Introduction
Atherosclerosis (AS) is a chronic cardiovascular
disease characterized by high morbidity and mortality rates
worldwide. AS can lead to the thickening and hardening of the walls
of arteries, and to the formation of atherosclerotic plaques
containing immune cells, mesenchymal cells and lipids. Once the
unstable atherosclerotic plaques are ruptured, this may then
contribute to thrombosis and may interrupt blood flow. At present,
it has been found that the formation of atherosclerotic plaques
involves multiple factors, including the presence of lipid
metabolism disorders, vascular endothelial cell injury, vascular
smooth muscle cell (VSMC) proliferation, platelet adhesion and
others (1). VSMC proliferation
has been revealed to play a pivotal role in the pathogenesis of AS,
as VSMCs are closely associated with the key initiating factor,
oxidized low-density lipoprotein (ox-LDL), which induces the
phenotypic trans-formation of contractile VSMCs into synthetic
VSMCs (2,3). Synthetic VSMCs proliferate, migrate
and produce collagen more easily than contractile VSMCs (4,5),
thus accelerating the progression of AS. Thus, it remains of utmost
priority to identify the molecular mechanisms regulating the
abnormal proliferation and migration of VSMCs.
Thousands of non-coding RNAs (ncRNAs), including
long ncRNAs (lncRNAs), microRNAs (miRNAs/miRs) and small
interfering RNAs (siRNAs), have been identified in the human genome
(6). Each type of ncRNA has been
identified to be involved in the progression of numerous types of
disease, which provides a potential promising approach for the
treatment of AS (7-10). lncRNAs are ncRNAs of >200 bp in
length (11). A previous study
revealed that lncRNAs played an important role in regulating the
proliferation and migration of VSMCs (12-14). In particular, the lncRNA, small
nucleolar RNA host gene 7-003 (SNHG7-003), has been observed to
inhibit the lipopolysaccharide-induced activation of the NF-κB
signaling pathway and regulate the inflammatory response in human
monocytes and macrophages, which were downregulated in the blood of
patients with coronary artery disease with unstable plaques; it was
therefore indicated to be a potential novel lncRNA biomarker for
the diagnosis of unstable plaques (15).
Additional previous studies have indicated that
miR-1306-5p may play a role in numerous types of cardiovascular
disease. For example, miR-1306 expression levels were discovered to
be upregulated in a miRNA expression profile of AS obliterans
samples (16-18). Furthermore, a previous study by
the authors confirmed that sirtuin 7 (SIRT7) modulated VSMC
proliferation and migration via the Wnt/β-Catenin signaling pathway
(19), and binding sites between
miR-1306-5p and SIRT7 were predicted using the StarBase v2.0
database. However, to the best of our knowledge, the effects of
SNHG7-003, miR-1306-5p and SIRT7 on VSMC have not yet been studied
and remain unknown. Thus, the present study aimed to investigate
the role of SNHG7-003 in VSMC proliferation and migration, as well
as to clarify the potential underlying mechanisms.
Materials and methods
Cells, cell culture and treatment
VSMCs were obtained from the China Center for Type
Culture Collection. VSMCs were cultured in DMEM (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.), and maintained in a humidified incubator
with 95% air and 5% CO2 at 37°C. When cells grew to a
certain density, cells in logarithmic phase were digested and
passaged with 0.25% trypsin-EDTA (Gibco; Thermo Fisher Scientific,
Inc.). Referring to a previous study (19), to establish the cell model of AS,
VMSCs were incubated with 25, 50 and 100 µg/ml ox-LDL
(Yiyuan Biotechnologies) for 24 h and the most suitable
concentrations of ox-LDL were screened.
Bioinformatics prediction
The Starbase v2.0 website (http://starbase.sysu.edu.cn) was used to predict the
potential miR-lncRNA binding partners and the downstream target
genes of these miRs.
Cell transfection
The pcDNA-SNHG7-003 (SNHG7-003) and its blank
control pcDNA-NC (vector) were constructed by Shanghai GenePharma
Co. Ltd. The miR-1306-5p mimic and miR-NC were purchased from
Guangzhou RiboBio Co.Ltd. The SIRT7 overexpression plasmid (SIRT7)
and overexpression negative control (OE-NC) were obtained from
BioVector. VSMCs in the logarithmic growth phase were seeded into
6-well plates and upon reaching 60-70% confluence, were transfected
with the vectors or mimics and the respective NCs at a final
concentration of 100 nM using Lipofectamine® 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) for 8 h,
according to the manufacturer's protocol. The transfection
efficiency was determined using reverse transcription-quantitative
PCR (RT-qPCR) or western blot analysis after 48 h. Cells were
randomly assigned to the following groups: The control group,
ox-LDL group, ox-LDL + vector group, ox-LDL + SNHG7-003 group,
ox-LDL + SNHG7-003 + miR-NC group, ox-LDL + SNHG7-003 + miR-1306-5p
group, ox-LDL + SNHG7-003 + miR-1306-5p + OE-NC group and ox-LDL +
SNHG7-003 + miR-1306-5p + SIRT7 group.
RT-qPCR
Total RNA was extracted using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and reverse
tran-scribed into cDNA using the QuantiTect RT kit (Qiagen),
according to the manufacturer's protocol. qPCR was subsequently
performed using an Applied Biosystems 7500 Real-Time PCR system
(Thermo Fisher Scientific, Inc.). The following primers were used
for the qPCR: SNHG7-003 forward, 5′-CTT CGC CTG TGA TGG ACT TC-3′
and reverse, 5′-CCT GCC CAT CCC TTT ATT CC-3′; and GAPDH forward,
5′-CTC ACC GGA TGC ACC AAT GTT -3′ and reverse, CGC GTT GCT CAC AAT
GTT CAT -3′. Total RNA was extracted and reverse transcribed into
cDNA using the miRNAeasy Mini kit and miScript II RT kit (Qiagen
GmbH), respectively. miR-1306-5p expression levels were detected
using specific primers for miRNAs (Qiagen GmbH). The mRNA and miRNA
expression levels were quantified using the 2−ΔΔCq
method (20) and normalized to
GAPDH or the small nuclear RNA U6, respectively as previously
described.
Cell viability assay
A Cell Counting kit-8 (CCK-8; Beyotime Institute of
Biotechnology, Inc.) assay was used to analyze the proliferation of
the VSMCs, according to the manufacturer's protocol. Briefly, at 24
h post-transfection, the VSMCs were plated into a 96-well plate at
a density of 2×103 cells/well. Following 24 h of
incubation, 10 µl CCK-8 solution were added to each well and
incubated for further 4 h. A microplate reader (Bio-Rad,
Laboratories, Inc.) was used to measure the absorbance at 450
nm.
Wound-healing assay
A wound-healing assay was used to analyze the
migratory ability of the VSMCs. Briefly, 1×105
transfected VSMCs were seeded into 12-well plates containing
RPMI-1640 medium without FBS and were cultured to 80% confluence.
