Introduction
Osteosarcoma (OS) is the most common primary
malignant bone tumor that affects children, adolescents and young
adults (1,2). With the application of
chemotherapeutics, a combination of surgical resection and
multi-chemotherapy has become a standard clinical treatment
strategy for almost all patients with OS, which has significantly
improved patient survival (3).
Despite extensive advances achieved in OS therapy, the overall
survival rate of patients with distant metastasis remains poor
(4,5). To date, the biological
characteristics of OS are well understood, but it is urgent to
explore the mechanisms of OS progression, which may contribute to
the development of effective strategies for the diagnosis,
treatment and prognosis of patients with OS.
Long non-coding RNAs (lncRNAs) belong to the
non-coding RNA family and are generally comprised of ~200
nucleotides (6,7). Abnormal expression levels of lncRNAs
contribute to tumor initiation, growth and metastasis (8-11).
Recently, several lncRNAs have been reported to be involved in OS
progression, such as ODRUL (12),
LINC01278 (13) and lncRNA DLEU1
(14). However, the functions of
lncRNAs in OS require further investigation. Tumor suppressor
candidate 7 (TUSC7) is a lncRNA that has been reported to be a
cancer suppressor gene in numerous types of human cancer, such as
colorectal cancer (15),
pancreatic carcinoma (16) and
esophageal squamous cell carcinoma (17). Recently, TUSC7 has been also
identified as a tumor suppressor in OS (18). However, the regulatory mechanisms
of TUSC7 in OS require additional investigation.
In addition to lncRNAs, microRNAs (miRNAs/miRs) are
another group of non-coding but short-length (<25 nucleotides)
RNAs. Previous studies have demonstrated that miR-181a serves
important roles in the development and progression of OS (19-21). In recent years, the discovery of
crosstalk between lncRNAs and miRNAs has revealed a new mechanism
of protein-coding gene modulation (22,23). Specifically, it has been
demonstrated that lncRNAs work with miRNAs by acting as competitive
endogenous RNAs (ceRNAs) or as miRNA sponges (24,25). Accordingly, the present study
hypothesized a regulatory mechanism for lncRNA-miRNA-mRNA and
investigated its functional roles in OS.
The present study investigated TUSC7 expression in
OS and the vital role of TUSC7 in the proliferation, migration and
invasion of OS both in vitro and in vivo.
Materials and methods
Clinical OS specimens
A total of 45 pairs of OS tissue samples and
adjacent normal tissue samples (<2 cm from tumor) were collected
between November 2018 and October 2019 from patients diagnosed with
OS at the Affiliated Hospital of Inner Mongolia Medical University
(Hohhot, China). There were 34 males and 11 females, with a median
age of 19.4 years (age range, 10-25 years). The pathological
diagnoses of OS were confirmed by two independent pathologists.
Patients receiving chemotherapy or radiotherapy treatment were
excluded. Written informed consent was provided by all patients
with OS enrolled in the study. The present study was approved by
the Ethics Committee of the Affiliated Hospital of Inner Mongolia
Medical University (approval no. Y K D2017142). All tissue samples
were immediately frozen in liquid nitrogen and then stored at −80°C
until further use.
Cell culture
Human OS cell lines (Saos2, U2OS, MG63 and 143B) and
a human osteoblast cell line (hFOB 1.19) were purchased from the
American Type Culture Collection. All cell lines were cultured in
DMEM supplemented with 10% FBS and 1% antibiotics (penicillin, 100
IU/ml and streptomycin, 10 mg/ml; all Gibco; Thermo Fisher
Scientific, Inc.) at 37°C and 5% CO2.
Cell transfection
For transient transfection, cells were seeded into
6- or 96-well plates (5×106 or 104
cells/well, respectively). U2OS and MG63 cells were transfected
with 2 µg pcDNA3.1-TUSC7 (TUSC7 overexpression vector), 2
µg pcDNA3.1-NC (empty vector), 100 nM non-targeting negative
control (NC)-mimic or 100 nM miR-181a mimic. The synthesized
sequences were 5′-AAC AUU CAA CGC UGU CGG UGA GU-3′ for the
miR-181a mimic and 5′-UUC UCC GAA CG UGU CAC GUT T-3′ for the
NC-mimc. In the present study, pcDNA3.1-TUSC7, pcDNA3.1-NC, 2
µg small interfering (si)-RASSF6 inhibitor (si-RASSF6),
si-NC, miR-181a mimic and NC-mimic were provided by Shanghai
GenePharma Co., Ltd. Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) was used for transfection according
to the manufacturer's protocol. The RASSF6 siRNA target sequence
was 5′-GAC CCA GAU UCC UAU GUC U-3′, while the si-NC target
sequence was 5′-UUC UCC GAA CGU GUC ACG UTT-3′. After transfection
for 2 days at the room temperature, the cells were collected for
subsequent use.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA from tissues and cells was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) following the manufacturer's protocol. RNA (1 µg) was
reverse transcribed to cDNA using the Prime Script RT Master Mix
kit (Takara Bio, Inc.) according to the manufacturer's protocol.
After RT, qPCR analysis was conducted using SYBR Premix Ex Taq™ II
(Takara Bio, Inc.) on a StepOnePlus™ Real-Time PCR System (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The PCR protocol was
set at 95°C for 10 min, followed by 40 cycles at 95°C for 10 sec
and 60°C for 1 min. The 2−ΔΔCq method (26) was used to calculate relative mRNA
expression. The expression levels of TUSC7 and RASSF6 were
normal-ized to those of GAPDH, while miR-181a expression was
normalized to that of U6. The following gene-specific primers were
used: TUSC7 forward, 5′-CAC TGC CTA TGT GCA CGA CT-3′ and reverse,
5′-AGA GTC CGG CAA GAA GAA CA-3′; miR-181a forward, 5′-ACA CTC CAG
CTG GGA ACA TTC AAC GCT GTC G-3′ and reverse, 5′-GGT GTC GTG GAG
TCG GCA ATT CAG TTG AG-3′; RASSF6 forward, 5′-AGG CCA GAC AGC TCT
GAT GT-3′ and reverse, 5′-AGG CCA GAC AGC TCT GAT GT-3′; U6
forward, 5′-CTC GCT TCG GCA GCA CA-3′ and reverse, 5′-AAC GCT TCAC
GAA TTT GCG T-3′; GAPDH forward, 5′-TGA CTT CAA CAG CGA CAC CCA-3′
and reverse, 5′-CAC CCT GTT GCT GTA GCC AAA-3′; and β-actin
forward, 5′-CTC CAT CCT GGC CTC GCT GT-3′ and reverse, 5′-GCT GTC
ACC TTC ACC GTT CC-3′.
Western blot analysis
Total proteins in tissues and cells were lysed using
RIPA lysate buffer with protease inhibitor cocktail (Roche Applied
Science). The protein concentration was quantified using the BCA
assay. SDS-PAGE (10%) was conducted to isolate 20 µg/lane of
protein and transfer it to PVDF membranes (EMD Millipore). After
blocking with 5% skimmed milk solution at room temperature for 1 h,
membranes were incubated overnight with primary antibodies at 4°C
and then incubated with HRP-conjugated secondary antibody (cat. no.
A8419; 1:5,000; Sigma-Aldrich; Merck KGaA) at room temperature for
1 h. All primary anti-bodies were purchased from Abcam, including
anti-RASSF6 (cat. no. ab220111; 1:1,000), anti-Bax (cat. no.
ab32503; 1:1,000), anti-Bcl2 (cat. no. ab182858; 1:2,000),
anti-Caspase-3 (cat. no. ab13847; 1:500), anti-Caspase-8 (cat. no.
ab32397; 1:500) and anti-GAPDH (cat. no. ab9485, 1:2,500). Target
protein levels were normalized to GAPDH, which served as the
control. Protein expression was measured using enhanced
chemiluminescence (EMD Millipore), and the software used for
densitometry was ImageJ v1.8.0 (National Institutes of Health).
