Introduction
Deep vein thrombosis (DVT) is a form of a blood
clotting in the deep veins, which can occur in any part of the
body, but typically occurs in the legs (1). DVT, as a common peripheral vascular
disease, is often accompanied by endothelial injury and may develop
into pulmonary embolism, which is related to mortality (2,3).
Additionally, >20% of affected patients may develop
post-thrombotic syndrome, which can seriously affect the quality of
life of patients in the later stages of DVT (4). The incidence of DVT is 1 in 1,000
adults and increases with age (5,6);
there are 10 million diagnosed cases in China (5,7).
DVT is a complex event and the disease pathogenesis remains to be
elucidated. Moreover, to date, thrombus-related damage to the vein
wall cannot be effectively inhibited by pharmacological methods
(8). Evidence has demonstrated
that inflammatory markers, including the inflammatory cytokines,
tumor necrosis factor (TNF)-α, interleukin (IL)-6 and IL-8
(9) contributed to the formation
of DVT. However, the underlying molecular mechanisms are not yet
fully understood.
Coagulation factor XII (FXII), a type of serine
protease zymogen, is a key coagulation factor and the initiating
factor of the intrinsic coagulation pathway. It has been reported
that FXII may activate coagulation pathway related-proteins via the
factors XI and IX (10,11). FXII has been reported to be
involved in various physiological processes, including coagulation,
fibrinolysis and angiogenesis. It has also been demonstrated that
FXII protein may play an important role in regulating thrombosis
(12). Furthermore,
FXII-deficient animals may effectively prevent thrombus formation
without impaired hemostasis (13-15). Additionally, FXII protein is
closely associated with inflammatory disorders, including
rheumatoid arthritis and colitis. FXII protein may promote the
inflammatory response by upregulating the protein expression of
TNF-α, IL-6 and IL-8 in lungs affected by acute respiratory
distress syndrome (16). The
phosphoinositide 3-kinase (PI3K)/AKT signaling pathway is an
important signaling pathway in the regulation of the inflammatory
response. Of note, FXII protein may induce the activation of AKT
signaling (17). These data
indicate that FXII protein may play an important role in the
formation of DVT. However, to the best of our knowledge, the
effects of the FXII-mediated activation of the PI3K/AKT signaling
pathway on the formation of DVT have not been reported to date.
The present study thus attempted to investigate the
mechanisms of the FXII-mediated PI3K/AKT signaling pathway in the
formation of DVT. A mouse model of DVT was used to assess whether
FXII protein and the PI3K/AKT signaling pathway are involved in the
regulation of the formation of DVT, and to further reveal the
molecular mechanisms of the FXII-mediated activation of PI3K/AKT
signaling in the formation of DVT. These findings may provide
theoretical evidence and a potential approach with which to
attenuate the formation of DVT.
Materials and methods
Drugs and antibodies
LY294002 (a PI3K inhibitor) was purchased from
Sigma-Aldrich; Merck KGaA. Anti-PI3K (#AF5121) and
anti-phosphorylated (p)-PI3K (#AF3241) anti-bodies were purchased
from Affinity Biosciences. Anti-factor XII (#12551-1-AP) was
purchased from ProteinTech Group, Inc. Anti-AKT (#4685), anti-p-AKT
(#4060) and anti-GAPDH (#2118) antibodies and HRP-conjugated
secondary antibodies were purchased from Cell Signaling Technology,
Inc.
Animal experiments
A total of 66 7-week-old male C57BL/6 mice (weighing
20-21 g) were purchased from the Experimental Animal Center of
Xi'an Jiaotong University. All mice were randomly assigned to the
following groups: The control group (A), the sham-operated group
(B) and the model group (C), with 6 mice in each group. All mice
were kept in a specific pathogen-free facility for 1 week of
adaptive feeding. The mouse model of DVT was established as
previously described (18). The
mice were anesthetized using an animal isoflurane anesthesia
machine (R520, RWD Life Science, anesthesia was induced by 4%
isoflurane and maintained by 1.5% isoflurane) and fixed on an
operating table. The abdomen of the mice was opened in the middle
of the abdominal white line, and the small bowel and other organs
in the abdominal cavity were exteriorized onto a piece of sterile
gauze pre-soaked with normal saline. After exposing the inferior
vena cava (IVC) and its branches, the IVC was carefully separated
and all the visible branches of the IVC were ligated. Once the
proximal and distal IVC were clamped using a vascular clamp for 30
sec, the IVC was passed through the rear with a 6-0 suture. The IVC
was ligated using a 4-0 suture, and the 6-0 suture was then rapidly
drawn out. After confirming that the respiratory circulation of the
mice was stable, the abdominal incision of the mice was closed
layer by layer and the mice were fed normally. The mice in the
model groups also were randomly assigned to 4 subgroups as follows:
C, model group; D, model group transfected with adenovirus vector
pAd-pG2.1; E, model group transfected with adenovirus vector
pAd-pG2.1-small interfering RNA (siRNA) FXII; and F, model group
treated with LY294002 (50 mg/kg): The mice were injected with
treated with LY294002 (50 mg/kg) through the tail vein (twice, 0.5
h prior to the establishment of the DVT model and 24 h after the
establishment of the DVT model). In the present study, 18 mice were
used for the experiments shown in Figs. 1-3 (3 groups, 6 mice in each group), 30
mice were used for the experiments shown in Figs. 4-6 (5 groups, 6 mice in each group) and
18 mice were used for the experiments shown in Figs. 7 and 8 (3 groups, 6 mice in each group). The
mice were intraperitoneally injected with 1 ml 3% pentobarbital
sodium (30 mg/kg) and the femoral vein tissue of mice was excised
at 48 h following the establishment of the DVT model. At the end of
the experiment (48 h following the establishment of the DVT model),
the mice underwent euthanasia via cervical dislocation to confirm
death. The experimental procedures and protocols were reviewed and
approved by the Animal Investigation Ethics Committee of the First
Affiliated Hospital of Xi'an Jiaotong University and were performed
in accordance with the Guidelines for the Care and Use of
Laboratory Animals from the National Institutes of Health,
China.
