Introduction
Despite accounting for only a small percentage
(5-8%) of liver cells, hepatic stellate cells (HSCs) are important
for liver cell regeneration and development of liver fibrosis
(1,2). Specifically, liver injuries
facilitate the release of cytokines such as tumor necrosis factor-α
(TNF-α) and transforming growth factor-β (TGF-β) from hepatocytes
and Kupffer cells, as well as the release of reactive oxygen
species (ROS), causing resting HSCs to transform into α-smooth
muscle actin (α-SMA)-expressing myofibroblast-like cells. As a
consequence, HSCs proliferate aggressively and produce large
quantities of extracellular matrix proteins, primarily collagen,
which ultimately leads to liver fibrosis (3,4).
Liver fibrosis treatment is often delayed, and there is a high
incidence of liver cancer in patients with liver fibrosis (5). It is therefore crucial to
understand the mechanisms of HSC activation and inflammation, in
order to develop novel diagnostic and treatment strategies for
liver fibrosis.
Extensive research underlines the link between long
non-coding RNAs (lncRNAs) and liver fibrosis development,
suggesting that lncRNAs have potential value in diagnosis,
prognosis and treatment of liver fibrosis (6). For example, lncRNA H19 and liver
fibrosis-associated lncRNA were reported to accelerate or
decelerate liver fibrosis progression, respectively, by altering
TGF-β1/Smad signaling and collagen production (7-10). Furthermore, lncRNA metastasis
associated lung adenocarcinoma transcript 1 (MALAT1), also known as
nuclear enriched abundant transcript 2, was reported to be a
negative regulator of sirutin 1 (11), upregulated expression of which
induces Smad3 deacetylation and deactivation of HSC (12). MALAT1 also promotes HSC
activation by targeting microRNA (miR/miRNA)-101b and increasing
Rac1 expression (13), and
increasing collagen 1 (COL1)A2 expression by downregulating miR-26b
(14). miR-181a, although
sponged by MALAT1 in myeloma cells (15), was reported to rescue the
aberrant inflammatory cascade and over-proliferation of HSCs by
decreasing TLR4/NF-κB signaling, thereby limiting liver fibrosis
progression (16,17). However, there is no convincing
evidence suggesting a role for the lncRNA MALAT1/miR-181a axis in
the etiology of liver fibrosis, to the best of our knowledge.
Therefore, the aim of the present study was to elucidate the
contribution of the MALAT1/miR-181a axis in the abnormal
inflammation and HSC activation underlying the pathogenesis of
liver fibrosis.
Materials and methods
Clinical samples
In total, 151 patients with liver fibrosis or
chronic hepatitis B virus (HBV)-associated liver fibrosis were
recruited at the Affiliated Hospital of Guizhou Medical University
(Guiyang, China), between October 2018 and December 2019. The age
range of the patient cohort was 25-68 years, with a median age of
51. The cohort consisted of 116 males and 35 females. The inclusion
criteria were: i) Liver fibrosis was histopathologically confirmed
(Fig. S1); ii) Hepatitis B
antigen remained positive for >6 months; iii) HBV DNA was ≥105
IU/ml; iv) alanine aminotransferase (ALT) levels were ~1 fold
higher than the normal range; and v) patients were fully informed
about this program. The present study was approved by the Ethics
Committee of the Affiliated Hospital of Guizhou Medical University.
Conversely, participants were excluded if: i) They were infected
with hepatitis C virus, hepatitis A virus, hepatitis D virus or
other hepatotropic viruses; ii) their livers were damaged due to
alcohol or drug use, or immune disorders; iii) they had diabetes,
hypertension, malignant tumors or any other severe lesions in the
heart, kidneys or lungs; iv) they received anti-viral treatment
within the preceding 6 months; or v) they suffered from acute HBV
infection, liver failure or cirrhosis. Concurrently, 93 healthy
individuals without a family history of HBV and with normal liver
function were recruited as controls. The age range of control group
was 22-73, with a median age of 46, consisting of 67 males and 26
females. Written informed consent was obtained from all
participants included in the present study.
Venous blood was taken from the medical cubital vein
of each subject on an empty stomach, and serum levels of ALT,
aspartate aminotransferase (AST) and albumin were determined using
an automatic biochemical analyzer (model: 7600P, Hitachi, Ltd.),
according to the manufacturer's protocol (Beijing Solarbio Science
& Technology Co., Ltd.). Serum levels of hyaluronic acid (HA),
procollagen III (PC-III), collagen type IV (C-IV) and laminin (LN)
were detected by chemiluminescence immunoassay (CL-2000I; Mindray
Medical International, Ltd.). Serum levels of tumor necrosis
factor-α (TNF-α), interleukin-6 (IL-6) and high-sensitivity
C-reactive protein (hs-CRP) were determined using specific ELISA
kits (cat. nos. 1749-48T, A100084-96T and A099284-96T;
Affandi-e).
Cell culture
The human LX2 HSC cell line (Shanghai Advanced
Research Institute, The Cell Bank of Type Culture Collection of The
Chinese Academy of Sciences) was cultured in DMEM with 2% FBS in a
humidified incubator at 37°C incubator supplied with 5%
CO2.
Cell treatment with TGF-β1 or LPS
LX2 cells were treated with recombinant human TGF-β1
(PeproTech, Inc.) at 5, 10 and 15 ng/ml for 24 h, as described
previously (18). LX2 cells were
treated with LPS (Sigma-Aldrich; Merck KGaA) at a final
concentrations of 0.01, 0.1 and 1 µg/ml for 24 h.
