Introduction
Heart failure (HF), as a heterogeneous clinical
syndrome, mainly develops as a consequence of cardiac overload and
injury (1). HF results in low
quality of life given its high mortality and incidence rate
(2). Elderly individuals over 65
years of age and women are more likely to develop HF (3,4).
In addition, HF patients may experience breathing problems,
fatigue, poor endurance during exercise and fluid retention
(5). Chronic inflammatory
processes and lipid metabolism disorders are implicated in HF
(6). Risk factors, including
diseases (ischemic heart disease, hypertension and diabetes) and
unhealthy living habits (tobacco use and high-fat diets), function
as predictors for HF incidence and severity (7). Although progress has been made in
the development of new drugs for the treatment of HF, the efficacy
is not satisfactory (8).
Therefore, given the significant health risks caused by HF, it is
crucial to identify new and more effective methods to treat HF.
Recently, some long noncoding RNAs (lncRNAs) have been demonstrated
to be involved in the HF-related lncRNA-mRNA network, demonstrating
potent diagnostic value for HF (9). Circulating lncRNAs are also useful
biomarkers for myocardial infarction and other adult heart diseases
(10). Therefore, the aim of the
present study was to identify a novel lncRNA-based therapy for HF
from the aspects of lipid metabolism and inflammatory
responses.
LncRNAs, which are RNA molecules greater than 200
nucleotides in length (11),
regulate cholesterol [(mostly oxidized low-density lipoprotein
(oxLDL)] accumulation and inflammation in macrophages in the
progression of atherosclerosis (AS) (12) which is closely related to HF
(13). LncRNA
metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) is
highly conserved and widely expressed in mammalian tissue cells
(14). MALAT1 is involved in the
pathogenesis of AS (15) and
regulates oxLDL-induced endothelial inflammation during AS
(16). However, studies on the
regulatory mechanism of MALAT1 in HF are limited. In addition, it
has been shown that lncRNAs interacting with microRNAs (miRNAs) to
regulate mRNA are strongly linked to the development of
cardiovascular diseases, including HF (17). miRNAs, which are recognized as
small noncoding RNAs, modulate gene expression, and potentially
serve as biomarkers for HF (18).
miR-532-3p is involved in atherothrombotic events that may lead to
HF (19,20). miR-532-3p is of great value in the
regulation of diverse metabolic disorders, including inflammation
and obesity (21). Nevertheless,
the direct role of miR-532-3p in HF has not been reported.
Therefore, it was reasonable to hypothesize that
MALAT1 played an underlying role in regulating lipid metabolism and
inflammatory responses in HF through the miRNA-mRNA system.
Consequently, we performed a series of histological and molecular
experiments to identify the lncRNA-miRNA-mRNA network of MALAT1 and
study the underlying molecular machinery with the aim of providing
some novel therapies against HF.
Materials and methods
Ethics approval
This study was approved and supervised by the human
ethics committee of Yantai Yuhuangding Hospital. All the subjects
signed the informed consent. All experiments were approved by the
animal ethics committee of Yantai Yuhuangding Hospital.
Clinical samples
From May 2018 to June 2019, 57 HF patients (39 males
and 18 females, aged 48-73 years) diagnosed via coronary
angiography were enrolled in Yantai Yuhuangding Hospital. In
addition, 48 healthy controls (28 males and 20 females, aged 50-73
years) were included during the same period. Venous blood was
extracted from the subjects under fasting conditions and stored at
-80°C, and relevant physiological and biochemical indexes were
detected in the patients and controls. Inclusion criteria for
patients with HF were as follows: i) At least one major coronary
artery exhibited >60% stenosis; ii) informed consent was signed.
Exclusion criteria were: i) Unstable angina pectoris or myocardial
infarction; ii) complicated with another organic heart disease;
iii) presence of severe hypertension, diabetes, shock, arrhythmia,
liver and kidney dysfunction, familial hypercholesterolemia, and/or
malignant tumor or inflammatory diseases.
Animal treatment and grouping
A total of 40 healthy Sprague-Dawley (SD) male rats
[220-260 g, SYXK (Zhejiang) 2019-0021] purchased from Hangzhou
Qingda Kerui Biotechnology Co., Ltd. (Hangzhou, Zhejiang, China)
were housed under standard conditions for one week before the
experiments. All experimental animals were kept in a specific
pathogen-free animal room (22-24°C, light/dark=12/12 h, humidity
50%), and could freely obtain standard food and water.
After rats were anesthetized using intraperitoneal
injection of 60 mg/kg pentobarbital sodium, the left anterior
descending coronary artery was ligated to construct a rat model of
HF (22). Rats in the sham group
were only threaded without ligation. The rat model was identified
using an electrocardiogram (ECG) 3 days after the operation. HF
model rats were assigned to 3 groups: HF group (injection of the
same amount of normal saline via the tail vein), HF + small
hairpin-negative control (sh-NC) group (injection of 50 nM sh-NC
via the tail vein), and HF + sh-MALAT1 group (injection of 50 nM
sh-MALAT1 via the tail vein). The normal saline, sh-NC and
sh-MALAT1 groups were injected equally twice a week for 4 weeks.
The above adenovirus vectors were purchased from Guangzhou RiboBio
Co., Ltd.
Cardiac function and hemodynamics
Cardiac function and hemodynamics were determined as
described previously (22).
Briefly, the rats (6 rats in each group) were anesthetized by
inhalation of 1.5-2% isoflurane prior to scanning using the
isoflurane anesthesia system (JD Medical Dist Co Inc.). Heart rate
(HR) was measured by inserting a right ventricular catheter into
the right ventricle via the right jugular vein. Left ventricular
ejection fraction (LVEF) and fraction shortening (FS) were detected
dynamically using a cardiac color Doppler ultrasonic cardiogram
(ACSON512, Philips). Then, the BL-420S biological function
experimental system (Taimeng Technology Co., Ltd.) was used to
monitor the cardiac function of rats. Left ventricular systolic
pressure (LVSP), left ventricular end-diastolic pressure (LVEDP),
and left ventricular maximum rates of rise and fall (±dp/dtmax) of
rats in each group were recorded.
Sample collection
After the detection of cardiac function and
hemodynamics, rats were euthanized by intraperitoneal injection of
excessive pentobarbital sodium (800 mg/kg) (23), and serum was collected and stored.
Some rats in each group were used to generate homogenate, and the
myocardial tissues of the remaining rats were fixed in 4%
paraformaldehyde buffer. Paraffin-embedded tissues were routinely
sectioned at 4 µM for tissue staining.
Hematoxylin and eosin (H&E)
staining
After euthanasia, the rats were perfused with
phosphate-buffered saline (PBS) from the left ventricle. Myocardial
tissues were isolated, fixed with 4% paraformaldehyde at 25°C for 4
h and embedded in paraffin. Paraffin-embedded sections were
deparaffinized and dehydrated. Then, sections at 4 µm
underwent treatment with xylene I, xylene II and gradient alcohol
successively for 5 min each. Following 2-min washing with water,
the sections were stained with H&E for 8 min at 25°C, washed
with water, differentiated using 1% hydrochloric acid alcohol for 5
sec, and then washed 3 times with water. The samples were washed
with 0.25% ammonia water for 1 min followed by 3 washes with water
for 2 min each. Next, the sections were stained with 1% eosin for
30 sec, washed with water three times, and immersed in gradient
alcohol, xylene I and xylene II for 3 min separately. After the
xylene on the slide was completely dried, the slide was sealed with
neutral gum, covered by a clean cover slide, and observed under an
optical microscope (BX 51, Olympus Optical Co., Ltd.).
Terminal deoxynucleotidyl transferase
(TdT)-mediated dUTP nick end labeling (TUNEL) staining
After paraffin-embedded sections were
deparaffinized, surgery was performed according to the instructions
of TUNEL apoptosis detection kit (Beyotime Biotechnology Co.,
Ltd.). TUNEL-positive cells were observed under the optical
microscope after development using 3,3′-diaminobenzidine (DAB).
After dewaxing and hydration, the paraffin sections
were added with proteinase K working solution at 37°C for 20 min.
After PBS washing, the sections were immersed in sealing solution
(3% H2O2 dissolved in methanol) at room
temperature (25°C) for 10 min. Then the sections were incubated
with TUNEL reaction solution at 37°C for 60 min, and then
2,4-diaminobutyric acid (DAB) color-developing solution was added.
After color development, 4′,6-diamidino-2-phenylindole (DAPI) (100
ng/ml, at 25°C) was added for nuclear staining for 10 min, and then
observed under fluorescence microscope (Olympus, BX51). At least
three sections were observed in each tissue. Three visual fields
(×400) were randomly selected for statistical analysis.
