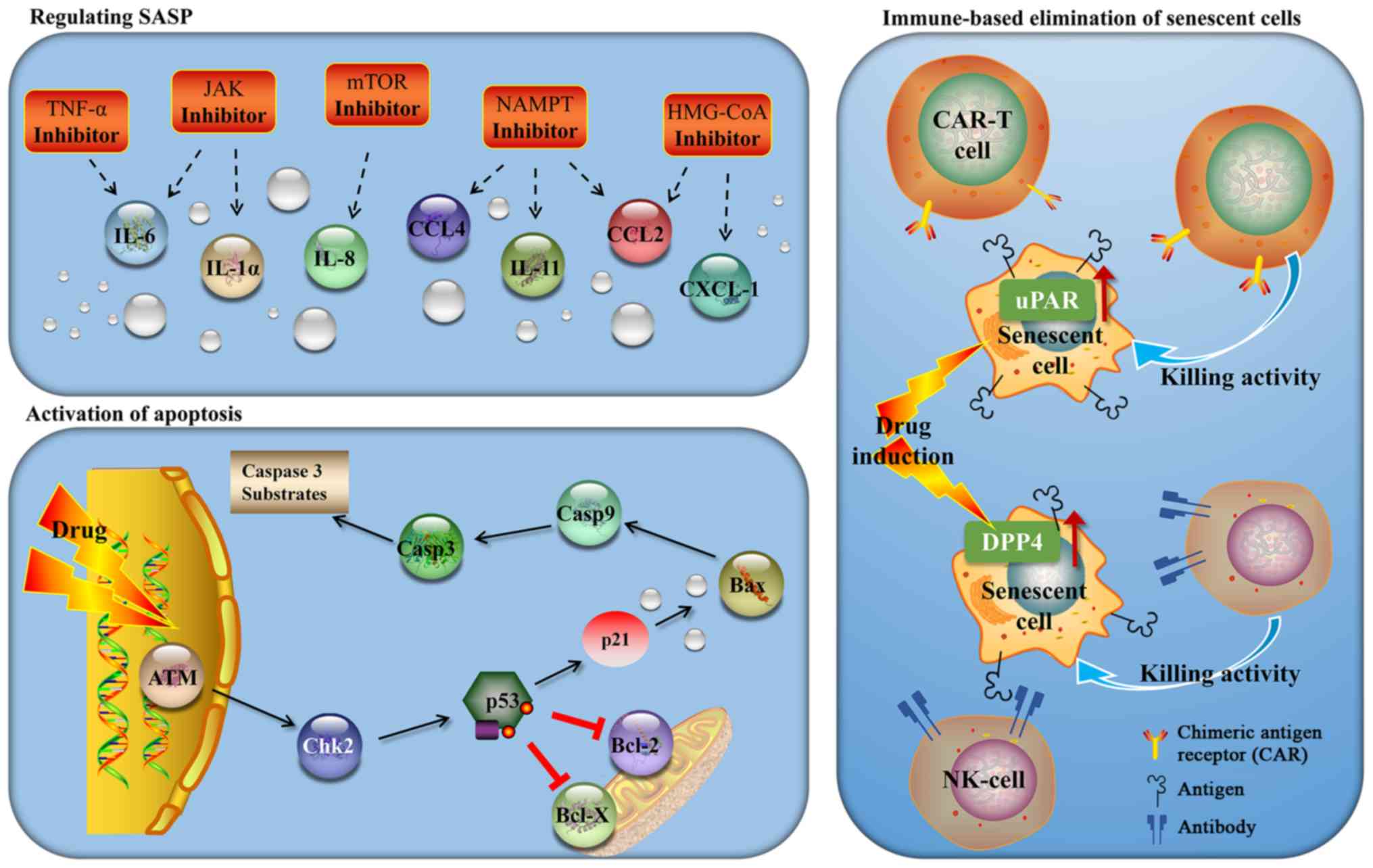
important targeting strategies (Fig.
