Introduction
Osteoporosis is the most prevalent systemic skeletal
system disease, leading to increased bone fragility and
vulnerability to fractures (1,2).
Due to the microarchitectural destruction in bone tissue, fracture
healing in osteoporotic patients is often delayed and compromised
compared with non-osteoporotic individuals (3,4).
Osteoporosis usually results from menopause, aging, metabolic
diseases and drug therapies with the precise cellular and molecular
mechanism remaining to be elucidated (2,5).
Currently, dysregulated bone turnover, particularly
the dysregulated differentiation of bone marrow-derived mesenchymal
stem cells (BMSCs) has been discovered as an important factor of
osteoporotic fractures (6,7).
In detail, the osteoblastic differentiation of BMSCs is impaired,
with a greater number of BMSCs being differentiated into adipocytes
in osteoporotic patients, leading to progressive dysregulated bone
turnover and bone loss (5,6).
Therefore, approaches for fracture healing based on the activation
of the osteogenic differentiation and the inhibition of the
adipogenic differentiation of BMSCs are innovative and appealing
(8,9).
The nuclear factor of activated T-cells (NFAT) is a
substrate of the Ca2+-dependent transcription factor
family, which has been shown to regulate cell differentiation and
organ development, podocyte injury and cell apoptosis. In addition,
NFAT signaling has been found to play critical roles in bone
metabolism, such as in the process of osteoblast and osteoclast
formation (10). In detail, the
activation of the NFAT signaling pathway has been shown to lead to
the inhibition of osteoprogenitor cell formation (11). By contrast, the blocking of NFAT
signaling will contribute to the osteoblastic differentiation of
MSCs (11,12), which suggests that NFAT is a
potential target for the treatment of osteoporotic fractures.
Peptide 11R-VIVIT is a NFAT-specific inhibitor without affecting
upstream calcineurin (Cn). Due to its high specificity against
NFAT2, peptide 11R-VIVIT has been widely examined and few adverse
effects have been reported (13-15). A Previous study demonstrated that
11R-VIVIT can stimulate bone formation by decreasing NFATc1
expression and regulating the expression of inflammation-related
molecules (10). However,
whether peptide 11R-VIVIT can affect the healing process of
osteoporotic fractures via the downregulation of NFATc1 remains
unknown.
The present study aimed to elucidate the effects and
underlying molecular mechanisms of peptide 11R-VIVIT in fracture
healing. The results indicated that 11R-VIVIT promoted autophagy to
enhance the osteogenic differentiation of osteoporotic BMSCs to
promote fracture healing in osteoporotic rats through the protein
kinase B (AKT)/NFATc1 signaling pathway, which may provide new
insight for the BMSC-based treatment of osteoporosis in the
future.
Materials and methods
Animals and ethics statement
Following the Animal Experimentation Ethics
Guidelines, the Ethics Committee of the Affiliated Hangzhou First
People's Hospital approved all the procedures in the study (no.
ZJCLA-IACUC-20080010). A total of 24 Sprague-Dawley female rats (12
weeks old, weighing 200-240 g) were purchased from the Beijing
Weitong Lihua Experimental Animal Technology Co. Ltd. (Beijing,
China). Then, the rats were housed in a specific pathogen-free
(SPF) facility (temperature, 22±2°C; humidity, 50±10%) with a
12/12-h light/dark cycle, and standard laboratory animal chow and
tap water were available ad libitum. To assess the effects
of peptide 11R-VIVIT on osteoporotic fracture healing, a rat model
of osteoporosis [model of ovariectomy (OVX)-induced osteoporosis]
was established at first by removing the ovaries bilaterally
(16). At 12 weeks after the OVX
surgery, a unilateral open femur fracture model was established in
all rats by an experienced operator to reduce pain in the animals.
Briefly, an osteotomy fracture was established using an oscillating
sagittal saw the middle of the left femur and 25-G needles were
inserted intramedullary for fixation. A total of four groups (n=6
for each group) were included in the study as follows: i) A group
of whole-stage saline treatment (Saline/Saline group, saline
treatment only during ovariectomy and sacrifice); ii) a group of
late-stage VIVIT (intraperitoneal injection, 100 µg/kg,
Tocris Bioscience) treatment (Saline/VIVIT group, saline treatment
following OVX and 11R-VIVIT treatment following OVX); iii) a group
of early-stage 11R-VIVIT following saline treatment (VIVIT/Saline
group, 11R-VIVIT treatment following OVX and saline treatment
following OVX); and iv) a group of whole-stage 11R-VIVIT treatment
(VIVIT/VIVIT group, 11R-VIVIT treatment only during OVX and
sacrifice). Food and water were provided without limits to all rats
during the fracture healing process. All animals were anesthetized
by an intraperitoneal injection (50 mg/kg) of 1% sodium
pentobarbital during the surgeries. The rats were then euthanized
by cervical dislocation under anesthesia. The femurs were collected
for X-ray, micro-CT and histological analyses. All wounds healed
well and no death or adverse events were observed at the
experimental endpoint.
X-ray and micro-CT evaluation and
histological staining
Femurs were fixed in 4% paraformaldehyde for 24 h
and then scanned with an X-ray scanner or a micro-CT system. The
region of interest was defined as 2 mm above and below the
fracture. All the slices derived from the micro-CT scanning were
collected for 3D reconstruction. The bone formation area, including
callus and cortex bone around the fracture line was selected to
measure fracture healing. Following X-ray and micro-CT evaluation,
the femurs were decalcified using EDTA, sliced into
4-µm-thick sections and stained with hematoxylin and eosin
(Sigma-Aldrich; Merck KGaA) at room temperature for 30 min.
Histological images were observed under a microscope (XSP-C204,
China Investment Corporation).
Isolation of rat BMSCs
The BMSCs were isolated and obtained by referring to
a previous study (17). In
brief, after the rats were anesthetized, the rat tibia was
separated and obtained. The ends of the rat tibia were cut with
scissors, and the tibia bone marrow contents were flushed and
washed with phosphate-buffered saline (PBS) supplemented with 5%
fetal bovine serum (FBS). Thereafter, the bone marrow contents were
centrifuged at 450 × g for 10 min at room temperature and the cell
pellet was re-suspended, which was then layered using the same
volume of Ficoll-Paque (inno-train Diagnostik GmbH) and centrifuged
at 850 g for 25 min at 4°C. Subsequently, BMSCs was collected and
re-suspended in α-MEM containing 10% FBS and 1%
penicillin-streptomycin (all from HyClone; Cytiva). Cell cultures
were maintained in a 37°C incubator with 5% CO2, and the
medium was replaced twice weekly in the present study. Cells at
passage 3-6 were utilized throughout the present study.
Characterization of BMSCs and
multi-lineage differentiation
Flow cytometric analysis for the characterization of
BMSCs was conducted by referring to the method described in
previous studies (18,19). Briefly, 1×106 BMSCs
were incubated with (FITC or PE)-conjugated antibody CD45 (cat. no.
559135, BD Biosciences), CD11b (cat. no. 554982, BD Biosciences),
CD73 (cat. no. 11-0739-42, Invitrogen; Thermo Fisher Scientific,
Inc.) and CD90 (cat. no. 554898, BD Biosciences) (1
µg/106 cells) in washing buffer for 30 min at
4°C. All fluorescently-labeled antibodies used in flow cytometric
analysis were purchased from BD Biosciences. Thereafter, both the
stained and unstained cells were washed, and their fluorescence
intensity was quantified using a FACSCalibur flow cytometer (BD
Biosciences). FlowJo software (Ver. 6.2) was utilized for data
analysis.