Myocardial cell culture and grouping
H9C2 myocardial cells of rats (Cell Bank of Chinese
Academy of Sciences) were cultured in Dulbecco's modified Eagle's
medium (DMEM) (Beijing Solarbio) containing 10% fetal bovine serum
(FBS) at 37°C with 5% CO2. When cells grew to 80%
confluency, the cells were allocated into the control group (H9C2
myocardial cells were cultured normally) and isoproterenol (ISO)
group (H9C2 myocardial cells were treated with 80 µM ISO for
48 h) (24). In addition, cells
were transfected with si-NC, si-MALAT1, inhibitor NC, miR-532-3p
inhibitor, miR-532-3p inhibitor + si-NC and miR-532-3p inhibitor +
si-MALAT1 (all from Guangzhou RiboBio) using Lipofectamine 2000
(Thermo Fisher Scientific) as per the manufacturer's instructions.
After transfection for 24 h, the cells were treated with 80
µM ISO for 48 h and collected for subsequent
experiments.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
TRIzol reagent was used to extract total RNA from
myocardial tissues and cells of rats in each group, and RNA was
reverse transcribed into cDNA using the PrimeScript RT reagent kit
(RR047A, Takara Bio Inc.) or NcodeTM miRNA First-Strand cDNA
Synthesis (Thermo Scientific Fisher). The synthesized cDNA was
detected by RT-qPCR using the Fast SYBR-Green PCR kit (Applied
Biosystems, Inc.) and the ABI PRISM 7300 RT-PCR system (Applied
Biosystems). The reaction system (20 µl) included 2X SYBR
Green (10 µl), upstream and downstream primers (20
µM; 0.3 µl for each), cDNA (1.0 µl) and
RNase-free dH2O (8.4 µl). The reaction procedure
was 95°C for 5 min followed by 40 cycles of 95°C for 30 sec and
60°C for 1 min. Each sample was assessed in triplicate. U6 or
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) served as internal
references. The relative expression of related genes was analyzed
using the 2−ΔΔCq method as follows: ΔΔCq=(Cq target
gene in the experimental group-Cq internal reference in
the experimental group)-(Cq target gene in the control
group-Cq internal reference in the control group)
(25). Primers are presented in
Table I.
 | Table IRT-qPCR primer sequences. |
Table I
RT-qPCR primer sequences.
| Gene | Primer
sequence |
|---|
|
hsa-MALAT1 | F:
5′-AAAGCAAGGTCTCCCCACAAG-3′ |
| R:
5′-GGTCTGTGCTAGATCAAAAGGCA-3′ |
|
rno-MALAT1 | F:
5′-AGCGGAAGAACGAATGTAAC-3′ |
| R:
5′-GAACAGAAGGAAGAGCCAAG-3′ |
|
rno-miR-532-3p | F:
5′-ACGGCTTTTCTCTTCCATGCCT-3′ |
| R:
5′-CAGTGCAGGGTCCGAGGTAT-3′ |
| rno-U6 | F:
5′-CTCGCTTCGGCAGCACA-3′ |
| R:
5′-AACGCTTCACGAATTTGCGT-3′ |
|
rno-LDLR | F:
5′-GTGGCAGTAGTGAGTGTATC-3′ |
| R:
5′-ATCCCAAAGACGAGAAGT-3′ |
|
hsa-GAPDH | F:
5′-GCAACTAGGATGGTGTGGCT-3′ |
| R:
5′-TCCCATTCCCCAGCTCTCATA-3′ |
|
rno-GAPDH | F:
5′-CTCCTCGAAGTACCCTGTGC-3′ |
| R:
5′-CATGGTGCAGCGATGCTTTA-3′ |
Western blot analysis
Total protein in myocardial tissues and cells of
rats in each group was extracted, and the protein concentration was
measured using a bicinchoninic acid (BCA) protein quantification
kit (Boster Biological Technology Co., Ltd.). Protein (30
µg) was separated by 10% SDS-PAGE, transferred to
polyvinylidene fluoride membranes and blocked with 5% bovine serum
albumin at room temperature for 2 h to block non-specific binding.
Membranes were then incubated with a primary antibody against
low-density lipoprotein receptor (LDLR) (1/200, no. ab30532) at 4°C
overnight and a secondary immunoglobulin G (IgG) antibody (1
µg/ml, no. ab8226) (Abcam Inc.) at room temperature for 1 h.
Then, the membranes were washed with 1X PBS containing 0.5% Triton
X-100 (PBST), developed and visualized using enhanced
chemiluminescence solution (EMD Millipore Corporation). Each band
in the western blot images was analyzed for gray value
quantification using ImageJ (National Institute of Health). β-actin
(1/2000, ab205718) was used as an internal reference.
Nuclear and cytoplasmic fractionation
assay
According to the instructions of NE-PER™ nuclear and
cytoplasmic fractionation kit (Thermo Scientific Fisher), MALAT1
expression in the isolated cytoplasm and nuclear extract was
detected using RT-qPCR.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay
After transfection, when the cell confluency reached
80%, H9C2 cells were washed twice with PBS solution and detached
with 0.25% trypsin to prepare a single cell suspension. Then,
3×103-6×103 cells/well were seeded into
96-well plates (0.2 ml/well; repeated for 6 wells) and incubated in
incubators at 37°C containing 5% CO2. Next, the plates
were removed at 24, 48 and 72 h, and the medium was replaced with
medium containing 10% MTT solution (5 g/l) (GD-Y1317, Guduo
Biotechnology Company) for further culture for 4 h at 37°C. Then,
the supernatant was absorbed, and 100 µl dimethyl sulfoxide
(D5879-100ML, Sigma-Aldrich) was added to each well followed by 10
min of gentle agitation and mixing to fully dissolve the formazan
crystal produced by living cells. The optical density (OD) at 490
nm of each well was detected using a microplate reader (Nanjing
Detie Experimental Equipment Co., Ltd.), and the cell survival rate
was calculated. The cell survival rate (%) was calculated as: OD
value in the experimental group/OD value in the control group
×100%. The cell survival rate of the control group was set as
100%.
Flow cytometry
Briefly, cells were collected by centrifugation at
4,500 × g for 5 min at 4°C, and the culture medium was discarded.
Next, the cells were washed twice with cold PBS and suspended with
400 µl 1X binding buffer. Then, 5 µl Annexin
V-fluorescein isothiocyanate (FITC) was added to the cell
suspension, and the suspension was gently mixed and incubated at
4°C for 15 min in the dark. Next, 10 µl propidium iodide
(PI) was added to the cells, and the suspension was gently mixed
for 5 min at 4°C. Cells were assessed within 1 h using a flow
cytometer (FACS Calibur, BD Biosciences). Each experiment was
repeated three times.
Fluorescence in situ hybridization
assay
The localization of MALAT1 in cells was predicted by
online analysis website (http://www.csbio.sjtu.edu.cn/cgi-bin/lncLocator.py).
According to the instructions of the FISH kit (Guangzhou Ribobio),
cells were fixed with 4% paraformaldehyde. PBST was added, and the
samples were blocked with prehybridization solution at 37°C. Cells
were hybridized with lncRNA FISH probe (DNA) overnight at 37°C,
washed with the hybridizing solution at 42°C in the dark and
stained using 4′,6-diamidino-2-phenylindole (DAPI) for 10 min.
Next, the cells were fixed on the glass slide with a sealing agent
in the dark and observed under a fluorescence microscope (BX53,
Olympus).
Dual-luciferase reporter gene assay
Starbase (http://starbase.sysu.edu.cn/index.php) predicted the
binding relationship between MALAT1 and miR-532-3p as well as
miR-532-3p and LDLR. Briefly, 293T cells were seeded into the
6-well plates (2×105 cells/well). After the cells
adhered to the wall, MALAT1-wild type (WT), MALAT1-mutant (MUT),
LDLR-WT, and LDLR-MUT were co-transfected with NC mimic and
miR-552-3p mimic, separately, into 293T cells using Lipofectamine
2000 (Invitrogen) as per the manufacturer's instructions. After
successful transfection, the cells were cultured for 48 h and
collected. Changes in luciferase activity in the cells were
detected according to the protocol of the dual-luciferase detection
kit (D0010, Beijing Solarbio Science & Technology Co., Ltd.).
The fluorescence intensity was detected using a GloMax 20/20
luminometer (Promega). Each experiment was repeated three
times.
RNA pull-down
RNA pull-down was carried out as previously
described (26,27). H9C2 myocardial cells were
transfected with 50 nM of biotinylated Bio-miR-542-3p-WT,
Bio-miR-542-3p-MUT and the corresponding Bio-NC. After 48 h, the
cells were collected, washed with PBS, and vortexed. Next, the
cells were incubated with the specific lysis buffer (Ambion, Inc.)
for 10 min and then underwent a 3-h incubation with M-280
streptavidin magnetic beads precoated with RNase-free and yeast
tRNA (Sigma-Aldrich) at 4°C followed by 2 washes with precooled
lysis buffer, 3 washes with low-salt buffer and one wash with
high-salt buffer. Finally, RNA was purified using TRIzol, and
MALAT1 expression was detected using RT-qPCR.