2).
Regulating SASP
Tumor necrosis factor (TNF) is the main component of
SASP, and adalimumab is a monoclonal antibody that directly
inhibits TNF. It has previously been demonstrated that adalimumab
can inhibit the secretion of SASP and markedly reduce the release
of IL-6, attenuating the cancer-promoting effect of SASP (40). Xu et al (41) demonstrated that JAK inhibitors
reduce the secretion of SASP in adipocyte precursor cells and human
umbilical vein endothelial cells (HUVECs), suggesting that the JAK
pathway is a potential target for anti-senescence-associated
treatments. Mammalian target of rapamycin (mTOR) controls SASP by
regulating the translation of IL-1 and MAPK-activated protein
kinase 2 (MAPKAPK2) (42,43). Rapamycin, an inhibitor of mTOR,
reduces the mRNA levels of IL-6 and other cytokines, and
selectively inhibits the translation of membrane-bound cytokine
IL-1, regulating SASP. A previous study confirmed that rapamycin
improved aging-associated diseases and tumors by inhibiting
aging-associated inflammation (44). Simvastatin is an inhibitor of
β-hydroxy β-methylglutaryl-CoA (HMG-CoA) reductase, and Liu et
al (45) demonstrated that
simvastatin reduced the SASP of aging human fibroblasts by
inhibiting protein isopentenylation without affecting the
stagnation of age growth. The study by Nacarelli et al
(36) used a Ras-induced IMR90
cell senescence model to demonstrate that the down-regulation of
HMGA1 or NAMPT, or using FK886 in senescent cells, may inhibit the
expression of SASP. They confirmed that NAD+ promotes
the activity of NF-κB to mediate SASP by inhibiting the
AMP-activated protein kinase (AMPK) signaling pathway, thereby
inhibiting the proliferation of tumor cells. They postulated that
targeting NAMPT may be an effective strategy to inhibit
proinflammatory and tumor microenvironment promotion in tumors with
high expression levels of HMGA1 for patients who are undergoing
lifespan-extending cancer treatment (such as chemotherapy or
radiotherapy).
Activation of apoptosis
Wang et al(46) demonstrated that a cell division
cycle kinase 7 (CDC7) inhibitor (XL413) specifically induced aging
in TP53 mutant liver cancer cells, whereas sertraline, a drug used
to treat depression, specifically promoted the apoptosis of aging
liver cancer cells by down-regulating the mTOR signaling pathway.
Guerrero et al (47)
discovered that cardiac glycosides are anti-aging drugs that can
selectively kill senescent cells in the body, thereby avoiding the
toxic side-effects of conventional drug therapies. It has been
demonstrated that senescent cells exert a sensitizing effect with
regard to ouabain-induced apoptosis, which is partly mediated by
inducing the apoptosis-promoting Bcl-2 family protein, NOXA.
Hickson et al (48)
demonstrated for the first time that anti-aging drugs may
successfully remove senescent cells in human trials, where networks
of senescent cell-resisting apoptosis pathways (SCAPs) were
discovered. The anti-aging drugs, dasatinib and quercetin (a
flavonoid), have been used to target these SCAP nodes: Dasatinib
targets tyrosine kinase, whereas the flavonoid quercetin targets
Bcl-2, hypoxia-inducible factor-1, phosphoinositide 3-kinase and
p21, promoting apoptosis in senescent cells and allowing cancer
cells to start the process of apoptosis for self-destructive
purposes (49).
