Introduction
Diabetic encephalopathy (DE) is one of the main
chronic complications of diabetes, and is characterized by acquired
cognitive and behavioral defects, accompanied by changes in brain
morphology and abnormal neurophysiological functions (1–3).
The clinical manifestations of DE include decreased learning and
memory ability, impaired comprehension and judgment, and slow
thinking (4,5). It is of great importance to
determine the pathogenesis of DE and to identify effective
preventive and treatment strategies.
MicroRNAs (miRNAs/miRs) are a group of short
non-coding RNAs composed of 20–22 nucleotides, which play a key
role in mRNA suppression and degradation (6). Previous studies have demonstaated
that miRNAs are widely involved in the development of
diabetes-related complications (7–10). miR-130b is downregulated in the
serum of patients with diabetic nephropathy (DN), and has been
reported to improve epithelial-interstitial transition in DN and
ameliorate tubulointerstitial fibrosis (11,12). The expression of miR-130b is
decreased in the serum of patients with diabetes with coronary
heart disease complications (13). However, the expression pattern
and role of miR-130b in DE is currently unknown.
Previous studies have indicated that phosphatase and
tensin homolog (PTEN) is one of the target genes of miR-130b
(14–16). PTEN is an antioncogene involved
in the regulation of cell proliferation, migration, differentiation
and invasion in various malignant tumor cells (17). PTEN exerts its biological
function in part by inhibiting the PI3K/Akt signaling pathway
(18). The PI3K/Akt signaling
pathway is a key signaling pathway involved in cell metabolism,
growth, proliferation, survival, transcription and protein
synthesis (19). In addition,
the PI3K/Akt signaling pathway has also been shown to be associated
with oxidative stress (20–22), which has been regarded as one of
the pathogenesis of DE (23).
Thus, the present study aimed to evaluate whether miR-130b
activates Akt activity to protect against oxidative stress-induced
injury via the regulation of PTEN in DE.
Materials and methods
Cell culture and high glucose
treatment
PC12 (no. CL-0480) and hippocampal cells (no.
CP-R107) were obtained from Procell Life Science & Technology
Co., Ltd. PC12 and hippocampal cells were cultured in RPMI-1640
medium (Gibco; Thermo Fisher Scientific, Inc.) containing 11.11 mM
glucose (Sigma-Aldrich LLC.), 15% fetal bovine serum (Gibco; Thermo
Fisher Scientific, Inc.) and 1% penicillin-streptomycin (Gibco;
Thermo Fisher Scientific, Inc.), and placed in a cell incubator at
37°C with 5% CO2. For high glucose conditions, cells
were cultured in the aforementioned medium but containing 75 mM
glucose for 72 h (24).
Animals and treatment
Male Wistar rats (n=40; weight, 180–220 g) were
purchased from Chengdu Dashuo Experimental Animal Co., Ltd.
Following one week of adaptive feeding, the rats were randomly
divided into four groups (n=10/group) and kept in their cages in a
controlled environment, at constant room temperature (18–25°C) and
humidity (40–60%) under a normal 12-h light/dark cycle. Rats in the
control group received an intraperitoneal injection of 0.9% saline.
Rats in the other groups were administered streptozotocin (STZ;
Sigma-Aldrich; Merck KGaA) in a single intraperitoneal injection at
a dose of 60 mg/kg for the induction of diabetes. At 72 h after the
STZ injection, 200 μl blood were collected from the tail
vein to measure blood glucose, and blood glucose levels ≥16.7
mmol/l were considered to indicate successful modeling. The
STZ-treated rats were divided into three groups at three days after
the STZ injection. The model group was left untreated and served as
the untreated diabetic group. The agomir-NC and agomir-130b groups
received agomir-NC (miR4N0000001-4-5; Guangzhou RiboBio Co., Ltd.)
or agomir-130b (miR40000837-4-5; Guangzhou RiboBio Co., Ltd.) via
intranasal administration once a week at a dose of 5 nmol (25). These treatments were continued
until the end of the study period (8 weeks). The study protocol was
approved by the Animal Experimental Ethics Committee of Henan
Provincial People's Hospital (approval no. 2020408A).
Preparation of hippocampal samples
Following an intraperitoneal injection of 3%
pentobarbital sodium (50 mg/kg) for anesthesia, the animals were
sacrificed by cervical dislocation, and the hippocampal tissues of
the rats were collected immediately. One part of the collected
hippocampus tissues was initially flash-frozen in liquid nitrogen
for RT-qPCR, western blot analysis and biochemical examination. The
remaining hippocampal tissues were collected for pathological
examination.
Cell transfection
PC12 and hippocampal cells (1×106
cells/well) were cultured in 6-well plates until reaching 70%
confluence. Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to perform cell transfection. Cells were
transfected with 24 nM miR-130b mimics (miR10000837-1-5; Guangzhou
RiboBio Co., Ltd.), negative control (NC) of miRNA mimic (NC mimic;
miR1N0000001-1-5; Guangzhou RiboBio Co., Ltd.), miR-130b inhibitor
(miR20000837-1-5; Guangzhou RiboBio Co., Ltd.) or NC of miRNA
inhibitor (NC inhibitor; miR2N0000001-1-5; Guangzhou RiboBio Co.,
Ltd.), or 30 nM small interfering RNA (siRNA/si) against PTEN
(si-PTEN; siB1142121626-1-5; Guangzhou RiboBio Co., Ltd.) or NC
siRNA (si-NC; siN0000002-1-5; Guangzhou RiboBio Co., Ltd.). The
cells were collected for further analysis following transfection
for 48 h.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from the PC12 cells,
hippocampal cells and hippocampal tissue using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). Extracted RNA was
reverse transcribed into cDNA for miRNA and mRNA detection using
the SuperScript™ IV First-Strand Synthesis system (Invitrogen;
Thermo Fisher Scientific, Inc.) and Mir-X miRNA First-Strand
Synthesis kit (Takara Bio, Inc.), respectively. The expression of
miR-130b and PTEN was determined by RT-qPCR using a Mir-X miRNA
qRT-PCR TB Green® kit or TB Green® Fast qPCR mix (Takara Bio,
Inc.), respectively. U6 and GAPDH were used for the normalization
of miRNA and mRNA levels, respectively. The results of the RT-qPCR
were calculated using the 2−ΔΔCq method (26). The thermocycling conditions were
as follows: Initial denaturation at 97°C for 3 min, followed by 40
cycles at 95°C for 15 sec and 60°C for 30 sec, and melting curve
stage at taking fluorescent measurements every 0.5°C for 15 sec
from 55°C until 95°C. GAPDH was used as a standardized internal
control. The primer sequences are presented in Table I.
 | Table ISequences of primers used for
RT-qPCR. |
Table I
Sequences of primers used for
RT-qPCR.
| Name | Sequences
(5′-3′) |
|---|
| miR-130b | Sense:
ACTCTTTCCCTGTTGCACTAC
Universal antisense: Mir-X miRNA qRT-PCR TB Green® kit (cat. no.
638314; Clontech, Takara Bio, Inc.) provided |
| PTEN | Sense:
CAATGTTCAGTGGCGAACTT
Antisense: GGCAATGGCTGAGGGAACT |
| U6 | Mir-X miRNA qRT-PCR
TB Green® kit (cat. no. 638314; Clontech, Takara Bio, Inc.)
provided |
| GAPDH | Sense:
TTCCTACCCCCAATGTATCCG
Antisense: CATGAGGTCCACCACCCTGTT |
Dual-luciferase reporter assay
TargetScan (www.targetscan.org) was utilized to predict the
binding site between PTEN and miR-130b. Wild-type (Wt) or mutant
(Mut) fragments of the 3′-untranslated region (UTR) from PTEN was
synthesized by HanBio Co., Ltd., which included the potential
miR-130b binding sites. The fragments were then ligated into the
psiCheck2 vector (Promega Corporation), and the constructs were
named PTEN-Wt and PTEN-Mut, respectively.
For the dual-luciferase reporter assay, 293T cells
(cat. no. CL-0005; Procell Life Science & Technology Co., Ltd.)
were seeded at a density of 2×104 cells/well in 24-well
plates and cultured in RPMI-1640 medium containing 15% fetal bovine
serum and 1% penicillin-streptomycin for 24 h. The cells were then
co-transfected with 24 nM miR-130b mimic/NC mimic and 50 nM
PTEN-Wt/Mut reporter plasmids using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). Cells were collected
following transfection for 24 h. The Dual-Luciferase Reporter Assay
system (Promega Corporation) was used to monitor the luciferase
activity. Relative luciferase activity was normalized as the ratio
of firefly luciferase activity to Renilla luciferase
activity. Each experiment was performed in triplicate.
Cell viability assay
Cell viability was detected using the Cell Counting
Kit-8 (CCK-8) assay (Dojindo Molecular Technologies, Inc.). PC12
and hippocampal cells (6,000 cells/well) were seeded into 96-well
plates and treated as aforementioned for processing for 48 h. The
cells were then incubated in 10% CCK-8 solution at 37°C for 1 h,
and the absorbance value was measured at 450 nm using a microplate
reader (Thermo Fisher Scientific, Inc.). Each experiment was
performed in triplicate.
Apoptosis assay
Cell apoptosis was detected using an Annexin
V-FITC/PI Apoptosis Detection kit (Beyotime Institute of
Biotechnology) and flow cytometry. Cells were digested with 0.25%
Trypsin-EDTA (Gibco; Thermo Fisher Scientific, Inc.) at 37°C for 1
min. The cells were then washed with PBS buffer solution (Gibco;
Thermo Fisher Scientific, Inc.), centrifugated at 200 × g at 4°C
for 5 min and double-stained with 10 μl Annexin V-FITC/PI
mix for 15 min at room temperature in the dark. Subsequently, the
cells were evaluated by a FACSCalibur™ Flow Cytometer (BD
Biosciences) and analyzed using Cell Quest 3.3 software (BD
Biosciences). The total apoptotic rate of the cells was defined as
the sum of both early apoptosis (Annexin V-FITC-positive,
PI-negative) and late apoptosis (Annexin V-FITC- and PI-positive).
Each experiment was performed in triplicate.