Cell transfection
LX2 cells in the logarithmic growth phase, were
seeded at a density of 6×105 cells/well, were separately
transfected with pcDNA3.1-MALAT1, small interfering (si) RNA
against MALAT1-1 (si-MALAT1-1), si-MALAT1-2 miR-181a-5p mimic or
miR-181a-5p inhibitor (all synthesized by Shanghai GenePharma, Co.,
Ltd.) for 48 h, using Lipofectamine 2000 according to the
manufacturer's protocol (Invitrogen; Thermo Fisher Scientific,
Inc.). The working concentration of plasmids was 100 nM. The
sequences of the constructs used were: si-MALAT1-1: 5′-CCU CAG ACA
GGU AUC UCU U-3′; si-MALAT1-2 5′-GAU CCA UAA UCG GUU UCA A-3′;
miR-181a-5p mimic, 5′-AAC AUU CAA CGC UGU CGG UGA GU-3′ and 3′-UUG
UAA GUU GCG ACA GCC ACU CA-5′; miR-mimic NC, 5′-UUU GUA CUA CAC AAA
AGU ACU G-3′ and 3′-AAA CAU GAU GUG UUU UCA UGA C-5′; miR-181a-5p
inhibitor 5′-ACU CAC CGA CAG CGU UGA AUG UU-3′; and miR-inhibitor
NC 5′-CAG UAC UUU UGU GUA GUA CAA A-3′.
MTT assay
LX2 cells were seeded into 96-well plates at a
density of 4×103 cells/well. Once they had adhered to
the plate, 20 µl MTT (5 mg/ml) was added for 4 h.
Subsequently, 150 µl DMSO (Sinopharm Chemical Reagent Co.,
Ltd.) was added to each well, and the mixture was shaken at 37°C
for 10 min. Absorbance was measured on a microplate reader (Elx800,
BioTek Instruments Inc.) at 490 nm.
Transwell assays
LX2 cells were resuspended in serum-free medium
(1×105 cells/ml) and seeded in the upper chamber of
24-well plate Transwell inserts (Corning, Inc.). A total of 500
µl supplemented medium was added to the lower chamber. After
24 h of incubation, the polycarbonate membranes and the cells
remaining in the upper chamber were removed, and the chambers were
immersed in crystal violet solution (Merck KGaA) for 10 min at
37°C, until cells in the lower chamber were dyed violet.
Wound healing assay
LX2 cells were seeded into 6-well plates and
cultured until 80% confluent. Culture medium was replaced with
serum-free medium and the cell layer was scratched using a 200
µl pipette tip, along the center of the wells. After 24 h,
images of the culture plates were taken using a light microscope.
(magnification, ×200).
Flow cytometry
After centrifugation at 42.62 × g for 5 min at room
temperature, LX2 cells were re-suspended in 1X binding buffer at a
density of 1× 106/ml. A total of 5 µl Annexin
V/FITC and 5 µl propidium iodide (BD Biosciences) were added
to 100 µl cell suspension, followed by incubation in the
dark at room temperature for 15 min. The proportion of apoptotic
cells was determined by flow cytometry (Coulter epics-XL; Beckman
Coulter, Inc.) and analyzed using CellQuest version 3.2 software
(BD Bioscience).
Colony formation assay
LX2 cells (2×103 cells/well) were seeded
in 6-well plates and cultured at 37°C in 5% CO2 for 12
days. Cells were fixed using 4% paraformaldehyde (Wuhan Boster
Biological Technology, Ltd.) for 20 min and then stained with 1%
crystal violet (Wuhan Boster Biological Technology, Ltd.) for 20
min, both at room temperature. Colonies consisting of >50 cells
were counted manually under a microscope (magnification, ×20;
BioTek Instruments, Inc.).
Reverse transcription-quantitative
(q)PCR
Total RNA was extracted from blood samples of
participants and LX2 cells using an easyPureBlood RNA kit (TransGen
Biotech, Co. Ltd.). RNA was reverse transcribed into cDNA using a
HiScript III 1st Strand cDNA Synthesis kit (Biotech Co., Ltd.)
according to the manufacturer's protocol, after which qPCR was
performed using SYBR-Green (Roche Diagnostics), with the following
thermocycling parameters: 95°C for 15 min; followed by 40 cycles of
94°C for 15 sec and 55°C for 30 sec; and a final extension step of
70°C for 60 sec. The primer sequences used are listed in Table SI. Expression of MALAT1 and
miR-181a-5p were normalized to GAPDH or U6, respectively, using the
2−∆∆Cq method (19).
Western blotting
Proteins (50 µg), extracted from blood
samples and LX2 cells using a protein extraction kit according to
the manufacturer's protocol (Applygen Technologies Inc.), were
loaded on a 10% SDS-gel, resolved using SDS-PAGE and then
transferred to PVDF membranes. The protein concentration was
determined using a BCA protein assay kit (Beyotime Institute of
Biotechnology). After blocking with 5% skimmed milk powder for 1.5
h at room temperature, the protein samples were incubated with
primary antibodies against NF-κB (1:1,000; cat. no. 8242; Cell
Signaling Technology, Inc.), toll like receptor (TLR) 4 (1:500;
cat. no. ab13556; Abcam), α-SMA (1:1,000; cat. no. 19245; Cell
Signaling Technology, Inc.), COL1 (1:100; cat. no. 72026; Cell
Signaling Technology, Inc.), tissue inhibitor of metalloprotease
(TIMP)-1 (1:1,000; cat. no. ab211926; Abcam), Bax (1:2,000; cat.
no. ab32503; Abcam), Bcl-2 (1:1,000; cat. no. ab32124; Abcam),
cleaved caspase-3 (1:500; cat. no. ab2302; Abcam), E-cadherin
(1:25; cat. no. ab227639; Abcam), Desmin (1:1,000; cat. no.
ab227224; Abcam), Vimentin (1:2,000; cat. no. ab92547; Abcam) and
GAPDH (1:2,500; cat. no. ab9485; Abcam) at 4°C overnight, and were
then incubated with HRP-conjugated goat anti-rabbit IgG (1:3,000;
cat. no. ab6721; Abcam) for 1 h at room temperature. Following
chemiluminescent detection, protein bands were quantified using the
Gel-pro software (version 3.1; Media Cybernetics, Inc.).