Cell Counting Kit-8 (CCK-8) assay
CCK-8 (Beyotime Institute of Biotechnology) assays
were conducted according to the manufacturer's protocol to assess
cell proliferation. U2OS and MG63 cells (104 cells/well)
were seeded into 96-well plates and transfected with TUSC7
overexpression vector or control vector, or miR-181a mimic or
NC-mimic, as aforementioned. Cells were incubated with 10% CCK-8
solution added to each well in the dark for 2 h at 37°C.
Proliferation rates were determined at 24, 48 and 72 h after
transfection. The optical density (OD) of cells was measured using
an ultraviolet spectrophotometer (ELX800; BioTek Instruments, Inc.;
Agilent Technologies, Inc.) at a wavelength of 450 nm.
Colony formation assay
After transfection, cells were incubated in 6-well
plates (500 cells/well) for colony formation assays. After 2 weeks
of culture at 37°C, colonies were fixed with 10% formaldehyde at
room temperature for 15 min and stained with 0.1% crystal violet
solution at room temperature for 10 min. A colony was defined as
containing >50 cells. The colony number in each well was counted
and imaged under a light microscope (magnification, ×20; Olympus
Corporation).
Transwell assay
The migratory and invasive abilities of U2OS and
MG63 cells were estimated using Transwell assays. For the Transwell
migration assay, transfected U2OS and MG63 cells (1×105)
were seeded into the upper chamber of 8-µm pore size
membranes (Merck KGaA) containing serum-free medium. For the
Transwell invasion assay, U2OS and MG63 cells (2×105)
were added in serum-free medium to the upper chambers pre-coated
with diluted Matrigel® (1:5; BD Biosciences) at 37°C for
5 h. A total of 500 µl medium containing 10% FBS was added
into the lower chamber. After incubation at 37°C for 48 h, cells on
the upper membrane were removed using a cotton swab, and cells that
had traversed the membrane were stained using 0.1% crystal violet
at room temperature for 10 min and counted under a light microscope
(magnification, ×100; Olympus Corporation) in five randomly chosen
microscopic fields. The number of cells that entered the lower
chamber reflected the migratory or invasive ability of tumor
cells.
Apoptosis assay
Flow cytometry analysis was used to evaluate
apoptosis. Briefly, U2OS and MG63 cells (2×105
cells/well) were seeded into 6-well plates, and after transfection
for 48 h, cells were treated with Annexin V-FITC (5 µl) and
PI (5 µl) using an Annexin V-FITC Apoptosis Detection kit
(Invitrogen; Thermo Fisher Scientific, Inc.). The apoptosis rate
was detected using a CytoFLEX flow cytometry (cat. no. C02945;
Beckman Coulter, Inc.) following the manufacturer's protocol and
analyzed using ModFit LT v5.0 (Verity Software House, Inc.).
Dual luciferase reporter assay
Firstly, the starBase data-base (http://starbase.sysu.edu.cn/) was used to predict the
binding sites between miR-181a and TUSC7 or RASSF6, and luciferase
reporter assay was used to confirm this association. For the
luciferase reporter assay, sequences of TUSC7 and RASSF6 containing
wild-type (WT) binding sites to miR-181a or mutant (Mut) sites were
amplified and cloned into the luciferase reporter vector pGL3
(Promega Corporation), called TUSC7-WT and RASSF6-WT, and TUSC7-Mut
and RASSF6-Mut, respectively. Cells were transfected with
TUSC7-WT/Mut or RASSF6-WT/Mut, as well as with miR-181a mimic or
NC-mimic using Lipofectamine 2000 as aforementioned. After ~48 h,
luciferase activity was measured using the Dual-Luciferase Reporter
Assay System (Promega Corporation) following the manufacturer's
protocol. Renilla luciferase activity was used as an
internal reference.
Nuclear/cytoplasmic RNA
fractionation
The cytoplasmic/nuclear fraction isolation assay was
performed using a PARIS kit (Thermo Fisher Scientific, Inc.)
following the manufacturer's protocol. After purification and DNase
I treatment, RNA from the isolated nuclear and cytoplasmic
fractions was reverse transcribed, and qPCR was used to evaluate
the relative expression levels of TUSC7 and GAPDH in each sample,
as aforementioned.
Hematoxylin and eosin (H&E)
staining
Samples were collected from mouse subcutaneous
tumors. First, samples were fixed in 4% paraformaldehyde solution
at room temperature for 48 h, embedded in paraffin and transversely
cut into 5-µm-thick sections. Second, paraffin sections were
deparaffinized using xylene I and II for 10 min and rehydrated
using a descending alcohol series (100, 90, 80 and 70% alcohol for
5 min each, followed by washing with water for 5 min). Finally,
slices were subjected to H&E staining at room temperature for
15 min and observed under a light microscope (magnification, ×100;
Olympus Corporation).
Immunohistochemistry
Mouse subcutaneous tumors were removed and fixed in
4% formaldehyde at room temperature for 48 h, embedded in paraffin
and transversely cut into 5-µm-thick sections. Subsequently,
sections were washed three times with 0.1 M PBS after
deparaffinization using xylene I and II for 10 min and rehydration
as aforementioned, and blocked with blocking buffer (Dual
Endogenous Enzyme Block; Dako; Agilent Technologies, Inc.) at room
temperature for 5 min. Sections were incubated with goat anti-Ki67
primary antibody (cat. no. ab15580; 1:100; Abcam) for 24 h at 4°C,
followed by incubation with an HRP-conjugated rabbit anti-goat IgG
secondary antibody (cat. no. ab6741; 1:500; Abcam) for 30 min at
room temperature. Finally, sections were stained with
3,3-diaminobenzidine (Wuhan Servicebio Technology Co., Ltd.) and
visualized using a light microscope (magnification, ×100; Olympus
Corporation).
In vivo mouse xenograft tumor assay
A total of 30 male BALB/c nude mice (weight, 18-20
g; age, 4-6 weeks) from Beijing Vital River Laboratory Animal
Technology Co., Ltd., were kept in a 12-h day/night cycle and in a
temperature-controlled room (temperature, 18-22°C; humidity,
45-65%) with free standard food and tap water. There were 6 mice in
each group. Each experiment was repeated 3 times. Animal
experiments were approved by the Animal Experimentation Ethics
Committee of the Affiliated Hospital of Inner Mongolia Medical
University and conducted following the Guide for the Care and Use
of Laboratory Animals (27). U2OS
cells stably expressing TUSC7 or control vector were propagated,
and 1×106 cells/100 µl culture medium were
subcutaneously inoculated into the right side of the posterior
flank in nude mice. Animals were sacrificed 5 weeks
post-inoculation by intraperitoneal injection of pentobarbital
sodium (100 mg/kg). Tumors were surgically dissected and
weighed.
Statistical analysis
All data were collected from at least three
independent experiments and are reported as the mean ± SD.
Statistical significance between normal and tumor tissues were
analyzed using a paired Student's t-test, while other comparisons
between two groups were analyzed using an unpaired Student's
t-test. One-way ANOVA followed by Tukey's post-hoc test was used
for multiple comparisons using SPSS version 21.0 software (IBM
Corp.). P<0.05 was considered to indicate a statistically
significant difference.
