Introduction
At present, infections of severe acute respiratory
syndrome coronavirus 2 (SARS-CoV-2) are a global life threatening
disease (1). Updated from weekly
report of WHO on 13 July 2021, the cumulative deaths from
SARS-CoV-2 have surpassed 4 million people and the cumulative cases
was up to 186 million people globally (2). S-(S) glycoprotein (spike protein),
nucleocapsid protein, membrane protein and envelope protein are all
structural proteins of SARS-CoV-2 (3). S-protein is prominent in the viral
membrane (3). SARS-CoV-2 uses
S-protein binding to angiotensin-converting enzyme 2 (ACE2) to
invade the host cells (4). ACE2
is a transmembrane protein with its active site in the N-terminal
domain (5), which is also the
SARS-CoV binding site. Lung, heart, vessels, gut, testis and brain
all express ACE2 receptors (6-9).
In particular, ACE2 receptors are abundantly expressed in the
kidney (10), which can be
infected by SARS-CoV-2 (11).
A disintegrin and metalloproteases (ADAMs) are
membrane-bound proteins with disintegrin and metalloproteinase
domains (12). A disintegrin and
metalloprotease domain 17 (ADAM17) is one of members of the ADAM
family of transmembrane proteins containing an N-terminal domain
(12). ADAMs are involved in
ectodomain shedding of enzymes including ACE2 (12). Cellular (membrane-bound) ACE2 can
be cleaved to release the ACE2 ectodomain into the circulation by
ADAM17 (13). An increase of
ectodomain ACE2 may exacerbate the RAS (renin-angiotensin system)
imbalance (ACE/ACE2) in patients with specific comorbidities, such
as cardiovascular disease, hypertension, diabetes and chronic lung
disease (12). ACE2 is also
downregulated by viral transcription (14) and endocytosis together with the
virus (15). In addition, a
decrease in ADAM17 expression inhibits ACE2 shedding, while
introduction of ADAM17 cDNA restores SARS-S-induced ACE2 shedding
(16). High levels of shed ACE2
can reduce SARS-CoV virus infection (13,17).
MicroRNAs (miRNAs) are non-coding RNAs which can
directly bind to the 3′ untranslated regions (3′-UTRs) of target
mRNAs to regulate gene expression posttranscriptionally. miRNAs are
powerful regulators of numerous cell processes, including cell
growth, differentiation, development and apoptosis (18). miRNAs are involved in various
viral infectious diseases, such as human immunodeficiency virus-1
(HIV-1) infection (19), human
papillomavirus infection (20),
SARS-CoV infection (21) and
hepatitis B virus infection (20-22). As mature miRNAs, miR-28-3p and
miR-28-5p are derived from the 3′ and 5′ ends, respectively, of
pre-miR-28 and then go on to target different mRNAs, such as WD
repeat and SOCS box-containing 2 and forkhead box O1 (23-27). There is evidence indicating that
miRNAs may block RNA virus replication during viral infection
(28). Purohit et al
(29) demonstrated that
downregulation of miRNA-28 expression increased HIV infection by
promoting viral replication (29) and it has also been demonstrated
that miR-28-3p inhibits human T cell leukemia virus type 1
replication and virus infection (23). Although the expression and
biological function of these miRNAs can suppress the replication of
some viruses, the effects of miR-28-3p on SARS-Cov-2 infection
remain unknown. In the present study, it was hypothesized that
miR-28-3p may exert a regulatory role in ADAM17-dependent ACE2
ectodomain shedding during SARS-Cov-2 infection. Since the present
study demonstrated the direct interaction between miR-28-3p and
ADAM17, upregulation of miR-28-3p may provide a novel therapeutic
strategy for treatment of patients with SARS-CoV-2.
Materials and methods
Cell culture and S-protein treatment
293T cells, with high transfection efficiency and
stably expressing hACE2, which are termed tool cells and were used
in some studies of SARS-Cov-2 virus (30-33), were purchased from the American
Type Culture Collection. Cells were cultured in DMEM (Gibco; Thermo
Fisher Scientific Inc.) supplemented with 10% fetal bovine serum
(FBS) (Gibco; Thermo Fisher Scientific Inc.) and 100 mg/ml
penicillin/streptomycin (Gibco; Thermo Fisher Scientific Inc.).
293T cells were treated with 100 ng/ml of S-protein for 48 h at
37°C in a humidified incubator with 5% CO2 (cat. no.
bs-46008P; BIOSS) (34,35). Cells were maintained in an
incubator at 37°C with 5% CO2, and were passaged every 3
days.
Transfection
293T cells were seeded in 6-well plates
(1×105 cells/well) and maintained in an incubator at
37°C with 5% CO2 for 12 h to obtain a cell confluence of
30-35%. For transfection, the cell culture medium was replaced with
2 ml Opti-MEM medium (Gibco; Thermo Fisher Scientific Inc.) without
FBS and antibiotics 12 h before transfection. Then, a total of 10
µl of Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) was diluted with 200 µl of Opti-MEM
and the miR-28-3p mimic, miR-28-3p inhibitor, mimic negative
control (NC) or inhibitor NC, or with small interfering (si)RNA or
siRNA NC, and the mixture was incubated for 5 min at room
temperature. Subsequently, the cells were transfected with a final
concentration of 20 nM miR-28-3p mimic/inhibitor or miR-28-3p
mimic/inhibitor NC, or with 100 pmol si-ADAM17 or siRNA NC
(scrambled) at 37°C with 5% CO2 for 16 h, after
which the medium was replaced with complete DMEM supplemented with
10% FBS for 72 h in a humidified incubator at 37°C with 5%
CO2. At 24, 48 and 72 h after transfection, the
transfection efficiency was evaluated by using inverted
fluorescence microscope (Ti-S; Nikon Corporation) to observe and
count the GFP-positive cells. The miR-28-3p mimic, miR-28-3p mimic
NC, miR-28-3p inhibitor and miR-28-3p inhibitor NC were obtained
from Guangzhou RiboBio Co. Ltd. The miR-28-3p mimic and miR-28-3p
mimic NC, miR-28-3p inhibitor and miR-28-3p inhibitor NC sequences
were as follows: miR-28-3p mimic, 5′-UAG AUC ACA GUC CUU UGU
UAU-3′; miR-28-3p mimic NC, 5′-AUC UAG UCA GUC CUU UGU UUA U-3′;
miR-28-3p inhibitor, 5′-AUC UAG UGU CAG GAA ACA AAU A-3′; and
miR-28-3p inhibitor NC, 5′-UAG AUC AGU CAG GAA ACA AAU A-3′. The
si-ADAM17 or siRNA NC were obtained from Shanghai GenePharma Co.
Ltd. si-ADAM17 and siRNA NC sequences were as follows: si-ADAM17,
5′-TGA GGC AGT CTC TCC TAT TCC TGA CCA GC-3′; and siRNA NC
(scramble), 5′-TGA CCA CCC TGA CCT ACG GCG TGC AGT GC-3′.
Western blotting (WB)
293T cells were seeded in 6-well plates
(1×105 cells/well) and cultured at 37°C with 5%
CO2 for 48 h after transfection (34). Then, cell culture supernatants
were collected. Total protein of cell supernatants was concentrated
by using Amicon® Ultra Centrifugal Filters
(MilliporeSigma). As a control, 10 µg of ovalbumin (OVA) was
added to each sample of supernatant and the precipitates were
probed with anti-OVA and anti-ACE2 antibodies. Total protein was
extracted from cell culture using RIPA buffer (Beyotime Institute
of Biotechnology). Protein concentrations were determined using
Bradford assays (Thermo Fisher Scientific, Inc.). Total protein (40
µg/lane) was separated by 10% SDS-PAGE and transferred to
PVDF membranes. The membranes were blocked with 5% fat-free skimmed
milk for 2 h at room temperature and subsequently incubated with
anti-ADAM17 (cat. no. ab39163), anti-GAPDH (cat. no. Ab8245),
anti-ACE2 (cat. no. ab108252) and anti-OVA (anti-ovalbumin) (cat.
no. ab236590) (all 1:1,000; Abcam) at 4°C overnight and then washed
with TBST buffer (0.1% Tween-20 in 1X TBS buffer) three times. The
secondary antibody goat anti-rabbit IgG H&L horseradish
peroxidase (HRP) (1:3,000; cat. no. ab6721; Abcam) was incubated
with the membranes for 2 h at room temperature. The membranes were
washed thrice, 5 min each time in TBST, then incubated in ECL
substrate chemiluminescent detection reagent (Thermo Fisher
Scientific, Inc.) for 5 min. The chemiluminescent blots were imaged
first with the ChemiDoc MP imager (Bio-Rad, Inc.) and then on film.