Dual-luciferase reporter gene assay
MALAT1 fragments that incorporated binding sites for
miR-181a-5p were amplified using PCR and cloned into a pmirGLO
vector (Promega Corporation). These were then mutated to remove the
predicted miR-181a-5p binding sites using site-directed mutagenesis
to construct a plasmid termed pmiRGLO-MALAT1-Mut. pmirGLO-MALAT1-Wt
or pmirGLO-MALAT1-Mut were co-transfected with miR-181a-5p mimic or
miR-NC into LX2 cells using Lipofectamine 2000, according to the
manufacturer's protocol (Invitrogen; Thermo Fisher Scientific,
Inc.). Renilla and firefly luciferase activities were detected
using a dual-luciferase reporter assay system, according to the
manufacturer's protocol (Promega Corporation).
RNA immunoprecipitation (RIP) assay
RIP from LX2 cells was performed using a Magna RIP™
RNA-Binding Protein Immunoprecipitation kit (MilliporeSigma), and
cell lysates were incubated with RIP buffer containing magnetic
beads conjugated with human anti-Ago2 antibody (MilliporeSigma) or
IgG (MilliporeSigma) at 4°C overnight. The resulting protein-RNA
complexes were digested with proteinase K to release RNA, which was
then qualified by RT-qPCR as described above.
Fluorescence in situ hybridization (FISH)
for MALAT1 and miR-181a-5p in LX2 cells
RNA FISH assays were performed using a FISH kit
(Creative Bioarray) according to the manufacturer's protocol. LX2
cells were incubated with 0.2 mol/l HCl and 0.3% Triton X-100 at
room temperature. After fixing cells with 4% formaldehyde at room
temperature for 10 min, RNA hybridization buffer was added to the
cells, and pre-hybridization was performed at 55°C for 2 h.
Subsequently, RNA probes, having been denatured at 85°C for 5 min
and 37°C for 2 min, were added and hybridization was performed in
the dark at 37°C overnight. The length of MALAT1 probe is 20 bp,
and the sequence is 5′-TATGACACTTTCCTTGCCCA-3′. The length of the
miR-181a-5p probe is 23 nt, and the sequence is 5′-TTG TTT GTT GCG
TCT GCC TCT CT-3′ with 23 bp. Images were collected on a confocal
laser scanning microscope (magnification: ×600; FV1000; Olympus
Corporation).
Statistical analysis
All data were statistically analyzed using SPSS
version 22.0 (IBM, Corp.). Continuous data (mean ± standard
deviation) were compared between groups using an unpaired Student's
t-test, and count data were compared using a χ2 test.
Pearson's correlation coefficients were calculated to assess
correlations between serum levels of lncRNA MALAT1, miR-181a-5p,
