Introduction
Chronic kidney disease (CKD), which is defined by
the estimated glomerular filtration rate (eGFR) and/or albuminuria,
is recognized as a global health issue, the incidence, morbidity
and mortality rates of which are increasing over the past two
decades, and affects 10-16% of adults worldwide (1). Several patients with CKD also suffer
from cardiovascular complications, which are major contributors to
mortality in patients with late-stage CKD (2). Therefore, timely prevention,
diagnosis and treatment of CKD, including the treatment of medical
conditions and diseases that lead to CKD, are critical (1). Increased vascular calcification (VC)
in patients with CKD resulted in poor overall survival as well as
cardiovascular mortality and morbidity (3). In addition, hyperphosphatemia, (high
levels of phosphorus in the blood), may severely damage the
kidneys, bone, and heart (4). As
calcium is leached from bones in hyperphosphatemia, it can result
in weakened bones as well as calcium deposition in other tissues.
In particular, calcium deposition in the vasculature may increase
the risk of VC and CKD (4).
As a hormone that is primarily secreted by
osteocytes, fibroblast growth factor 23 (FGF23) regulates the
metabolism of vitamin D and phosphate. In particular, the serum
levels of phosphorus are regulated by FGF23, which also regulates
the production and secretion of parathyroid hormone (5). A previous study reported that both
Klotho and canonical FGF receptors are required during FGF23
signalling (6). In addition, the
serum levels of FGF23 gradually increases as kidney function
declines (7). For example, the
serum levels of FGF23 are only 2-5 times the physiological levels
during the early stages of CKD, but may be >200 times the normal
levels during the later stages of CKD (8). In addition, clinical studies have
shown a positive correlation between the concentration of FGF23 and
the levels of VC, cardiovascular risk and mortality in patients
with CKD (8,9).
It has been shown that that the expression of both
extracellular signal-related kinase (ERK) and AKT are increased by
FGF23 (10,11). Furthermore, the activation of ERK
accelerates VC, whereas the activation of both AKT and ERK inhibits
VC (12,13). In addition, FGF23 activates ERK
1/2 in the parathyroid, kidney and Klotho-expressing cells
(6). It is hypothesized that
Klotho regulates FGF23 signalling through FGFR1 activation, which
in-turn increases the phosphorylation of ERK1/2 and the
transcription of the early growth response-1 gene in
Klotho-expressing tissues, such as the parathyroid gland, pituitary
gland and kidney (10). However,
tissues which do not express Klotho are not affected by FGF23 (for
example, skeletal muscle, adrenal gland, thymus, small intestine,
stomach, spleen, lung, heart and liver) (6).
In the present study, an animal model of renal
failure was established, which was then treated with a phosphorus
lowering agent to evaluate its effect on VC. Furthermore, the
effects of different phosphorus reducing treatments in an animal
model of VC were assessed.
Materials and methods
Animal experiments
Male Wistar rats aged 12 weeks (72 rats, with an
average weight of 400 g) were used in the present study; 24 rats
were continuously fed the same diet and were used as non-uremic
controls (Adenine-free group), and the remaining 48 rats received
0.75% adenine in addition to the same diet. All rats were treated
for 4 weeks to initiate uraemia.
After oral administration and metabolism of adenine,
it becomes 2,8-dihydroxyadenine, which can form tubular crystals
and damage renal tissues (11).
The 48 rats in the two treatment groups were administered either
vehicle (Adenine-vehicle group, n=24) or lanthanum
(Adenine-lanthanum group, n=24) within 24 h of application of
adenine. Lanthanum was dissolved in a solution of 0.85% NaCl, 10 mM
succinic acid and 0.9% benzyl alcohol (pH 4.5), and was
subsequently given via subcutaneous (SC) injection at a dose of 0.3
mg/kg. The SC injection was given once a day for a total of 4
weeks. Furthermore, the 24 rats on the adenine diet were
administered empty vehicles by SC once every day for a total of 4
weeks. All animal studies were performed in strict compliance with
the Guide for the Care and Use of Laboratory Animals (14) and ethical approval was granted by
the Ethics Committee of Dongguan People's Hospital Affiliated to
Southern Medical University. No rats were lost during the study.
The animals were sacrificed by a lethal intravenous dose of sodium
pentobarbital (100 mg/kg) after completion of the experiment. Death
was confirmed by lack of a pulse, breathing, corneal reflex,
response to toe pinch and a lack of respiratory sounds and
heartbeat. In addition, percutaneous cardiac puncture can be
performed after the animal is unconscious. If the needle and
attached syringe do not move after insertion into the heart
(aspiration of blood provides evidence of correct location), this
suggests indicates lack of cardiac muscle movement and thus,
death.
Serum and plasma analysis
A final blood sample of 2 ml was obtained from the
left jugular vein of each subject. The plasma levels of PTH were
measured using a PTH ELISA kit (cat. no. 60-2305; Immunotopics,
Inc.). In addition, serum levels of FGF23 and PTH were measured
using a PTH-FGF23 magnetic bead assay (EMD Millipore) according to
the manufacturer's protocol. The serum levels of creatinine were
measured using a creatinine enzymatic assay (cat. no. c7548; Pointe
Scientific, Inc.). The levels of total calcium, blood urea nitrogen
(BUN), and phosphorus were measured using a blood chemistry
analyser (Olympus AU 400; Olympus Corporation).
Histology of the kidneys
After 30 days post-operation, the animals were
anaesthetized using a mixture of ketamine (35 mg/kg), xylazine (5
mg/kg), and acepromazine (1 mg/kg) and the kidney cortex tissue
samples were removed and fixed in 10% neutral buffered formalin
(NBF) for 48 h at room temperature, embedded in paraffin, sliced
into 4 µm sections, and stained with haematoxylin and eosin.
Animals were sacrificed using a lethal intravenous dose of sodium
pentobarbital (100 mg/kg). Kidney tubulointerstitial damage was
evaluated semi-quantitatively under a light microscope, and a score
of 0-5 was assigned to each sample based on the magnitude of
tubulointerstitial damage in the renal cortex: 0, normal; grade 1,
<10%; grade 2, 10-25%; grade 3, 25-50%; grade 4, 50-75%; and
grade 5, 75-100%.
Histology of aortic tissues
After 30 days post-operation, the animals were
anaesthetized and the aortic arch containing 1 cm of descending
aortic tissue was harvested from each animal and fixed in 10% NBF.
Subsequently, paraffin-embedded longitudinal sections (4-µm
in thickness) were prepared and subjected to von Kossa staining
(15) to assess the condition of
calcified lesions. Semi-quantitative histological scoring of aortic
wall mineralization was performed based on the following criteria:
Grade 1, minimal = detectible patchy focal vascular mineralization
of the tunica media; grade 2, mild=<10% of the entire aortic
sample was mineralized; grade 3, moderate=10-50% of the entire
aortic sample was mineralized; and grade 4, significant >50% of
the entire sample was mineralized.