GAPDH and OVA were used as the loading controls for cell lysate and
cell culture supernatants, respectively. The band analysis tools of
ImageJ v.1.8.0 software (National Institutes of Health) were used
to calculate the gray value of the bands after subtracting the
background density.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
293T cells were seeded in 6-well plates
(1×105 cells/well) and cultured at 37°C with 5%
CO2. Following transfection for 48 h, TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.) was used to extract
total RNA on ice and the concentration and purity of RNA were
detected using Nanodrop 2000 (Thermo Fisher Scientific Inc.). DNA
contamination was removed using DNase I (cat. no. AMPD1,
Sigma-Aldrich; Merck KGaA) and then the miRNAs were reversed
transcribed into cDNA using the PrimeScript RT kit (Takara Bio
Inc.). The reverse transcription program was as follows: 65°C for 5
min, on ice immediately followed by the addition of reverse
transcriptase, annealing at 50°C for 15 min followed by extension
at 95°C for 5 min and 4°C hold. miScript HiSpec buffer was used to
prepare the cDNA and real-time PCR was carried out using iTaq™
SYBR® Green Supermix (Beijing Solarbio Science &
Technology Co., Ltd.) according to the manufacturer's protocol. The
reaction conditions were as follows: 95°C for 3 min followed by 40
cycles of 3-step PCR (98°C for 10 sec, 60°C for 30 sec and 72°C for
60 sec), 72°C for 10 min, then hold at 4°C. U6 was used as the
internal control. The expression of miRNAs was determined using the
2−ΔΔCq method (36).
Primer sequences were as follows: miR-28-3p forward, 5′-CGC GCA CTA
GAT TGT GAG CT-3′ and reverse, 5′-AGT GCA GGG TCC GAG GTA TT-3′;
and U6 forward, 5′-ATT GGA ACG ATA CAG AGA AGA TT-3′ and reverse,
5′-GGA ACG CTT CAC GAA TTT G-3′.
MTT assay
293T cells were seeded in 6-well plates
(1×104 cells/well). Following transfection for 48 h, 20
µl MTT (5 mg/ml, Sigma-Aldrich; Merck KGaA) was added to
each well and the cells were cultured for another 4 h in an
incubator at 37°C with 5% CO2. The solution was
discarded after incubation for 4 h and then 150 µl DMSO
(Sigma-Aldrich; Merck KGaA) was added to each well and incubated in
the dark for 10 min with shaking. A microplate reader (Bio-Rad,
Inc.) was used to detect the optical density at 490 nm.
TUNEL assay
TUNEL assay was carried out by using DNA
Fragmentation Imaging kit (cat. no. 6432344001; Roche Diagnostics;
Merck KGaA). 293T cells were seeded in 6-well plates
(1×104 cells/well). Following transfection for 48 h,
cells were fixed for 30 min in 4% paraformaldehyde in
phosphate-buffered saline solution (PBS) at 4°C, then incubated
with 0.1% Triton X-100 in PBS for 2 min on ice. After washing the
cells with PBS three times, the cells were incubated with 45
µl of the Reaction Solution, which was combined the Enzyme
Solution with the Label Solution, for 60 min in the dark at 37°C
and then washed with PBS three times at room temperature. Then, 150
µl of the Nuclei Dye mixture was added and cells were
incubated for 5 min at room temperature; a coverslip was added and
images were captured under a fluorescence microscope (80i; Nikon
Corporation; magnification, ×200). At least 100 cells were counted
in 10 random fields and the percentage of positive apoptotic cells
was calculated.
Dual-Luciferase activity assay
ADAM17 (3′ UTR) was predicted as the binding site of
miR-28-3p by TargetScan v.7.2 (www.targetscan.org) online prediction software. 293T
cells were seeded in 6-well plates (1×105 cells/well),
and then co-transfected with a pmir-GLO dual-luciferase miRNA
target expression vector (Promega Corporation), containing a
wild-type (WT) or mutant (Mut) ADAM17 untranslated region (3′UTR)
and miR-28-3p mimic (5′-UAG AUC ACA GUC CUU UGU UAU-3′) or mimic-NC
(5′-AUC UAG UCA GUC CUU UGU UUA U-3′) using Lipofectamine
2000® (Invitrogen; Thermo Fisher Scientific, Inc.).
Following 48 h of incubation, a dual-luciferase reporter assay
(Promega Corporation) was conducted to detect luciferase activity
according to manufacturer's instructions. Firefly
luciferase activity was normalized by comparison with
Renilla luciferase activity.
Statistical analysis
SPSS 18.0 software (IBM Corp.) was used for
statistical analyses. Data were shown as means ± standard deviation
(SD) from 3 independent experiments. The differences between 2
groups were determined using paired Student's t-tests or one-way
analysis of variance followed by the post hoc Tukey's test. The
correlation test used was Pearson's correlation test.
*P<0.05 was considered to indicate a statistically
significant difference.
Results
miR-28-3p expression and ADAM17 and cell
viability in S-protein-treated 293T cells
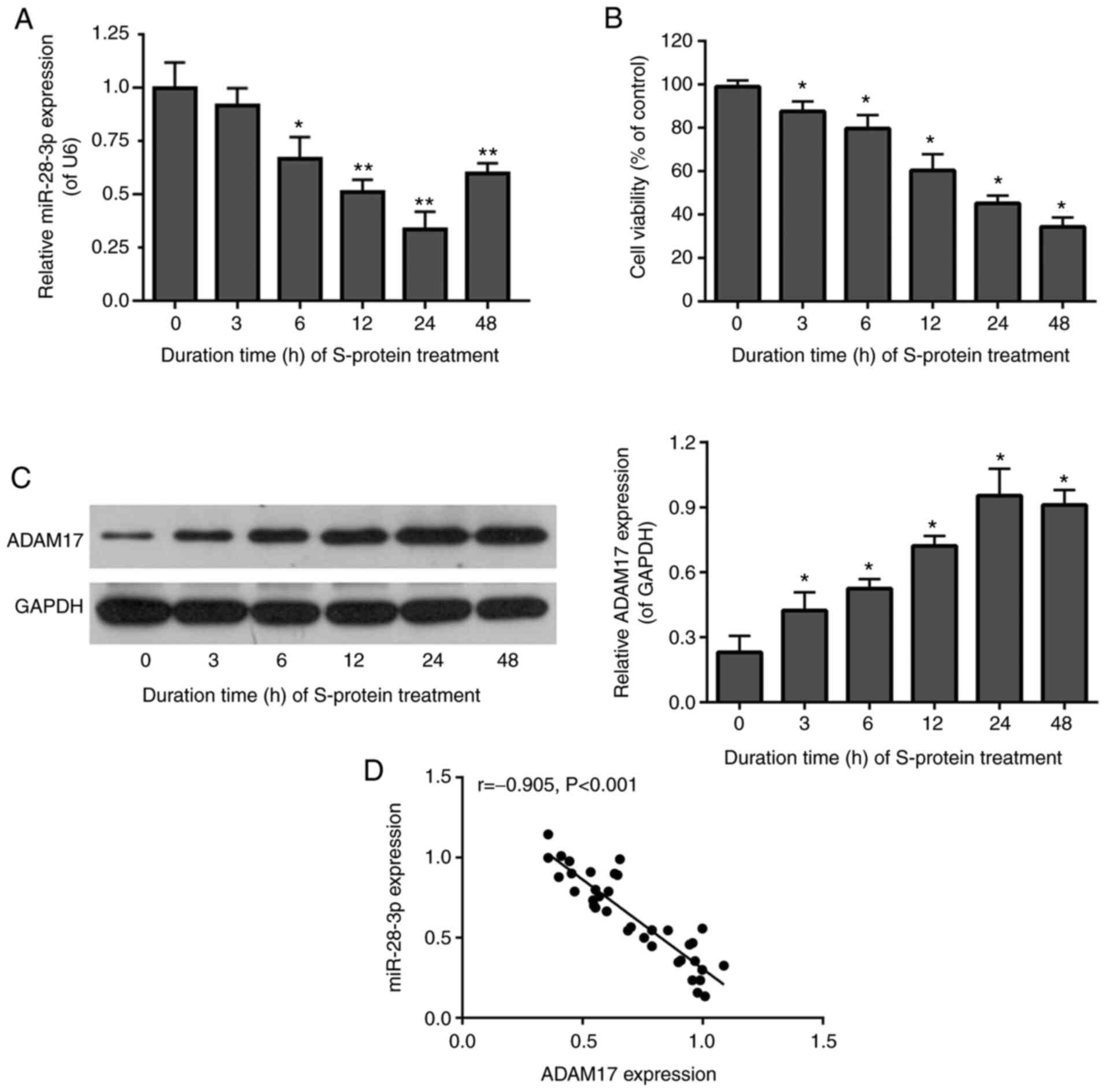
To determine whether miR-28-3p and ADAM17 were
related in S-protein-treated 293T cells, miR-28-3p and ADAM17
expression were assessed by RT-qPCR and WB in 293T cells after
S-protein treatment at 37°C for 0, 3, 6, 12, 24 and 48 h at 37°C.