inflammatory factors and hepatic fibrosis indices, as well as
receiver operating characteristic (ROC) curves were constructed to
evaluate the diagnostic value of lncRNA MALAT1 and miR-181a-5p for
patients with liver fibrosis. P<0.05 was considered to indicate
a statistically significant difference.
Results
Clinical significance of lncRNA MALAT1
and miR-181a-5p in liver fibrosis
Patients with liver fibrosis had higher serum levels
of ALT and AST and lower serum levels of albumin than the healthy
volunteers, and the two populations did not have significantly
different mean ages or sex ratios (P>0.05; Table I). Serum levels of inflammatory
factors, including TNF-α, IL-6 and hs-CRP, as well as liver
fibrosis indicators, including HA, PC-III, C-IV and LN, were higher
in patients with liver fibrosis compared with the healthy controls
(P<0.05). Furthermore, the levels of MALAT1 in the serum of
patients with liver fibrosis was ~2X higher than that in the
healthy subjects (P<0.05), whereas serum miR-181a-5p levels were
significantly reduced in patients with liver fibrosis relative to
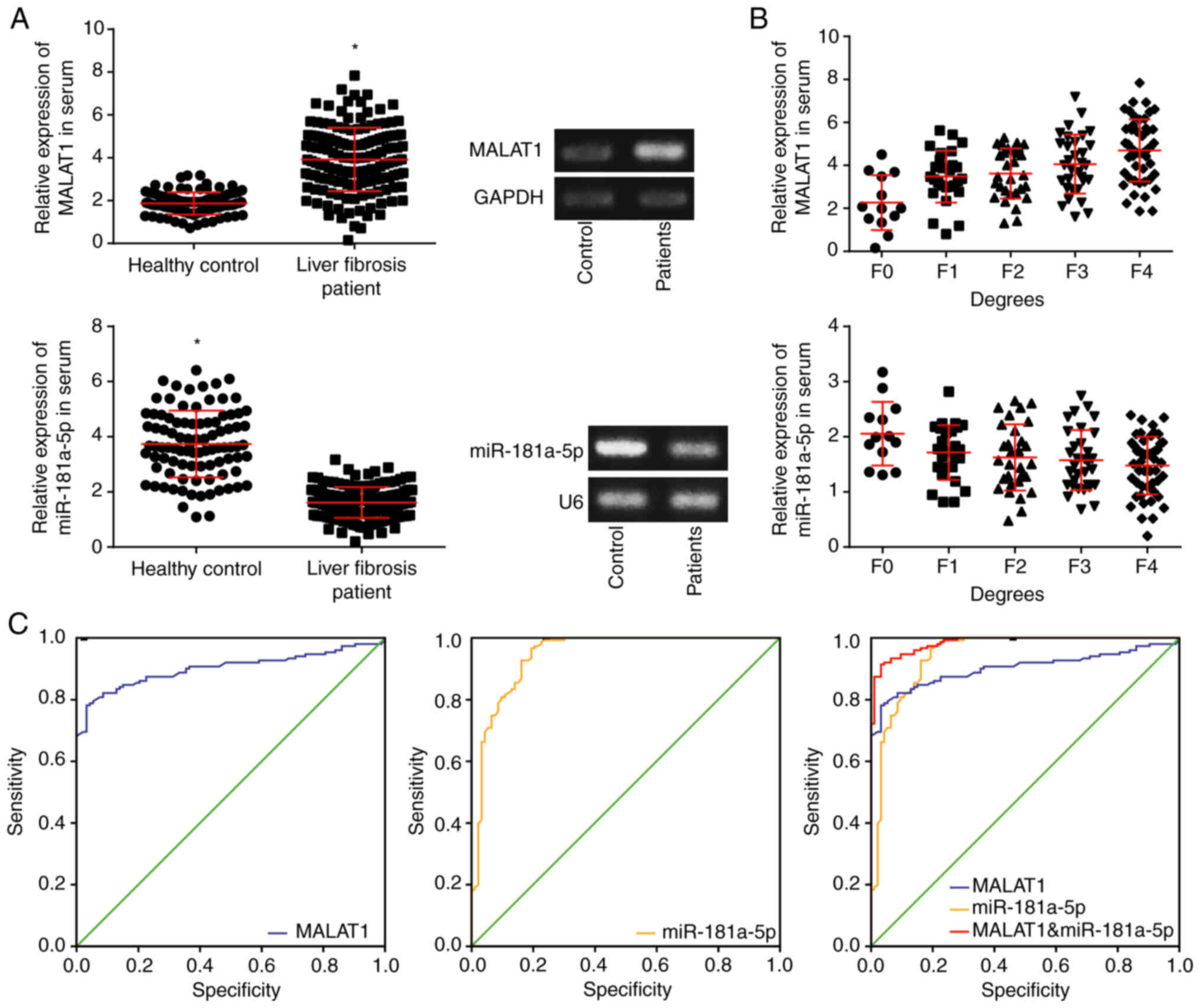
the healthy controls (P<0.05; Fig. 1A). MALAT1 and miR-181a-5p
expression levels were positively and negatively correlated,
respectively, with the severity of liver fibrosis (Fig. 1B). It was noteworthy that both
serum MALAT1 and miR-181a-5p had high area under ROC curve (AUC)
values of 0.901 and 0.944, respectively, suggesting that they may
be useful for diagnosis of liver fibrosis. MALAT1 in combination
with miR-181a-5p resulted in an even higher AUC value, 0.984
(Fig. 1C; Table II).
 | Table IComparison of baseline features
between patients with liver fibrosis and healthy volunteers. |
Table I
Comparison of baseline features
between patients with liver fibrosis and healthy volunteers.
|
Characteristics | Patients | Healthy
control |
χ2/t | P-value |
|---|
| Age, year | 43.96±10.35 | 43.58±11.29 | 0.27c | 0.788 |
| Sex | | | 6.94d | 0.008a |
| Male | 116 | 67 | | |
| Female | 35 | 26 | | |
| Stage | | | | |
| F0-F1 | 40 | - | - | - |
| F2-F3 | 62 | - | - | - |
| F4 | 49 | - | - | - |
| Alanine
transaminase, µg/l | 35.91±16.74 | 14.71±7.52 | 11.51c | <0.001b |
| Aspartate
aminotransferase, µg/l | 51.45±18.31 | 17.65±5.36 | 17.34c | <0.001b |
| Albumin, g/l | 45.26±16.82 | 76.38±4.05 | 17.52c | <0.001b |
| HBV-DNA,
log10 copies/ml | 5.25±1.83 | - | - | - |
| Tumor necrosis
factor-α, ng/l | 263.56±39.15 | 96.89±18.8 | 38.40c | <0.001b |
| Interleukin-6,