Subsequently, the cell monolayer was scratched with a 200-µl
plastic pipette tip. The cells were washed 3 times with PBS to
remove the suspended cells prior to incubation in serum-free medium
in a 12-well plate for 24 h in an incubator. The wound was
photographed (magnification, ×200) at 0 and 24 h using an inverted
microscope (Olympus Corporation) and the extent of wound closure
was semi-quantified using ImageJ software 2.0 (National Institutes
of Health) at 0 and 24 h.
Transwell assay
The cell invasive ability was analyzed using a
Transwell chamber (Corning, Inc.). Briefly, 1×106
transfected cells suspended in free-serum medium were seeded into
the upper chamber of the plate pre-coated with Matrigel (Corning,
Inc.) for 30 min at 37°C (2 mg/ml; 15 µl). RPMI-1640 medium
supplemented with 10% FBS was added to the lower chambers.
Following incubation for 24 h at 37°C, the invasive cells were
fixed with methanol and stained with 0.1% crystal violet (Sigma
Aldrich; Merck KGaA) for 30 min at room temperature. Finally, the
cells were visualized under a light microscope (magnification,
×100; Olympus Corporation).
Western blot analysis
Total protein was extracted from the cells using
RIPA lysis buffer (Beyotime Institute of Biotechnology), according
to the manufacturer's protocol. Total protein was quantified using
the BCA method and the protein samples were separated via 10%
SDS-PAGE (Beyotime Institute of Biotechnology). A total of 30
µg separated protein/lane was subsequently transferred onto
PVDF membranes (EMD Millipore) for different periods of time
according to the different molecular weight of the proteins and
then blocked with 5% skimmed milk overnight at 4°C. The membranes
were incubated at 4°C overnight with the following primary
antibodies: Anti-α-SMA (1:1,000; cat. no. 19245; Cell Signaling
Technology, Inc.), anti-MMP-2 (1:1,000; cat. no. 40994; Cell
Signaling Technology, Inc.), anti-MMP-9 (1:1,000; cat. no. 13667;
Cell Signaling Technology, Inc.), anti-SIRT7 (1:1,000; cat. no.
5360; Cell Signaling Technology, Inc.) and anti-GAPDH (1:1,000;
cat. no. 5174; Cell Signaling Technology, Inc.). Following
incubation with the primary antibodies, the membranes were
incubated with HRP-conjugated goat anti-rabbit secondary antibody
(1:5,000; cat. no. A0208; Beyotime Institute of Biotechnology) for
1 h at room temperature. Protein bands were visualized using an ECL
detection kit (Bio-Rad Laboratories, Inc.) and the expression
levels were analyzed using Image Lab software (Bio-Rad
Laboratories, Inc.).
Dual luciferase reporter assay
SNHG7-003-wild-type (WT) and SNHG7-003-mutant (Mut),
SIRT7-WT and SIRT7-Mut vectors were obtained from Promega
Corporation. SNHG7-003-WT was constructed using the luciferase
reporter vector containing SNHG7-003 sequences and the predicted
miR-1306-5p binding site. SNHG7-003 with a Mut sequence was
synthesized and subcloned into the vector, yielding SNHG7-003-Mut.
The construction of the SIRT7-WT and SIRT7-Mut vectors was
identical to the protocol described above. Subsequently, VSMCs were
co-transfected with 50 ng SNHG7-003-WT/Mut (or SIRT7-WT/Mut) and
200 ng miR-1306-5p mimic or miR-NC using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
provided protocol. The relative luciferase activity was measured
using a Dual Luciferase Reporter assay system (Promega Corporation)
at 48 h following transfection. Firefly luciferase activity was
normalized to Renilla luciferase activity.
Statistical analysis
Statistical analysis was performed using SPSS 22.0
software (IBM Corp.) and data are presented as the means ± SD of ≥3
experiments. A Student's t-test or one-way ANOVAs followed by a
Tukey's test were used to determine statistical differences between
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
Overexpression of SNHG7-003 inhibits the
proliferation, migration and invasion of VSMCs
A previous study reported that the expression levels
of SNHG7-003 were downregulated in patients with coronary artery
disease and unstable plaques (15). In the present study, to determine
the role of SNHG7-003 in AS, VSMCs were stimulated with ox-LDL at
various concentrations (0, 25, 50 and 100 µg/ml) for 24 h.
The results revealed that the expression levels of SNHG7-003 were
downregulated in a dose-dependent manner, with a significant
difference being observed following treatment with the
concentration of 50 or 100 µg/ml (Fig. 1A). The expression level of
SNHG7-003 was the lowest in VSMCs exposed to ox-LDL at the higher
concentration (100 µg/ml). Thus the concentration of 100
µg/ml ox-LDL was selected for use in the subsequent
experiments. Subsequently, VSMCs were stimulated with 100
µg/ml ox-LDL for 24 h for subsequent experiments. The
pcDNA-SNHG7-003 vector was constructed and the overexpression
efficiency was verified by RT-qPCR; VSMCs transfected with
pcDNA-SNHG7-003 exhibited upregulated expression levels of
SNHG7-003 compared with the control vector (Fig. 1B). Subsequently, the roles of
SNHG7-003 in VSMC proliferation and migration were investigated.
The OE of SNHG7-003 reduced the increased proliferative ability of
VSMCs induced by ox-LDL (Fig.
1C), and the observed downregulated expression levels of the
VSMC-specific marker protein, α-SMA, following SNHG7-003 OE also
validated this finding (Fig.
1D).
