Introduction
Aging is a universal biological phenomenon known to
induce a declines in the physiological capacity of a number of
tissues, including the cardiovascular system. As the elderly
population is increasing worldwide, age has become the dominant
risk factor for the development of cardiovascular diseases
(1). To prevent age-related
cardiac diseases, further insight should be gained from the
investigation of the processes involved in cardiac aging.
The mitochondria constitute the central metabolic
hub in cells and are the gateway to aerobic metabolism (2). The mitochondria have also been
proven to be intimately involved in signal transduction and aging
processes, particularly in the aging of the heart (3,4).
The mitochondria also undergo several alternations in aged
cardiomyocytes, such as reductions in the number and area of the
inner membrane, an increase in the volume fraction and alternations
in shape (5-9).
Cytochrome c oxidase (COX), also known as complex
IV, is the terminal enzyme of the electron transport chain,
catalyzing the reduction of oxygen to water. COX is composed of at
least 15 subunits, 12 of which are encoded by nuclear DNA and 3 of
which are encoded by mitochondrial DNA (mtDNA) (10). The mtDNA-encoded subunits, COX1
(MT-COI), COX2 (MT-COII) and COX3 (MT-COIII), form the catalytic
core of the enzyme (11,12).
Similar to the methylation of genomic DNA, mtDNA has
been found to be CpG-methylated, which plays an important role in
mitochondrial gene regulation (13,14). A previous study by the authors
reported a phenomenon of decreased methylation at several
methylation hotspots on the mtDNA of senescent human heart
mesenchymal stem cells (HMSCs) (15). In the present study, using
bisulfite genomic sequencing (BSP), the methylation of the
COX2 gene was detected in aged HMSCs. In addition, the
effects of COX2 on cell senescence and proliferation were
investigated. The present study may provide a deeper understanding
of the mechanisms through which the mitochondria function during
the aging process.
Materials and methods
Cell culture
HMSCs were isolated and were a kind gift from Dr
Ji-Fan Hu of Stanford University (15). The study group of Dr Ji-Fan Hu
obtained the ethical approval and informed consent for the
isolation of the HMSCs. The characteristics of the HMSCs were also
measured and described in this previous study (15). HMSCs (G0) were cultured in normal
DMEM (Biological Industries, Inc.) containing 10% fetal bovine
serum (FBS, Biological Industries, Inc.) and antibiotics
(Biological Industries, Inc.) and incubated at 37°C in a 5%
CO2 humidified atmosphere.
5-Aza-2′-deoxycytidine (AdC, MCE Corporation) was
used as a DNA methylation inhibitor. AdC solution was prepared with
DMSO (Sigma-Aldrich; Merck KGaA) and the final working
concentration in the culture medium was 1 µM, as described
in a previous study (16).
Establishment of replicative senescence
(RS) and stress-induced premature senescence (SIPS) models
For the RS model, HMSCs at the G0 phase were
continuously cultured in normal DMEM in 10 cm dishes. Cells were
detached and passaged at the ratio of 1:2 when the confluence
reached 70%. The generation of HMSCs (Gn) was defined using the
cell passage number. A strategy was adopted by chronically exposing
HMSCs to low levels of oxidative stress (50 µM
H2O2; Sigma-Aldrich; Merck KGaA) and a low
serum environment (5% FBS) to establish the SIPS model, which was
also described in a previous study by the authors (15).
Staining of senescence-associated
β-galactosidase (SA-β-Gal)
Cellular senescence was visualized and quantified by
measuring the activity of β-Gal. HMSCs in a 6-well plate were fixed
with 3% formaldehyde (Beijing Chemical Works) for 3 min at room
temperature. After washing with PBS for 2×5 min, cell senescence
was determined with the Senescence β-Galactosidase Staining kit
according to the manufacturer's protocol (Beyotime Institute of
Biotechnology, Inc.). Following overnight incubation at 37°C, HMSCs
with the SA-β-Gal staining were assessed using a microscope-mounted
camera (IXplore, Olympus Corporation).
Detection of telomere length in senescent
HMSCs
The relative length of telomeres in the HMSCs was
measured by real-time fluorescence quantitative polymerase chain
reaction (qPCR) as previously described (17,18), with certain modifications.
Briefly, total cellular DNA was extracted using the Qiagen DNeasy
Blood & Tissue kit (Qiagen, Inc.). DNA was diluted to 35
ng/µl and was heated at 95°C for 5 min and chilled on ice
for 5 min. The DNA samples were then placed in a 20 µl qPCR
reaction system containing 10 µl 2X SYBR premixed buffer
(Genstar Technologies Company Inc.), and 2 µl forward and
reverse primers. The sequences of the primers were as follows:
Telomere forward, 5′-GGT TTT TGA GGG TGA GGG TGA GGG TGA GGG TGA
GGG T-3′ (100 nM) and reverse, 5′-TCC CGA CTA TCC CTA TCC CTT CCC
TAT CCC TAT CCC TA-3′ (300 nM); β-globin forward, 5′-GCT TCT GAC
ACA ACT GTG TTC ACT AGC -3′ (150 nM) and reverse, 5′-CAC CAA CTT
CAT CCA CGT TCA CC-3′ (150 nM). The PCR amplification process was
as follows: One cycle at 95°C for 10 min, 40 cycles at 95°C for 15
sec, 56°C for 30 sec, and 72°C for 30 sec (ABI SepOnePlus).
Telomere length was estimated by calculating the relative ratio
between the copies of telomere and the copies of β-globin. The qPCR
results were calculated using 2−ΔΔCq method (19).
Relative qualification of mtDNA copy
number
Total cellular DNA was extracted as described above.
The change in mtDNA copy number was evaluated by qPCR targeting the
β-globin gene and the mitochondrial ND1 gene as previously
described by Ding et al (20). Primers and probes were as follows:
For the β-globin gene: Forward, 5′-CTG GGC ATG TGG AGA CAG AGA AGA
CT-3′ and reverse, 5′-AGG CCA TCA CTA AAG GCA CCG AGC -3′; probe:
5′(FAM)-CCC TTA GGC TGC TGG TGG TCT ACCCTT-(TAMRA)3′. For
mitochondrial ND1 gene: Forward, 5′-GAC GCC ATA AAA CTC TTC ACC
AA-3′ and reverse, 5′-AGG TTG AGG TTG ACC AGG GG-3′; probe: 5′
(FAM)-CCA TCA CCC TCT ACA TCA CCG CCC -(TAMRA)3′. The reaction was
conducted in a 20 µl system containing 2 µl DNA (~70
ng), 10 µl Faststart TaqMan Probe Master (ROX) (Roche
Diagnostics, Inc.), 300 nM forward and reverse primers and 100 nM
TaqMan probe. The PCR amplification process was as follows: One
cycle at 95°C for 10 min, 40 cycles at 95°C for 15 sec and 60 °C
for 1 min. The qPCR results were calculated using the
2−ΔΔCq method.