BMSCs were pre-treated with 0.1, 1 and 10 µM
11R-VIVIT, 10 µM LY294002 (Sigma-Aldrich; Merck KGaA) and
200 nM rapamycin (Beijing Solarbio Science & Technology Co.,
Ltd.) for 1 h.
To detect the osteogenic ability, BMSCs were
inoculated at approximately 1×104 cells/cm2
in 24-well plates and induced in osteogenic induction medium (OIM;
cat. no. RASMX-90021, Cyagen Biosciences). Likewise, commercial
standard adipogenic induction medium (AIM; cat. no. RASMX-90031,
Cyagen Biosciences,) was employed for the induction of
adipogenesis, which revealed the adipogenic ability of the
BMSCs.
ALP staining and ALP activity assays were performed
using BMSCs that had been cultured in OIM for 7 days. For ALP
staining, BMSCs were washed with PBS, fixed in 4% paraformaldehyde
for 20 min at room temperature, and then stained with an ALP
staining kit (Nanjing Jiancheng Bioengineering Institute) for 30
min at room temperature in the dark. For ALP activity
quantification, osteogenically differentiated BMSCs were examined
using an EscAPeTM SEAP Chemiluminescence kit (Clontech
Laboratories, Inc.), the value of which was normalized to the total
cellular protein concentrations. Alizarin Red S staining and Oil
Red O staining were performed at day 14 following differentiation.
For Alizarin Red S staining, BMSCs were fixed with 4%
paraformaldehyde for 20 min, washed PBS, and then stained with
Alizarin Red (Cyagen Biosciences) for 5 min at room temperature.
Images obtained following staining were analyzed using ImageJ
software (ver. 1.6, Wayne Rasband, National Institutes of Health)
and the Alizarin Red S percentage of the positively stained area
was determined. Finally, Oil Red O staining was applied to
determine the adipogenic ability of the BMSCs. In brief, BMSCs were
fixed in 3.7% formaldehyde for 30 min, followed by staining with
Oil Red O (Sigma-Aldrich; Merck KGaA) for 30 min. All images were
acquired using a light microscope (Dmi8, Leica Microsystems
GmbH).
Reverse transcription and quantitative
polymerase chain reaction (RT-qPCR) and semi-quantitative PCR
Total RNA was extracted from the BMSCs with
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). Reverse transcription was carried out using the Script™
RT-PCR kit (Takara Bio, Inc.), followed by PCR using SYBR PreMix Ex
TaqTM (Takara Bio, Inc.) on the CFX96TM Real-time PCR Detection
System (Applied Biosystems). GAPDH mRNA was used as an internal
control. The reaction conditions were as follows: 95°C for 30 sec,
followed by 40 cycles of 94°C for 5 sec and 60°C for 35 sec. The
2−ΔΔCq method (20)
was used to determine the relative expression levels. The primers
for rat mRNAs are listed in Table
I. The amplified products were separated on 1% agarose gel
stained with ethidium bromide and visualized by UV light
 | Table ISequences of primers used for
PCR. |
Table I
Sequences of primers used for
PCR.
| Gene | Primer pair
sequence (5′-3′) |
|---|
| ALP |
AAGGCTTCTTCTTGCTGGTG |
|
GCCTTACCCTCATGATGTCC |
| OCN |
GGCAGCGAGGTAGTGAAGAG |
|
CTGGAGAGGAGCAGAACTGG |
| OPN |
CATCAGAGCCACGAGTTTCA |
|
TCAGGGCCCAAAACACTATC |
| PPARγ |
GGCTTCATGACAAGGGAGTTTC |
|
AACTCAAACTTGGGCTCCATAAAG |
| LPL |
CTGGACGGTAACAGGAATGTATGAG |
|
CATCAGGAGAAAGACGACTCGG |
| GAPDH |
TCGACAGTCAGCCGCATCTTCTTT |
|
GCCCAATACGACCAAATCCGTTGA |
Western blot analysis
Western blot analysis was carried out by referring
to the protocol described in a previous study (21). Total proteins were isolated from
the BMSCs using RIPA-lysis buffer (Sigma-Aldrich; Merck KGaA) on
ice, and protein concentrations were measured using a BCA protein
assay kit (Sigma-Aldrich; Merck KGaA). A total of 20 µg of
total protein were electrophoresed on 12% SDS-PAGE and transferred
to nitrocellulose membranes. The membranes were blocked using
non-fat milk for 1 h at room temperature and incubated overnight at
4°C with primary polyclonal antibodies including anti-LC3
(ab192890, 1:2,000), anti-p62 (ab109012, 1:10,000), anti-p-AKT
(ab38449, 1:1,000), anti-AKT (ab8805, 1:500), anti-p-glycogen
synthase kinase-3β (GSK-3β, ab131097, 1:1,000), anti-GSK-3β
(ab93926, 1:1,000), anti-NFATc1 (ab25916, 1:2,000), anti-Lamin B
(ab32532, 1:500), anti-Runt-related transcription factor 2 (Runx2;
ab76956, 1:2,000) (all from Abcam), anti-ALP (PA1004, 1:1,000,
Boster Biological Technology Co., Ltd.) and anti-GAPDH (ab8245,
1:1,000, Abcam). The proteins were then visualized following
incubation with goat anti-rat IgG (H&L) secondary antibody
(ab182018, 1:2,000, Abcam) for 60 min at 25°C, with extensively
washing with TBST and detecting using a chemiluminescence system
(Pierce; Thermo Fisher Scientific, Inc.). The intensity of protein
bands was measured by ImageJ 1.6 software and the signal intensity
of each band was normalized to its corresponding GAPDH or Lamin B
control.
CCK-8 assay
The viability of BMSCs was measured using a CCK-8
kit (Dojindo Molecular Technologies, Inc.). In detail, normal
BMSCs, osteoporotic BMSCs and osteoporotic BMSCs treated with
11R-VIVIT (10 µM) in 96-well plates were collected at the
time points of 0, 24, 48 and 72 h. CCK-8 kit solution (15
µl) were added to each well, and the cells were incubated at
37°C for an additional 3 h. The solution was finally removed, and
the absorbance at 450 nm was recorded using a microplate
spectrophotometer (Bio-Tek Instruments, Inc.).
Enzyme-linked immunosorbent assay
(ELISA)
The tumor necrosis factor (TNF)-α and interleukin
(IL)-1β concentrations of the normal BMSCs, osteoporotic BMSCs and
osteoporotic BMSCs treated with 11R-VIVIT (10 µM) were
measured by ELISA. The cells were cultured in 96-well plates for 3
days and the culture media were harvested and centrifuged at 1,200
× g for 10 min at room temperature. The TNF-α and IL-1β
concentrations in the supernatants were detected using rat TNF-α
(EK0526) and IL-1β (EK0393) ELISA kits (Boster Biological
Technology Co., Ltd.), according to the standard instructions. The
value at 450 nm was read using a microplate reader (SpectraMax 190,
Molecular Devices, LLC). The concentrations were calculated using
the standard curve.
Statistical analysis
All data were presented as the mean ± SD.