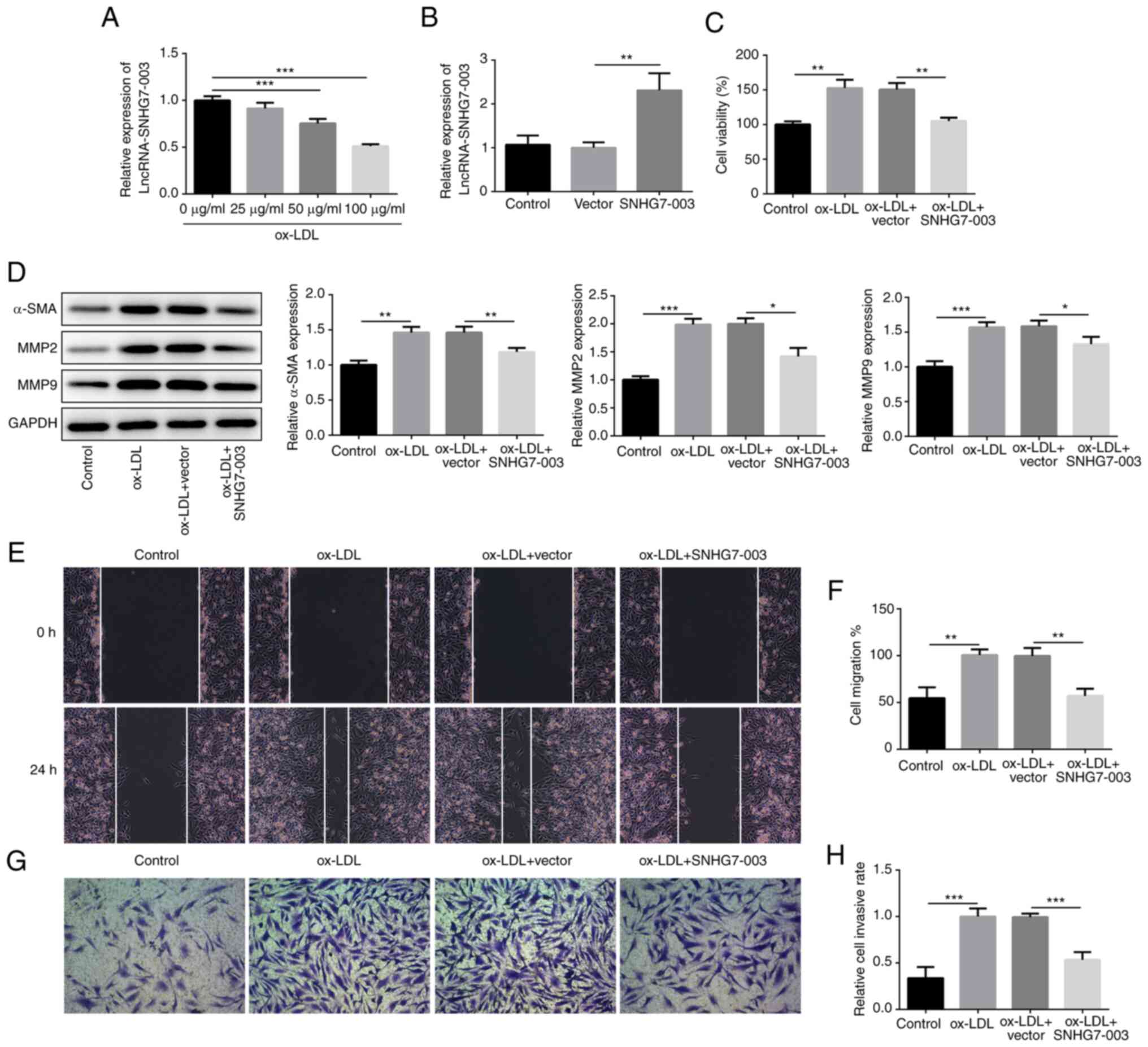 | Figure 1Overexpression of SNHG7-003 inhibits
the proliferation, migration and invasion of VSMCs. (A) The
expression level of SNHG7-003 in VSMCs stimulated with ox-LDL (0,
25, 50 and 100 µg/ml) was measured by RT-qPCR. (B) The
transfection efficiency was evaluated by RT-qPCR. (C) CCK-8 assay
was employed to assess cell viability. (D) The protein levels of
α-SMA, MMP2 and MMP9 were measured by western blot analysis. (E and
F) Wound-healing assay was conducted to assess the cell migration;
magnification, ×100. (G and H) Transwell assay was performed to
estimate the capability of cell invasion; magnification, ×100.
*P<0.05, **P<0.01,
***P<0.001. SNHG7-003, small nucleolar RNA host gene
7-003; VSMCs, vascular smooth muscle cells; ox-LDL, oxidized
low-density lipoprotein; MMP, matrix metalloproteinase; α-SMA, α
smooth muscle actin. |
MMPs are known to decompose extracellular matrix
proteins, consequently stimulating VSMC migration. As shown in
Fig. 1D, the protein expression
levels of MMP2 and MMP9 were upregulated by ox-LDL, whereas the OE
of SNHG7-003 abrogated the effects induced by ox-LDL. The results
of wound-healing and Transwell assays both demonstrated that ox-LDL
stimulation promoted cell migration and invasion, respectively,
whereas, these effects were impeded by the pcDNA-SNHG7-003 vector
(Fig. 1E-H). These findings
suggest that SNHG7-003 overexpression inhibits the proliferation,
migration and invasion of VSMCs stimulated by ox-LDL.
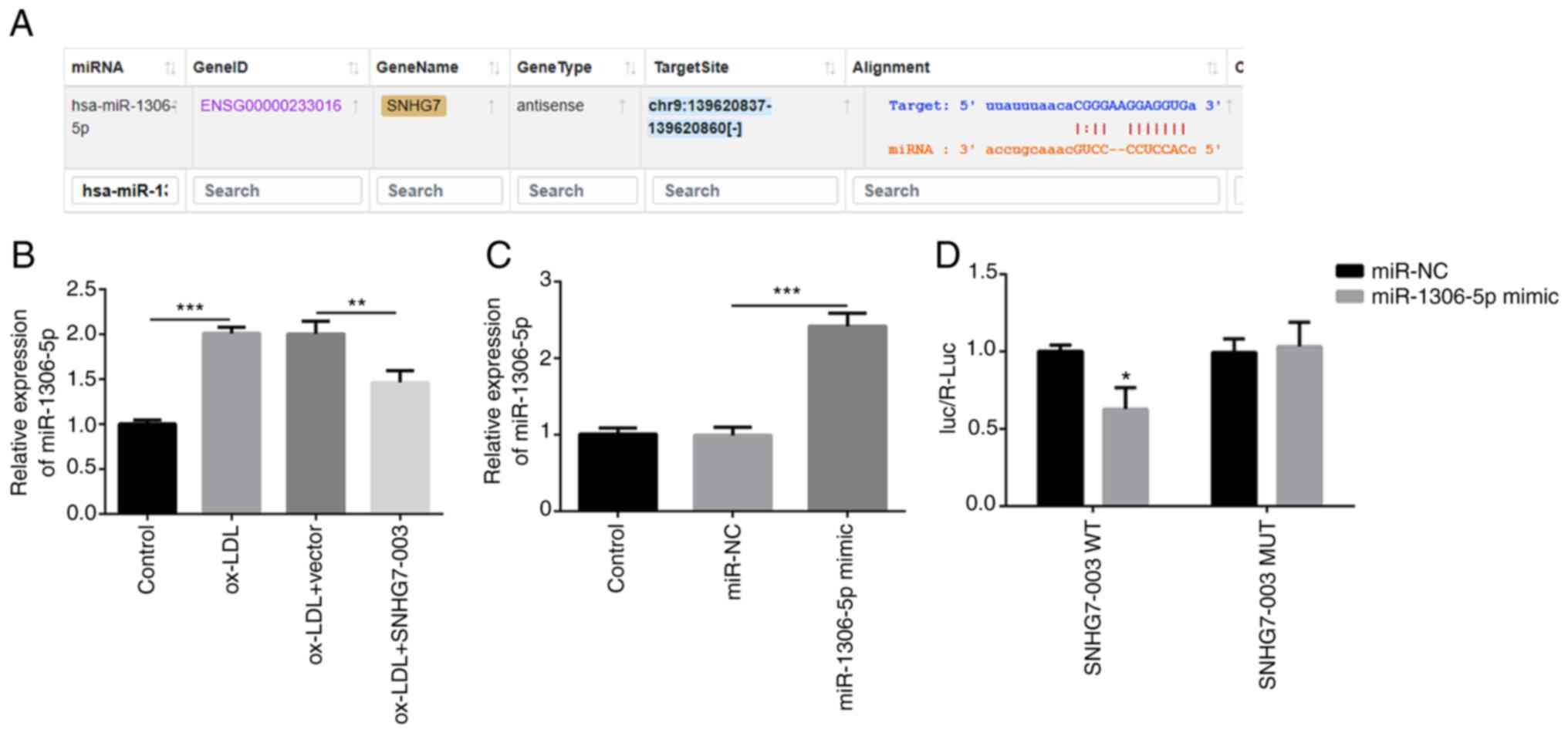
miR-1306-5p is a direct target of
SNHG7-003
First, the target genes of SNHG7-003 were predicted
using the Starbase v2.0 website (http://starbase.sysu.edu.cn). Through target gene
screening and an extensive literature research (16-19), miR-1306-5p was considered to be
the best candidate target gene of SNHG7-003 (Fig. 2A). Thus, the expression levels of
miR-1306-5p in VSMCs were investigated in the present study. The
results revealed that the miR-1306-5p expression levels were
upregulated by ox-LDL. However, in the VSMCs transfected with
pcDNA-SNHG7-003, the expression levels of miR-1306-5p were
downregulated (Fig. 2B).