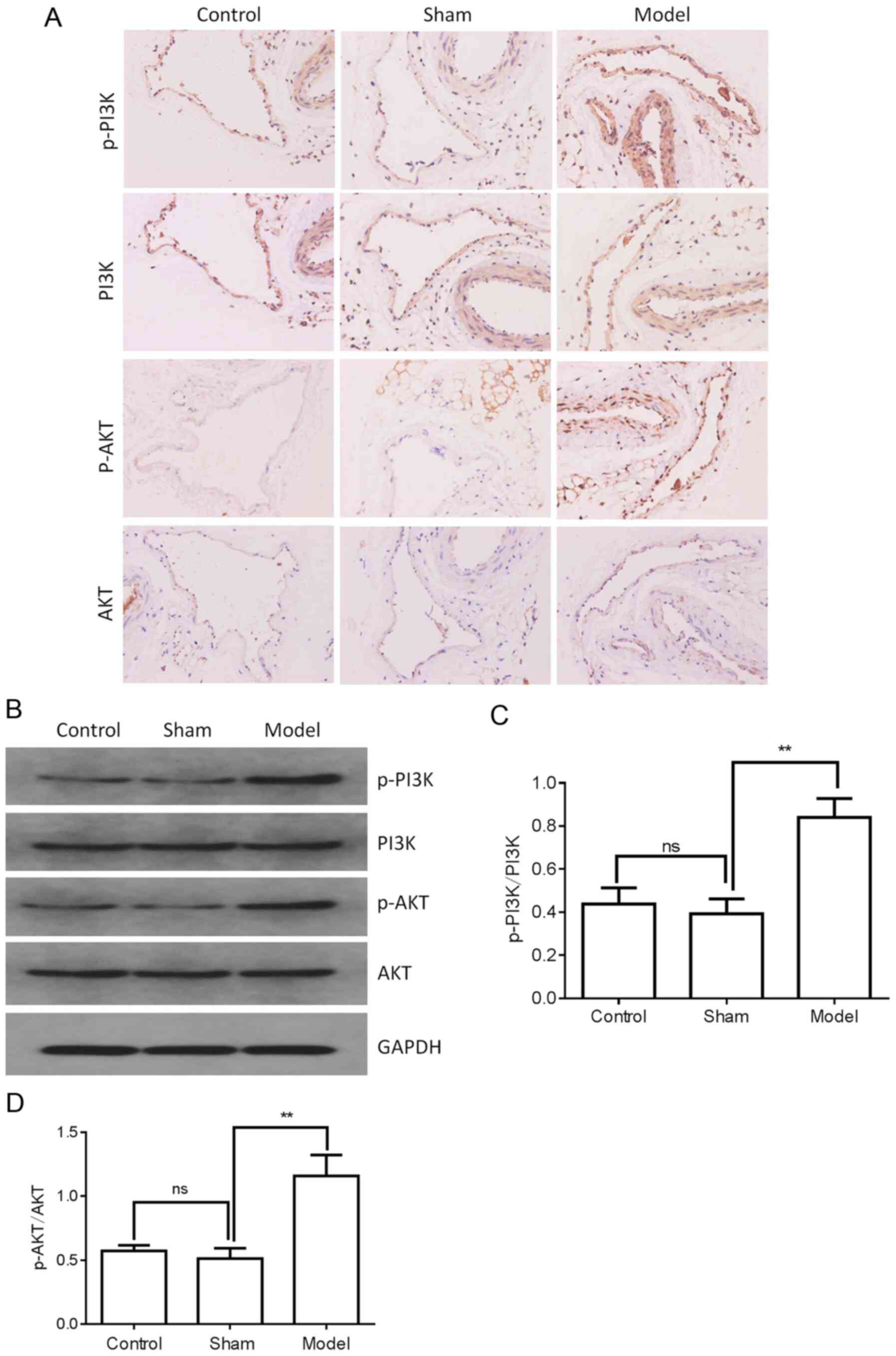
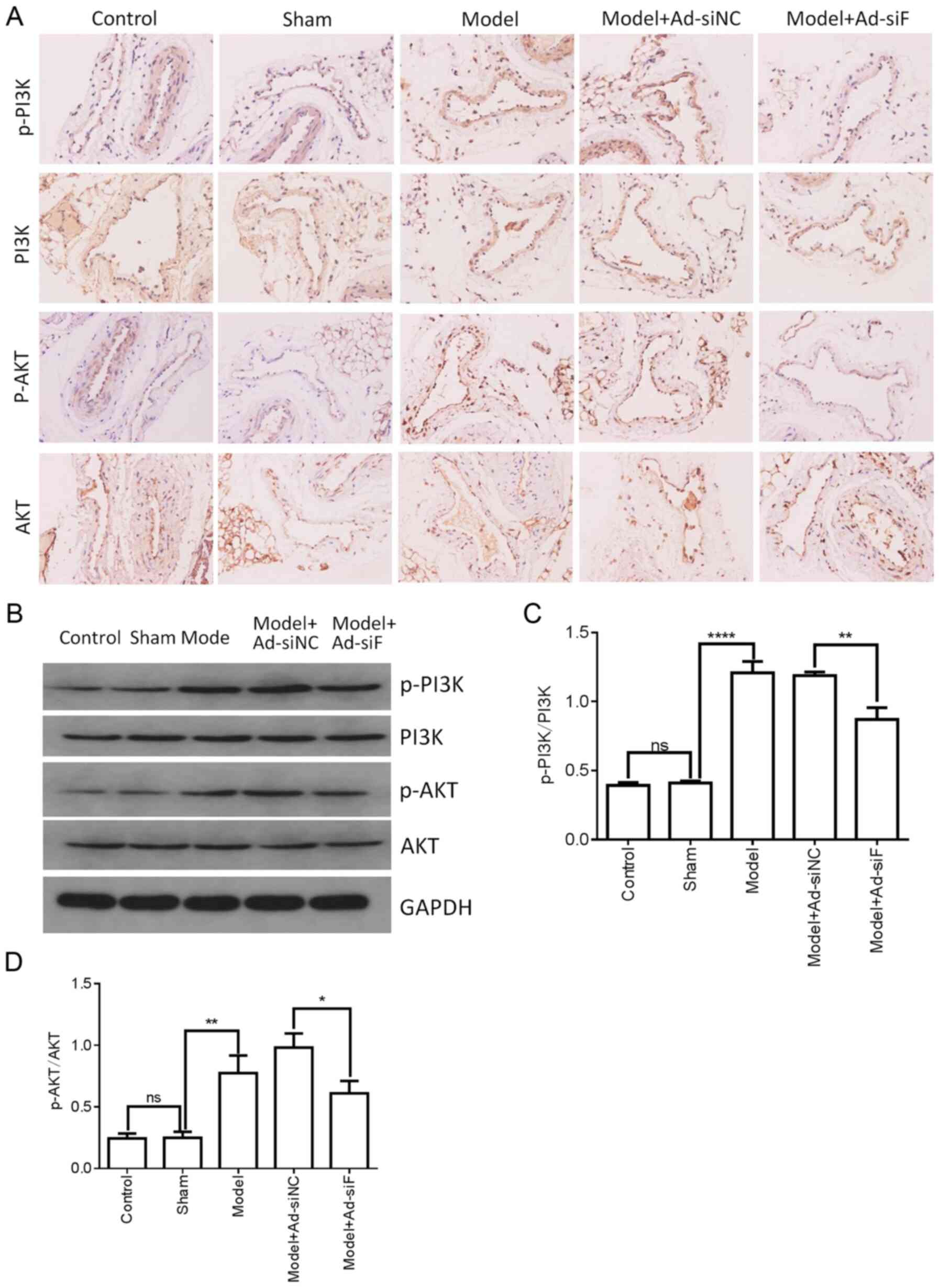 | Figure 6Effects of FXII protein on the
activation of PI3K/AKT signaling in the femoral vein tissue of DVT
mice. FXII protein was knocked down by transfection with
pAd-pG2.1-siRNA FXII (11×108 pfu/mice). (A and B) The
levels of PI3K, p-PI3K, AKT and p-AKT in the femoral vein tissue of
different treatment groups (n=6 mice in each group) were determined
by western blotting and immunohistochemical staining. (C and D)
Western blot analysis quantification of PI3K, p-PI3K, AKT and p-AKT
levels. *P<0.05, **P<0.01 and
****P<0.0001; ns, not significant. FXII, coagulation
factor XII; DVT, deep vein thrombosis; siRNA, small interfering
RNA; pfu, plaque-forming units; p, phosphorylated. |
Humane endpoints
The humane endpoints of the animal experiments were
performed in accordance with the Association of Primate
Veterinarians' Humane Endpoint Guidelines for Nonhuman Primates in
Biomedical Research, 2020 and Institutional Animal Care and Use
Committee Guidebook (http://grants.nih.gov/grants/olaw/GuideBook.pdf).
Humane endpoints were at 48 h after establishment of DVT model.
However, humane endpoints involved immediate intervention under the
following conditions: i) The body weight of the mice decreased
15-20% following the establishment of the DVT model; ii) the mice
lost complete interest in eating for 24 h following the
establishment of the DVT model; iii) the mice could not drink and
eat by themselves; iv) the mice could not stand for 24 h following
the establishment of the DVT model; v) abnormal central nervous
responses (such as convulsion and paralysis) and inability to
control pain in mice following the establishment of the DVT model;
vi) other abnormal phenomena in mice, such as persistent
self-mutilation, persistent hypothermia following the establishment
of the DVT model.
Histological analysis
The femoral vein injury was evaluated by hematoxylin
and eosin (H&E) staining. To assess the histological
characterization of the femoral vein tissue, the femoral vein
tissue was excised from the mice and fixed with 4% paraformaldehyde
at 48 h after surgery. The femoral vein tissue was dehydrated in
graded concentrations of ethanol and then embedded in paraffin. The
femoral vein tissue was cut into sections at a thickness of
5-µm. Following dewaxing and rehydration, the slices were
stained with H&E by using a Hematoxylin and Eosin Staining kit
(Beyotime Institute of Biotechnology) according to the
manufacturer's instructions. The final stained sections were
photographed using an BX-50 Olympus light microscope (Olympus
Corporation) equipped with a DP22 camera at ×400 magnification.