Human subjects
A total of 48 subjects with a median age of 52.8
years (range, 46-50 years) were enrolled in the present study and
observed for 24 weeks. All subjects were aged 18 years or older,
and the inclusion criteria were as follows: i) Subjects have been
treated with haemodialysis for ≥3 months; ii) plasma concentration
of intact PTH (iPTH) >300 pg/ml; iii) plasma concentration of
bio-iPTH (biPTH) >160 pg/ml; iv) in the case of iPTH and biPTH
concentrations of 150-300 and 80-160 pg/ml, respectively, the
albumin-corrected value for the calcium-phosphorus product was
>50 mg2/dl2 during the treatment with
vitamin D sterols; and v) albumin-corrected serum calcium ≥8.4
mg/dl. The exclusion criteria were as follows: i) Subjects that had
been previously treated with lanthanum carbonate or calcium-free
phosphate-binding substances, including aluminium hydroxide,
lanthanum carbonate, or sevelamer hydrochloride within 30 days
prior to enrolment; ii) subjects that were previously treated with
bisphosphonates, 3-hydroxy-3-methylglutaryl-coenzyme A reductase
inhibitors and other cholesterol reducing drugs within 30 days
prior to enrolment; iii) subjects with a thoracic aortic aneurysm,
cardiac transplantation, aortic/cardiac valve replacement, coronary
artery stent, coronary artery bypass grafting, or active atrial
fibrillation; iv) subjects who had undergone parathyroidectomy
within the past 3 months or were anticipating parathyroidectomy in
the following 6 months; v) subjects scheduled for kidney
transplantation; vi) subjects with digestive disorders that may
affect their ability to receive oral medications; and vii) subjects
with known allergies to lanthanum carbonate. The present study was
approved by the Institutional Ethics Review Committee of Dongguan
People's Hospital Affiliated to Southern Medical University. The
characteristics of all participants are presented in Table I. Written informed consent was
obtained from each subject prior to the start of this study. This
study was performed in accordance with the principles of the
Declaration of Helsinki (12).
 | Table IPatient characteristics. |
Table I
Patient characteristics.
|
Characteristics | Regular group,
n=40 | Fortified group,
n=40 | P-value |
|---|
| Age, years | 52.5±5.3 | 53.1±6.2 | 0.61 |
| Sex, male, n
(%) | 27 (67.5) | 28 (70.0) | 0.84 |
| CKD etiology, n
(%) | | | 0.81 |
| Diabetes | 5 (12.5) | 6 (15.0) | |
| Hypertension | 5 (12.5) | 5 (12.5) | |
| IgAN | 19 (47.5) | 20 (50.0) | |
| Chronic GN
(excluding IgAN) | 3 (7.5) | 3 (7.5) | |
| Others | 8 (20.0) | 6 (15.0) | |
| Systolic BP,
mmHg | 120.8±10.4 | 125.8±13.5 | 0.73 |
| Diastolic BP,
mmHg | 72.3±3.9 | 74.3±5.6 | 0.64 |
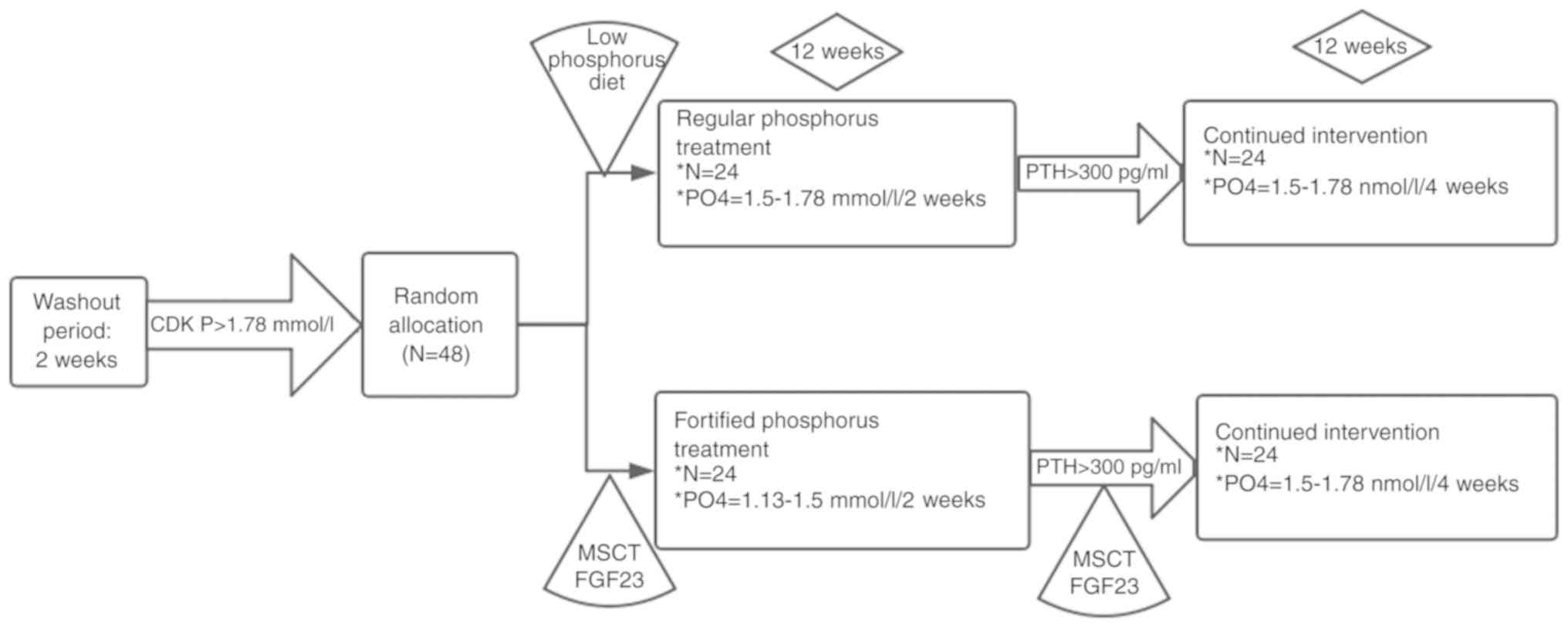
Design of the human study
Subjects were randomly divided into two groups, the
regular phosphorus treatment group (regular group, n=24) and the
fortified phosphorus treatment group (fortified group, n=24). The
subjects in both groups were exclusively treated with lanthanum
carbonate to minimize possible confounding effects caused by other
phosphate-binding drugs. In the regular group, the subjects were
treated with 750 mg/day lanthanum carbonate. In the fortified
group, the dose of lanthanum carbonate started at 750 mg/day, and
increased sequentially to 1,500, 2,250 and 3,000 mg/day in 1-week
intervals. The flow chart of the design of the human study is shown
in Fig. 1. During the study
period, no subjects dropped out of the research. Therefore, all 48
subjects were included for analysis.
Evaluation of human subjects
Serum samples were collected from all subjects, and
the levels of FGF23, calcium, phosphorus and PTH were determined
using a blood chemistry analyser. The status of vascular
calcification was evaluated and scored using multi-slice spiral
computed tomography (MSCT) as described previously (13).
Western blot analysis
To evaluate the effect of FGF23 on the levels of
phospho- (p-)ERK in humans and rats, VSMCs were treated as follows:
i) 5 and 10 ng FGF23 and evaluated at 30 min; ii) 5 ng FGF23 and
evaluated after 0, 15 and 30 min. Tissue and cell samples were
first frozen in liquid nitrogen and then lysed using a lysis buffer
(Vazyme Biotech Co., Ltd.). Subsequently, the samples were
centrifuged at 241.49 x g for 15 min at 4°C to remove debris. The
supernatant from each sample was collected, and its protein
concentration was measured using a bicinchoninic acid kit (Beyotime
Institute of Biotechnology). An appropriate quantity of protein
sample was then mixed with 2X SDS loading buffer and boiled at
100°C for 5 min. Subsequently, 50 µg protein sample was
loaded per lane onto a 10% SDS gel and resolved by SDS-PAGE,
transferred to a PVDF membrane, which was then blocked with 5%
skimmed milk for 1 h at room temperature and incubated overnight at
4°C with diluted (1:100) primary antibodies against total- (t-)ERK
(cat. no. 4695, 1:5,000; Cell Signalling Technology), p-ERK (cat.
no. 4370, 1:5,000; Cell Signalling Technology), Msx2, Osx (cat. no.
sc-393325, 1:5,000; Santa Cruz Biotechnology) or β-actin (cat. no.
3700, 1:5,000; Cell Signalling Technology). Subsequently, the
membrane was washed 3 times with Tris(-HCl)-buffered saline +
Tween-20, 0.1%, and incubated at room temperature for 1 h with a
goat-anti-rabbit horseradish peroxidase-conjugated secondary
antibody (cat. no. ab6721; 1:12,000; Abcam). The membrane was then
soaked in an enhanced chemiluminescence reagent (Biomiga, Inc.).