Detection of biochemical indexes
The levels of creatine kinase isoenzyme (CK-MB),
cardiac troponin I (cTn-I), reactive oxygen species (ROS),
superoxide dismutase (SOD), malondialdehyde (MDA), lipid
metabolism-related indexes [endothelin (ET), total cholesterol
(TC), triglyceride (TG), LDL cholesterol (LDL-C), and high-density
lipoprotein cholesterol (HDL-C)], and inflammatory-related
cytokines [tumor necrosis factor alpha (TNF-α), interleukin-1 beta
(IL-1β), IL-6, transforming growth factor beta (TGF-β), IL-4 and
IL-10] in serum of clinical patients and normal volunteers and in
serum and cells of rats in each group were determined,
respectively, according to the instructions of the ELISA kits
(Nanjing Jiancheng Bioengineering Institute). In addition, ROS in
cells was detected using the DCFDA/H2DCFDA-Cellular ROS assay kit
(cat. no. ab113851, Abcam). The samples were incubated with
DCFDA/H2DCFDA for 20 min. After PBS washing, DAPI (100 ng/ml) was
added for nuclear staining at 25°C for 10 min and the samples were
observed under fluorescence microscope.
Statistical analysis
All data were analyzed by SPSS 21.0 statistical
software (IBM Corp.). The data exhibited a normal distribution as
demonstrated by the Kolmogorov-Smirnov test. The results were
expressed by mean ± standard deviation. One-way or two-way analysis
of variance (ANOVA) was used for the comparison among multiple
groups followed by Tukey's multiple comparison test. In addition,
receiver operating characteristic (ROC) curves were used to
evaluate the diagnostic value of MALAT1. The Chi-square test was
used for gender data analysis, and t-test was used for other
clinical data analysis. The P-value was calculated using a
bilateral test, and P<0.05 indicates that the difference was
statistically significant.
Results
LncRNA MALAT1 exhibited important value
in predicting HF patients
To investigate the diagnostic value of serum lncRNA
MALAT1 levels in HF, we enrolled 48 healthy volunteers and 57 HF
patients and compared their general characteristics (Table II). No significant differences in
sex, age, body mass index (BMI), smoking, alcohol and hypertension,
as well as indexes related to lipid metabolism (TC, TG, and LDL-C)
were identified between the two groups. However, HDL-C in HF
patient serum was reduced compared with that in normal volunteers,
and myocardial injury-related indexes (cTn-I and CK-MB) in HF
patient serum were increased compared with those in normal
volunteers (all P<0.01). To examine the relationship between
MALAT1 and HF, MALAT1 expression in serum of normal volunteers and
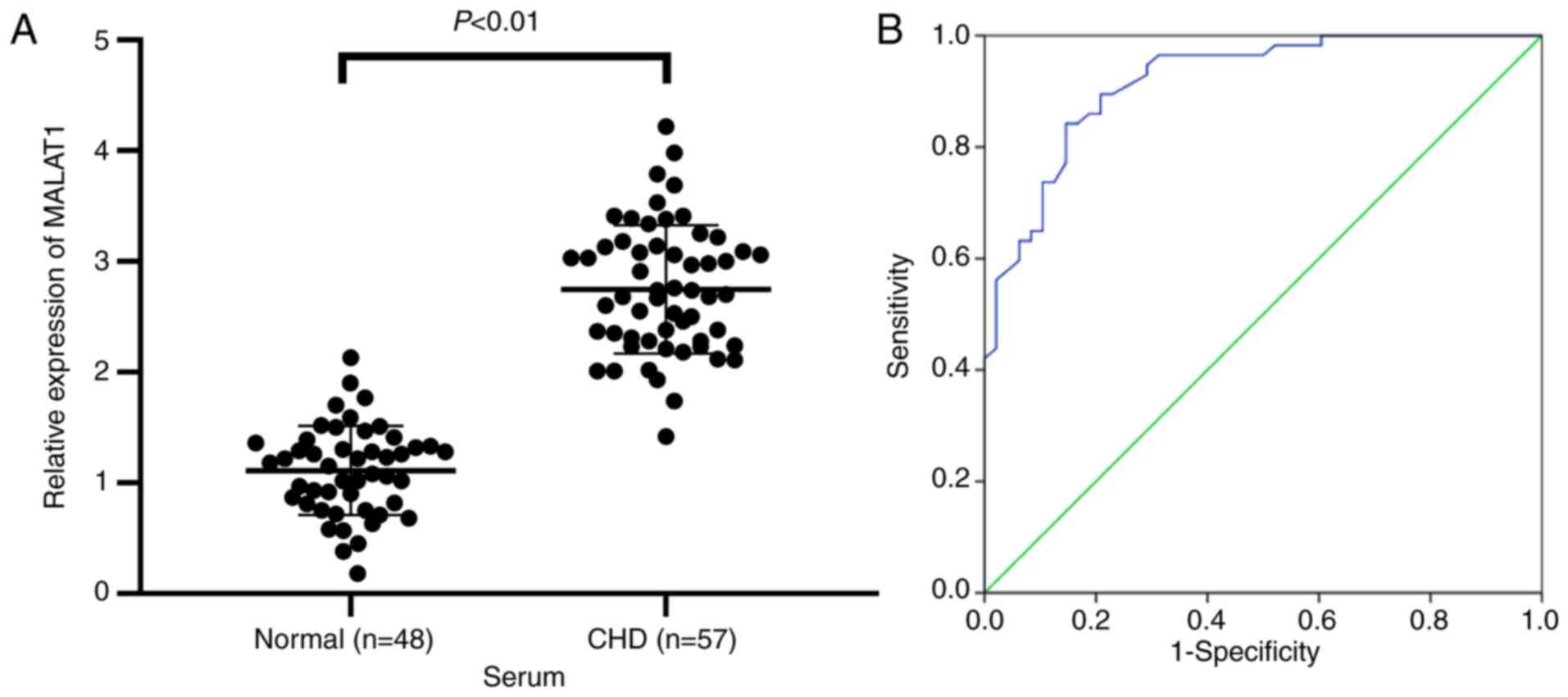
HF patients was measured. The results showed that MALAT1 expression
in serum of HF patients was markedly upregulated compared with that
in the normal controls (P<0.01, Fig. 1A). The diagnostic value of MALAT1
levels in HF patients was analyzed using ROC curves. The area under
the ROC curve was 0.918, the 95% confidence interval was
0.868-0.969. The sensitivity and specificity were 84.2 and 85.4%,
respectively (Fig. 1B). The
results showed that lncRNA MALAT1 exhibited great value in the
prediction of HF patients.
| Table IIBasic clinical characteristics of the
patient. |
Table II
Basic clinical characteristics of the
patient.
| Characteristic | Normal (n=48) | HF (n=57) | P-value |
|---|
| Sex | | | 0.46 |
| Male | 28 (58.33%) | 39 (68.42%) | |
| Female | 20 (41.67%) | 18 (31.58%) | |
| Age | 60.21±13.32 | 61.95±12.16 | 0.49 |
| BMI | 24.21±3.14 | 25.33±3.26 | 0.08 |
| Smoking | 29/19 | 39/18 | 0.22 |
| Alcohol | 25/23 | 33/24 | 0.36 |
| Hypertension | 27/21 | 38/19 | 0.31 |
| TC (mmol/l) | 1.68±0.56 | 1.76±0.61 | 0.49 |
| TG (mmol/l) | 4.21±1.05 | 4.38±1.12 | 0.43 |
| LDL-C (mmol/l) | 3.21±0.86 | 3.56±0.92 | 0.16 |
| HDL-C (mmol/l) | 1.15±0.21 | 0.81±0.16 | <0.01 |
| cTn-I
(µg/l) | 0.17±0.06 | 1.55±0.26 | <0.01 |
| CK-MB (U/l) | 15.37±6.18 | 48.51±6.62 | <0.01 |
Inhibition of lncRNA MALAT1 reduced
myocardial injury in HF rats
To examine the molecular mechanism of MALAT1 in HF,
we first constructed an HF rat model. ECG detection revealed that
compared with the sham-operated rats, HF rats exhibited an
increased ST segment in ECG (Fig.
2A), and MALAT1 expression was clearly upregulated in the
tissues of HF rats (P<0.01, Fig.
2B). To further study the effect of MALAT1 on HF, HF rats were
injected with sh-NC or sh-MALAT1 via the tail vein, and the
relevant indexes were detected. Compared with sh-NC-treated HF
rats, sh-MALAT1-treated HF rats exhibited significantly decreased
expression of MALAT1 (P<0.01, Fig.
2B). After the establishment of the HF model, HR results
revealed no significant difference in HR between the experimental
group and the control group (Fig.
S1). LVEF, FS, LVSP and ±dp/dtmax levels were reduced, and
LVEDP levels were obviously increased in HF rats (Fig. 2C). However, MALAT1 inhibition
yielded opposite results (all P<0.01, Fig. 2C). ELISA showed that the serum
levels of cTn-I and CK-MB in HF rats were significantly increased
compared with sham-operated rats, and these levels were reversed
after inhibition of MALAT1 (P<0.01, Fig. 2D). H&E staining showed that
the myocardial cells of sham-operated rats exhibited a normal shape
and an ordered arrangement, whereas those of HF rats were swollen,
disordered and extremely damaged. After MALAT1 inhibition, the
degree of myocardial injury in the rats was reduced (Fig. 2E). TUNEL staining showed that the
number of TUNEL-positive cells in HF rats was increased compared
with sham-operated rats; the above results were reversed after
MALAT1 knockdown (P<0.01, Fig.