Western blot analysis
Total protein was isolated from the PC12 cells,
hippocampal cells and hippocampal tissue using
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology). Bicinchoninic acid assay (Thermo Fisher Scientific,
Inc.) was used to quantify proteins. Subsequently, proteins (30
μg per lane) were mixed with loading buffer (Beijing
Solarbio Science & Technology Co., Ltd.), boiled for 15 min and
separated by 10% SDS-PAGE (Beyotime Institute of Biotechnology).
The proteins were transferred to PVDF membranes (Millipore Sigma).
After blocking with 5% skim milk for 1 h at room temperature, the
PVDF membranes were incubated at 4°C overnight with the
corresponding primary antibodies, including anti-PTEN (1:1,000;
cat. no. ab267787; Abcam), anti-Akt (1:500; cat. no. ab8805;
Abcam), anti-phosphorylated (p)-Akt (1:500; cat. no. ab38449;
Abcam), anti-Bcl-2 (1:500; cat. no. ab59348; Abcam), anti-Bax
(1:2,000; cat. no. ab32503; Abcam), anti-cleaved caspase-3 (1:500;
cat. no. ab49822; Abcam), anti-caspase-3 (1:1,000; cat. no. 9662;
Cell Signaling Technology, Inc.) and anti-β-actin (1:1,000; cat.
no. ab8227; Abcam). The PVDF membranes were then washed and further
incubated with a secondary antibody conjugated to horseradish
peroxidase (1:2,000; cat. no. ab205718; Abcam) for 1 h at room
temperature. SuperSignal West Pico Plus (Thermo Fisher Scientific,
Inc.) was employed to visualize the protein bands. Image-Pro Plus
6.0 (Media Cybernetics, Inc.) was used to semi-quantify the
relative band intensities. β-actin was employed for normalization
of relative protein expression. Each experiment was performed in
triplicate.
Biochemical detection
PC12 and hippocampal cells were lysed at 4°C using
an ultrasonic cell disruptor and centrifuged at 200 × g at 4°C for
10 min. A total of 100 μl supernatant of each group was
obtained to detect the optical density values using a microplate
reader (spectra max PLUS 384, Molecular Devices, LLC) according to
the instructions of the superoxide dismutase (SOD) assay kit
(Nanjing Jiancheng Bioengineering Institute) (450 nm) and the
malondialdehyde (MDA) assay kit (Nanjing Jiancheng Bioengineering
Institute) (532 nm).
The hippocampal tissues collected as aformenetioned
were shredded using an ophthalmic scissor. Normal saline was then
added to produce a 10% brain tissue homogenate. The homogenate was
centrifuged at 200 × g at 4°C for 10 min, and the supernatant was
obtained for future use. The activity of SOD and the content of MDA
were detected using a colorimetric method according to the
instructions provided with the aforementioned kits.
Behavioral assessment
A step-down passive avoidance test was used to
detect the cognitive competence of the rats (27,28). The test was performed in a
plexiglass cage (20×20×20 cm) with a rubber platform (8.0×8.0×4.5
cm) in the corner. In the training session, the rats were placed in
the plexiglass cage for 3 min to explore the box freely. The rats
were then exposed to an electric shock (36 V) until they stepped
onto the rubber platform. After 24 h, the rats were again placed
onto the rubber platform, and the step-down latency and number of
errors within 5 min were recorded.
Pathological examination
The hippocampal tissues were fixed with 4%
paraformaldehyde for 48 h and prepared into 5-μm-thick
paraffin sections. The pathological changes in the hippocampus
tissues were stained using a hematoxylin and eosin (H&E)
staining kit (Beyotime Institute of Biotechnology). Briefly, the
sections were stained with hematoxylin for 2 min and stained with
eosin for 10 min at room temperature. Following dehydration with
gradient alcohol, the sections were sealed using neutral resin and
observed under a microscope (BA400Digital; Motic Group Co., Ltd.).
The apoptotic cells in the hippocampal tissues were stained using a
TUNEL assay kit (Roche Applied Science). Briefly, the sections were
incubated with protease K for 30 min at 25°C and washed by PBS
twice. The sections were then incubated with TUNEL reaction mixture
at 37°C for 1 h and washed three times with PBS. Thereafter, the
sections were incubated with converter-POD at 37°C for 30 min.
Following three PBS washes, the sections were added to Substrate
solution and incubated at 20°C for 25 min. Ultrastructural damage
of the hippocampus was observed using a transmission electron
microscope (TEM). Briefly, the hippocampal tissues were pre-fixed
with 3% glutaraldehyde (Sigma-Aldrich LLC.) and then fixed with 1%
osmium (VIII) oxide (Electron Microscopy China, EMCN™).
Subsequently, the hippocampal tissues were dehydrated with acetone,
embedded in epoxy resins (EMCN™) and sliced for 50-nm-thick slices
by an ultratome (EM UC7, Leica Inc.). The ultra-thin slices were
loaded on a formvar/carbon coated grid (EMCN™), and stained with
uranium acetate (EMCN™) and lead citrate (EMCN™) at room
temperature for 15–20 min. Finally, the slices were observed under
a JEM-1400 Flash electron microscope (JEOL Ltd.).
Statistical analysis
Data are presented as the mean ± standard deviation.
Experiments were repeated three times. Statistical evaluations and
calculations were performed with SPSS 22.0 (IBM Corp.). Differences
of data were evaluated using an independent-samples t-test between
two groups or one-way analysis of variance followed by a Turkey's
post hoc analysis among multiple groups. P<0.05 was considered
to indicate a statistically significant difference.
Results
High glucose stimulation reduces the
expression of miR-130b in PC12 and hippocampal cells
To examine the effects of high glucose on the
expression of miR-130b in PC12 and hippocampal cells, RT-qPCR was
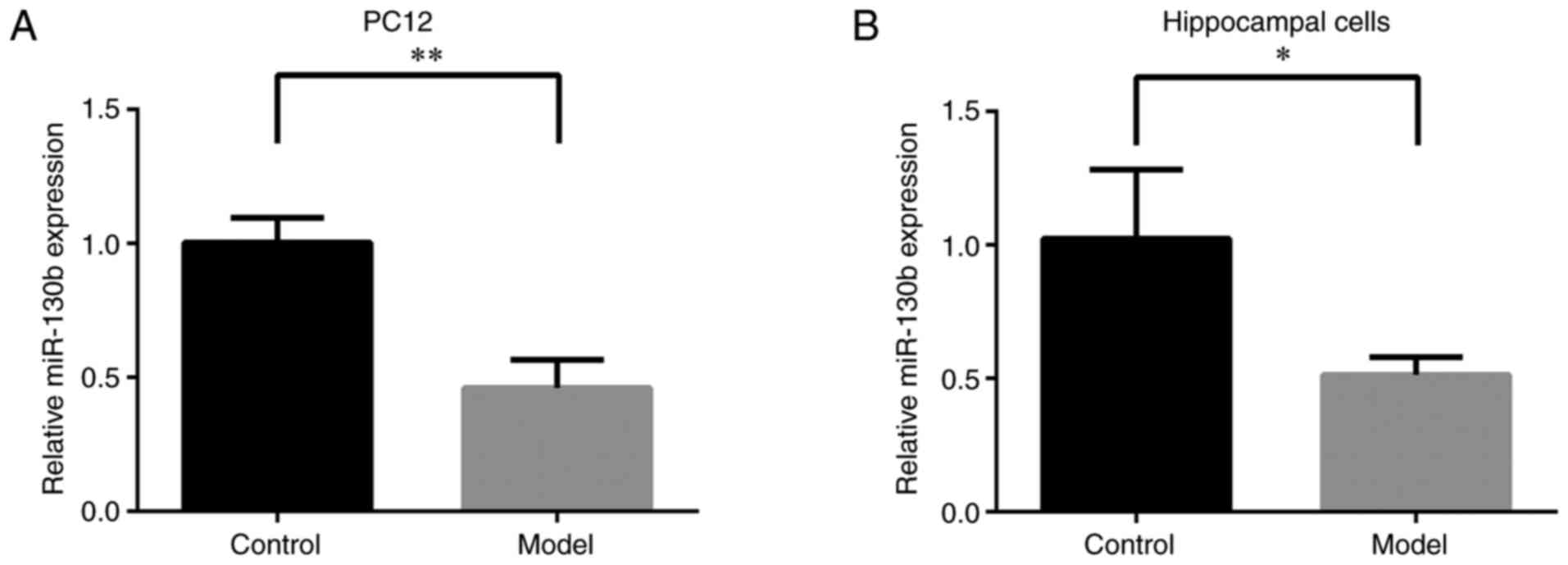
performed to detect the miR-130b levels. As shown in Fig. 1A and B, compared with that of the
control group, the expression of miR-130b in PC12 and hippocampal
cells under a high glucose environment was significantly decreased
(P<0.01 or P<0.05, respectively). These results indicated
that high glucose stimulation inhibited miR-130b expression in PC12
and hippocampal cells.