Immune-based elimination of senescent
cells
Researchers have indicated that the body removes
senescent cells by recruiting immune cells. NRAS-H12V can cause
oncogene-induced cell senescence, and these senescent cells can be
removed via both the innate and adaptive immune systems, such as
natural killer (NK) cells and macrophages. Ta et al
(50) demonstrated that
dipeptidyl-peptidase 4 (DPP4; also known as CD26) was highly
expressed in aging human fibroblasts, and senescent cells were
preferentially cleared by NK cells that recognize anti-DPP4
antibodies. Removing aging and damaged cells provides a favorable
microenvironment for tissue regeneration, which stimulates the
proliferation and differentiation of neighboring cells, including
tissue stem cells, enabled damaged tissues to be replenished with
healthy cells, aiding in tissue repair. The results revealed that,
in different models, the immune system possesses a variety of
mechanisms concerning the clearance of senescent cells. Amor et
al (51) analyzed the
expression of transmembrane proteins found in human and mouse
senescent cells, and demonstrated that urokinase-type
proenzyme-activating receptor (uPAR) was highly expressed in
senescent cells both in vivo and in vitro. Of note, a
soluble uPAR (suPAR) that lacked the transmembrane region was shown
to be a component secreted during the SASP response. The presence
of suPAR is a hallmark of several chronic diseases, including
diabetes and nephropathy. Subsequently, the investigators designed
CAR-T cells targeting uPAR, and tested these in mouse models of
several aging-associated diseases, including cancer and liver
fibrosis. It was found that CAR-T cell therapy targeting uPAR led
to the elimination of senescent cells in mouse models of liver and
lung cancer. Similarly, CAR-T cell therapy improved the survival
rate in a mouse model of lung cancer when used in association with
a drug previously shown to induce senescence in this cancer type
(Navitoclax).
As research has progressed in this area, Childs
et al (52) subsequently
demonstrated that a series of drugs can be targeted to induce the
death of senescent cells, although they exert minimal effects on
non-senescent cells; these drugs have been termed 'senolytics', and
they have been used to remove senescent cells from the human body.
A number of senolytic drugs have been successively developed, such
as dasatinib, mistletoe extracts, navitoclax (ABT263), piperine,
phenanthrone, A1331852 and A1155463, and numerous other drugs are
undergoing clinical studies and are expected to be successively
applied in the future (53,54) (https://clini-caltrials.gov/ct2/results?pg=1&load=cart&id=NCT04063124).
Jeon et al (55) collected
chondrocyte samples from patients with severe osteoarthritis and
cultured 3D-stereoscopic cartilage tissue models in vitro,
and their results revealed that the drug under investigation,
UBX0101, was able to both selectively remove senescent cells from
the joints and effectively prevent the generation and development
of osteoarthritis. The results of an interim report of a clinical
trial for the treatment of dysfunction in diabetic patients with
chronic kidney disease indicated that the use of dasatinib and
quercetin in combination chemotherapy removed senescent cells from
tissues, delayed physical dysfunction and reduced inflammatory
cytokine secretion associated with human aging (48). Similarly, another study (56) featuring a clinical trial study
conducted on a small group of patients with pulmonary fibrosis
revealed that dasatinib and quercetin selectively removed senescent
cells to treat idiopathic pulmonary fibrosis. Clinical trials of
another senolytic drug, UBX1967, for the treatment of ocular
diseases in the elderly are also advancing (57).
5. Conclusion
As the understanding of cell aging gradually
develops, and the mechanisms underlying the etiology of
aging-associated diseases are better understood, researchers will
also better understand the pathogenesis of aging-associated
diseases, and this knowledge should provide novel therapeutic
targets, giving rise to novel therapeutic approaches in the
treatment of aging-associated diseases.
Funding
The present study was supported by funding from
National Key R&D Program of China (grant no. 2016YFC1303100),
and the National Natural Science Foundation of China (grant nos.
31570803, 81773090, 81272879 and 81402151).
Availability of data and materials
Not applicable.
Authors' contributions
ZW, JG, YO and CX were involved in the conception of
the study. JG and HL were involved in the literature search and
critical reviewing of the manuscript. ZW and JG were involved in
the preparation of the draft of the manuscript. ZW, JG, YO and CX
were involved in the revising and editing of the manuscript. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgements
Not applicable.