The level of miR-28-3p was downregulated (Fig. 1A), while ADAM17 protein was
upregulated (Fig. 1C) in
S-protein-treated 293T cells. Expression of miR-28-3p and ADAM17
was lowest and highest, compared with 0 h group respectively, after
treatment for 24 h (Fig. 1A and
C). In addition, cell viability of S-protein-treated 293T cells
was significantly diminished at 24 h compared with the 0 h group
(Fig. 1B). miR-28-3p and ADAM17
were highly and inversely correlated by Pearson's correlation test
(Fig. 1D). The above results
suggested that S-protein treatment was able to affect cell
viability and regulate the expression of both miR-28-3p and ADAM17.
In addition, the expression of miR-28-3p and ADAM17 was negatively
correlated.
To investigate the effects of miR-28-3p mimic and
si-ADAM17 on the viability and apoptosis of S-protein-treated 293T
cells, the S-protein-treated 293T cells were transfected with
miR-28-3p mimic and si-ADAM17. The transfection efficiency of
miR-28-3p mimic (Fig. 2A) and
si-ADAM17 (Fig. 2B) were
analyzed by RT-qPCR and WB, respectively. For cell viability, 293T
treated with S-protein demonstrated lower cell viability when
compared with control 293T cells (Fig. 2C). However, both miR-28-3p mimic
and si-ADAM17 promoted cell viability in the S-protein-treated 293T
cells when compared with mimic-NC or siRNA-NC, respectively
(Fig. 2C). For apoptosis, 293T
cells treated with S-protein demonstrated a higher apoptosis rate
when compared with control 293T cells (Fig. 2D) However, both miR-28-3p mimic
and si-ADAM17 could inhibit apoptosis rate in the S-protein-treated
293T cells when compared with mimic-NC or siRNA-NC, respectively.
The aforementioned results demonstrated that S-protein treatment
affected cell viability and cell apoptosis through both miR-28-3p
and ADAM17.
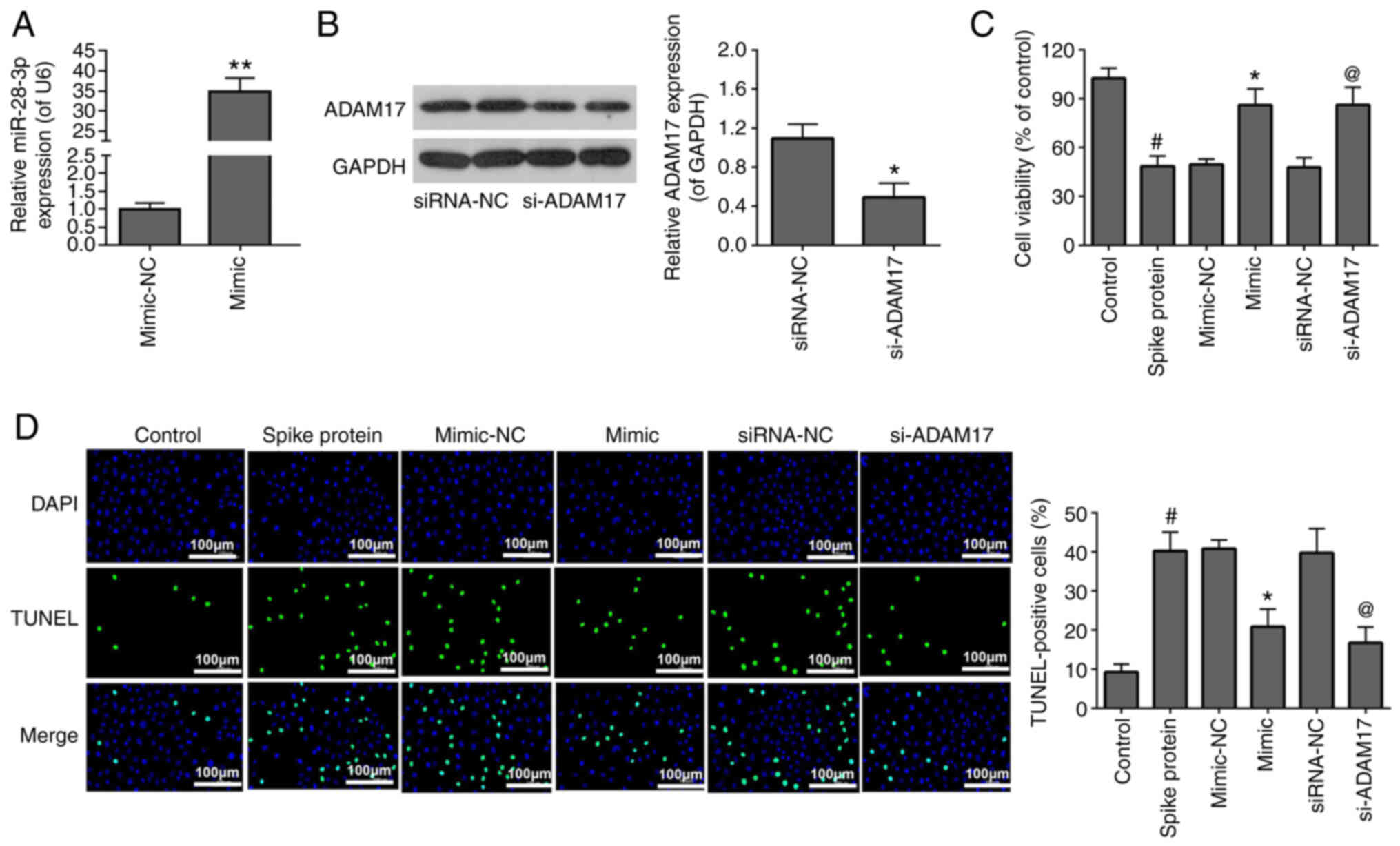 | Figure 2miR-28-3p mimic and si-ADAM17 reverse
the effects of S-protein on proliferation and apoptosis in 293T
cells. (A) Expression of miR-28-3p mimic and mimic-NC was measured
by RT-qPCR. (B) Expression of si-ADAM17 and siRNA-NC was measured
by WB. (C) Cell proliferation in S-protein-treated 293T cells
transfected with miR-28-3p mimic, si-ADAM17, mimic-NC or siRNA-NC
were measured using the MTT assay. (D) Cell apoptosis in
S-protein-treated 293T cells transfected with miR-28-3p mimic,
si-ADAM17, mimic-NC or siRNA-NC measured by the TUNEL assay.