Overexpression of miR-130b promotes cell
viability and inhibits apoptosis, while miR-130b silencing inhibits
cell viability and promotes the apoptosis of PC12 and hippocampal
cells under an elevated glucose environment
To further investigate the role of miR-130b in PC12
and hippocampal cells in a high glucose environment, miR-130b was
overexpressed using a miR-130b mimic, and miR-130b was silenced
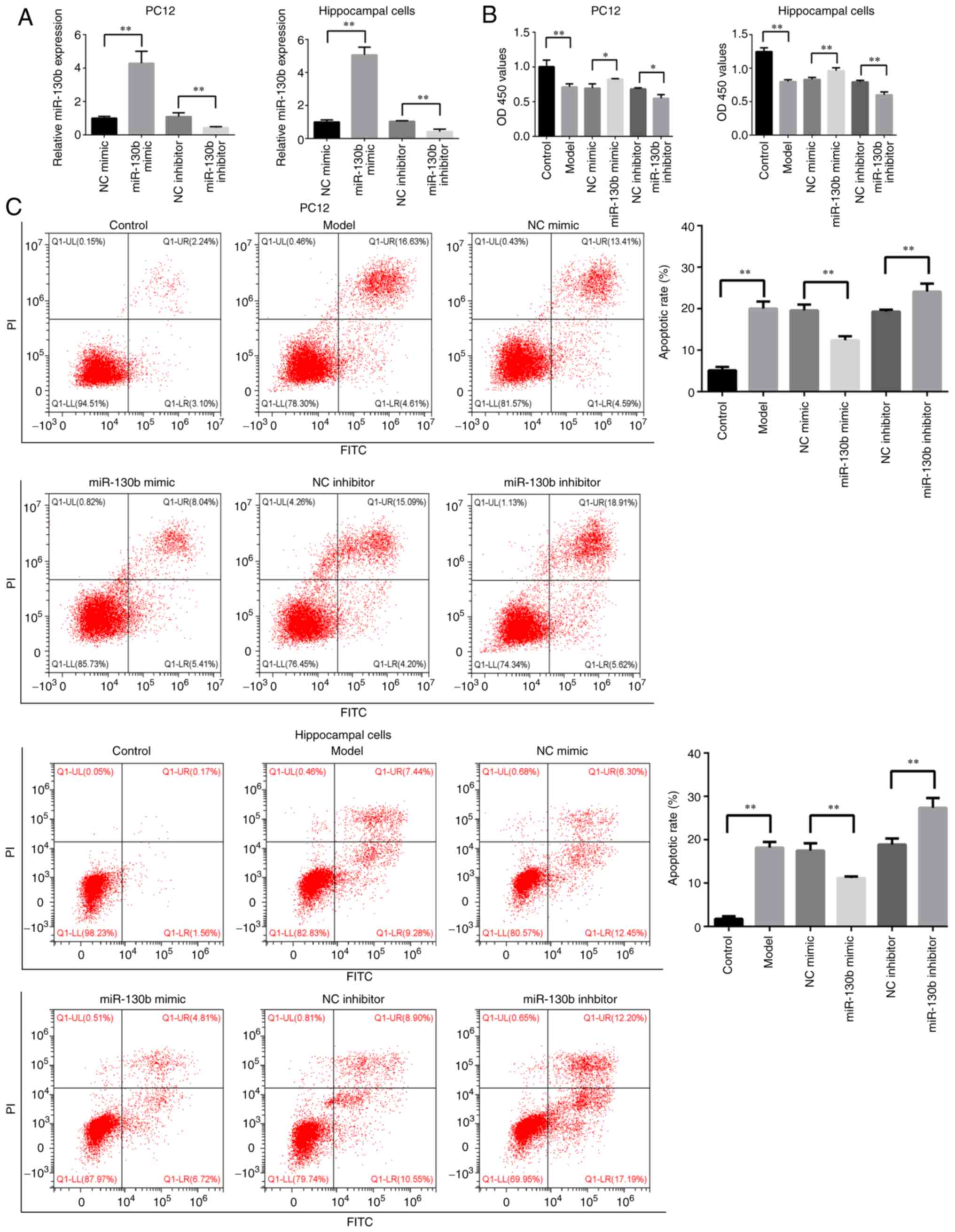
using a miR-130 inhibitor. As shown in Fig. 2A, the expression of miR-130b in
PC12 and hippocampal cells was increased following miR-130b mimic
transfection (P<0.01). The expression of miR-130b was decreased
following incubation with miR-130b inhibitor (P<0.01).
CCK-8 assay was performed to detect the effects of
miR-130b on the viability of PC12 and hippocampal cells in a high
glucose environment. As shown in Fig. 2B, compared with that of the
control group, cell viability was significantly decreased in the
model group (P<0.01). The overexpression of miR-130b increased
cell viability, while the inhibition of miR-130b reduced the
viability of PC12 and hippocampal cells in a high glucose
environment (P<0.05).
The present study also explored the effects of
miR-130b on the apoptosis of PC12 and hippocampal cells under high
glucose conditions. As shown in Fig.
2C, compared with that of the control group, the apoptotic rate
of the model group was significantly increased (P<0.01). The
overexpression of miR-130b decreased the apoptotic rate, while the
inhibition of miR-130b increased the apoptotic rate of PC12 and
hippocampal cells in a high glucose environment (P<0.01). These
results demonstrated that miR-130b attenuated high glucose-induced
cell damage.
PTEN is the target gene of miR-130b in
PC12 and hippocampal cells
To further explore the mechanisms of action of
miR-130b, TargetScan was used to predict the potential targets of
miR-130b. It was found that PTEN was a potential target of miR-130b
(Fig. 3A). To confirm the
potential targeting association between miR-130b and PTEN, a
dual-luciferase reporter assay was performed. miR-130b mimic and
PTEN-Wt or Mut were co-transfected into 293T cells. The data
indicated that the overexpression of miR-130b reduced the
luciferase activity of PTEN-Wt (P<0.01), but not that of
PTEN-Mut (Fig. 3B). Furthermore,
to examine the effects of miR-130b on PTEN mRNA expression in PC12
and hippocampal cells, a RT-qPCR assay was performed. As shown in
Fig. 3C, miR-130b mimic
significantly inhibited the expression of PTEN (P<0.01), while
miR-130b inhibitor enhanced the mRNA expression of PTEN
(P<0.05). These results demonstrated that PTEN was the target
gene of miR-130b in PC12 and rat hippocampal cells.
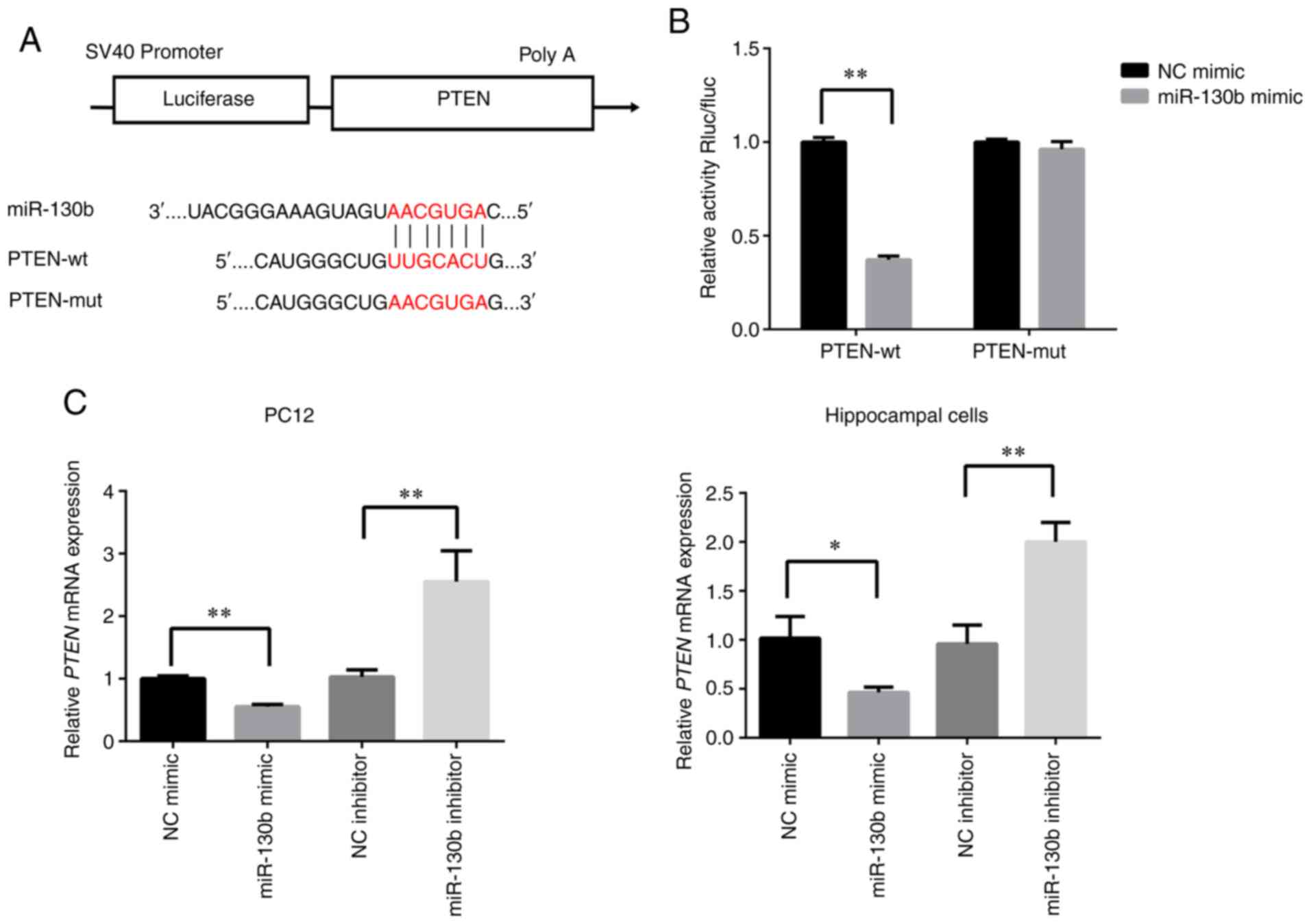 | Figure 3PTEN is the direct target of miR-130b
in PC12 and hippocampal cells. (A) Sequence of binding sites
between PTEN-3′-UTR and miR-130b. (B) Relative luciferase
activities were detected in 293T cells co-transfected with PTEN
3′-UTR (Wt or mut) reporter plasmid, miR-130b mimic or NC mimic.
(C) Reverse transcription-quantitative PCR analysis of PTEN mRNA
expression in PC12 and hippocampal cells transfected with miR-130b
mimic, NC mimic, miR-130b inhibitor and NC inhibitor. U6 served as
the reference gene. *P<0.05, **P<0.01.
