Introduction
Diabetes mellitus (DM) is a major metabolic disorder
that exerts a negative effect on global public health (1). As a particular microvascular
complication of diabetes, diabetic retinopathy (DR) can be
classified as non-proliferative diabetic retinopathy (NPDR) and
proliferative diabetic retinopathy (PDR), involving retinal
neovascularization following ischemia (2). DR has become the leading cause of
visual impairment in adults in a number of countries and is a major
burden on patients and society (3,4).
Thus, it is necessary to examine the pathogenesis of DR and
identify effective measures for its prevention.
MicroRNAs (miRNAs or miRs) are a class of highly
conserved small-molecule non-coding RNAs that regu-late gene
expression by binding to complementary target sequences of the 3′
untranslated regions (3′-UTR) of mRNAs (5). Recently, studies have focused on
miRNAs as important regulatory mediators for diabetic microvascular
and macrovascular complications (6,7).
miR-126 (also known as miR-126-3p), is an endothelial cell-specific
miRNA that is highly expressed in endothelial cells, such as
capillaries and large blood vessels. It plays a critical role in
regulating angiogenic signaling and maintaining vascular integrity
in endothelial cells (8). A
previous study by the authors found that the serum miR-126 level in
patients with DR, particularly those with PDR, was significantly
lower than that in healthy individuals (9). However, the precise mechanisms of
action of miR-126 in DR remain poorly understood.
Polo-like kinase (PLK) is a type of serine/threonine
kinase that plays a key regulator in various cellular events during
cell divisions, allowing cells to divide and grow normally. From
previous research, it was found that miR-126 directly targets PLK4
in hepatocellular carcinoma (10). PLK4 is a unique member of the PLK
family and serves as a primary regulator of centrosome duplication
and mitotic progression. It has been demonstrated that PLK4
expression is increased in highly proliferating tissues (11).
Thus, it was hypothesized that miR-126 can affect DR
by regulating PLK4 expression. In the present study, rat models of
STZ-induced diabetes and high glucose (HG)-induced HRCECs were used
to explore the role and mechanisms of miR-126 in DR, with an aim of
providing potential therapeutic targets.
Materials and methods
Animals
Male SD rats weighing 200-250 g (n=55) were
purchased from the Laboratory Animal Centre of Southern Medical
University (Guangzhou, China) and housed with free access to food
and water in the Medical Research Center of the Third Affiliated
Hospital of Southern Medical University. The environment was
maintained at 20-26°C, relative humidity of 40-60%, a 12-h
light/dark cycle, and with natural and mechanical ventilation. All
animal experiments were approved by the Animal Ethics Committee of
Southern Medical University.
Diabetes induction
The rats were randomly divided into the non-DM (NDM)
and diabetic groups (DM). After being allowed to eat freely for 3
days to adapt to their new living environment, the rats in the DM
group (n=35) were administered a single intraperitoneal injection
of STZ (60 mg/kg; Sigma-Aldrich; Merck KGaA) dissolved in citrate
buffer (pH 4.4) to induce diabetes. The same dose of citrate buffer
was used for treating the rats in the NDM group (n=20). One week
later, the blood glucose level was measured using a blood sample of
the tail vein, and values ≥16.7 mmol/l were considered to indicate
diabetes. Rat weight and blood glucose levels were assessed once
every 2 weeks.
Intravitreal injection
Lentivirus expressing rno-miR-126 (lenti-miR-126,
TCG TAC CGT GAG TAT AAT GCG) and negative control oligonucleotide
with a scrambled sequence (lenti-NC, TTC TCC GAA CGT GTC ACG T)
were obtained from GenePharma, Inc. At 1 week following the
induction of diabetes, rats were randomly grouped and anesthetized
with an intraperitoneal injection of 3% pentobarbital sodium (45
mg/kg) and then administered an intravitreal injection of 4
µl of lentivirus to each of the two eyes using a 33-gauge
needle (Hamilton™, GuangZhou Jetway Biotech Co., Ltd.). Following
the injection, levofloxacin hydrochloride eye gel obtained from EBE
Pharmaceutical Co., Ltd. was used to prevent infection. Animal
health and behavior were monitored on a daily basis. Of the 55 rats
purchased, 1 rat failed to acquire diabetes, 3 rats died following
anesthesia during intravitreal injection, and 3 rats died of
diabetes. After 3 months, the remaining rats (n=48) were
anesthetized with pentobarbital sodium (45 mg/kg), and
approximately 3-4 ml of blood was drawn from each rat. The rats
were then euthanized by cervical dislocation. The eyeballs were
removed from the dead rats after confirming the cessation of the
heartbeat and breathing, stiff limbs, whitening lips and the
disappearance of blinking reflexes.
Cells and cell culture
Human retinal capillary endothelial cells (HRCECs,
JNO-121) were purchased from Guangzhou Jennio Biotech Co., Ltd.,
and cultured in endothelial cell medium (ECM, Sciencell, Inc.) at
37°C in a humidified incubator with 5% CO2. When the
HRCECs reached a confluence of 80-90%, they were digested with
0.25% trypsin without EDTA and sub-cultured. The HRCECs were
cultured in normal glucose (NG, 5.5 mM) or a high concentration of
glucose (HG, 25 mM) in ECM for 48 h to examine the mechanisms of
DR. Cells were then cultured using CFI-400945 fumarate
(MedChemExpress, New Jersey, USA) at a concentration of 200 nM for
24 h.
Cell transfection
miR-126 mimic (sense 5′-UCG UAC CGU GAG UAA UAA UGC
G-3′ and antisense 5′-CAU UAU UAC UCA CGG UAC GAU U-3′), miR-126
inhibitor (5′-CGC AUU AUU ACU CAC GGU ACG A-3′) were purchased from
Sangon Biotech Co., Ltd. They were separately transfected into
HRCECs at a concentration of 50 nM using Lipofectamine 3000 reagent
(Life Technologies; Thermo Fisher Scientific, Inc.) as instructed
by the manufacturer for 6 h. The cells were then collected for use
in subsequent experiments following 24 h of transfection.
RNA extraction and RT-qPCR
Total serum RNA was extracted using the miRNeasy
Serum/Plasma kit (Qiagen, Inc.). RNA was extracted from the mouse
retinas or HRCECs using RNA isolater Total RNA Extraction Reagent
(TRIzol reagent; R401-01, Nanjing Vazyme Medical Technology Co.,
Ltd.). RT-qPCR was performed using the miRNA 1st Strand cDNA
Synthesis kit (by stem-loop), miRNA Universal SYBR qPCR Master Mix,
HiScript® II Q RT SuperMix for qPCR and ChamQ™
SYBR® qPCR Master Mix (all from Nanjing Vazyme Medical
Technology Co., Ltd.). All the above-mentioned reactions were
executed according to the protocols supplied by the manufacturer.
Relative quantities of miR-126 were normalized to U6, and PLK4 mRNA
was normalized to GAPDH. The thermocycling conditions are as
follows: A single cycle of initial denaturation at 95°C/5 min (for
miRNA) or 95°C/30 sec (for RNA), then 40 cycles of denaturation at
95°C/10 sec, annealing at 56°C/30 sec, and extension at 72°C/30
sec. After the last cycle, the reaction was completed by a melting
curve step at 95°C/15 sec, 60°C/60 sec, and 95°C/15 sec. The primer
sequences were as follows: miR-126 RT, 5′-GTC GTA TCC AGT GCA GGG
TCC GAG GTA TTC GCA CTG GAT ACG ACC GCA TT-3′ and forward, 5′-CGC
GTC GTA CCG TGA GTA AT-3′; U6 RT, 5′-GTC GTA TCC AGT GCA GGG TCC
GAG GTA TTC GCA CTG GAT ACG ACT TGG CG-3′ and forward, 5′-GGA GAC
ACG CAA ACG GAA G-3′; human PLK4 forward, 5′-TCC AAG AGG CAG AAG
AAA GA CC-3′ and reverse, 5′-GCT GAG TGA CAT CGT TCC ATT G-3′. All
primers and GAPDH primer solutions were purchased from Sangon
Biotech (Shanghai) Co., Ltd. Relative gene expression was
calculated using the 2−ΔΔCq method (12). Three independent experiments were
carried out.