Results
TUSC7 expression is significantly
downregulated in OS tissues and cell lines
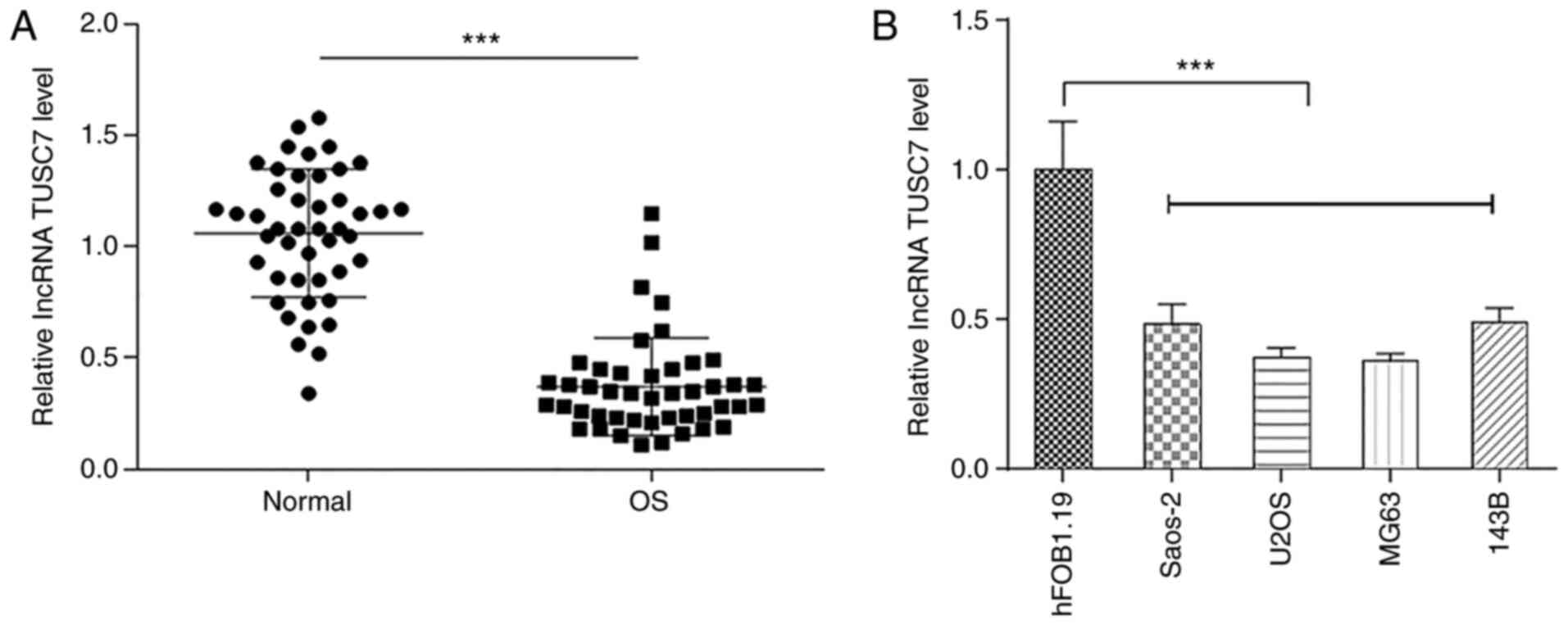
To investigate the role of TUSC7 in OS progression,
the expression levels of TUSC7 in OS and normal bone tissues were
measured by RT-qPCR. Results demonstrated that TUSC7 expression was
significantly lower in OS tissues compared with in normal bone
tissues (Fig. 1A). In addition,
TUSC7 expression was significantly decreased in OS cell lines
compared with in the human osteoblast hFOB1.19 cell line (Fig. 1B), especially in U2OS and MG63
cells, which were therefore used for subsequent experiments.
TUSC7 overexpression inhibits the
progression of OS
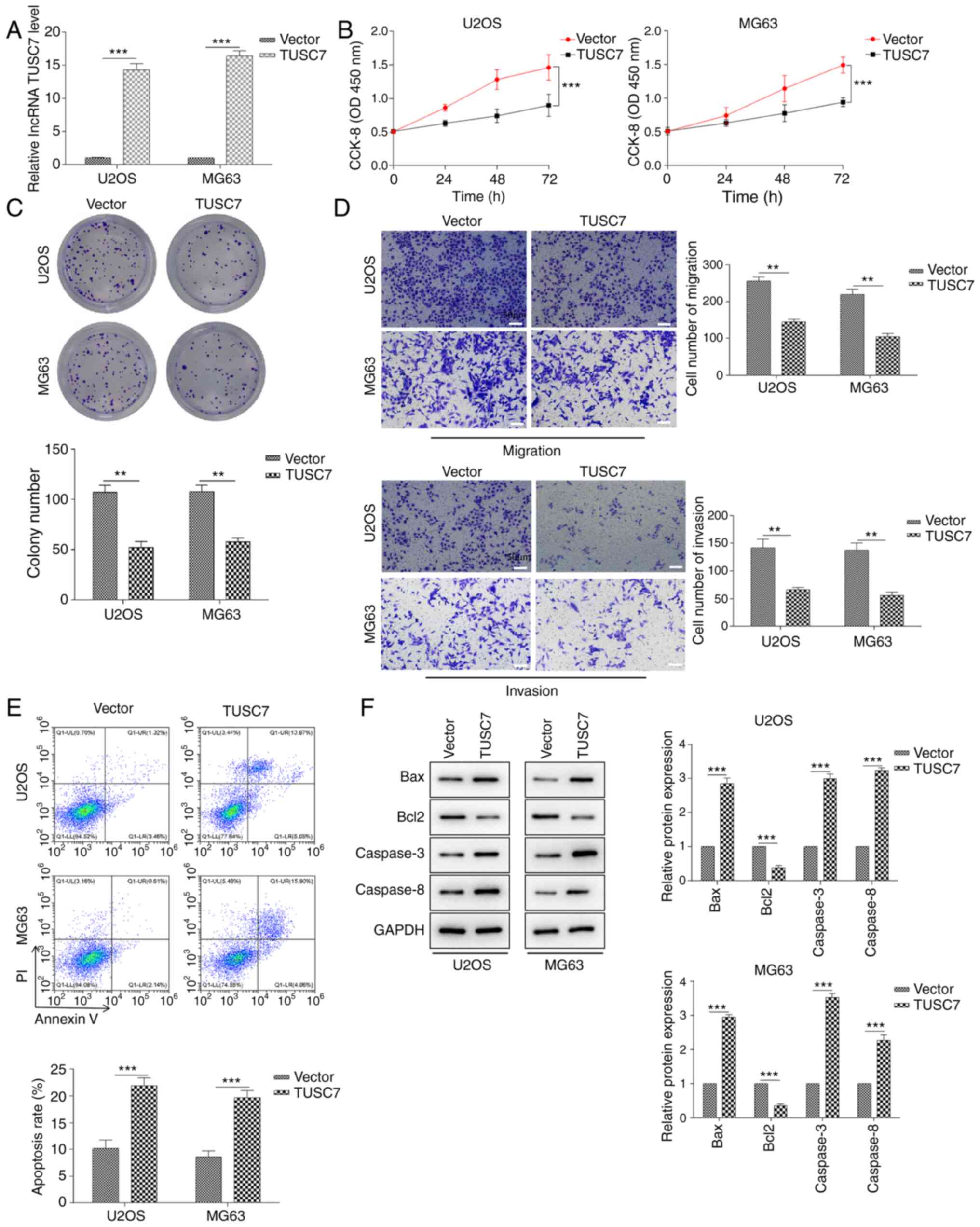
To explore the functional roles of TUSC7 in OS cell
proliferation, migration, invasion and apoptosis, U2OS and MG63
cells were transfected with TUSC7-overexpressing plasmid or control
vector. RT-qPCR assays revealed that the expression levels of TUSC7
in U2OS and MG63 cells were significantly increased in response to
TUSC7 transfection (Fig. 2A).