Determination of inflammatory cytokine
levels
The femoral vein tissue of the mice was homogenized,
and supernatant samples were collected and stored at −80°C prior to
use. The mice were anesthetized using an animal isoflurane
anesthesia machine (R520, RWD Life Science, anesthesia was induced
by 4% isoflurane and maintained by 1.5% isoflurane). A total of 200
µl each of mouse plasma was also obtained by mouse
retro-orbital sinus blood collection and stored at −80°C prior to
use. The concentrations of IL-8, and the cytokine levels of IL-6
and TNF-α in femoral vein tissue or plasma were examined using
corresponding specific ELISA kits (Elabscience, Inc.) according to
the manufacturer's instructions. Each sample was tested in
triplicate. The absorbance at a wavelength of 450 nm was examined
using a Multiskan MK3 microplate reader (Thermo Fisher Scientific,
Inc.).
Determination of malondialdehyde (MDA)
and superoxide dismutase (SOD) levels
The femoral vein tissue of mice was homogenized on
ice in sterile saline by using TissueLyser-24 tissue homogenizer
(Shanghai Jingxin Industrial Development Co., Ltd.), and
supernatant samples were collected. The concentration of MDA and
SOD in the femoral vein tissue was examined using corresponding
specific assay kits (Nanjing Jiancheng Bioengineering Institute)
according to the manufacturer's instructions. The MDA and SOD
samples were tested in triplicate. The absorbance at wavelengths of
532 nm (MDA) and 450 nm (SOD) was examined using a Multiskan MK3
microplate reader (Thermo Fisher Scientific, Inc.).
TUNEL assay and immunohistochemical
staining
The femoral vein tissue was dehydrated in graded
concentrations of ethanol and embedded in paraffin. The femoral
vein tissue was cut into sections with a thickness of 4 µm.
Following dewaxing and rehydration, cell apoptosis of the femoral
vein tissue was examined using a TUNEL Apoptosis Detection kit
(Alexa Fluor 640; Shanghai Yeasen Biotechnology Co., Ltd.)
according to the manufacturer's instructions. The femoral vein
tissue was stained with 2 µg/ml DAPI (Beyotime Institute of
Biotechnology) for 5 min at 25°C in a dark room. Sections of the
femoral vein tissue were also incubated with anti-PI3K (1:100),
anti-p-PI3K (1:100), anti-AKT (1:100), anti-p-AKT (1:100) and
anti-factor XII (1:100) antibodies overnight at 4°C, followed by
incubation with HRP-conjugated secondary antibodies (1:50) for 45
min at 37°C. After the final wash, the sections were observed using
a Dako REAL EnVision Detection system.
Construction of adenoviral vectors
encoding siRNA FXII
Antisense FXII cDNA was constructed according to the
FXII sequence (GCA AGA GTC TGT CTT CGA T) was constructed.
Antisense FXII cDNA was inserted into the plasmids pG2.1 (Biofavor
Biotechnology Services Co. Ltd.) using 2 cDNA primers (5′-CAC CGC
AAG AGT CTG TCT TCG ATT TCA AGA CGA TCG AAG ACA GAC TCT TGC TTT TTT
G-3′ and 5′-AGC TCAAAAAAG CAAGAGT CTGTCTT CGATC GTC TTGAAA TCG AAG
ACA GAC TCT TGC-3′). Briefly, antisense FXII cDNA was obtained from
2 primers annealed at 94°C and cooled to room temperature. pG2.1
plasmid was digested with BsaI and were ligated with
antisense FXII cDNA at 22°C in a water bath overnight. The entire
pG2.1-FXII siRNA was sub-cloned into the adenoviral vector,
pAd/PL-DEST (Invitrogen; Thermo Fisher Scientific, Inc.) by Gateway
LR Clonase™ II (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Recombinant
adenovirus (pAd-pG2.1-siRNA FXII) was obtained from 293 cells
(Procell Co. Ltd.). Following 1 week of adaptive feeding, the mice
in the model groups were injected with 1×108 plaque
forming units per mouse.
Western blot analysis
The femoral vein tissues were homogenized using RIPA
lysis buffer (Beyotime Institute of Biotechnology) with 1 mM PMSF
on ice for 30 min. Following centrifugation at 12,000 × g for 10
min at 4°C, the supernatants were collected. The protein
concentration was determined using a BCA Protein Test kit (Beyotime
Institute of Biotechnology). Total protein (20 µg/well) were
separated via 10% SDS-PAGE and transferred onto PVDF membranes (EMD
Millipore). Membranes were blocked with 5% non-fat milk at 25°C for
2 h and incubated with primary antibodies against PI3K (1:1,000),
p-PI3K (1:1,000), AKT (1:1,000), p-AKT (1:1,000) and GAPDH
(1:2,000) at 4°C overnight. Subsequently, membranes were incubated
with HRP-conjugated goat anti-rabbit secondary antibody (1:2,000)
at 25°C for 2 h. Protein bands were visualized using enhanced
chemiluminescence detection reagent (Thermo Fisher Scientific,
Inc.). Densitometric analysis was performed using ImageJ 1.38X
software (National Institutes of Health).
Statistical analysis
GraphPad Prism 7 software (GraphPad Software, Inc.)
was used to perform statistical analysis. All experiments were
performed at least in triplicate and data are expressed as the mean
± SD. Different variables between the 2 groups were analyzed using
a Student's t-test and different variables between three or more
groups were analyzed using one-way ANOVA followed by the Tukey's
post-hoc tests. P<0.05 was considered to indicate a
statistically significant difference.
Results
Upregulation of FXII protein in the
femoral vein tissue of mice with DVT
To assess FXII protein expression in the femoral
vein tissue of mice, a mouse model of DVT was established and the
histological characterization of the femoral vein tissue was
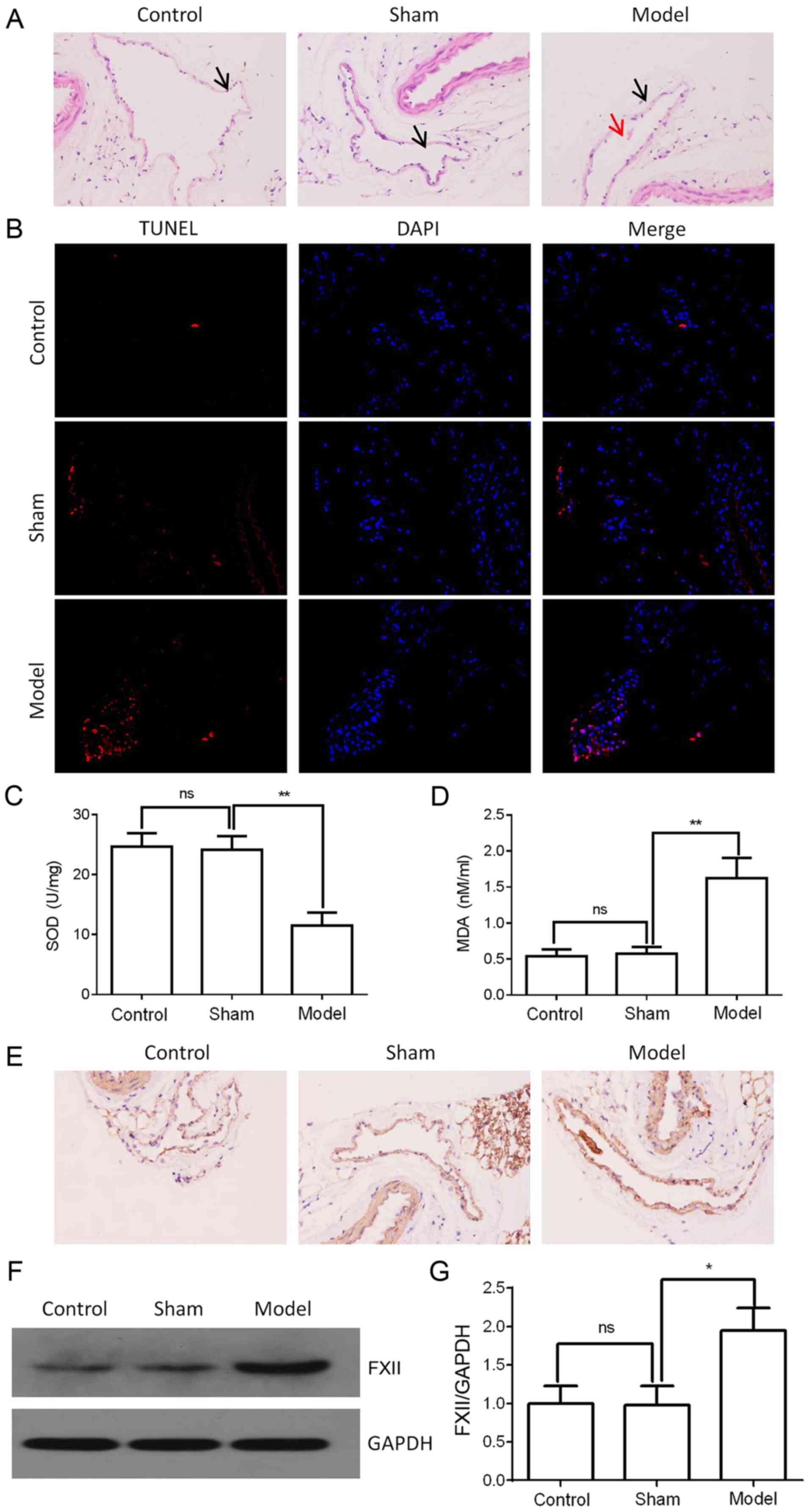
performed by H&E staining. As shown in Fig. 1A, thrombosis was significantly
induced in the femoral vein tissue of the model group compared with
the control or sham groups. Femoral vein tissue damage was examined
by TUNEL assay, As shown in Fig.