#P<0.05 vs. control; *P<0.05 vs.
mimic-NC, **P<0.01 vs. mimic-NC;
@P<0.05 vs. siRNA-NC. WB, western blotting;
S-protein, spike protein; ADAM17, A disintegrin and
metalloproteinase 17; RT-qPCR, reverse-transcription quantitative
polymerase chain reaction; NC, negative control; miR, microRNA; si,
small interfering. |
Both miR-28-3p mimic and si-ADAM17
inhibit shedding of the ACE2 ectodomain in 293T cells treated with
S-protein
To examine the effects of miR-28-3p mimic and
si-ADAM17 on ACE2 shedding in S-protein-treated 293T cells, WB was
used to detect the expression of ACE2 in the supernatants and whole
cells. As shown in Fig. 3, for
supernatants, 293T treated with S-protein demonstrated a higher
level of ACE2 when compared with control 293T cells and both
miR-28-3p mimic and si-ADAM17 could inhibit ACE2 expression in the
S-protein-treated 293T cells when compared with their respective
controls (Fig. 3). The
expression of ACE2 in the whole cell lysate demonstrated a reverse
trend compared with supernatants. 293T cells were treated with
S-protein, the expression of ACE2 in cells was lower compared with
the control. While 293T cells transfected with miR-28-3p mimic or
si-ADAM17, the expression of ACE2 in cells was higher compared with
mimic-NC or siRNA-NC, respectively (Fig. 3). These results suggested that
S-protein treatment promoted ACE2 shedding. miR-28-3p mimic or
si-ADAM17 downregulated ACE2 shedding.
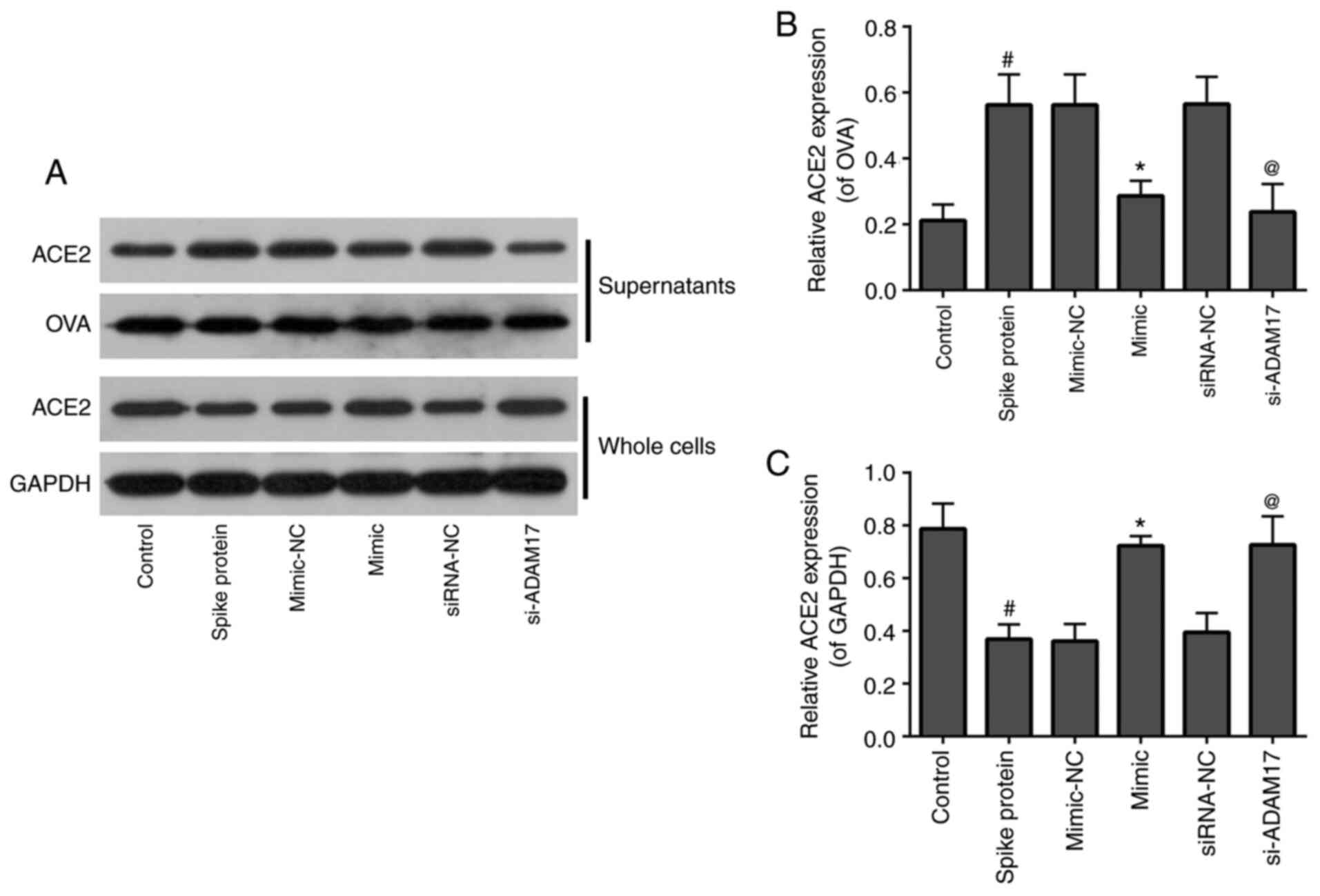 | Figure 3miR-28-3p mimic and si-ADAM17 inhibit
shedding of the ACE2 ectodomain in S-protein-treated 293T cells.
(A) Western blotting results of ACE2, OVA, GAPDH expression in
supernatants and whole cells of control, spike protein, mimic-NC,
mimic, siRNA-NC, si-ADAM17 group. (B) Quantification of ACE2/OVA in
supernatants of control, spike protein, mimic-NC, mimic, siRNA-NC
and si-ADAM17 groups. (C) Quantification graph of ACE2/GAPDH in
whole cells of control, spike protein, mimic-NC, mimic, siRNA-NC
and si-ADAM17 groups. #P<0.05 vs. control;
*P<0.05 vs. mimic-NC; @P<0.05 vs.
siRNA-NC. ACE2, angiotensin-converting enzyme 2; S-protein, spike
protein; ADAM17, A disintegrin and metalloproteinase 17; NC,
negative control; miR, microRNA; si, small interfering; OVA,
ovalbumin. |
ADAM17 is a potential target of
miR-28-3p
TargetScan v.7.2 was used to predict the potential
target gene of miR-28-3p. It was found that the ADAM17 3′-UTR
contained a potential binding site for miR-28-3p (Fig. 4A). The miR-28-3p mimic
significantly suppressed WT, but not Mut 3′-UTR luciferase activity
in S-protein-treated 293T cells in dual-luciferase activity assay
(Fig. 4B). To verify that ADAM17
is a direct target of miR-28-3p, S-protein-treated 293T cells were
transfected with miR-28-3p mimic or miR-28-3p inhibitor. First of
all, the inhibitory effect of miR-28-3p inhibitor was verified by
RT-qPCR and the inhibitory efficiency was ~50% (Fig. 4C). Subsequently, the ADAM17
expression level was assessed in S-protein-treated-293T cells
transfected with miR-28-3p mimic or miR-28-3p inhibitor,
respectively. 293T cells treated with S-protein demonstrated higher
ADAM17 level when compared with control 293T cells (Fig. 4D). miR-28-3p mimic inhibited
ADAM17 expression, while miR-28-3p inhibitor upregulated ADAM17
expression in the S-protein-treated 293T cells when compared with
mimic-NC or inhibitor-NC, respectively. These results demonstrated
that ADAM17 is a direct target of miR-28-3p during S-protein
treatment.