References
|
1
|
Kuwano K, Araya J, Hara H, Minagawa S,
Takasaka N, Ito S, Kobayashi K and Nakayama K: Cellular senescence
and autophagy in the pathogenesis of chronic obstructive pulmonary
disease (COPD) and idiopathic pulmonary fibrosis (IPF). Respir
Investig. 54:397–406. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shimizu H, Bolati D, Adijiang A, Muteliefu
G, Enomoto A, Nishijima F, Dateki M and Niwa T: NF-κB plays an
important role in indoxyl sulfate-induced cellular senescence,
fibrotic gene expression, and inhibition of proliferation in
proximal tubular cells. Am J Physiol Cell Physio. 301:C1201–C1212.
2011. View Article : Google Scholar
|
|
3
|
Lansu K and Gentile S: Potassium channel
activation inhibits proliferation of breast cancer cells by
activating a senescence program. Cell Death Dis. 4:e6522013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ma J, Hu X, Liao C, Xiao H, Zhu Q, Li Y,
Liu Z, Tao A, He Z, Xu C and Zheng K: Gypenoside L inhibits
proliferation of liver and esophageal cancer cells by inducing
senescence. Molecules. 24:10542019. View Article : Google Scholar :
|
|
5
|
Schosserer M, Grillari J and Breitenbach
M: The dual role of cellular senescence in developing tumors and
their response to cancer therapy. Front Oncol. 7:2782017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kurz DJ, Decary S, Hong Y and Erusalimsky
JD: Senescence-associated (beta)-galactosidase reflects an increase
in lysosomal mass during replicative ageing of human endothelial
cells. J Cell Sci. 113(Pt 20): 3613–3622. 2000.PubMed/NCBI
|
|
7
|
Dimri GP, Lee X, Basile G, Acosta M, Scott
G, Roskelley C, Medrano EE, Linskens M, Rubelj I, Pereira-Smith O,
et al: A biomarker that identifies senescent human cells in culture
and in aging skin in vivo. Proc Natl Acad Sci USA. 92:9363–9367.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sosińska P, Mikuła-Pietrasik J, Ryżek M,
Naumowicz E and Książek K: Specificity of cytochemical and
fluorescence methods of senescence-associated β-galactosidase
detection for ageing driven by replication and time.
Biogerontology. 15:407–413. 2014. View Article : Google Scholar
|
|
9
|
Coppé JP, Patil CK, Rodier F, Sun Y, Muñoz
DP, Goldstein J, Nelson PS, Desprez PY and Campisi J:
Senescence-associated secretory phenotypes reveal
cell-nonautonomous functions of oncogenic RAS and the p53 tumor
suppressor. PLoS Biol. 6:2853–2868. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ovadya Y and Krizhanovsk V: Senescent
cells: SASPected drivers of age-related pathologies.
Biogerontology. 15:627–642. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Donate LE and Blasco MA: Telomeres in
cancer and ageing. Philos Trans R Soc Lond B Biol Sci. 366:76–84.
2011. View Article : Google Scholar :
|
|
12
|
Muller HJ: The Remaking of Chromosomes.
The Collecting Net, Woods Hole. 13:181–198. 1938.
|
|
13
|
Leontieva OV and Blagosklonny MV: Tumor
promoter-induced cellular senescence: Cell cycle arrest followed by
geroconversion. Oncotarget. 5:12715–12727. 2014. View Article : Google Scholar
|
|
14
|
Chien M, Rinker-Schaeffer C and Stadler
WM: A G2/M growth arrest response to low-dose intermittent H2O2 in
normal uroepithelial cells. Int J Oncol. 17:425–432.
2000.PubMed/NCBI
|
|
15
|
Campisi J and Fabrizio FD: Cellular
senescence: When bad things happen to good cells. Nat Rev Mol Cell
Biol. 8:729–740. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin HK, Chen Z, Wang G, Nardella C, Lee
SW, Chan CH, Yang WL, Wang J, Egia A, Nakayama KI, et al: Skp2
targeting suppresses tumorigenesis by Arf-p53-independent cellular
senescence. Nature. 464:374–379. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vigneron A and Vousden KH: p53, ROS and
senescence in the control of aging. Aging (Albany NY). 2:471–474.
