Introduction
Hepatocellular carcinoma (HCC) is a type of primary
liver cancer characterized by high incidence and mortality rates
(1). In recent years, although
significant progress has been made in the treatment of HCC, the
overall prognosis remains dismal (2). Surgery is the first and most
effective method for the treatment of patients with HCC; however,
chemotherapy is considered the most appropriate method for some
patients with advanced disease (3,4).
Sorafenib is a multi-target anti-tumor drug, which is currently the
first-line treatment drug for patients with advanced HCC (5). However, due to the development of
drug resistance during treatment, sorafenib becomes less effective
(6,7). Furthermore, immune escape has been
demonstrated to be one of the main obstacles to the treatment of
HCC patients (8). Therefore, it
is crucial to investigate the molecular mechanisms underlying the
development of sorafenib resistance and immune escape in HCC
cells.
Long non-coding RNAs (lncRNAs) are a family of
transcripts >200 nucleotides in length, which do not have the
ability to encode proteins (9).
LncRNAs have been verified to be crucial regulators in several
biological processes, such as cell metastasis, differentiation,
inflammation and immune escape (10–12). Currently, lncRNA KCNQ1 overlapping
transcript 1 (KCNQ1OT1) was proven to be associated with drug
resistance in diverse types of cancer. For example, Zhang et
al reported that KCNQ1OT1 was increased in cisplatin-resistant
tongue squamous cell carcinoma (TSCC) and promoted chemoresistance
of TSCC cells (13). Ren et
al demonstrated that the expression of KCNQ1OT1 was higher in
paclitaxel-resistant lung adenocarcinoma (LAD) tissues and cells,
and KCNQ1OT1 knockdown enhanced the sensitivity of LAD to
paclitaxel (14). However, there
are yet no reports on the role and mechanism of action of KCNQ1OT1
in sorafenib resistance and immune escape in HCC cells.
LncRNAs may act as microRNA (miRNA) sponges to
regulate the expression and activities of miRNAs (15). As a class of RNA molecules without
protein-coding ability, miRNAs contain 18–22 nucleotides and mainly
regulate gene expression by recognizing the 3′ untranslated region
(UTR) of their target mRNAs (16). In particular, miR-506 has been
demonstrated to act as a tumor suppressor in diverse human cancers,
such as colorectal (17),
cervical (18) and ovarian
(19) cancers. Moreover, Zhou
et al confirmed that miR-506 was able to reduce oxaliplatin
resistance in colorectal cancer cells (20). Wang et al suggested that
miR-506 was decreased in the serum of patients with
sorafenib-resistant thyroid carcinoma, and that miR-506
overexpression could enhance the sensitivity of thyroid carcinoma
cells to sorafenib (21). These
findings indicated that miR-506 plays a key role in the development
and chemoresistance of several tumors. Moreover, miRNAs can
participate in the regulation of a number of biological processes,
including immune escape of tumor cells (22). However, the exact regulatory
mechanisms of miR-506 in sorafenib resistance and immune escape in
HCC remain unclear.
Programmed death-ligand-1 (PD-L1) plays a key role
in inhibiting tumor immunity and promoting tumor progression after
binding to the receptor programmed death 1 (PD-1), which is
expressed on the surface of T lymphocytes (23). To date, a variety of miRNAs have
been confirmed to affect the oncogenesis and drug resistance of
human cancers by regulating PD-L1 expression. For example, miR-34a
reduced chemoresistance of glioma cells by targeting PD-L1 in
(24), and the miR-200/PD-L1 axis
reduced immunosuppression and metastasis of CD8+ T cells
in lung cancer (25). However,
whether miR-506 can target PD-L1 in HCC cells remains to be
elucidated.
The aim of the present study was to investigate the
levels of KCNQ1OT1, miR-506 and PD-L1 in sorafenib-resistant HCC
tissues and cells, and to explore the roles of KCNQ1OT1 and miR-506
in sorafenib-resistant HCC cell proliferation, apop-tosis and
metastasis. Moreover, the effects of KCNQ1OT1 and miR-506 on the
tumor microenvironment and T-cell apoptosis were investigated.
Materials and methods
Tissue collection
Sorafenib-sensitive (n=25) and sorafeni-resistant
(n=38) HCC tissue sections were collected from the Sanquan College
of Xinxiang Medical University. All the collected samples were
immediately placed in liquid nitrogen and then stored in a −80°C
refrigerator for RNA extraction. The protocol of the present study
was approved by the Ethics Committee of the Sanquan College of
Xinxiang Medical University, and all the patients provided written
informed consent.
Cell culture
SK-HEP-1 and Huh-7 cells were purchased from the
Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). The corresponding sorafenib-resistant HCC cells
(SK-HEP-1/sorafenib and Huh-7/sorafenib) were constructed by
exposing SK-HEP-1 and Huh-7 cells to gradually increasing
concentrations of sorafenib and then continuously culturing in a
sorafenib-containing medium for 2 months. The maximum dose of
sorafenib was 20 μM (26).
All cells were kept in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
and 1% penicillin/streptomycin (Gibco; Thermo Fisher Scientific,
Inc.) in an atmosphere of 5% CO2 and 37°C.
Cell transfection
Short hairpin RNA (shRNA) against KCNQ1OT1 and its
negative control (sh-NC), mimics of miR-506 (miR-506) and its
matched control (miR-NC), inhibitors of miR-506 (anti-miR-506) and
its matched control (anti-NC), KCNQ1OT1 overexpression vector
(KCNQ1OT1) and its corresponding control (pcDNA), were synthesized
by GenePharma. These oligonucleotides or vectors were transfected
into cells using Lipofectamine® 3000 Transfection
Reagent (Invitrogen; Thermo Fisher Scientific, Inc.) based on the
manufacturer's instructions.
RNA extraction and quantitative PCR
(RT-qPCR) analysis
TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) was used to isolate total RNA from HCC
tissues and cells. The concentrations of extracted RNA samples were
detected by NanoDrop 2000c (Thermo Fisher Scientific, Inc.).
Subsequently, RNAs were reversely transcribed into cDNAs by
PrimeScript RT reagent kit (Takara Biotechnology Co., Ltd.) or
TaqMan Reverse Transcription Reagents (Thermo Fisher Scientific,
Inc.). Next, qPCR analysis was conducted using FastStart Universal
SYBR Green Master (Roche Diagnostics) on a Real-Time PCR Detection
System (Thermo Fisher Scientific, Inc.). The thermocycling
conditions were as follows: i) 95°C for 5 min; ii) 40 cycles at
95°C for 30 sec, 60°C for 45 sec and 72°C for 30 sec; iii)
dissolving curve at 94°C for 90 sec, 60°C for 180 sec and 94°C for
10 sec. The relative expression of KCNQ1OT1, PD-L1 and miR-506 was
examined by the 2−ΔΔCq method (27), with GAPDH or U6 as an internal
reference. The sequences of the primers were as follows: KCNQ1OT1
F: 5′-CTTTGCAGCAACCTCCTTGT-3′ and R: 5′-TGGGGTGAGGGATCTGAA-3′;
miR-506 F: 5′-ATCCAG TGCGTGTCGTG-3′ and R:
5′-TGCTTAAGGCACCCTTCT-3′; PD-L1 F: 5′-TCCACTCAATGC CTCAAT-3′ and R:
5′-GAAGACCTCACA GAC TCAA-3′; GAPDH F:
5′-CCATGTTCGTCATGGGTGTGAACCA-3′ and R: 5′-GCCAGTAGAGGCAGG GAT GAT
GTT C-3′; and U6 F: 5′-AGAGCCTGTGGTGTCCG-3′ and R:
5′-CATCTTCAAAGCACTTCCCT-3′.
Sorafenib resistance assay
Sorafenib resistance was evaluated by the Cell
Counting Kit-8 (CCK-8) assay. SK-HEP-1, Huh-7, SK-HEP-1/sorafenib
or Huh-7/sorafenib cells were incubated in 96-well plates at a
density of 3.0×103cells/well. The cells were then
treated with different doses of sorafenib for 48 h. Subsequently,
10 μl CCK-8 solution (5 mg/ml; Beijing Solarbio Science
& Technology Co., Ltd.) was added into each well and maintained
for 4 h at 37°C. At last, the absorbance at 450 nm was examined
using a microplate reader (Bio-Rad Laboratories, Inc.). The value
of IC50 was determined by relative survival curve.
Immunohistochemistry (IHC) assay
After being fixed with 4% paraformaldehyde (Beyotime
Institute of Biotechnology) for 48 h at room temperature,
sorafenib-sensitive or -resistant HCC tissue samples were embedded
in paraffin and cut into 4-μm sections. Then, the sections
were deparaffinized, hydrated through a graded ethanol series and
treated with H2O2 in methanol for 15 min at
room temperature. Thereafter, the slides were washed three times (5
min each) using PBS (Beijing Solarbio Science & Technology Co.,
Ltd.) and interacted with normal goat serum for 20 min followed by
incubation with anti-PD-L1 (bs-10159R; 1:2,000; BIOSS) at 4°C
overnight and the indicated secondary antibody (bs-0308R; 1:5,000;
BIOSS) for 30 min at room temperature. After staining with
diaminobenzidine (Beyotime Institute of Biotechnology) for 5 min at
room temperature and counterstaining with hematoxylin for 50 sec at
room temperature, the slides were photographed using a digital
microscope camera (Nikon Corporation) at a magnification of
×100.