Cell proliferation was measured using CCK-8 and colony formation
assays. The results demonstrated that cell proliferation rate and
colony formation ability in the TUSC7 group were significantly
lower compared with those in the control vector group (Fig. 2B and C). Additionally, the results
of the Transwell assay revealed that migration and invasion of U2OS
and MG63 cells were significantly decreased after TUSC7
overexpression (Fig. 2D). By
contrast, the rate of early and late apoptotic cells in the TUSC7
transfection group was significantly increased (Fig. 2E). In addition, the protein
expression levels of the anti-apoptotic protein Bcl2 were
significantly decreased, while those of the pro-apoptotic proteins
Bax, Caspase-3 and Caspase-8 were significantly increased in the
TUSC7 group (Fig. 2F). The
current results indicated that TUSC7 overexpression significantly
inhibited proliferation, migration and invasion, and promoted
apoptosis in U2OS and MG63 cells.
TUSC7 competitively binds to miR-181a and
suppresses its expression in OS cell lines
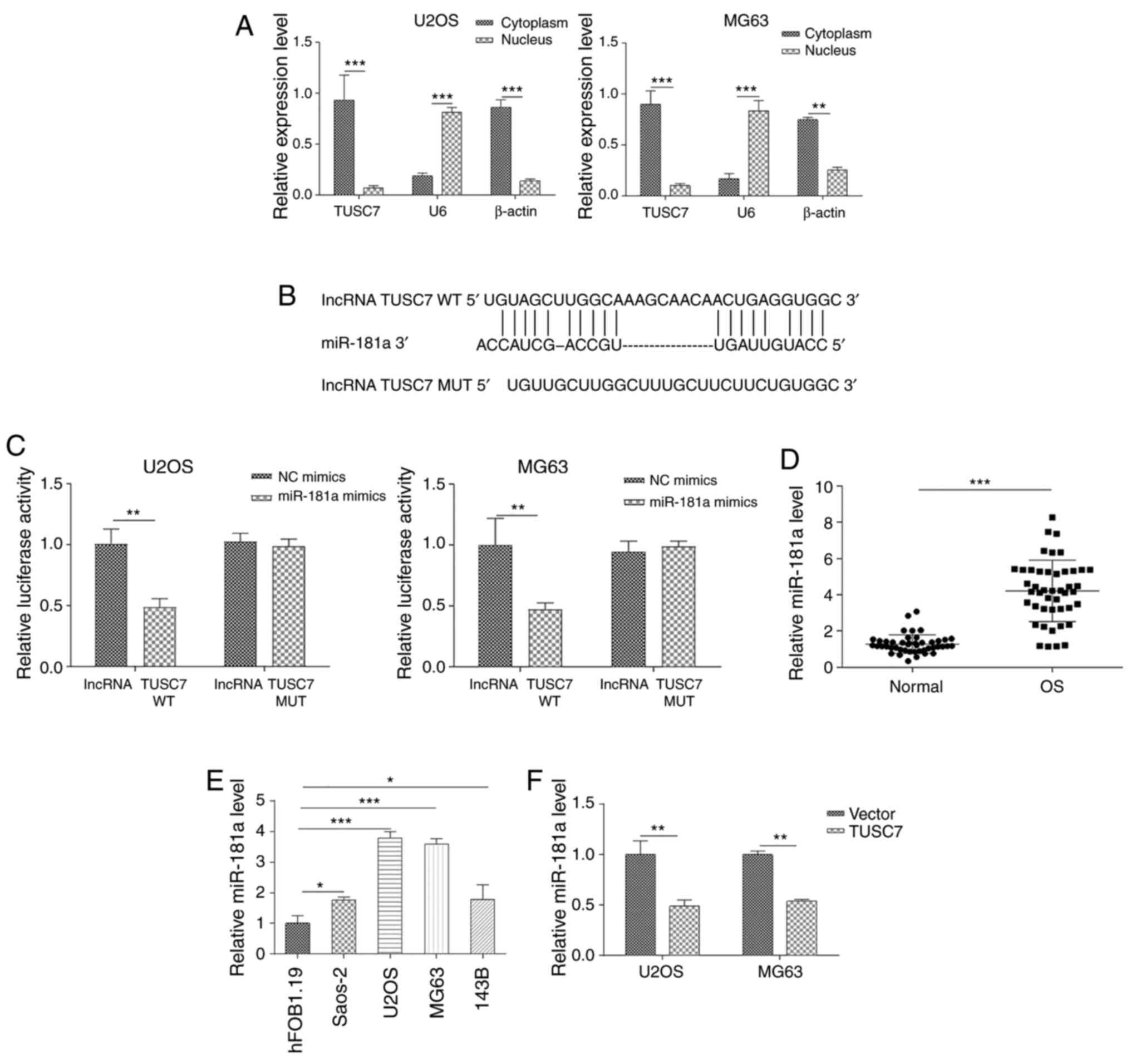
To examine the role of TUSC7 in OS progression, the
subcellular localization of TUSC7 in U2OS and MG63 cells was
analyzed. RT-qPCR revealed that TUSC7 was primarily localized in
the cytoplasm (Fig. 3A),
indicating its role in post-transcriptional regulation of gene
expression. Previous studies have proven that lncRNA expression in
the cytoplasm is vital for pathophysiological processes and
partially occurs by sponging miRNAs (28,29). Since the promoting role of
miR-181a in OS progression is well known and its expression levels
have been reported to be aberrantly elevated in OS tissues and
cells (20), the present study
examined whether TUSC7 inhibited OS progression by sponging
miR-181a expression. First, it was revealed that miR-181a was
directly bound to the 3′-untranslated regions (UTR) of TUSC7 using
the star-Base database (Fig. 3B).
Subsequently, a luciferase reporter vector ligated with TUSC7-WT
and TUSC7-Mut sequences was constructed. The results revealed that
luciferase activities of TUSC7-WT, but not TUSC7-Mut, were
significantly decreased following transfection with miR-181a mimics
(Fig. 3C). In addition, miR-181a
expression was significantly increased in both OS tissues and cell
lines (Fig. 3D and E). As shown
in Fig. 3F, the expression levels
of miR-181a were significantly downregulated by TUSC7
overexpression compared with the control vector. In summary, the
present results indicated that TUSC7 regulated OS progression by
binding to miR-181a.