1B, the cell apoptotic rate in the femoral vein tissue was
significantly increased in the DVT model. SOD, a member of
metalloproteinases, is a major free radical scavenger which
significantly reduces free radical damage to the body and protects
tissues from injury. MDA is a type of unsaturated fatty acid which
can damage the biofilm and reflect the extent of lipid peroxidation
and tissue injury (19-21). Thus, the SOD and MDA content
plays an important role in vein thrombosis. Hence, in the present
study, the SOD and MDA concentrations in the femoral vein tissue
were measured using ELISA. The results revealed that the SOD
concentrations were decreased, while the MDA concentrations were
increased in the femoral vein tissue of the model group (Fig. 1C and D). Additionally, FXII
protein expression was found to be upregulated in the femoral vein
tissue of the model group, as shown by western blot analysis and
immunohistochemical staining (Fig.
1E-G). These data thus indicated that FXII protein may be
associated with the formation of DVT.
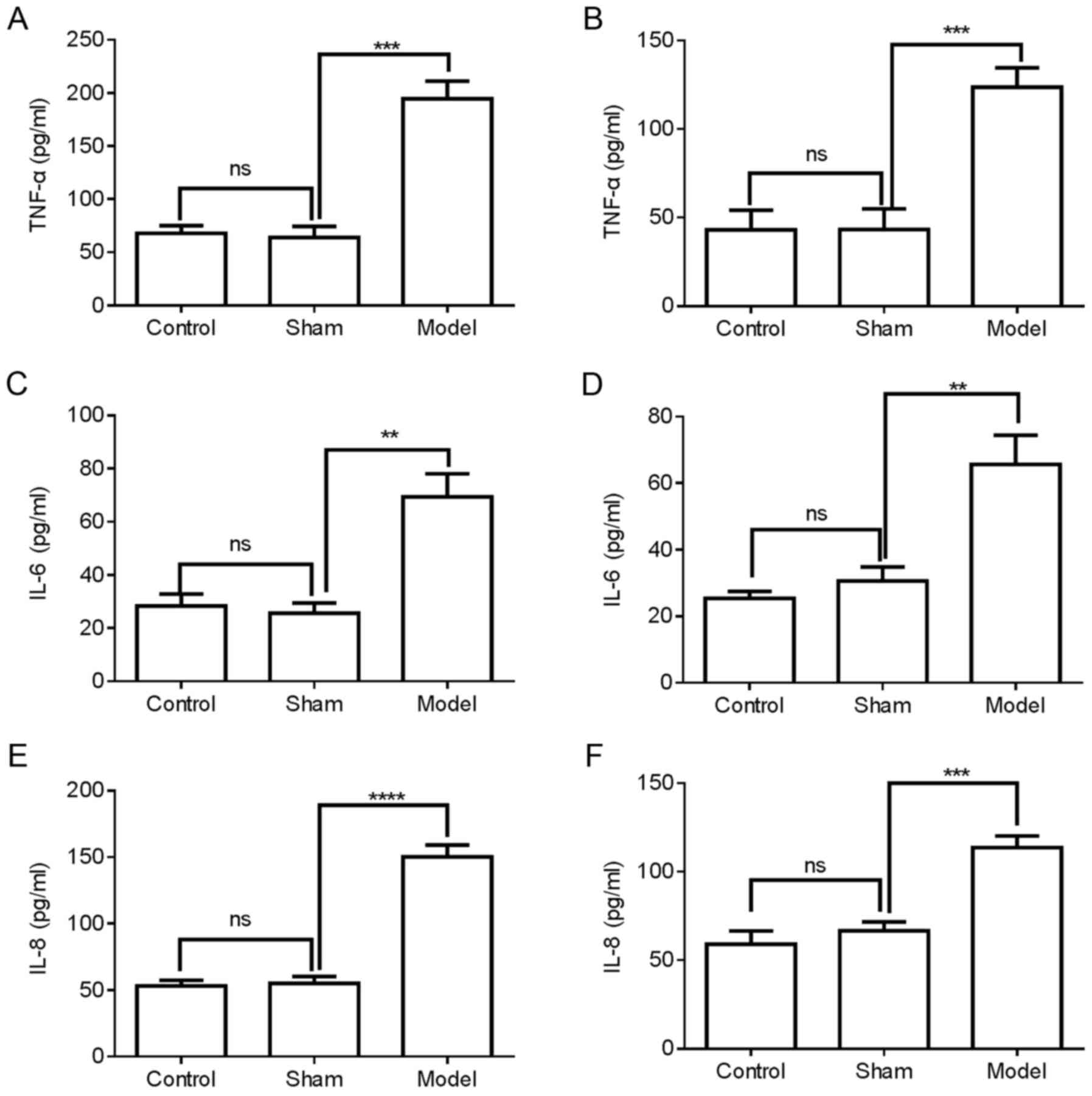
Upregulation of inflammatory cytokines in
the femoral vein tissue and plasma of mice with DVT
The expression of inflammatory cytokines in the
femoral vein tissue of mice was then investigated. The protein
expression levels of TNF-α, IL-6 and IL-8 in the femoral vein
tissue and plasma of mice were examined using ELISA. Compared with
the control or sham groups, the protein expression levels of TNF-α,
IL-6 and IL-8 were significantly increased in the femoral vein
tissue of the model mice (Fig. 2A, C
and E) and plasma of the model mice (Fig. 2B, D and F). These data suggested
that inflammatory cytokines TNF-α, IL-6 and IL-8 might be involved
in DVT formation.
PI3K/AKT signaling is activated in the
femoral vein tissue of mice with DVT
The activation of the PI3K/AKT signaling pathway was
then determined. The levels of PI3K, p-PI3K, AKT and p-AKT were
examined by western blot analysis and immunohistochemical staining.
As shown in Fig. 3A and B, the
protein expression of PI3K and AKT was not markedly altered in the
femoral vein tissue of the model group compared with the control or
sham groups. However, the p-PI3K and p-AKT levels were
significantly upregulated in the femoral vein tissue of the model
group (Fig. 3A-D). These results
suggested that PI3K/AKT signaling may be involved in the regulation
of DVT formation.