2F). The above results showed that HF rats exhibited severe
myocardial injury, which was relieved by MALAT1 inhibition.
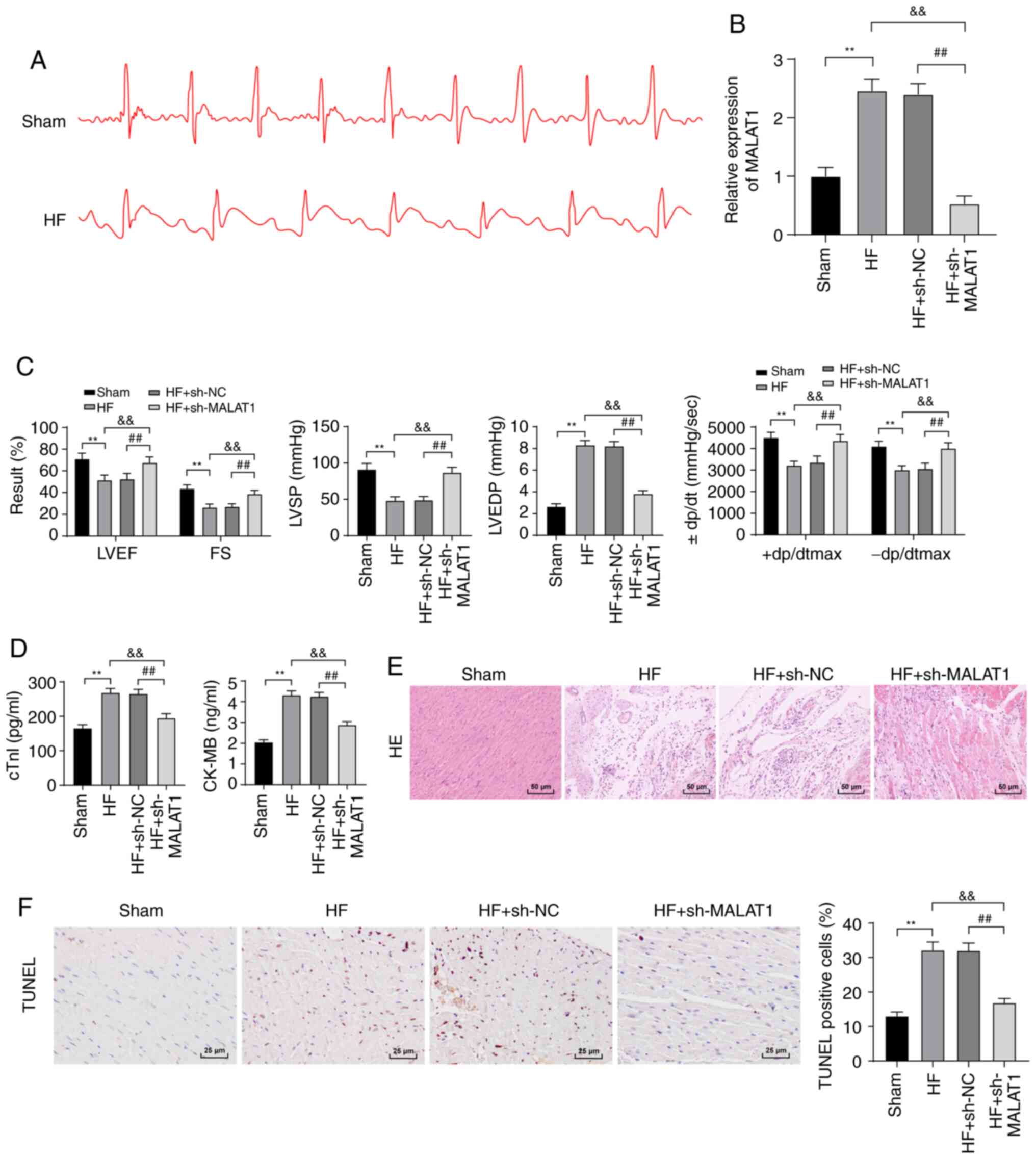 | Figure 2Inhibition of lncRNA MALAT1 reduced
myocardial injury in HF rats. (A) ECG detection of rats, n=10. (B)
Expression of MALAT1 in rat tissues was detected by RT-qPCR, n=4.
(C) Detection of cardiac function and left ventricular hemodynamics
in rats, n=10. (D) ELISA detection of myocardial injury-related
indexes in rats, n=4. (E) Observation of the pathological changes
of rat tissues using H&E staining. Magnification, ×200, n=6.
(F) Detection of myocardial cell apoptosis in rats using TUNEL
staining, Magnification, ×200, n=6; scale, 25 µm. Three
independent repeated tests were conducted. The data were expressed
as the mean ± standard deviation and analyzed using one-way or
two-way ANOVA followed by Tukey's multiple comparison test.
**P<0.01, compared with the sham group;
&&P<0.01, compared with the HF group;
##P<0.01, compared with the HF + sh-NC group. |
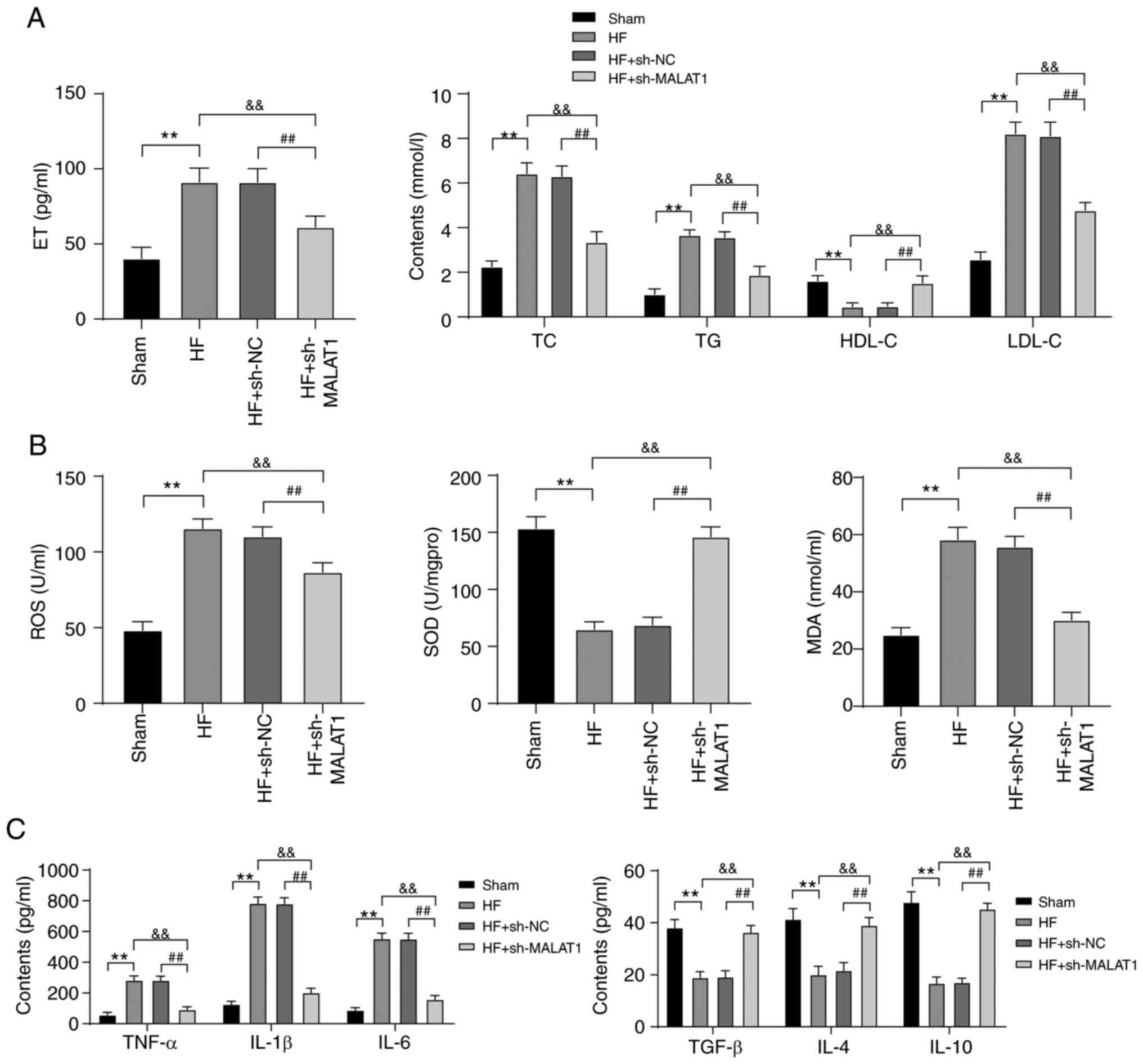
Inhibition of MALAT1 improved lipid
metabolism disorders and relieved inflammation in HF rats
Lipid metabolism disorders and inflammatory
responses are important pathogenic conditions of HF (28,29). Therefore, lipid metabolism and
inflammatory responses in HF rats were detected. Compared with rats
in the sham group, HF rats exhibited lipid metabolism disorders.