Western blot analysis
In order to obtain protein samples, the retinas or
HRCECs were placed in RIPA buffer with a protease inhibitor
(phenylmethanesulfonyl fluoride), crushed using an ultrasonic cell
pulverizer, centrifuged (5,000 × g/2 min at room temperature) to
obtain the supernatant protein, and quantified using a BCA Protein
Assay kit (Nanjing KeyGen Biotech. Co., Ltd.). Equivalent amounts
of protein (20 µg) from each sample were run at 10% SDS-PAGE
and transferred to 0.22 µm polyvinylidene fluoride membranes
(EMD Millipore). After blocking with 5% non-fat dried milk solution
at room temperature for 2 h, the membranes were first incubated
overnight at 4°C with polyclonal rabbit anti-vascular endothelial
growth factor (VEGF) antibody (A12303, ABclonal; 1:3,000),
polyclonal rabbit anti-pigment epithelium-derived factor (PEDF)
antibody (A11782, ABclonal; 1:3,000), poly-clonal rabbit anti-PLK4
antibody (12952-1-AP, ProteinTech Group, Inc.; 1:1,000) or
monoclonal mouse anti-β-tubulin antibody (CW0098, ComWin Biotech;
1:5,000), and then with appropriate peroxidase-labeled secondary
anti-rabbit or anti-mouse antibodies (RM3002, or RM3001, Beijing
Ray Antibody Biotech; 1:3,000) at room temperature for 1 h.
Chemiluminescent signals were visualized using Ncm-ECL Ultra (New
Cell & Molecular Biotech Co., Ltd.), while ImageJ software was
used to determine the gray strip values.
Paraffin sections and histological
assessment
Rat enucleated eyeballs separated from the anterior
segments were immediately fixed in 4% paraformaldehyde solution for
24 h, transferred to gradient dehydration (from 50 to 100%
ethanol), made transparent in xylene, embedded in paraffin, divided
into 4-µm-thick slices, and attached to slides. Following
deparaffinization and rehydration (xylene I, xylene II, 100%
ethanol I, 100% ethanol II each for 10 min, and 95, 90, 80 and 70%
ethanol each for 5 min), the vertical paraffin sections around
optic nerve (20 to 60 µm apart) were stained with
hematoxylin for 3 min and with eosin for 30 sec (H&E, Beijing
Solarbio Science & Technology Co., Ltd.) at room temperature
and then examined under a light microscope (Olympus
Corporation).
Preparation of retinal vasculature by
Trypsin digestion
The retinal vasculature was prepared according to
the protocol described in the study by Chou et al (13). A fluorescence microscope (Carl
Zeiss AG) was used to obtain images of the prepared retinal
vessels, and 5 images were selected from each group. The number of
endothelial cells and pericytes from these images were counted
manually, and the ratio (E/P) was then calculated to determine the
extent of retinopathy.
Immunofluorescence staining
The steps for the preparation of the
paraffinembedded sections were the same as those described above.
Following deparaffinization and rehydration (xylene I, xylene II,
100% ethanol I, 100% ethanol II each for 10 min, and 95, 90, 80 and
70% ethanol each for 5 min), the sections were incubated in 0.01%
trypsin (Beijing Solarbio Science & Technology Co., Ltd.) for
antigen retrieval at 37°C for 15 min, rinsed 3 times with 0.01 M
PBS for 5 min/time, blocked with standard goat serum blocking
solution (Wuhan Boster Biological Technology, Ltd.) for 1 h at room
temperature, and incubated overnight at 4°C with monoclonal rabbit
anti-CD34 (ab81289, Abcam, 1:200) or polyclonal rabbit anti-PLK4
(12952-1-AP, ProteinTech Group, Inc., 1:150). The sections were
then incubated for 1 h at room temperature with donkey anti-rabbit
IgG (H+L) highly cross-adsorbed secondary antibody, Alexa Fluor 594
(A21207, Thermo Fisher Scientific, Inc.; 1:200) antibody and
covered with ProLong™ Diamond Antifade Mountant with DAPI (Thermo
Fisher Scientific, Inc.). Images of these sections were obtained
using a fluorescence microscope (Carl Zeiss AG).
Dual-luciferase activity assay
PCR was used to amplify the wild-type (Wt), and
mutant (Mut) fragments of the PLK4 3′UTR containing the predicted
miR-126-binding site. To construct luciferase reporter gene
vectors, the target fragments were inserted into the pGL3-Promotor
(Promega Corporation) by Nanjing Zhifan Biotechnology Co., Ltd. The
constructs were co-transfected into HRCECs along with miR-126 mimic
(50 nM) and negative control mimic (50 nM) using Lipofectamine 3000
reagent (Life Technologies; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. The sequences were as
follows: miR-126 mimic (sense, 5′-UCG UAC CGU GAG UAA UAA UGC G-3′
and antisense, 5′-CAU UAU UAC UCA CGG UAC GAU U-3′), negative
control mimic (sense, 5′-UUC UCC GAA CGU GUC ACG UTT-3′ and
antisense, 5′-ACG UGA CAC GUU CGG AGA ATT). Following 48 h of
transfection, the Dual-Luciferase Reporter Assay System (Promega
Corporation) was used following the instructions provided by the
manufacturer to determine the luciferase activity compared with
Renilla luciferase of the samples.
EdU assay
HRCECs were inoculated into 15 mm glass-bottomed
dishes. They were treated separately with different interventions
after being adherent and achieving a confluence of 60-70%.
Following 48 h of treatment, the cell proliferative capacity was
analyzed with a kFluor488-EdU assay kit (KGA337, Nanjing KeyGen
Biotech. Co., Ltd.) following the instructions provided by the
manufacturer. In detail, samples were incubated at 37°C for 2 h
with a working solution of 50 µM EdU. The nuclei were
stained blue with Hoechst 33342, protected from light at room
temperature for 30 min. The cells were imaged using a fluorescence
micro-scope (Olympus Corporation). A total of 5 fields of view were
selected randomly to calculate the positive rate.
Transwell assay
Following treatment of the HRCECs with different
interventions for 48 h, 1×105/ml cells were diluted with
serum-free ECM and transferred into the upper chamber of a
Transwell insert (Costar; Corning, Inc.). An ECM containing 5% FBS
was added to the lower chamber. The non-migrating cells were
removed gently from the upper chamber following 24 h of incubation
in a CO2 incubator at 37°C. Cells that had already
migrated through the membrane were fixed with 4% paraformaldehyde
for 20 min and stained with a solution of 0.5% crystal violet
(Beijing Solarbio Science & Technology Co., Ltd.) at room
temperature for 10 min. The migrated cells were imaged using an
inverted optical microscope (Olympus Corporation) and 5 fields of
view were randomly selected to count the cell numbers.