Western blot analysis
First, total protein was extracted from SK-HEP-1,
Huh-7, SK-HEP-1/sorafenib or Huh-7/sorafenib cells using RIPA lysis
buffer (Beijing Solarbio Science & Technology Co., Ltd.). Then,
extracted protein samples were quantified by BCA Protein Assay kit
(Beijing Solarbio Science & Technology Co., Ltd.), separated by
10% SDS-PAGE (Beijing Solarbio Science & Technology Co., Ltd.)
and transferred onto PVDF membranes (Pall Life Sciences). The
membranes were then blocked with skimmed milk for 1 h at room
temperature. Subsequently, the membranes were incubated overnight
at 4°C with primary antibody anti-PD-L1 (bs-10159R; 1:2,000; BIOSS)
followed by incubation with matched horseradish
peroxidase-conjugated secondary antibody (bs-0308R; 1:5,000; BIOSS)
for 2 h at room temperature. The protein bands were detected via
RapidStep ECL Reagent (EMD Millipore).
Transwell assay
For the detection of cell migration, transfected
SK-HEP-1/sorafenib and Huh-7/sorafenib cells (2.0×104
cells) were suspended in serum-free DMEM (Gibco; Thermo Fisher
Scientific. Inc.) and seeded into the upper chamber of a Transwell
insert (8-μm pore size; Corning Inc.). DMEM (Gibco; Thermo
Fisher Scientific. Inc.) supplemented with 10% FBS (Gibco; Thermo
Fisher Scientific. Inc.) was added into the lower chamber. After
incubation for 24 h, cells that did not migrate were removed and
cells that migrated were fixed in methanol for 30 min at 37°C and
then stained with 0.5% crystal violet solution for 15 min at 37°C
(Beijing Solarbio Science & Technology Co., Ltd.). Stained
cells were examined under a light microscope (Nikon Corporation) at
a magnification of ×100. For the detection of cell invasion, the
protocols were the same as those in cell migration assay, except
that the upper chamber was pre-coated with Matrigel (Beijing
Solarbio Science & Technology Co., Ltd.) for 30 min at
37°C.
Sorafenib-resistant HCC/T-cell co-culture
model
First, the separation of peripheral blood
mononuclear cells (PBMCs) was conducted by density gradient
centrifugation. Separated PBMCs (2.0×106 per well) were
seeded into 6-well plates. Subsequently, T cells were activated by
incubating PBMCs with SK-HEP-1/sorafenib or Huh-7/sorafenib cell
lysates, anti-CD3e (bsm-52386R; 1:2,000; BIOSS) and anti-CD28
(bs-8865R; 1:2,000; BIOSS) for 48 h. Subsequently, activated T
cells were co-cultured with SK-HEP-1/sorafenib or Huh-7/sorafenib
cells at 37°C for 16 h at a ratio of 10:1. Finally, co-culture
media were collected for the following experiments.
ELISA
The levels of tumor necrosis factor (TNF)-α,
interferon (IFN)-γ, interleukin (IL)-2, IL-10 and transforming
growth factor (TGF)-β secreted by T cells were detected using ELISA
Max kits (SEKH-0047; SEKH-0046; SEKH-0018; SEKH-0316; Beijing
Solarbio Science & Technology Co., Ltd.) according to the
manufacturer's instructions. In brief, cytokine capture antibodies
were coated in 96-well plates at 4°C overnight, and the wells were
then blocked at room temperature for 1 h. Cell supernatant (100
μl) was added into each well and then incubated for 2 h at
37°C. Next, 100 μl of detection antibody and diluted
streptavidin peroxidase were added in succession. Finally, the
reaction was stopped by adding 3,3′,5,5′-tetramethylbenzidine
substrate (Beijing Solarbio Science & Technology Co., Ltd.) and
sulfuric acid (Sigma-Aldrich; Merck KGaA). The optical density
value at 450 nm was measured via a microplate reader (Bio-Rad
Laboratories, Inc.).
Flow cytometry analysis
At 48 h post-transfection, the apoptosis of
SK-HEP-1/sorafenib and Huh-7/sorafenib cells was evaluated by using
Annexin V-FITC/PI Apoptosis Detection kit (Vazyme Biotech). In
brief, SK-HEP-1/sorafenib and Huh-7/sorafenib cells were harvested
and resuspended in binding buffer at a density of
1.0×105 cells/ml. Then, Annexin V-FITC and PI were added
to the cell suspension and maintained for 15 min at room
temperature in the dark to stain the cells. Finally, flow cytometry
(FACSCalibur; Thermo Fisher Scientific, Inc.) was used to detect
cell apoptosis. The apoptotic cells were analyzed with software
FlowJo 7.6.1 (FlowJo LLC).
To analyze the apoptosis of CD8+T cells,
10 μl/ml anti-PD-L1 (bs-10159R; 1:2,000; BIOSS) was added
into the well of sorafenib-resistant HCC/T-cell co-culture media.
Then, activated T cells were collected and PE-conjugated, and PD-1,
Alexa Fluor 488-conjugated Annexin V and APC-conjugated CD8
antibody were added to stain the PBMCs. The percentage of Annexin
V+ cells in the gated PD-1+/CD8+
population was used to evaluate the apoptosis of CD8+ T
cells.
Dual-luciferase reporter assay
The sequences of KCNQ1OT1 or 3′UTR of PD-L1
containing the putative complementary sequences of wild-type (WT)
or mutant (MUT) miR-506 were amplified and inserted into pcDNA3.1
vector (Guangzhou RiboBio Co., Ltd.) to establish luciferase
reporter vectors (WT-KCNQ1OT1, MUT-KCNQ1OT1, PD-L1 3′UTR-WT and
PD-L1 3′UTR-MUT, respectively). The indicated luciferase reporter
vector and miR-506 or miR-NC were transfected into
SK-HEP-1/sorafenib and Huh-7/sorafenib cells using
Lipofectamine® 3000 Transfection Reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). Subsequently, the luciferase
activity was detected by Dual-Luciferase Reporter Assay kit (Vazyme
Biotech) and normalized to the Renilla luciferase activity.
Statistical analysis
All analyses were conducted by GraphPad Prism 7
(GraphPad Software, Inc.). The data obtained from three independent
experiments are presented as means ± standard deviation. The
significance of the differences was analyzed by Student's t-test or
one-way ANOVA followed by Tukey's post hoc test. The correlations
among KCNQ1OT1, miR-506 and PD-L1 were analyzed by Pearson's
correlation analysis. P<0.05 was considered to indicate
statistically significant differences.
Results
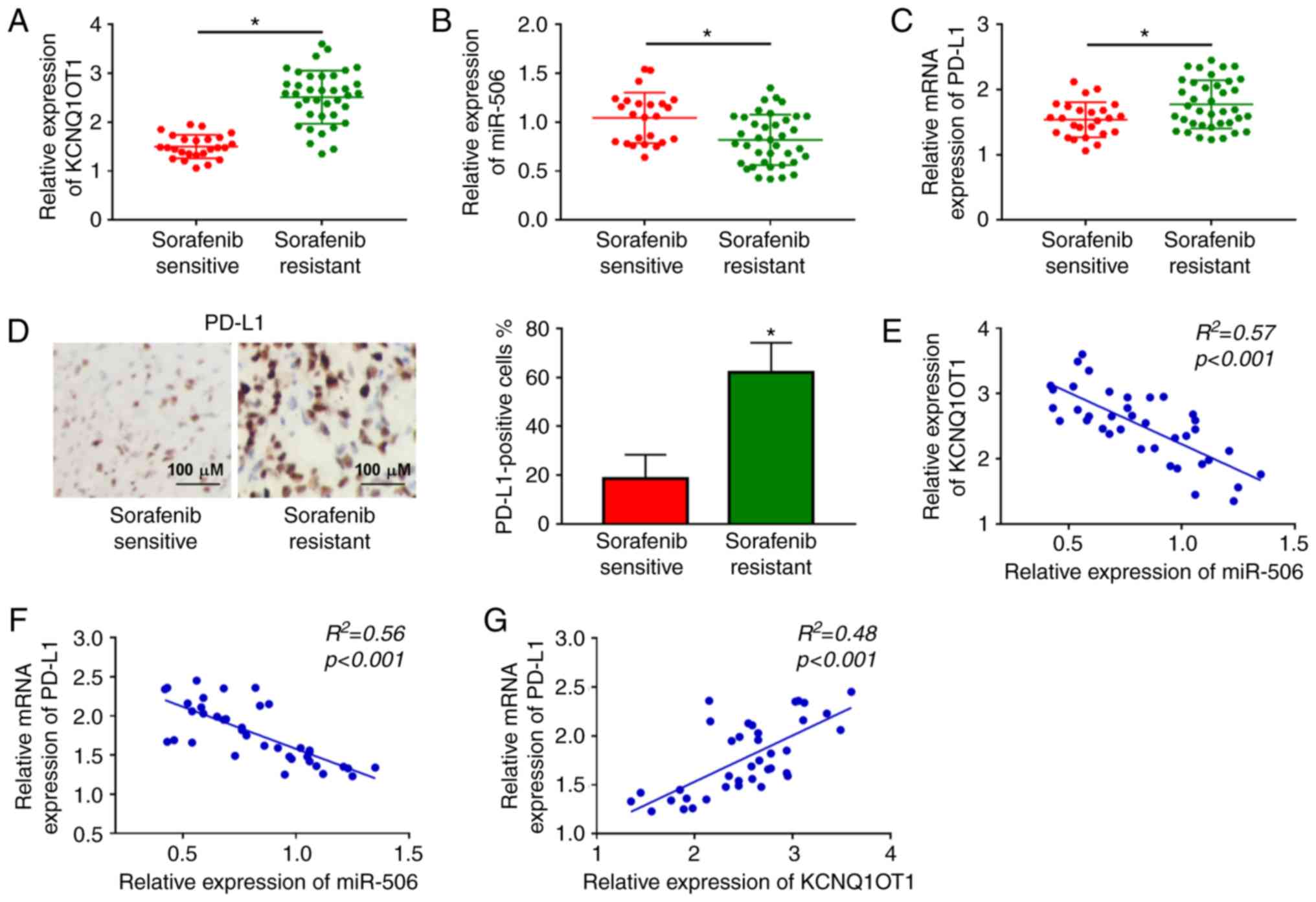
KCNQ1OT1 and PD-L1 are upregulated and
miR-506 is downregulated in sorafenib-resistant HCC tissues
Initially, the relative expression levels of
KCNQ1OT1 and miR-506 in 38 sorafenib-resistant HCC tissue samples
and 25 sorafenib-sensitive HCC tissue samples were determined by
RT-qPCR analysis. The results revealed that the expression level of
KCNQ1OT1 was notably increased, and the expression level of miR-506
was markedly decreased in sorafenib-resistant HCC tissues compared
to those in sorafenib-sensitive HCC tissues (Fig. 1A and B). Moreover, through RT-qPCR
and IHC assays, PD-L1 was shown to be upregulated in
sorafenib-resistant HCC tissues in comparison with
sorafenib-sensitive HCC tissues (Fig.