FXII protein promotes the formation of
DVT by activating PI3K/AKT signaling and upregulating inflammatory
cytokine levels
Based on the aforementioned results, it was then
determined whether FXII is involved in regulating the formation of
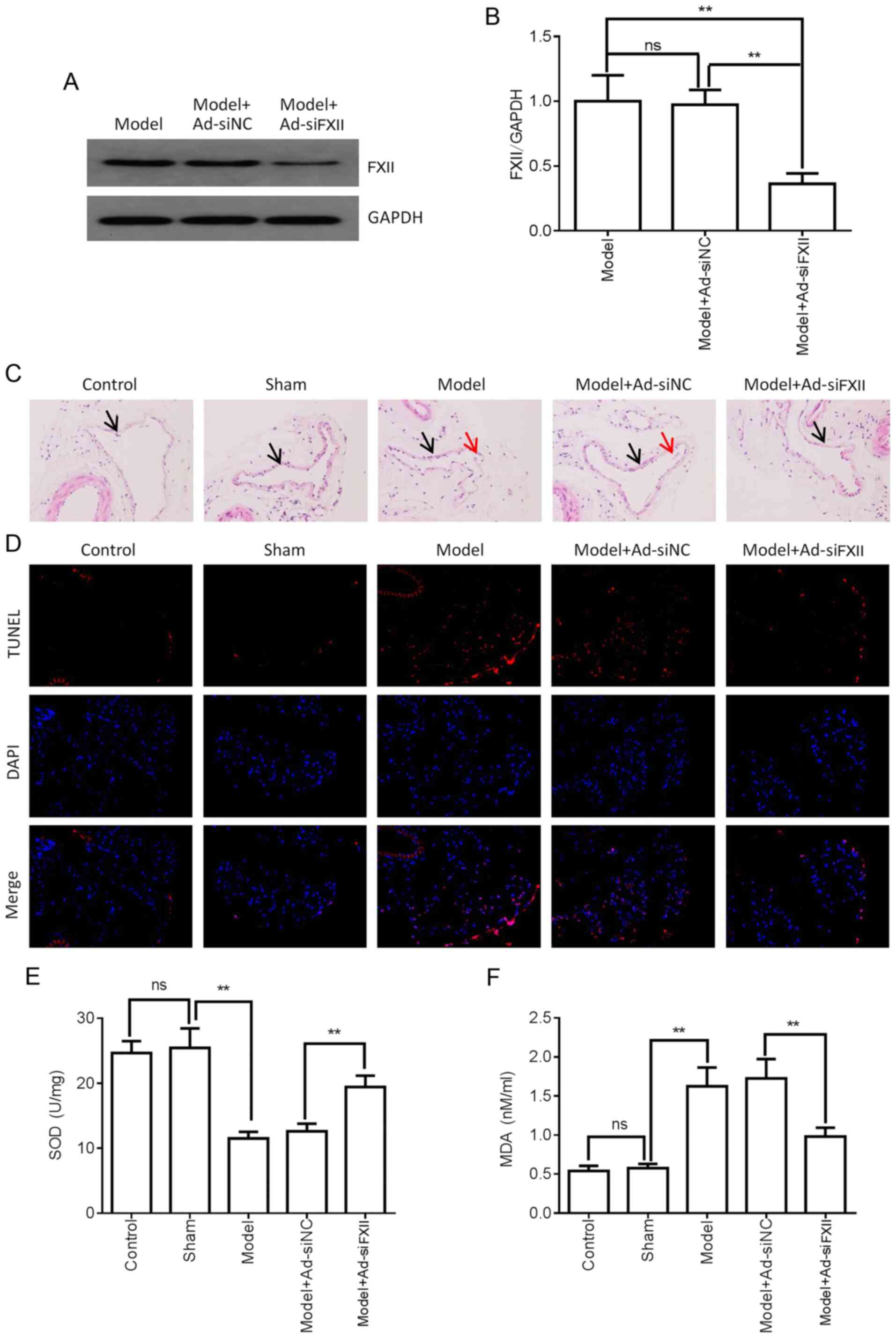
DVT. FXII knockdown was achieved using adenoviral particle
pAd-pG2.1-siRNA FXII in mice with DVT. The transfection efficiency
of pAd-pG2.1-siRNA FXII in mice was first assayed by detecting the
protein expression level of FXII following transfection with
pAd-pG2.1-siRNA FXII in the femoral vein tissue. The results
revealed that FXII protein expression was downregulated in the
femoral vein tissue of the pAd-pG2.1-siRNA FXII model group
compared with model or pAd-pG2.1-siRNA NC model groups (Fig. 4A and B). H&E staining
revealed that pAd-pG2.1-siRNA FXII transduction significantly
attenuated thrombosis compared with pAd-pG2.1-NC transduction in
the femoral vein tissue of mice with DVT (Fig. 4C). Femoral vein tissue damage was
then examined by TUNEL assay. As shown in Fig. 4D, FXII knockdown significantly
decreased cell apoptosis in the femoral vein tissue of mice with
DVT. FXII knockdown also significantly downregulated the MDA
concentrations and upregulated the SOD concentrations in the
femoral vein tissue of mice with DVT (Fig. 4E and F). Furthermore, ELISA was
performed to assess the effects of FXII knockdown on inflammatory
cytokine expression. As shown in Fig. 5A, C and E, FXII knockdown
significantly downregulated the expression TNF-α, IL-6 and IL-8
protein in the femoral vein tissue and plasma of mice with DVT. As
shown in Fig. 5B, D and F, FXII
knockdown significantly downregulated the protein expression of
TNF-α, IL-6 and IL-8 in the plasma of mice with DVT. Additionally,
the effects of FXII knockdown on the PI3K/AKT signaling pathway
were assessed by western blot analysis and immunohistochemical
staining. As shown in Fig. 6A and
B, FXII knockdown did not markedly alter PI3K and AKT protein