Subsequently, miR-1306-5p mimic was transfected into the VSMCs to
determine the expression levels of miR-1306-5p. As shown in
Fig. 2C, miR-1306-5p mimic
markedly upregulated the expression levels of miR-1306-5p.
Subsequently, SNHG7-003-WT or SNHG7-003-Mut vectors were
synthesized and co-transfected into the VSMCs with either the
miR-1306-5p mimic or miR-NC. Co-transfection with the miR-1306-5p
mimic markedly decreased the relative luciferase activity of the
VSMCs transfected with SNHG7-003-WT, while it had no effect on the
relative luciferase activity of the cells transfected with the
SNHG7-003-Mut vector (Fig. 2D).
Thus, these results suggest that SNHG7-003 targets and modulates
the expression of miR-1306-5p.
miR-1306-5p is involved in the effects of
SNHG7-003 on VSMCs
The present study subsequently aimed to deter-mine
the role of miR-1306-5p in the SNHG7-003-regulated proliferation
and migration of VSMCs. While SNHG7-003 inhibited the proliferation
of VSMCs, this effect was counteracted by transfection with
miR-1306-5p mimic (Fig. 3A).
Furthermore, the expression levels of α-SMA, MMP2 and MMP9 were
also investigated in cells transfected with pcDNA-SNHG7-003 and/or
miR-1306-5p mimic. Transfection with miR-1306-5p mimic was found to
reverse the downregulated expression levels of α-SMA, MMP2 and MMP9
mediated by pcDNA-SNHG7-003 (Fig.
3B). Similar trends were observed following the analysis of
VSMC migration and invasion (Fig.
3C-F). Taken together, these data suggest that miR-1306-5p
inhibits the effects of SNHG7-003 on the proliferation, migration
and invasion of VSMCs.
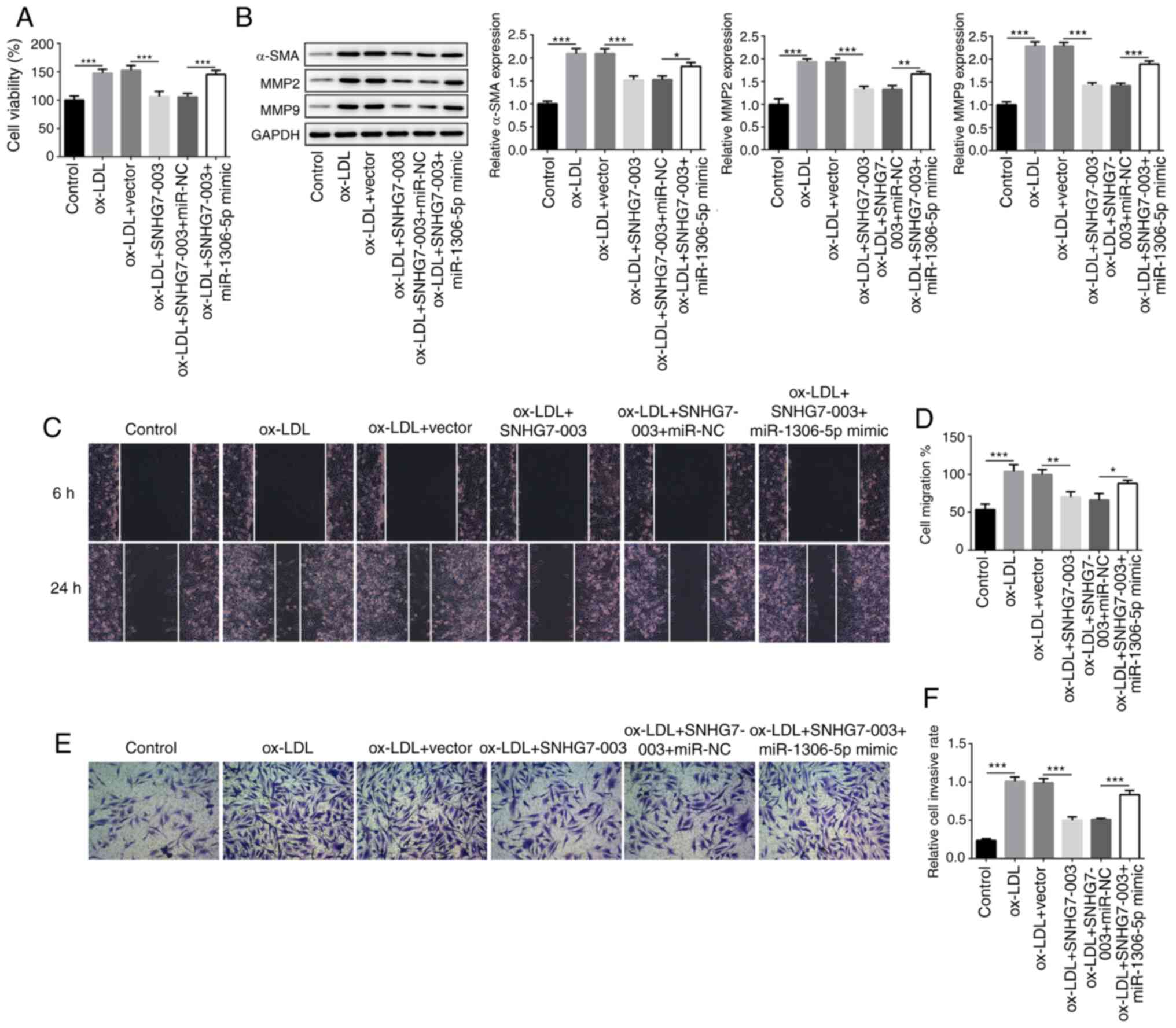 | Figure 3miR-1306-5p involves in the effects
of SNHG7-003 on VSMCs. (A) The cell viability was measured by CCK-8
assay. (B) The protein levels of α-SMA, MMP2 and MMP9 were measured
by western blot analysis. (C and D) Wound healing assay was
conducted to assess the cell migration; magnification, ×100. (E and
F) Transwell assay was performed to estimate the capability of cell
invasion; magnification, ×100. *P<0.05,
**P<0.01, ***P<0.001. SNHG7-003, small
nucleolar RNA host gene 7-003; VSMCs, vascular smooth muscle cells;
ox-LDL, oxidized low-density lipoprotein; MMP, matrix
metalloproteinase; α-SMA, α smooth muscle actin. |