RASSF6 is a direct target of miR-181a in
OS cell lines
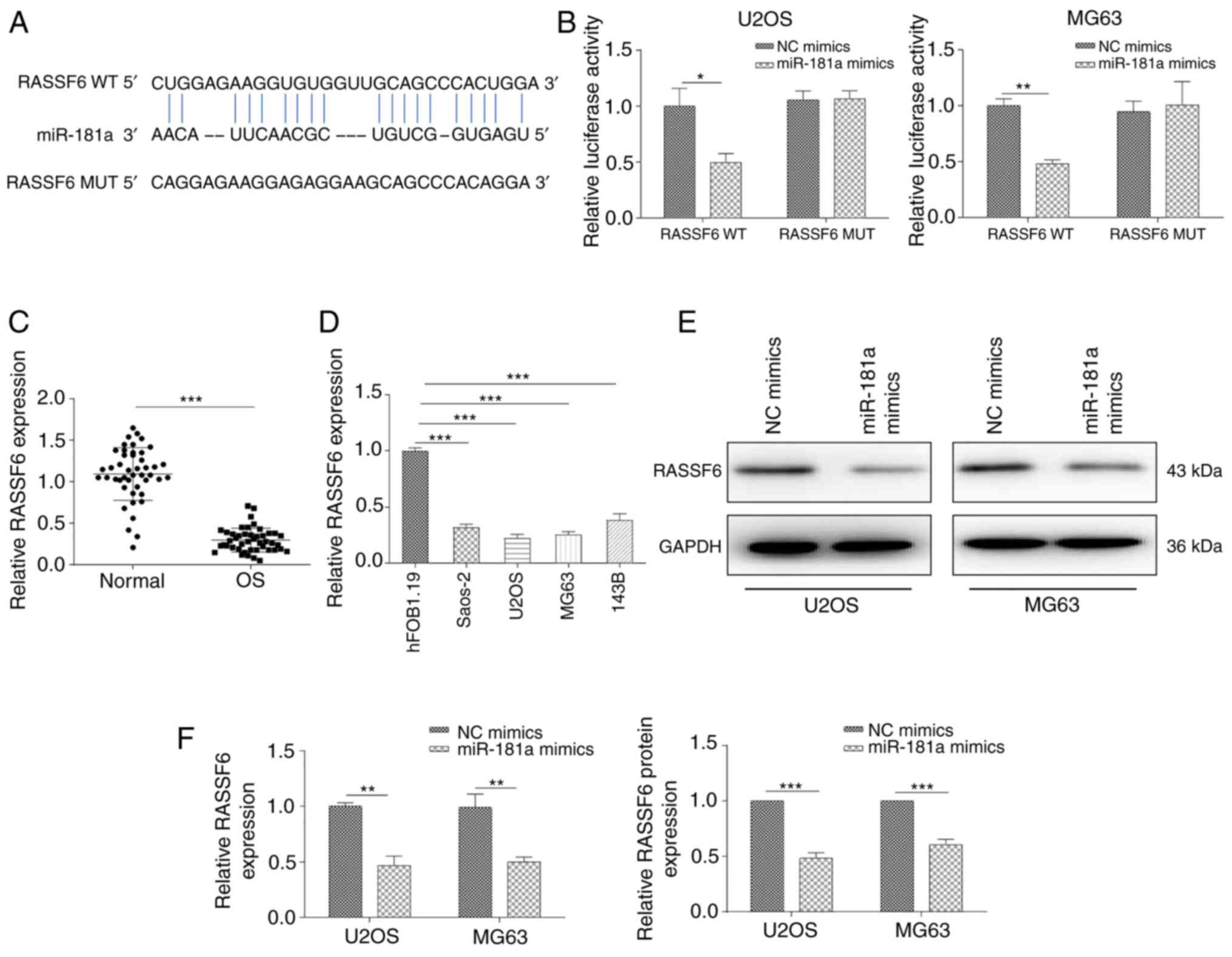
Having determined the potential competitive
mechanism between miR-181a and TUSC7, the present study attempted
to identify critical direct targets underlying the mechanistic
contribution of miR-181a to OS progression. The predicted binding
site of miR-181a at the 3′-UTR of RASSF6 was determined using the
starBase online database. The potential binding sequences are shown
in Fig. 4A. A dual-luciferase
reporter assay was further conducted to validate direct binding of
the 3′-UTR of RASSF6 mRNA with miR-181a. The results revealed that
miR-181a mimics significantly suppressed luciferase activity of the
WT 3′-UTR of RASSF6, but not of the Mut 3′-UTR of RASSF6 (Fig. 4B). In addition, RT-qPCR assays
demonstrated that RASSF6 expression was significantly decreased in
OS tissues and cell lines (Fig. 4C
and D). RASSF6 expression at both the protein and mRNA levels
was significantly suppressed after transfection with the miR-181a
mimic (Fig. 4E and F). In
summary, the current findings suggested that RASSF6 may act as a
down-stream effector of miR-181a in OS progression.
TUSC7/miR-181a/RASSF6 regulates the
progression of OS
The present study demonstrated that TUSC7 served as
an endogenous sponge for miR-181a in OS. Thus, whether TUSC7
inhibited OS cell proliferation, migration and invasion by
inhibiting miR-181a expression was investigated. As shown in
Fig. 5A, miR-181a expression was
significantly increased in both U2OS and MG63 cells after
transfection with the miR-181a mimic compared with the NC mimic.
CCK-8 and colony formation assays demonstrated that miR-181a
significantly reversed TUSC7-induced inhibition of cell
proliferation in cells co-transfected with miR-181a mimic and TUSC7
vector (Fig. 5B and C).
Additionally, Transwell assay revealed that TUSC7 overexpression
significantly decreased cell migration and invasion, which was
rescued by the addition of miR-181a (Fig. 5D). Furthermore, flow cytometry
assays demonstrated that miR-181a overexpression rescued
TUSC7-induced apoptosis in OS cells (Fig. 5E) and reversed TUSC7-induced
decreases in Bcl2 expression and increases in Bax, Caspase-3 and
Caspase-8 expression, as shown by western blotting (Fig. 5F). RT-qPCR and western blot assays
revealed that the expression levels of RASSF6 were significantly
increased following TUSC7 overexpression, and this effect was
significantly reversed by the addition of the miR-181a mimic
(Fig. 5G and H).
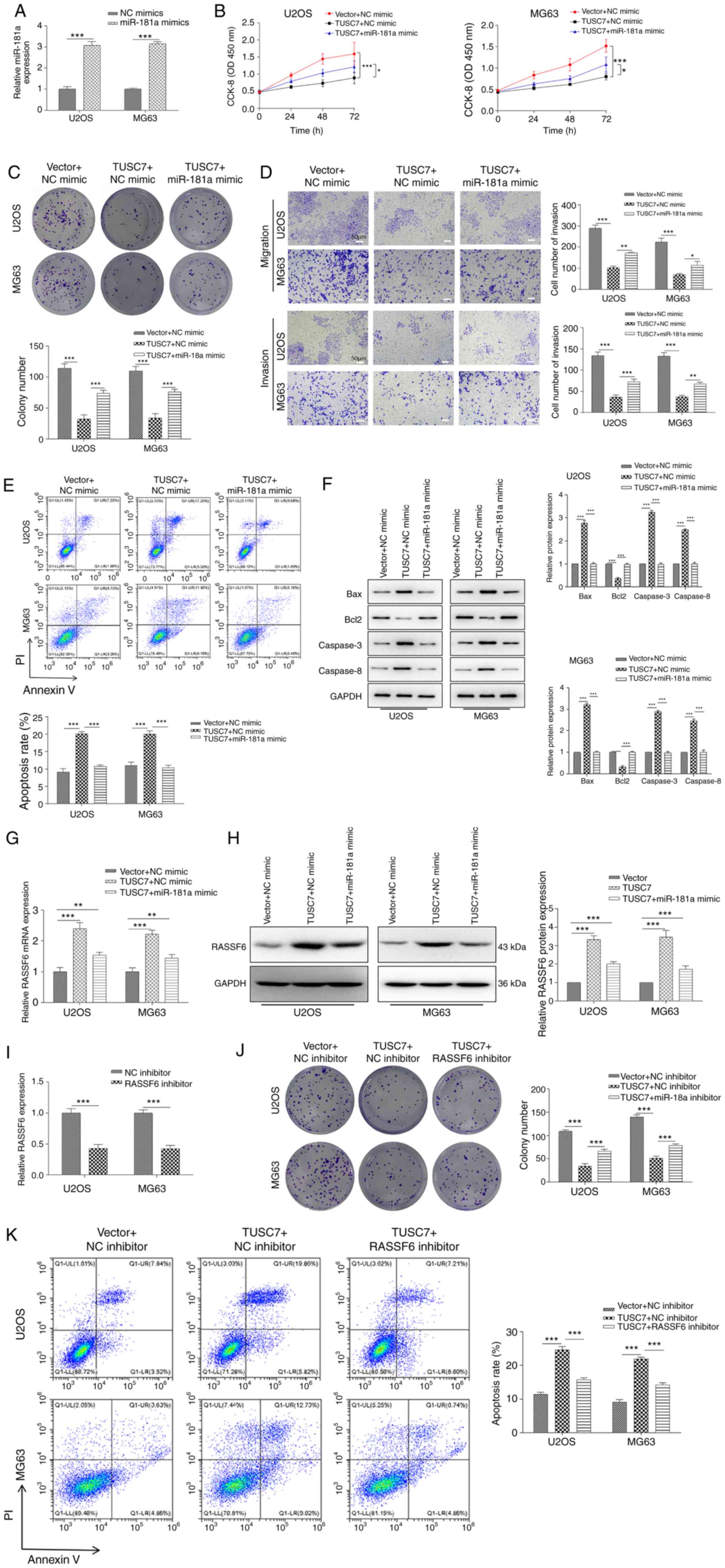 | Figure 5TUSC7/miR-181a/RASSF6 regulates the
progression of OS. (A) Relative miR-181a expression measured via
RT-qPCR in miR-181a mimics-transfected U2OS and MG63 cells.