Reverse transcription PCR (RT-PCR)
The age-related gene expression levels of
p16, p27, p53 and p21 were determined
by RT-PCR. Total RNA was extracted from the HMSCs using the
EasyPure RNA Purification kit (TransGen Biotech). In total, 1,000
ng RNA was reverse transcribed with TransScript All-in-One
First-Strand cDNA Synthesis SuperMix for qPCR (One-Step gDNA
Removal) (TransGen Biotech). The primers were as follows:
p16 forward, 5′-ACC AGA GGC AGT AAC CAT GC-3′ and reverse,
5′-GAC CTT CGG TGA CTG ATG ATC -3′; p27 forward, 5′-GCC TCA
GAA GA CGT CAA ACG T-3′ and reverse, 5′-TGT CCA TTC CAT GAA GTC AGC
-3′; p53 forward, 5′-CTC AGC ATC TTA TCC GAG TGG -3′ and
reverse, 5′-GCA GGA ACT GTT ACA CAT GTA G-3′; p21 forward,
5′-GTG GAC CTG TCA CTG TCT TGT AC-3′ and reverse, 5′-GCT TCC TCT
TGG AGA AGA TCA GC-3′; β-actin forward, 5′-CAG GTC ATC ACC
ATT GGC AAT GAG C-3′ and reverse, 5′-CGG ATG TCC ACG TCA CAC TTC
ATG A-3′. RT-PCR was conducted on an Eppendorf Mastercycler
(Eppendorf Shanghai International Trade Co. Ltd.) and was based on
the instructions provide with RealStar Green Fast Mixture
(GenStar). The reaction mixture contained the following: 2
µl cDNA, 4 µl 2X Taq PCR StarMix buffer and 2
µl of each primer. The thermocycling conditions were as
follows: Initial denaturation at 98°C for 3 min, followed by 25
cycles (for β-actin) or 31 cycles (for other genes) of 95°C
for 15 sec, 62°C for 15 sec and 72°C for 15 sec, and a final
extension step at 72°C for 5 min. The reaction products were
separated on a 3% agarose gel and visualized by GelStain
(TransGen). Densitometric analysis was performed using Quantity One
software (version 4.6; Bio-Rad Laboratories, Inc.).
Gene synthesis, plasmid construction and
transfection
The COX2 gene was optimized according to
nuclear format and commercially synthesized (Sangon Biotech
(Shanghai) Co., Ltd.). The synthesized gene was sub cloned into the
Sal1/Xho1 restriction site of the mitochondria
targeting plasmid vector, pCMV-myc-mito (Invitrogen; Thermo Fisher
Scientific, Inc.), to form a new pCMV-COX2-myc-mito plasmid. HMSCs
were transfected with 2 µg plasmid in a 6-well plate using
Lipofectamine 3000 reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. Empty vector was
also transfected into HMSCs as a negative control. Stable
transfectants were selected using neomycin (Invitrogen; Thermo
Fisher Scientific, Inc.) within 2 weeks, and after that, subsequent
experiments were carried out.
Protein extraction and western blot
analysis
Whole cell protein was extracted using the Protein
Extraction kit (Beyotime Institute of Biotechnology, Inc.). Equal
amounts of 25 µg protein from each sample were separated by
5-12% SDS polyacrylamide gel electrophoresis and were transferred
to 0.45 µm PVDF membranes (EMD Millipore). Membranes were
incubated in 5% BSA/TBST solution at 37°C for 1 h and washed in
TBST for 3×5 min. Membranes were then incubated with anti-COX2
(1:1,000 ab110258, Abcam), anti-myc-Tag (1:1,000, 2267, Cell
signaling Technology, Inc.) and anti-β-actin (1:3,000, ab8226,
Abcam) antibodies at 4°C overnight. Following incubation with the
primary antibodies, the membranes were incubated with HRP-coupled
goat anti-mouse secondary antibody (1:3,000, A0216, Beyotime
Institute of Biotechnology, Inc.) at room temperature for 1 h.
Immunoreactivity was detected using ECL luminescence reagent
[Sangon Biotech (Shanghai) Co., Ltd.]. Densitometric analysis was
performed using Quantity One software (version 4.6.2, Bio-Rad
Laboratories, Inc.).
MtDNA methylation analysis
HMSC mitochondria were isolated using the Tissue
Mitochondria Isolation kit (Beyotime Institute of Biotechnology,
Inc.) and mtDNA was extracted. mtDNA methylation was detected by
BSP. According to the manufacturer's protocol, 1,000 ng mtDNA in
total was used for bisulfite conversion with the EpiTect Bisulfite
kit (Qiagen, Inc.). The normal and converted COX2 mtDNA was
amplified by PCR reaction using the following primers: Forward
primer, 5′-ATT TTG TTA AAG TTA AAT TAT AGG -3′ and reverse primer,
5′-TTA ATT CTC TTA ATC TTT AAC TTA -3′. The thermocycling
conditions were as follows: Initial denaturation at 98°C for 5 min,
followed by 30 cycles of 95°C for 20 sec, 62°C for 20 sec and 72°C
for 100 sec, and a final extension step at 72°C for 10 min. The
reaction products were separated on a 1% agarose gel and recycled
using AxyPrep DNA Gel Extraction kit (Hangzhou Axygen
Biotechnology).
The recycled DNA molecules were linked with the
pJET1.2/blunt vector supplied with the CloneJET PCR Cloning kit
(K1231, Thermo Fisher Scientific, Inc.) and sequenced using Sanger
method [Sangon Biotech (Shanghai) Co., Ltd.]. The methylation
status of the 29th CpG site on the COX2 gene was analyzed.
The percentage of methylated CpG=the number of unconverted C in
CpG/the total number of CpG ×100%.
Cell proliferation assay
Cell proliferation was determined by WST-1 assay
(Beyotime Institute of Biotechnology, Inc.). Briefly,
5×103 HMSCs were planted into a well of 96-well plate
and cultured at 37°C for 24 h. According to the manufacturer's
instructions, 20 µl WST-1 were added to 200 µl cell
culture medium and incubated at 37°C in the dark for 2.5 h.
OD450 and OD630 were measured using a
microplate reader (BioTek Instruments, Inc.).
Statistical analysis
SPSS 19.0 (SPSS Inc.) was used for the analysis of
the data. All data were obtained from at least 3 independent
experiments and expressed as the means ± standard deviation (SD).