PTEN, phosphatase and tensin homolog; UTR, untranslated region;
miR, microRNA; Wt, wild-type; mut, mutant; NC, negative
control. |
Silencing of PTEN promotes the viability,
and inhibits the apoptosis of PC12 and hippocampal cells in a high
glucose environment
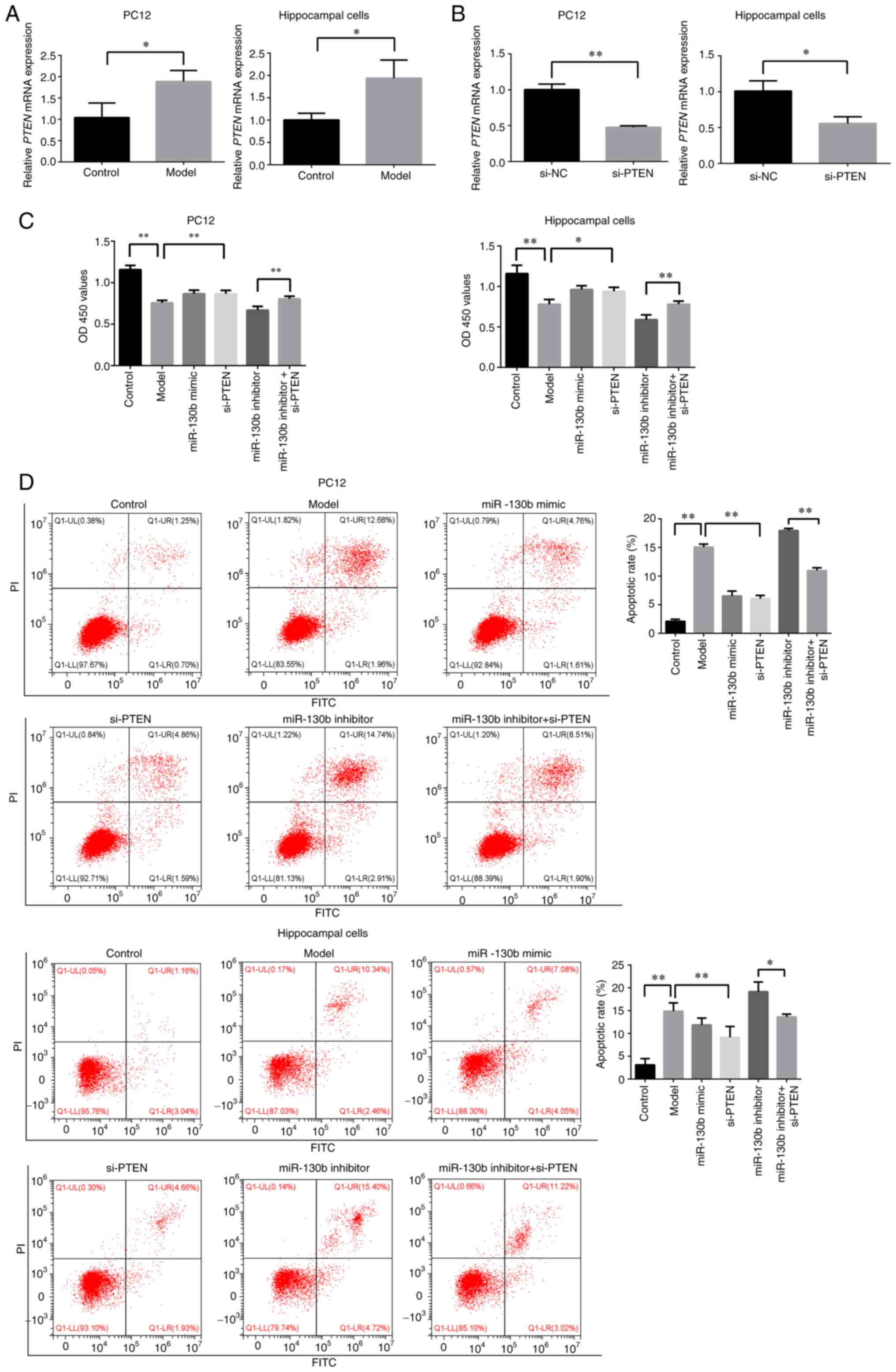
To examine the effects of high glucose on the
expression of PTEN in PC12 and hippocampal cells, RT-qPCR was
performed to detect the PTEN levels. As shown in Fig. 4A, compared with that of the
control group, the expression of PTEN in PC12 and hippocampal cells
in a high glucose environment was significantly increased
(P<0.05). Subsequently, si-PTEN was synthesized, and it was
determined that si-PTEN could effectively interfere with PTEN mRNA
expression in PC12 and hippocampal cells (P<0.05 or P<0.01,
respectively; Fig. 4B). CCK-8
assay was then performed to detect the effects of PTEN on the
viability of PC12 and hippocampal cells in a high glucose
environment. As shown in Fig.
4C, the inhibition of PTEN increased the viability of PC12 and
hippocampal cells in a high glucose environment (P<0.05 or
P<0.01, respectively), and partly reversed the effects of the
miR-130b inhibitor on the reduction of cell viability (P<0.01).
Furthermore, the present study explored the role of PTEN in the
apoptosis of PC12 and hippocampal cells under high glucose
conditions. As shown in Fig. 4D,
the inhibition of PTEN reduced the apoptotic rate of PC12 and
hippocampal cells in a high glucose environment, and partly
abrogated the promoting effects of miR-130b inhibitor on apoptosis
(P<0.05 or P<0.01, respectively). These results indicated
that miR-130b regulated PTEN to attenuate high glucose-induced cell
damage.
miR-130b regulates PTEN to strengthen the
antioxidant capacity of PC12 and hippocampal cells in an elevated
glucose environment
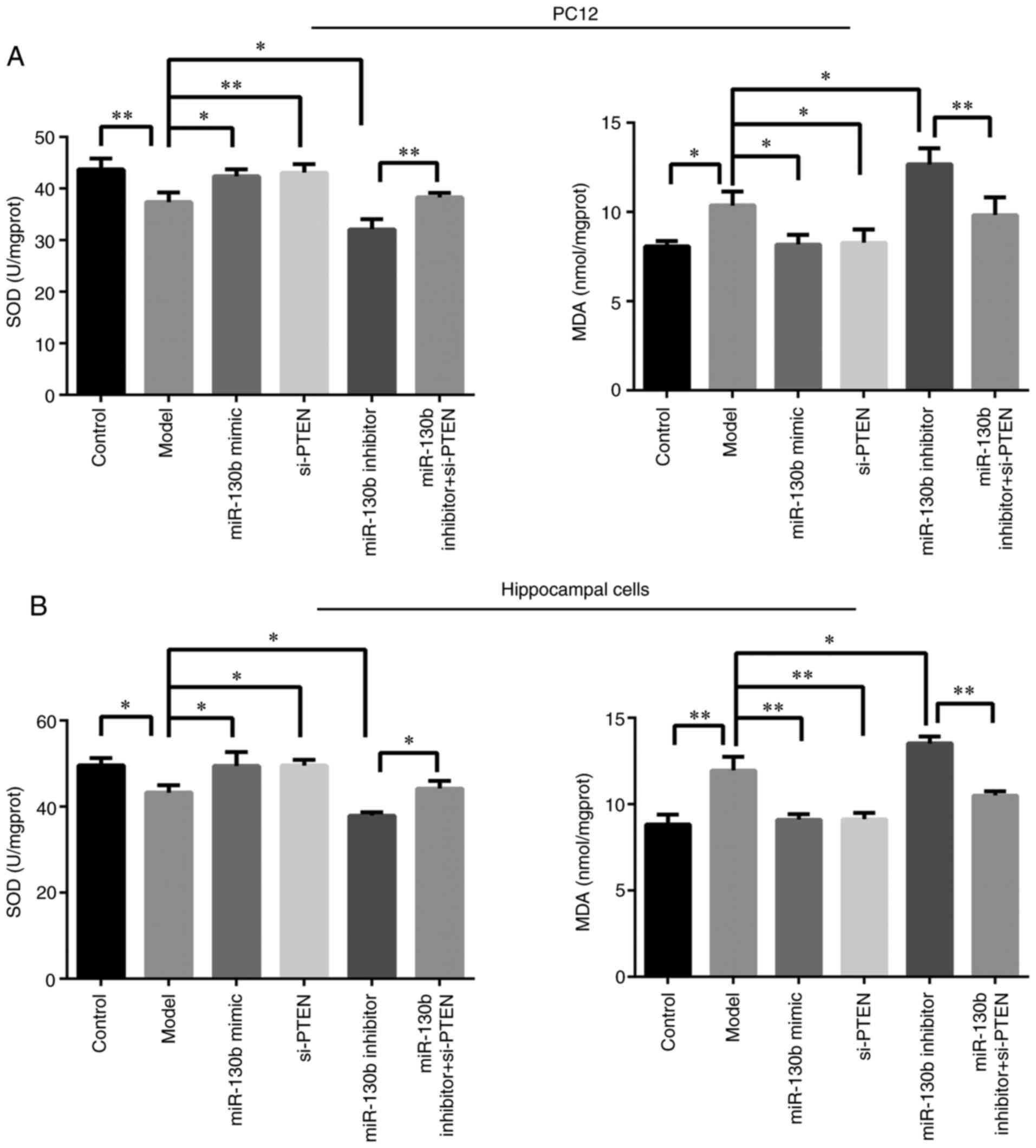
As shown in Fig. 5A
and B, compared with that of the control group, the activity of
SOD in PC12 and hippocampal cells of the model group was decreased
(P<0.05 or P<0.01, respectively), while the content of MDA
was markedly increased (P<0.05 or P<0.01, respectively).