ng/l | 283.51±48.17 | 53.28±12.64 | 45.11c | <0.001b |
| High-sensitivity
C-reactive protein, ng/l | 349.34±52.14 | 66.25±19.52 | 50.20c | <0.001b |
| Serum hyaluronic
acid, µg/l | 106.34±31.47 | 15.06±4.63 | 27.77c | <0.001b |
| Procollagen III,
µg/l | 35.49±18.78 | 3.41±1.85 | 15.93c | <0.001b |
| Collagen type IV,
µg/l | 32.22±17.24 | 6.53±3.29 | 13.30c | <0.001b |
| Laminin,
µg/l | 10.17±2.44 | 3.31±1.12 | 25.49c | <0.001b |
| Table IIDiagnostic potential of lncRNA MALAT1
and miR-181-5p for liver fibrosis. |
Table II
Diagnostic potential of lncRNA MALAT1
and miR-181-5p for liver fibrosis.
| Biomarkers | Value | Sensitivity | Specificity | Area under the
curve | 95% confidence
interval |
|---|
| lncRNA MALAT1 | 2.81 | 0.781 | 0.968 | 0.901 | 0.862-0.941 |
|
microRNA-181-5p | 2.62 | 0.967 | 0.806 | 0.944 | 0.912-0.976 |
| lncRNA MALAT1 and
miR-181-5p | - | 0.914 | 0.968 | 0.984 | 0.973-0.995 |
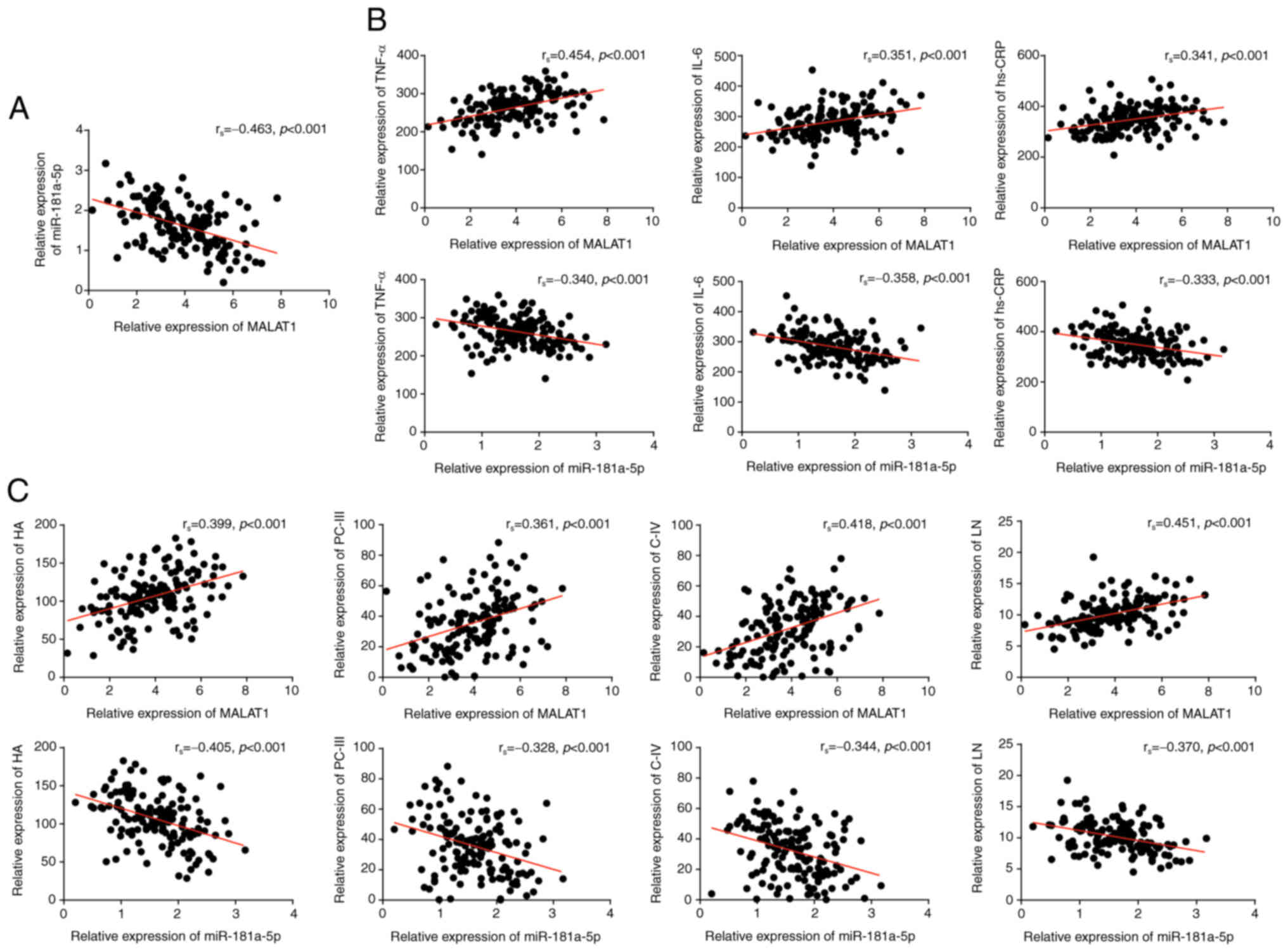
Serum levels of MALAT1 were negatively correlated
with serum levels of miR-181a-5p in the patients with liver
fibrosis (rs=−0.463; Fig.
2A). MALAT1 was positively correlated with serum TNF-α
(rs=0.454), IL-6 (rs=0.351) and hs-CRP
(rs=0.341), whereas serum miR-181a-5p levels were
negatively correlated with TNF-α (rs=−0.340), IL-6
(rs=−0.358) and hs-CRP (rs=−0.333) (Fig. 2B). Serum levels of the liver
fibrosis indicators HA, PC-III, C-IV and LN were positively
correlated with serum MALAT1 (HA, rs=0.399; PC-III,
rs= 0.361; C-IV, rs=0.418; LN,
rs=0.451), and negatively correlated with serum
miR-181a-5p levels (HA, rs=−0.405; PC-III,
rs= −0.328; C-IV, rs=−0.344; LN,
rs=−0.370) (Fig.
2C).
MALAT1 and miR-181a-5p regulate the
inflammatory response of HSCs
FISH data (Fig.
S2) suggested that MALAT1 (red fluorescence) and miR-181a-5p
(green fluorescence) were co-localized in LX2 cells. MALAT1 was
primarily localized to the nucleus, whereas miR-181a-5p was present
both in the nucleus and the cytoplasm.
Upon stimulation with 0.01, 0.1 and 1 µg/ml
LPS, MALAT1 expression in the LX2 cells increased (P<0.05),
whereas miR-181a-5p expression decreased (P<0.05; Fig. 3A). MALAT1 expression in LX2 cells
was increased after transfection of pcDNA3.1-MALAT1 (P<0.05),
and both si-MALAT1-1 and si-MALAT1-2 transfection, significantly
decreased MALAT1 expression (P<0.05). miR-181a-5p expression in
LX2 cells increased when miR-181a-5p mimics were transfected
(P<0.05), and decreased when miR-181a-5p inhibitor was
transfected (P<0.05; Fig.
3B).