After the removal of excess liquid, signals were visualized using
X-ray film. The ratio between the mean optical density (OD) value
of the target protein and the mean OD value of β-actin was
calculated as the relative expression of the p-ERK and t-ERK. Each
experiment was performed three times. Densitometry analysis was
performed using Quantity One (version 4.6.6; Bio-Rad Laboratories,
Inc.) software.
Statistical analysis
Statistical analysis was performed using SPSS
version 21.0 (IBM, Corp.). Data are shown as the mean ± standard
deviation. The differences between two groups were compared using a
Student's t-test, whereas the differences among several groups were
compared using a one-way ANOVA, with a post-hoc Tukey's test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Serum levels of creatinine, BUN, FGF23,
calcium, phos- phorus and the plasma levels of PTH were compared
among the different animal groups
As shown in Fig.
2, the initial serum levels of creatinine (Fig. 2A) and BUN (Fig. 2B) were comparable among the
Adenine-free, Adenine-vehicle and Adenine-lanthanum groups. After 2
weeks of adenine administration, the serum levels of creatinine
(Fig. 2A) and BUN (Fig. 2B) in the Adenine-vehicle and
Adenine-lanthanum groups were significantly increased. After 4
weeks of adenine administration, the levels of serum creatinine
(Fig. 2A) and BUN (Fig. 2B) in all rats on the adenine diet
were significantly higher compared with the values after 2 weeks.
Furthermore, the levels of serum creatinine (Fig. 2A) and BUN (Fig. 2B) in the Adenine-lanthanum group
were significantly lower compared with the Adenine-vehicle group,
suggesting that lanthanum treatment alleviated uraemia. As shown in
Fig. 2C, both uremic groups
displayed significantly higher levels of plasma PTH compared with
those of the non-uremic controls in week 4. Furthermore, the
Adenine-lanthanum group exhibited significantly lower levels of
plasma PTH compared with the Adenine-vehicle group in week 4, when
the serum levels of FGF23 in uremic rats was significantly higher
compared with the non-uremic rats (Fig. 2D). In addition, the serum levels
of FGF23 were significantly reduced in the Adenine-lanthanum group
compared with the Adenine-vehicle group, and the serum levels of
total calcium in the Adenine-lanthanum group was significantly
reduced in week 4 (Fig. 2E).
Finally, in both weeks 2 and 4, the serum levels of phosphorus in
uremic rats was significantly higher compared with the non-uremic
rats, although the serum phosphorus levels were the highest in the
Adenine-vehicle group (Fig.
2F).
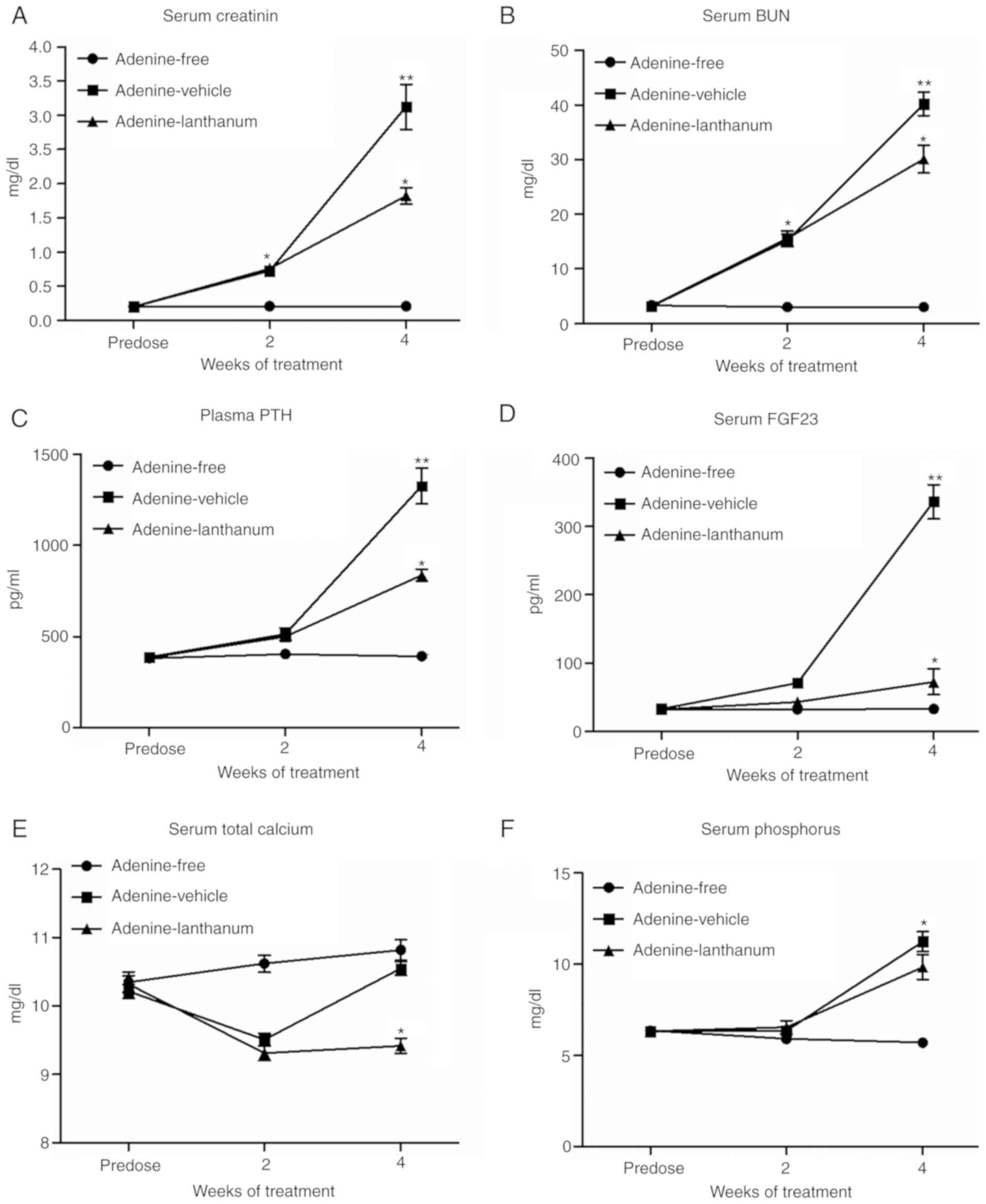 | Figure 2Serum levels of creatinine, BUN,
FGF23, calcium and phosphorus, and the plasma levels of PTH were
compared among the different groups. (A) Serum levels of
creatinine, (B) BUN, (C) FGF23, (D) calcium and (E) phosphorus were
compared among the Adenine-free, Adenine-vehicle and
Adenine-lanthanum groups. (F) Plasma levels of PTH were compared
among the Adenine-free, Adenine-vehicle and Adenine-lanthanum
groups. BUN, blood urea nitrogen; FGF23, fibroblast growth factor
23; PTH, parathyroid hormone. *P<0.05,
**P<0.05 vs. Adenine-lanthanum group. |
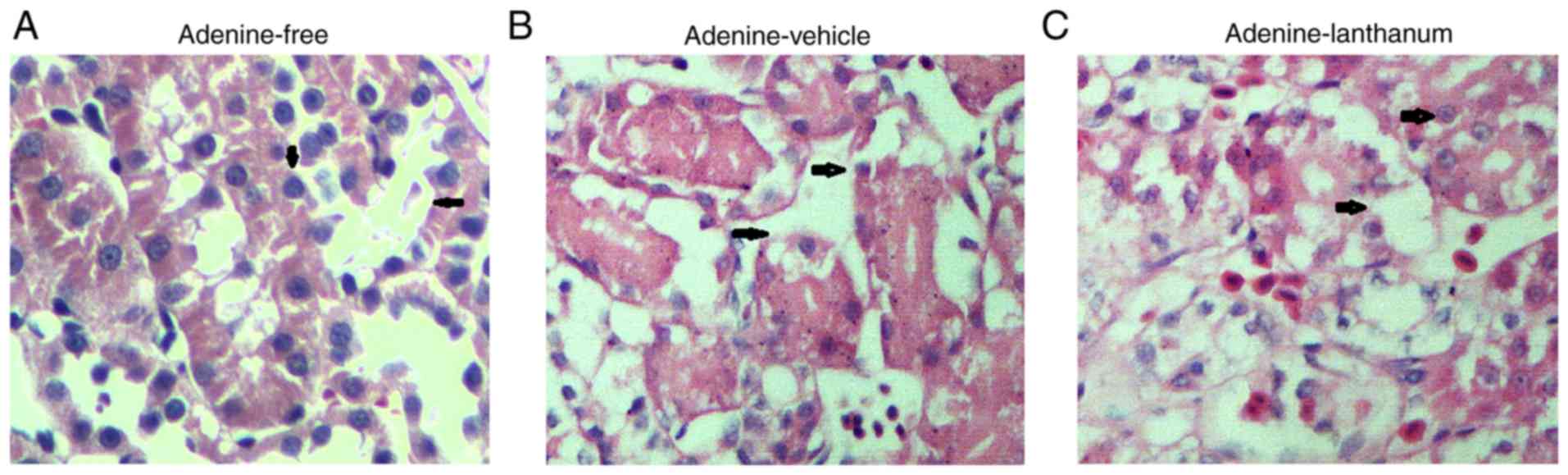
Kidney histology among the four animal
groups
The damage to kidney tissues presented as tubular
necrosis, degeneration, dilation, intratubular cellular debris,
epithelial attenuation, and crystal deposition. The kidney tissues
collected from the rats fed the adenine diet (Fig. 3B and C) exhibited a similar
pattern of renal cortex damage and moderate-to-severe diffuse
tubulointerstitial injuries (Fig.