Compared with those of the model group, the miR-130b mimic and
si-PTEN groups displayed a reduced content of MDA (P<0.05 or
P<0.01, respectively) and an increased SOD activity (P<0.05
or P<0.01, respectively), while the miR-130b inhibitor group
exhibited an increased MDA content (P<0.05) and a decreased
activity of SOD (P<0.05). Compared with those of the miR-130b
inhibitor group, the miR-130b inhibitor + si-PTEN group displayed
an increased SOD activity (P<0.05 or P<0.01, respectively)
and a reduced content of MDA (P<0.01). These results indicated
that miR-130b regulated PTEN and promoted the antioxidant capacity
of PC12 and hippocampal cells in an elevated glucose
environment.
miR-130b regulates PTEN expression to
activate Akt and inhibit the mitochondria-mediated apoptosis
pathway in PC12 and hippocampal cells in a high glucose
environment
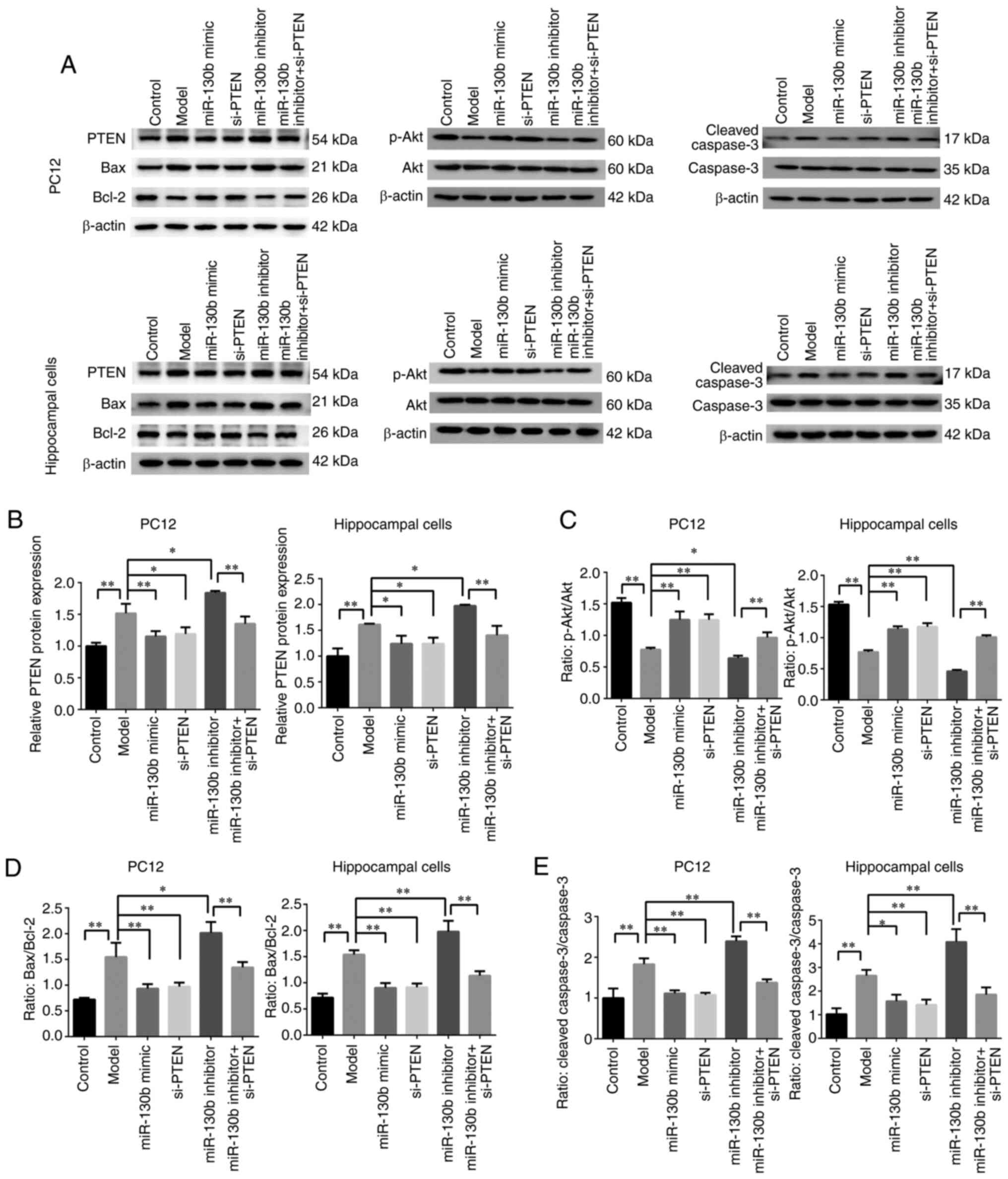
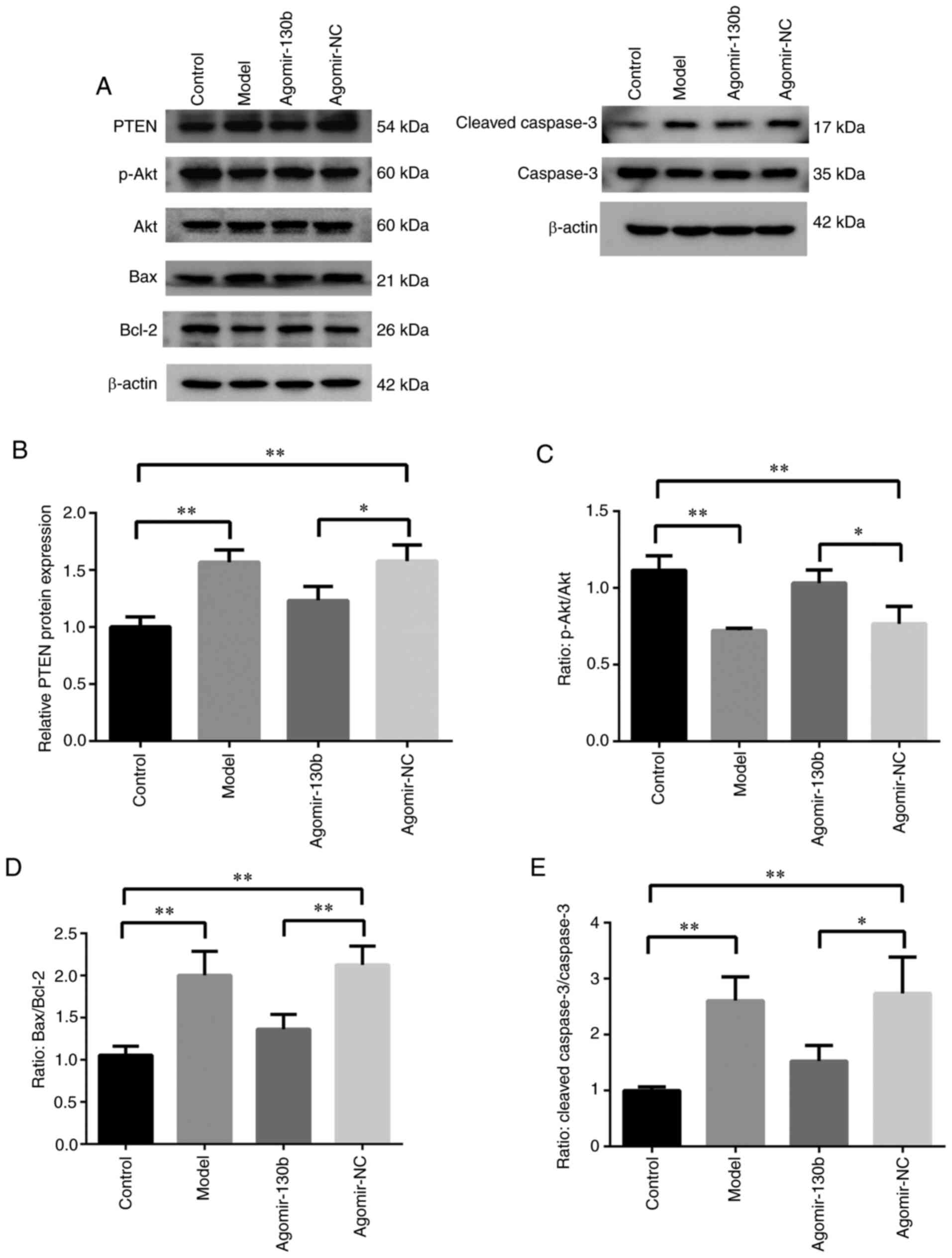
The results of western blot analysis are illustrated
in Fig. 6A. Compared with that
of the control group, the expression of PTEN was significantly
increased (P<0.01; Fig. 6B),
while the ratio of p-Akt/Akt was decreased in the model group
(P<0.01; Fig. 6C). Compared
with that of the model group, the expression of PTEN was
significantly decreased (P<0.05 or P<0.01, respectively;
Fig. 6B), and the ratio of
p-Akt/Akt was increased (P<0.01; Fig. 6C) in the miR-130b mimic and
si-PTEN groups, while the expression of PTEN was increased
(P<0.05 or P<0.01, respectively; Fig. 6B) and the ratio of p-Akt/Akt was
significantly reduced (P<0.05 or P<0.01, respectively;
Fig. 6C) in the miR-130b
inhibitor group. Compared with that of the miR-130b inhibitor
group, the expression of PTEN was decreased (P<0.01; Fig. 6B) and the ratio of p-Akt/Akt was
increased in the miR-130b inhibitor + si-PTEN group (P<0.01;
Fig. 6C). Furthermore, the
expression of mitochondria-mediated apoptosis pathway-related
proteins was detected. Compared with that of the control group, the
ratios of Bax/Bcl-2 and cleaved caspase-3/caspase-3 were
significantly increased in the model group (P<0.01; Fig. 6D and E). Compared with that of
the model group, the ratios of Bax/Bcl-2 and cleaved
caspase-3/caspase-3 were decreased in the miR-130b mimic and
si-PTEN groups (P<0.01 or P<0.05, respectively; Fig. 6D and E), while the ratios of
Bax/Bcl-2 and cleaved caspase-3/caspase-3 were significantly
increased in the miR-130b inhibitor group (P<0.01 or P<0.05,
respectively; Fig. 6D and E).
Compared with that of the miR-130b inhibitor group, the ratios of
Bax/Bcl-2 and cleaved caspase-3/caspase-3 were decreased in the
miR-130b inhibitor + si-PTEN group (P<0.01; Fig. 6D and E). These results indicated
that the activity of Akt was inhibited and the
mitochondria-mediated apoptosis pathway was promoted in PC12 and
hippocampal cells under a high glucose environment, and that
miR-130b could inhibit PTEN to reverse the high glucose-induced
decreased Akt activity and mitochondria-mediated apoptosis.