Proliferation of U2OS and MG63 cells co-transfected with TUSC7 and
miR-181a mimics identified by (B) CCK-8 and (C) colony formation
assays. (D) Migration and invasion of U2OS and MG63 cells
co-transfected with TUSC7 and miR-181a mimics measured by Transwell
assays (scale bar, 50 µm). (E) Apoptosis of OS cells
determined by flow cytometry in TUSC7- and miR-181a
mimics-transfected U2OS and MG63 cells. (F) Expression levels of
the apoptosis-associated proteins Bcl2, Bax, Caspase-3 and
Caspase-8 detected via western blot assays in TUSC7- and miR-181a
mimics-transfected U2OS and MG63 cells. (G) RT-qPCR and (H) western
blot assays of the expression levels of RASSF6 in TUSC7- and
miR-181a mimics-transfected U2OS and MG63 cells. (I) Relative
RASSF6 expression measured by RT-qPCR in RASSF6 inhibitor-treated
U2OS and MG63 cells. (J) Proliferation of U2OS and MG63 cells
co-transfected with TUSC7 and RASSF6 inhibitor detected by colony
formation assay. (K) Apoptosis of OS cells determined by flow
cytometry in TUSC7- and RASSF6 inhibitor-transfected U2OS and MG63
cells. Data are presented as the mean ± SD. *P<0.05,
**P<0.01 and ***P<0.001. CCK-8, Cell
Counting Kit-8; OD, optical density; RT-qPCR, reverse
transcription-quantitative PCR; NC, negative control; miR,
microRNA; OS, osteosarcoma; RASSF6, Ras association domain family
member 6; TUSC7, tumor suppressor candidate 7. |
In addition, to further detect the functions of
RASSF6 on the effects induced by TUSC7, a RASSF6 inhibitor was used
to treat OS cells. As shown in Fig.
5I, the RASSF6 inhibitor significantly attenuated mRNA
expression levels of RASSF6 in U2OS and MG63 cells compared with
the NC inhibitor. Colony formation assays revealed that the RASSF6
inhibitor significantly reversed the TUSC7-induced decrease in cell
proliferation (Fig. 5J). Flow
cytometry demonstrated that TUSC7 overexpression significantly
increased apoptosis, while the RASSF6 inhibitor significantly
reversed the effects of TUSC7 (Fig.
5K). The current data illustrated that TUSC7 directly regulated
RASSF6 expression by competitively binding to miR-181a as a miRNA
sponge.
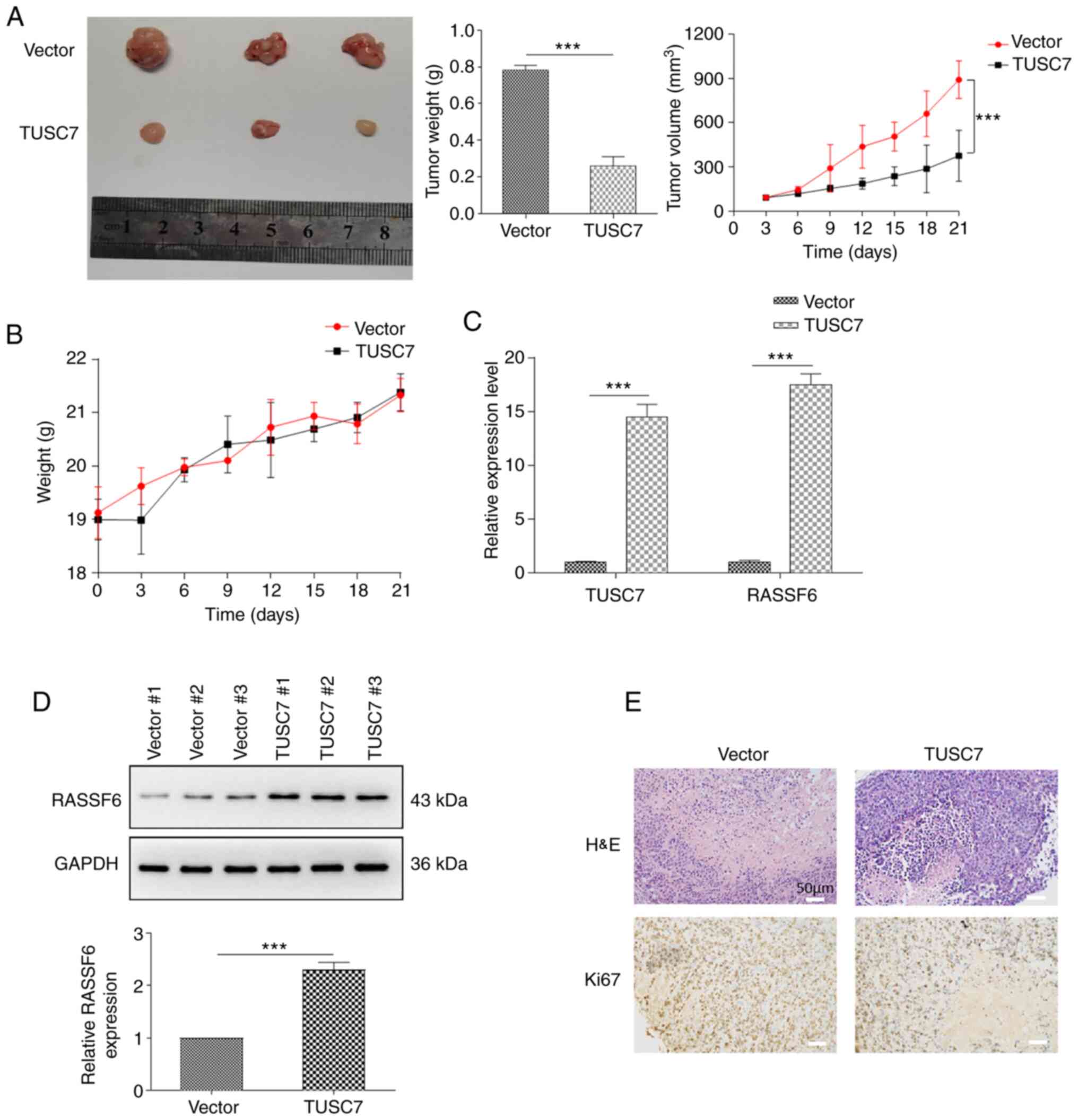
TUSC7 overexpression inhibits tumor
growth in vivo
To further explore the roles of TUSC7 in OS
tumorigenesis, U2OS cells were transfected with control or TUSC7
over-expression vectors for in vivo analysis of tumor
growth. Cells were subcutaneously injected into BALB/c nude mice,
which were euthanized for tumor collection after 5 weeks.