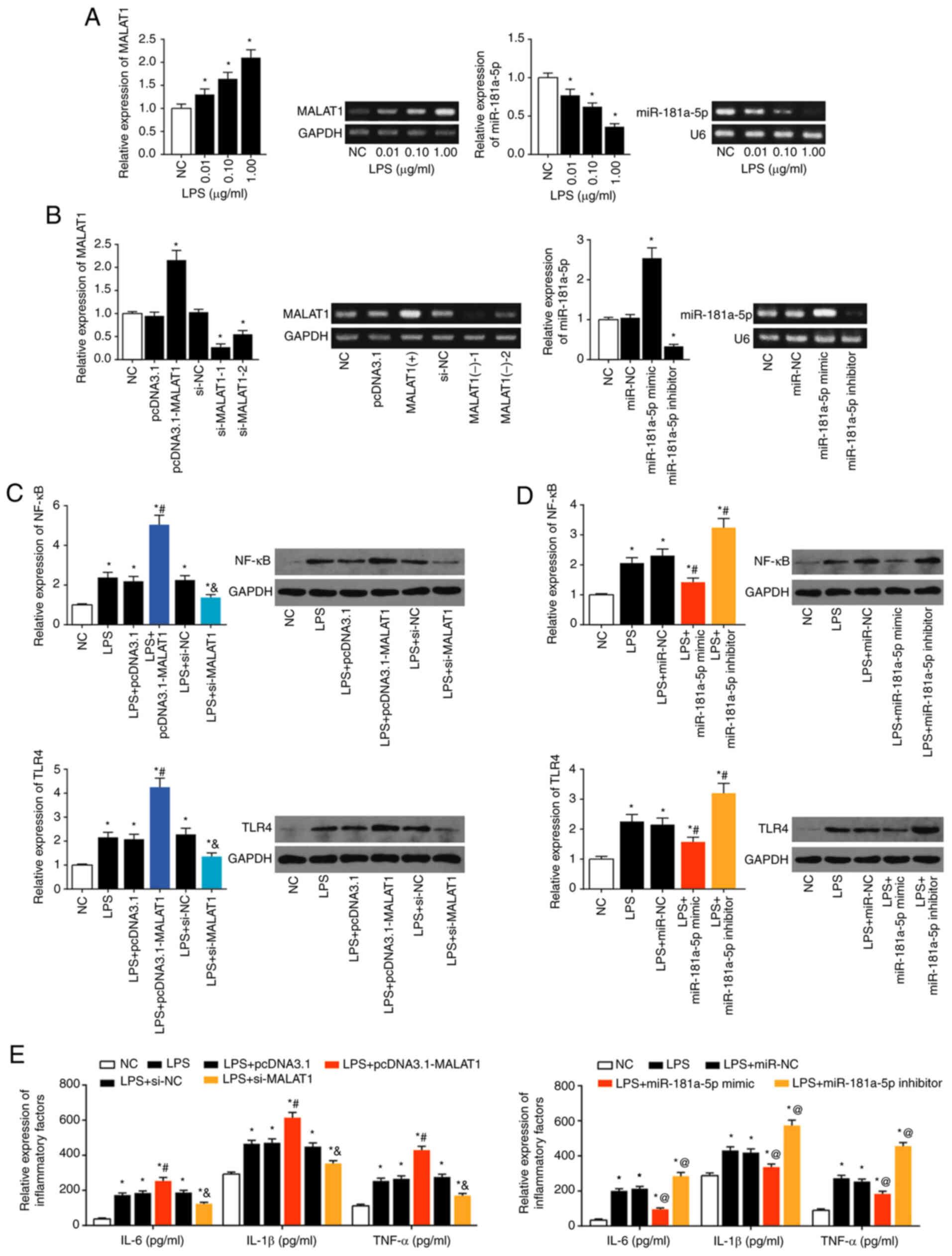 | Figure 3Effect of the lncRNA
MALAT1/miR-181a-5p axis on NF-κB/TLR4 signaling. (A) MALAT1 and
miR-181a-5p expressions were determined in LX2 cells with different
concentrations of LPS. *P<0.05 vs. NC. (B) MALAT1 and
miR-181a-5p expression was assessed after transfection of
pcDNA3.1-MALAT1, si-MALAT1-1, si-MALAT1-2, miR-181a-5p mimic and
miR-181a-5p inhibitor. *P<0.05 vs. NC. (C) NF-κB and
TLR4 expression levels were detected in the LX2 cells treated with
LPS, LPS+pcDNA3.1, LPS+pcDNA3.1-MALAT1, LPS+si-NC and
LPS+si-MALAT1. *P<0.05 vs. NC; #P<0.05
vs. LPS+pcDNA3.1; &P<0.05 vs. LPS+si-NC. (D)
NF-κB and TLR4 expression levels were compared in the LX2 cells
treated with, LPS, LPS+miR-NC, LPS+miR-181a-5p mimic and
LPS+miR-181a-5p inhibitor. *P<0.05 vs. NC;
#P<0.05 vs. LPS+miR-NC. (E) The levels of
inflammatory cytokines were assessed in LX2 cells treated with LPS,
LPS+pcDNA3.1, LPS+pcDNA3.1-MALAT1, LPS+si-NC and LPS+si-MALAT1 as
well as, LPS+miR-NC, LPS+miR-181a-5p mimic and LPS+miR-181a-5p
inhibitor. *P<0.05 vs. NC; #P<0.05 vs.
LPS+pcDNA3.1; &P<0.05 vs. LPS+si-NC;
@P<0.05 vs. miR-NC. lncRNA MALAT1, long non-coding
RNA metastasis associated lung adenocarcinoma transcript 1; miR,
microRNA; NF-κB, nuclear factor-κB; TLR4, toll-like receptor-4;
LPS, lipopolysaccharide; siRNA, small interfering RNA; NC, negative
control. |
Expression levels of TLR4 and NF-κB were increased
in LX2 cells after LPS exposure (P<0.05) (Fig. 3C). TLR4 and NF-κB were both
downregulated in the LPS+si-MALAT1 and LPS+miR-181a-5p mimic group,
yet were upregulated in the LPS+pcDNA3.1-MALAT1 and LPS+miR-181a-5p
inhibitor group, when compared with the LPS group (P<0.05;
Fig. 3C and D). IL-6, IL-1β and
TNF-α levels in LX2 cells were increased in the LPS group compared
to the control group (P<0.05; Fig. 3E). Levels of these cytokines were
increased in the LPS+pcDNA3.1-MALAT1 and LPS+miR-181a-5p inhibitor
group, and were decreased in the LPS+si-MALAT1 and LPS+miR-181a-5p
mimic group compared with the LPS group (P<0.05; Fig. 3E).
MALAT1 sponges miR-181a-5p and activates
TLR4/NF-κB signaling in HSCs
Using RIP assays with an anti-Ago2 antibody, it was
shown that MALAT1 interacted with Ago2, a vital constituent of the
RNA-induced silencing complex (Fig.
4A). Furthermore, luciferase activity of LX2 cells transfected
with pmirGLO-MALAT1-Wt+miR-181a-5p mimics was significantly reduced
compared with both pmirGLO-MALAT1-Wt+miR-NC and pmirGLO+miR-181a-5p
mimics (both P<0.05; Fig.
4B), suggesting that miR-181a-5p was sponged by MALAT1 in LX2
cells.