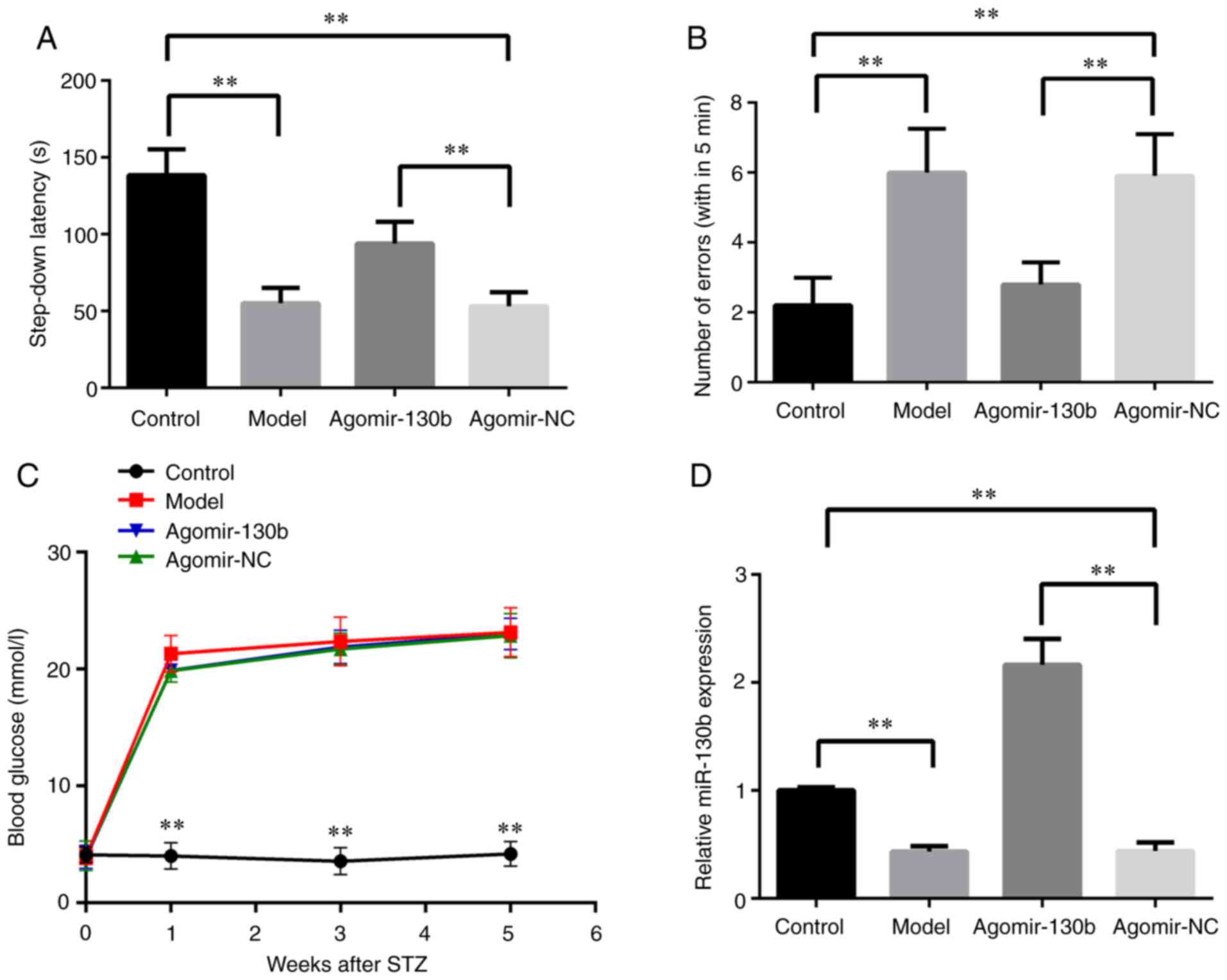
miR-130b enhances the learning ability of
rats with DE
The step-down passive avoidance test was performed
to detect the learning ability of rats with DE following miR-130b
treatment. As shown in Fig. 7A and
B, compared with those of the control group, the model and
agomir-NC groups showed decreased latency time and increased error
numbers. Compared with the findings in the agomir-NC group, the
step-down latency time was increased, while the number of errors
was reduced, in the agomir-130b group (P<0.01). The present
study then examined the effect of miR-130b on the blood glucose
content of rats with DE. As shown in Fig. 7C, compared with that of the
control group, the content of blood glucose was increased in the
model, agomir-NC and agomir-130b groups (P<0.01). In addition,
the expression of miR-130b in the hippocampus of rats was detected.
As shown in Fig. 7D, compared
with that of the control group, the model and agomir-NC groups
exhibited a decreased miR-130b expression (P<0.01). Compared
with that of the agomir-NC group, the expression of miR-130b was
increased in the agomir-130b group (P<0.01). These results
indicated that miR-130b promoted the learning ability, whereas it
did not affect the content of blood glucose of rats with DE.
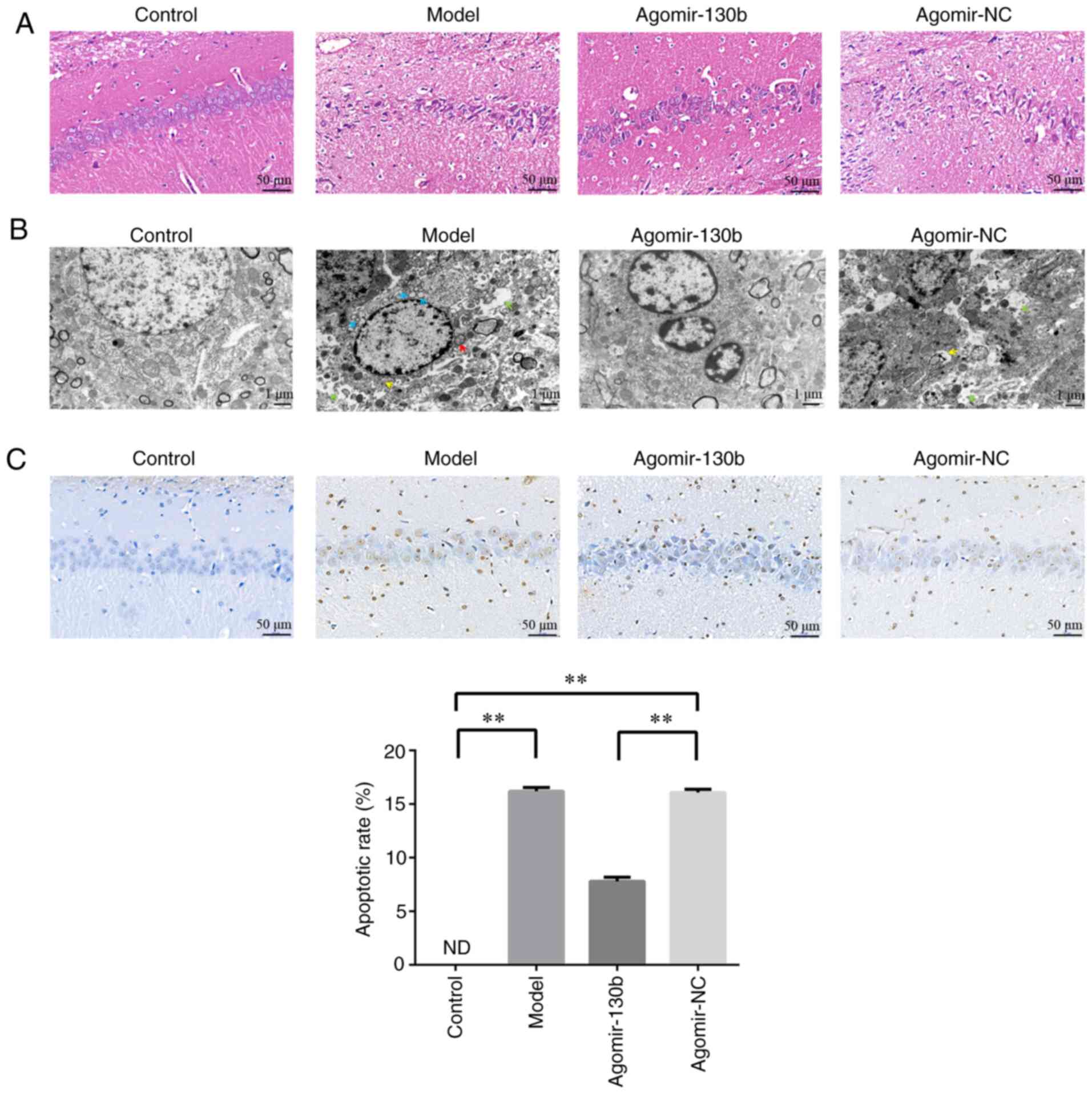
miR-130b attenuates hippocampus neuronal
damage in rats with DE
Pathological changes in the hippocampus were
detected using H&E staining (Fig. 8A). Compared with those of the
control and agomir-130b groups, the model and agomir-NC groups
exhibited a substantially reduced numer of pyramidal cells, and the
nucleus was solidified and exhibited increased positive staining.
The ultrastructural damage to the hippocampus was observed using
TEM (Fig. 8B). The control and
agomir-130 groups exhibited a clear nuclear membrane, obvious
nucleolus, abundant organelles and a complete synaptic structure.
The model and agomir-NC groups exhibited an irregular nucleolus,
mitochondrial swelling, rough endoplasmic reticulum dilatation,
autophagy and a damaged synaptic structure. Apoptotic cells were
detected using TUNEL staining (Fig.
8C). Compared with that of the control group, the apoptotic
rate of the model and agomir-NC groups was significantly increased
(P<0.01). Compared with that of the agomir-NC group, the
apoptotic rate of the agomir-130b group was significantly reduced
(P<0.01). These results demonstrated that miR-130b attenuated
damage to hippocampal neurons and the apoptosis of hippocampal
cells in rats with DE.