Statistical analysis
The experimental data are represented as the means ±
SD. Significant differences between 2 groups were evaluated with a
t-test using GraphPad Prism 7.0; one-way ANOVA and Tukey's test
following ANOVA were used to evaluated the differences between ≥3
groups for multiple comparisons. A value of P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-126 is downregulated in serum and
retinas of rats with STZ-induced diabetes
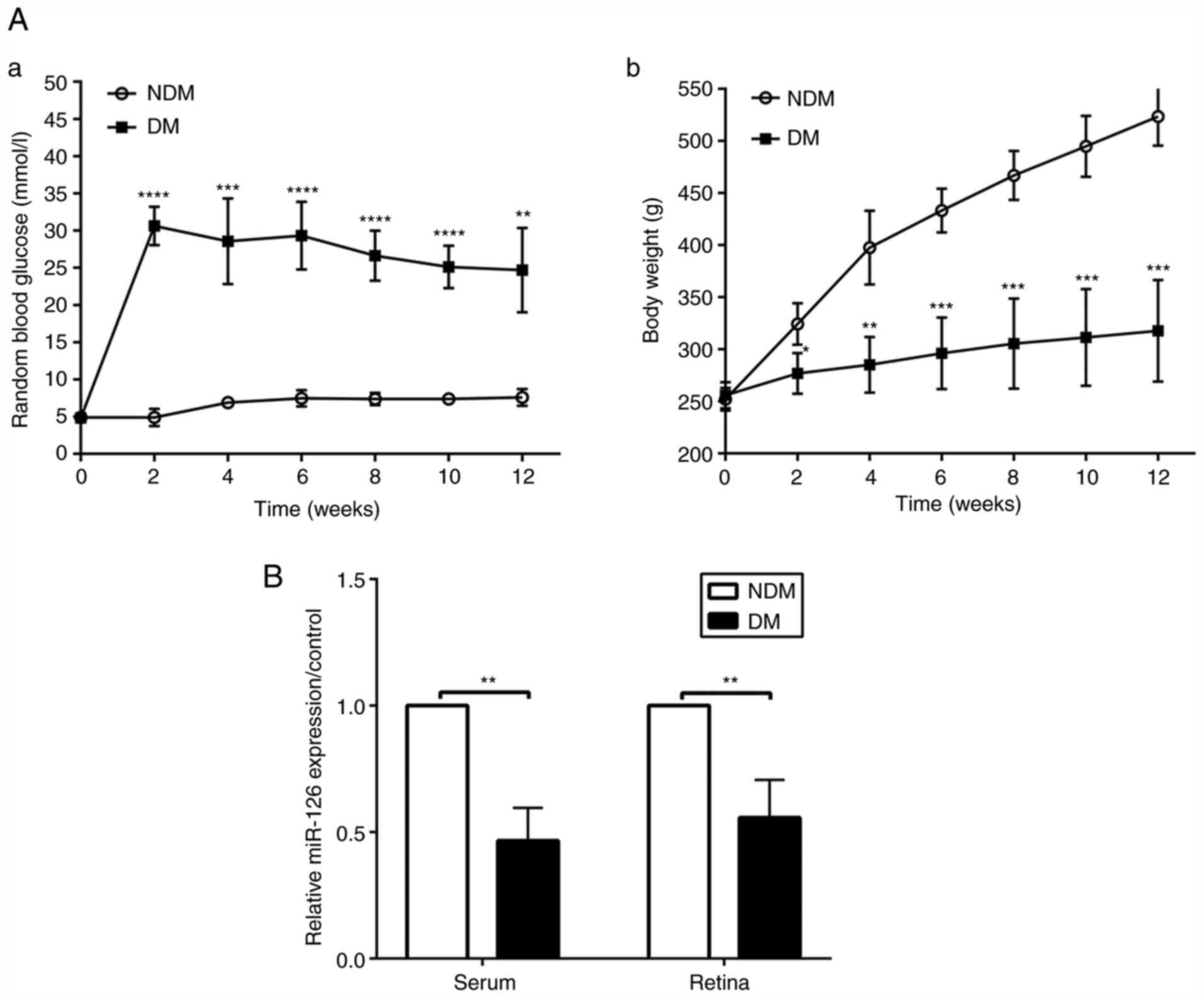
At the established time points of 2, 4, 6, 8, 10 and
12 weeks following the induction of diabetes, blood glucose levels
in the rats with DM were higher than those in the NDM group (both
P<0.05, Fig. 1A-a), while the
bodyweight of the rats with DM was lower than that of the rats in
the NDM group (both P<0.05, Fig.
1A-b). Random levels of blood glucose in the rats with DM were
>16.7 mmol/l and did not return to the normal concentration of
glucose, proving the successful induction of diabetes. The results
of RT-qPCR revealed that the miR-126 level in the serum and retinas
was decreased in the diabetic rats at 12 weeks, as compared with
the rats in the NDM group (both P<0.05, Fig. 1B).
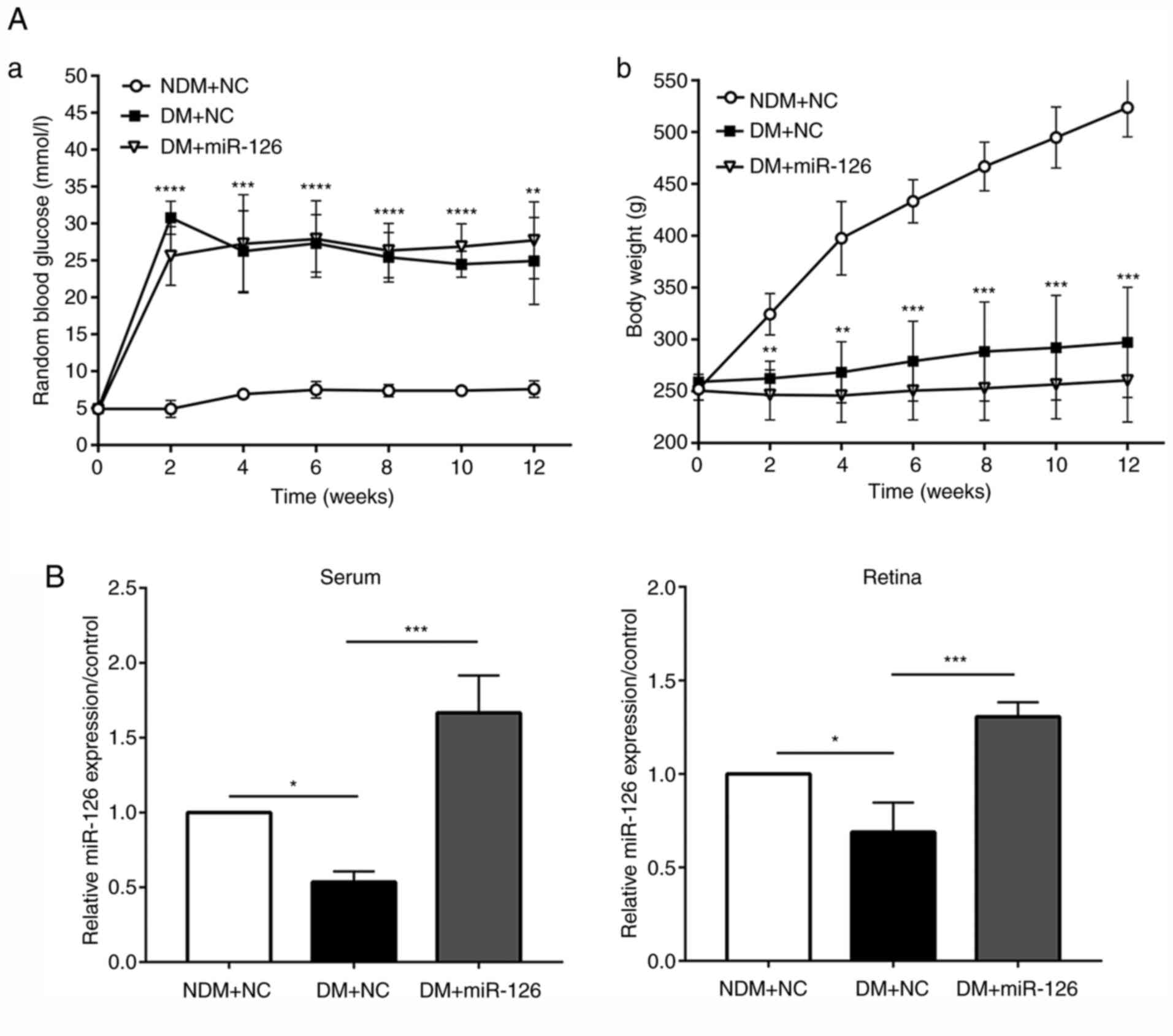
Intravitreal injection of lenti-miR-126
increases the level of miR-126 without affecting blood glucose
levels and weight of the diabetic rats
The lenti-miR126 or its negative control lenti-NC
were injected intravitreally into the eyes of rats in the DM and
NDM groups at 1 week following the induction of diabetes. Blood
glucose levels in the rats with DM injected with lenti-NC (DM+NC)
were higher than those in the rats in the NDM group injected with
lenti-NC (NDM+NC) at 2, 4, 6, 8, 10 and 12 weeks following the
induction of diabetes. The body weight of the rats in the DM+NC
group was, however, lower than that of the rats in the NDM+NC group
(both P<0.05). No significant differences in body weight and
blood glucose levels were observed between the rats with DM
injected with lenti-miR-126 (DM+miR-126) and the rats in the DM+NC
group, suggesting that the intravitreal injection of lentivirus did
not affect the physical conditions of rats, as presented in
Fig. 2A. At 3 months after the
STZ injection, the retinas were harvested and RT-qPCR was
performed. The results revealed a decrease in both serum and
retinal miR-126 levels in the rats in the DM+NC group compared to
those in the NDM+NC group. As shown in Fig. 2B, in the serum and retinas of the
rats with DM injected with lenti-miR-126, the level of miR-126 was
upregulated as compared with the rats with DM injected with
lenti-NC (both P<0.05).