Specifically, ET, TC, TG and LDL-C levels were clearly increased,
whereas the level of HDL-C was significantly reduced. However,
inhibition of MALAT1 reversed the above results (all P<0.01)
(Fig. 3A). In addition, in HF
rats, ROS and MDA levels were increased, while SOD levels were
decreased. In addition, the levels of anti-inflammatory cytokines
(TGF-β, IL-4 and IL-10) were reduced, whereas the levels of
pro-inflammatory cytokines (TNF-α, IL-1β and IL-6) were increased.
MALAT1 inhibition yielded the completely opposite results (all
P<0.01, Fig. 3B and C). The
above results showed that HF rats displayed lipid metabolism
disorders and increased levels of lipid oxidation and inflammation
responses, which were alleviated after MALAT1 downregulation.
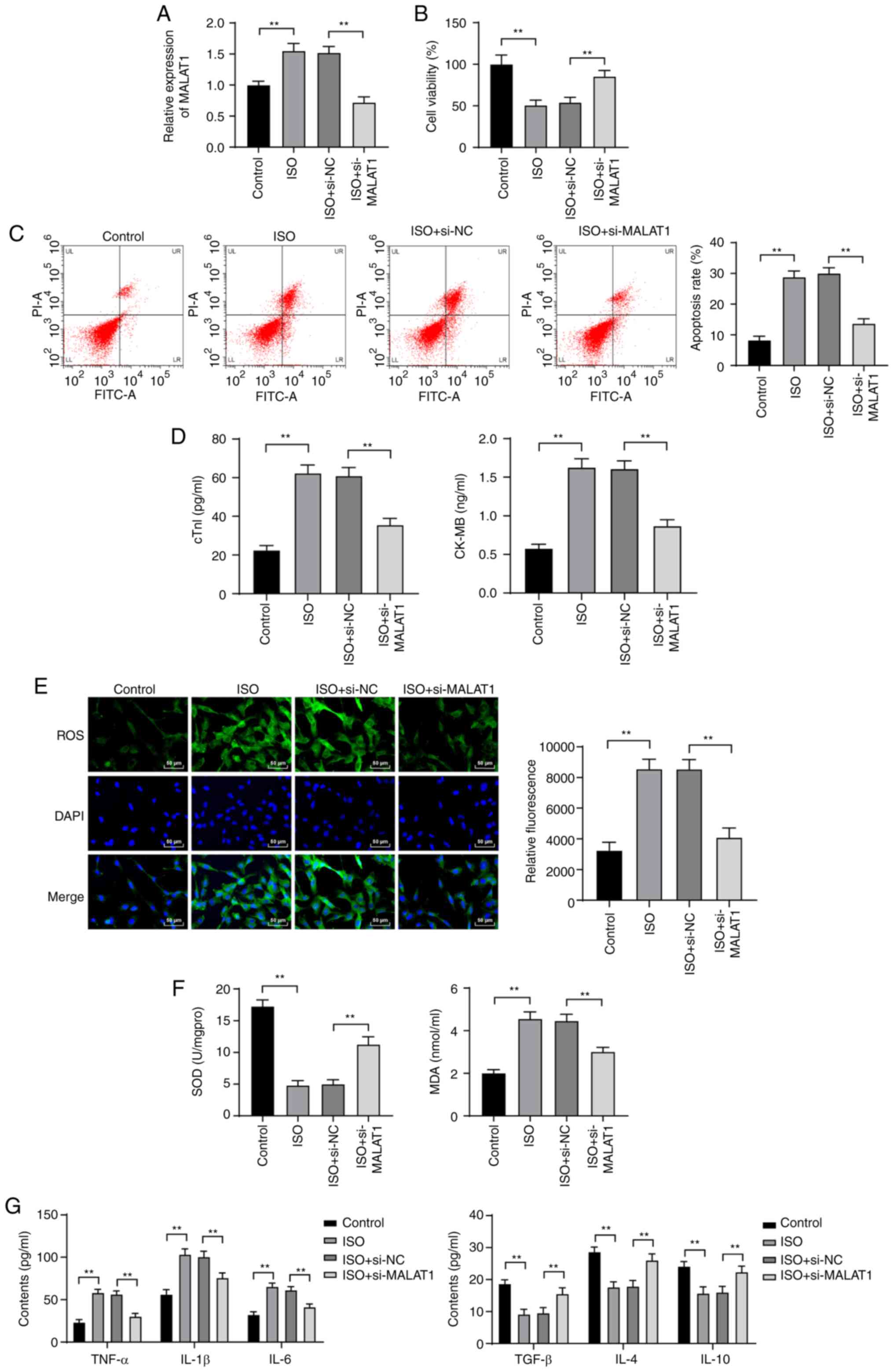
Inhibition of MALAT1 reduced ISO-induced
H9C2 myocardial cell injury
To further investigate the effect of MALAT1 on HF,
si-NC or si-MALAT1 was transfected into ISO-induced H9C2 cells.
MALAT1 expression was distinctly increased in H9C2 cells after ISO
treatment. Compared with that in H9C2 cells treated with si-NC,
MALAT1 expression was significantly decreased in H9C2 cells after
MALAT1 inhibition (P<0.01, Fig.
4A). The viability of ISO-induced H9C2 cells was obviously
decreased. The apoptosis rate cTn-I and CK-MB levels were
significantly increased. However, silencing MALAT1 expression
reversed the above results (all P<0.01, Fig. 4B-D). ISO treatment increased ROS,
MDA, TNF-α, IL-1β and IL-6 levels and reduced SOD, TGF-β, IL-4 and
IL-10 levels; however, MALAT1 knockdown exhibited the opposite
trend (all P<0.01, Fig.
4E-G).
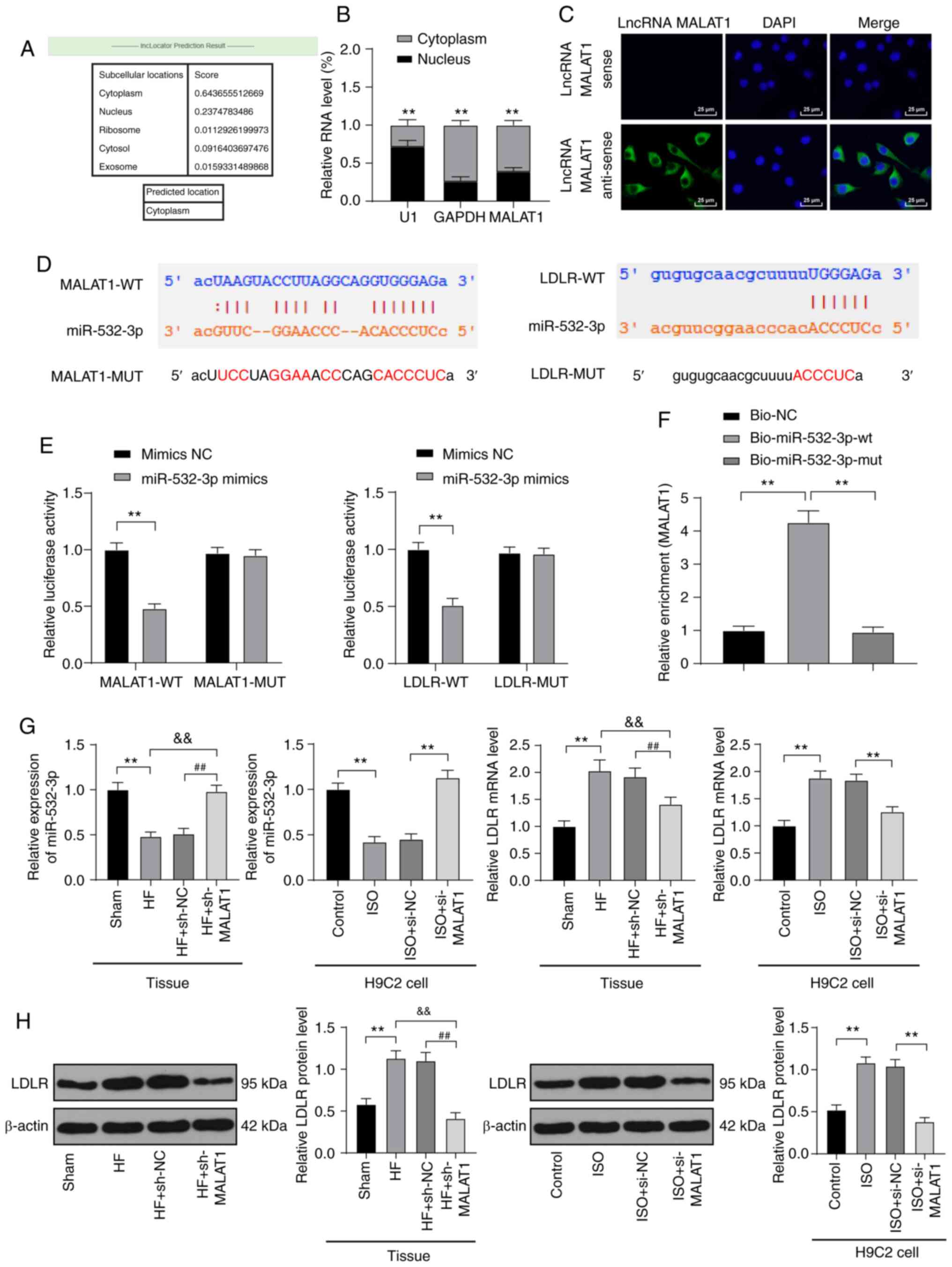
LncRNA MALAT1 competitively bound to
miR-532-3p to upregulate LDLR
In the above in vitro and in vivo
experimental results, we verified that inhibition of MALAT1 reduced
injury in HF rats and ISO-induced H9C2 cells, but the downstream
action mechanism of MALAT1 was still unclear. The mechanism of
action of lncRNA depended on its subcellular location. The online
website (http://www.csbio.sjtu.edu.cn/cgi-bin/lncLocator.py)
(30) predicted that MALAT1 is
located in the cytoplasm of myocardial cells (Fig. 5A). Nuclear and cytoplasmic
fractionation assays (P<0.01, Fig.