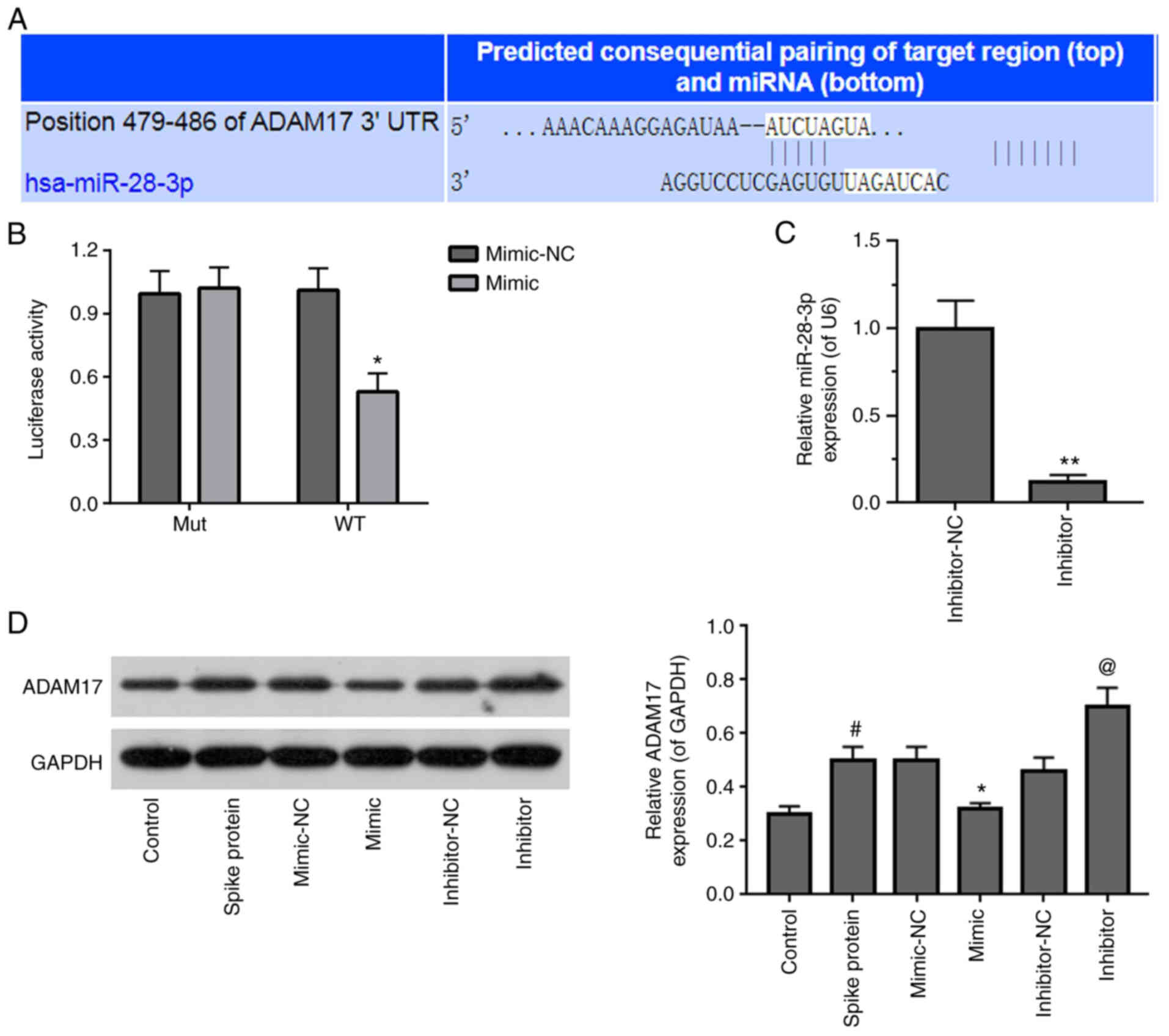 | Figure 4Identification of ADAM17 as a target
of miR-28-3p. (A) Pairing of the 3′-UTR sequence of ADAM17 and
miR-28-3p sequence using TargetScan v.7.2. (B) Luciferase assay in
S-protein-treated 293T cells with WT-ADAM17 or Mut-ADAM17 and
miR-28-3p mimic or mimic-NC. (C) Expression of miR-28-3p inhibitor
and inhibitor-NC in 293T cells was measured by RT-qPCR. (D)
Expression of ADAM17 in S-protein-treated 293T cells transfected
with miR-28-3p mimic, miR-28-3p inhibitor, mimic-NC or inhibitor-NC
was detected by WB. #P<0.05 vs control;
*P<0.05 vs. mimic-NC, **P<0.01 vs.
mimic-NC; @P<0.05 vs. inhibitor-NC. WB, western
blotting; S-protein, spike protein; ADAM17, A disintegrin and
metalloproteinase 17; RT-qPCR, reverse-transcription quantitative
polymerase chain reaction; NC, negative control; miR, microRNA; si,
small interfering; WT, wild-type; Mut, mutant; UTR, untranslated
region. |
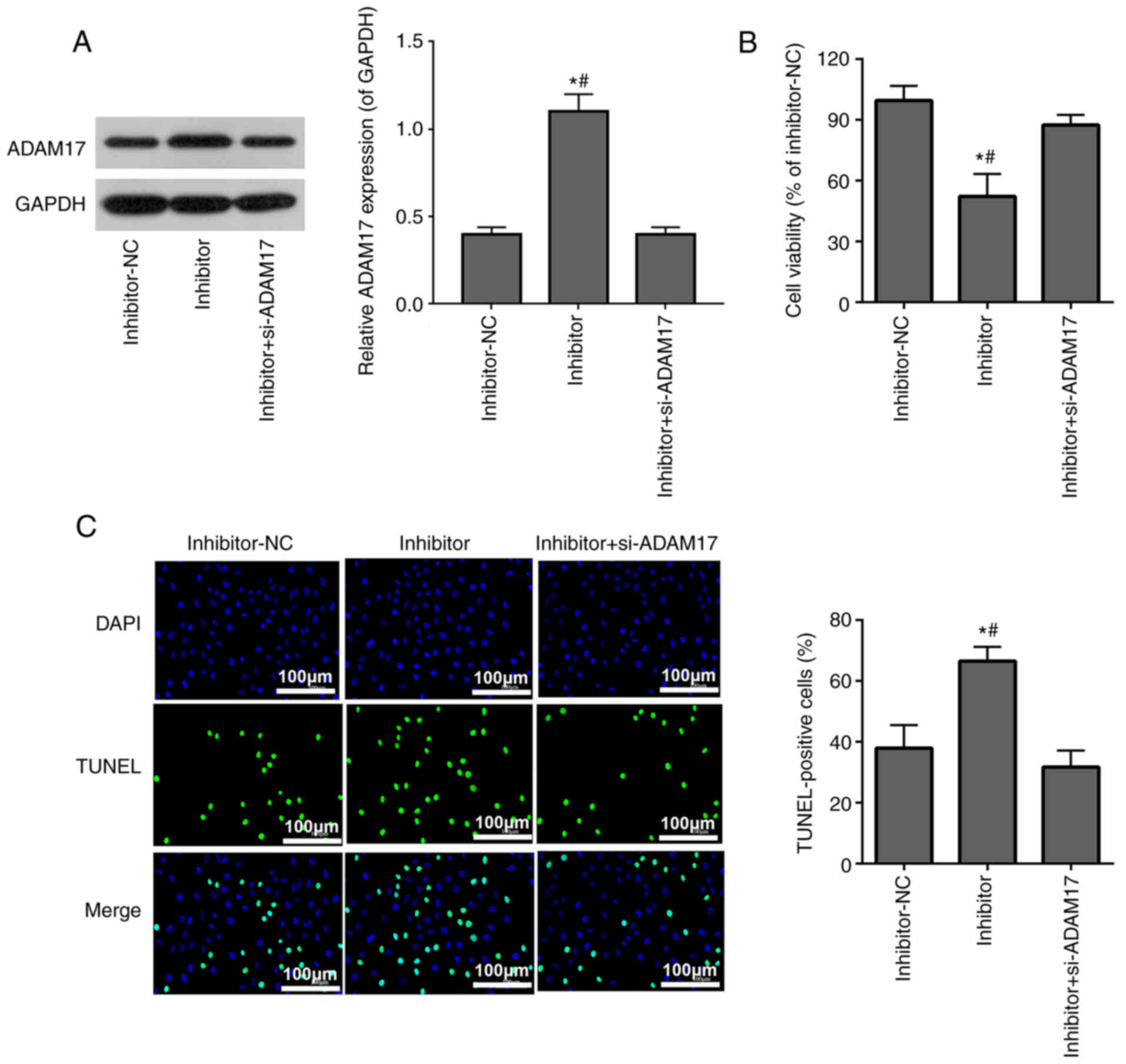
si-ADAM17 inhibits the effects of
miR-28-3p inhibitor on cell viability and apoptosis in 293T cells
treated with the S-protein
293T cells treated with the S-protein were
transfected with miR-28-3p inhibitor alone or co-transfected with
si-ADAM17 and miR-28-3p inhibitor. WB confirmed that miR-28-3p
inhibitor upregulated ADAM17 protein expression, while
co-transfection with si-ADAM17 and miR-28-3p inhibitor successfully
reversed the effect of miR-28-3p inhibitor on ADAM17 expression
(Fig. 5A). MTT assay
demonstrated that miR-28-3p inhibitor inhibited S-protein-treated
293T cell proliferation, while co-transfection with si-ADAM17 and
miR-28-3p inhibitor weakened the inhibitory effect of miR-28-3p
inhibitor on S-protein-treated 293T cell proliferation (Fig. 5B). TUNEL assay demonstrated that
miR-28-3p inhibitor promoted apoptosis of S-protein-treated 293T
cells, while co-transfection with si-ADAM17 and miR-28-3p inhibitor
weakened the promoted effect of miR-28-3p inhibitor on the
apoptosis of S-protein-treated 293T cells (Fig. 5C). The aforementioned results
demonstrated that miR-28-3p inhibitor inhibited cell viability and
promoted cell apoptosis during S-protein treatment and this
inhibition could be abolished by ADAM17 knockdown, which meant that
miR-28-3p exerts its function on both cell viability and cell
apoptosis through ADAM17.