Intraocular delivery of miR-126
alleviates retinopathy in rats with diabetes
H&E staining revealed a typical standard
architecture of retinal layers arranged regularly in the rats in
the NDM group injected with lenti-NC (NDM+NC). However, in the rats
with DM injected with lenti-NC, the structures of the ganglion cell
layer (GCL), inner nuclear layer (INL) and outer nuclear layer
(ONL) were loosened and disordered compared with those in the rats
with DM injected with lenti-miR-126 (DM+miR-126) (Fig. 3A). As also shown in Fig. 3A, the rats in the DM+NC group also
appeared to have more blood vessels (black arrow) compared with
those in the NDM+NC and DM+miR-126 groups.
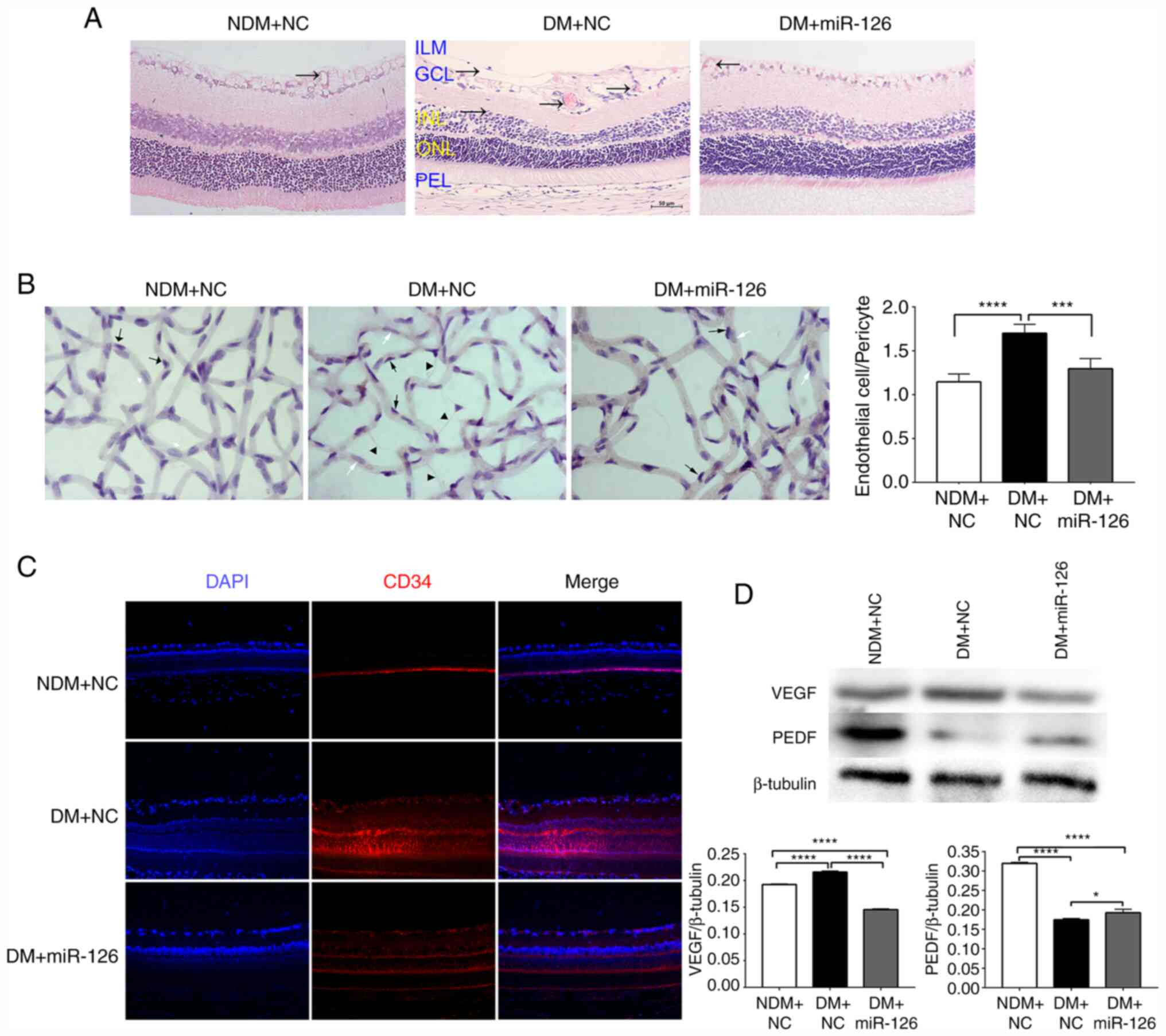 | Figure 3Intraocular delivery of miR-126
alleviates experimental diabetic retinopathy in diabetic rats. (A)
H&E staining of retinal paraffin-embedded sections
(magnification, ×200). ILM, inner limiting membrane; GCL, ganglion
cell layer; INL, inner nuclear layer; ONL, outer nuclear layer;
PEL, pigment epithelial layer. (B) The prepared retinal vasculature
by trypsin digest (magnification, ×400). Endothelial cells are
indicated by white arrows, pericytes are indicated by black arrows
and acellular capillaries are indicated by triangles. (C) Retinal
immunofluorescence staining for CD34. (D) The protein contents of
VEGF and PEDF in the rat retina. VEGF, vascular endothelial growth
factor; PEDF, pigment epithelium-derived factor; NDM+NC,
non-diabetic rats injected with lenti-NC; DM+NC, diabetic rats
injected with lenti-NC; DM+miR-126, diabetic rats injected with
lenti-miR-126. *P<0.05; ***P<0.001;
****P<0.0001. |
As shown by the retinal vasculature, the nucleus of
endothelial cells was large and oval-shaped, parallel to the
vasculature in the long axis (white arrows); the nuclei of
pericytes were small and triangle-shaped, located at one side of
the capillary (black arrows). In the rats in the DM+NC group, the
ratio of the number of endothelial cells to the number of pericytes
(E/P) was significantly higher compared to that in the NDM+NC and
DM+miR-126 groups (P<0.05). Furthermore, the retinas from the
rats in the DM+NC group exhibited more acellular capillaries
(triangles) than those from the rats in the NDM+NC and DM+miR-126
groups (Fig. 3B).