5B) and FISH assays (Fig. 5C)
verified that MALAT1 is mainly located in the cytoplasm of H9C2
cells, indicating that MALAT1 plays a regulatory role in myocardial
cells through the mechanism of ceRNA. Subsequently, Starbase
(http://starbase.sysu.edu.cn/index.php) predicted that
MALAT1 and miR-532-3p as well as miR-532-3p and LDLR had targeted
binding sites (Fig. 5D), which
were subsequently confirmed by dual-luciferase reporter gene assays
and RNA pull-down assays (P<0.01, Fig. 5E-F). In myocardial tissues of HF
rats, miR-532-3p expression was obviously decreased, and LDLR mRNA
and protein levels were obviously increased. After MALAT1
inhibition, these results were reversed. The detection results were
consistent with that in H9C2 cells (all P<0.01, Fig. 5G-H).
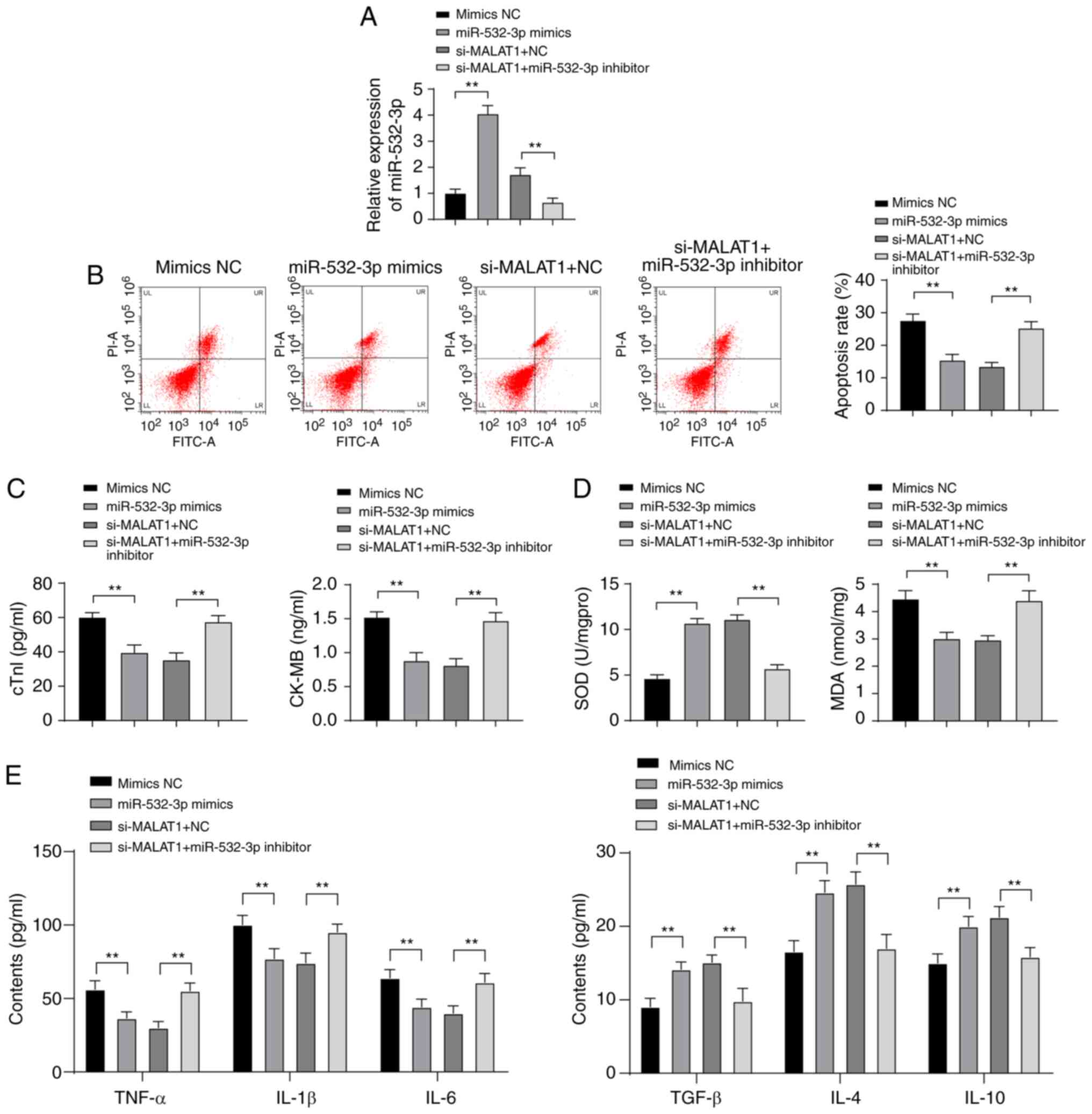
Inhibition of miR-532-3p weakened the
protective effect of downregulated MALAT1 against H9C2 myocardial
cell injury
To further investigate the effect of miR-532-3p on
H9C2 myocardial cell injury and the mechanism by which MALAT1
regulates myocardial injury through miR-532-3p, mimic NC or
miR-532-3p mimic was transfected into ISO-treated H9C2 myocardial
cells, and a functional rescue experiment was established via the
combined treatment of si-MALAT1 + NC and si-MALAT1 + miR-532-3p
inhibitor. After overexpression of miR-532-3p, miR-532-3p
expression was significantly increased in H9C2 myocardial cells.
Compared with cells treated with si-MALAT1 + NC, the expression of
miR-532-3p in H9C2 myocardial cells was clearly decreased in the
si-MALAT1 + miR-532-3p inhibitor group (P<0.01, Fig. 6A). miR-532-3p overexpression
reduced the apoptosis rate and levels of cTn-I, CK-MB, MDA, TNF-α,
IL-1β and IL-6 in H9C2 myocardial cells and increased levels of
SOD, TGF-β, IL-4 and IL-10. Compared with cells treated with
miR-532-3p inhibitor + si-NC, ISO-treated H9C2 myocardial cells in
the miR-532-3p inhibitor + si-MALAT1 group were more damaged (all
P<0.01, Fig. 6B-E).
Overexpression of miR-532-3p reduced injury of ISO-treated H9C2
myocardial cells, and inhibition of miR-532-3p weakened the
protective effect of downregulated MALAT1 against ISO-treated H9C2
myocardial cell injury.
Discussion
HF is a chronic disease, and its rate of incidence
is likely to continue to increase in the next few years (31). Previous findings have shown that
lncRNAs play an important role in regulating mammalian
cardiogenesis (32). A study has
revealed the association of MALAT1 expression profiles in
peripheral blood of coronary artery disease patients with previous
cardiac events (33). In this
study, we demonstrated that lncRNA MALAT1 was highly expressed in
HF patients and exhibited great value for HF diagnosis. MALAT1
targets miR-532-3p to upregulate LDLR in HF, and inhibition of
MALAT1 alleviated myocardial injury, lipid metabolism disorders and
inflammation responses in HF rats.
As evidenced, HF may develop as a result of
myocardial injury (34). MALAT1
is involved in HF (35). Both
cTn-T and CK-MB are biomarkers of myocardial injury, and
upregulation of these biomarkers is indicative of aggravated
myocardial injury (36). It is
well established that LVEF, FS, LVSP, LVEDP and ±dp/dtmax can be
used to evaluate myocardial injury (37,38). Our study revealed that MALAT1 was
highly expressed in HF patients and rats, and suppression of MALAT1
reduced the myocardial injury of HF rats. These conclusions were
confirmed by the significantly reduced serum levels of cTn-I, CK-MB
and LVEDP; increased LVEF, FS, LVSP, and ±dp/dtmax levels, and
decreased TUNEL-positive cells. Consistently, MALAT1 knockdown
helps to improve the cardiac function and decrease myocardial
injury and apoptosis (39).