1C and D). In addition, the correlations among KCNQ1OT1,
miR-506 and PD-L1 were analyzed by Pearson's correlation analysis.
The results revealed that miR-506 expression was negatively
correlated with KCNQ1OT1 expression and PD-L1 mRNA expression
(Fig. 1E and F); moreover, the
mRNA expression of PD-L1 was positively correlated with the
expression of KCNQ1OT1 in sorafenib-resistant HCC tissues (Fig. 1G). These data suggested that the
abnormal expression of KCNQ1OT1, miR-506 and PD-L1 may be
associated with sorafenib resistance in HCC.
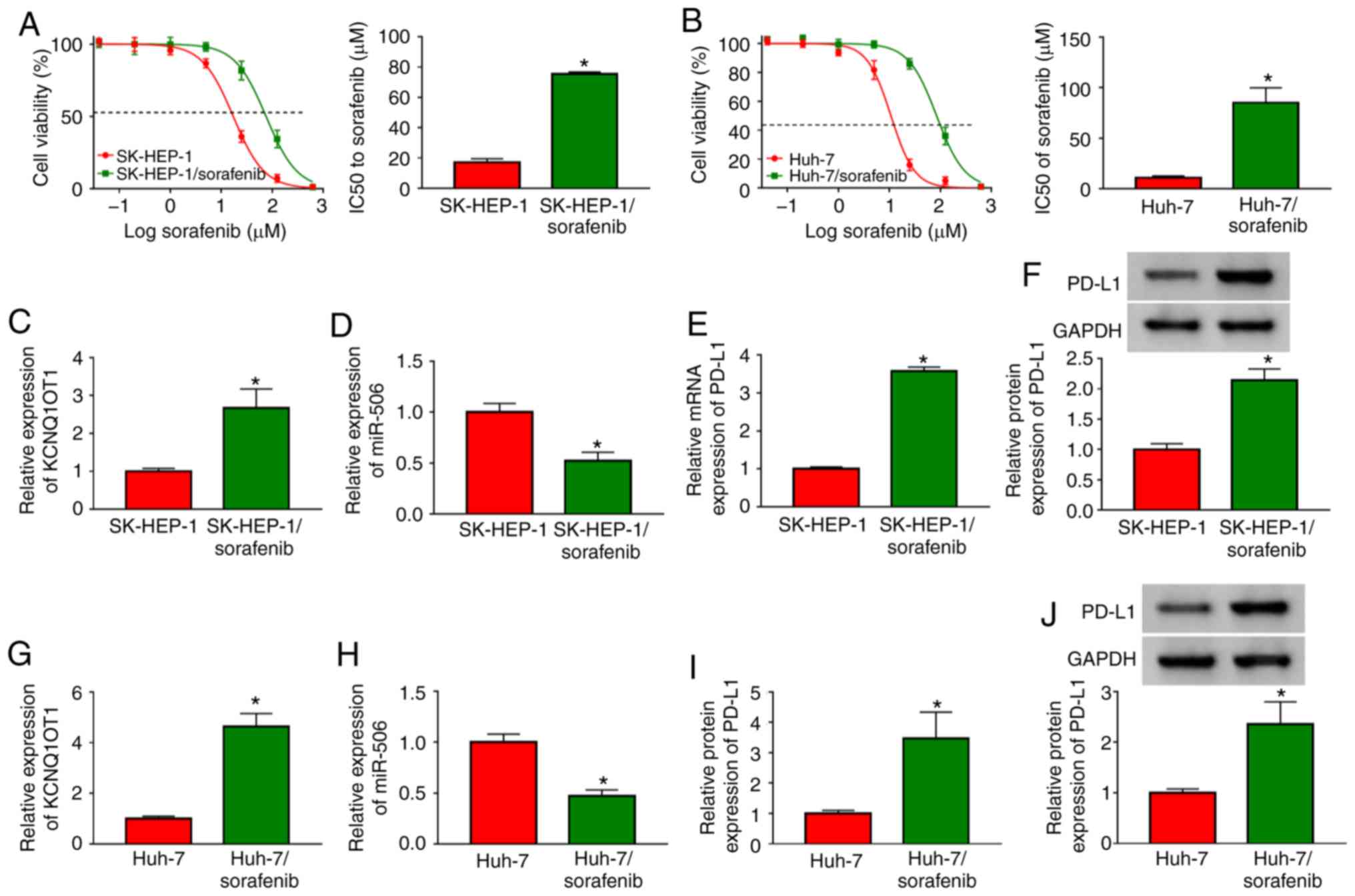
High expression of KCNQ1OT1 and PD-L1 and
low expression of miR-506 is observed in sorafenib-resistant HCC
cells
To explore the mechanism of sorafenib resistance in
HCC cells, two sorafenib-resistant HCC cell lines
(SK-HEP-1/sorafenib and Huh-7/sorafenib) were constructed.
Subsequently, the level of sorafenib resistance in
SK-HEP-1/sorafenib and Huh-7/sorafenib cells was evaluated by CCK-8
assay after exposing sorafenib-resistant HCC cells to different
concentrations of sorafenib for 48 h. The data demonstrated that
IC50 values in SK-HEP-1/sorafenib cells and
Huh-7/sorafenib cells were notably higher compared with those in
SK-HEP-1 and Huh-7 cells (Fig. 2A and
B), indicating that sorafenib resistance developed in
SK-HEP-1/sorafenib and Huh-7/sorafenib cells. Subsequently, the
expression levels of KCNQ1OT1, miR-506 and PD-L1 mRNA in
SK-HEP-1/sorafenib cells were assessed by RT-qPCR analysis and the
protein expression of PD-L1 was analyzed via western blotting. The
results demonstrated that the levels of KCNQ1OT1, and the mRNA and
protein expression of PD-L1 were all obviously upregulated, whereas
miR-506 was significantly downregulated in SK-HEP-1/sorafenib cells
compared with SK-HEP-1 cells (Fig.
2C-F). Similarity, the levels of KCNQ1OT1, and those of PD-L1
mRNA and protein were increased, and the level of miR-506 was
decreased in Huh-7/sorafenib cells (Fig. 2G-J). Taken together, the results
mentioned above indicate that KCNQ1OT1, miR-506 and PD-L1 may be
involved in the sensitivity of HCC cells to sorafenib.