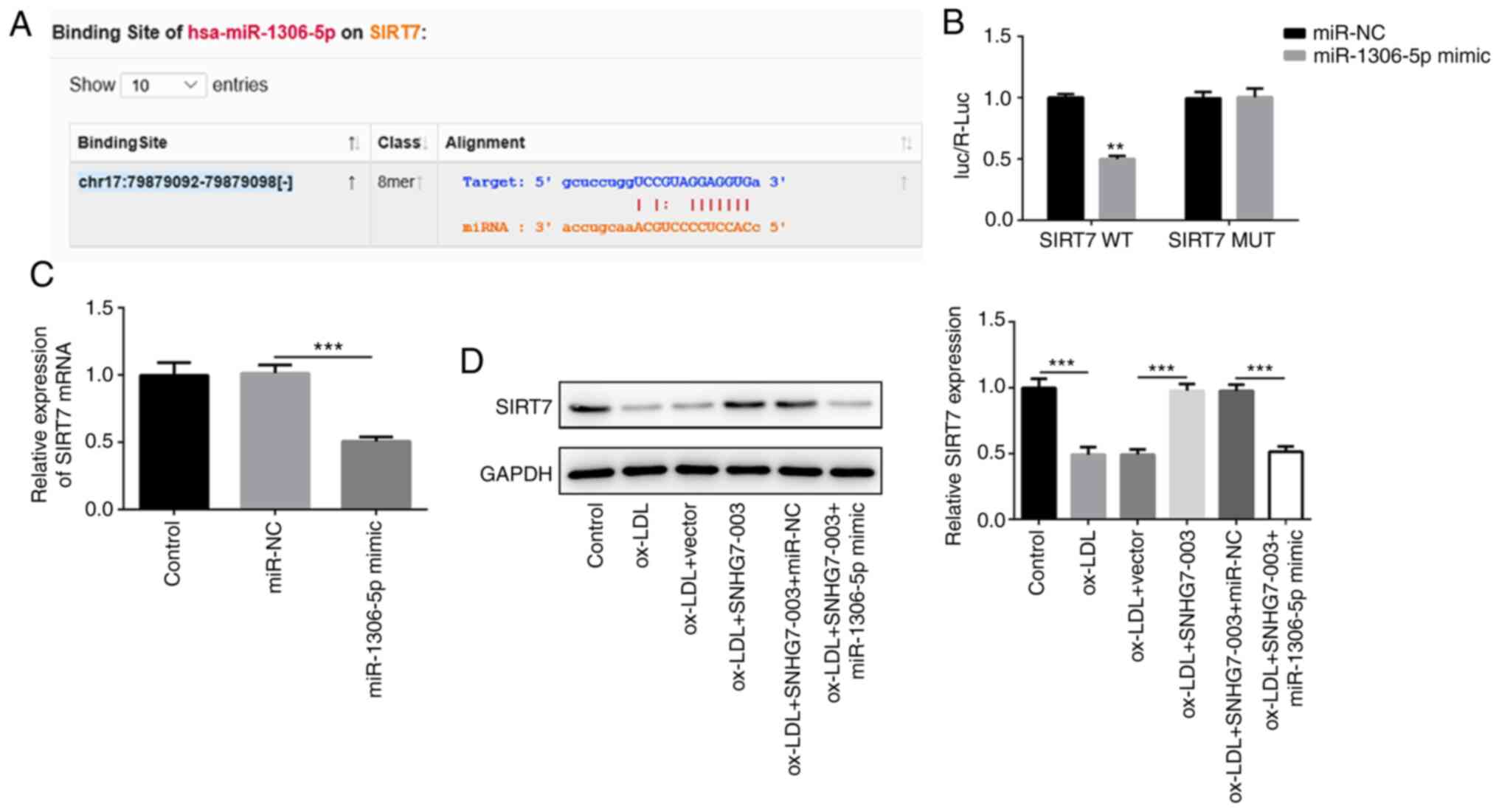
SIRT7 is the target gene of miR-1306-5p
in VSMCs
To further elucidate the underlying mechanisms
through which the SNHG7-003/miR-1306-5p axis modulates VSMC
proliferation and migration, the StarBase v2.0 website was used to
search for miR-1306-5p target genes and the binding sites of
miR-1306-5p and SIRT7 were predicted. Following a comprehensive
comparison, SIRT7 was considered to be the best candidate target
gene of miR-1306-5p (Fig. 4A).
SIRT7-WT or SIRT7-Mut were then constructed and co-transfected with
miR-1306-5p mimic or miR-NC into VSMCs. The results of the dual
luciferase reporter assay validated that miR-1306-5p could directly
bind to SIRT7 (Fig. 4B).
Consequently, the OE of miR-1306-5p down-regulated the mRNA
expression levels of SIRT7 (Fig.
4C). Notably, the downregulated expression levels of SIRT7 in
ox-LDL-stimulated VSMCs were reversed following transfection with
pcDNA-SNHG7-003, while transfection with miR-1306-5p mimic weakened
the effects of SNHG7-003 on SIRT7 (Fig. 4D). Overall, these results
suggested that SIRT7 may be a target gene of miR-1306-5p and may be
regulated by SNHG7-003.
SNHG7-003/miR-1306-5p/SIRT7 axis is
involved in VSMC proliferation, migration and invasion
To further elucidate whether the underlying
mechanisms of VSMC proliferation and migration are associated with
SIRT7, OE-SIRT7 or OE-NC vectors were constructed and transduced
into VSMCs successfully, which was demonstrated by the upregulated
expression levels of SIRT7 at the mRNA and protein level (Fig. 5A and B). However, following
transfection with the OE-SIRT7 vector, the proliferative ability of
the cells was markedly inhibited, preventing the synergism of
SNHG7-003 and miR-1306-5p on VSMCs simulated by ox-LDL (Fig. 5C). The expression levels of α-SMA,
MMP2 and MMP9 were also analyzed and the results revealed that
OE-SIRT7 attenuated the effects of the SNHG7-003/miR-1306-5p axis
on VSMC proliferation and migration (Fig. 5D). A similar trend was observed
upon analyzing the VSMC migratory and invasive abilities (Fig. 5E-H). In summary, these data
indicate that the SNHG7-003/miR-1306-5p/SIRT7 axis may be involved
in the proliferation, migration and invasion of VSMCs.