Lipid metabolism disorders are critical in the
pathogenesis of HF (40).
Assessment of the changes of TC, HDL and LDL may provide a better
understanding of lipid metabolism (41). The current results showed that
inhibition of MALAT1 decreased levels of ET, TC, TG, and LDL-C and
increased the level of HDL-C in HF rats, indicating improvements in
lipid metabolism disorders. Consistent with our study, decreased
ET, TC, TG, and LDL-C levels and increased HDL-C levels signified
reduced blood lipid levels (42).
MALAT1 was highly expressed in hepatocytes of obese mice, and
downregulated MALAT1 significantly inhibited lipid accumulation
(43). Moreover, inflammation is
widely perceived as a dominating risk factor for HF (44). In the current study, downregulated
MALAT1 decreased levels of pro-inflammatory cytokines (TNF-α, IL-1β
and IL-6) and increased levels of anti-inflammatory cytokines
(TGF-β, IL-4 and IL-10) in HF rats, indicating mitigation of
inflammation. Numerous studies have reported results similar to
this study. For example, suppressed MALAT1 helps to relieve
inflammation caused by hypoxia reoxygenation stimulation by
decreasing transcripts and production of pro-inflammatory cytokines
(45). MALAT1 knockdown
alleviates inflammation after brain ischemia (46). Additionally, lipid oxidation is
also strongly associated with the progression of HF (47). Results of the present study showed
that MDA and ROS levels were increased, and SOD levels were reduced
in HF rats. These effects were reversed after MALAT1 inhibition.
Consistently, silencing MALAT1 contributes to downregulation of ROS
and MDA levels and elevated SOD levels in hypoxia-induced cell
injury (48). The results
indicated that MALAT1 inhibition alleviated lipid oxidation in HF
rats.
The above analyses revealed that inhibition of
MALAT1 reduced myocardial injury and relieved lipid metabolism
disorders, inflammation and lipid oxidation in HF rats. These
results were further verified in ISO-induced H9C2 cells in
vitro. However, the downstream regulatory mechanism of MALAT1
was still unclear. MALAT1 functions as a ceRNA and plays an
essential role in cardiac progenitor cell growth via miR-mediated
regulation, providing a potential method for HF therapy (49). miR-532 is enriched in mitochondria
in HF (50). In this study,
targeted binding relationships between MALAT1 and miR-532-3p as
well as miR-532-3p and LDLR were verified. miR-532-3p was decreased
and LDLR was upregulated in HF rats and ISO-induced H9C2 cells, and
inhibition of miR-532-3p weakened the protective effect of
downregulated MALAT1 against H9C2 myocardial cell injury.
Consistent with this study, miR-532 exerts cardioprotective effects
on myocardial infarction and subsequent HF (51). LDLR can serve as a risk predictor
in HF (52). In addition,
miR-532-3p participates in the regulation of atherosclerotic plaque
formation, significantly contributing to atherothrombotic events,
including heart attack, via the involvement of LDLR (20). However, the interactions between
MALAT1 and miR-532-3p have not been elucidated, demonstrating the
innovation of this study. Briefly, the results indicated that
MALAT1 acts as a ceRNA to sponges miR-532-3p to upregulate LDLR,
thereby aggravating myocardial injury in HF.
Altogether, our study revealed that suppression of
MALAT1 relieved myocardial injury, lipid metabolism disorders and
inflammation in HF. These results identified a novel lncRNA-based
therapy for HF, and inhibition of the MALAT1/miR-532-3p/LDLR axis
could potentially be developed as a promising therapeutic approach
for HF treatment. Considering the limitation of animal ethics and
experimental funding, no dose gradient was established, and no
double-labeling method for identification of specific
TUNEL-positive cells was used in this study. In addition,
predisposing mutations associated with MALAT1 gene, vascularity,
fibrosis and hypertrophy were not explored. The above questions are
to be addressed in future research. Although the present study
provides information on the therapeutic value of MALAT1 in the
treatment of HF, the experimental results and clinical applications
need to be further verified.
Supplementary Data
Funding
No funding was received.
Availability of data and materials
All the data generated or analyzed during this study
are included in this published article.
Authors' contributions
PZ and YKW are the guarantors of integrity of the
entire study. PZ and YKW contributed to the study concepts, study
design, and definition of intellectual content. LPZ and JHZ
contributed to the literature research and data acquisition. YKW
contributed to manuscript editing and review. HQW contributed to
the clinical studies. NL contributed to the experimental studies
and data acquisition. LPZ contributed to the data analysis and
statistical analysis. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
This study was approved and supervised by the human
ethics committee of Yantai Yuhuangding Hospital. All subjects
signed the informed consent. All experiments were approved by the
animal ethics committee of Yantai Yuhuangding Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors have no conflicts of interest to
declare.
Acknowledgments
Not applicable.
References
|
1
|
Snipelisky D, Chaudhry SP and Stewart GC:
The many faces of heart failure. Card Electrophysiol Clin.
11:11–20. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tomasoni D, Adamo M, Lombardi CM and Metra
M: Highlights in heart failure. ESC Heart Fail. 6:1105–1127. 2019.
View Article : Google Scholar
|
|
3
|
Bozkurt B and Khalaf S: Heart failure in
women. Methodist Debakey Cardiovasc J. 13:216–223. 2017.
|
|
4
|
Melander S and Miller S: Heart failure:
Overcoming the physiologic dilemma through evidence-based practice.
Nurs Clin North Am. 51:13–27. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ziaeian B and Fonarow GC: Epidemiology and
aetiology of heart failure. Nat Rev Cardiol. 13:368–378. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Martinelli AEM, Maranhao RC, Carvalho PO,
Freitas FR, Silva BMO, Curiati MNC, Kalil Filho R and
Pereira-Barretto AC: Cholesteryl ester transfer protein (CETP), HDL
capacity of receiving cholesterol and status of inflammatory
cytokines in patients with severe heart failure. Lipids Health Dis.
17:2422018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bui AL, Horwich TB and Fonarow GC:
Epidemiology and risk profile of heart failure. Nat Rev Cardiol.
8:30–41. 2011. View Article : Google Scholar :
|
|
8
|
Rosik J, Szostak B, Machaj F and Pawlik A:
Potential targets of gene therapy in the treatment of heart
failure. Expert Opin Ther Targets. 22:811–816. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fan Z, Gao S, Chen Y, Xu B, Yu C, Yue M
and Tan X: Integrative analysis of competing endogenous RNA
networks reveals the functional lncRNAs in heart failure. J Cell
Mol Med. 22:4818–4829. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Turton N, Swan R, Mahenthiralingam T,
Pitts D and Dykes IM: The functions of long non-coding RNA during
embryonic cardiovascular development and its potential for
diagnosis and treatment of congenital heart disease. J Cardiovasc
Dev Dis. 6:212019. View Article : Google Scholar :
|
|
11
|
Zhao Y, Wu J, Liangpunsakul S and Wang L:
Long non-coding RNA in liver metabolism and disease: Current
status. Liver Res. 1:163–167. 2017. View Article : Google Scholar
|
|
12
|
Yan Y, Song D, Wu J and Wang J: Long
non-coding RNAs link oxidized low-density lipoprotein with the
inflammatory response of macrophages in atherogenesis. Front
Immunol. 11:242020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sirtori CR, Ruscica M, Calabresi L, Chiesa
G, Giovannoni R and Badimon JJ: HDL therapy today: From
atherosclerosis, to stent compatibility to heart failure. Ann Med.
51:345–359. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu Q and Yi X: Down-regulation of long
noncoding RNA MALAT1 protects hippocampal neurons against excessive
autophagy and apoptosis via the PI3K/Akt signaling pathway in rats
with epilepsy. J Mol Neurosci. 65:234–245. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu L, Tan L, Yao J and Yang L: Long
non-coding RNA MALAT1 regulates cholesterol accumulation in
ox-LDL-induced macrophages via the microRNA-17-5p/ABCA1 axis. Mol
Med Rep. 21:1761–1770. 2020.PubMed/NCBI
|
|
16
|
Wang L, Qi Y, Wang Y, Tang H, Li Z, Wang
Y, Tang S and Zhu H: LncRNA MALAT1 suppression protects endothelium
against oxLDL-induced inflammation via inhibiting expression of
MiR-181b target gene TOX. Oxid Med Cell Longev. 2019:82458102019.
View Article : Google Scholar
|
|
17
|
Huang Y: The novel regulatory role of
lncRNA-miRNA-mRNA axis in cardiovascular diseases. J Cell Mol Med.
22:5768–5775. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shah P, Bristow MR and Port JD: MicroRNAs
in heart failure, cardiac transplantation, and myocardial recovery:
Biomarkers with therapeutic potential. Curr Heart Fail Rep.
14:454–464. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bauersachs R, Zeymer U, Briere JB, Marre
C, Bowrin K and Huelsebeck M: Burden of coronary artery disease and
peripheral artery disease: A literature review. Cardiovasc Ther.