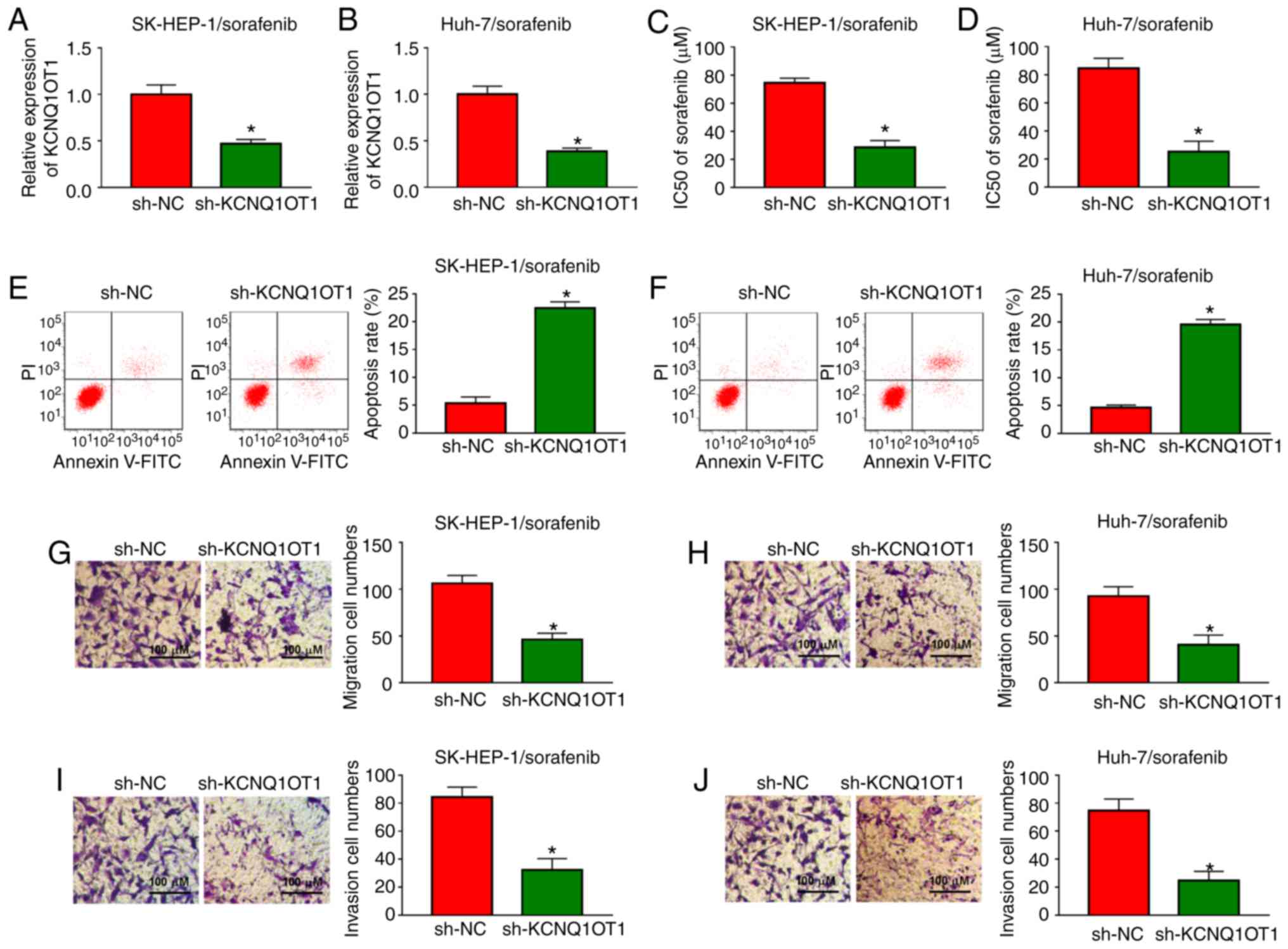
KCNQ1OT1 knockdown enhances sorafenib
sensitivity, promotes apoptosis and inhibits migration and invasion
in sorafenib-resistant HCC cells
In order to investigate the functional roles of
KCNQ1OT1 in sorafenib-resistant HCC cells, a loss-of-function
experiment was carried out by transfecting sh-KCNQ1OT1 into
SK-HEP-1/sorafenib and Huh-7/sorafenib cells. Knockdown efficiency
was evaluated by RT-qPCR and the results demonstrated that
sh-KCNQ1OT1 transfection markedly decreased the expression of
KCNQ1OT1 in both SK-HEP-1/sorafenib and Huh-7/sorafenib cells
(Fig. 3A and B). Then,
SK-HEP-1/sorafenib and Huh-7/sorafenib cells were treated with
different concentrations of sorafenib and IC50 value was
examined by CCK-8 assay. The data demonstrated that the
IC50 value of sorafenib was drastically reduced in
SK-HEP-1/sorafenib and Huh-7/sorafenib cells transfected with
sh-KCNQ1OT1 (Fig. 3C and D),
indicating that the sensitivity of SK-HEP-1 and Huh-7 cells to
sorafenib was enhanced. Furthermore, cell apoptosis was accelerated
by sh-KCNQ1OT1 transfection in SK-HEP-1/sorafenib and
Huh-7/sorafenib cells compared with sh-NC transfected cells
(Fig. 3E and F). KCNQ1OT1
knockdown distinctly inhibited cell migration and invasion in both
SK-HEP-1/sorafenib and Huh-7/sorafenib cells, as illustrated by the
Transwell assay (Fig. 3G-J).
Taken together, these data demonstrated that knockdown of KCNQ1OT1
sensitized HCC cells to sorafenib, induced cell apoptosis, and
suppressed the migration and invasion of sorafenib-resistant HCC
cells.
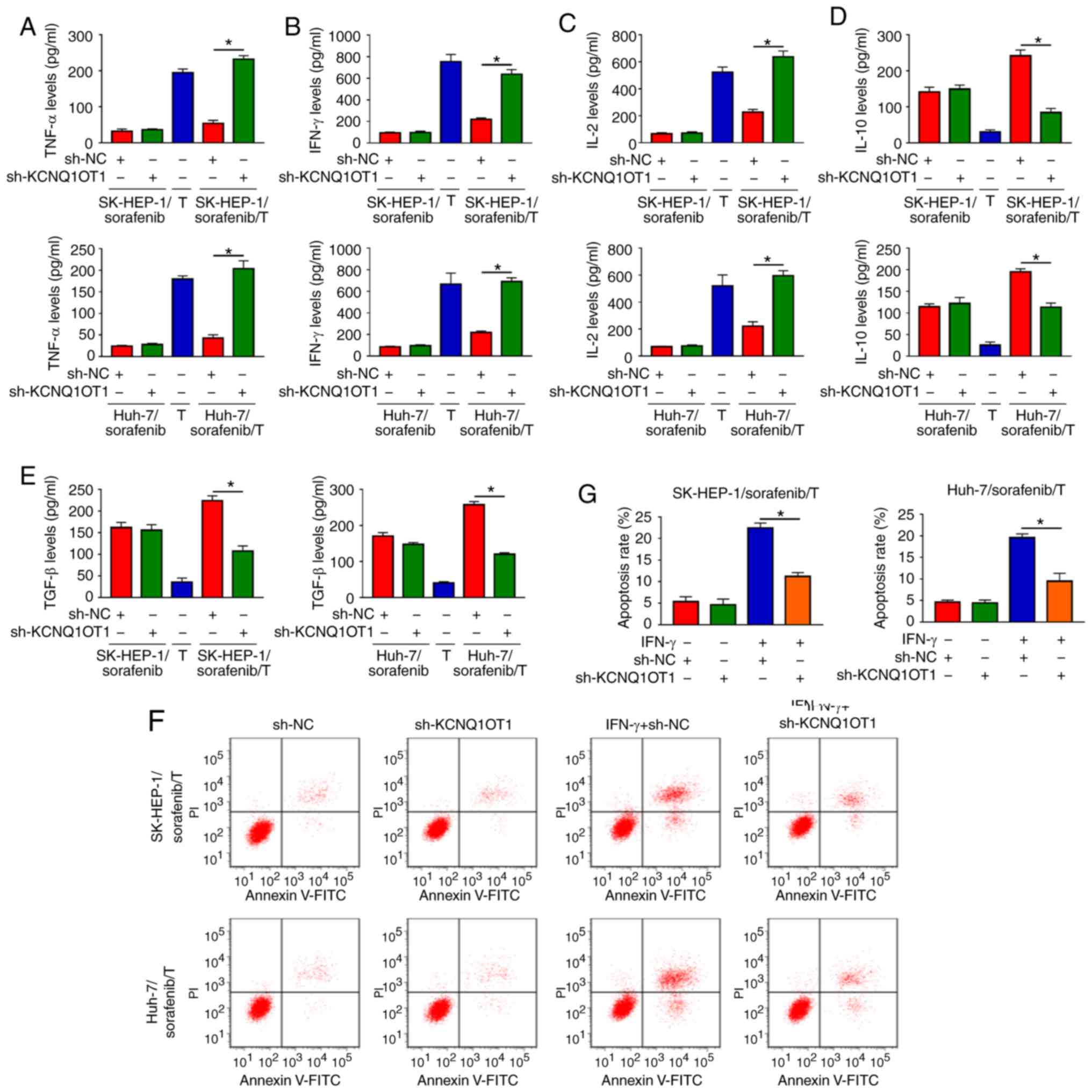
KCNQ1OT1 affects cytokine secretion and
T-cell apoptosis in a sorafenib-resistant HCC/T-cell co-culture
model
To explore the function of KCNQ1OT1 in the immune
surveillance of T cells, two sorafenib-resistant HCC/T-cell
co-culture models we established, namely SK-HEP-1/sorafenib/T and
Huh-7/sorafenib/T. Subsequently, the levels of cytokines secreted
by T cells were detected via ELISA. The results indicated that the
levels of TNF-α, IFN-γ and IL-2 were markedly increased, and the
levels of IL-10 and TGF-β were markedly decreased in
SK-HEP-1/sorafenib cells transfected with sh-KCNQ1OT1 and T-cell
co-culture group compared with those in SK-HEP-1/sorafenib cells
transfected with sh-NC and T-cell co-culture group (Fig. 4A-E). As predicted, similar results
were observed in Huh-7/sorafenib cells transfected with sh-KCNQ1OT1
and co-cultured with T cells (Fig.