expression. However, the p-PI3K and p-AKT levels significantly
decreased following FXII knockdown in the femoral vein tissue of
mice with DVT (Fig. 6A-D). Taken
together, the data indicated that FXII induced DVT formation by
regulating PI3K/AKT signaling and inflammatory cytokines.
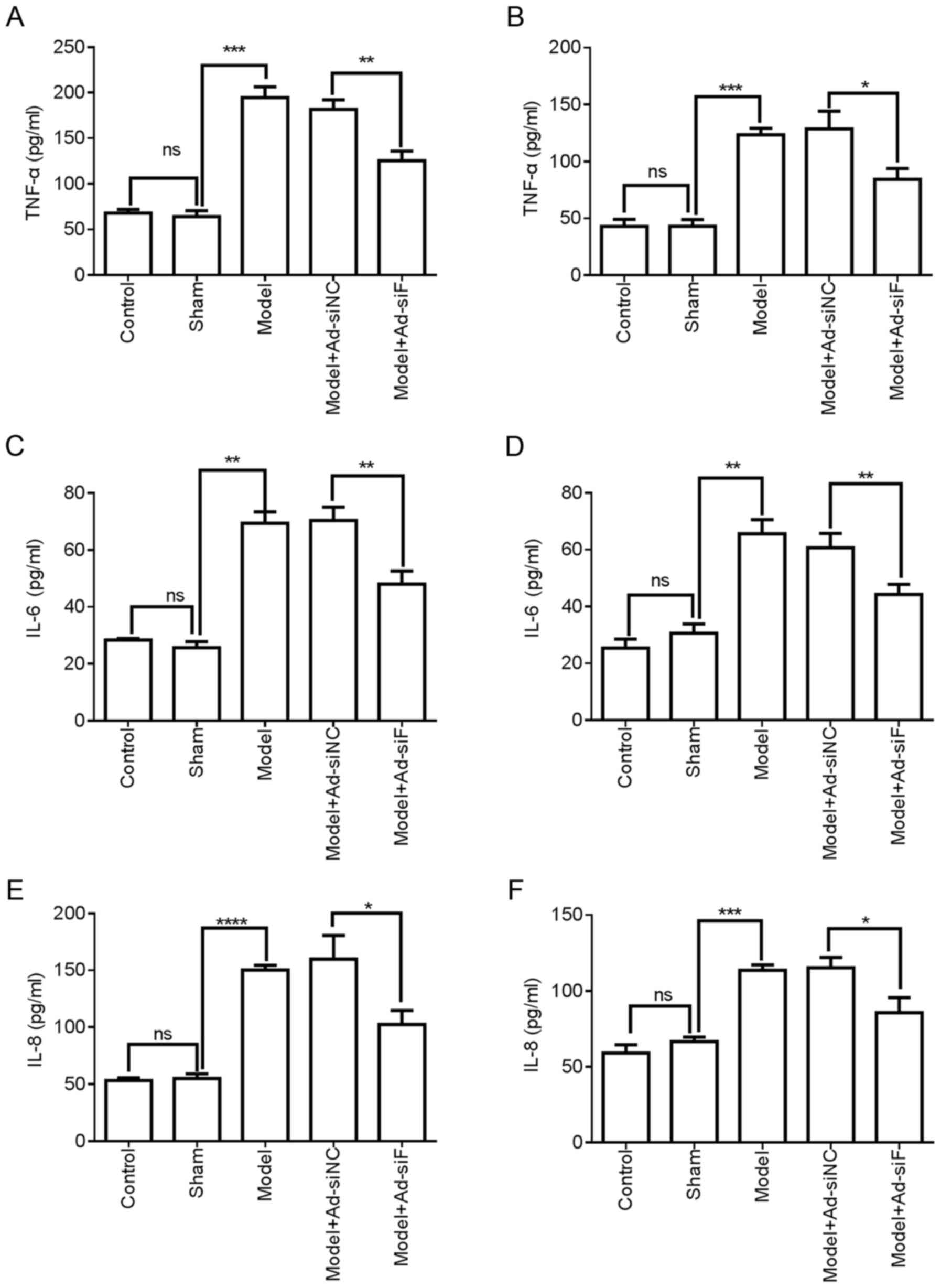 | Figure 5Effects of FXII protein on TNF-α, IL-6
and IL-8 protein in the femoral vein tissue and plasma of mice with
DVT. FXII protein was knocked down by transfection with
pAd-pG2.1-siRNA FXII (1×108 pfu/mice). (A, C and E) The
levels of TNF-α, IL-6 and IL-8 in the femoral vein tissue of
different treatment groups (n=6 mice in each group) were detected
using ELISA. (B, D, and F) The levels of TNF-α, IL-6 and IL-8 in
the plasma of mice in the different treatment groups (n=6 mice in
each group) were detected using ELISA. *P<0.05,
**P<0.01, ***P<0.001 and
****P<0.0001; ns, not significant. FXII, coagulation
factor XII; DVT, deep vein thrombosis; siRNA, small interfering
RNA; pfu, plaque-forming units. |
PI3K/AKT signaling promotes DVT formation
by upregulating inflammatory cytokine levels
The present study also investigated whether the
activation of PI3K/AKT signaling induces DVT formation. The results
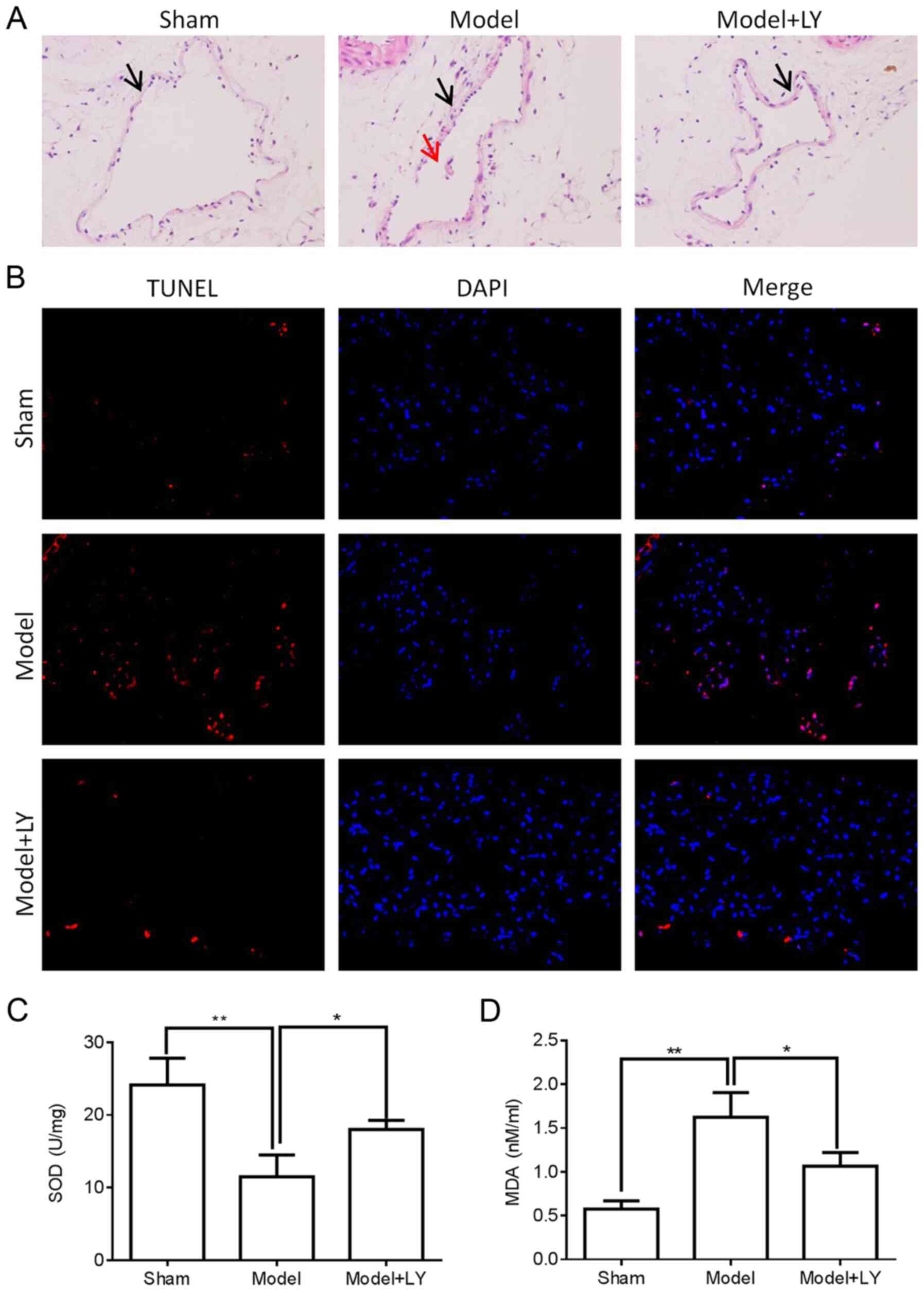
of H&E staining demonstrated that pre-treatment with LY294002,
a PI3K inhibitor, markedly decreased thrombosis in the femoral vein
tissue of mice with DVT (Fig.
7A). Furthermore, femoral vein tissue damage was assessed by
TUNEL assay and ELISA. As shown in Fig. 7B-D, LY294002 pre-treatment
markedly decreased cell apoptosis and the MDA concentration, and
increased the SOD concentration in the femoral vein tissue of mice
with DVT. In addition, LY294002 pre-treatment significantly
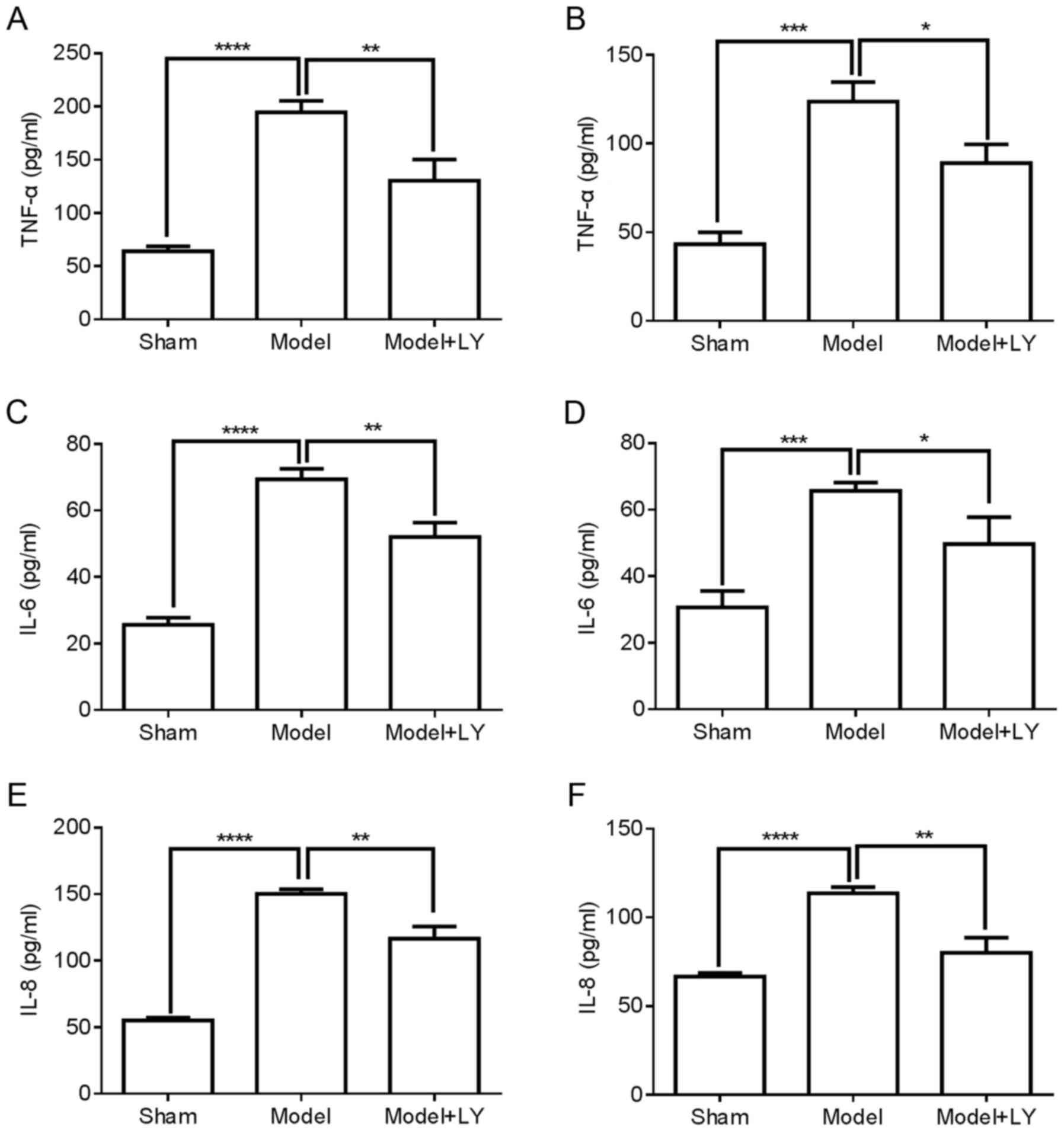
attenuated the protein expression of TNF-α, IL-6 and IL-8 in the
femoral vein tissue of mice with DVT (Fig. 8A, C and E) and plasma of mice
with DVT (Fig. 8B, D and F).