Statistical analyses were calculated using GraphPad Prism (version
8.0) software. The Student's t-test was adopted for two-sample
comparisons. One-way ANOVA followed by Tukey's post hoc analysis
were adopted for multiple comparisons. All data were determined by
at least three independent experiments. P<0.05 was considered to
indicate a statistically significant difference.
Results
11R-VIVIT accelerates fracture
healing
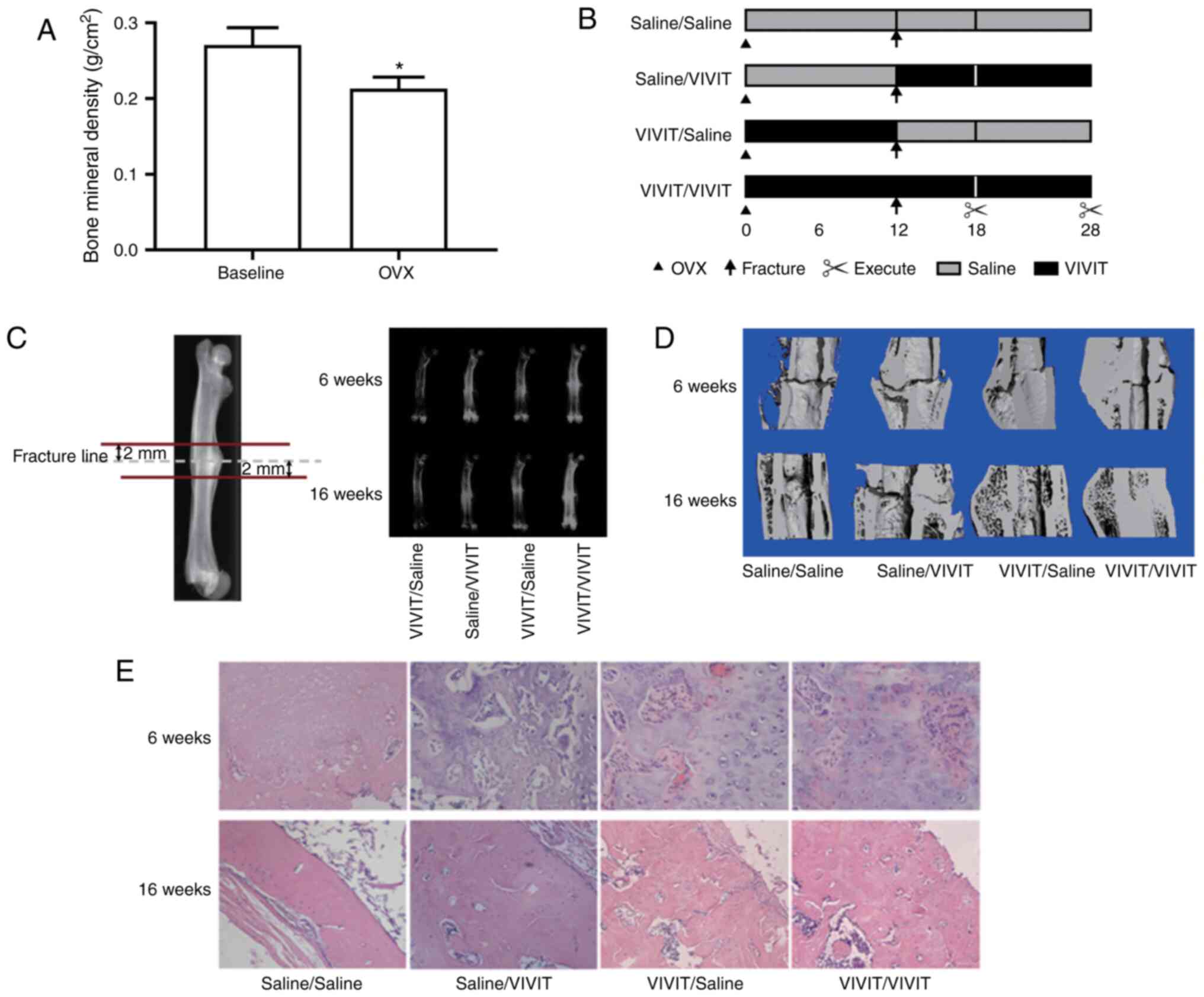
To determine whether the model of OVX was
successfully established, bone mineral density in the rats was
detected. The results revealed that rats subjected to OVX had a
lower bone mineral density than the rats at baseline (Fig. 1A). Thereafter, the effects of
11R-VIVIT on the osteoporotic fracture healing process were
examined (Fig. 1B). Micro-CT
scans results revealed that the rats treated with 11R-VIVIT had a
higher bone volume compared with those treated with saline
(Fig. 1C and D) in at 6 and 16
weeks after the fracture. The reconstruction results of micro-CT
analysis revealed that at 6- and 16-weeks post-fracture, the
11R-VIVIT-treated rats (particularly the rats from the VIVIT/VIVIT
group) exhibited smaller fracture calluses and fracture gaps. At 16
weeks post-fracture, all fracture gaps in the whole-stage
11R-VIVIT-treated rats had almost disappeared, while the other
groups exhibited obvious fracture gaps. H&E staining analysis
also revealed that whole-stage 11R-VIVIT treatment increased the
formation of bony connective junctions between the fracture gaps,
which also suggested that 11R-VIVIT accelerated bone remodeling
process in osteoporotic fracture healing (Fig. 1E).