4A-E). Furthermore, the apoptosis of CD8+ T cells
was evaluated by flow cytometric analysis. We found that co-culture
of T cells and SK-HEP-1/sorafenib cells or Huh-7/sorafenib cells
treated with IFN-γ and sh-KCNQ1OT1 markedly suppressed the
apoptosis of CD8+ T cells when compared to the control
group (Fig. 4F and G). These data
demonstrated that KCNQ1OT1 knockdown enhanced immune surveillance
of T cells in a sorafenib-resistant HCC/T-cell co-culture
model.
KCNQ1OT1 regulates PD-L1 expression by
sponging miR-506
As dysregulated expression of KCNQ1OT1, miR-506 and
PD-L1 was observed in sorafenib-resistant HCC tissues and cells,
their targeting relationships were predicted using the online
software starBase 3.0. As shown in Fig. 5A, miR-506 was predicted to be a
target of KCNQ1OT1. miR-506 transfection markedly elevated the
expression of miR-506 in SK-HEP-1/sorafenib and Huh-7/sorafenib
cells, indicating that miR-506 was successfully transfected
(Fig. S1A). Next,
dual-luciferase reporter assay was conducted to verify this
prediction. The data demonstrated that co-transfection of miR-506
and WT-KCNQ1OT1 significantly suppressed the luciferase activity in
both SK-HEP-1/sorafenib and Huh-7/sorafenib cells compared with
miR-NC and WT-KCNQ1OT1 co-transfected cells, while co-transfection
with miR-506 or miR-NC and MUT-KCNQ1OT1 did not affect the
luciferase activity (Fig. 5B and
C). After KCNQ1OT1 was successfully transfected into
SK-HEP-1/sorafenib and Huh-7/sorafenib cells (Fig. S1B), the impact of KCNQ1OT1 on
miR-506 expression was investigated. The results revealed that
KCNQ1OT1 knockdown notably increased the expression of miR-506, and
KCNQ1OT1 overexpression notably decreased the expression of miR-506
in both SK-HEP-1/sorafenib and Huh-7/sorafenib cells (Fig. 5D and E). However, miR-506
overexpression did not alter the expression of KCNQ1OT1 in
SK-HEP-1/sorafenib and Huh-7/sorafenib cells (Fig. S1D). Moreover, PD-L1 was
identified as a target of miR-506 using the online software
starBase 3.0 and the complementary sequences are shown in Fig. 5F. Dual-luciferase reporter assay
revealed that, compared to PD-L1 3′UTR-WT and miR-NC co-transfected
groups, the luciferase activity was markedly inhibited in PD-L1
3′UTR-WT and miR-506 co-transfected SK-HEP-1/sorafenib and
Huh-7/sorafenib cells, but the luciferase activity was not affected
in the PD-L1 3′UTR-MUT groups (Fig.
5G and H). As shown in Fig.
S1C, anti-miR-506 transfection markedly decreased the level of
miR-506 in SK-HEP-1/sorafenib and Huh-7/sorafenib cells.
Subsequently, sh-KCNQ1OT1, sh-NC, sh-KCNQ1OT1+anti-miR-506 or
sh-KCNQ1OT1+anti-NC were transfected into SK-HEP-1/sorafenib and
Huh-7/sorafenib cells and the expression of PD-L1 was measured. The
results of the RT-qPCR and western blot assays indicated that the
mRNA and protein expression levels of PD-L1 were notably decreased
by sh-KCNQ1OT1, whereas anti-miR-506 reversed this effect in
SK-HEP-1/sorafenib cells and Huh-7/sorafenib cells (Fig. 5I-L). All these data demonstrated
that KCNQ1OT1 knockdown may inhibit the expression of PD-L1 by
targeting miR-506 in sorafenib-resistant HCC cells.
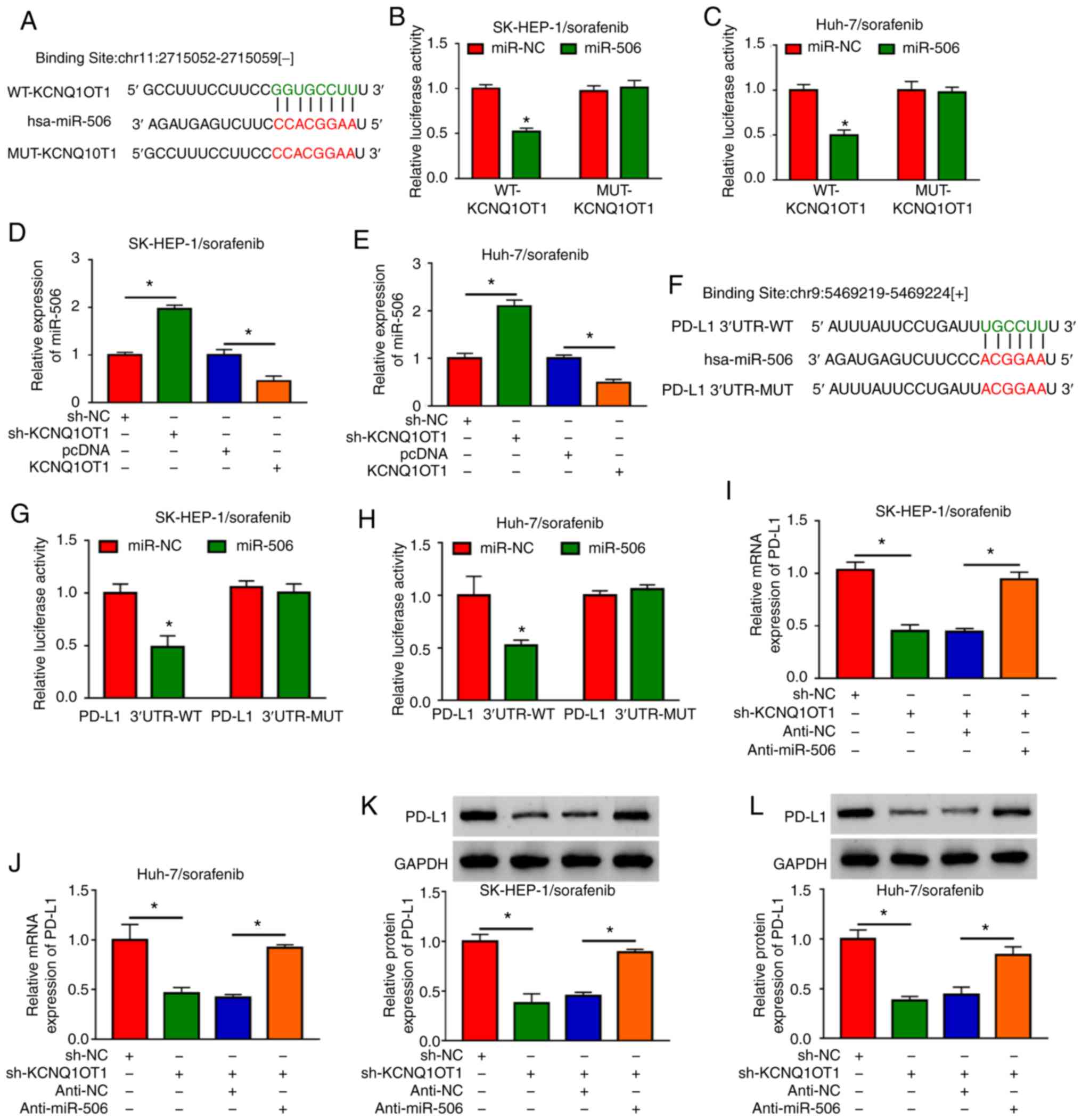 | Figure 5KCNQ1OT1 regulated PD-L1 expression
via sponging miR-506 in sorafenib-resistant HCC cells. (A) The
putative binding sites between KCNQ1OT1 and miR-506 were predicted
by starBase 3.0. (B and C) The luciferase activity in miR-506 or
miR-NC and WT-KCNQ1OT1 or MUT-KCNQ1OT1 co-transfected
SK-HEP-1/sorafenib and Huh-7/sorafenib cells was determined by
dual-luciferase reporter assay. (D and E) The expression level of
miR-506 in sh-KCNQ1OT1, sh-NC, KCNQ1OT1 or pcDNA-transfected
SK-HEP-1/sorafenib and Huh-7/sorafenib cells was detected by
RT-qPCR. (F) Complementary sequences between miR-506 and PD-Ll. (G
and H) The luciferase activity in miR-506 or miR-NC and PD-L1
3′UTR-MUT or PD-L1 3′UTR-WT co-transfected SK-HEP-1/sorafenib and
Huh-7/sorafenib cells was determined by dual-luciferase reporter
assay. (I-L) The mRNA and protein expression levels of PD-L1 in
sh-KCNQ1OT1, sh-NC, sh-KCNQ1OT1+anti-miR-506 or sh-KCNQ1OT1+anti-NC
transfected SK-HEP-1/sorafenib and Huh-7/sorafenib cells were
detected by RT-qPCR and western blotting, respectively.