2010. View Article : Google Scholar
|
|
18
|
Kochetkova EY, Blinova GI, Bystrova OA,
Martynova MG, Pospelov VA and Pospelova TV: Targeted elimination of
senescent Ras-transformed cells by suppression of MEK/ERK pathway.
Aging (Albany NY). 9:2352–2375. 2017. View Article : Google Scholar
|
|
19
|
Mooi WJ and Peeper DS: Oncogene-induced
cell senescence-halting on the road to cancer. N Engl J Med.
355:1037–1046. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Braig M and Schmitt CA: Oncogene-induced
senescence: Putting the brakes on tumor development. Cancer Res.
66:2881–2884. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen Z, Trotman LC, Shaffer D, Lin HK,
Dotan ZA, Niki M, Koutcher JA, Scher HI, Ludwig T, Gerald W, et al:
Crucial role of p53-dependent cellular senescence in suppression of
Pten-deficient tumorigenesis. Nature. 436:725–730. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Alimonti A, Nardella C, Chen Z, Clohessy
JG, Carracedo A, Trotman LC, Cheng K, Varmeh S, Kozma SC, Thomas G,
et al: A novel type of cellular senescence that can be enhanced in
mouse models and human tumor xenografts to suppress prostate
tumorigenesis. J Clin Invest. 120:681–693. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Peeper DS: PICS-ure This: Prosenescence
Therapy? Cancer Cell. 17:219–210. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tato-Costa J, Casimiro S, Pacheco T, Pires
R, Fernandes A, Alho I, Pereira P, Costa P, Castelo HB, Ferreira J
and Costa L: Therapy-induced cellular senescence induces
epithelial-to-mesenchymal transition and increases invasiveness in
rectal cancer. Clin Colorectal Cancer. 15:170–178. 2016. View Article : Google Scholar
|
|
25
|
Gewirtz DA, Holt SE and Elmore LW:
Accelerated senescence: An emerging role in tumor cell response to
chemotherapy and radiation. Biochem Pharmacol. 76:947–957. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tonnessen-Murray CA, Lozano G and Jackson
JG: The regulation of cellular functions by the p53 protein:
Cellular senescence. Cold Spring Harb Perspect Med. 7:a0261122017.
View Article : Google Scholar
|
|
27
|
Rodier F and Campisi J: Four faces of
cellular senescence. J Cell Biol. 192:547–556. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Meiyanto E, Putri H, Larasati YA, Utomo
RY, Jenie RI, Ikawati M, Lestari B, Yoneda-Kato N, Nakamae I,
Kawaichi M and Kato JY: Anti-proliferative and anti-metastatic
potential of curcumin analogue, pentagamavunon-1 (PGV-1), toward
highly metastatic breast cancer cells in correlation with ROS
generation. Adv Pharm Bull. 9:445–452. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dasgupta J, Kar S, Liu R, Joseph J,
Kalyanaraman B, Remington SJ, Chen C and Melendez JA: Reactive
oxygen species control senescence-associated matrix
metalloproteinase-1 through c-Jun-N-terminal kinase. J Cell
Physiol. 225:52–62. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Acosta JC and Gil J: A role for CXCR2 in
senescence, but what about in cancer? Cancer Res. 69:2167–2170.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ortiz-Montero P, Londoño-Vallejo A and
Vernot JP: Senescence-associated IL-6 and IL-8 cytokines induce a
self- and cross-reinforced senescence/inflammatory milieu
strengthening tumorigenic capabilities in the MCF-7 breast cancer
cell line. Cell Commun Signal. 15:172017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Carrière C, Gore AJ, Norris AM, Gunn JR,