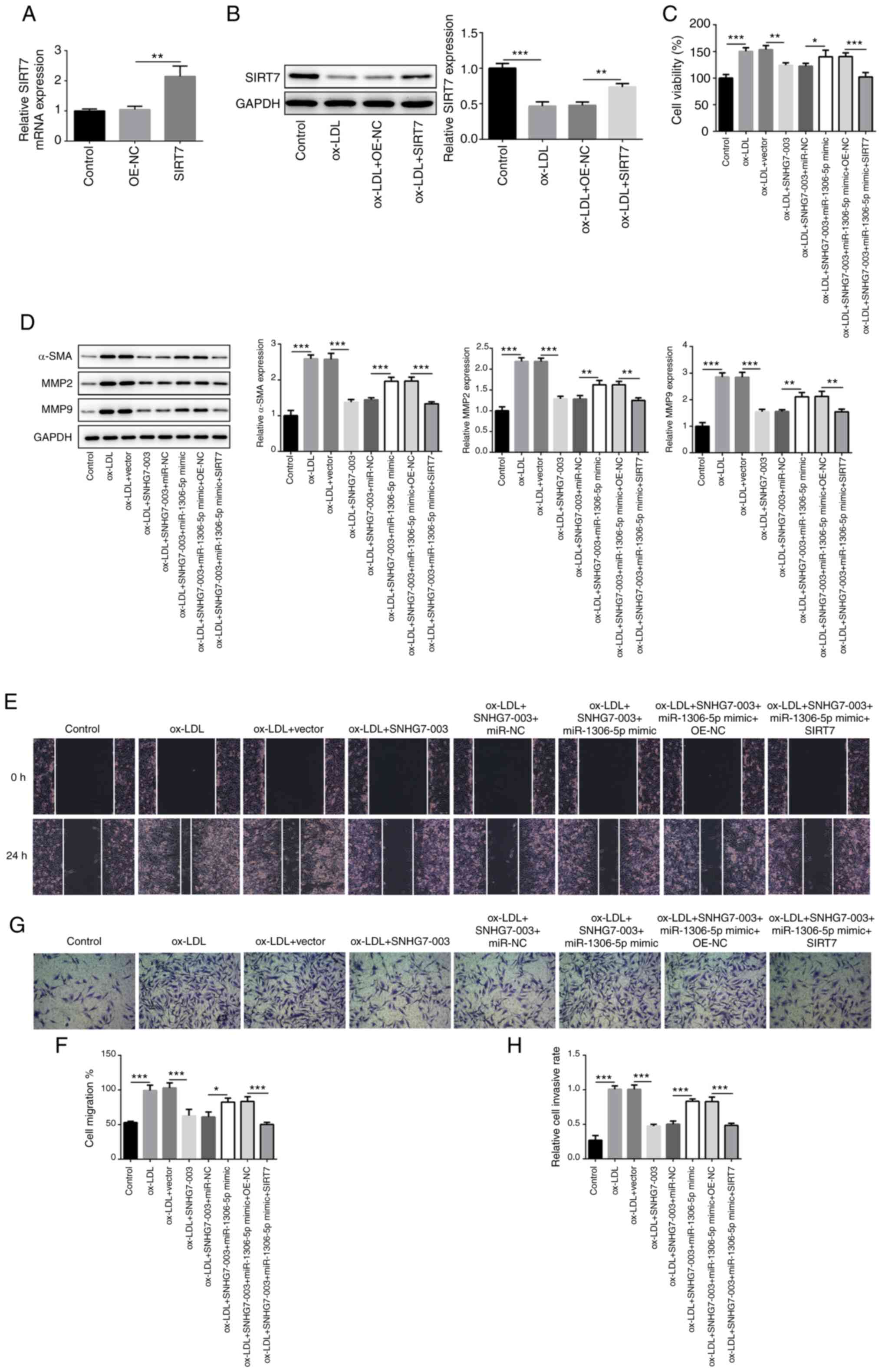 | Figure 5SNHG7-003/miR-1306-5p/SIRT7 axis
plays a role in VSMCs proliferation, migration and invasion. (A)
The mRNA expression of SIRT7 was exam-ined in VSMCs transfected
with OE- SIRT7 or OE-NC by RT-qPCR. (B) The protein level of SIRT7
in VSMCs transfected with OE- SIRT7 was estimated by western blot
analysis. (C) Cell viability was measured by CCK-8. (D) The protein
levels of α-SMA, MMP2 and MMP9 were measured by western blot
analysis. (E and F) Wound-healing assay was conducted to assess the
cell migration; magnification, ×100. (G and H) Transwell assay was
performed to estimate the capability of cell invasion;
magnification, ×100. *P<0.05, **P<0.01,
***P<0.001. SNHG7-003, small nucleolar RNA host gene
7-003; VSMCs, vascular smooth muscle cells; SIRT7, sirtuin 7;
ox-LDL, oxidized low-density lipoprotein; MMP, matrix
metalloproteinase; α-SMA, α smooth muscle actin. |
Discussion
AS is a chronic degenerative disease commonly
accompanying numerous types of cardiovascular disease, with its
pathogenesis being closely associated to the aberrant proliferation
and migration of VSMCs (21).
Accumulating evidence has reported that ncRNAs served a significant
role in AS progression, which suggested their potential to be
developed into therapeutic agents and targets for AS (22,23). For example, the lncRNA LEF1-AS1
has been discovered to regulate the migration and proliferation of
VSMCs through the miR-544a/PTEN axis (24). Another study revealed that the
lncRNA ENST00000602558.1 mediated ABCG1 expression levels and the
cholesterol efflux to HDL from VSMCs via binding to p65 (25). In addition, the lncRNA SNHG16 has
been found to promote the proliferation and inflammatory response
of macrophages through miR-17-5p/NF-κB in patients with AS
(26). These reports highlighted
the significant role that ncRNAs play in AS. However, at present,
the role of lncRNAs in AS pathogenesis is still poorly studied, and
further research is required to fully understand the function of
lncRNAs in AS.
SNHG is a class of lncRNA that plays an important
role in biological regulation of malignant tumors. The abnormal
expression of SNHG has been detected in several types of tumors
gastric cancer and breast cancer and is closely associated with the
occurrence and development of tumors (27,28). The most widely studied of these is
SNHG7. SNHG7 is located on chromosome 9 and the average length of
these transcripts was 2,176 bp, which can guide the
post-translational modification of small nucleolar RNA (snoRNA),
while snoRNA can directly participate in the progression of cancer
(29). A number of studies have
demonstrated that SNHG7 promotes the proliferation, migration and
invasion of multiple tumor cells, including malignant pleural
mesothelioma, chromophobe renal cell carcinoma and lung cancer
(30-32). However, to the best of our
knowledge, there are no studies available to date on the role of
SNHG7 in AS. Notably, a previous study used RNA-Seq to screen out
differentially expressed lncRNAs between a stable plaque group and
the unstable plaque, and the expression levels in the blood samples
from each group were determined; it was identified that SNHG7-003
expression in AS plaques was decreased significantly, and SNHG7-003
suppressed the NF-κB signaling pathway and reduced the secretion of
inflammatory factors, suggesting that blood SNHG7-003 may be a
potential biomarker for patients with coronary artery diseases
(15). In view of the
above-mentioned studies, it was hypothesized that SNHG7-003 may act
as a tumor suppressor gene in the development of AS. In the present
study, ox-LDL was used to induce VSMC proliferation and it was
discovered that the expression levels of SNHG7-003 were
downregulated following ox-LDL treatment in a dose- and
time-dependent manner, while the overexpression of SNHG7-003
counteracted the promoting effects of ox-LDL on cell proliferation
and migration.
lncRNAs can exert their biological functions by
competitive binding with miRNA to inhibit the negative regulation
of miRNA on target genes (33,34). Therefore, the authors aimed to
further study the regulatory mechanisms of SNHG7-003 in AS. The
Starbase v2.0 website predicted several miRNAs that may target
SNHG7-003 and miR-1306-5p was determined as a target following
screening. By searching the literature, it was found that
miR-1306-5p was involved in numerous types of cardio-vascular
diseases and that miR-1306 was specifically upregulated in AS
obliterans samples (16-18). These results suggested that
miR-1306-5p may play an important role in the pathogenesis of AS;
however, the role of miR-1306-5p in AS has not been studied to
date, at least to the best of our knowledge. Thus, the present
study investigated the association between miR-1306-5p and
SNHG7-003. The results of dual-luciferase reporter assay revealed
that SNHG7-003 acted as a sponge towards miR-1306-5p in VSMCs.
miR-1306-5p overexpression reversed the inhibitory effects of
SNHG7-003 overexpression on the proliferation, migration and
invasion of VSMCs induced by ox-LDL. These results revealed that
SNHG7-003 inhibited the proliferation and metastasis of VSMCs by
downregulating miR-1306-5p.