Consistent with the in vitro results, overexpression of
TUSC7 resulted in significantly smaller tumors (Fig. 6A). Moreover, tumor volumes of
TUSC7-overexpressing mice were significantly smaller compared with
those in the control vector group, while there was no significant
difference between the body weights of TUSC7-overexpressing mice
and the control vector group (Fig. 6A
and B). In addition, RASSF6 expression at both the protein and
mRNA levels was significantly increased in response to
overexpression of TUSC7 (Fig. 6C and
D). Additionally, H&E and Ki-67 staining revealed that the
TUSC7-overexpressing group exhibited a decrease in proliferating
cells (Fig. 6E). Overall, the
present results demonstrated that TUSC7 negatively influenced OS
progression.
Discussion
lncRNAs and miRNAs are reportedly dysregulated in
numerous types of cancer, including OS (12,30,31). lncRNAs act as carcinogenic factors
or tumor suppressor factors by regulating different mRNAs by
sponging them (32,33). Several lncRNAs have been reported
to be involved in OS progression (18,34,35). For example, as oncogenes, lncRNA
PGM5-AS1 promotes epithelial-mesenchymal transition, invasion and
metastasis of OS cells by impairing miR-140-5p-mediated fibrillin-1
inhibition (34). On the other
hand, lncRNAs can act as tumor suppressors. lncRNA CEBPA-AS1
expression is decreased in OS tissues and cell lines, and
overexpression of CEBPA-AS1 inhibits proliferation and migration
while enhancing apoptosis in OS cells through the
miR-10b-5p/nuclear receptor corepressor 2/Notch signaling pathway
(35). lncRNA TUSC7 has been
previously demonstrated to be a tumor suppressor in OS (18). The present study confirmed that
TUSC7 was expressed at low levels in OS tissues and cell lines,
while overexpression of TUSC7 inhibited proliferation and invasion,
and promoted apoptosis in OS cells.
Functionally, the current study revealed that
overexpression of TUSC7 inhibited cell proliferation and migration,
while promoting apoptosis both in vitro and in vivo.
Numerous studies have indicated that lncRNAs function as ceRNAs by
competitively binding miRNAs, eliminating the inhibition of miRNAs
on their target gene transcripts (36,37). The present study demonstrated that
TUSC7 was primarily localized in the cytoplasm, and a luciferase
reporter assay revealed that TUSC7 competitively bound to miR-181a.
miR-181a has previously been reported to significantly promote
proliferation and inhibit apoptosis in OS cells (19). However, to the best of our
knowledge, there are no studies on the role of lncRNAs on miR-181a
or its mechanism in OS. In the current study, RT-qPCR assays
revealed that miR-181a expression was significantly increased in OS
tissues and cell lines, and was negatively associated with TUSC7
expression.
Multiple members of the RASSF exhibit anticancer
effects (38). RASSF6 is a novel
tumor suppressor that serves an important role in the pathogenesis
of various types of cancer (39,40). For instance, miR-181a-5p promotes
the progression of gastric cancer via RASSF6-mediated MAPK
signaling activation (41).
Moreover, miR-496 promotes migration and epithelial-mesenchymal
transition by targeting RASSF6 in colorectal cancer (42). However, the roles of RASSF6 and
its regulatory mechanism in OS remain unclear. To explore the
association between miRNA and RASSF6, a luciferase reporter assay
was used to identify miR-181a-specific binding to RASSF6. Moreover,
RASSF6 expression was downregulated in OS tissues and cell lines,
and was negatively regulated by miR-181a. The current data
suggested that RASSF6 was a direct target gene of miR-181a.
Additionally, a critical finding of the present
study was that overexpression of miR-181a rescued the inhibitory
effect of TUSC7 on OS cell proliferation, migration and invasion.
Moreover, overexpression of miR-181a significantly decreased the
expression levels of RASSF6 induced by TUSC7 over-expression. The
current data indicated that TUSC7 directly regulated RASSF6
expression by competitively binding to miR-181a as a miRNA
sponge.
In conclusion, the present study defined a novel
role for TUSC7 as a tumor suppressor in OS development by sponging
miR-181a, leading to RASSF6 upregulation. Overall, TUSC7 may
represent a potential therapeutic and prognostic target for OS,
although its clinical value should be further consolidated in
future studies.
Funding
The present study was supported by the National
Natural Scientific Foundation of China (grant no. Bai891018).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
RB, AZ and WL conceived the study. XC, NW and YW
wrote the manuscript and performed the experiments. LS, HX and LW
analyzed the data and designed the figures. SC and YY contributed
to the resources, interpreted the data and critically revised the
manuscript. All authors reviewed the paper and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Affiliated Hospital of Inner Mongolia Medical
University (approval no. Y K D2017142; Hohhot, China). All
procedures involving human participants were performed in
accordance with the ethical standards of the Institutional and
National Research Committee and with the Declaration of Helsinki.
Written informed consent was provided by all patients enrolled in
the study. All mice were treated according to the Guide for the
Care and Use of Laboratory Animals published by the US National
Institutes of Health, and animal experiments were approved by the
Animal Experimentation Ethics Committee of the Affiliated Hospital
of Inner Mongolia Medical University (approval no. Y K
D2017142).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Mirabello L, Troisi RJ and Savage SA:
Osteosarcoma incidence and survival rates from 1973 to 2004: Data
from the surveillance, epidemiology, and end results program.
Cancer. 115:1531–1543. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Isakoff MS, Bielack SS, Meltzer P and
Gorlick R: Osteosarcoma: Current treatment and a collaborative
pathway to success. J Clin Oncol. 33:3029–3035. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Iwamoto Y, Tanaka K, Isu K, Kawai A,
Tatezaki S, Ishii T, Kushida K, Beppu Y, Usui M, Tateishi A, et al:
Multiinstitutional phase II study of neoadjuvant chemotherapy for
osteosarcoma (NECO study) in Japan: NECO-93J and NECO-95J. J Orthop
Sci. 14:397–404. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhou W, Hao M, Du X, Chen K, Wang G and
Yang J: Advances in targeted therapy for osteosarcoma. Discov Med.
17:301–307. 2014.PubMed/NCBI
|
|
5
|
Simpson S, Dunning MD, de Brot S,
Grau-Roma L, Mongan NP and Rutland CS: Comparative review of human
and canine osteosarcoma: Morphology, epidemiology, prognosis,
treatment and genetics. Acta Vet Scand. 59:712017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kornienko AE, Guenzl PM, Barlow DP and
Pauler FM: Gene regulation by the act of long non-coding RNA
transcription. BMC Biol. 11:592013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schmitt AM and Chang HY: Long noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li Z, Yu X and Shen J: Long non-coding
RNAs: Emerging players in osteosarcoma. Tumour Biol. 37:2811–2816.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu S, Gong Y, Yin Y, Xing H and Zhang N:
The multiple function of long noncoding RNAs in osteosarcoma
progression, drug resistance and prognosis. Biomed Pharmacother.
127:1101412020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bhan A, Soleimani M and Mandal SS: Long
noncoding RNA and cancer: A new paradigm. Cancer Res. 77:3965–3981.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lim LJ, Wong SYS, Huang F, Lim S, Chong
SS, Ooi LL, Kon OL and Lee CG: Roles and regulation of long
noncoding RNAs in hepatocellular carcinoma. Cancer Res.