si-ADAM17 reverses the effects of
miR-28-3p inhibitor on the shedding of the ACE2 ectodomain in 293T
cells treated with the S-protein
To further verify that miR-28-3p regulated the ACE2
ectodomain shedding of S-protein-treated-293T cells by targeting
ADAM17, miR-28-3p inhibitor alone or miR-28-3p inhibitor combined
with si-ADAM17 was transfected into S-protein-treated 293T cells.
WB demonstrated that the miR-28-3p inhibitor elevated ACE2 level in
supernatants of S-protein-treated 293T cells and reduced ACE2 level
of whole cell lysates (Fig. 6).
Co-transfection with miR-28-3p inhibitor and si-ADAM17 reversed the
effect of transfected miR-28-3p inhibitor alone on the ACE2 level
both in supernatants and in whole cell lysates (Fig. 6). This suggested that miR-28-3p
inhibitor enhanced ACE2 shedding of S-protein-treated 293T cells,
while si-ADAM17 efficiently blocked miR-28-3p inhibitor-induced
ACE2 shedding of S-protein-treated 293T cells. The aforementioned
results demonstrated that miR-28-3p inhibitor promoted ACE2
ectodomain shedding during S-protein treatment, which could be
abolished by ADAM17 knockdown. miR-28-3p exerts its function on
ACE2 ectodomain shedding through ADAM17.
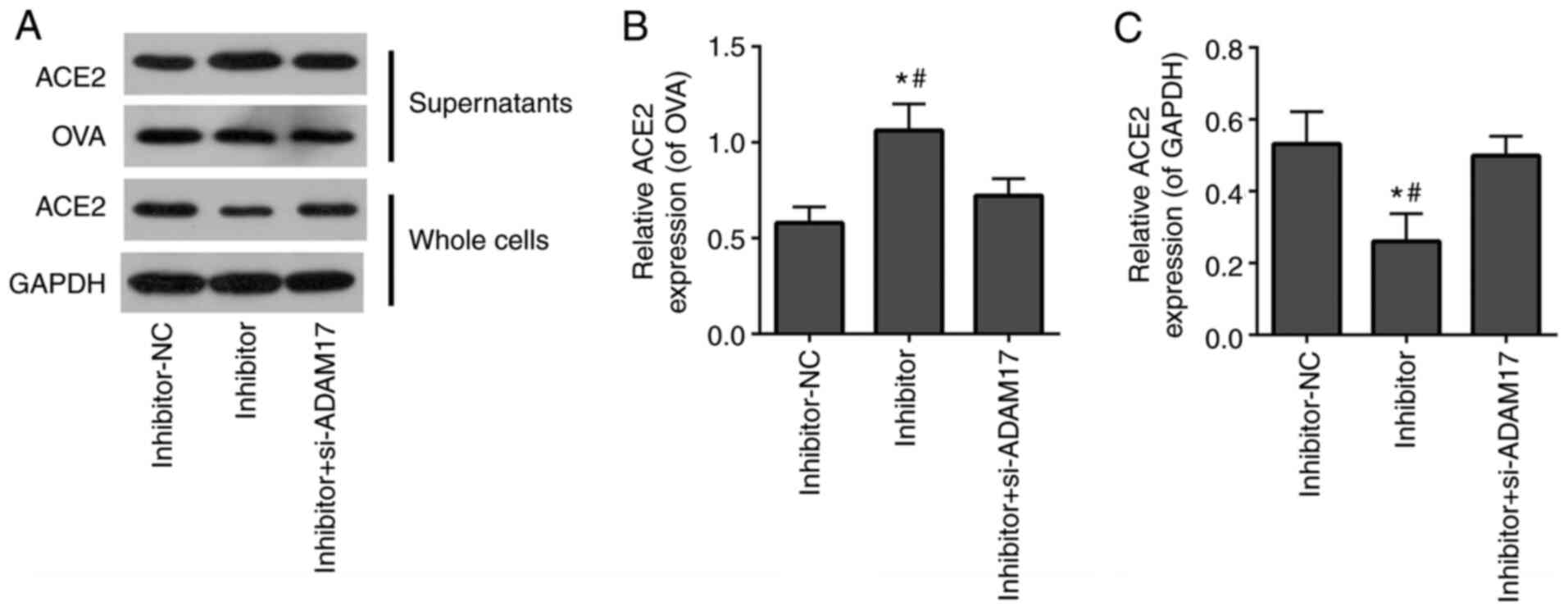 | Figure 6si-ADAM17 inhibits the effects of
miR-28-3p inhibitor on the shedding of the ACE2 ectodomain in 293T
cells treated with S-protein. (A) Western blotting results of ACE2,
OVA, GAPDH expression in supernatants and whole cells of
inhibitor-NC, inhibitor and inhibitor + si-ADAM17 groups. (B)
Quantification of ACE2/OVA in supernatants of inhibitor-NC,
inhibitor and inhibitor + si-ADAM17 groups. (C) Quantification of
ACE2/GAPDH in whole cells inhibitor-NC, inhibitor and inhibitor +
si-ADAM17 groups. #P<0.05 vs. inhibitor+si-ADAM17;
*P<0.05 vs. mimic-NC. ACE2, angiotensin-converting
enzyme 2; S-protein, spike protein; ADAM17, A disintegrin and
metalloproteinase 17; NC, negative control; si, small interfering;
OVA, ovalbumin. |
Discussion
ACE2, a homologue of ACE, serves a pivotal role in
balancing responses initiated from ACE, hydrolyses Angiotensin (Ang
I) to generate Ang-(1-9) and Ang II to generate Ang-(1-7),
which maintain the normal functioning of the cardiovascular system,
kidney and lung (37). In
addition, ACE2 is a receptor for the S-protein of SARS-CoV-2
(38). The binding affinity of
SARS-CoV-2 with ACE2 appears stronger compared with that of
SARS-CoV and has been reported to be 10- to 20-fold higher
(9,39), which aids the quick spreading of
the SARS-CoV-2 virus (40).
Meanwhile, similar to SARS-CoV, SARS-CoV-2 invades cells via the
binding of S-protein to the ACE2, followed by subsequent
downregulation of surface ACE2 expression (40). Previous studies have demonstrated
that ADAM17 is involved in regulating of ACE2 ectodomain shedding
during SARS-CoV-2 invading cells (3,41,42). ADAM17 cleaves ACE2, which results
in shedding of the ACE2 ectodomain into cell culture supernatants
and S-protein stimulates ADAM17-dependent ACE2 cleavage (43). ADAM17 is a member of the ADAM
family of proteinases, which also includes ADAM10, ADAM9 and ADAM12
(44). Lambert et al
(13) found that shedding of the
ACE2 ectodomain was significantly inhibited using a mixed
ADAM10/ADAM17 inhibitor (GW280264X), but was unaffected by a
selective ADAM10 inhibitor (GI254023X) alone. Upregulation of
ADAM-17 activity facilitated SARS-CoV viral entry, while knockdown
of ADAM17 by siRNA severely attenuated SARS-CoV cellular entry
(9). These results suggested
ADAM17 may serve a major role in the cleavage and shedding of the
ACE2 ectodomain (9,13). However, it is unknown which
molecular mechanisms initiate ADAM17 to serve such an important
role in ACE2 ectodomain shedding during SARS-CoV-2 virus
infection.