In order to construct a complete regulatory axis, we
once again predicted the target gene SIRT7 for miR-1306-5p by
Starbase v2.0 website. SIRT7 is the full-length 1.7-kb mRNA located
on chromosome 17 and belongs to classIII of histone deacetylase,
which is widely involved in a variety of cell progression,
including cell cycle, inflammation and apoptosis (35). Previous study showed that SIRT7
plays a role served as a tumor suppressor or oncogene in different
tumors or diseases (36). For
example, SIRT7 is highly expressed in gastric cancer, and the
expression level of SIRT7 is correlated positively with the
severity of the disease (37). In
papillary thyroid carcinoma, SIRT7 promotes tumorigenesis via
activating the Akt pathway by targeting DBC1 (38). In breast cancer, SIRT7 inhibits
the lung metastasis of breast cancer by suppressing
epithelial-mesenchymal transition (39). However, to date, there are limited
studies available on SIRT7 in non-neoplastic diseases, at least to
the best of our knowledge. A previous study demonstrated that the
SIRT7 expression levels were downregulated in VSMCs stimulated with
ox-LDL and that SIRT7 regulated VSMC proliferation and migration
via the Wnt/β-Catenin signaling pathway (19). However, the study of SIRT7 in AS
warrants further in-depth investigations. Thus, in the present
study, SIRT7 overexpression plasmids were synthesized, and
functional experiments were performed. The expression of SIRT7 was
downregulated in VMSCs cells stimulated with ox-LDL. SIRT7
overexpression abrogated the effects of the SNHG7-003/miR-1306-5p
axis on VSMC proliferation, migration and invasion. However, there
were several limitations to the present study. Firstly, the
specific mechanisms of SNHG7-003 regulating the miR-1306/SIRT7
pathway require further investigations. Secondly, the present study
only performed in vitro cellular experiments, and animal
experiments are required to verify the results of the current
study. In the future, the authors aim to conduct an in-depth study
on the in vivo functions and clinical relevance of
SNHG-003.
In conclusion, the findings of the present study
indicated that the expression levels of SNHG7-003 were
down-regulated in ox-LDL-stimulated VSMCs. SNHG7-003 was identified
to be able to directly target miR-1306-5p, which proceeded to
affect the expression levels of the downstream target gene, SIRT7,
exerting an inhibitory effect on cell proliferation, migration and
invasion. Thus, it was suggested that SNHG7-003 may inhibit the
proliferation and migration of VSMCs through the miR-1306-5p/SIRT7
pathway, and plays a protective role in the progression of AS. The
results enrich the biological function of SNHG7-003 in the
progression of AS, provide a new molecular mechanism through which
SNHG7-003 regulated AS, and promote the development of
lncRNA-directed diagnostics and therapeutics against AS.
Funding
The present study was supported by the Sichuan
Provincial Health Office Fund (no. 10035).
Availability of data and materials
The data used to support the findings of this study
are available from the corresponding author upon request.
Authors' contributions
JZ, QT and HC contributed to the acquisition of
data. KC, HW and ZC contributed to the analysis and interpretation
of data, YX and HY contributed to the conception and design of the
study. KL and YL performed the research. All authors read and
approved the final version of the manuscript to be published.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Ross R: The pathogenesis of
atherosclerosis: A perspective for the 1990s. Nature. 362:801–809.
1993. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shah P, Bajaj S, Virk H, Bikkina M and
Shamoon F: Rapid progression of coronary atherosclerosis: A review.
Thrombosis. 2015:6349832015. View Article : Google Scholar
|
|
3
|
Pidkovka NA, Cherepanova OA, Yoshida T,
Alexander MR, Deaton RA, Thomas JA, Leitinger N and Owens GK:
Oxidized phospholipids induce phenotypic switching of vascular
smooth muscle cells in vivo and in vitro. Circ Res. 101:792–801.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Owens GK, Kumar MS and Wamhoff BR:
Molecular regulation of vascular smooth muscle cell differentiation
in development and disease. Physiol Rev. 84:767–801. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ang AH, Tachas G, Campbell JH, Bateman JF
and Campbell GR: Collagen synthesis by cultured rabbit aortic
smooth-muscle cells. Alteration with phenotype Biochem J.
265:461–469. 1990.
|
|
6
|
Alevizos I and Illei GG: MicroRNAs as
biomarkers in rheumatic diseases. Nat Rev Rheumatol. 6:391–398.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cheng Q, Zhang M, Zhang M, Ning L and Chen
D: Long non-coding RNA LOC285194 regulates vascular smooth muscle
cell apoptosis in atherosclerosis. Bioengineered. 11:53–60. 2020.
View Article : Google Scholar :
|
|
8
|
Hung J, Scanlon JP, Mahmoud AD, Rodor J,
Ballantyne M, Fontaine MAC, Temmerman L, Kaczynski J, Connor KL,
Bhushan R, et al: Novel plaque enriched long noncoding RNA in
atherosclerotic macrophage regulation (PELATON). Arterioscler
Thromb Vasc Biol. 40:697–713. 2020. View Article : Google Scholar :
|
|
9
|
Wang L, Zheng Z, Feng X, Zang X, Ding W,
Wu F and Zhao Q: circRNA/lncRNA-miRNA-mRNA network in oxidized,
low-density, lipoprotein-induced foam cells. DNA Cell Biol.
38:1499–1511. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bai HL, Lu ZF, Zhao JJ, Ma X, Li XH, Xu H,
Wu SG, Kang CM, Lu JB, Xu YJ, et al: Microarray profiling analysis
and validation of novel long noncoding RNAs and mRNAs as potential
biomarkers and their functions in atherosclerosis. Physiol
Genomics. 51:644–656. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xing YH, Bai Z, Liu CX, Hu SB, Ruan M and
Chen LL: Research progress of long noncoding RNA in China. IUBMB
Life. 68:887–893. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ahmed ASI, Dong K, Liu J, Wen T, Yu L, Xu
F, Kang X, Osman I, Hu G, Bunting KM, et al: Long noncoding RNA
NEAT1 (nuclear paraspeckle assembly transcript 1) is critical for
phenotypic switching of vascular smooth muscle cells. Proc Natl
Acad Sci USA. 115:E8660–E8667. 2018. View Article : Google Scholar
|
|
13
|
Leung A, Stapleton K and Natarajan R:
Functional long non-coding RNAs in vascular smooth muscle cells.