Statistical significance between groups was determined using the
Student's t-test or one-way ANOVA followed by the LSD or Tukey's
post hoc test. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
Senescence of HMSCs in the RS and SIPS
model
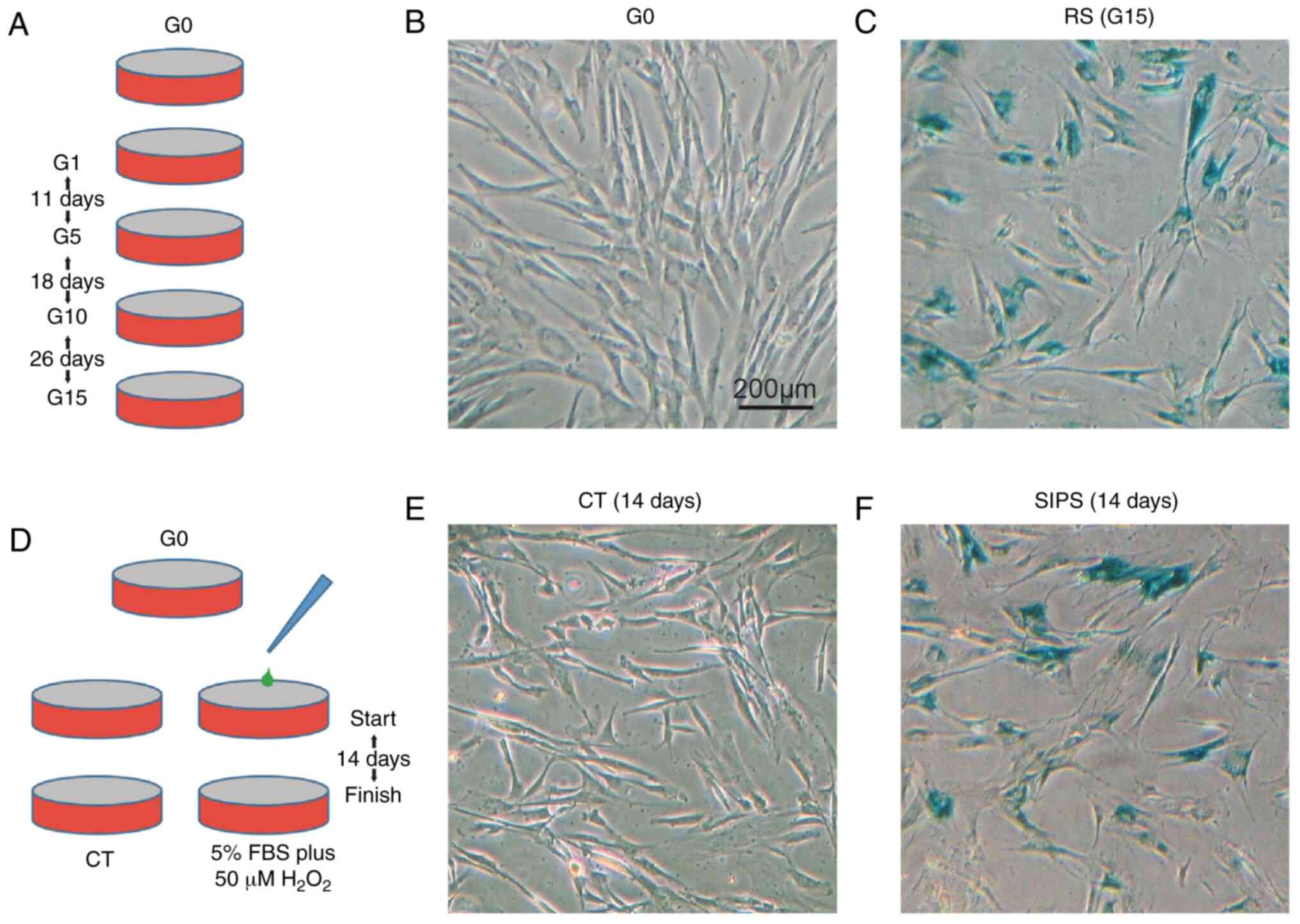
In the RS model, the HMSCs were continually cultured
from G0 to G15, lasting for 55 days (Fig. 1A). In the SIPS model, the HMSCs
were cultured in low FBS plus 50 µM
H2O2 for 14 days (Fig. 1D). The HMSCs from the G0/G15 and
CT/SIPS groups were stained for SA-β-Gal. As shown in Fig. 1B, C, E and F, the HMSCs from the
G15 and SIPS groups exhibited a typical senescence-like morphology
and stained positive for β-Gal.
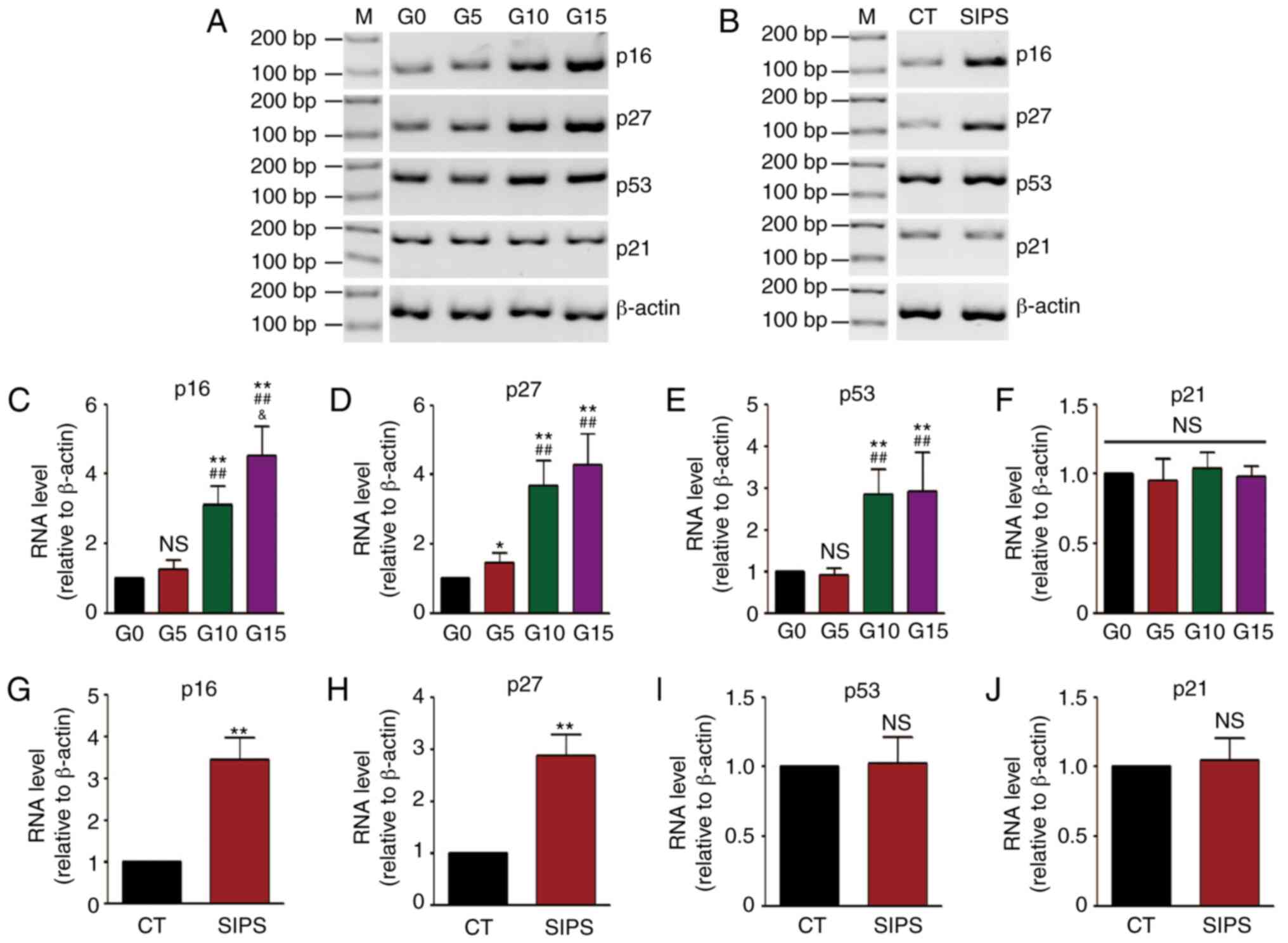
Subsequently, age-related gene expression levels
were detected. The mRNA levels of 4 genes, including p16,
p27, p53 and p21 were measured by RT-PCR. The
results of RT-PCR of the HMSCs in the RS and SIPS models are shown
in Fig. 2A and B, and the results
of densitometric analysis of RT-PCR are shown in Fig. 2C-J. It was found that the mRNA
levels of the p16, p27 and p53 genes were
increased along with cell senescence in the RS model, while the
p16 and p27 levels were increased in the SIPS
model.