2019:82950542019. View Article : Google Scholar
|
|
20
|
Huang R, Cao Y, Li H, Hu Z, Zhang H, Zhang
L, Su W, Xu Y, Liang L, Melgiri ND, et al: miR-532-3p-CSF2RA axis
as a key regulator of vulnerable atherosclerotic plaque formation.
Can J Cardiol. 36:1782–1794. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Arola-Arnal A and Blade C:
Proanthocyanidins modulate microRNA expression in human HepG2
cells. PLoS One. 6:e259822011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gao K, Zhang J, Gao P, Wang Q, Liu Y, Liu
J, Zhang Y, Li Y, Chang H, Ren P, et al: Qishen granules exerts
cardioprotective effects on rats with heart failure via regulating
fatty acid and glucose metabolism. Chin Med. 15:212020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zatroch KK, Knight CG, Reimer JN and Pang
DS: Refinement of intraperitoneal injection of sodium pentobarbital
for euthanasia in laboratory rats (Rattus norvegicus). BMC Vet Res.
13:602017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fan C, Tang X, Ye M, Zhu G, Dai Y, Yao Z
and Yao X: Qi-Li-Qiang-Xin alleviates isoproterenol-induced
myocardial injury by inhibiting excessive autophagy via activating
AKT/mTOR pathway. Front Pharmacol. 10:13292019. View Article : Google Scholar :
|
|
25
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao W, Geng D, Li S, Chen Z and Sun M:
LncRNA HOTAIR influences cell growth, migration, invasion, and
apoptosis via the miR-20a-5p/HMGA2 axis in breast cancer. Cancer
Med. 7:842–855. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou YX, Wang C, Mao LW, Wang YL, Xia LQ,
Zhao W, Shen J and Chen J: Long noncoding RNA HOTAIR mediates the
estrogen-induced metastasis of endometrial cancer cells via the
miR-646/NPM1 axis. Am J Physiol Cell Physiol. 314:C690–C701. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Choi SS, Kim ES, Koh M, Lee SJ, Lim D,
Yang YR, Jang HJ, Seo KA, Min SH, Lee IH, et al: A novel
non-agonist peroxisome proliferator-activated receptor γ (PPARγ)
ligand UHC1 blocks PPARγ phosphorylation by cyclin-dependent kinase
5 (CDK5) and improves insulin sensitivity. J Biol Chem.
289:26618–26629. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Madej-Pilarczyk A, Niezgoda A, Janus M,
Wojnicz R, Marchel M, Fidziańska A, Grajek S and
Hausmanowa-Petrusewicz I: Limb-girdle muscular dystrophy with
severe heart failure overlapping with lipodystrophy in a patient
with LMNA mutation p.Ser334del. J Appl Genet. 58:87–91. 2017.
View Article : Google Scholar :
|
|
30
|
Cao Z, Pan X, Yang Y, Huang Y and Shen HB:
The lncLocator: A subcellular localization predictor for long
non-coding RNAs based on a stacked ensemble classifier.
Bioinformatics. 34:2185–2194. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rogers C and Bush N: Heart failure:
Pathophysiology, diagnosis, medical treatment guidelines, and
nursing management. Nurs Clin North Am. 50:787–799. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Scheuermann JC and Boyer LA: Getting to
the heart of the matter: Long non-coding RNAs in cardiac
development and disease. EMBO J. 32:1805–1816. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Toraih EA, El-Wazir A, Alghamdi SA,
Alhazmi AS, El-Wazir M, Abdel-Daim MM and Fawzy MS: Association of
long non-coding RNA MIAT and MALAT1 expression profiles in
peripheral blood of coronary artery disease patients with previous
cardiac events. Genet Mol Biol. 42:509–518. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hartupee J and Mann DL: Neurohormonal
activation in heart failure with reduced ejection fraction. Nat Rev
Cardiol. 14:30–38. 2017. View Article : Google Scholar :
|
|
35
|
Guo X, Wu X, Han Y, Tian E and Cheng J:
LncRNA MALAT1 protects cardiomyocytes from isoproterenol-induced
apoptosis through sponging miR-558 to enhance ULK1-mediated
protective autophagy. J Cell Physiol. 234:10842–10854. 2019.
View Article : Google Scholar
|
|
36
|
Sadoh WE, Eregie CO, Nwaneri DU and Sadoh
AE: The diagnostic value of both troponin T and creatinine kinase
isoenzyme (CK-MB) in detecting combined renal and myocardial
injuries in asphyxiated infants. PLoS One. 9:e913382014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang X, Wang J, Tu T, Iyan Z, Mungun D,
Yang Z and Guo Y: Remote ischemic postconditioning protects against
myocardial ischemia-reperfusion injury by inhibition of the
RAGE-HMGB1 pathway. Biomed Res Int. 2018:45656302018.PubMed/NCBI
|
|
38
|
Yan B, Liu S, Li X, Zhong Y, Tong F and
Yang S: Preconditioning with endoplasmic reticulum stress
alleviated heart ischemia/reperfusion injury via modulating
IRE1/ATF6/RACK1/PERK and PGC-1α in diabetes mellitus. Biomed
Pharmacother. 118:1094072019. View Article : Google Scholar
|
|
39
|
Fan YZ, Huang H, Wang S, Tan GJ and Zhang
QZ: Effect of lncRNA MALAT1 on rats with myocardial infarction
through regulating ERK/MAPK signaling pathway. Eur Rev Med
Pharmacol Sci. 23:9041–9049. 2019.PubMed/NCBI
|
|
40
|
Abdalla S, Fu X, Elzahwy SS, Klaetschke K,
Streichert T and Quitterer U: Up-regulation of the cardiac lipid
metabolism at the onset of heart failure. Cardiovasc Hematol Agents
Med Chem. 9:190–206. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kurpińska AK, Jarosz A, Ożgo M and
Skrzypczak WF: Changes in lipid metabolism during last month of
pregnancy and first two months of lactation in primiparous
cows-analysis of apolipoprotein expression pattern and changes in
concentration of total cholesterol, HDL, LDL, triglycerides. Pol J
Vet Sci. 18:291–298. 2015. View Article : Google Scholar
|
|
42
|
Li J, Lei HT, Cao L, Mi YN, Li S and Cao
YX: Crocin alleviates coronary atherosclerosis via inhibiting lipid
synthesis and inducing M2 macrophage polarization. Int
Immunopharmacol. 55:120–127. 2018. View Article : Google Scholar
|
|
43
|
Yan C, Chen J and Chen N: Long noncoding
RNA MALAT1 promotes hepatic steatosis and insulin resistance by
increasing nuclear SREBP-1c protein stability. Sci Rep.
6:226402016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Shirazi LF, Bissett J, Romeo F and Mehta
JL: Role of inflammation in heart failure. Curr Atheroscler Rep.
19:272017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang Y, Zhang H, Zhang Z, Li S, Jiang W,
Li X and Lv J: LncRNA MALAT1 cessation antagonizes
hypoxia/reoxygenation injury in hepatocytes by inhibiting apoptosis
and inflammation via the HMGB1-TLR4 axis. Mol Immunol. 112:22–29.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cao DW, Liu MM, Duan R, Tao YF, Zhou JS,
Fang WR, Zhu JR, Niu L and Sun JG: The lncRNA Malat1 functions as a
ceRNA to contribute to berberine-mediated inhibition of HMGB1 by
sponging miR-181c-5p in poststroke inflammation. Acta Pharmacol
Sin. 41:22–33. 2020. View Article : Google Scholar :
|
|
47
|
Fragasso G: Deranged cardiac metabolism
and the pathogenesis of heart failure. Card Fail Rev. 2:8–13. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yang L, Xu F, Zhang M, Shang XY, Xie X, Fu
T, Li JP and Li HL: Role of LncRNA MALAT-1 in hypoxia-induced PC12
cell injury via regulating p38MAPK signaling pathway. Neurosci
Lett. 670:41–47. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Li L, Wang Q, Yuan Z, Chen A, Liu Z, Wang
Z and Li H: LncRNA-MALAT1 promotes CPC proliferation and migration
in hypoxia by up-regulation of JMJD6 via sponging miR-125. Biochem
Biophys Res Commun. 499:711–718. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang X, Song C, Zhou X, Han X, Li J, Wang
Z, Shang H, Liu Y and Cao H: Mitochondria associated MicroRNA
expression profiling of heart failure. Biomed Res Int.
2017:40425092017.PubMed/NCBI
|
|
51
|
Bayoumi AS, Teoh JP, Aonuma T, Yuan Z,
Ruan X, Tang Y, Su H, Weintraub NL and Kim IM: MicroRNA-532
protects the heart in acute myocardial infarction, and represses
prss23, a positive regulator of endothelial-to-mesenchymal
transition. Cardiovasc Res. 113:1603–1614. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Bayes-Genis A, Núñez J, Zannad F, Ferreira
JP, Anker SD, Cleland JG, Dickstein K, Filippatos G, Lang CC, Ng
LL, et al: The PCSK9-LDL receptor axis and outcomes in heart
failure: BIOSTAT-CHF subanalysis. J Am Coll Cardiol. 70:2128–2136.
2017. View Article : Google Scholar : PubMed/NCBI
|