The findings of the present study demonstrated that
ADAM17 expression was upregulated in S-protein-treated 293T cells
compared with the non-treatment control group. In addition, it was
also demonstrated that ACE2 ectodomain shedding was increased
following treatment with S-protein, while knockdown of ADAM17
expression by siRNA led to a reduction in ACE2 ectodomain shedding,
which was consistent with previous research (9,34). The findings of the present study
confirmed the previous conclusions that inhibition of ADAM17 may
reduce shedding of the ACE2 ectodomain and have a protective effect
against SARS-CoV infection. Targeting ADAM17 expression may be a
potential treatment strategy for SARS-CoV-2.
It has been known that the S-protein stimulates
ADAM17-dependent ACE2 cleavage, which drives virus entry into cells
(43). However, there is little
known about how the S-protein regulates ADAM17-dependent ACE2
cleavage (12). There has been a
recent explosion of knowledge in the field of miRNAs regulating
viral infection. For example, miR-27b is widely known as an
antiviral miRNA which reduces HIV-1 replication (45) and miR-122 can inhibit hepatitis C
viral replication (46). In one
study, miR-1246 regulated the expression of ACE2 in patients with
SARS-CoV-2 infection (47).
The present study first observed that miR-28-3p
expression was downregulated in S-protein treated 293T cells,
miR-28-3p expression was negatively correlated with ADAM17
expression (Fig. 1D) and
miR-28-3p mimic reduced ACE2 ectodomain shedding (Fig. 3A). Further bioinformatics
analysis demonstrated that ADAM17 was a potential target of
miR-28-3p (Fig. 4A). It was
hypothesized that miR-28-3p participated in ADAM17-dependent ACE2
cleavage by S-protein stimulation. Further experimental findings of
the present study suggested that miR-28-3p did participate in
ADAM17-dependent ACE2 cleavage by S-protein stimulation.
In conclusion, the present study demonstrated that
the S-protein of SARS-Cov-2 promoted ADAM17-dependent ACE2
ectodomain shedding through downregulation of miR-28-3p, which may
provide a novel therapeutic strategy for treatment of SARS-CoV-2
infections. However, it remains unclear whether miR-28-3p inhibits
ACE2 ectodomain shedding in 293T cells in the absence of S-protein
and is miR-28-3p the only one miRNA that is downregulated by
S-protein during SARS-Cov-2 invading cells. Future studies should
perform infectivity experiments to further verify the findings of
the present study and to elucidate the mechanisms of how the
S-protein downregulates miR-28-3p.
Availability of data and materials
The datasets use/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YX was responsible for the concept and study design
and provided critical input and reviewed the manuscript for
important intellectual content. YL performed the experiments,
collected, analyzed and interpreted the data and wrote the
manuscript. YX and XL confirmed the authenticity of all the raw
data. All authors have read and approved the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
No funding was received.
Abbreviations:
|
SARS-CoV-2
|
syndrome coronavirus 2
|
|
ACE2
|
angiotensin-converting enzyme 2
|
|
S-protein
|
spike protein
|
|
ADAM17
|
A disintegrin and metalloproteinase
17
|
|
RT-qPCR
|
reverse-transcription quantitative
polymerase chain reaction
|
|
WB
|
western blotting
|
|
3′-UTR
|
3′ untranslated regions
|
|
WT
|
wild type
|
|
Mut
|
mutant
|
References
|
1
|
Horton R: Offline: 2019-nCoV
outbreak-early lessons. Lancet. 395:3222020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
World Health Organization (WHO):
Coronavirus disease (COVID-2019) situation report. WHO; Geneva:
2021, https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports
Accessed July 19, 2021.
|
|
3
|
Wan MY, Zhao R, Gao LJ, Gao XF, Wang DP
and Cao JM: SARS-CoV-2: Structure, biology, and structure-based
therapeutics development. Front Cell Infect Microbiol.
10:5872692020. View Article : Google Scholar
|
|
4
|
Hoffmann M, Kleine-Weber H, Schroeder S,
Krüger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu NH,
Nitsche A, et al: SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2
and is blocked by a clinically proven protease inhibitor. Cell.
181:271–280.e8. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xiao L, Sakagami H and Miwa N: ACE2: The
key molecule for understanding the pathophysiology of severe and
critical conditions of COVID-19: Demon or angel? Viruses.
12:4912020. View Article : Google Scholar
|
|
6
|
Kuba K, Imai Y and Penninger JM: Multiple
functions of angiotensin-converting enzyme 2 and its relevance in
cardiovascular diseases. Circ J. 77:301–308. 2013. View Article : Google Scholar
|
|
7
|
Patel VB, Zhong JC, Grant MB and Oudit GY:
Role of the ACE2/angiotensin 1-7 axis of the renin-angiotensin
system in heart failure. Circ Res. 118:1313–1326. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Turner AJ, Hiscox JA and Hooper NM: ACE2:
From vasopeptidase to SARS virus receptor. Trends Pharmacol Sci.
25:291–294. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gheblawi M, Wang K, Viveiros A, Nguyen Q,
Zhong JC, Turner AJ, Raizada MK, Grant MB and Oudit GY:
Angiotensin-converting enzyme 2: SARS-CoV-2 receptor and regulator
of the renin-angiotensin system: Celebrating the 20th anniversary
of the discovery of ACE2. Circ Res. 126:1456–1474. 2020. View Article : Google Scholar
|
|
10
|
Danilczyk U and Penninger JM:
Angiotensin-converting enzyme II in the heart and the kidney. Circ
Res. 98:463–471. 2006. View Article : Google Scholar
|
|
11
|
Su H, Yang M, Wan C, Yi LX, Tang F, Zhu
HY, Yi F, Yang HC, Fogo AB, Nie X and Zhang C: Renal
histopathological analysis of 26 postmortem findings of patients
with COVID-19 in China. Kidney Int. 98:219–227. 2020. View Article : Google Scholar
|
|
12
|
Zipeto D, Palmeira JDF, Argañaraz GA and
Argañaraz ER: ACE2/ADAM17/TMPRSS2 interplay may be the main risk
factor for COVID-19. Front Immunol. 11:5767452020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lambert DW, Yarski M, Warner FJ, hornhill
P, Parkin ET, Smith AI, Hooper NM and Turner AJ: Tumor necrosis
factor-alpha convertase (ADAM17) mediates regulated ectodomain
shedding of the severe-acute respiratory syndrome-coronavirus
(SARS-CoV) receptor, angiotensin-converting enzyme-2 (ACE2). J Biol
Chem. 280:30113–30119. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kuba K, Imai Y, Rao S, Gao H, Guo F, Guan
B, Huan Y, Yang P, Zhang Y, Deng W, et al: A crucial role of
angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced
lung injury. Nat Med. 11:875–879. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang S, Guo F, Liu K, Wang H, Rao S, Yang
P and Jiang C: Endocytosis of the receptor-binding domain of
SARS-CoV spike protein together with virus receptor ACE2. Virus
Res. 136:8–15. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Patel VB, Clarke N, Wang Z, Fan D,
Parajuli N, Basu R, Putko B, Kassiri Z, Turner AJ and Oudit GY:
Angiotensin ii induced proteolytic cleavage of myocardial ACE2 is
mediated by TACE/ADAM-17: A positive feedback mechanism in the RAS.