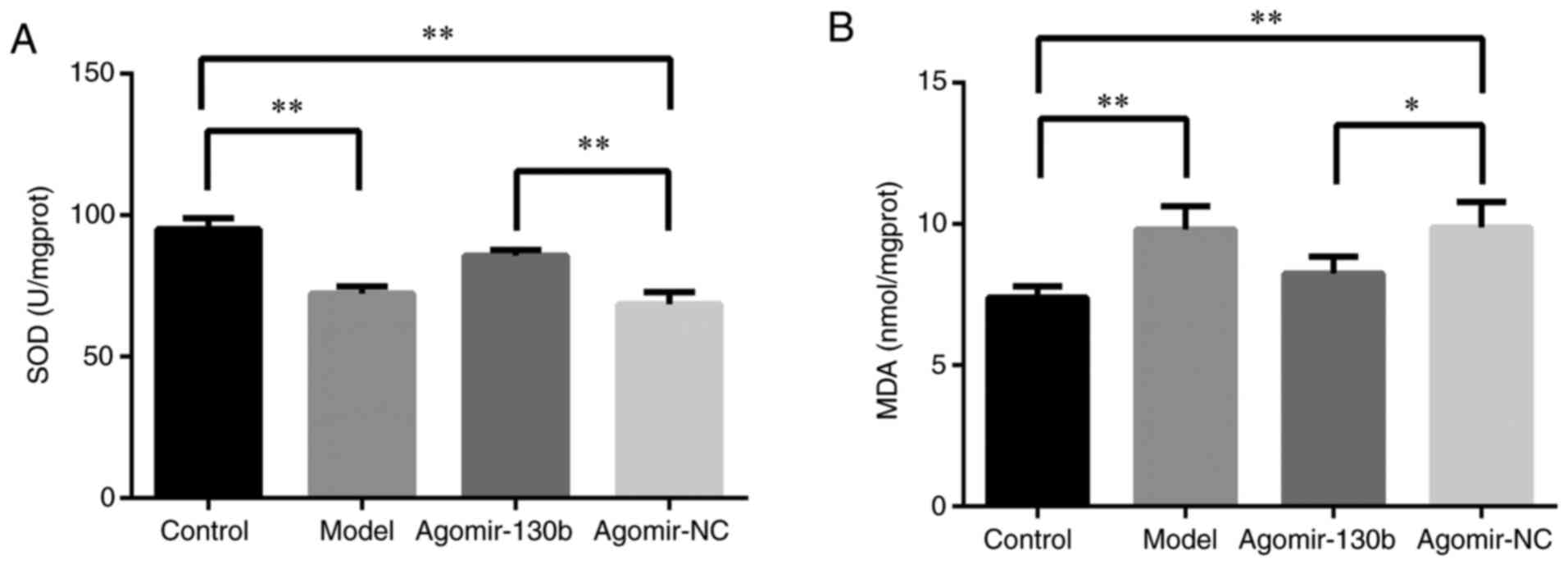
miR-130b improves oxidative stress in the
hippocampus of rats with DE
As shown in Fig. 9A
and B, compared with that of the control group, the activity of
SOD in the hippocampus of the model and agomir-NC groups was
markedly decreased, while the content of MDA was increased
(P<0.01). Compared with those of the agomir-NC group, the
agomir-130 group showed increased activity of SOD (P<0.01) and
decreased content of MDA (P<0.05). These data indicated that
miR-130b may improve oxidative stress in the hippocampus of rats
with DE.
miR-130b activates Akt protein and
inhibits the mitochondria-mediated apoptosis pathway in the
hippocampus of rats with DE
The results of western blot analysis are presented
in Fig. 10A. Compared with that
of the control group, the expression of PTEN in the model and
agomir-NC groups was markedly increased, while the ratio of
p-Akt/Akt was reduced (P<0.01). Compared with that of the
agomir-NC group, the expression of PTEN was significantly increased
(P<0.05; Fig. 10B) and the
ratio of p-Akt/Akt was decreased in the agomir-130b group
(P<0.05; Fig. 10C). In
addition, compared with that of the control group, the ratios of
Bax/Bcl-2 and cleaved caspase-3/caspase-3 were significantly
increased in the model and agomir-NC groups (P<0.01). Compared
with that of the agomir-NC group, the ratios of Bax/Bcl-2 and
cleaved caspase-3/caspase-3 were decreased in the agomir-130b group
(P<0.01 or P<0.05, respectively; Fig. 10D and E). These results
demonstrated that miR-130b regulated PTEN expression to activate
Akt and inhibit the mitochondria-mediated apoptosis pathway in
vivo.
Discussion
DE is a chronic complication of diabetes mellitus
that damages the central nervous system and causes cognitive
impairment (29). A high glucose
environment can lead to the accumulation of advanced glycation end
products, which can induce oxidative stress and subsequent damage
to hippocampal neurons (30).
The present study aimed to examine the effects of miR-130b on
oxidative stress injury in rats with DE. The present data
demonstrated that miR-130b could activate Akt by inhibiting PTEN,
thereby improving oxidative stress damage in rats with DE.
Previous studies have demonstrated that miR-130b is
an oncogene, which has been found to promote cancer cell
proliferation, invasion and metastasis in hepatocellular carcinoma
(31), breast cancer (32), gastric cancer (33), esophageal cancer (16) and glioma (34). A previous study found that
miR-130b reduced fat deposition in C57BL/6 mice, reversed glucose
tolerance and improved high-fat diet-induced obesity (35). In addition, miR-130b has been
shown to attenuate kidney fibrosis and damage induced by diabetes
(36). The results of the
present study indicated that the expression level of miR-130b was
reduced in PC12 and hippocampal cells stimulated with high glucose.
The overexpression of miR-130b reduced the high glucose-induced
apoptosis of PC12 and hippocampal cells, and improved cell
viability, while miR-130b inhibitor increased apoptosis and reduced
cell viability.
PTEN is the target gene of miR-130b in various
cancer cells, and miR-130b promotes the development of cancer by
regulating the expression of PTEN (14,37–39). PTEN can inhibit the PI3K/Akt
signaling pathway, which can attenuate oxidative stress-induced
damage by activating downstream antioxidant proteins (40). Therefore, it was hypothesized
that miR-130b could attenuate high glucose-induced oxidative stress
damage by inhibiting PTEN and activating Akt. SOD is an antioxidant
enzyme involved in free radical scavenging, while MDA is a product
of lipid peroxidation, and its content can reflect the level of
free radicals. These two indicators are commonly used to assess the
level of oxidative stress (41).
The present study also confirmed that PTEN is the target gene of
miR-130b, and showed that PTEN was involved in the high
glucose-induced oxidative stress injuries of PC12 and hippocampal
cells. The downregulation of the expression of PTEN may improve
cell viability and inhibit cell apoptosis by increasing SOD
activity and reducing MDA content, indicating that miR-130b can
regulate PTEN to ameliorate oxidative stress-related cell
injury.
When cells are subjected to oxidative stress, the
metabolic and functional characteristics of mitochondria change,
and the intrinsic apoptotic pathway mediated by mitochondria is
initiated (42). The Bcl-2
family is a key regulatory protein family in the mitochondrial
apoptosis pathway. This family includes the anti-apoptotic protein,
Bcl-2, and the pro-apoptotic protein, Bax. The Bax/Bcl-2 ratio has
been used as a marker to represent the effect of apoptosis
(43). Caspase-3 is one of the
members of the caspase protein family that mediates apoptosis.
Caspase-3 is cleaved into cleaved caspase-3 and then activated to
cause apoptosis (44). The data
of the present study demonstrated that miR-130b regulated PTEN to
increase the p-Akt/Akt ratio, and reduce the Bax/Bcl-2 and the
cleaved caspase-3/caspase-3 ratios in PC12 cells hippocampal cells,
suggesting that miR-130b could prevent cell apoptosis.
The present study established a rat model of DE to
further evaluate the effects of miR-130b. The expression level of
miR-130b was downregulated in the hippocampal tissue of rats with
DE, and the overexpression of miR-130b attenuated cognitive
impairment and hippocampal damage in DE model rats. The
overexpression of miR-130b in vivo increased SOD activity
and the p-Akt/Akt ratio, and reduced cell apoptosis, the MDA
content, the ratio of Bax to Bcl-2 and the expression of cleaved
caspase-3 in the hippocampus of rats with DE. This suggests that
miR-130b may attenuate cognitive impairment by protecting the
hippocampus from oxidative stress-induced damage in rats with
DE.
In conclusion, the results of the present study
demonstrated that miR-130b improved oxidative stress-induced injury
in rats with DE and in high glucose-induced PC12 and hippocampal
cells by inhibiting PTEN and activating the PI3K/Akt pathway. The
role of PTEN in DE remains unclear. Further research is thus
required to focus on the role of PTEN in DE and other cognitive
disorders. In addition, the present study only partially discussed
the role of PTEN as a target gene of miR-130b in the treatment of
DE. However, miR-130b may also target other genes to protect nerve
cells, which warrants investigation in future studies.
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL, MY, HL and JL designed and performed the
experiments. MY and RX collected and analyzed the data. YL wrote
the manuscript. HL, RX and JL assisted in the drafting of the
manuscript. YL, RX and MY confirm the authenticity of all the raw
data. All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The study protocol was approved by the Animal
Experimental Ethics Committee of Henan Provincial People's Hospital
(approval no. 2020408A).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by the Natural Science
Foundation of China (grant no. 81372617).
References
|
1
|
Sima AA and Li ZG: The effect of C-peptide
on cognitive dysfunction and hippocampal apoptosis in type 1
diabetic rats. Diabetes. 54:1497–1505. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tirassa P, Maccarone M, Florenzano F,
Cartolano S and De Nicolò S: Vascular and neuronal protection
induced by the ocular administration of nerve growth factor in
diabetic-induced rat encephalopathy. CNS Neurosci Ther. 19:307–318.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Samarghandian S, Azimi-Nezhad M and Samini
F: Ameliorative effect of saffron aqueous extract on hyperglycemia,
hyperlipidemia, and oxidative stress on diabetic encephalopathy in
streptozotocin induced experimental diabetes mellitus. Biomed Res
Int. 2014:9208572014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Reinhard H, Garde E, Skimminge A, Åkeson
P, Ramsøy TZ, Winther K, Parving HH, Rossing P and Jacobsen PK:
Plasma NT-proBNP and white matter hyperintensities in type 2
diabetic patients. Cardiovasc Diabetol. 11:1192012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sima AA: Encephalopathies: The emerging
diabetic complications. Acta Diabetol. 47:279–293. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shi B, Wang Y, Zhao R, Long X, Deng W and
Wang Z: Bone marrow mesenchymal stem cell-derived exosomal miR-21
protects C-kit+ cardiac stem cells from oxidative injury through
the PTEN/PI3K/Akt axis. PLoS One. 13:e01916162018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nandi SS, Zheng H, Sharma NM, Shahshahan
HR, Patel KP and Mishra PK: Lack of miR-133a decreases
contractility of diabetic hearts: A role for novel cross talk
between tyrosine aminotransferase and tyrosine hydroxylase.
Diabetes. 65:3075–3090. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kölling M, Kaucsar T, Schauerte C, Hübner
A, Dettling A, Park JK, Busch M, Wulff X, Meier M, Scherf K, et al:
Therapeutic miR-21 silencing ameliorates diabetic kidney disease in
mice. Mol Ther. 25:165–180. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Y, Song C, Liu J, Bi Y and Li H:
Inhibition of miR-25 aggravates diabetic peripheral neuropathy.