Immunofluorescence staining also revealed that the retinas from the
rats in the DM+NC group exhibited prominent CD34 staining compared
to those from the rats in the NDM+NC and DM+miR-126 groups
(Fig. 3C).
The results obtained from western blot analysis are
depicted in Fig. 3D. It was found
that the content of retinal VEGF protein was higher in the rats in
the DM+NC group than those in the NDM+NC group and lower in the
rats in the DM+miR-126 compared with the rats in the DM+NC group.
Furthermore, the PEDF retinal content was lower in the rats in the
DM+NC group than in those in the NDM+NC group, while it was higher
in the rats in the DM+miR-126 group compared with the rats in the
DM+NC group.
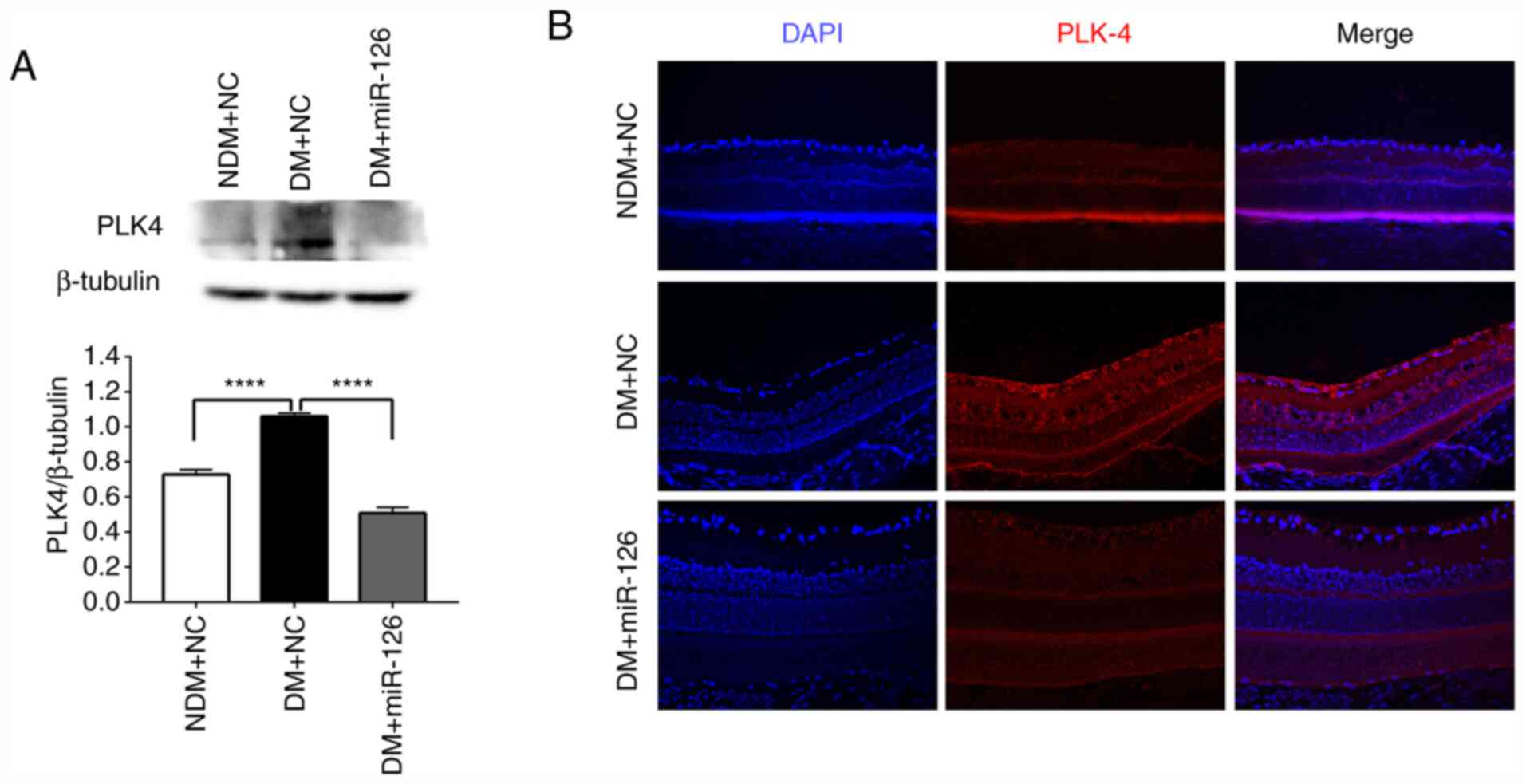
Intraocular delivery of miR-126
downregulates PLK4 expression in the retinas of diabetic rats
The results of western blot analysis revealed that
the protein content of PLK4 in the rats with DM injected with
lenti-NC (DM+NC) was higher than that in the rats in the NDM group
injected with lenti-NC (NDM+NC) (Fig.
4A). The intravitreal injection of lenti-miR-126 downregulated
PLK4 expression in the retinas of the rats with DM compared those
of the rats injected with lenti-NC (Fig. 4A). In addition, immunofluorescence
staining demonstrated that the retinas of the rats in the DM+NC
group exhibited prominent PLK4 staining compared to the those of
the rats in the NDM+NC and DM+miR-126 groups (Fig. 4B).