11R-VIVIT increases the osteogenic
potential of osteoporotic BMSCs
As the osteogenic potential of BMSCs is a key factor
during osteoporotic fracture healing, the normal and osteoporotic
BMSCs were used to induce osteogenesis in vitro to determine
whether 11R-VIVIT would affect the bone formation ability of
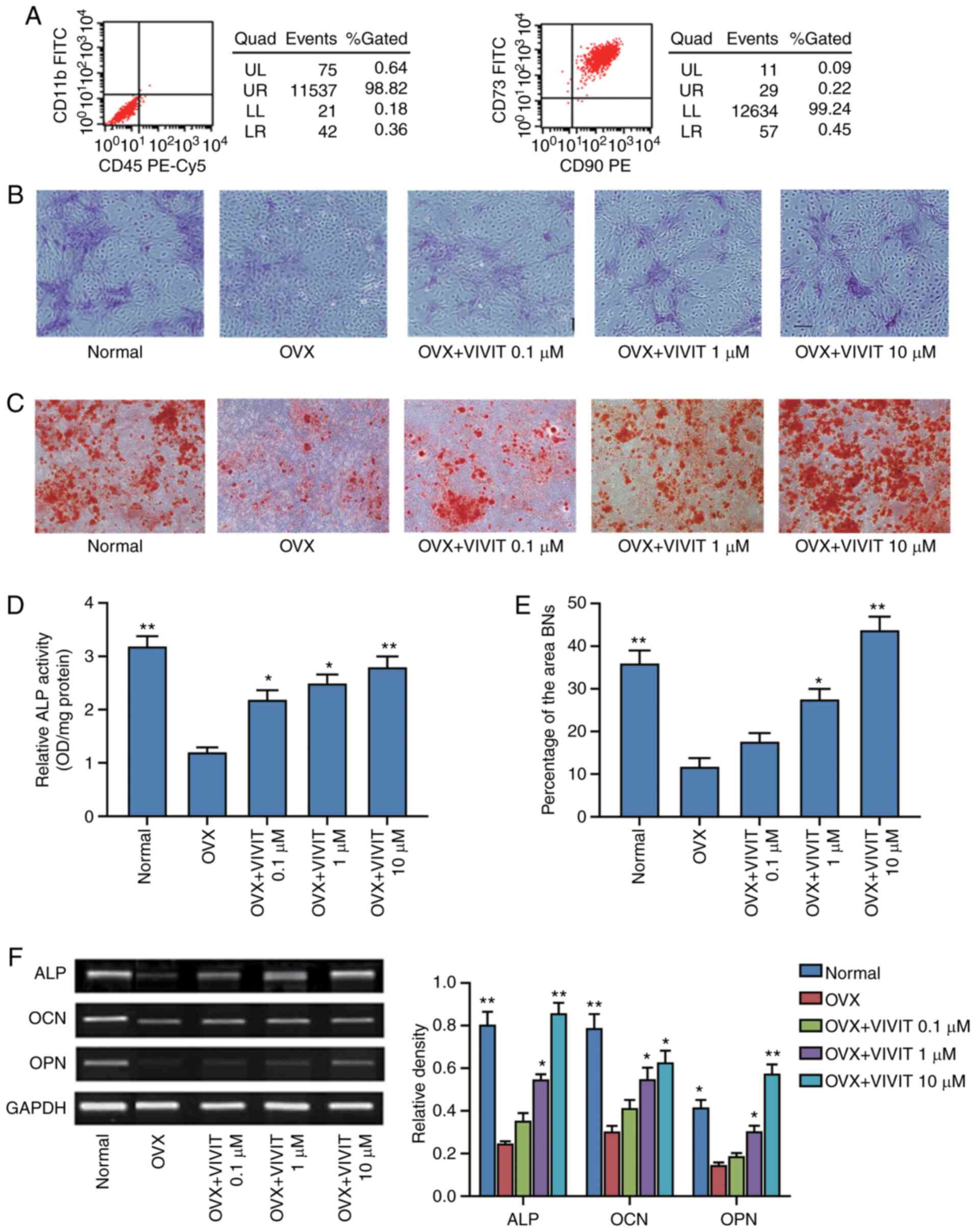
osteoporotic BMSCs. The results of flow cytometric analysis
revealed that the isolated cells were positive for CD73 and CD90,
whereas they were negative for CD11b and CD45, indicating that the
isolated cells were BMSCs (Fig.
2A). ALP staining, ALP activity assays and Alizarin Red
staining were then performed. The osteoporotic BMSCs treated with
11R-VIVIT exhibited more mineralized nodules in the extracellular
matrix than the cells from the OVX group (Fig. 2B and D). The results of Alizarin
Red S staining revealed that the BMSCs from the OVX group had less
calcium deposition than those from the normal group; treatment
with11R-VIVIT increased calcium deposition (Fig. 2C and E). The results of
semi-quantitative PCR revealed that the normal BMSCs expressed
higher levels of ALP, osteocalcin (OCN) and osteopontin (OPN) than
the osteoporotic BMSCs (Fig.
2F). The osteoporotic BMSCs were then treated with 11R-VIVIT,
at the concentration of 0.1-10 µM. ALP staining and Alizarin
Red S staining revealed that the ALP activity and the mineralized
nodules of the osteogenic BMSCs gradually increased in response to
the increasing 11R-VIVIT concentration in a concentration-dependent
manner. Semi-quantitative PCR revealed higher expression levels of
ALP, OCN and OPN mRNA in response to 11R-VIVIT treatment compared
with the OVX control group. Hence, 11R-VIVIT at a concentration of
10 µM was employed for further analyses in the present
study.
11R-VIVIT impairs the adipogenic
potential of osteoporotic BMSCs
As the adipogenic potential of BMSCs also plays
vital roles in osteoporotic fracture healing, the normal and
osteoporotic BMSCs were used to induce adipogenesis in vitro
to determine whether 11R-VIVIT can affect the adipogenic ability of
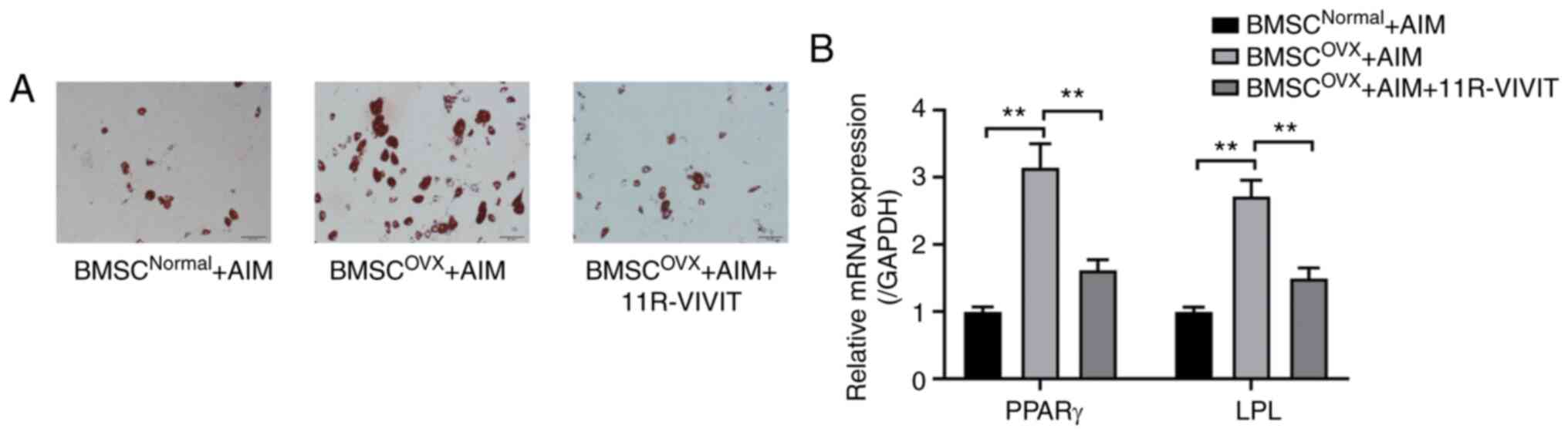
osteoporotic BMSCs. The results of RT-qPCR and Oil Red O staining
revealed that the osteoporotic BMSCs had a greater adipogenic
capacity than the normal BMSCs (Fig.
3). After the osteoporotic BMSCs were treated with 11R-VIVIT,
Oil Red O staining revealed that the adipogenic activity of the
osteoporotic BMSCs was inhibited (Fig. 3A). Likewise, the results of
RT-qPCR revealed lower mRNA expression levels of peroxisome
proliferator-activated receptor (PPAR)γ and lipoprotein lipase
(LPL) in response to 11R-VIVIT treatment compared with the OVX
control group (Fig. 3B).
11R-VIVIT attenuates the inflammatory
levels of osteoporotic BMSCs and increases cell viability
ELISA and CCK-8 assay were used to determine whether
11R-VIVIT can affect the viability and inflammatory levels of
osteoporotic BMSCs. CCK-8 assay revealed that the viability of
osteoporotic BMSCs was markedly lower than that of normal BMSCs. In
addition, 11R-VIVIT treatment enhanced the viability of
osteoporotic BMSCs compared with the BMSCs from the OVX group
(Fig. 4A). Finally, the results
of ELISA revealed a same pattern as CCK-8 assay. In detail,
11R-VIVIT treatment eliminated the levels of inflammatory factors
in osteoporotic BMSCs (Fig. 4B and
C).