Young AL, Longnecker DS and Korc M: Deletion of Rb accelerates
pancreatic carcinogenesis by oncogenic kras and impairs senescence
in premalignant lesions. Gastroenterology. 141:1091–1101. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Malaquin N, Vercamer C, Bouali F, Martien
S, Deruy E, Wernert N, Chwastyniak M, Pinet F, Abbadie C and
Pourtier A: Senescent fibroblasts enhance early skin carcinogenic
events via a paracrine MMP-PAR-1 axis. PLoS One. 8:e636072013.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen F, Long Q, Fu D, Zhu D, Ji Y, Han L,
Zhang B, Xu Q, Liu B, Li Y, et al: Targeting SPINK1 in the damaged
tumour micro-environment alleviates therapeutic resistance. Nat
Commun. 9:43152018. View Article : Google Scholar
|
|
35
|
Zhang B, Fu D, Xu Q, Cong X, Wu C, Zhong
X, Ma Y, Lv Z, Chen F, Han L, et al: The senescence-associated
secretory phenotype is potentiated by feedforward regulatory
mechanisms involving Zscan4 and TAK1. Nat Commun. 9:17232018.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nacarelli T, Lau L, Fukumoto T, Zundell J,
Fatkhutdinov N, Wu S, Aird KM, Iwasaki O, Kossenkov AV, Schultz D,
et al: NAD+ metabolism governs the proinflammatory
senescence-associated secretome. Nat Cell Biol. 21:397–407. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu Q, Long Q, Zhu D, Fu D, Zhang B, Han L,
Qian M, Guo J, Xu J, Cao L, et al: Targeting amphiregulin (AREG)
derived from senescent stromal cells diminishes cancer resistance
and averts programmed cell death 1 ligand (PD-L1)-mediated
immunosuppression. Aging Cell. 18:e130272019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang ST, Huang SW, Liu KT, Lee TY, Shieh
JJ and Wu CY: Atorvastatin-induced senescence of hepatocellular
carcinoma is mediated by downregulation of hTERT through the
suppression of the IL-6/STAT3 pathway. Cell Death Discov. 6:172020.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Miyazoe Y, Miuma S, Miyaaki H, Kanda Y,
Nakashiki S, Sasaki R, Haraguchi M, Shibata H, Honda T, Taura N and
Nakao K: Extracellular vesicles from senescent hepatic stellate
cells promote cell viability of hepatoma cells through increasing
EGF secretion from differentiated THP-1 cells. Biomed Rep.
12:163–170. 2020.PubMed/NCBI
|
|
40
|
Prattichizzol F, Giuliani A, Recchioni R,
Bonafè M, Marcheselli F, De Carolis S, Campanati A, Giuliodori K,
Rippo MR, Brugè F, et al: Anti-TNF-α treatment modulates SASP and
SASP-related microRNAs in endothelial cells and in circulating
angiogenic cells. Oncotarget. 7:11945–11958. 2016. View Article : Google Scholar
|
|
41
|
Xu M, Tchkonia T, Ding H, Ogrodnik M,
Lubbers ER, Pirtskhalava T, White TA, Johnson KO, Stout MB, Mezera
V, et al: JAK inhibition alleviates the cellular senescence
associated secretory phenotype and frailty in old age. Proc Natl
Acad Sci USA. 112:E6301–E6310. 2015. View Article : Google Scholar
|
|
42
|
Alimbetov D, Davis T, Brook AJ, Cox LS,
Faragher RG, Nurgozhin T, Zhumadilov Z and Kipling D: Suppression
of the senescence-associated secretory phenotype (SASP) in human
fibroblasts using small molecule inhibitors of p38 MAP kinase and
MK2. Biogerontology. 17:305–315. 2016. View Article : Google Scholar :
|
|
43
|
Herranz N, Gallage S, Mellone M,
Wuestefeld T, Klotz S, Hanley CJ, Raguz S, Acosta JC, Innes AJ,
Banito A, et al: mTOR regulates MAPKAPK2 translation to control the
senescence-associated secretory phenotype. Nat Cell Biol.