These results suggested that PI3K/AKT signaling induced DVT
formation by regulating inflammatory cytokine levels.
Discussion
The present study found that thrombosis and damage
significantly increased in the femoral vein tissue of mice with DVT
compared with the control or sham groups. It was also found that
FXII protein, PI3K/AKT signaling and the protein levels of the
inflammatory cytokines, TNF-α, IL-6 and IL-8, were upregulated in
the femoral vein tissue of mice with DVT. Mechanistic analyses
further demonstrated that the effects of FXII protein on thrombosis
and femoral vein tissue damage in mice with DVT was associated with
PI3K/AKT signaling and the inflammatory cytokines, TNF-α, IL-6 and
IL-8. FXII protein promoted DVT formation by upregulating the
protein expression of the inflammatory cytokines, TNF-α, IL-6 and
IL-8, via PI3K/AKT signaling.
Accumulating evidence has demonstrated that the
imbalance of inflammatory cytokine expression was closely
associated with DVT formation (22,23). TNF-α, IL-6 and IL-8 proteins are
important pro-inflammatory cytokines. It has been reported that the
upregulation of miRNA-9-5p can alleviate thrombosis in rats with
DVT by decreasing the expression of TNF-α, IL-6 and IL-8 (24). Zhang et al demonstrated
that anti-IL-6 antibody decreased IL-6 expression and alleviated
DVT formation in a mouse model (7). The present study found that the
levels of TNF-α, IL-6 and IL-8 protein were significantly
upregulated in the femoral vein tissue and plasma of mice with DVT
compared with the control or sham groups, which is consistent with
the findings of previous studies (7,24). These data further illustrated
that the inflammatory cytokines, TNF-α, IL-6 and IL-8, play a role
in DVT formation.
Numerous studies have indicated that the PI3K/AKT
signaling pathway is an important intracellular signaling pathway
that plays a role in a variety of cellular physiological processes,
including cell proliferation, cell apoptosis and inflammatory
response (25-27). Chang et al reported that
7-ketocholesterol contributed to thrombosis via the induction of
endothelial damage, apoptosis and inflammatory responses, which
were associated with the activation of PI3K/AKT signaling (28). Su et al also reported that
the pyrrolidinoindoline alkaloid, Psm2, alleviated platelet
aggregation and thrombus formation by inhibiting PI3K/AKT signaling
(29). Additionally, it has been
reported that plantamajoside inhibited the lipopolysaccharide
(LPS)-induced expression of IL-6 and IL-8 by inhibiting PI3K/AKT
signaling in human gingival fibroblasts (30). Vitamin D has been shown to
attenuate the LPS-induced upregulation of TNF-α and IL-6 by
inhibiting PI3K/AKT signaling in human umbilical vein endothelial
cells (31). The results of the
present study suggested that PI3K/AKT signaling was activated in
the femoral vein tissue of mice with DVT. Furthermore, LY294002
pre-treatment markedly decreased thrombosis and femoral vein tissue
damage in mice with DVT. LY294002 pre-treatment also significantly
attenuated the protein expression TNF-α, IL-6 and IL-8 in the
femoral vein tissue and plasma of mice with DVT. The results
presented herein indicated that PI3K/AKT signaling induced DVT
formation by inducing the inflammatory response.
FXII protein is an important member of the
coagulation pathway and plays a key role in coagulation,
fibrinolysis and angiogenesis. Increasing evidence has demonstrated
that FXII protein is involved in regulating thrombosis (12,32). Trauma-induced microvascular
thrombus formation could be minimized by the inhibition of
activated FXII in mice (33).
The results of the present study also demonstrated that FXII
protein expression was significantly increased in the femoral vein
tissue of DVT mice compared with control or sham groups. The
knockdown of FXII protein significantly attenuated thrombosis and
femoral vein tissue damage in mice with DVT. Furthermore, LaRusch
et al demonstrated that FXII induced the activation of AKT
signaling in human umbilical vein endothelial cells (17). The data of the present study
demonstrated that the knockdown of FXII protein significantly
downregulated the activation of PI3K/AKT signaling in the femoral
vein tissue of mice with DVT. Additionally, it has been
demonstrated that the accumulation of FXII can induce the
production and release of pro-inflammatory TNF-α, IL-6 and IL-8
proteins in lungs affected by acute respiratory distress syndrome
(16). The present study also
found that FXII protein knockdown significantly downregulated the
protein expression TNF-α, IL-6 and IL-8 in the femoral vein tissue
and plasma of mice with DVT. These results suggested that FXII
protein may induce DVT formation by activating PI3K/AKT signaling
and promoting the inflammatory response. However, the present study
did not examine the expression of other coagulation factors (XI,
IX, VIII, VII and others) following the knockdown of FXII. The
authors aim to clarify the effects of FXII on the expression of
other coagulation factors in the future.
Taken together, the present study demonstrated that
the upregulation of FXII protein may contribute to the formation of
DVT in mice. The results further demonstrated that FXII protein
promoted DVT formation by inducing the inflammatory response
through the activation of PI3K/AKT signaling in mice with DVT.
Therefore, the inhibition of FXII protein may be a potential
strategy for the treatment of DVT.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YM and HT conducted the experiments and analyzed the
data. YM, QY, QM and HT made substantial contributions to the
design of the present study, and prepared the manuscript. HQ, BZ,
HP and JZ performed the western blot analysis and analyzed the
data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental procedures and protocols were
reviewed and approved by the Animal Investigation Ethics Committee
of the First Affiliated Hospital of Xi'an Jiaotong University and
were performed in accordance with the Guidelines for the Care and
Use of Laboratory Animals from the National Institutes of Health,
China.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
No funding was received.
References
|
1
|
Strijkers RH, Cate-Hoek AJ, Bukkems SF and
Wittens CH: Management of deep vein thrombosis and prevention of
post-thrombotic syndrome. BMJ. 343:d59162011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sun J, Zhang Z, Ma T, Yang Z, Zhang J, Liu
X, Lu D, Shen Z, Yang J and Meng Q: Endothelial progenitor
cell-derived exosomes, loaded with miR-126, promoted deep vein
thrombosis resolution and recanalization. Stem Cell Res Ther.