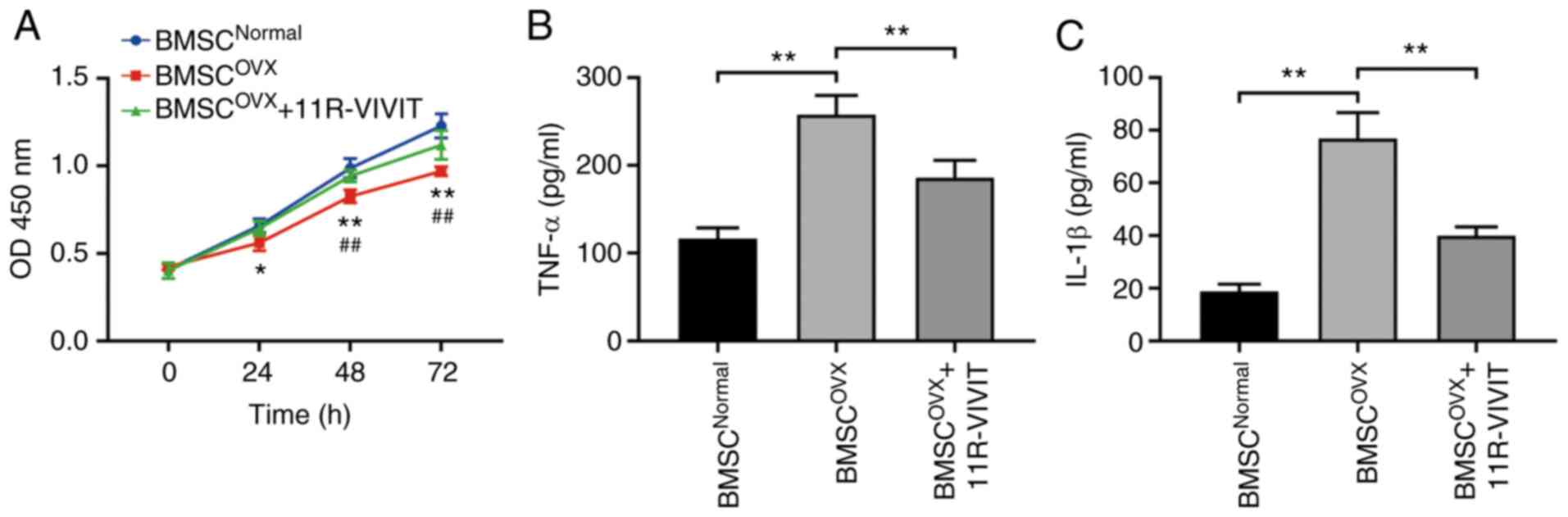 | Figure 411R-VIVIT alleviates the inflammatory
levels of osteoporotic BMSCs and increased their viability. (A)
Cell viability of BMSCs in the OVX group was decreased when
compared with the normal group, and 11R-VIVIT treatment relieved
the decreased cell viability caused by OVX. *P<0.05
and **P<0.01, normal group vs. OVX.
##P<0.01, OVX vs. OVX + 11R-VIVIT. (B and C)
Expression levels of the inflammatory factors, TNF-α and IL-1β, in
BMSCs in the OVX group were increased, and 11R-VIVIT relieved the
increase in the levels of TNF-α and IL-1β induced by OVX.
**P<0.01. BMSC, bone marrow-derived mesenchymal stem
cell; OVX, ovariectomy; TNF-α, tumor necrosis factor α; IL-1β,
interleukin 1β. |
11R-VIVIT promotes autophagy by
inactivating AKT/NFATc1 signaling
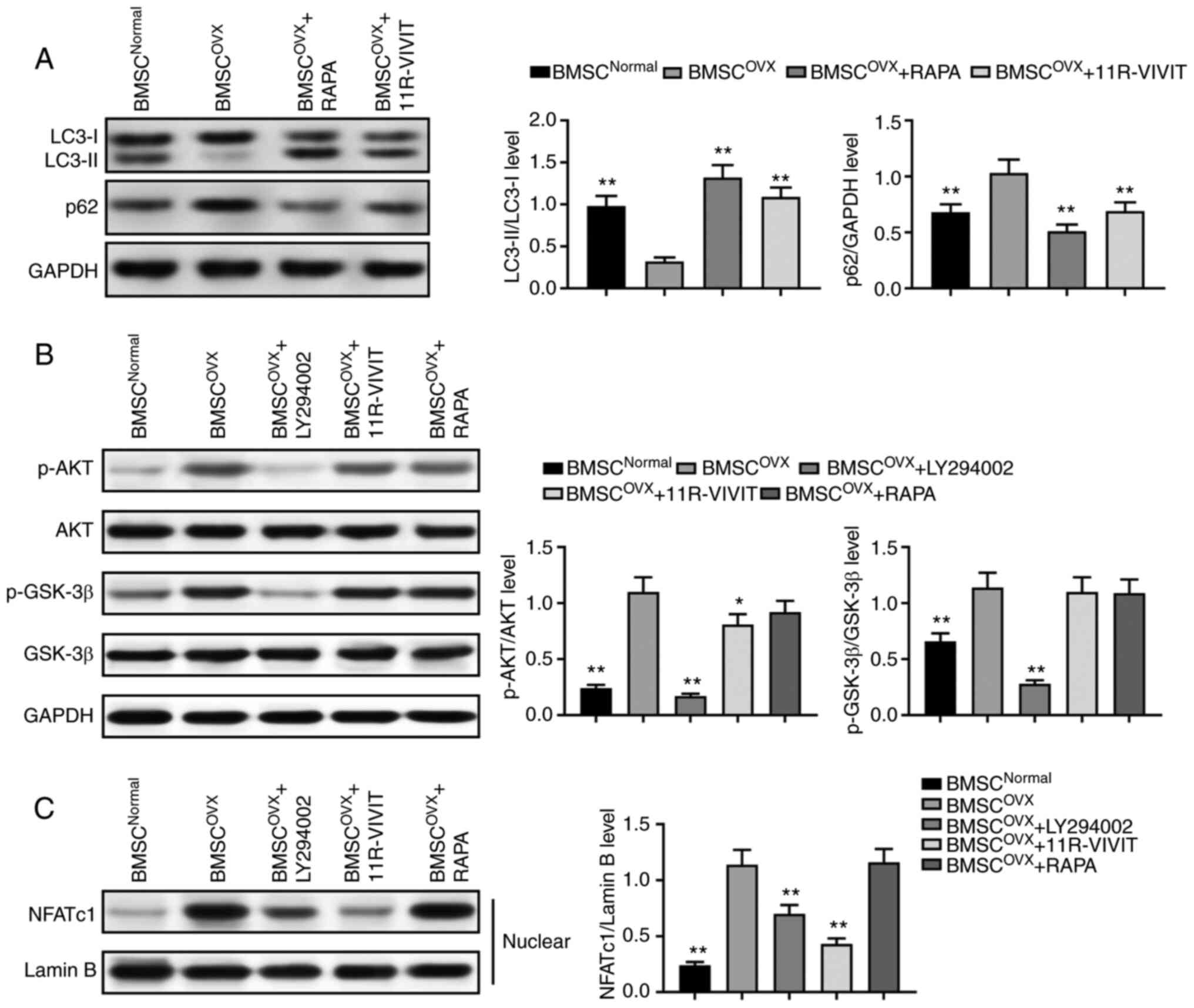
Subsequently, western blot analysis was performed to
determine whether autophagy is involved in the reversion of
osteogenic capacity under 11R-VIVIT treatment. The results revealed
a significant downregulation in the LC3-II/LC3-I ratio and the
upregulation of p62 protein expression in osteoporotic BMSCs during
osteogenesis compared with the normal ones, which indicated that
osteoporosis impaired the autophagy of BMSCs. By contrast, a
significant upregulation in the LC3-II/LC3-I ratio and the
downregulation of p62 activity were observed in response to
treatment with 11R-VIVIT (Fig.