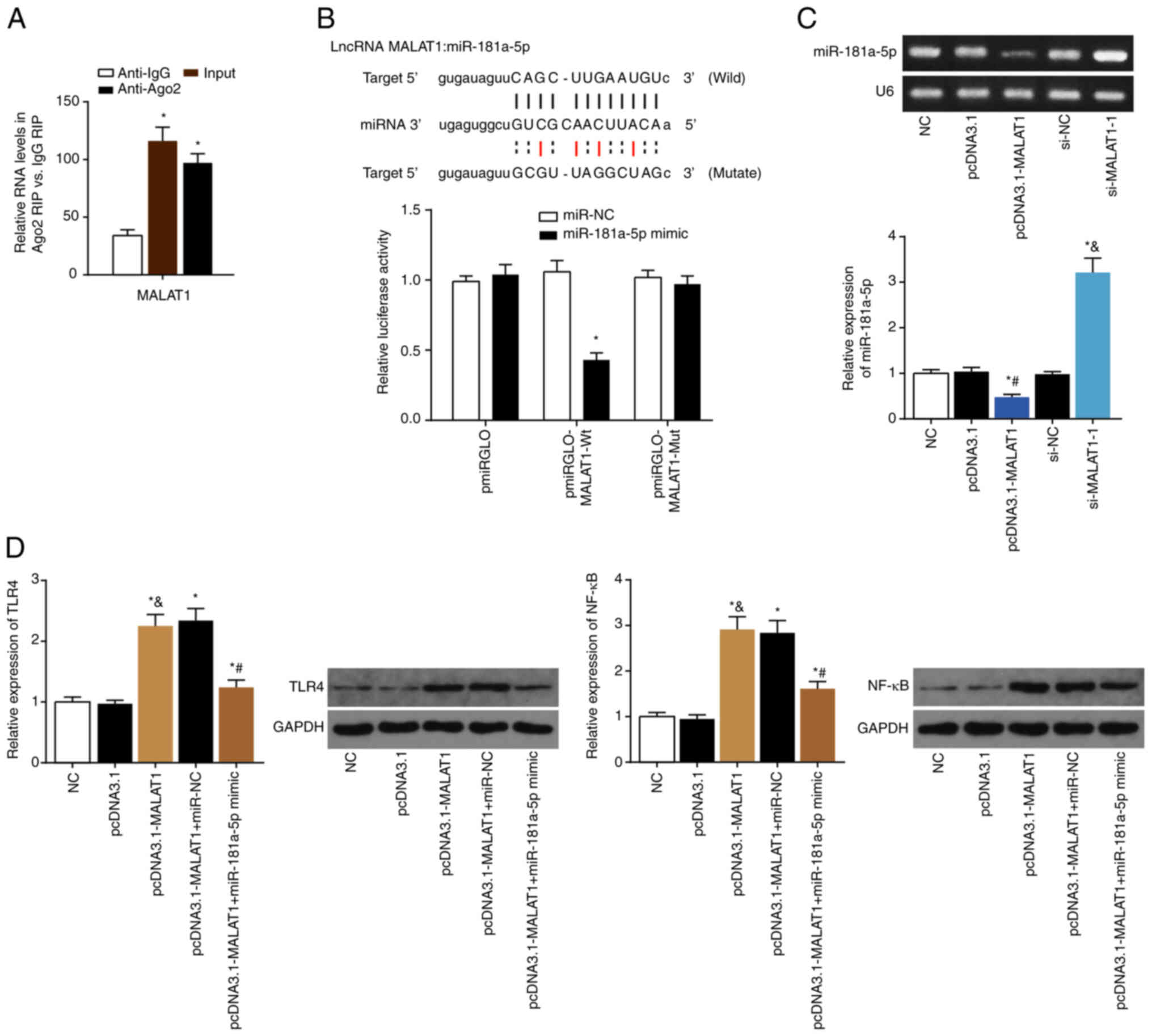 | Figure 4Sponging relationship between MALAT1
and miR-181a-5p in LX2 cells. (A) RNA immunoprecipitation assay was
performed in LX2 cell lysates with antibodies against Ago2/IgG, and
expression of MALAT1 was detected via PCR. (B) Luciferase activity
of LX2 cells was compared between the pmiRGLO-MALAT1-Wt+miR-181a-5p
mimic group and pmiRGLO-MALAT1-Wt+miR-NC group.
*P<0.05 vs. pmiRGLO-MALAT1-Wt+miR-NC. (C) miR-181a-5p
expression was determined in LX2 cells treated with pcDNA3.1,
pcDNA3.1-MALAT1, si-NC and si-MALAT1-1. *P<0.05 vs.
NC; #P<0.05 vs. pcDNA3.1; &P<0.05
vs. si-NC. (D) NF-κB and TLR4 expression levels in LX2 cells
transfected with pcDNA3.1, pcDNA3.1-MALAT1, pcDNA3.1-MALAT1+miR-NC
or pcDNA3.1-MALAT1+miR-181a-5p mimic. *P<0.05 vs. NC;
#P<0.05 vs. pcDNA3.1-MALAT1+miR-NC;
&P<0.05 vs. pcDNA3.1. MALAT1, metastasis
associated lung adenocarcinoma transcript 1; miR, microRNA; NF-κB,
nuclear factor-κB; TLR4, toll-like receptor-4; siRNA, small
interfering RNA; NC, negative control. |
Furthermore, miR-181a-5p expression was decreased in
LX2 cells transfected with pcDNA3.1-MALAT1 compared with the
pcDNA3.1 control (P<0.05), whereas miR-181a-5p expression was
increased in cells transfected with si-MALAT1 compared to the si-NC
control (P<0.05; Fig. 4C).
Expression of TLR4 and NF-κB were increased in LX2 cells
transfected with pcDNA3.1-MALAT1 (P<0.05), whereas transfection
of pcDNA3.1-MALAT1+miR-181a-5p mimic counteracted this increase
(P<0.05; Fig. 4D).