*P<0.05. HCC, hepatocellular carcinoma; KCNQ1OT1,
KCNQ1 overlapping transcript 1; PD-L1, programmed death-ligand-1;
RT-qPCR, reverse transcription-quantitative PCR analysis; WT,
wild-type; MUT, mutant; UT, untranslated region. |
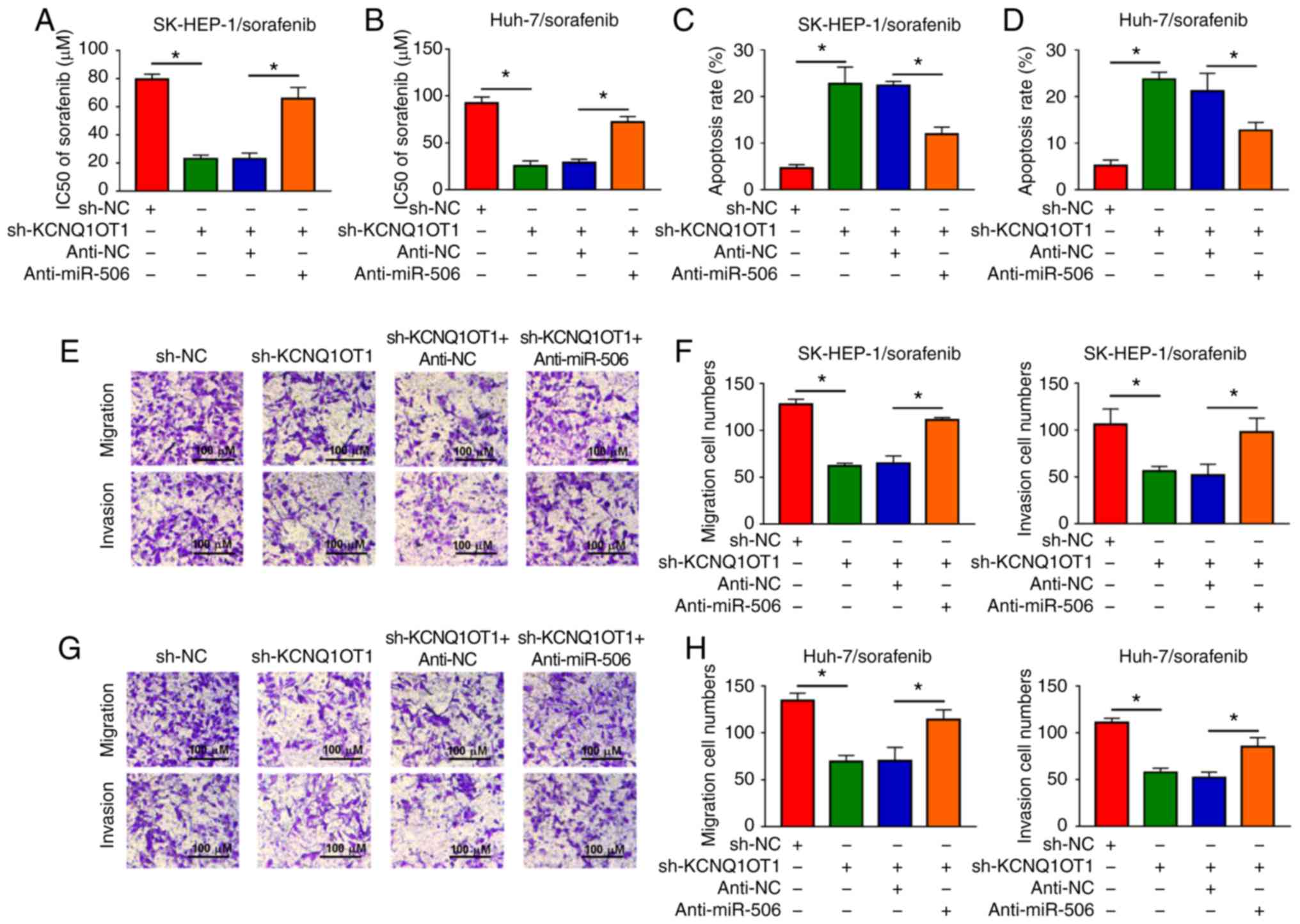
Inhibition of miR-506 partially reverses
the effects on sorafenib sensitivity, cell apoptosis, cell
migration and cell invasion mediated by KCNQ1OT1 knockdown in
sorafenib-resistant HCC cells
Based on the results mentioned above, it was further
investigated whether KCNQ1OT1 affects sorafenib sensitivity, cell
apoptosis, migration and invasion by targeting miR-506 in
sorafenib-resistant HCC cells. It was observed that the reduced
IC50 value caused by sh-KCNQ1OT1 was partly abolished
following anti-miR-506 transfection in SK-HEP-1/sorafenib and
Huh-7/sorafenib cells (Fig. 6A and
B). Flow cytometry revealed that downregulation of miR-506
inhibited the apoptosis-promoting effect of KCNQ1OT1 knockdown in
both SK-HEP-1/sorafenib and Huh-7/sorafenib cells (Fig. 6C and D). In addition, the
migration and invasion of SK-HEP-1/sorafenib and Huh-7/sorafenib
cells were suppressed by knocking down the expression of KCNQ1OT1,
while these effects were partially reversed by the depletion of
miR-506, as shown by the Transwell assay (Fig. 6E-H). These results demonstrated
that KCNQ1OT1 affected sorafenib sensitivity, HCC cell apoptosis,
migration and invasion via regulating the expression of
miR-506.
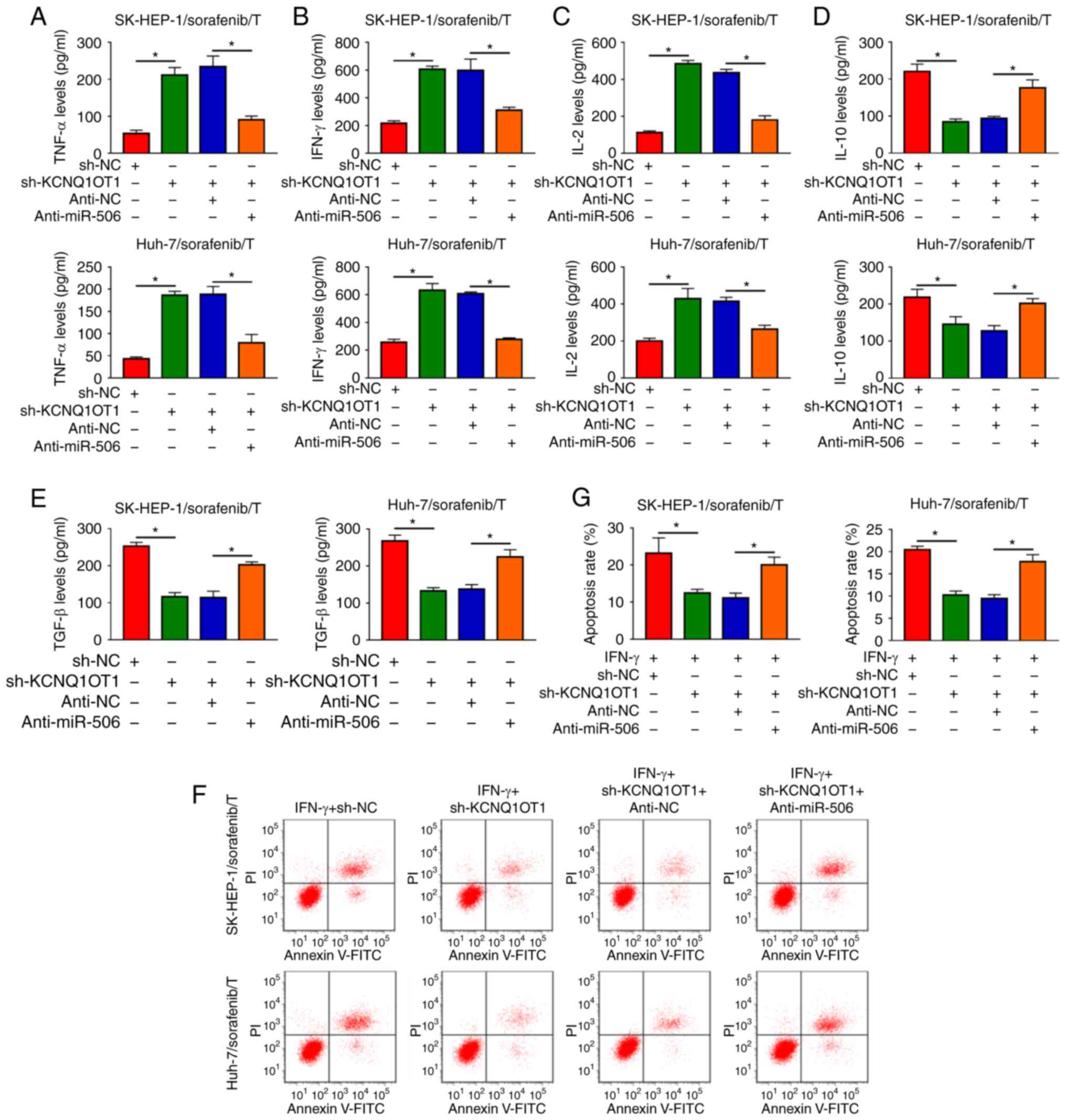
KCNQ1OT1 affects the production of
cytokines and the apoptosis of T cells by sponging miR-506 in a
sorafenib-resistant HCC/T-cell co-culture model
It was next investigated whether KCNQ1OT1 could
regulate the immune escape of sorafenib-resistant HCC cells. The
data of ELISA revealed that the inhibitory effects of KCNQ1OT1
knockdown on TNF-α, IFN-γ and IL-2 levels and the promoting effects
on IL-10 and TGF-β levels were all partly abolished by miR-506
inhibition in SK-HEP-1/sorafenib/T and Huh-7/sorafenib/T models
(Fig. 7A-E). Flow-cytometric
analysis revealed that inhibition of miR-506 attenuated the
suppression of KCNQ1OT1 knockdown on CD8+ T-cell
apoptosis in both SK-HEP-1/sorafenib/T and Huh-7/sorafenib/T models
(Fig. 7F and G). Collectively,
the data mentioned above indicate that KCNQ1OT1 regulated the
production of cytokines and the apoptosis of CD8+ T
cells by targeting miR-506 in a sorafenib-resistant HCC/T-cell
co-culture model.