5A), suggesting that 11R-VIVIT acted as an activator of
autophagy in osteoporotic BMSCs.
The present study then attempted to elucidate the
molecular mechanisms through which 11R-VIVIT led to the enhancement
of autophagy and the osteogenesis of osteoporotic BMSCs. As the
AKT/GSK-3β signaling pathway plays a vital role in the autophagy
and osteogenic differentiation of BMSCs, the present study
determined whether there was a crosstalk between 11R-VIVIT and
AKT/GSK-3β signaling. Western blot analysis revealed a significant
upregulation of p-AKT and p-GSK-3β in osteoporotic BMSCs during
osteogenesis compared with the normal BMSCs (Fig. 5B). Thereafter, the osteoporotic
BMSCs were treated with 11R-VIVIT, which resulted in the
downregulation of p-AKT in osteoporotic BMSCs, while the level of
p-GSK-3β exhibited no significant changes in the osteoporotic
BMSCs. In order to confirm these results, LY294002 was used to
inhibit AKT signaling and rapamycin (RAPA) was used to activate
autophagy. Western blot analysis revealed that the activation of
autophagy and the inhibition of AKT signaling exerted similar
effects as observed with 11R-VIVIT treatment. Finally, western blot
analysis revealed that the nuclear expression of NFATc1 was
significantly increased in osteoporotic BMSCs compared with the
normal ones. The nuclear expression of NFATc1 was decreased in the
11R-VIVIT treatment group (Fig.
5C), indicating that 11R-VIVIT inhibited the activation of
NFATc1. Combined with AKT expression inhibition and autophagy
activation, these results demonstrated that 11R-VIVIT activated
autophagy and inhibited NFATc1 activation by inactivating AKT
expression.
Subsequently, Alizarin Red S and ALP staining were
performed to confirm the osteogenic potential of the BMSCs after
the various treatments. The osteoporotic BMSCs treated with
11R-VIVIT, LY294002 and RAPA exhibited an increased osteogenic
potential when compared with the osteoporotic BMSCs without
treatment (Fig. 6A-D). Western
blot analysis of Runx2 and ALP also revealed that the activation of
autophagy and the inhibition of AKT signaling exerted similar
effects as those observed with 11R-VIVIT treatment (Fig. 6E).
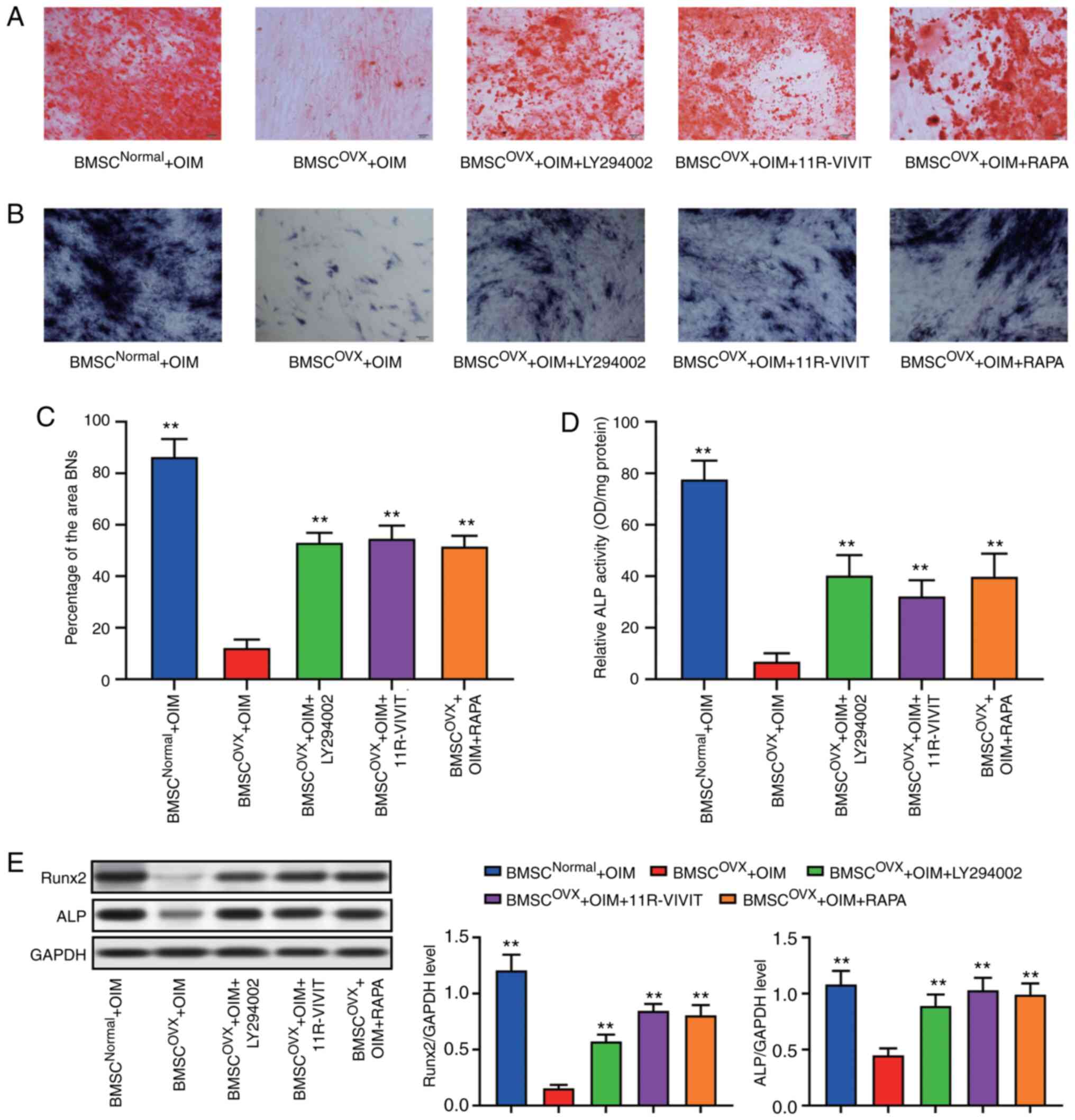 | Figure 6AKT/NFATc1 signaling pathway
regulates the osteogenic differentiation of BSMCs. (A) ALP staining
results. (B) Alizarin Red S staining results. (C) Quantitative
results of ALP staining. (D) Quantitative results of Alizarin Red S
staining. (E) Following osteogenic induction, the levels of
osteogenic marker genes Runx2 and ALP of BMSCs in the OVX group
were significantly reduced, while LY294002 and 11R-VIVIT promoted
their expression. **P<0.01, compared with BMSCs from
rats subjected to OVX. BMSC, bone marrow-derived mesenchymal stem
cell; OVX, ovariectomy; ALP, alkaline phosphatase; AKT, protein
kinase B; GSK-3β, glycogen synthase kinase 3β; NFAT, nuclear factor
of activated T-cells; RAPA, rapamycin. |
Discussion
The most salient finding of the present study was
that the inhibition of NFATc1 with 11R-VIVIT significantly enhanced
osteoporotic fracture healing. In addition, the present study also
revealed that 11R-VIVIT treatment impaired adipogenesis and
enhanced the osteoblast differentiation of the osteoporotic BMSCs
for the first time. Furthermore, the defective osteogenic phenotype
was confirmed in osteoporotic BMSCs, which was reversed by
11R-VIVIT treatment. Moreover, 11R-VIVIT acted as an activator of
autophagy in osteoporotic BMSCs. Taken together, the findings
indicated that 11R-VIVIT promoted autophagy and osteoblast
differentiation for fracture healing through regulating the
AKT/NFATc1 signaling pathway.