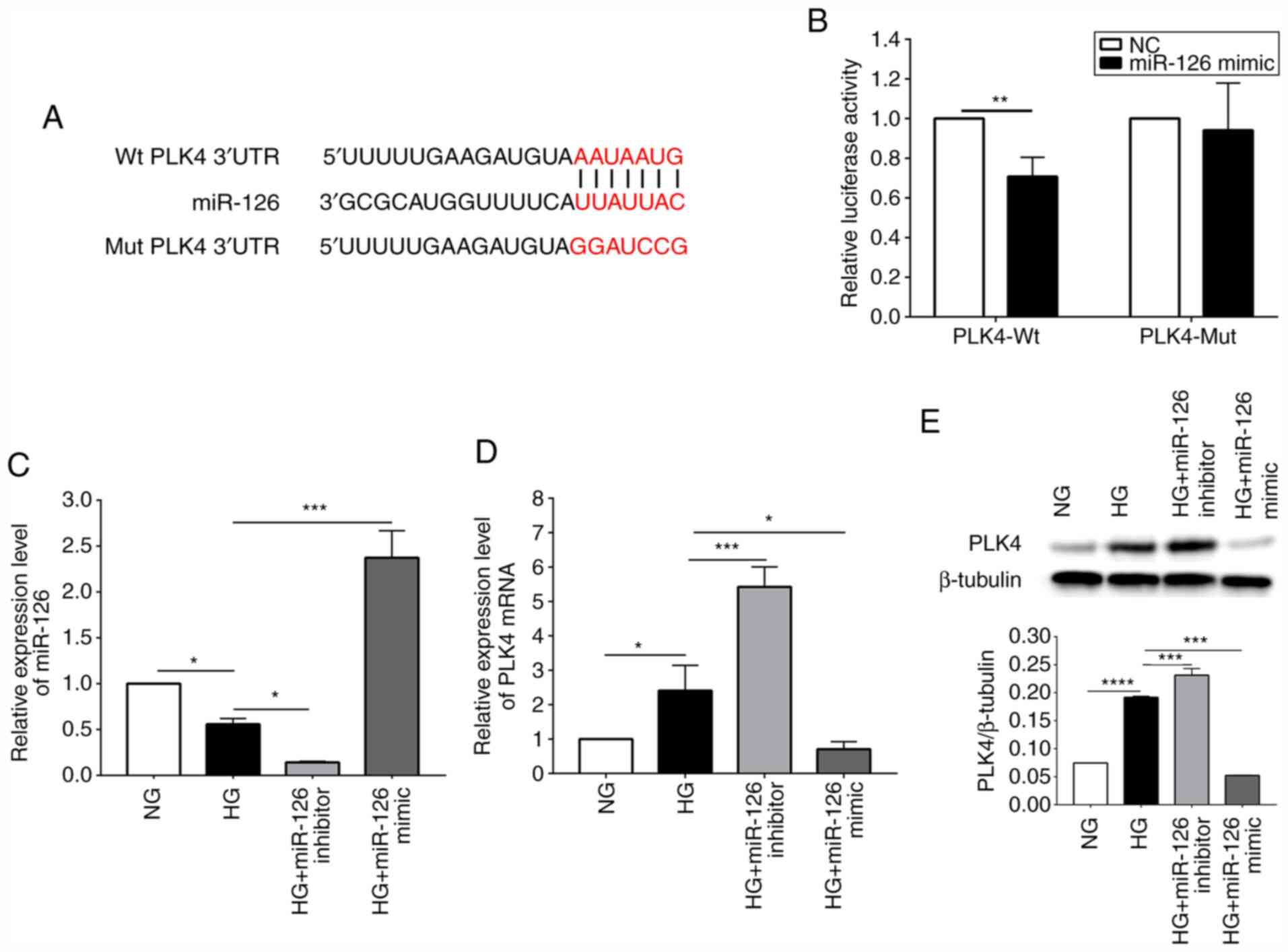
miR-126 negatively regulates the
expression of PLK4 in HG-induced HRCECs
In order to confirm that miR-126 targets PLK4,
luciferase reporter assays were performed in which the Wt or Mut
PLK4 3′-UTR bound with miR-126 and was fused to the luciferase gene
(Fig. 5A). Luciferase reporter
assay revealed that transfection with miRNA-126 mimic effectively
inhibited the luciferase activity in HRCECs expressing the Wt
reporter, but not in those expressing the Mut (Fig. 5B). Furthermore, the results of
RT-qPCR indicated that the miR-126 level in HG-induced HRCECs was
lower than that of the standard control (P<0.05). The miR-126
level was reduced (P<0.05) following transfection of the
HG-induced HRCECs with a miR-126 inhibitor. By contrast,
transfection with miR-126 mimic increased the miR-126 level
(P<0.0001) (Fig. 5C). As shown
in Fig. 5D, in the HG-induced
HRCECs, the mRNA expression of PLK4 was upregulated compared with
the normal control (P<0.05). Transfection with miR-126 inhibitor
upregulated PLK4 expression (P<0.001); however, transfection
with miR-126 mimic downregulated PLK4 expression (P<0.05). The
results of western blot analysis revealed that the PLK4 protein
content was higher in the HG-induced HRCECs compared to the
standard control and was negatively regulated by miR-126 (Fig. 5E).
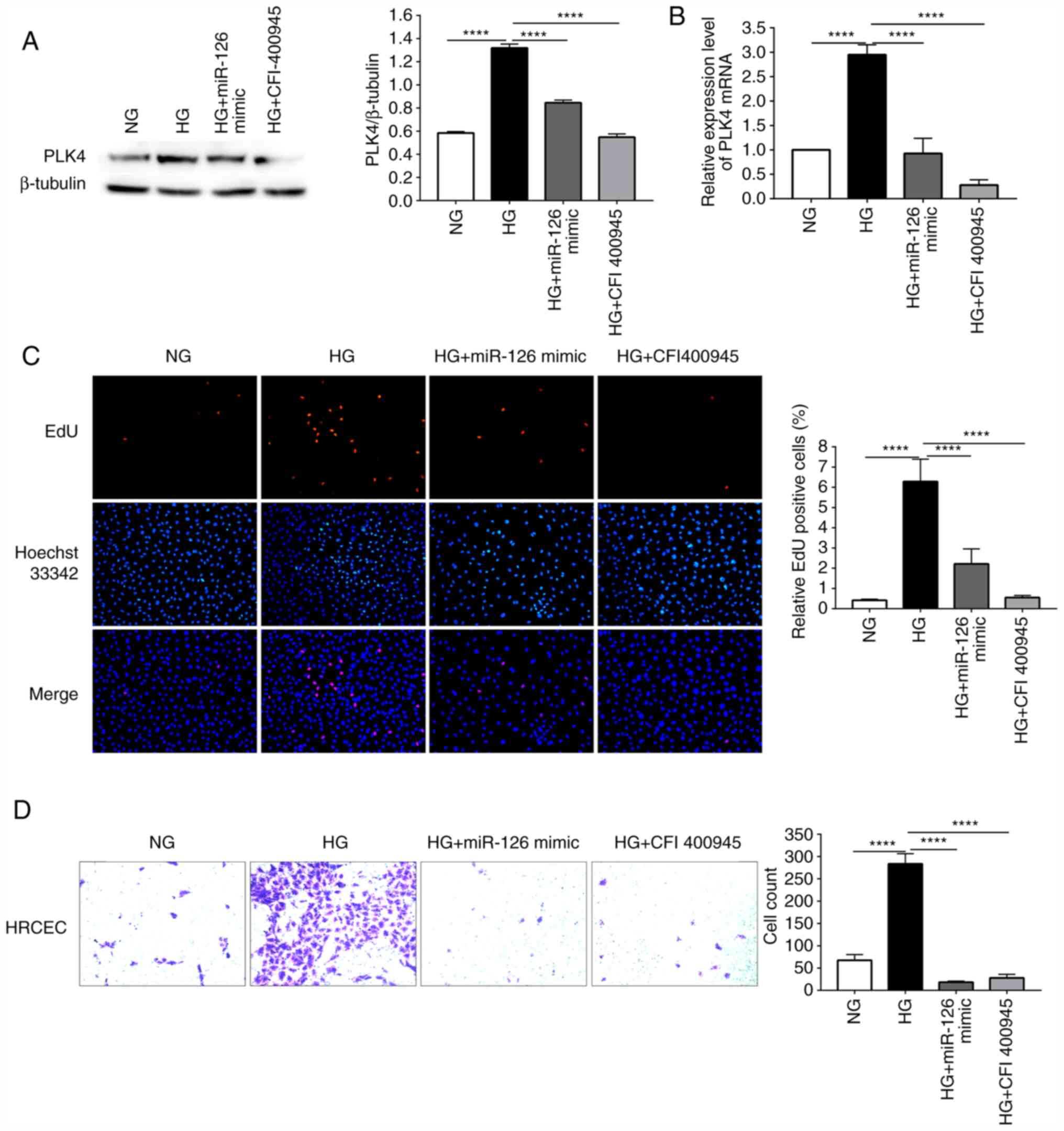
Overexpression of miR-126 suppresses the
proliferation and migration of HG-induced HRCECs by inhibiting PLK4
expression
miR-126 mimic was used to transfect HG-induced
HRCECs in order to examine the role of miR-126 in the angiogenesis
of DR. EdU and Transwell assays were used to evaluate the cell
proliferative and migratory abilities. CFI-400945 fumarate, a PLK4
inhibitor, was added to the HRCECs to inhibit the expression of
PLK4. The results of western blot analysis revealed that the PLK4
protein content was higher in the HG-induced HRCECs than in the NG
group. Transfection with miR-126 mimic and treatment with
CFI-400945 fumarate suppressed the HG-induced upregulation of PLK4
protein expression (Fig. 6A). In
the HG-induced HRCECs, the PLK4 mRNA level was upregulated compared
to the NG control, whereas it was down-regulated by transfection
with miR-126 mimic or treatment with CFI-400945 fumarate (Fig. 6B). In the EdU assay, HG treatment
was shown to result in a significant increase in cell proliferation
compared to the NG group. At the same time, miR-126 mimic, or
CFI-400945 fumarate suppressed the HG-induced increase in cell
proliferation (Fig. 6C). In the
Transwell assay, HG was shown to increase the migratory ability of
the HRCECs, which was suppressed by transfection with miR-126 mimic
or treatment with CFI-400945 fumarate (Fig. 6D).