3A). The interstitium in all groups was expanded and
infiltrated by fibroblasts and inflammatory cells. Treatment with
lanthanum failed to reduce the severity and incidence of kidney
injuries.
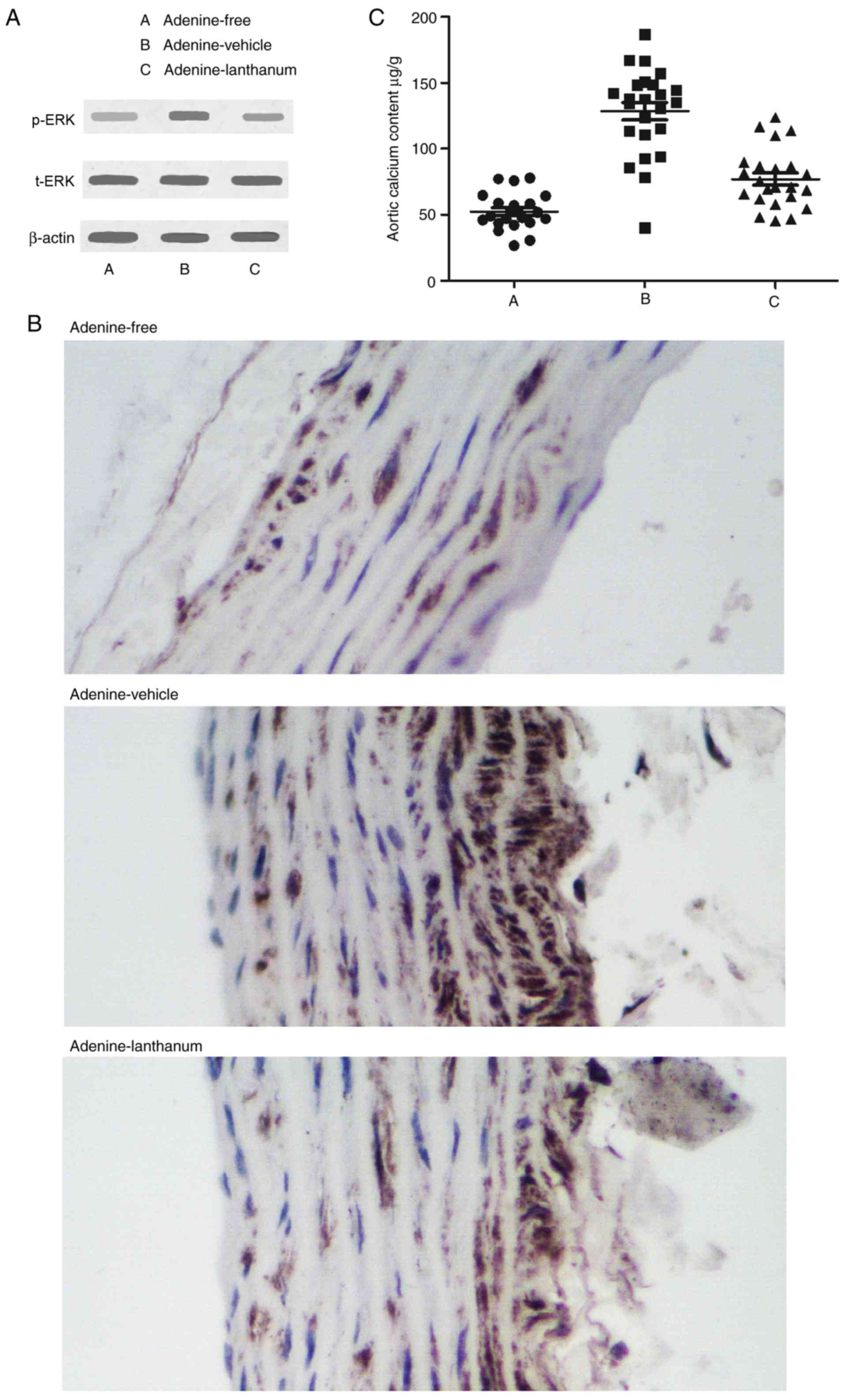
Levels of t-ERK, p-ERK and aortic calcium
among the different animal groups
The protein levels of t-ERK in kidney cortex tissues
collected from the Adenine-free, Adenine-vehicle and
Adenine-lanthanum groups were comparable (Fig. 4A). The levels of p-ERK (Fig. 4A) and aortic calcium (Fig. 4C) in the Adenine-vehicle group
were significantly higher compared with the Adenine-lanthanum
group. In addition, the levels of p-ERK in the Adenine-lanthanum
group was slightly but not significantly higher compared with the
Adenine-free group.