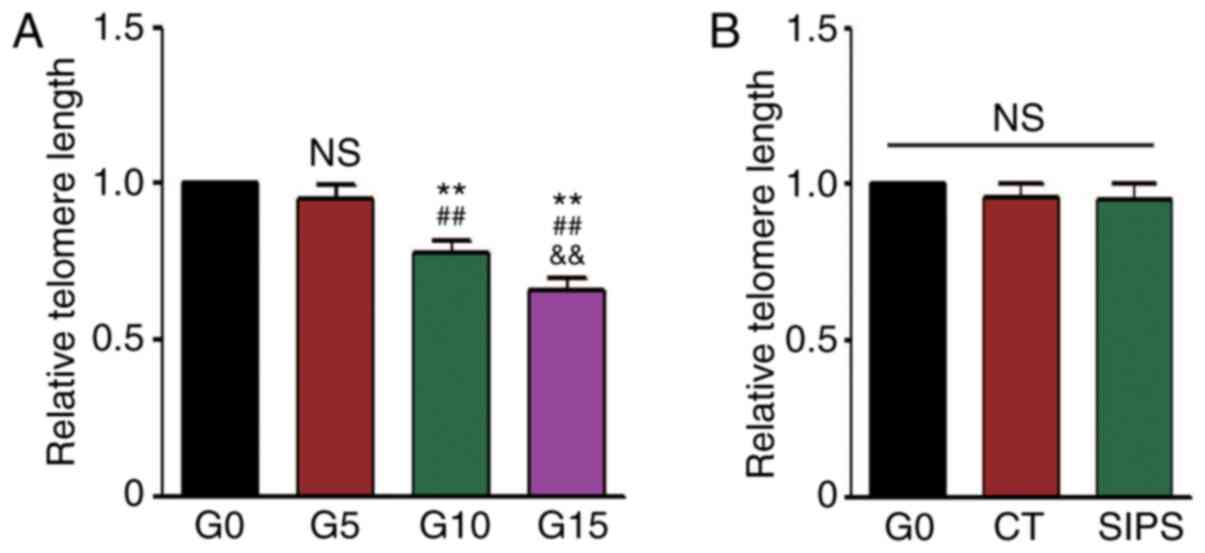
Since telomere length is considered a biomarker of
chronological aging (21), the
present study also detected this the RS and SIPS models. The
results of qPCR revealed that the telomere length of the HMSCs
gradually became shorter in the RS model (Fig. 3A), but not in the SIPS model
(Fig. 3B).
COX2 methylation and protein expression
in senescent HMSCs
It has been reported that COX plays a critical role
in age-associated cell dysfunction (10). In a previous study, the authors
demonstrated that COX1 methylation decreased and COX1 mRNA
expression was significantly upregulated following senescence
(15). The present study wished
to determine the mechanisms through COX2 mtDNA methylation is
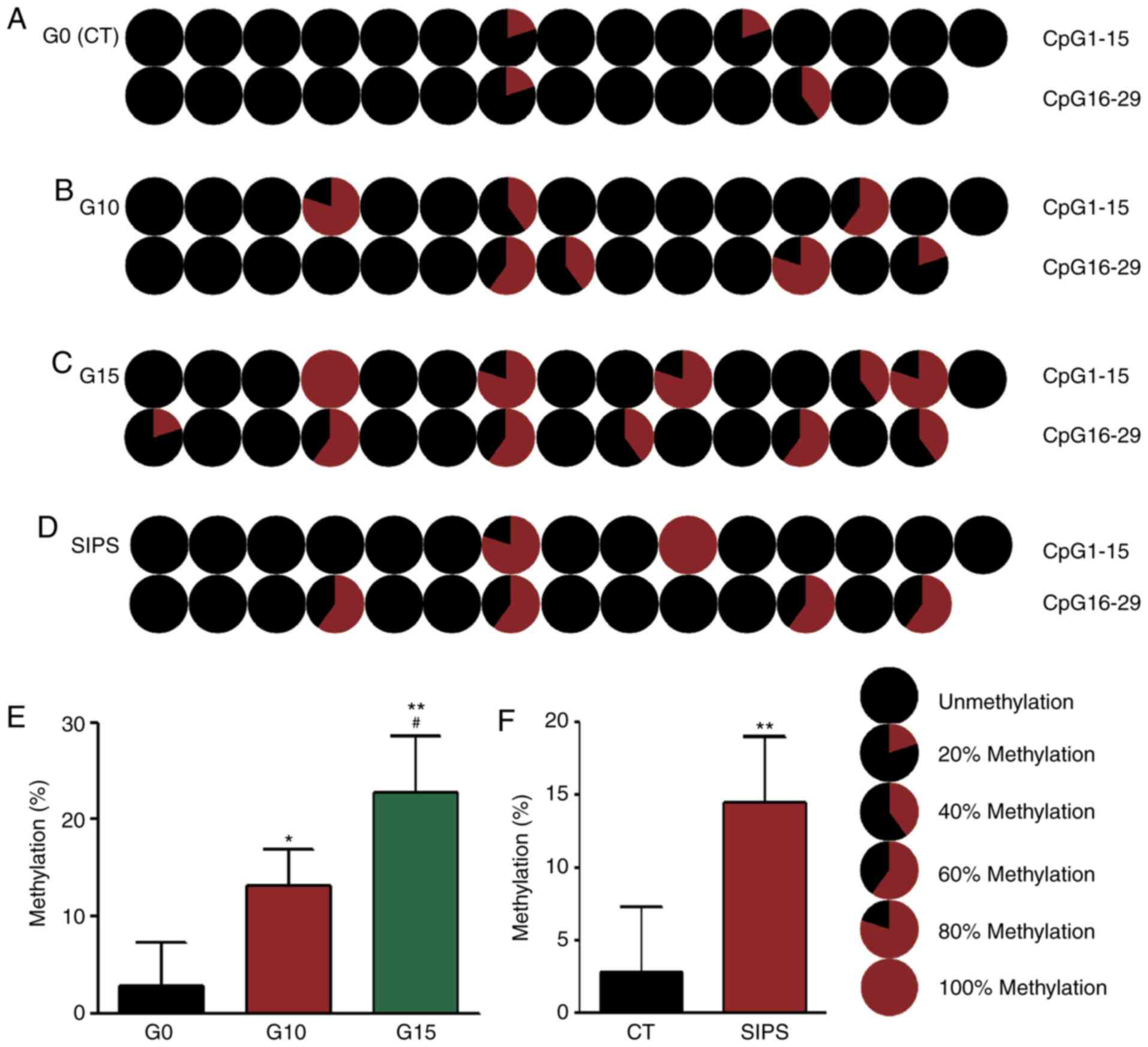
altered in senescent HMSC. It was found that in the RS model, the
results of BSP revealed that the methylation rate of the G0 HMSCs
was 2.76% (Fig. 4A), that of G10
HMSCs was 13.1% (Fig. 4B) and
that of G15 HMSCs was 22.76% (Fig.