17:1205–1217. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tsubouchi K, Araya J and Kuwano K:
PINK1-PARK2-mediated mitophagy in COPD and IPF pathogeneses.
Inflamm Regen. 38:182018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu S, Uppal H, Demaria M, Desprez PY,
Campisi J and Kapahi P: Simvastatin suppresses breast cancer cell
proliferation induced by senescent cells. Sci Rep. 5:178952015.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang C, Vegna S, Jin H, Benedict B,
Lieftink C, Ramirez C, de Oliveira RL, Morris B, Gadiot J, Wang W,
et al: Inducing and exploiting vulnerabilities for the treatment of
liver cancer. Nature. 574:268–272. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Guerrero A, Herranz N, Sun B, Wagner V,
Gallage S, Guiho R, Wolter K, Pombo J, Irvine EE, Innes AJ, et al:
Cardiac glycosides are broad-spectrum senolytics. Nat Metab.
1:1074–1088. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hickson LJ, Langhi Prata LGP, Bobart SA,
Evans TK, Giorgadze N, Hashmi SK, Herrmann SM, Jensen MD, Jia Q,
Jordan KL, et al: Senolytics decrease senescent cells in humans:
Preliminary report from a clinical trial of Dasatinib plus
Quercetin in individuals with diabetic kidney disease.
EBioMedicine. 47:446–456. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kovacovicova K, Skolnaja M, Heinmaa M,
Mistrik M, Pata P, Pata I, Bartek J and Vinciguerr M: Senolytic
cocktail dasatinib+quercetin (D+Q) does not enhance the efficacy of
senescence-inducing chemotherapy in liver cancer. Front Oncol.
8:4592018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ta NN, Li Y, Schuyler CA, Lopes-Virella MF
and Huang Y: DPP-4 (CD26) inhibitor alogliptin inhibits
TLR4-mediated ERK activation and ERK-dependent MMP-1 expression by
U937 histiocytes. Atherosclerosis. 213:429–435. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Amor C, Feucht J, Leibold J, Ho YJ, Zhu C,
Alonso-Curbelo D, Mansilla-Soto J, Boyer JA, Li X, Giavridis T, et
al: Senolytic CAR T cells reverse senescence-associated
pathologies. Nature. 583:127–132. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Childs BG, Gluscevic M, Baker DJ, Laberge
RM, Marquess D, Dananberg J and van Deursen JM: Senescent cells: An
emerging target for diseases of ageing. Nat Rev Drug Discov.
16:718–735. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhu Y, Doornebal EJ, Pirtskhalava T,
Giorgadze N, Wentworth M, Fuhrmann-Stroissnigg H, Niedernhofer LJ,
Robbins PD, Tchkonia T and Kirkland JL: New agents that target
senescent cells: The flavone, fisetin, and the BCL-XL inhibitors,
A1331852 and A1155463. Aging (Albany NY). 9:955–963. 2017.
View Article : Google Scholar
|
|
54
|
Kirkland JL, Tamara T, Yi Z, Niedernhofer
LJ and Robbins PD: The clinical potential of senolytic drugs. J Am
Geriatr Soc. 65:2297–2301. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Jeon OH, Kim C, Laberge RM, Demaria M,
Rathod S, Vasserot AP, Chung JW, Kim DH, Poon Y, David N, et al:
Local clearance of senescent cells attenuates the development of
post-traumatic osteoarthritis and creates a pro-regenerative
environment. Nat Med. 23:775–781. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Justice JN, Nambiar AM, Tchkonia T,
LeBrasseur NK, Pascual R, Hashmi SK, Prata L, Masternak MM,
Kritchevsky SB, Musi N and Kirkland JL: Senolytics in idiopathic
pulmonary fibrosis: Results from a first-in-human, open-label,
pilot study. EBioMedicine. 40:554–563. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Van Deursen JM: Senolytic therapies for
healthy longevity. Science. 364:636–637. 2019. View Article : Google Scholar : PubMed/NCBI
|