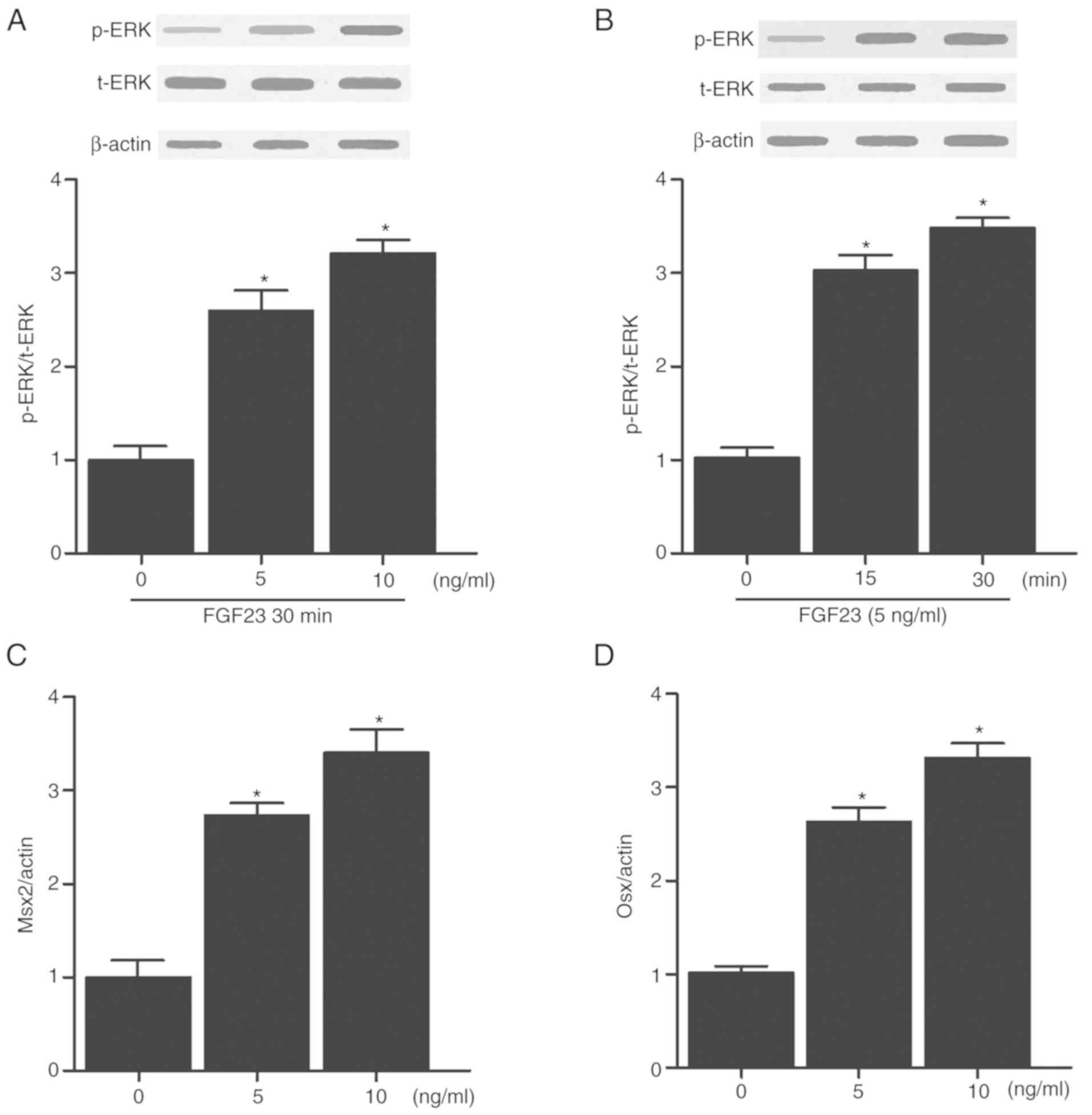
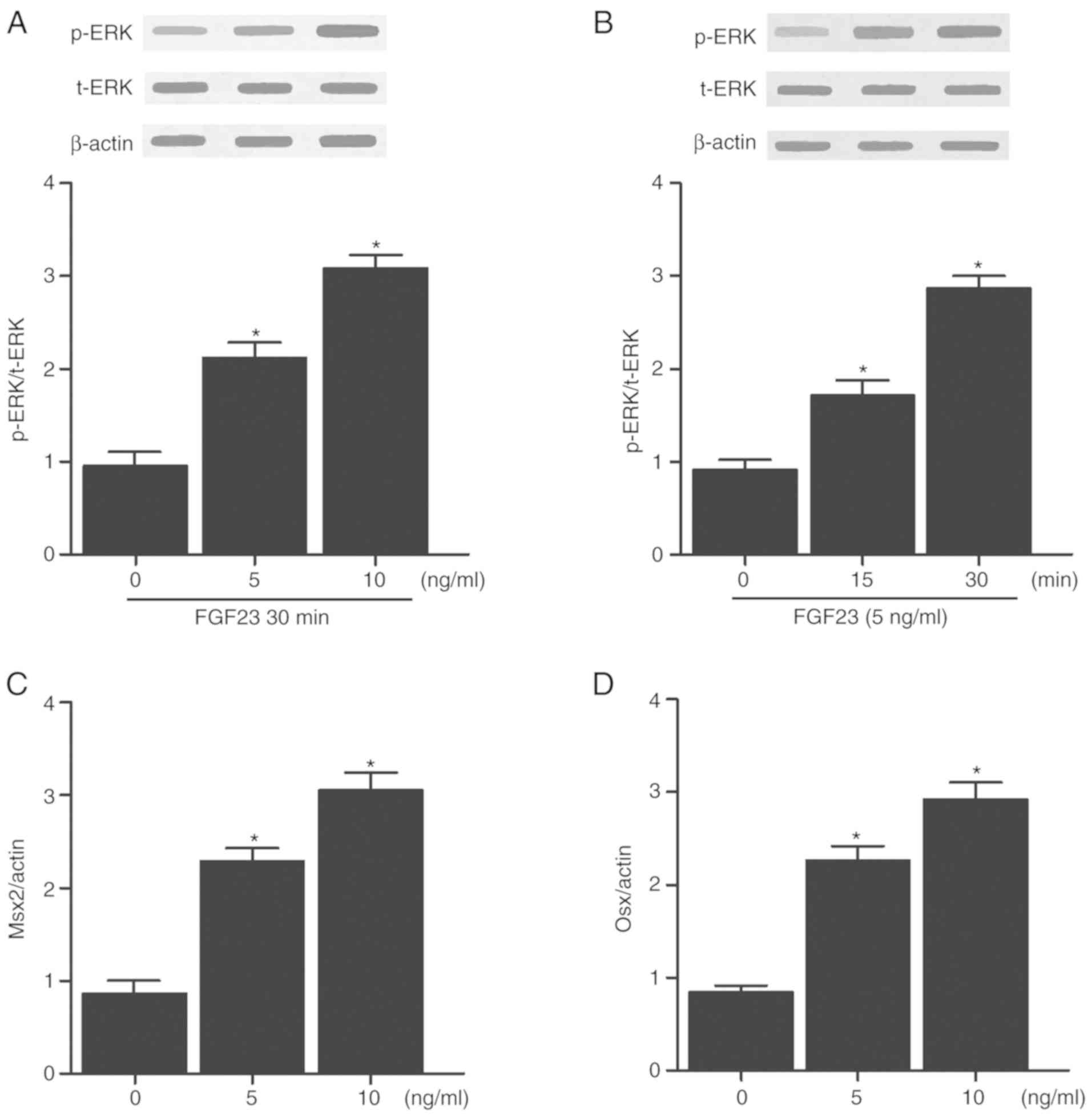
Effect of FGF23 on ERK phosphorylation
and osteoblast differentiation
The effects of FGF23 on ERK phosphorylation in human
and rat vascular smooth muscle cells (VSMCs) were evaluated by
treating the cells with different doses of FGF23 (5 or 10 ng). In
addition, the effects of FGF23 on the levels of p-ERK in human and
rat VSMCs were evaluated at different time points (0, 15 and 30
min) by treating the cells with 5 ng of FGF23. As shown in Figs. 5A and 6A, FGF23 increased the protein
expression levels of p-ERK in a dose-dependent manner in both human
(Fig. 5A) and rat (Fig. 6A) VSMCs. In addition, FGF23
treatment significantly increased the protein expression levels of
p-ERK in both human (Fig. 5B) and
rat (Fig. 6B) VSMCs in a
time-dependent manner. To further determine whether FGF23 affected
the levels of osteoblastic markers, the cells were treated with
different doses of FGF23 and the levels of Msx2 and Osx were
measured. The protein expression levels of Msx2 (Figs. 5C and 6C) and Osx (Figs. 5D and 6D) were increased in both human and rat
VSMCs in a dose-dependent manner.