Discussion
Sorafenib is a molecular-targeted drug that is
useful for the treatment of patients with HCC. However, patients
may develop sorafenib resistance during the treatment process, for
which there is currently no effective solution (5). Immune evasion from T-cell
surveillance was confirmed to play a key role in tumor development
(28). A growing body of
literature indicates that lncRNAs are associated with drug
resistance and immune escape in human cancers (29). The present study focused on the
role and mechanism of action of KCNQ1OT1 in sorafenib resistance
and immune escape in HCC. It was observed that the expression of
KCNQ1OT1 was markedly increased in sorafenib-resistant HCC tissues
and cells. Furthermore, the KCNQ1OT1/miR-506 axis was shown to play
a key role in the sensitivity of HCC cells to sorafenib and the
apoptosis of T cells mediated by PD-L1.
KCNQ1OT1 has been found to be aberrantly expressed
in drug-resistant tumors and to play crucial roles in the
regulation of drug resistance. For example, Xian et al
reported that KCNQ1OT1 was increased in methotrexate
(MTX)-resistant colorectal cancer (CRC) cells, and demonstrated
that KCNQ1OT1 knockdown reduced MTX resistance of CRC cells
(30). Hu et al suggested
that KCNQ1OT1 was highly expressed in oxaliplatin-resistant HCC
tissues and cells, and oxaliplatin-resistant HCC cell metastasis
was suppressed by knocking down KCNQ1OT1 expression (31). Consistently with these findings,
the present study demonstrated that the KCNQ1OT1 level was markedly
higher in sorafenib-resistant HCC tissues and cells in comparison
with that in sorafenib-sensitive HCC tissues and cells. KCNQ1OT1
knockdown partly reversed sorafenib resistance in
sorafenib-resistant HCC cells. Apoptosis is a tightly coordinated
cellular process, and dysregulation of the apoptotic pathway alters
the course of carcinogenesis; therefore, it appears reasonable for
apoptosis to be considered as a strategy of targeted treatment in
cancer (32). The cell migration
and invasion abilities are essential regulators in tumors and may
provide new targets for cancer therapy (33). Thus, the effects of KCNQ1OT1 on
HCC cell apoptosis and motility were investigated in the present
study. The results demonstrated that KCNQ1OT1 downregulation
promoted apoptosis and inhibited the metastasis of
sorafenib-resistant HCC cells. These data demonstrated that
KCNQ1OT1 knockdown improved sorafenib sensitivity in
sorafenib-resistant HCC cells.
LncRNAs were reported to regulate T-cell apoptosis
and induce immune escape in tumors (34). It was previously demonstrated that
immune escape can accelerate the occurrence and development of HCC,
and T-cell apoptosis is an important mechanism through which tumors
achieve immune escape (35).
CD8+ T-cell exhaustion is commonly observed in the tumor
microenvironment (36).
Furthermore, the levels of cytokines are associated with T-cell
immunity (37,38). Thus, the levels of cytokines
produced by T cells and the apoptosis of CD8+ T cells
were investigated in a sorafenib-resistant HCC/T-cell co-culture
model. The data revealed that TNF-α, IFN-γ and IL-2 were increased,
whereas IL-10 and TGF-β were decreased following KCNQ1OT1 knockdown
in SK-HEP-1/sorafenib/T and Huh-7/sorafenib/T media. In addition,
KCNQ1OT1 knockdown reduced the apoptosis of CD8+T cells
in the sorafenib-resistant HCC/T-cell co-culture model. These data
indicated that the immune surveillance ability of T cells on
sorafenib-resistant HCC cells was enhanced following KCNQ1OT1
knockdown.
miR-506 was identified as a target of KCNQ1OT1, and
an inverse correlation was observed between KCNQ1OT1 and miR-506
expression in sorafenib-resistant HCC tissues. Accumulating
evidence has confirmed that miR-506 is weakly expressed in
drug-resistant tumors, and miR-506 upregulation enhanced the drug
sensitivity of tumor cells (20,21). In line with these data, miR-506
was found to be markedly decreased in sorafenib-resistant HCC
tissues and cells, whereas miR-506 inhibition abolished the effects
of KCNQ1OT1 downregulation on sorafenib resistance, apoptosis and
metastasis in sorafenib-resistant HCC cells, demonstrating that
miR-506 reduced the resistance of sorafenib-resistant HCC cells to
sorafenib. Moreover, the secretion of cytokines and the apoptosis
of T cells mediated by KCNQ1OT1 knockdown were reversed by miR-506
inhibition, suggesting that miR-506 inhibition weakens the immune
surveillance of T cells on sorafenib-resistant HCC cells.
PD-L1 was verified to be a target gene of miR-506 in
sorafenib-resistant HCC cells. Bishop et al found that PD-L1
was upregulated in enzalutamide-resistant prostate cancer cells
(39). Liu et al
demonstrated that PD-L1 was upregulated in sorafenib-resistant HCC
cells (40). In the present
study, PD-L1 was notably upregulated in sorafenib-resistant HCC
tissues and cells. Moreover, PD-L1 inhibited the activation of T
cells, playing a crucial role in the immune escape of tumors
(41). However, the exact role of
PD-L1 in sorafenib resistance and immune escape of
sorafenib-resistant HCC cells requires further investigation.
Based on these data, it was inferred that KCNQ1OT1
knockdown enhances sorafenib sensitivity, induces apoptosis and
inhibits the metastasis of sorafenib-resistant HCC cells via
targeting miR-506. Moreover, the KCNQ1OT1/miR-506 axis may regulate
PD-L1-mediated immune escape of sorafenib-resistant HCC cells.
These findings may improve our understanding of the molecular
mechanism underlying the role of KCNQ1OT1 in sorafenib resistance
in HCC, and may help with designing methods to overcome it.
However, in vivo animal experiments must be further
conducted to verify the results of the present study.
Supplementary Data
Funding
No funding was received.
Availability of data and materials
The datasets generated and/or analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
Conceptualization and methodology: XZ and XM; formal
analysis and data curation: ZY and MH; validation and
investigation: JZ and XM; original draft preparation, review and
editing: JZ, XZ and XM. All the authors have read and approved the
final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics Review
Committee of The Sanquan College of Xinxiang Medical
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Bruix J and Sherman M: American
Association for the Study of Liver Diseases Management of
hepatocellular carcinoma: An update. Hepatology. 53:1020–1022.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cidon EU: Systemic treatment of
hepatocellular carcinoma: Past, present and future. World J
Hepatol. 9:797–807. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hollebecque A, Malka D, Ferté C, Ducreux M
and Boige V: Systemic treatment of advanced hepatocellular
carcinoma: From disillusions to new horizons. Eur J Cancer.