Discussion
Rat models of STZ-induced diabetes have frequently
been used in studies related to diabetes and the associated
complications (14-16). In the present study, a rat model
of STZ-induced diabetes was established. RT-qPCR revealed that both
serum and retinal miR-126 levels in the diabetic rats were
decreased compared to those in the rats in the NDM group, which is
consistent with the findings of a previous study on the human body
by the authors (9). Moreover,
miRNAs are usually packaged in microvesicles to protect against
endogenous RNase activity in the circulation and are present in
this stable form. Thus, the serum miR-126 level can reflect its
level in the retinal tissues and act as a biomarker for DR
screening.
The activation of the angiogenesis process in
endothelial cells is essential for normal embryonic development and
physiological angiogenesis in adults. However, the formation of new
blood vessels in cancer and DR is both pathological and harmful. It
has been indicated that miRNAs participate in the maintenance of
vascular homeostasis and the prevention of terminal organ sequelae
caused by diabetes (17). miR-126
is the only miRNA known to be expressed explicitly in endothelial
cell lineage, hematopoietic progenitor cells, and endothelial cell
lines (18).
Lentivirus vectors can be stably integrated into the
host cell genome for effective exogenous gene expression without
any alterations of the cloned gene and have a high gene
transduction efficiency. In the present study, lenti-miR-126 was
injected intravitreally into the eyes of rats with DM to
investigate its function in DR. It was found that the intravitreal
injection of lenti-miR-126 increased the miR-126 level in the serum
and retinas of rats. Pericytes play a vital role in angiogenesis
and vessel stabilization. The condition of reduced retinal pericyte
coverage is a well-established fact of DR pathology (19). In the retinal vasculature, the
ratio of the number of endothelial cells to pericytes (E/P) was
used to assess DR. The results revealed that the diabetic rats
exhibited a higher E/P ratio and more acellular capillaries as
compared to the rats in the NDM group. However, the intraocular
delivery of miR-126 reduced the E/P ratio and acellular capillaries
number. In addition, the endothelial marker (CD34) was examined by
immunofluorescence to identify capillary endothelium. Although
vertical sections around the optic nerve (20 to 60 µm apart)
were selected for H&E staining and immunofluorescence, there
are still deficiencies in using these methods for the detection of
retinopathy. Therefore, the authors intend to make further progress
in this matter in future studies.
VEGF is an essential pro-angiogenic growth factor
that increases vascular permeability and promotes the proliferation
of the vascular endothelial cell. Studies have found that VEGF
plays a vital role in the development of DR (20,21). By contrast, PEDF is a potent
anti-angiogenic factor with neuroprotective effects, inhibiting the
pathological angiogenesis (22).
The protein content of VEGF and PEDF in the retina was used as an
auxiliary judgment of retinopathy. The results obtained from the
present study prove that the intraocular delivery of miR-126 can
alleviate retinopathy in diabetic rats. Furthermore, the results of
western blot analysis and immunofluorescence staining demonstrated
that the intraocular delivery of miR-126 downregulated the
diabetes-induced upregulation of PLK4 in vivo, indicating
that overexpression of miR-126 may alleviate DR by regulating PLK4
expression.
DR is a microangiopathy in which vascular
endothelial cells are the primary cellular targets. Thus, HRCECs
were cultured in ECM with a glucose concentration of 25 mM to
establish an in vitro DR model and were used to investigate
the mechanisms of miR-126 in DR (23,24). Luciferase reporter assay proved
that miR-126 could directly bind to PLK4. In HG-induced HRCECs,
PLK4 expression was increased compared to the NG group, although
the overexpression of miR-126 reduced PLK4 expression. It can thus
be concluded that miR-126 negatively regulates PLK4 expression in
DR. EdU and Transwell assays were used to investigate whether
miR-126 alleviates retinal angiogenesis during the DR process. The
miR-126 level was decreased in HG-induced HRCECs, and increased
PLK4 expression, as compared to the NG control. Cellular
proliferative and migratory abilities were also enhanced. Both
miR-126 mimic and CFI-400945 fumarate (a PLK4 inhibitor) reduced
PLK4 expression and cellular proliferation and migration. As a
result, miR-126 inhibited the proliferation and migration of
HG-induced HRCECs by regulating PLK4 expression.
According to the above-mentioned results, miR-126 is
decreased in DR and may reduce retinal angiogenesis by targeting
PLK4 expression. miR-126 also decreased in tissues and cells of
various cancers (10,25-28). It can inhibit proliferation,
migration, invasion and cell survival by inhibiting a range of
essential target genes, thereby inhibiting tumor progression
(29,30). The invention of miRNA agents that
are safe, effective, and tolerable in humans would have a positive
effect on the proper treatment of DR.
PLK4 has become a potential therapeutic target for
various types of advanced cancers (31). CFI-400945 fumarate is a highly
selective oral inhibitor of PLK4. Clinical studies have
demonstrated that a continuous daily oral dose of CFI-400945 is
feasible and safe for patients with advanced solid tumors (32). It was hypothesized that PLK4
inhibitors may play a major role in the treatment of DR. Future
studies using rats with DM with CFI-400945 fumarate may, therefore,
be performed to confirm the effects of PLK4 during the development
of DR and to explore the appropriate therapeutic dose of CFI-400945
fumarate.
In conclusion, the present study found that the
serum miR-126 level can act as a distinctive signature for DR
screening. The overexpression of miR-126 may inhibit endothelial
cell proliferation and migration by directly targeting PLK4
expression, thus alleviating diabetic retinopathy. These results
may set new targets for DR screening and treatment.
Funding
The present study was supported by the Science and
Technology Planning Project of Guangdong Province (grant no.