Curr Top Microbiol Immunol. 394:127–141. 2016.
|
|
14
|
Cui C, Wang X, Shang XM, Li L, Ma Y, Zhao
GY, Song YX, Geng XB, Zhao BQ, Tian MR and Wang HL: lncRNA 430945
promotes the proliferation and migration of vascular smooth muscle
cells via the ROR2/RhoA signaling pathway in atherosclerosis. Mol
Med Rep. 19:4663–4672. 2019.PubMed/NCBI
|
|
15
|
Pan Z, Fan Z, Ma J, Liu H, Shen L, He B
and Zhang M: Profiling and functional characterization of
circulation LncRNAs that are associated with coronary
atherosclerotic plaque stability. Am J Transl Res. 11:3801–3815.
2019.PubMed/NCBI
|
|
16
|
Chatterjee S, Gupta SK, Bär C and Thum T:
Noncoding RNAs: Potential regulators in cardioncology. Am J Physiol
Heart Circ Physiol. 316:H160–H168. 2019. View Article : Google Scholar :
|
|
17
|
Soler-Botija C, Gálvez-Montón C and
Bayés-Genís A: Epigenetic biomarkers in cardiovascular diseases.
Front Genet. 10:9502019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang M, Li W, Chang GQ, Ye CS, Ou JS, Li
XX, Liu Y, Cheang TY, Huang XL and Wang SM: MicroRNA-21 regulates
vascular smooth muscle cell function via targeting tropomyosin 1 in
arteriosclerosis obliterans of lower extremities. Arterioscler
Thromb Vasc Biol. 31:2044–2053. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zheng J, Chen K, Wang H, Chen Z, Xi Y, Yin
H, Lai K and Liu Y: SIRT7 regulates the vascular smooth muscle
cells proliferation and migration via Wnt/β-catenin signaling
pathway. Biomed Res Int. 2018:47695962018. View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
21
|
Dzau VJ, Braun-Dullaeus RC and Sedding DG:
Vascular proliferation and atherosclerosis: New perspectives and
therapeutic strategies. Nat Med. 8:1249–1256. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Holdt LM, Kohlmaier A and Teupser D: Long
noncoding RNAs of the arterial wall as therapeutic agents and
targets in atherosclerosis. Thromb Haemost. 119:1222–1236. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Skuratovskaia D, Vulf M, Komar A,
Kirienkova E and Litvinova L: Promising directions in
atherosclerosis treatment based on epigenetic regulation using
microRNAs and long noncoding RNAs. Biomolecules. 9:2262019.
View Article : Google Scholar :
|
|
24
|
Zhang L, Zhou C, Qin Q, Liu Z and Li P:
LncRNA LEF1-AS1 regulates the migration and proliferation of
vascular smooth muscle cells by targeting miR-544a/PTEN axis. J
Cell Biochem. 120:14670–14678. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cai C, Zhu H, Ning X, Li L, Yang B, Chen
S, Wang L, Lu X and Gu D: LncRNA ENST00000602558.1 regulates ABCG1
expression and cholesterol efflux from vascular smooth muscle cells
through a p65-dependent pathway. Atherosclerosis. 285:31–39. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
An JH, Chen ZY, Ma QL, Wang HJ, Zhang JQ
and Shi FW: LncRNA SNHG16 promoted proliferation and inflammatory
response of macrophages through miR-17-5p/NF-κB signaling pathway
in patients with atherosclerosis. Eur Rev Med Pharmacol Sci.
23:8665–8677. 2019.PubMed/NCBI
|
|
27
|
Zhao L, Guo H, Zhou B, Feng J, Li Y, Han
T, Liu L, Li L, Zhang S, Liu Y, et al: Long non-coding RNA SNHG5
suppresses gastric cancer progression by trapping MTA2 in the
cytosol. Oncogene. 5:5770–5780. 2016. View Article : Google Scholar
|
|
28
|
Lee J, Jung JH, Chae YS, Park HY, Kim WW,
Lee SJ, Jeong JH and Kang SH: Long noncoding RNA snaR regulates
proliferation, migration and invasion of triple-negative breast
cancer cells. Anticancer Res. 36:6289–6295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Romano G, Veneziano D, Acunzo M and Croce
CM: Small non-coding RNA and cancer. Carcinogenesis. 38:485–491.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
He HT, Xu M, Kuang Y, Han XY, Wang MQ and
Yang Q: Biomarker and competing endogenous RNA potential of
tumor-specific long noncoding RNA in chromophobe renal cell
carcinoma. Onco Targets Ther. 9:6399–6406. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Quinn L, Finn SP, Cuffe S and Gray SG:
Non-coding RNA repertoires in malignant pleural mesothelioma. Lung
Cancer. 90:417–426. 2015. View Article : Google Scholar
|
|
32
|
She K, Huang J, Zhou H, Huang T, Chen G
and He J: lncRNA-SNHG7 promotes the proliferation, migration and
invasion and inhibits apoptosis of lung cancer cells by enhancing
the FAIM2 expression. Oncol Rep. 36:2673–2680. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhu H, Lu J, Zhao H, Chen Z, Cui Q, Lin Z,
Wang X, Wang J, Dong H, Wang S and Tan J: Functional long noncoding
RNAs (lncRNAs) in clear cell kidney carcinoma revealed by
reconstruction and comprehensive analysis of the lncRNA-miRNA-mRNA
regulatory network. Med Sci Monit. 24:8250–8263. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pan R, He Z, Ruan W, Li S, Chen H, Chen Z,
Liu F, Tian X and Nie Y: lncRNA FBXL19-AS1 regulates osteosarcoma
cell proliferation, migration and invasion by sponging miR-346.
Onco Targets Ther. 11:8409–8420. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Houtkooper RH, Pirinen E and Auwerx J:
Sirtuins as regulators of metabolism and healthspan. Nat Rev Mol
Cell Biol. 13:225–238. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ryu D, Jo YS, Lo Sasso G, Stein S, Zhang
H, Perino A, Lee JU, Zeviani M, Romand R, Hottiger MO, et al: A
SIRT7-dependent acetylation switch of GABPβ1 controls mitochondrial
function. Cell Metab. 20:856–869. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang S, Chen P, Huang Z, Hu X, Chen M, Hu
S, Hu Y and Cai T: Sirt7 promotes gastric cancer growth and
inhibits apoptosis by epigenetically inhibiting miR-34a. Sci Rep.
5:97872015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li H, Tian Z, Qu Y, Yang Q, Guan H, Shi B,
Ji M and Hou P: SIRT7 promotes thyroid tumorigenesis through
phosphorylation and activation of Akt and p70S6K1 via DBC1/SIRT1
axis. Oncogene. 38:345–359. 2019. View Article : Google Scholar
|
|
39
|
Tang X, Shi L, Xie N, Liu Z, Qian M, Meng
F, Xu Q, Zhou M, Cao X, Zhu WG and Liu B: Sirt7 antagonizes TGF-β
signaling and inhibits breast cancer metastasis. Nat Commun.
8:3182017. View Article : Google Scholar
|