J Mol Cell Cardiol. 66:167–176. 2014. View Article : Google Scholar
|
|
17
|
Palau V, Riera M and Soler MJ: ADAM17
inhibition may exert a protective effect on COVID-19. Nephrol Dial
Transplant. 35:1071–1072. 2020. View Article : Google Scholar
|
|
18
|
Saliminejad K, Khorram Khorshid HR,
Soleymani Fard S and Ghaffari SH: An overview of microRNAs:
Biology, functions, therapeutics, and analysis methods. J Cell
Physiol. 234:5451–5465. 2019. View Article : Google Scholar
|
|
19
|
Biswas S, Haleyurgirisetty M, Lee S,
Hewlett I and Devadas K: Development and validation of plasma miRNA
biomarker signature panel for the detection of early HIV-1
infection. EBioMedicine. 43:307–316. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Park S, Eom K, Kim J, Bang H, Wang HY, Ahn
S, Kim G, Jang H, Kim S, Lee D, et al: MiR-9, miR-21, and miR-155
as potential biomarkers for HPV positive and negative cervical
cancer. BMC cancer. 17:6582017. View Article : Google Scholar :
|
|
21
|
Khan MA, Sany MRU, Islam MS and Islam
ABMMK: Epigenetic regulator mirna pattern differences among
SARS-CoV, SARS-CoV-2, and SARS-CoV-2 World-Wide isolates delineated
the mystery behind the epic pathogenicity and distinct clinical
characteristics of pandemic COVID-19. Front Genet. 11:7652020.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang G, Dong F, Xu Z, Sharma S, Hu X, Chen
D, Zhang L, Zhang J and Dong Q: MicroRNA profile in HBV-induced
infection and hepatocellular carcinoma. BMC cancer. 17:8052017.
View Article : Google Scholar :
|
|
23
|
Bai XT and Nicot C: miR-28-3p is a
cellular restriction factor that inhibits human T cell leukemia
virus, type 1 (HTLV-1) replication and virus infection. J Biol
Chem. 290:5381–5390. 2015. View Article : Google Scholar
|
|
24
|
Ma L, Zhang YF and Hu F: miR-28-5p
inhibits the migration of breast cancer by regulating WSB2. Int J
Mol Med. 46:1562–1570. 2020.PubMed/NCBI
|
|
25
|
Zhu G, Wang Z, Mijiti M, Du G, Li Y and
Dangmurenjiafu G: MiR-28-5p promotes human glioblastoma cell growth
through inactivation of FOXO1. Int J Clin Exp Pathol. 12:2972–2980.
2019.
|
|
26
|
Fan HN, Liao XH, Zhang J and Zheng HM:
Macrophages promote cell proliferation in colorectal cancer via
IL-1β-mediated downregulation of miR-28-3p. J Biol Regul Homeost
Agents. 34:1657–1668. 2020.PubMed/NCBI
|
|
27
|
Zhou X, Wen W, Shan X, Qian J, Li H, Jiang
T, Wang W, Cheng W, Wang F, Qi L, et al: MiR-28-3p as a potential
plasma marker in diagnosis of pulmonary embolism. Thromb Res.
138:91–95. 2016. View Article : Google Scholar
|
|
28
|
Tiwari A, Mukherjee B and Dixit M:
MicroRNA key to angiogenesis regulation: MiRNA biology and therapy.
Curr Cancer Drug Targets. 18:266–277. 2018. View Article : Google Scholar
|
|
29
|
Purohit V, Rapaka RS, Rutter J and
Shurtleff D: Do opioids activate latent HIV-1 by down-regulating
anti-HIV microRNAs? J Neuroimmune Pharmacol. 7:519–523. 2012.
View Article : Google Scholar
|
|
30
|
Tandon R, Sharp JS, Zhang F, Pomin VH,
Ashpole NM, Mitra D, Jin W, Liu H, Sharma P and Linhardt RJ:
Effective inhibition of SARS-CoV-2 entry by heparin and enoxaparin
derivatives. J Virol. 95:e019872021. View Article : Google Scholar
|
|
31
|
Zhang Y, Hu S, Wang J, Xue Z, Wang C and
Wang N: Dexamethasone inhibits SARS-CoV-2 spike pseudotyped virus
viropexis by binding to ACE2. Virology. 554:83–88. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hu S, Wang J, Zhang Y, Bai H, Wang C, Wang
N and He L: Three salvianolic acids inhibit 2019-nCoV spike
pseudovirus viropexis by binding to both its RBD and receptor ACE2.
J Med Virol. 93:3143–3151. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gao J, Ding Y, Wang Y, Liang P, Zhang L
and Liu R: Oroxylin A is a severe acute respiratory syndrome
coronavirus 2-spiked pseudotyped virus blocker obtained from Radix
Scutellariae using angiotensin-converting enzyme II/cell membrane
chromatography. Phytother Res. 35:3194–3204. 2021. View Article : Google Scholar
|
|
34
|
Haga S, Yamamoto N, Nakai-Murakami C,
Osawa Y, Tokunaga K, Sata T, Yamamoto N, Sasazuki T and Ishizaka Y:
Modulation of TNF-alpha-converting enzyme by the spike protein of
SARS-CoV and ACE2 induces TNF-alpha production and facilitates
viral entry. Proc Natl Acad Sci USA. 105:7809–7814. 2008.
View Article : Google Scholar
|
|
35
|
Yang ZY, Huang Y, Ganesh L, Leung K, Kong
WP, Schwartz O, Subbarao K and Nabel GJ: pH-dependent entry of
severe acute respiratory syndrome coronavirus is mediated by the
spike glycoprotein and enhanced by dendritic cell transfer through
DC-SIGN. J Virol. 78:5642–5650. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
37
|
Wu Y: Compensation of ACE2 function for
possible clinical management of 2019-nCoV-induced acute lung
injury. Virol Sin. 35:256–258. 2020. View Article : Google Scholar :
|
|
38
|
Moore MJ, Dorfman T, Li W, Wong SK, Li Y,
Kuhn JH, Coderre J, Vasilieva N, Han Z, Greenough TC, et al:
Retroviruses pseudotyped with the severe acute respiratory syndrome
coronavirus spike protein efficiently infect cells expressing
angiotensin-converting enzyme 2. J Virol. 78:10628–10635. 2004.
View Article : Google Scholar
|
|
39
|
Wrapp D, Wang N, Corbett KS, Goldsmith JA,
Hsieh CL, Abiona O, Graham BS and McLellan JS: Cryo-EM structure of
the 2019-nCoV spike in the prefusion conformation. Science.
367:1260–1263. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang Q, Zhang Y, Wu L, Niu S, Song C,
Zhang Z, Lu G, Qiao C, Hu Y, Yuen KY, et al: Structural and
functional basis of SARS-CoV-2 entry by using Human ACE2. Cell.
181:894–904.e9. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Taneera J, El-Huneidi W, Hamad M, Mohammed
AK, Elaraby E and Hachim MY: Expression profile of SARS-CoV-2 host
receptors in human pancreatic islets revealed upregulation of ACE2
in diabetic donors. Biology (Basel). 9:2152020.
|
|
42
|
de Loyola MB, Dos Reis TTA, de Oliveira
GXLM, da Fonseca Palmeira J, Argañaraz GA and Argañaraz ER:
Alpha-1-antitrypsin: A possible host protective factor against
Covid-19. Rev Med Virol. 31:e21572021. View Article : Google Scholar
|
|
43
|
Heurich A, Hofmann-Winkler H, Gierer S,
Liepold T, Jahn O and Pöhlmann S: TMPRSS2 and ADAM17 cleave ACE2
differentially and only proteolysis by TMPRSS2 augments entry
driven by the severe acute respiratory syndrome coronavirus spike
protein. J Virol. 88:1293–1307. 2014. View Article : Google Scholar :
|
|
44
|
Zunke F and Rose-John S: The shedding
protease ADAM17: Physiology and pathophysiology. Biochim Biophys
Acta Mol Cell Res. 1864:2059–2070. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Das G, Mukherjee N and Ghosh S:
Neurological insights of COVID-19 pandemic. ACS Chem Neurosci.
11:1206–1209. 2020. View Article : Google Scholar
|
|
46
|
Trobaugh DW and Klimstra WB: MicroRNA
regulation of RNA virus replication and pathogenesis. Trends Mol
Med. 23:80–93. 2017. View Article : Google Scholar
|
|
47
|
Zhang H, Rostami MR, Leopold PL, Mezey JG,
O'Beirne SL, Strulovici-Barel Y and Crystal RG: Expression of the
SARS-CoV-2 ACE2 receptor in the human airway epithelium. Am J
Respir Crit Care Med. 202:219–229. 2020. View Article : Google Scholar : PubMed/NCBI
|