51:327–339. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hsueh KC, Lee TY, Kor CT, Chen TM, Chang
TM, Yang SF and Hsieh CB: The role of liver transplantation or
resection for patients with early hepatocellular carcinoma. Tumour
Biol. 37:4193–4201. 2016. View Article : Google Scholar
|
|
5
|
Colagrande S, Regini F, Taliani GG, Nardi
C and Inghilesi AL: Advanced hepatocellular carcinoma and
sorafenib: Diagnosis, indications, clinical and radiological
follow-up. World J Hepatol. 7:1041–1053. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chow AK, Ng L, Lam CS, Wong SK, Wan TM,
Cheng NS, Yau TC, Poon RT and Pang RW: The Enhanced metastatic
potential of hepatocellular carcinoma (HCC) cells with sorafenib
resistance. PLoS One. 8:e786752013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu B, Cao Y, Jiang H and Mao A: Autophagy
facilitates the sorafenib resistance of hepatocellular carcinoma
cells. West Indian Med J. 62:698–700. 2013.
|
|
8
|
Ruiz de Galarreta M, Bresnahan E,
Molina-Sánchez P, Lindblad KE, Maier B, Sia D, Puigvehi M, Miguela
V, Casanova-Acebes M, Dhainaut M, et al: β-catenin activation
promotes immune escape and resistance to anti-PD-1 therapy in
hepatocellular carcinoma. Cancer Discov. 9:1124–1141. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang G, Lu X and Yuan L: LncRNA: A link
between RNA and cancer. Biochim Biophys Acta. 1839:1097–1109. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fatica A and Bozzoni I: Long non-coding
RNAs: New players in cell differentiation and development. Nat Rev
Genet. 15:7–21. 2014. View
Article : Google Scholar
|
|
11
|
Ponting CP, Oliver PL and Reik W:
Evolution and functions of long noncoding RNAs. Cell. 136:629–641.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wapinski O and Chang HY: Long noncoding
RNAs and human disease. Trends Cell Biol. 21:354–361. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang S, Ma H, Zhang D, Xie S, Wang W, Li
Q, Lin Z and Wang Y: LncRNA KCNQ1OT1 regulates proliferation and
cisplatin resistance in tongue cancer via miR-211-5p mediated
Ezrin/Fak/Src signaling. Cell Death Dis. 9:7422018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ren K, Xu R, Huang J, Zhao J and Shi W:
Knockdown of long non-coding RNA KCNQ1OT1 depressed chemoresistance
to paclitaxel in lung adenocarcinoma. Cancer Chemother Pharmacol.
80:243–250. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang P, Ning S, Zhang Y, Li R, Ye J, Zhao
Z, Zhi H, Wang T, Guo Z and Li X: Identification of
lncRNA-associated competing triplets reveals global patterns and
prognostic markers for cancer. Nucleic Acids Res. 43:3478–3489.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gu S, Jin L, Zhang F, Sarnow P and Kay MA:
Biological basis for restriction of microRNA targets to the 3′
untranslated region in mammalian mRNAs. Nat Struct Mol Biol.
16:144–150. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang M, Xie X, Song X, Gu S, Chang X, Su
T, Liang B and Huang D: MiR-506 suppresses colorectal cancer
development by inhibiting orphan nuclear receptor NR4A1 expression.
J Cancer. 10:3560–3570. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wen SY, Lin Y, Yu YQ, Cao SJ, Zhang R,
Yang XM, Li J, Zhang YL, Wang YH, Ma MZ, et al: miR-506 acts as a
tumor suppressor by directly targeting the hedgehog pathway
transcription factor Gli3 in human cervical cancer. Oncogene.
34:717–725. 2015. View Article : Google Scholar
|
|
19
|
Sun Y, Hu L, Zheng H, Bagnoli M, Guo Y,
Rupaimoole R, Rodriguez-Aguayo C, Lopez-Berestein G, Ji P, Chen K,
et al: MiR-506 inhibits multiple targets in the
epithelial-to-mesenchymal transition network and is associated with
good prognosis in epithelial ovarian cancer. J Pathol. 235:25–36.
2015. View Article : Google Scholar
|
|
20
|
Zhou H, Lin C, Zhang Y, Zhang X, Zhang C,
Zhang P, Xie X and Ren Z: miR-506 enhances the sensitivity of human
colorectal cancer cells to oxaliplatin by suppressing MDR 1/P-gp
expression. Cell Prolif. 50:e123412017. View Article : Google Scholar
|
|
21
|
Wang Z, Dai J, Yan J, Zhang Y and Yin Z:
Targeting EZH 2 as a novel therapeutic strategy for
sorafenib-resistant thyroid carcinoma. J Cell Mol Med.
23:4770–4778. 2019.PubMed/NCBI
|
|
22
|
Svoronos AA, Engelman DM and Slack FJ:
OncomiR or tumor suppressor? The duplicity of microRNAs in cancer.
Cancer Res. 76:3666–3670. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Topalian SL, Drake CG and Pardoll DM:
Targeting the PD-1/B7-H1 (PD-L1) pathway to activate anti-tumor
immunity. Curr Opin Immunol. 24:207–212. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang Y and Wang L: miR-34a attenuates
glioma cells progression and chemoresistance via targeting PD-L1.
Biotechnol Lett. 39:1485–1492. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen L, Gibbons DL, Goswami S, Cortez MA,
Ahn YH, Byers LA, Zhang X, Yi X, Dwyer D, Lin W, et al: Metastasis
is regulated via microRNA-200/ZEB1 axis control of tumour cell
PD-L1 expression and intratumoral immunosuppression. Nat Commun.
5:52412014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Llovet JM, Ricci S, Mazzaferro V, Hilgard
P, Gane E, Blanc JF, de Oliveira AC, Santoro A, Raoul JL, Forner A,
et al: Sorafenib in advanced hepatocellular carcinoma. N Engl J
Med. 359:378–390. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
28
|
Igney FH and Krammer PH: Immune escape of
tumors: Apoptosis resistance and tumor counterattack. J Leukoc
Biol. 71:907–920. 2002.PubMed/NCBI
|
|
29
|
Chen QN, Wei CC, Wang ZX and Sun M: Long
non-coding RNAs in anti-cancer drug resistance. Oncotarget.
8:1925–1936. 2017. View Article : Google Scholar :
|
|
30
|
Xian D and Zhao Y: LncRNA KCNQ1OT1
enhanced the methotrexate resistance of colorectal cancer cells by
regulating miR-760/PPP1R1B via the cAMP signalling pathway. J Cell
Mol Med. 23:3808–3823. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hu H, Yang L, Li L and Zeng C: Long
non-coding RNA KCNQ1OT1 modulates oxaliplatin resistance in
hepatocellular carcinoma through miR-7-5p/ABCC1 axis. Biochem
Biophys Res Commun. 503:2400–2406. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wong RS: Apoptosis in cancer: From
pathogenesis to treatment. J Exp Clin Cancer Res. 30:872011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Eccles SA, Box C and Court W: Cell
migration/invasion assays and their application in cancer drug
discovery. Biotechnol Annu Rev. 11:391–421. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang D, Chen J, Yang L, Ouyang Q, Li J,
Lao L, Zhao J, Liu J, Lu Y, Xing Y, et al: NKILA lncRNA promotes
tumor immune evasion by sensitizing T cells to activation-induced
cell death. Nat Immunol. 19:1112–1125. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ghanem I, Riveiro ME, Paradis V, Faivre S,
de Parga PM and Raymond E: Insights on the CXCL12-CXCR4 axis in
hepatocellular carcinoma carcinogenesis. Am J Transl Res.
6:340–352. 2014.PubMed/NCBI
|
|
36
|
Li KK and Adams DH: Antitumor CD8+ T cells
in hepatocellular carcinoma: Present but exhausted. Hepatology.
59:1232–1234. 2014. View Article : Google Scholar
|
|
37
|
Barrera L, Montes-Servín E, Barrera A,
Ramírez-Tirado LA, Salinas-Parra F, Bañales-Méndez JL,
Sandoval-Ríos M and Arrieta Ó: Cytokine profile determined by
data-mining analysis set into clusters of non-small-cell lung
cancer patients according to prognosis. Ann Oncol. 26:428–435.
2015. View Article : Google Scholar
|
|
38
|
Garris CS, Arlauckas SP, Kohler RH, Trefny
MP, Garren S, Piot C, Engblom C, Pfirschke C, Siwicki M,
Gungabeesoon J, et al: Successful anti-PD-1 cancer immunotherapy
requires T cell-dendritic cell crosstalk involving the cytokines
IFN-γ and IL-12. Immunity. 49:1148–1161. 2018. View Article : Google Scholar
|
|
39
|
Bishop JL, Sio A, Angeles A, Roberts ME,
Azad AA, Chi KN and Zoubeidi A: PD-L1 is highly expressed in
Enzalutamide resistant prostate cancer. Oncotarget. 6:234–242.
2015. View Article : Google Scholar :
|
|
40
|
Liu J, Liu Y, Meng L, Liu K and Ji B:
Targeting the PD-L1/DNMT1 axis in acquired resistance to sorafenib
in human hepatocellular carcinoma. Oncol Rep. 38:899–907. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Topalian SL, Hodi FS, Brahmer JR,
Gettinger SN, Smith DC, McDermott DF, Powderly JD, Carvajal RD,
Sosman JA, Atkins MB, et al: Safety, activity, and immune
correlates of anti-PD-1 antibody in cancer. . N Engl J Med.
366:2443–2454. 2012. View Article : Google Scholar : PubMed/NCBI
|