4C), while in the SIPS model, the methylation rate of the
senescent HMSCs was 14.48% (Fig.
4D).
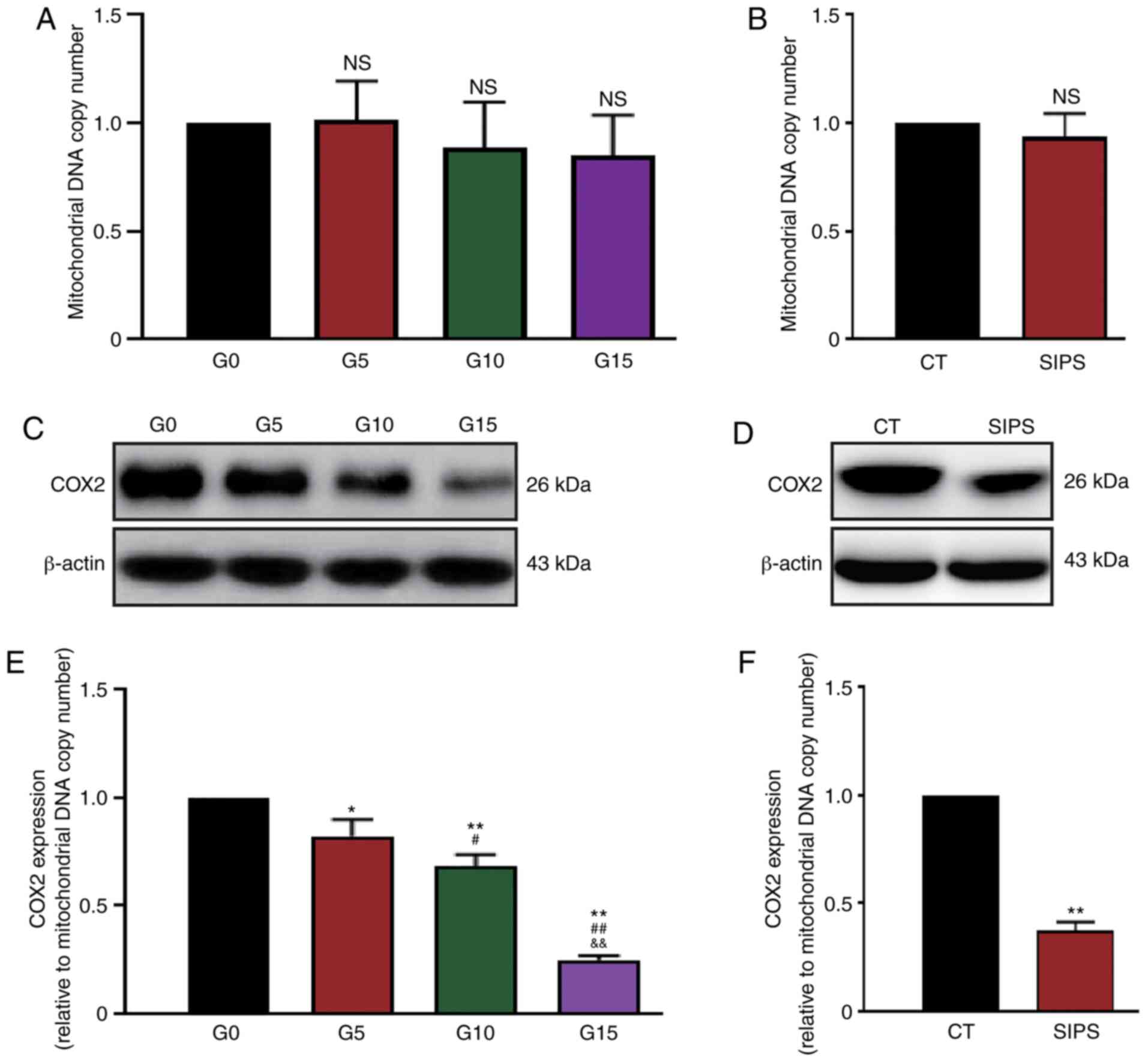
Subsequently, the expression status of the
COX2 gene was analyzed. Considering that the mitochondria
number may change in cells and thus affect protein quantity, the
mtDNA copy number was first measured by qPCR. It was found that
although the mtDNA copy numbers in G10, G15 and SIPS HMSCs
decreased slightly, this decrease was not significant (Fig. 5A and B). The present study then
detected the protein levels of COX2 in the RS and SIPS models by
western blot analysis (Fig. 5C and
D). The expression levels of COX2 were then evaluated using the
ratio between the gray value of blots and the mtDNA copy number. As
shown in Fig. 5E and F, the
results indicated that COX2 expression decreased along with HMSC
senescence in both the RS and SIPS models.