9:2232018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Giordano NJ, Jansson PS, Young MN, Hagan
KA and Kabrhel C: Epidemiology, pathophysiology, stratification,
and natural history of pulmonary embolism. Tech Vasc Interv Radiol.
20:135–140. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Appelen D, van Loo E, Prins MH, Neumann MH
and Kolbach DN: Compression therapy for prevention of
post-thrombotic syndrome. Cochrane Database Syst Rev.
9:CD0041742017.PubMed/NCBI
|
|
5
|
Yan Y, Yang H, Hu X, Zhang Z, Ge S, Xu Z,
Gao J, Liu J, White GC and Ma YQ: Kindlin-3 in platelets and
myeloid cells differentially regulates deep vein thrombosis in
mice. Aging (Albany NY). 11:6951–6959. 2019. View Article : Google Scholar
|
|
6
|
Sato K, Sakamoto K, Hashimoto Y, Hanzawa
K, Sueta D, Kojima S, Fukuda M, Usuku H, Kihara F, Hosokawa H, et
al: Risk factors and prevalence of deep vein thrombosis after the
2016 kumamoto earthquakes. Circ J. 83:1342–1348. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang Y, Zhang Z, Wei R, Miao X, Sun S,
Liang G, Chu C, Zhao L, Zhu X, Guo Q, et al: IL (Interleukin)-6
contributes to deep vein thrombosis and is negatively regulated by
miR-338-5p. Arterioscler Thromb Vasc Biol. 40:323–334. 2020.
View Article : Google Scholar :
|
|
8
|
Jia Z, Tu J, Zhao J, Ren B, Tian F, Wang
K, Li S and Jiang G: Aspiration thrombectomy using a large-size
catheter for acute lower extremity deep vein thrombosis. J Vasc
Surg Venous Lymphat Disord. 4:167–171. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Branchford BR and Carpenter SL: The role
of inflammation in venous thromboembolism. Front Pediatr.
6:1422018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
van Montfoort ML and Meijers JC: Recent
insights into the role of the contact pathway in
thrombo-inflammatory disorders. Hematology Am Soc Hematol Educ
Program. 2014:60–65. 2014. View Article : Google Scholar
|
|
11
|
Schmaier AH: Physiologic activities of the
contact activation system. Thromb Res. 133(Suppl 1): S41–S44. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zamolodchikov D, Renne T and Strickland S:
The Alzheimer's disease peptide beta-amyloid promotes thrombin
generation through activation of coagulation factor XII. J Thromb
Haemost. 14:995–1007. 2016. View Article : Google Scholar :
|
|
13
|
Renne T, Pozgajova M, Gruner S, Schuh K,
Pauer HU, Burfeind P, Gailani D and Nieswandt B: Defective thrombus
formation in mice lacking coagulation factor XII. J Exp Med.
202:271–281. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kleinschnitz C, Stoll G, Bendszus M, Schuh
K, Pauer HU, Burfeind P, Renné C, Gailani D, Nieswandt B and Renné
T: Targeting coagulation factor XII provides protection from
pathological thrombosis in cerebral ischemia without interfering
with hemostasis. J Exp Med. 203:513–518. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Matafonov A, Leung PY, Gailani AE, Grach
SL, Puy C, Cheng Q, Sun MF, McCarty OJ, Tucker EI, Kataoka H, et
al: Factor XII inhibition reduces thrombus formation in a primate
thrombosis model. Blood. 123:1739–1746. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hess R, Wujak L, Hesse C, Sewald K, Jonigk
D, Warnecke G, Fieguth HG, de Maat S, Maas C, Bonella F, et al:
Coagulation factor XII regulates inflammatory responses in human
lungs. Thromb Haemost. 117:1896–1907. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
LaRusch GA, Mahdi F, Shariat-Madar Z,
Adams G, Sitrin RG, Zhang WM, McCrae KR and Schmaier AH: Factor XII
stimulates ERK1/2 and Akt through uPAR, integrins, and the EGFR to
initiate angiogenesis. Blood. 115:5111–5120. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schonfelder T, Jackel S and Wenzel P:
Mouse models of deep vein thrombosis. Gefasschirurgie. 22:28–33.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tokgoz VY, Sipahi M, Keskin O, Guvendi GF
and Takir S: Protective effects of vitamin D on
ischemia-reperfusion injury of the ovary in a rat model. Iran J
Basic Med Sci. 21:593–599. 2018.PubMed/NCBI
|
|
20
|
Xu L, Yu Y, Sang R, Li J, Ge B and Zhang
X: Protective effects of Taraxasterol against Ethanol-Induced liver
injury by regulating CYP2E1/Nrf2/HO-1 and NF-κB signaling pathways
in mice. Oxid Med Cell Longev. 2018:82841072018. View Article : Google Scholar
|
|
21
|
Lu Y, Wu S, Xiang B, Li L and Lin Y:
Curcumin attenuates oxaliplatin-induced liver injury and oxidative
stress by activating the Nrf2 pathway. Drug Des Devel Ther.
14:73–85. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Roumen-Klappe EM, den Heijer M, van Uum
SH, van der Ven-Jongekrijg J, van der Graaf F and Wollersheim H:
Inflammatory response in the acute phase of deep vein thrombosis. J
Vasc Surg. 35:701–706. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tato F: Deep vein thrombosis-advances in
diagnosis and treatment. MMW Fortschr Med. 156(Spec no 2): 59–63;
quiz 64. 2014.In German.
|
|
24
|
Ou M, Zhang Y, Cui S, Zhao S and Tu J:
Upregulated MiR-9-5p protects against inflammatory response in rats
with deep vein thrombosis via inhibition of NF-κB p50.
Inflammation. 42:1925–1938. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li Y, Jiang Y, Wan Y, Zhang L, Tang W, Ma
J, Wu S and Cheng W: Medroxyprogestogen enhances apoptosis of
SKOV-3 cells via inhibition of the PI3K/Akt signaling pathway. J
Biomed Res. 27:43–50. 2013.PubMed/NCBI
|
|
26
|
Involvement of pro-inflammatory cytokines
in diabetic neuropathic pain via central PI3K/Akt/mTOR signal
pathway. Arch Physiol Biochem. Dec 11–2019.Epub ahead of print.
View Article : Google Scholar
|
|
27
|
Zhang HB, Tu XK, Chen Q and Shi SS:
Propofol reduces inflammatory brain injury after subarachnoid
hemorrhage: Involvement of PI3K/Akt pathway. J Stroke Cerebrovasc
Dis. 28:1043752019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chang MC, Chen YJ, Liou EJ, Tseng WY, Chan
CP, Lin HJ, Liao WC, Chang YC, Jeng PY and Jeng JH:
7-Ketocholesterol induces ATM/ATR, Chk1/Chk2, PI3K/Akt signalings,
cytotoxicity and IL-8 production in endothelial cells. Oncotarget.
7:74473–74483. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Su XL, Su W, Wang Y, Wang YH, Ming X and
Kong Y: The pyrrolidinoindoline alkaloid Psm2 inhibits platelet
aggregation and thrombus formation by affecting PI3K/Akt signaling.
Acta Pharmacol Sin. 37:1208–1217. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu F, Huang X, He JJ, Song C, Peng L,
Chen T and Wu BL: Plantamajoside attenuates inflammatory response
in LPS-stimulated human gingival fibroblasts by inhibiting PI3K/AKT
signaling pathway. Microb Pathog. 127:208–211. 2019. View Article : Google Scholar
|
|
31
|
Zhou W, Yuan G and Wang Q: Vitamin D
attenuates lipopoly-saccharide-induced inflammatory response in
endothelial cells through inhibition of PI3K/Akt/NF-κB signaling
pathway. Pharmazie. 74:412–417. 2019.PubMed/NCBI
|
|
32
|
Maas C and Renne T: Coagulation factor XII
in thrombosis and inflammation. Blood. 131:1903–1909. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hopp S, Albert-Weissenberger C, Mencl S,
Bieber M, Schuhmann MK, Stetter C, Nieswandt B, Schmidt PM,
Monoranu CM, Alafuzoff I, et al: Targeting coagulation factor XII
as a novel therapeutic option in brain trauma. Ann Neurol.
79:970–982. 2016. View Article : Google Scholar : PubMed/NCBI
|