Osteoblasts are essential for osteoporotic fracture
healing (3,22). During this complex and dynamic
process, the disorganized differentiation of BMSCs will lead to
abnormal fracture healing, even fracture non-unions. The
replacement of osteoblastogenic activity by adipogenic activity is
a typical feature of osteoporotic BMSCs, in which osteoblast
formation is impaired to a large extent. Therefore, the rescue of
the impaired osteogenic phenotype of osteoporotic BMSCs is helpful
for promoting bone formation and accelerating fracture healing
(23-25). In the present it was demonstrated
that osteoporotic rats with fractures exhibited evidently impaired
fracture healing at 6 and 16 weeks after the fracture, when treated
with saline alone. Furthermore, rats treated with whole-stage
11R-VIVIT exhibited an improved fracture healing process.
NFATc1 is regulated by
calmodulin/Ca2+-dependent signaling. Activated Cn
dephosphorylates NFATc1 protein and leads to its rapid nuclear
translocation. Likewise, the excessive activation of NFATc1 is
capable of by passing the requirement for RANKL and inducing
osteoclast differentiation. The selective inhibition of NFATc1
activation has been shown to result in impaired osteoclast
formation and reduced bone-resorption (26). Furthermore, it has been reported
that NFATc1 signaling negatively regulates the formation of
osteoprogenitor cells (11). The
selective blocking of NFATc1 signaling also favors osteoblast and
bone formation (11,12), which indicates the potential of
11R-VIVIT in regulating bone resorption and fracture unions.
11R-VIVIT has been reported to inhibit NFAT directly and to prevent
the nuclear translocation of NFAT and cell apoptosis (27). Consistently, in the present
study, 11R-VIVIT led to the significant inhibition of NFATc1
expression and nuclear translocation. Furthermore, the results
revealed that 11R-VIVIT stimulated bone formation and attenuated
osteoporosis by decreasing NFATc1 expression and the expression of
inflammation-related molecules, which was in line with the results
of the study by Li et al (10).
Previous findings have demonstrated that the
inactivation of NFATc1 by 11R-VIVIT enhances osteoblast
differentiation (28), which is
in accordance with the results of the present study that 11R-VIVIT
enhanced the osteogenic differentiation of osteoporotic BMSCs by
dysregulating the AKT/NFATc1 signaling pathway to promote the
fracture healing. The results also revealed that 11R-VIVIT
treatment restored the viability of osteoporotic BMSCs and
decreased osteoporosis-induced inflammation. In addition, the
results revealed that11R-VIVIT acted as an activator of autophagy
in osteoporotic BMSCs. 11R-VIVIT also resulted in the
downregulation of p-AKT in osteoporotic BMSCs, similar to the
effects of LY294002 and rapamycin. Furthermore, previous research
has demonstrated that 11R-VIVIT is a protective agent against a
number of pathological conditions. Cai et al (29) reported that the inhibition of
NFATc1 using 11R-VIVIT significantly reduced the development of
nerve injury-induced tactile allodynia. Taken together, the results
of the present study established 11R-VIVIT as a potential
therapeutic strategy against several common diseases, including
osteoporotic fractures.
However, certain limitations to the present study
should be mentioned. Firstly, only limited autophagy-related
proteins were detected due to limited time and attention. Further
autophagy-related proteins need to be detected in the future for
the better understanding of the association between 11R-VIVIT and
autophagy, as well as cell autophagy and the osteogenic
differentiation of osteoporotic BMSCs. Furthermore, other signaling
pathways, such as inflammation-related signaling pathways need to
be examined. Thus, further studies are required for the better
elucidation of the link between 11R-VIVIT and osteoporotic fracture
healing.
In conclusion, the present findings demonstrated
that 11R-VIVIT enhanced rat osteoporotic fracture healing by
modifying AKT/NFATc1 signaling. Moreover, 11R-VIVIT may also be
responsible for promoting the shift of osteoporotic BMSCs to an
osteoblastic phenotype, and may promote bone formation during
osteoporosis.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CH designed the study, performed the experiments,
collected and analyzed data and wrote the manuscript. XW, WJ, ZB
and LZ performed the experiments, and collected and analyzed data.
ML designed the study, conducted experiments and was involved in
manuscript preparation. CH and ML confirm the authenticity of all
the raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the
Institutional Animal Care and Use Committee of The Affiliated
Hangzhou First People's Hospital, Hangzhou, China (no.
ZJCLA-IACUC-20080010).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Abbreviations:
|
BMSCs
|
bone marrow-derived mesenchymal stem
cells
|
|
NFAT
|
nuclear factor of activated
T-cells
|
|
AKT
|
protein kinase B
|
|
GSK-3β
|
glycogen synthase kinase 3β
|
|
Cn
|
calcineurin
|
|
OIM
|
osteogenic induction medium
|
|
AIM
|
adipogenic induction medium
|
|
ALP
|
alkaline phosphatase
|
|
PCR
|
polymerase chain reaction
|
|
OCN
|
osteocalcin
|
|
SDS-PAGE
|
sodium dodecyl sulfate polyacrylamide
gel electrophoresis
|
References
|
1
|
Liang W, Zhuo X, Tang Z, Wei X and Li B:
Calcitonin gene-related peptide stimulates proliferation and
osteogenic differentiation of osteoporotic rat-derived bone
mesenchymal stem cells. Mol Cell Biochem. 402:101–110. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang X, Wang Y, Zhao H, Han X, Zhao T, Qu
P, Li G and Wang W: Extracellular vesicle-encapsulated miR-22-3p
from bone marrow mesenchymal stem cell promotes osteogenic
differentiation via FTO inhibition. Stem Cell Res Ther. 11:2272020.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ali D, Chen L, Kowal JM, Okla M,
Manikandan M, AlShehri M, AlMana Y, AlObaidan R, AlOtaibi N, Hamam
R, et al: Resveratrol inhibits adipocyte differentiation and
cellular senescence of human bone marrow stromal stem cells. Bone.