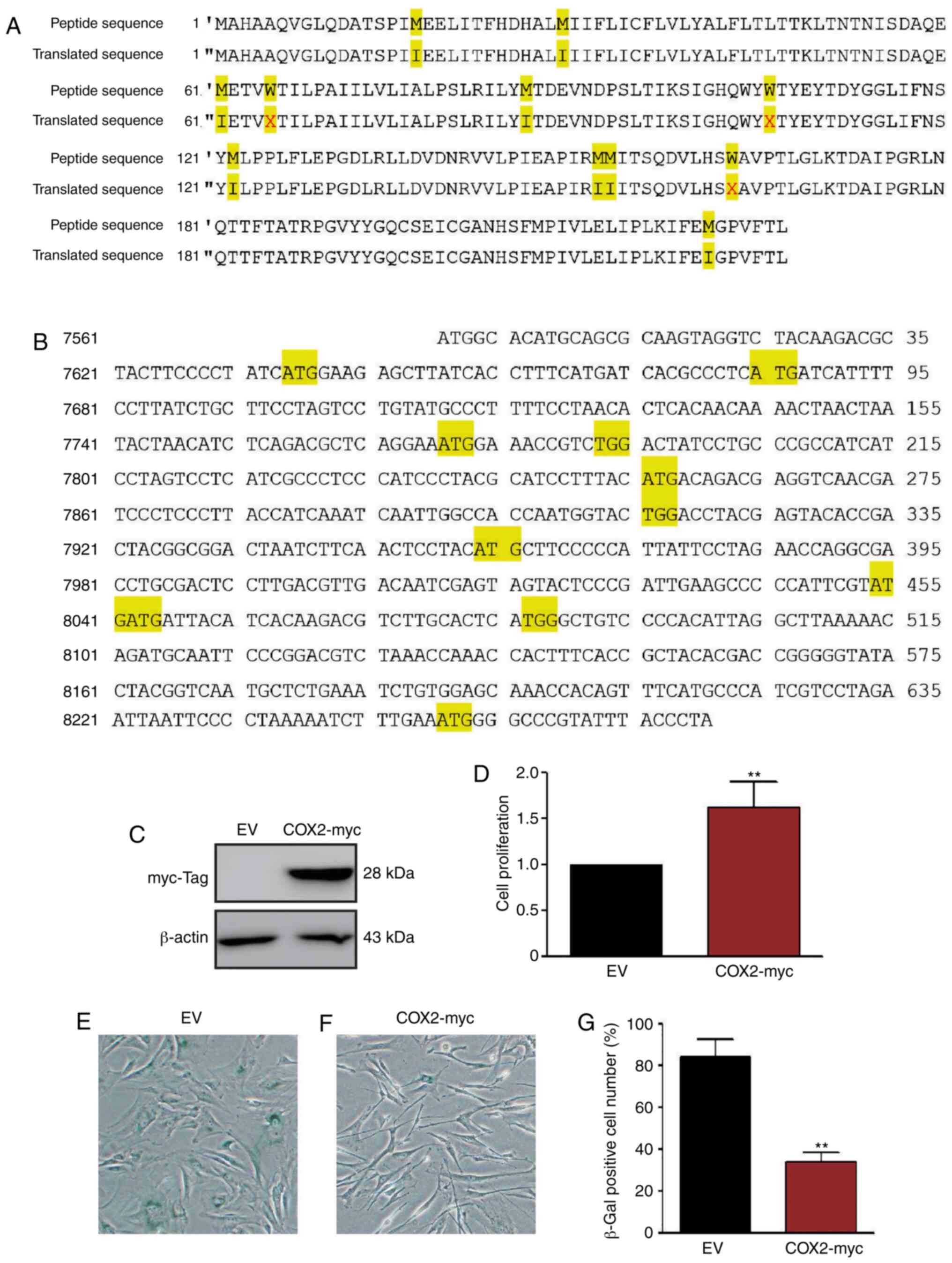
Overexpression of COX2 delays HMSC
senescence
Since the COX2 level decreased along with cell
senescence, exogenous COX2 protein was introduced into HMSC to
determine whether it delays the aging process of HMSCs. The COX2
expression plasmid, pCMV-COX2-myc-mito (Fig. 6A and B), was transfected into the
G1 HMSCs and stable transfectants were selected. The transfection
efficiency was confirmed by western blot analysis using anti-myc
antibodies (Fig. 6C). Stable
HMSCs were also cultured in 5% FBS and 50 µM
H2O2 conditioned medium for 14 days. The
results of WST-1 assay revealed that the COX2 transfectants grew at
a faster rate than the normal HMSCs (Fig. 6D). SA-β-Gal staining revealed that
the overexpression of COX2 significantly delayed the aging process
of HMSCs (Fig. 6E-G).
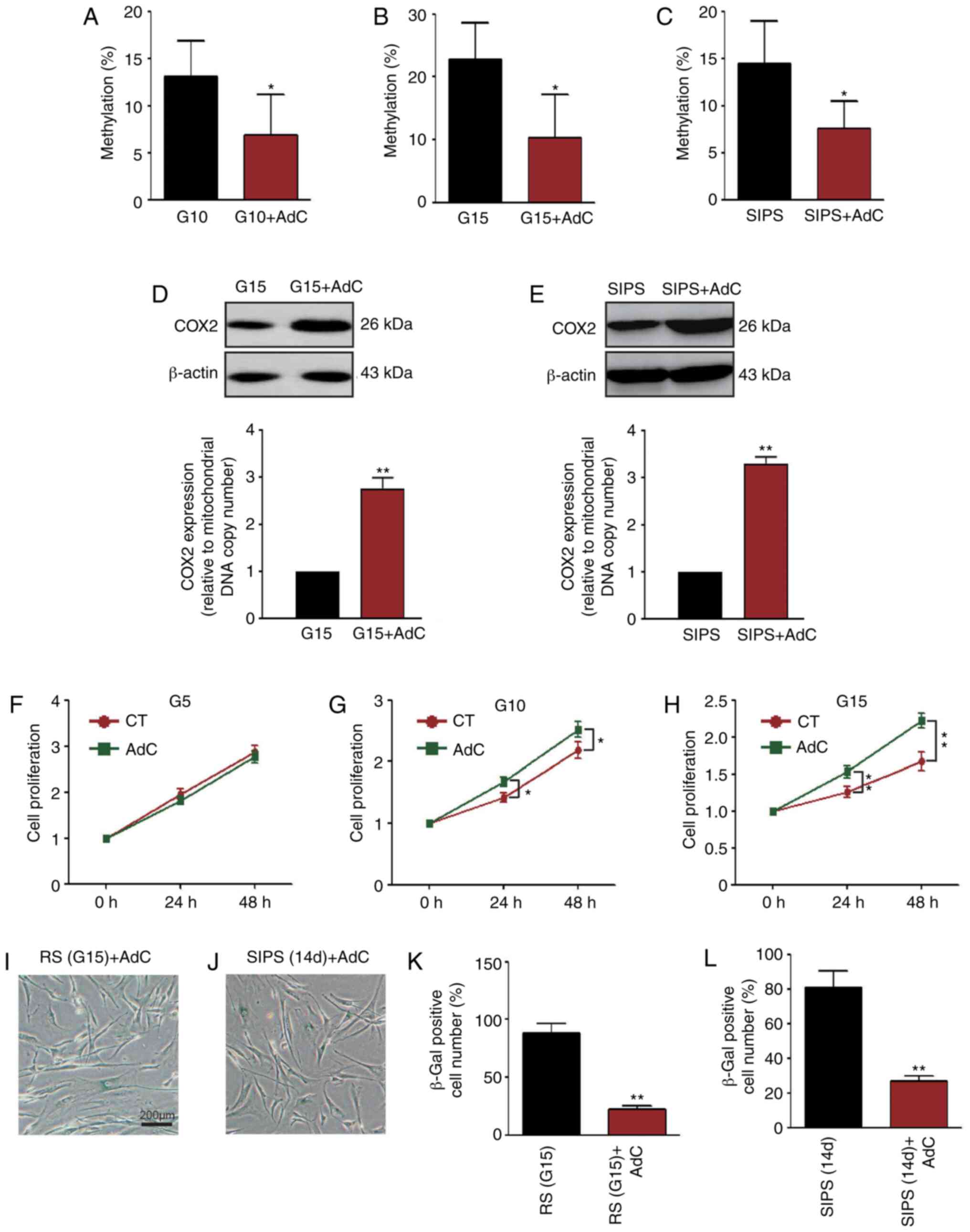
AdC decreases COX2 methylation and
attenuates HMSC senescence
To determine whether COX2 mtDNA methylation is
associated with cellular senescence, HMSCs were treated with AdC.