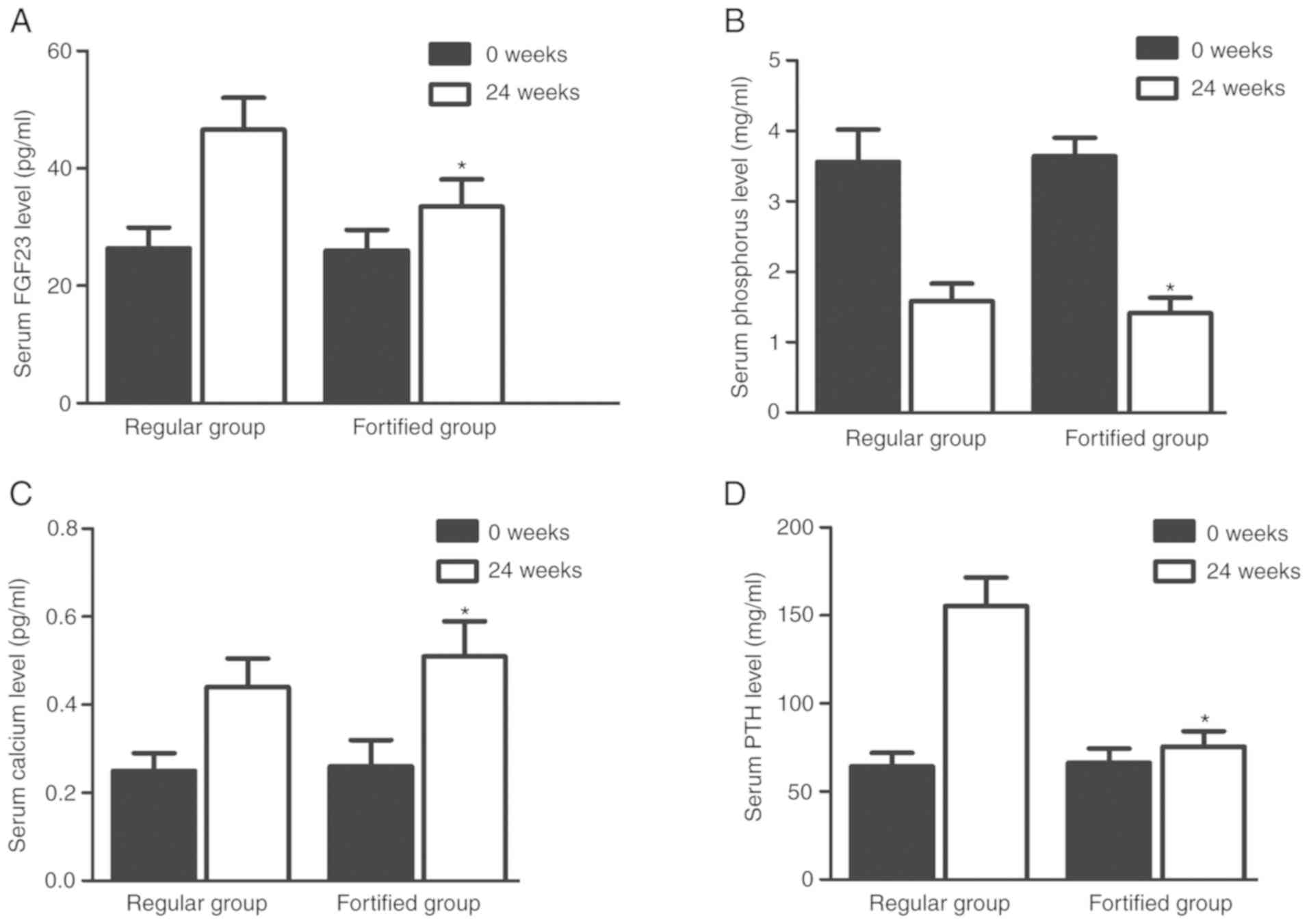
Serum levels of FGF23, calcium,
phosphorus and PTH were different between the two groups
Serum levels of FGF23 (Fig. 7A), calcium (Fig. 7B), phosphorus (Fig. 7C) and PTH (Fig. 7D) in the regular group were
similar to those in the fortified group at the beginning of the
experiment. However, the serum levels of FGF23 (Fig. 7A), phosphorus (Fig. 7B) and PTH (Fig. 7D) in the fortified group were
significantly lower compared with the regular group 24 weeks later,
whereas the serum levels of calcium (Fig. 7C) in the fortified group were
significantly higher compared with the regular group.
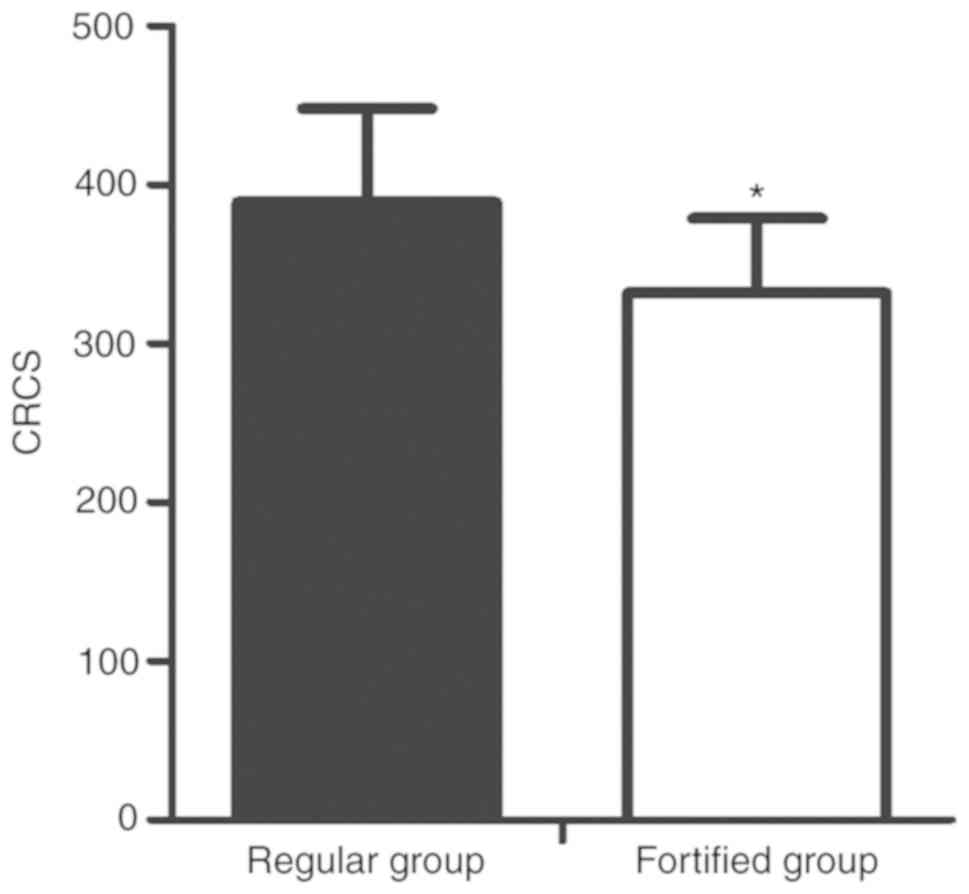
Effect of phosphorus-lowering treatment
on vascular calci- fication
The levels of vascular calcification were evaluated
by MSCT and scored accordingly. The preventative effect of
fortified phosphorus-lowering treatment was significantly greater
compared with the regular phosphorus-lowering treatment (Fig. 8).
Discussion
VC is characterized by arterial wall thickening and
loss of elasticity (16). VC
associated with CKD is an active and complex pathological process.
Inhibitors and promoters of VC are regulated by a wide range of
cellular processes (17).
Although patients with CKD are more likely to suffer from medial
calcification, VC is not a rare occurrence in CKD patients
(17). In particular, as CKD
progresses, the incidence of VC increases due to decreased kidney
functions (18). As VC has
several adverse effects, including ischaemic cardiac diseases, it
poses a significant risk to the health of patients with CKD
(19). During CKD, decreased
phosphorus elimination by renal filtration may result in an
increased phosphorus load during the early phase of CKD, although
symptoms of hyperphosphatemia are usually not apparent until the
later stages (20). In addition,
an increased level of phosphorus during CKD initiates a wide range
of different endocrine responses, such as increased secretion of
FGF23, which causes secondary hyperparathyroidism, cardiovascular
diseases, renal failure, ectopic calcification and metabolic bone
diseases (21). In addition,
hyperphosphatemia results in increased mortality rates among
patients undergoing dialysis (22). However, the restriction on dietary
phosphorus intake is no longer recommended as dietary phosphorus is
positively correlated with the nutritional status of end-stage
renal disease patients, and a reduction in phosphorus intake
benefits survival (23).