133:1152522020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
An Q, Wu D, Ma Y, Zhou B and Liu Q:
Suppression of Evi1 promotes the osteogenic differentiation and
inhibits the adipogenic differentiation of bone marrow-derived
mesenchymal stem cells in vitro. Int J Mol Med. 36:1615–1622. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang Z, Wang D, Yang D, Zhen W, Zhang J
and Peng S: The effect of icariin on bone metabolism and its
potential clinical application. Osteoporos Int. 29:535–544. 2018.
View Article : Google Scholar
|
|
6
|
Chen X, Zhi X, Cao L, Weng W, Pan P, Hu H,
Liu C, Zhao Q, Zhou Q, Cui J and Su J: Matrine derivate MASM
uncovers a novel function for ribosomal protein S5 in
osteoclastogenesis and postmenopausal osteoporosis. Cell Death Dis.
8:e30372017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mesner LD, Calabrese GM, Al-Barghouthi B,
Gatti DM, Sundberg JP, Churchill GA, Godfrey DA, Ackert-Bicknell CL
and Farber CR: Mouse genome-wide association and systems genetics
identifies Lhfp as a regulator of bone mass. PLoS Genet.
15:e10081232019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang Y, Wei L, Chang J, Miron RJ, Shi B,
Yi S and Wu C: Strontium-incorporated mesoporous bioactive glass
scaffolds stimulating in vitro proliferation and differentiation of
bone marrow stromal cells and in vivo regeneration of osteoporotic
bone defects. J Mater Chem B. 1:5711–5722. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
He LH, Liu M, He Y, Xiao E, Zhao L, Zhang
T, Yang HQ and Zhang Y: TRPV1 deletion impaired fracture healing
and inhibited osteoclast and osteoblast differentiation. Sci Rep.
7:423852017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li M, Wang X, Bian Z, Yao W, He Q, Tian F,
Zhang J and Zhu L: Peptide 11R-VIVIT stimulates osteoblastogenesis
through regulating the expression of nuclear factor of activated T
cells cytoplasmic 1. Cell Mol Biol (Noisy-le-grand). 63:46–52.
2017. View Article : Google Scholar
|
|
11
|
Boise LH, Petryniak B, Mao X, June CH,
Wang CY, Lindsten T, Bravo R, Kovary K, Leiden JM and Thompson CB:
The NFAT-1 DNA binding complex in activated T cells contains Fra-1
and JunB. Mol Cell Biol. 13:1911–1919. 1993.PubMed/NCBI
|
|
12
|
Mulero MC, Aubareda A, Orzaez M, Messeguer
J, Serrano-Candelas E, Martínez-Hoyer S, Messeguer A, Pérez-Payá E
and Pérez-Riba M: Inhibiting the calcineurin-NFAT (nuclear factor
of activated T cells) signaling pathway with a regulator of
calcineurin-derived peptide without affecting general calcineurin
phosphatase activity. J Biol Chem. 284:9394–9401. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li R, Zhang L, Shi W, Zhang B, Liang X,
Liu S and Wang W: NFAT2 mediates high glucose-induced glomerular
podocyte apoptosis through increased Bax expression. Exp Cell Res.
319:992–1000. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang L, Guan H, He J, Zeng L, Yuan Z, Xu
M, Zhang W, Wu X and Guan J: VEGF increases the proliferative
capacity and eNOS/NO levels of endothelial progenitor cells through
the calcineurin/NFAT signalling pathway. Cell Biol Int. 36:21–27.
2012. View Article : Google Scholar
|
|
15
|
Zhang L, Li R, Shi W, Liang X, Liu S, Ye
Z, Yu C, Chen Y, Zhang B, Wang W, et al: NFAT2 inhibitor
ameliorates diabetic nephropathy and podocyte injury in db/db mice.
Br J Pharmacol. 170:426–439. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sophocleous A and Idris AI: Rodent models
of osteoporosis. Bonekey Rep. 3:6142014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fathi E, Valipour B, Sanaat Z, Nozad
Charoudeh H and Farahzadi R: Interleukin-6, -8, and TGF-β secreted
from mesenchymal stem cells show functional role in reduction of
telomerase activity of leukemia cell via Wnt5a/β-catenin and P53
pathways. Adv Pharm Bull. 10:307–314. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fathi E, Farahzadi R, Valipour B and
Sanaat Z: Cytokines secreted from bone marrow derived mesenchymal
stem cells promote apoptosis and change cell cycle distribution of
K562 cell line as clinical agent in cell transplantation. PLoS One.
14:e02156782019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Farahzadi R, Fathi E and Vietor I:
Mesenchymal stem cells could be considered as a candidate for
further studies in cell-based therapy of Alzheimer's disease via
targeting the signaling pathways. ACS Chem Neurosci. 11:1424–1435.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
21
|
Fathi E, Farahzadi R, Javanmardi S and
Vietor I: L-carnitine extends the telomere length of the cardiac
differentiated CD117(+)-expressing stem cells. Tissue Cell.
67:1014292020. View Article : Google Scholar
|
|
22
|
Xia WF, Jung JU, Shun C, Xiong S, Xiong L,
Shi XM, Mei L and Xiong WC: Swedish mutant APP suppresses
osteoblast differentiation and causes osteoporotic deficit, which
are ameliorated by N-acetyl-L-cysteine. J Bone Miner Res.
28:2122–2135. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou Y, Zhu ZL, Guan XX, Hou WW and Yu HY:
Reciprocal roles between caffeine and estrogen on bone via
differently regulating cAMP/PKA pathway: The possible mechanism for
caffeine-induced osteoporosis in women and estrogen's antagonistic
effects. Med Hypotheses. 73:83–85. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu R, Fu Z, Liu X, Xiao T, Zhang P, Du Y,
Yuan H, Cheng J and Jiang H: Transplantation of osteoporotic bone
marrow stromal cells rejuvenated by the overexpression of SATB2
prevents alveolar bone loss in ovariectomized rats. Exp Gerontol.
84:71–79. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yu F, Wu F, Li F, Liao X, Wang Y, Li X,
Wang C, Shi Y and Ye L: Wnt7b-induced Sox11 functions enhance
self-renewal and osteogenic commitment of bone marrow mesenchymal
stem cells. Stem Cells. 38:1020–1033. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Crotti TN, Flannery M, Walsh NC, Fleming
JD, Goldring SR and McHugh KP: NFATc1 regulation of the human beta3
integrin promoter in osteoclast differentiation. Gene. 372:92–102.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bodmer D, Perkovic A, Sekulic-Jablanovic
M, Wright MB and Petkovic V: Pasireotide prevents nuclear factor of
activated T cells nuclear translocation and acts as a protective
agent in aminoglycoside-induced auditory hair cell loss. J
Neurochem. 139:1113–1123. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Maoqiang L, Zhenan Z, Fengxiang L, Gang W,
Yuanqing M, Ming L, Xin Z and Tingting T: Enhancement of osteoblast
differentiation that is inhibited by titanium particles through
inactivation of NFATc1 by VIVIT peptide. J Biomed Mater Res A.
95:727–734. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cai YQ, Chen SR and Pan HL: Upregulation
of nuclear factor of activated T-cells by nerve injury contributes
to development of neuropathic pain. J Pharmacol Exp Ther.
345:161–168. 2013. View Article : Google Scholar : PubMed/NCBI
|