As shown in Fig. 7A-C, COX2 mtDNA
methylation in the G10, G15 and SIPS HMSCs was decreased
(P<0.05, pie charts are not shown). The COX2 protein expression
levels were increased simultaneously (P<0.01, Fig. 7D and E; the mtDNA copy number is
not shown). It was also found that AdC treatment significantly
increased the proliferation of the senescent HMSC (Fig. 7F-H). Importantly, SA-β-Gal
staining revealed that AdC treatment also delayed the aging process
of the HMSCs (Figs. 1C and F, and
7I-L).
Discussion
Due to increasing longevity and declining fertility,
it has been predicted that almost one in four individuals will be
65 years or older by the year 2035 (22). There is a consensus that age is a
dominant risk factor for the development of cardiac diseases, which
is the leading cause of mortality worldwide (23). Therefore, there is an urgent need
to elucidate the molecular mechanisms of age-related cardiac
diseases and to develop novel intervention techniques.
In the present study, two types of aging models were
constructed, RS and SIPS. RS is a state where normal somatic cells
lose their replicative capacity in vitro and reach the
Hayflick limit (24). SIPS is
often used as a model to study aging as it shares the
characteristics of senescent cells, such as increased levels of
oxidative DNA damage, accelerated telomere shortening and changes
in cell-cycle regulation (25).
However, there are still some differences between RS and SIPS, such
as phenotypes of protein and gene expression (26). In the present study, 4 age-related
gene expression profiles were compared and it was found that the
trends of p16, p27 and p21 were consistent
between these two models, whereas that of p53 was not.
Additionally, telomere length shortening only occurred in RS and
not in SIPS. These data suggest that although RS and SIPS share
hallmarks of cellular senescence, such as β-Gal staining
positivity, they still differ in some cytological features.
Although the exact mechanisms responsible for
senescence are not currently fully understood, there is increasing
evidence to suggest that the mitochondria act as central regulators
of the aging process (27). In
cardiac cells, which are the most energy-consuming cells in the
human body, approximately one third of the cell volume is occupied
by mitochondria (28). Similar to
the methylation taking place in CpG sequences of nuclear DNA,
methylation has also been discovered in mtDNA (29). Despite recent progress in the
field of mtDNA methylation and its possible contribution to
age-related diseases, clear-cut evidence of its functionality is
still lacking (30).
In the present study, to reveal the mtDNA
methylation status during the cell senescence process, a
mitochondrial redox reaction-related gene, COX2, was selected, and
its methylation levels were measured in both the RS and SIPS
models. The results indicated that in the young G0 HMSCs, COX2
mtDNA exhibited a very low CpG methylation rate of 2.76%. Following
either continuous passaging or H2O2 exposure,
the COX2 methylation levels in senescent HMSCs were significantly
increased to 22.76 and 14.48%. In young G0 HMSCs, there were 4
methylated CpG sites (4/29, Fig.
4), whereas in the G15 and SIPS HMSCs, the number of sites
increased to 11 (11/29, Fig. 4)
and 6 (6/29, Fig. 4). These data
clearly indicate that COX2 methylation is a feature of the
senescence of HMSCs. A previous study provided evidence that mtDNA
methylation occurs not just at CpG sites: Dinucleotides CpT, CpA
and CpC were also methylated (31). Due to the limitations of
technology and equipment, these data were not in the present study.
However, the authors are still interested in investigating this
phenomenon in future studies.
Mitochondrial gene expression is closely associated
with mitochondria functionality. In the present study, it was
demonstrated that the expression level of COX2 decreased following
the senescence of HMSCs, and the addition of extraneous COX2
apparently delayed the aging process and promoted cell
proliferation. These data suggest that the COX2 level is essential
for maintaining HMSCs young. Normally, the D-loop is critical for
the transcription and replication of mtDNA; thus, D-loop
methylation is logically considered to control the mitochondrial
gene expression (32). However,
Ghosh et al demonstrated that methylcytosines distribute
constantly across the mitochondria and that the methylcytosines
near gene start sites can be temporally regulated, suggesting a
potential regulatory role for the mitochondrial genome (33). Herein, it was demonstrated that
there were 4 methylation hotspots on the COX2 gene during
the aging process: The seventh CpG, the 22nd CpG, the 27th CpG and
the 29th CpG. However, to reveal whether these CpGs methylation are
associated with COX2 downregulation and how they regulate
COX2 expression, further investigations are warranted.
To further reveal the association between mtDNA
methylation and cell senescence, AdC was used to treat the HMSCs.
AdC is an inhibitor of Dnmt1, which has a mitochondrial targeting
sequence and is the major enzyme responsible for the maintenance of
DNA methylation (34). It was
demonstrated that the administration of AdC may decrease the
methylation and expression of COX2. Moreover, AdC also promoted
proliferation and delayed the senescence of HMSCs. Mishra and
Kowluru demonstrated that Dnmt1 binds directly at the D-loop and
other regions (e.g., cytochrome b) of mtDNA and affects the
transcription of mitochondrial genes, including COX2 (34). The present study yielded similar
results to the study by Mishra and Kowluru. However, based on the
current data, it still cannot be determined if the decrease in the
COX2 level is regulated by the methylation of the D-loop or
the methylation of itself, or both. Therefore, this unresolved
issue will be the focus of future studies.
Funding
The present study was supported in part by grants
from the Jilin International Collaboration Grant (no. 20180414065GH
to DY and 20190701040GH to ZL), the Subject Arrangement Program
from Science and Technology Department of Jilin Province (no.
20200201123JC to DY), the Youth Development Programme of Health
Commission of Jilin Province (no. 2019Q007 to DY), the Health
Special Fund from Jilin Province Department of Finance (no.
2018SCZWSZX-048 to XS), the National Natural Science Foundation of
China (no. 81701270 to YL), the Transformation Fund of the First
Hospital of Jilin University (no. JDYYZH-1902033 to YL), the China
Postdoctoral Science Foundation (no. 171251 to LR), the
Science-technology Foundation of the Jilin Provincial Education
Department (no. JJKH20170823KJ to LR) and the Norman Bethune
Program of Jilin University (no. 2015326 to LR).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DY contributed to the conception and design of the
study. XS, ZW, XC and YL contributed to the acquisition, analysis
and interpretation of the data. DY drafted the article. ZL and LR
analyzed the data. TY was involved in performing the experiments.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank Dr Ji-Fan Hu and
Professor Andrew R. Hoffman in Stanford University for providing
technical advice on HMSC identification and culturing.
References
|
1
|
Lakatta EG and Levy D: Arterial and
cardiac aging: Major shareholders in cardiovascular disease
enterprises: Part II: The aging heart in health: Links to heart
disease. Circulation. 107:346–354. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Soro-Arnaiz I, Li QOY, Torres-Capelli M,
Meléndez-Rodríguez F, Veiga S, Veys K, Sebastian D, Elorza A, Tello
D, Hernansanz-Agustín P, et al: Role of mitochondrial complex IV in
age-dependent obesity. Cell Rep. 16:2991–3002. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Boengler K, Kosiol M, Mayr M, Schulz R and
Rohrbach S: Mitochondria and ageing: Role in heart, skeletal muscle
and adipose tissue. J Cachexia Sarcopenia Muscle. 8:349–369. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Frezza C: The role of mitochondria in the
oncogenic signal transduction. Int J Biochem Cell Biol. 48:11–17.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Corsetti G, Pasini E, D'Antona G, Nisoli
E, Flati V, Assanelli D, Dioguardi FS and Bianchi R: Morphometric
changes induced by amino acid supplementation in skeletal and
cardiac muscles of old mice. Am J Cardiol. 101:E26–E34. 2008.