Therefore, drugs that are capable of reducing the blood levels of
phosphorus are continuously used as the first-line treatment for
hyperphosphatemia (24).
FGF23 is a 32 kDa protein primarily expressed and
synthesized by osteocytes and osteoblasts (25). It has been shown that elevated
levels of FGF23 results in hereditary hypophosphatemic diseases,
such as autosomal dominant hypophosphatemic rickets, autosomal
recessive hypophosphatemic rickets and X-linked hypophosphatemia,
as well as acquired hypophosphatemic disorders, including
tumour-induced osteomalacia. In addition, inhibition of FGF23
results in hyperphosphatemia and tumoral calcinosis, which
manifests as increased levels of soft tissue calcifications and
1,25(OH)2D (26).
Furthermore, targeting FGFR 1, 3 and 4 receptor complexes,
transmembrane α-Kl and β glucuronidase suppresses the re-absorption
of phosphate in the kidneys by reducing the number of
Na+-dependent co-transporters (27). In addition, the inhibition of
Cyp27b1, an enzyme that converts 25(OH)D to 1,25(OH)2D,
and the stimulation of 1,25(OH)2D catabolism both reduce
the quantity of circulating 1,25(OH)2D by increasing the
activity of 24-hydroxylase (Cyp24) (28). FGF23 enhances the activity of
several signal transduction pathways by interacting with the
Klotho-FGF receptor complex (29). Furthermore, the levels of serum
FGF23 are increased during early stage CKD before any measurable
changes in serum levels of PTH and phosphate can be detected
(30). By regulating the
phosphate flux mediated by sodium/phosphate co-transporters 2a and
2c, Klotho protects cells against apoptosis and senescence
(31). In particular, it was
shown that Klotho reduces the severity of VC (32). Increased FGF23 expression and
Klotho deficiency induced arterial stiffness and VC in animal
experiments (10).
In the present study, the serum levels of
creatinine, BUN FGF23, calcium and phosphorus as well as the plasma
levels of PTH among three animal groups, the Adenine-free,
Adenine-vehicle and Adenine-lanthanum groups were measured. The
serum levels of creatinine, BUN, FGF23, calcium and phosphorus, and
the plasma levels of PTH were different among the different
groups.
It was previously shown both in vitro and
in vivo that FGF23 induced ERK1/2 phosphorylation to
subsequently enhance the activity of MAPK signalling and the
synthesis of early growth response 1 proteins (6,27).
As a member of the MAPK family, ERK is activated by a wide range of
inflammatory cytokines and environmental stressors (33-36). Upon activation, ERK participates
in the regulation of various cellular responses and intracellular
signalling pathways as well as the pathogenesis of several
diseases, such as rheumatoid arthritis (36). For example, in hereditary
osteoarthritis, activation of discoidin domain receptor 2 by type
II collagen increases the expression of MMP-13 through p38 and
Ras/Raf/MEK/ERK signalling (37).
In the present study, western blot analysis was performed to
compare the levels of t-ERK and p-ERK among the different groups.
The results showed that the levels of t-ERK were comparable among
the groups. However, the levels of p-ERK and aortic calcium in the
Adenine-vehicle group were significantly higher compared with the
Adenine-lanthanum group.
The process of VC is similar to the process of bone
mineralization, and the pathogenesis of VC also involves the
synthesis of bone-related proteins, including type-I collagen and
osteopontin, by VSMCs (38). It
was also shown that through the ERK pathway and oxidative stress,
VC is induced by advanced oxidation protein products (AOPPs) via
the promotion of osteoblast differentiation of VSMCs (39). In the present study, FGF23
increased the levels of p-ERK in a dose-dependent manner, and that
the effects of FGF23 increased in time-dependent manner.
Furthermore, FGF23 also increased the levels of Msx2 and Osx in a
dose-dependent manner in human and rat VSMCs. In 48 subjects
stratified into two groups based on treatment, a regular group and
a fortified group, serum levels of FGF23, phosphorus and PTH in the
fortified group were reduced, whereas the serum levels of calcium
in the fortified group was significantly increased. Thus, it was
hypothesized that the changes in the serum levels of FGF23 were
positively correlated with the changes in the serum levels of
calcium in the fortified group but not in the regular group.
However, in the animal experiment, lanthanum
treatment failed to ameliorate the severity and incidence of kidney
injuries, but the treatment did decrease the levels of Scr and BUN.
These paradoxical results may be attributed to the fact that
several other factors other than Scr and BUN may influence kidney
injuries. Therefore, in future studies, the involvement of
additional mechanisms will be further investigated. One limitation
of the present study was that the cohort recruited was limited in
ethnicity. Thus, in future studies, larger, more diverse cohorts
should be investigated.
In summary, it was shown that the reduction in
phosphorus by lanthanum reduced the levels of FGF23 and the
severity of VC in CKD. In addition, upregulation of FGF23
expression increased the serum levels of phosphorus. Furthermore,
increased serum phosphorus concentrations were associated with
progression of VC and increased risk of death in patients
undergoing dialysis. These results suggest that targeting FGF23 may
be beneficial in the management and prevention of VC in patients
with CKD.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analysed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JJ and GL designed the study. YL and DZ collected
the references. JJ, YL, DZ, ZW and HZ collected and analysed the
data. JJ and GL wrote the manuscript. All authors read and approved
the final version of the manuscript.
Ethics approval and consent to
participate
All animal studies were performed in strict
compliance with the Guide for the Care and Use of Laboratory
Animals and ethical approval was granted by the Ethics Committee of
Dongguan People's Hospital Affiliated to Southern Medical
University. Use of patient samples was approved by the
Institutional Ethics Review Committee of Dongguan People's Hospital
Affiliated to Southern Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Kohl LP, Shroff GR and Herzog CA:
Understanding risks associated with chronic kidney disease:
Translating observational data to patient care. Clin Chem.
59:876–879. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Moe SM: Vascular calcification and renal
osteodystrophy relationship in chronic kidney disease. Eur J Clin
Invest. 36(Suppl 2): 51–62. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Massy ZA, Mazière C, Kamel S, Brazier M,
Choukroun G, Tribouilloy C, Slama M, Andrejak M and Maziere JC:
Impact of inflammation and oxidative stress on vascular
calcifications in chronic kidney disease. Pediatr Nephrol.
20:380–382. 2005. View Article : Google Scholar
|
|
4
|
Calvo MS and Uribarri J: Public health
impact of dietary phosphorus excess on bone and cardiovascular
health in the general population. Am J Clin Nutr. 98:6–15. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fukumoto S and Yamashita T: FGF23 is a
hormone-regulating phosphate metabolism-unique biological
characteristics of FGF23. Bone. 40:1190–1195. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ben-Dov IZ, Galitzer H, Lavi-Moshayoff V,
Goetz R, Kuro-o M, Mohammadi M, Sirkis R, Naveh-Many T and Silver
J: The parathyroid is a target organ for FGF23 in rats. J Clin
Invest. 117:4003–4008. 2007.PubMed/NCBI
|
|
7
|
Yamazaki M, Ozono K, Okada T, Tachikawa K,
Kondou H, Ohata Y and Michigami T: Both FGF23 and extracellular
phosphate activate Raf/MEK/ERK pathway via FGF receptors in HEK293
cells. J Cell Biochem. 111:1210–1221. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gutiérrez OM, Mannstadt M, Isakova T,
Rauh-Hain JA, Tamez H, Shah A, Smith K, Lee H, Thadhani R, Jüppner
H and Wolf M: Fibroblast growth factor 23 and mortality among
patients undergoing hemodialysis. N Engl J Med. 359:584–592. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jean G, Bresson E, Terrat JC, Vanel T,
Hurot JM, Lorriaux C, Mayor B and Chazot C: Peripheral vascular
calcification in long-haemodialysis patients: Associated factors
and survival consequences. Nephrol Dial Transplant. 24:948–955.