79:5131–5139. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhu KP, Ma XL and Zhang CL: LncRNA ODRUL
contributes to osteosarcoma progression through the miR-3182/MMP2
axis. Mol Ther. 25:2383–2393. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qu Z and Li S: Long noncoding RNA
LINC01278 favors the progression of osteosarcoma via modulating
miR-133a-3p/PTHR1 signaling. J Cell Physiol. Jan 29–2020.Epub ahead
of print. View Article : Google Scholar
|
|
14
|
Chen X, Zhang C and Wang X: Long noncoding
RNA DLEU1 aggravates osteosarcoma carcinogenesis via regulating the
miR-671-5p/DDX5 axis. Artif Cells Nanomed Biotechnol. 47:3322–3328.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ren W, Chen S, Liu G, Wang X, Ye H and Xi
Y: TUSC7 acts as a tumor suppressor in colorectal cancer. Am J
Transl Res. 9:4026–4035. 2017.PubMed/NCBI
|
|
16
|
Yue L and Guo J: LncRNA TUSC7 suppresses
pancreatic carcinoma progression by modulating miR-371a-5p
expression. J Cell Physiol. 2019.Epub ahead of print. View Article : Google Scholar
|
|
17
|
Chang ZW, Jia YX, Zhang WJ, Song LJ, Gao
M, Li MJ, Zhao RH, Li J, Zhong YL, Sun QZ and Qin YR:
LncRNA-TUSC7/miR-224 affected chemotherapy resistance of esophageal
squamous cell carcinoma by competitively regulating DESC1. J Exp
Clin Cancer Res. 37:562018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cong M and Jing R: Long non-coding RNA
TUSC7 suppresses osteosarcoma by targeting miR-211. Biosci Rep.
39:BSR201902912019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhu ZJ, Huang P, Chong YX, Kang LX, Huang
X, Zhu ZX and Nie L: MicroRNA-181a promotes proliferation and
inhibits apoptosis by suppressing CFIm25 in osteosarcoma. Mol Med
Rep. 14:4271–4278. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ba Z, Gu L, Hao S, Wang X, Cheng Z and Nie
G: Downregulation of lncRNA CASC2 facilitates osteosarcoma growth
and invasion through miR-181a. Cell Prolif. 51:e124092018.
View Article : Google Scholar
|
|
21
|
Jones KB, Salah Z, Del Mare S, Galasso M,
Gaudio E, Nuovo GJ, Lovat F, LeBlanc K, Palatini J, Randall RL, et
al: miRNA signatures associate with pathogenesis and progression of
osteosarcoma. Cancer Res. 72:1865–1877. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Savary G, Dewaeles E, Diazzi S, Buscot M,
Nottet N, Fassy J, Courcot E, Henaoui IS, Lemaire J, Martis N, et
al: The long noncoding RNA DNM3OS Is a reservoir of FibromiRs with
major functions in lung fibroblast response to TGF-β and pulmonary
fibrosis. Am J Respir Crit Care Med. 200:184–198. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xiao G, Yao J, Kong D, Ye C, Chen R, Li L,
Zeng T, Wang L, Zhang W, Shi X, et al: The long noncoding RNA
TTTY15, which is located on the Y chromosome, promotes prostate
cancer progression by sponging let-7. Eur Urol. 76:315–326. 2019.
View Article : Google Scholar
|
|
24
|
Yamamura S, Imai-Sumida M, Tanaka Y and
Dahiya R: Interaction and cross-talk between non-coding RNAs. Cell
Mol Life Sci. 75:467–484. 2018. View Article : Google Scholar :
|
|
25
|
Lopez-Urrutia E, Bustamante Montes LP,
Ladron de Guevara Cervantes D, Perez-Plasencia C and Campos-Parra
AD: Crosstalk between long non-coding RNAs, Micro-RNAs and mRNAs:
Deciphering molecular mechanisms of master regulators in cancer.
Front Oncol. 9:6692019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
27
|
National Research Council (US): Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the Care and Use of Laboratory Animals. 8th
edition. National Academies Press; Washington, DC: 2011
|
|
28
|
Thomson DW and Dinger ME: Endogenous
microRNA sponges: Evidence and controversy. Nat Rev Genet.
17:272–283. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou Y, Li X and Yang H: LINC00612
functions as a ceRNA for miR-214-5p to promote the proliferation
and invasion of osteosarcoma in vitro and in vivo. Exp Cell Res.
392:1120122020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chi Y, Wang D, Wang J, Yu W and Yang J:
Long non-coding RNA in the pathogenesis of cancers. Cells.
8:10152019. View Article : Google Scholar :
|
|
31
|
Dong P, Xiong Y, Yue J, Xu D, Ihira K,
Konno Y, Kobayashi N, Todo Y and Watari H: Long noncoding RNA NEAT1
drives aggressive endometrial cancer progression via
miR-361-regulated networks involving STAT3 and tumor
microenvironment-related genes. J Exp Clin Cancer Res. 38:2952019.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao CC, Jiao Y, Zhang YY, Ning J, Zhang
YR, Xu J, Wei W and Kang-Sheng G: Lnc SMAD5-AS1 as ceRNA inhibit
proliferation of diffuse large B cell lymphoma via Wnt/β-catenin
pathway by sponging miR-135b-5p to elevate expression of APC. Cell
Death Dis. 10:2522019. View Article : Google Scholar
|
|
33
|
Liu F, Yuan JH, Huang JF, Yang F, Wang TT,
Ma JZ, Zhang L, Zhou CC, Wang F, Yu J, et al: Long noncoding RNA
FTX inhibits hepatocellular carcinoma proliferation and metastasis
by binding MCM2 and miR-374a. Oncogene. 35:5422–5434. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu W, Liu P, Gao H, Wang X and Yan M:
Long non-coding RNA PGM5-AS1 promotes epithelial-mesenchymal
transition, invasion and metastasis of osteosarcoma cells by
impairing miR-140-5p-mediated FBN1 inhibition. Mol Oncol.
14:2660–2677. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xia P, Gu R, Zhang W and Sun YF: lncRNA
CEBPA-AS1 over-expression inhibits proliferation and migration and
stimulates apoptosis of OS cells via notch signaling. Mol Ther
Nucleic Acids. 19:1470–1481. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Huang G, Wang M, Li X, Wu J, Chen S, Du N,
Li K, Wang J, Xu C, Ren H, et al: TUSC7 suppression of Notch
activation through sponging MiR-146 recapitulated the asymmetric
cell division in lung adenocarcinoma stem cells. Life Sci.
232:1166302019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Duan X, Wu Y, Zhang Z and Lu Z:
Identification and analysis of dysregulated lncRNA and associated
ceRNA in the pathogenesis of keloid. Ann Transl Med. 8:2222020.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Volodko N, Gordon M, Salla M, Ghazaleh HA
and Baksh S: RASSF tumor suppressor gene family: Biological
functions and regulation. FEBS Lett. 588:2671–2684. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Iwasa H, Jiang X and Hata Y: RASSF6; the
putative tumor suppressor of the RASSF family. Cancers (Basel).
7:2415–2426. 2015. View Article : Google Scholar
|
|
40
|
Zhu N, Si M, Yang N, Jing Y, Fu Y, Zhao X,
Lin Z and Yang G: Overexpression of RAS-association domain family 6
(RASSF6) inhibits proliferation and tumorigenesis in hepatocellular
carcinoma cells. Oncol Res. 25:1001–1008. 2017. View Article : Google Scholar
|
|
41
|
Mi Y, Zhang D, Jiang W, Weng J, Zhou C,
Huang K, Tang H, Yu Y, Liu X, Cui W, et al: miR-181a-5p promotes
the progression of gastric cancer via RASSF6-mediated MAPK
signalling activation. Cancer Lett. 389:11–22. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang H, Yan B, Zhang P, Liu S, Li Q, Yang
J, Yang F and Chen E: MiR-496-promotes migration and
epithelial-mesenchymal transition by targeting RASSF6 in colorectal
cancer. J Cell Physiol. 235:1469–1479. 2020. View Article : Google Scholar
|