View Article : Google Scholar
|
|
6
|
Cheng Z, Ito S, Nishio N, Thanasegaran S,
Fang H and Isobe K: Characteristics of cardiac aging in C57BL/6
mice. Exp Gerontol. 48:341–348. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mozet C, Martin R, Welt K and Fitzl G:
Cardioprotective effect of EGb 761 on myocardial ultrastructure of
young and old rat heart and antioxidant status during acute
hypoxia. Aging Clin Exp Res. 21:14–21. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
El'darov ChM, Vays VB, Vangeli IM,
Kolosova NG and Bakeeva LE: Morphometric examination of
mitochondrial ultrastructure in aging cardiomyocytes. Biochemistry
(Mosc). 80:604–609. 2015. View Article : Google Scholar
|
|
9
|
Müller-Höcker J, Schäfer S, Weis S,
Münscher C and Strowitzki T: Morphological-cytochemical and
molecular genetic analyses of mitochondria in isolated human
oocytes in the reproductive age. Mol Hum Reprod. 2:951–958. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xin MG, Zhang J, Block ER and Patel JM:
Senescence-enhanced oxidative stress is associated with deficiency
of mitochondrial cytochrome c oxidase in vascular endothelial
cells. Mech Ageing Dev. 124:911–919. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Abramson J, Svensson-Ek M, Byrne B and
Iwata S: Structure of cytochrome c oxidase: A comparison of the
bacterial and mitochondrial enzymes. Biochim Biophys Acta.
1544:1–9. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yoshikawa S, Shinzawa-Itoh K and Tsukihara
T: X-ray structure and the reaction mechanism of bovine heart
cytochrome c oxidase. J Inorg Biochem. 82:1–7. 2000. View Article : Google Scholar
|
|
13
|
Stoccoro A, Siciliano G, Migliore L and
Coppede F: Decreased methylation of the mitochondrial D-loop region
in late-onset Alzheimer's disease. J Alzheimers Dis. 59:559–564.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zheng LD, Linarelli LE, Brooke J, Smith C,
Wall SS, Greenawald MH, Seidel RW, Estabrooks PA, Almeida FA and
Cheng Z: Mitochondrial epigenetic changes link to increased
diabetes risk and early-stage prediabetes indicator. Oxid Med Cell
Longev. 2016:52906382016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu D, Du Z, Pian L, Li T, Wen X, Li W, Kim
SJ, Xiao J, Cohen P, Cui J, et al: Mitochondrial DNA
hypomethylation is a biomarker associated with induced senescence
in human fetal heart mesenchymal stem cells. Stem Cells Int.
2017:17645492017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Manev H and Uz T: DNA hypomethylating
agents 5-aza-2′-deoxycytidine and valproate increase neuronal
5-lipoxygenase mRNA. Eur J Pharmacol. 445:149–150. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Saferali A, Lee J, Sin DD, Rouhani FN,
Brantly ML and Sandford AJ: Longer telomere length in COPD patients
with α1-antitrypsin deficiency independent of lung function. PLoS
One. 9:e956002014. View Article : Google Scholar
|
|
18
|
el Bouazzaoui F, Henneman P, Thijssen P,
Visser A, Koning F, Lips MA, Janssen I, Pijl H, Willems van Dijk K
and van Harmelen V: Adipocyte telomere length associates negatively
with adipocyte size, whereas adipose tissue telomere length
associates negatively with the extent of fibrosis in severely obese
women. Int J Obes (Lond). 38:746–749. 2014. View Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
20
|
Ding Y, Xia BH, Zhang CJ and Zhuo GC:
Mutations in mitochondrial tRNA genes may be related to insulin
resistance in women with polycystic ovary syndrome. Am J Transl
Res. 9:2984–2996. 2017.PubMed/NCBI
|
|
21
|
Rizvi S, Raza ST and Mahdi F: Telomere
length variations in aging and age-related diseases. Curr Aging
Sci. 7:161–167. 2014. View Article : Google Scholar
|
|
22
|
Lakatta EG and Sollott SJ: Perspectives on
mammalian cardio-vascular aging: Humans to molecules. Comp Biochem
Physiol A Mol Integr Physiol. 132:699–721. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hashimoto H, Olson EN and Bassel-Duby R:
Therapeutic approaches for cardiac regeneration and repair. Nat Rev
Cardiol. 15:585–600. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hayflick L and Moorhead PS: The serial
cultivation of human diploid cell strains. Exp Cell Res.
25:585–621. 1961. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aan GJ, Hairi HA, Makpol S, Rahman MA and
Karsani SA: Differences in protein changes between stress-induced
premature senescence and replicative senescence states.
Electrophoresis. 34:2209–2217. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kural KC, Tandon N, Skoblov M,
Kel-Margoulis OV and Baranova AV: Pathways of aging: Comparative
analysis of gene signatures in replicative senescence and stress
induced premature senescence. BMC Genomics. 17(Suppl 14):
S10302016. View Article : Google Scholar
|
|
27
|
Payne BA and Chinnery PF: Mitochondrial
dysfunction in aging: Much progress but many unresolved questions.
Biochim Biophys Acta. 1847:1347–1353. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bravo-Sagua R, Parra V, López-Crisosto C,
Díaz P, Quest AF and Lavandero S: Calcium transport and signaling
in mitochondria. Compr Physiol. 7:623–634. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Vanyushin BF and Kirnos MD: Structure of
animal mitochondrial DNA (base composition, pyrimidine clusters,
character of methylation). Biochim Biophys Acta. 475:323–336. 1977.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zinovkina LA and Zinovkin RA: DNA
methylation, mitochondria, and programmed aging. Biochemistry
(Mosc). 80:1571–1577. 2015. View Article : Google Scholar
|
|
31
|
Bellizzi D, D'Aquila P, Scafone T,
Giordano M, Riso V, Riccio A and Passarino G: The control region of
mitochondrial DNA shows an unusual CpG and non-CpG methylation
pattern. DNA Res. 20:537–547. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
van der Wijst MG, van Tilburg AY, Ruiters
MH and Rots MG: Experimental mitochondria-targeted DNA methylation
identifies GpC methylation, not CpG methylation, as potential
regulator of mitochondrial gene expression. Sci Rep. 7:1772017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ghosh S, Sengupta S and Scaria V:
Comparative analysis of human mitochondrial methylomes shows
distinct patterns of epigenetic regulation in mitochondria.
Mitochondrion. 18:58–62. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mishra M and Kowluru RA: Epigenetic
modification of mitochondrial DNA in the development of diabetic
retinopathy. Invest Ophthalmol Vis Sci. 56:5133–5142. 2015.
View Article : Google Scholar : PubMed/NCBI
|