2009. View Article : Google Scholar
|
|
10
|
Jimbo R, Kawakami-Mori F, Mu S, Hirohama
D, Majtan B, Shimizu Y, Yatomi Y, Fukumoto S, Fujita T and
Shimosawa T: Fibroblast growth factor 23 accelerates
phosphate-induced vascular calcification in the absence of Klotho
deficiency. Kidney Int. 85:1103–1111. 2014. View Article : Google Scholar
|
|
11
|
Asadur R, Daisuke Y, Abu S, Kento K,
Hirofumi H, Daisuke N and Akira N: A novel approach to
adenine-induced chronic kidney disease associated anemia in
rodents. PLoS One. 13:e01925312018. View Article : Google Scholar
|
|
12
|
World Medical Association: World medical
association declaration of helsinki: Ethical principles for medical
research involving human subjects. JAMA. 310:2191–2194. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lim K, Lu TS, Molostvov G, Lee C, Lam FT,
Zehnder D and Hsiao LL: Vascular Klotho deficiency potentiates the
development of human artery calcification and mediates resistance
to fibroblast growth factor 23. Circulation. 125:2243–2255. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the Care and Use of Laboratory Animals: Guide
for the Care and Use of Laboratory Animals. 8th edition. National
Academies Press (US); Washington, DC: 2011
|
|
15
|
Shima WN, Ali AM, Subramani T, Mohamed
Alitheen NB, Hamid M, Samsudin AR and Yeap SK: Rapid growth and
osteogenic differentiation of mesenchymal stem cells isolated from
human bone marrow. Exp Ther Med. 9:2202–2206. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hunt JL, Fairman R, Mitchell ME, Carpenter
JP, Golden M, Khalapyan T, Wolfe M, Neschis D, Milner R, Scoll B,
et al: Bone formation in carotid plaques: A clinicopathological
study. Stroke. 33:1214–1219. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Giachelli CM: The emerging role of
phosphate in vascular calcification. Kidney Int. 75:890–897. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Goodman WG, Goldin J, Kuizon BD, Yoon C,
Gales B, Sider D, Wang Y, Chung J, Emerick A, Greaser L, et al:
Coronary-artery calcification in young adults with end-stage renal
disease who are undergoing dialysis. N Engl J Med. 342:1478–1483.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Blacher J, Guerin AP, Pannier B, Marchais
SJ and London GM: Arterial calcifications, arterial stiffness, and
cardiovascular risk in end-stage renal disease. Hypertension.
38:938–942. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Prié D, Torres PU and Friedlander G:
Latest findings in phosphate homeostasis. Kidney Int. 75:882–889.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Isakova T, Wahl P, Vargas GS, Gutiérrez
OM, Scialla J, Xie H, Appleby D, Nessel L, Bellovich K, Chen J, et
al: Fibroblast growth factor 23 is elevated before parathyroid
hormone and phosphate in chronic kidney disease. Kidney Int.
79:1370–1378. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Stevens LA, Djurdjev O, Cardew S, Cameron
EC and Levin A: Calcium, phosphate, and parathyroid hormone levels
in combination and as a function of dialysis duration predict
mortality: Evidence for the complexity of the association between
mineral metabolism and outcomes. J Am Soc Nephrol. 15:770–779.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lynch KE, Lynch R, Curhan GC and Brunelli
SM: Prescribed dietary phosphate restriction and survival among
hemodialysis patients. Clin J Am Soc Nephrol. 6:620–629. 2011.
View Article : Google Scholar :
|
|
24
|
Kestenbaum B: Phosphate metabolism in the
setting of chronic kidney disease: Significance and recommendations
for treatment. Semin Dial. 20:286–294. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu S, Guo R, Simpson LG, Xiao ZS, Burnham
CE and Quarles LD: Regulation of fibroblastic growth factor 23
expression but not degradation by PHEX. J Biol Chem.
278:37419–37426. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Quarles LD: Endocrine functions of bone in
mineral metabolism regulation. J Clin Invest. 118:3820–3828. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kurosu H, Ogawa Y, Miyoshi M, Yamamoto M,
Nandi A, Rosenblatt KP, Baum MG, Schiavi S, Hu MC, Moe OW and
Kuro-o M: Regulation of fibroblast growth factor-23 signaling by
klotho. J Biol Chem. 281:6120–6123. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Beckman MJ, Tadikonda P, Werner E, Prahl
J, Yamada S and DeLuca HF: Human 25-hydroxyvitamin
D3-24-hydroxylase, a multicatalytic enzyme. Biochemistry.
35:8465–8472. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Goetz R and Mohammadi M: Exploring
mechanisms of FGF signalling through the lens of structural
biology. Nat Rev Mol Cell Biol. 14:166–180. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Craver L, Marco MP, Martínez I, Rue M,
Borràs M, Martín ML, Sarró F, Valdivielso JM and Fernández E:
Mineral metabolism parameters throughout chronic kidney disease
stages 1-5-achievement of K/DOQI target ranges. Nephrol Dial
Transplant. 22:1171–1176. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dërmaku-Sopjani M, Sopjani M, Saxena A,
Shojaiefard M, Bogatikov E, Alesutan I, Eichenmüller M and Lang F:
Downregulation of NaPi-IIa and NaPi-IIb Na-coupled phosphate
transporters by coexpression of Klotho. Cell Physiol Biochem.
28:251–258. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ikushima M, Rakugi H, Ishikawa K, Maekawa
Y, Yamamoto K, Ohta J, Chihara Y, Kida I and Ogihara T:
Anti-apoptotic and anti-senescence effects of Klotho on vascular
endothelial cells. Biochem Biophys Res Commun. 339:827–832. 2006.
View Article : Google Scholar
|
|
33
|
Eliopoulos AG, Wang CC, Dumitru CD and
Tsichlis PN: Tpl2 transduces CD40 and TNF signals that activate ERK
and regulates IgE induction by CD40. EMBO J. 22:3855–3864. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
El-Najjar N, Chatila M, Moukadem H,
Vuorela H, Ocker M, Gandesiri M, Schneider-Stock R and
Gali-Muhtasib H: Reactive oxygen species mediate
thymoquinone-induced apoptosis and activate ERK and JNK signaling.
Apoptosis. 15:183–195. 2010. View Article : Google Scholar
|
|
35
|
Lannuzel A, Barnier JV, Hery C, Huynh VT,
Guibert B, Gray F, Vincent JD and Tardieu M: Human immunodeficiency
virus type 1 and its coat protein gp120 induce apoptosis and
activate JNK and ERK mitogen-activated protein kinases in human
neurons. Ann Neurol. 42:847–856. 2010. View Article : Google Scholar
|
|
36
|
Thiel MJ, Schaefer CJ, Lesch ME, Mobley
JL, Dudley DT, Tecle H, Barrett SD, Schrier DJ and Flory CM:
Central role of the MEK/ERK MAP kinase pathway in a mouse model of
rheumatoid arthritis: Potential proinflammatory mechanisms.
Arthritis Rheum. 56:3347–3357. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu L, Peng H, Wu D, Hu K, Goldring MB,
Olsen BR and Li Y: Activation of the discoidin domain receptor 2
induces expression of matrix metalloproteinase 13 associated with
osteoarthritis in mice. J Biol Chem. 280:548–555. 2005. View Article : Google Scholar
|
|
38
|
Wallin R, Wajih N, Greenwood GT and Sane
DC: Arterial calcification: A review of mechanisms, animal models,
and the prospects for therapy. Med Res Rev. 21:274–301. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
You H, Yang H, Zhu Q, Li M, Xue J, Gu Y,
Lin S and Ding F: Advanced oxidation protein products induce
vascular calcification by promoting osteoblastic
trans-differentiation of smooth muscle cells via oxidative stress
and ERK pathway. Ren Fail. 31:313–319. 2009. View Article : Google Scholar : PubMed/NCBI
|