2014A020212631), and the Science and Technology Planning Project of
Guangzhou (grant no. 201604020105).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YaZ, YiL and LW conducted majority of the
experiments and assisted with manuscript preparation. HX, ZL, and
YX wrote the manuscript and acquired data. WM and LY performed
immunofluorescence and analyzed data. DF, YeZ and KK fed the rats
and measured their body weight and blood glucose levels. YaL and MA
developed the concept, supervised the project, conceived the
experiments and critically reviewed the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Southern
Medical University Committee on the Use and Care of Animals and
were performed in accordance with the committee's guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Cho NH, Shaw JE, Karuranga S, Huang Y, da
Rocha Fernandes JD, Ohlrogge AW and Malanda B: IDF diabetes atlas:
Global estimates of diabetes prevalence for 2017 and projections
for 2045. Diabetes Res Clin Pract. 138:271–281. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cheung N, Mitchell P and Wong TY: Diabetic
retinopathy. Lancet. 376:124–136. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Flaxman SR, Bourne R, Resnikoff S, Ackland
P, Braithwaite T, Cicinelli MV, Das A, Jonas JB, Keeffe J, Kempen
JH, et al: Global causes of blindness and distance vision
impairment 1990-2020: A systematic review and meta-analysis. Lancet
Glob Health. 5:e1221–e1234. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ting DS, Cheung GC and Wong TY: Diabetic
retinopathy: Global prevalence, major risk factors, screening
practices and public health challenges: A review. Clin Exp
Ophthalmol. 44:260–277. 2016. View Article : Google Scholar
|
|
5
|
Lim LP, Lau NC, Garrett-Engele P, Grimson
A, Schelter JM, Castle J, Bartel DP, Linsley PS and Johnson JM:
Microarray analysis shows that some microRNAs downregulate large
numbers of target mRNAs. Nature. 433:769–773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Giannella A, Radu CM, Franco L, Campello
E, Simioni P, Avogaro A, de Kreutzenberg SV and Ceolotto G:
Circulating levels and characterization of microparticles in
patients with different degrees of glucose tolerance. Cardiovasc
Diabetol. 16:1182017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hulsmans M and Holvoet P:
MicroRNA-containing microvesicles regulating inflammation in
association with atherosclerotic disease. Cardiovasc Res. 100:7–18.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pishavar E and Behravan J: miR-126 as a
therapeutic agent for diabetes mellitus. Curr Pharm Design.
23:3309–3314. 2017. View Article : Google Scholar
|
|
9
|
Qin LL, An MX, Liu YL, Xu HC and Lu ZQ:
MicroRNA-126: A promising novel biomarker in peripheral blood for
diabetic retinopathy. Int J Ophthalmol. 10:530–534. 2017.PubMed/NCBI
|
|
10
|
Bao J, Yu Y, Chen J, He Y, Chen X, Ren Z,
Xue C, Liu L, Hu Q, Li J, et al: MiR-126 negatively regulates PLK-4
to impact the development of hepatocellular carcinoma via ATR/CHEK1
pathway. Cell Death Dis. 9:10452018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Maniswami RR, Prashanth S, Karanth AV,
Koushik S, Govindaraj H, Mullangi R, Rajagopal S and Jegatheesan
SK: PLK4: A link between centriole biogenesis and cancer. Expert
Opin Ther Targets. 22:59–73. 2018. View Article : Google Scholar
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
13
|
Chou JC, Rollins SD and Fawzi AA: Trypsin
digest protocol to analyze the retinal vasculature of a mouse
model. J Vis Exp. 76:e504892013.
|
|
14
|
Semkova I, Huemmeke M, Ho MS, Merkl B,
Abari E, Paulsson M, Joussen AM and Plomann M: Retinal localization
of the glutamate receptor GluR2 and GluR2-regulating proteins in
diabetic rats. Exp Eye Res. 90:244–253. 2010. View Article : Google Scholar
|
|
15
|
Zhuang P, Muraleedharan CK and Xu S:
Intraocular delivery of miR-146 inhibits diabetes-induced retinal
functional defects in diabetic rat model. Invest Ophthalmol Vis
Sci. 58:1646–1655. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu WL: MicroRNA-9 inhibits retinal
neovascularization in rats with diabetic retinopathy by targeting
vascular endothelial growth factor A. J Cell Biochem.
120:8032–8043. 2018. View Article : Google Scholar
|
|
17
|
Zampetaki A, Kiechl S, Drozdov I, Willeit
P, Mayr U, Prokopi M, Mayr A, Weger S, Oberhollenzer F, Bonora E,
et al: Plasma microRNA profiling reveals loss of endothelial
miR-126 and other microRNAs in type 2 diabetes. Circ Res.
107:359–810. 2010. View Article : Google Scholar
|
|
18
|
Meister J and Schmidt MHH: miR-126 and
miR-126*: New players in cancer. ScientificWorldJournal.
10:2090–2100. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Trost A, Lange S, Schroedl F, Bruckner D,
Motloch KA, Bogner B, Kaser-Eichberger A, Strohmaier C, Runge C,
Aigner L, et al: Brain and retinal pericytes: Origin, function and
role. Front Cell Neurosci. 10:202016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Song E, Dong Y, Han LN, Sui D, Xu Q, Wang
X and Wu J: Diabetic retinopathy: VEGF, bFGF and retinal vascular
pathology. Chinese Med J (Engl). 117:247–251. 2004.
|
|
21
|
Ekerbicer N, Gurpinar T, Sisman AR,
Guvendi G, Camsari UM and Uysal N: Statins reduce testicular and
ocular VEGF: A potential compromise to microcirculation. Microvasc
Res. 119:60–63. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Simó R and Hernández C; European
Consortium for the Early Treatment of Diabetic Retinopathy
(EUROCONDOR): Neurodegeneration in the diabetic eye: New insights
and therapeutic perspectives. Trends Endocrinol Metab. 25:23–33.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Karthikkeyan G, Nareshkumar RN, Aberami S,
Sulochana KN, Vedantham S and Coral K: Hyperglycemia induced early
growth response-1 regulates vascular dysfunction in human retinal
endothelial cells. Microvasc Res. 117:37–43. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yao J, Wang J, Yao Y, Wang K, Zhou Q and
Tang Y: miR-133b regulates proliferation and apoptosis in
high-glucose-induced human retinal endothelial cells by targeting
ras homolog family member A. Int J Mol Med. 42:839–850.
2018.PubMed/NCBI
|
|
25
|
Tavazoie SF, Alarcón C, Oskarsson T, Padua
D, Wang Q, Bos PD, Gerald WL and Massagué J: Endogenous human
microRNAs that suppress breast cancer metastasis. Nature.
451:147–152. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hua Y, Liang C, Miao C, Wang S, Su S, Shao
P, Liu B, Bao M, Zhu J, Xu A, et al: MicroRNA-126 inhibits
proliferation and metastasis in prostate cancer via regulation of
ADAM9. Oncol Lett. 15:9051–9060. 2018.PubMed/NCBI
|
|
27
|
Chen H, Li L, Wang S, Lei Y, Ge Q, Lv N,
Zhou X and Chen C: Reduced miR-126 expression facilitates
angiogenesis of gastric cancer through its regulation on VEGF-A.
Oncotarget. 5:11873–11885. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen M, Peng W, Hu S and Deng J:
miR-126/VCAM-1 regulation by naringin suppresses cell growth of
human non-small cell lung cancer. Oncol Lett. 16:4754–4760.
2018.PubMed/NCBI
|
|
29
|
Ebrahimi F, Gopalan V, Smith RA and Lam
AK: miR-126 in human cancers: Clinical roles and current
perspectives. Exp Mol Pathol. 96:98–107. 2014. View Article : Google Scholar
|
|
30
|
Zhai X and Xu W: Long noncoding RNA ATB
promotes proliferation, migration, and invasion in bladder cancer
by suppressing microRNA-126. Oncol Res. 26:1063–1072. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Press MF, Xie B, Davenport S, Zhou Y,
Guzman R, Nolan GP, O'Brien N, Palazzolo M, Mak TW, Brugge JS and
Slamon DJ: Role for polo-like kinase 4 in mediation of cytokinesis.
Proc Natl Acad Sci USA. 116:11309–11318. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Veitch ZW, Cescon DW, Denny T, Yonemoto
LM, Fletcher G, Brokx R, Sampson P, Li SW, Pugh TJ, Bruce J, et al:
Safety and tolerability of CFI-400945, a first-in-class, selective
PLK4 inhibitor in advanced solid tumours: A phase 1 dose-escalation
trial. Br J Cancer. 121:318–324. 2019. View Article : Google Scholar : PubMed/NCBI
|