Neuroreport. 29:945–953. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sun Z, Ma Y, Chen F, Wang S, Chen B and
Shi J: miR-133b and miR-199b knockdown attenuate TGF-β1-induced
epithelial to mesenchymal transition and renal fibrosis by
targeting SIRT1 in diabetic nephropathy. Eur J Pharmacol.
837:96–104. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bai X, Geng J, Zhou Z, Tian J and Li X:
MicroRNA-130b improves renal tubulointerstitial fibrosis via
repression of Snail-induced epithelial-mesenchymal transition in
diabetic nephropathy. Sci Rep. 6:204752016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lv C, Zhou YH, Wu C, Shao Y, Lu CL and
Wang QY: The changes in miR-130b levels in human serum and the
correlation with the severity of diabetic nephropathy. Diabetes
Metab Res Rev. 31:717–724. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yuan Y, Peng W, Liu Y and Xu Z:
Circulating miR-130 and its target PPAR-γ may be potential
biomarkers in patients of coronary artery disease with type 2
diabetes mellitus. Mol Genet Genomic Med. 7:e9092019. View Article : Google Scholar
|
|
14
|
Sekino Y, Sakamoto N, Sentani K, Oue N,
Teishima J, Matsubara A and Yasui W: miR-130b promotes sunitinib
resistance through regulation of PTEN in renal cell carcinoma.
Oncology. 97:164–172. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yuan B, Zou M, Zhao Y, Zhang K, Sun Y and
Peng X: Up-regulation of miR-130b-3p activates the
PTEN/PI3K/AKT/NF-κB pathway to defense against mycoplasma
gallisepticum (HS strain) infection of chicken. Int J Mol Sci.
19:21722018. View Article : Google Scholar
|
|
16
|
Yu T, Cao R, Li S, Fu M, Ren L, Chen W,
Zhu H, Zhan Q and Shi R: MiR-130b plays an oncogenic role by
repressing PTEN expression in esophageal squamous cell carcinoma
cells. BMC Cancer. 15:292015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ciuffreda L, Falcone I, Incani UC, Del
Curatolo A, Conciatori F, Matteoni S, Vari S, Vaccaro V, Cognetti F
and Milella M: PTEN expression and function in adult cancer stem
cells and prospects for therapeutic targeting. Adv Biol Regul.
56:66–80. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Worby CA and Dixon JE: PTEN. Annu Rev
Biochem. 83:641–669. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pompura SL and Dominguez-Villar M: The
PI3K/AKT signaling pathway in regulatory T-cell development,
stability, and function. J Leukoc Biol. Jan 22–2018.Online ahead of
print. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang M, Hu R, Wang Y, Liu L, You H, Zhang
J, Wu X, Pei T, Wang F, Lu L, et al: Atractylenolide III attenuates
muscle wasting in chronic kidney disease via the oxidative
stress-mediated PI3K/AKT/mTOR pathway. Oxid Med Cell Longev.
2019:18754712019.PubMed/NCBI
|
|
21
|
Zhang B, Zhao Z, Meng X, Chen H, Fu G and
Xie K: Hydrogen ameliorates oxidative stress via PI3K-Akt signaling
pathway in UVB-induced HaCaT cells. Int J Mol Med. 41:3653–3661.
2018.PubMed/NCBI
|
|
22
|
Wen Z, Hou W, Wu W, Zhao Y, Dong X, Bai X,
Peng L and Song L: 6′-O-galloylpaeoniflorin attenuates cerebral
ischemia reperfusion-induced neuroinflammation and oxidative stress
via PI3K/Akt/Nrf2 activation. Oxid Med Cell Longev.
2018:86782672018. View Article : Google Scholar
|
|
23
|
Mohamed AK, Bierhaus A, Schiekofer S,
Tritschler H, Ziegler R and Nawroth PP: The role of oxidative
stress and NF-kappaB activation in late diabetic complications.
Biofactors. 10:157–167. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen M, Zheng H, Wei T, Wang D, Xia H,
Zhao L, Ji J and Gao H: High glucose-induced PC12 cell death by
increasing glutamate production and decreasing methyl group
metabolism. Biomed Res Int. 2016:41257312016.PubMed/NCBI
|
|
25
|
Mai H, Fan W, Wang Y, Cai Y, Li X, Chen F,
Chen X, Yang J, Tang P, Chen H, et al: Intranasal administration of
miR-146a agomir rescued the pathological process and cognitive
impairment in an AD mouse model. Mol Ther Nucleic Acids.
18:681–695. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
27
|
Kameyama T, Nabeshima T and Kozawa T:
Step-down-type passive avoidance- and escape-learning method.
Suitability for experimental amnesia models. J Pharmacol Methods.
16:39–52. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Luo Y, Kuang S, Xue L and Yang J: The
mechanism of 5-lipoxygenase in the impairment of learning and
memory in rats subjected to chronic unpredictable mild stress.
Physiol Behav. 167:145–153. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Díaz-Gerevini GT, Daín A, Pasqualini ME,
López CB, Eynard AR and Repossi G: Diabetic encephalopathy:
Beneficial effects of supplementation with fatty acids ω3 and
nordihydroguaiaretic acid in a spontaneous diabetes rat model.
Lipids Health Dis. 18:432019. View Article : Google Scholar
|
|
30
|
Yaffe K, Lindquist K, Schwartz AV,
Vitartas C, Vittinghoff E, Satterfield S, Simonsick EM, Launer L,
Rosano C, Cauley JA and Harris T: Advanced glycation end product
level, diabetes, and accelerated cognitive aging. Neurology.
77:1351–1356. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tu K, Zheng X, Dou C, Li C, Yang W, Yao Y
and Liu Q: MicroRNA-130b promotes cell aggressiveness by inhibiting
peroxisome proliferator-activated receptor gamma in human
hepatocellular carcinoma. Int J Mol Sci. 15:20486–20499. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Miao Y, Zheng W, Li N, Su Z, Zhao L, Zhou
H and Jia L: MicroRNA-130b targets PTEN to mediate drug resistance
and proliferation of breast cancer cells via the PI3K/Akt signaling
pathway. Sci Rep. 7:419422017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lai KW, Koh KX, Loh M, Tada K, Subramaniam
MM, Lim XY, Vaithilingam A, Salto-Tellez M, Iacopetta B, Ito Y, et
al: MicroRNA-130b regulates the tumour suppressor RUNX3 in gastric
cancer. Eur J Cancer. 46:1456–1463. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tong L, Chu M, Yan B, Zhao W, Liu S, Wei
W, Lou H, Zhang S, Ma S, Xu J and Wei L: MTDH promotes glioma
invasion through regulating miR-130b-ceRNAs. Oncotarget.
8:17738–17749. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pan S, Yang X, Jia Y, Li Y, Chen R, Wang
M, Cai D and Zhao R: Intravenous injection of microvesicle-delivery
miR-130b alleviates high-fat diet-induced obesity in C57BL/6 mice
through translational repression of PPAR-γ. J Biomed Sci.
22:862015. View Article : Google Scholar
|
|
36
|
Liu Y, Yang Y, Wang Q, Kahaer A, Zhang J,
Liao J, Abudureyimu M, Yahefu R, Qi J, Zhao L and Zhu J: Regulatory
effect of 1,25(OH)2D3 on TGF-β1 and miR-130b expression in
streptozotocin-induced diabetic nephropathy in rats. Int J
Endocrinol. 2019:12313462019. View Article : Google Scholar
|
|
37
|
Zhang Q, Zhang B, Sun L, Yan Q, Zhang Y,
Zhang Z, Su Y and Wang C: MicroRNA-130b targets PTEN to induce
resistance to cisplatin in lung cancer cells by activating
Wnt/β-catenin pathway. Cell Biochem Funct. 36:194–202. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chang RM, Xu JF, Fang F, Yang H and Yang
LY: MicroRNA-130b promotes proliferation and EMT-induced metastasis
via PTEN/p-AKT/HIF-1α signaling. Tumour Biol. 37:10609–10619. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Egawa H, Jingushi K, Hirono T, Ueda Y,
Kitae K, Nakata W, Fujita K, Uemura M, Nonomura N and Tsujikawa K:
The miR-130 family promotes cell migration and invasion in bladder
cancer through FAK and Akt phosphorylation by regulating PTEN. Sci
Rep. 6:205742016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lee YJ, Jeong HY, Kim YB, Lee YJ, Won SY,
Shim JH, Cho MK, Nam HS and Lee SH: Reactive oxygen species and
PI3K/Akt signaling play key roles in the induction of Nrf2-driven
heme oxygenase-1 expression in sulforaphane-treated human
mesothelioma MSTO-211H cells. Food Chem Toxicol. 50:116–123. 2012.
View Article : Google Scholar
|
|
41
|
Wang Y, Wang Q, Li J, Lu G and Liu Z:
Glutamine improves oxidative stress through the Wnt3a/β-catenin
signaling pathway in Alzheimer's disease in vitro and in vivo.
Biomed Res Int. 2019:46902802019.
|
|
42
|
Askari H, Rajani SF, Poorebrahim M,
Haghi-Aminjan H, Raeis-Abdollahi E and Abdollahi M: A glance at the
therapeutic potential of irisin against diseases involving
inflammation, oxidative stress, and apoptosis: An introductory
review. Pharmacol Res. 129:44–55. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Del Principe MI, Dal Bo M, Bittolo T,
Buccisano F, Rossi FM, Zucchetto A, Rossi D, Bomben R, Maurillo L,
Cefalo M, et al: Clinical significance of bax/bcl-2 ratio in
chronic lymphocytic leukemia. Haematologica. 101:77–85. 2016.
View Article : Google Scholar :
|
|
44
|
Li H, Yin A, Cheng Z, Feng M, Zhang H, Xu
J, Wang F and Qian L: Attenuation of Na/K-ATPase/Src/ROS
amplification signal pathway with pNaktide ameliorates myocardial
ischemia-reperfusion injury. Int J Biol Macromol. 118:1142–1148.
2018. View Article : Google Scholar : PubMed/NCBI
|