MALAT1 promotes collagen production,
viability, proliferation, migration and adhesion of HSCs
LX2 cells exposed to 5, 10 and 15 ng/ml TGF-β1 all
expressed higher levels of MALAT1 and lower levels of miR-181a-5p
than the control group (P<0.05; Fig. 5A). Additionally, 5 ng/ml TGF-β1
significantly increased the expression of TLR4 and NF-κB compared
with the control group (P<0.05; Fig. 5A). In the presence of TGF-β1,
TLR4 and NF-κB expression were significantly increased by
transfection of pcDNA3.1-MALAT1 and were significantly suppressed
by transfection of si-MALAT1, compared with TGF-β1 alone
(P<0.05). Moreover, expression of α-SMA, COL1 and TIMP-1 were
increased in LX2 cells upon stimulation with 5 ng/ml TGF-β1
(P<0.05), and they were increased further by simultaneous
transfection with pcDNA3.1-MALAT1 (P<0.05; Fig. 5C).
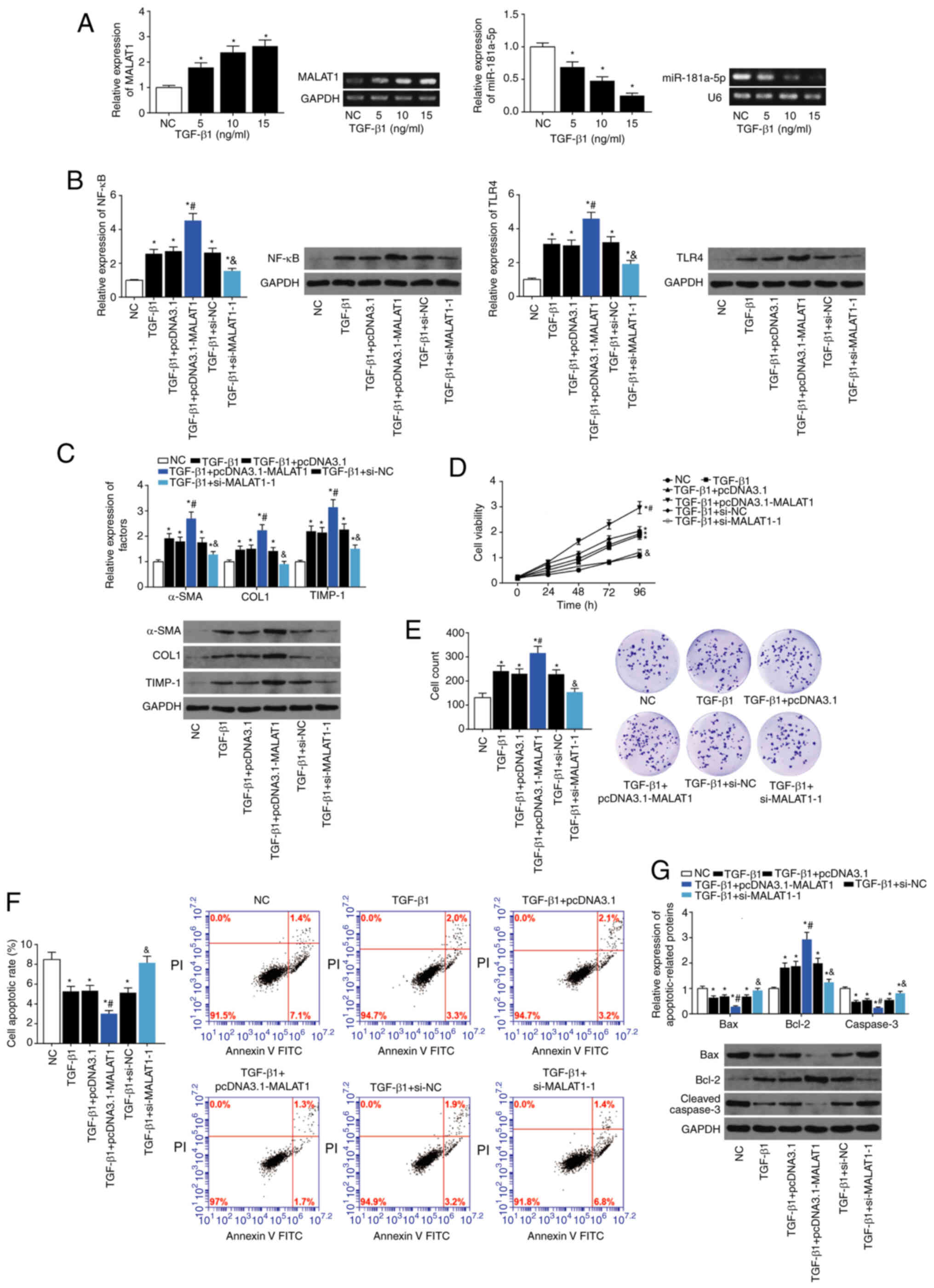 | Figure 5Impact of lncRNA MALAT1 on the
viability and proliferation of LX2 cells. (A) Expression levels of
MALAT1 and miR-181a-5p were estimated in LX2 cells treated with
different concentrations of TGF-β1. *P<0.05 vs. NC.
(B) Expression levels of NF-κB and TLR4 were detected in LX2 cells
treated with NC, TGF-β1, TGF-β1+pcDNA3.1, TGF-β1+pcDNA3.1-MALAT1,
TGF-β1+si-NC and TGF-β1+si-MALAT1. *P<0.05 vs. NC;
#P<0.05 vs. TGF-β1+pcDNA3.1;
&P<0.05 vs. TGF-β1+si-NC. (C) α-SMA, COL1 and
TIMP-1 levels were evaluated in LX2 treated with TGF-β1,
TGF-β1+pcDNA3.1, TGF-β1+pcDNA3.1-MALAT1, TGF-β1+si-NC and
TGF-β1+si-MALAT1. *P<0.05 vs. NC;
#P<0.05 vs. TGF-β1+pcDNA3.1;
&P<0.05 vs. TGF-β1+si-NC. (D) Viability, (E)
proliferation, (F) apoptosis and (G) apoptin expression were
determined in the TGF-β1, TGF-β1+pcDNA3.1, TGF-β1+pcDNA3.1-MALAT1,
TGF-β1+si-NC and TGF-β1+si-MALAT1 treated LX2 cells.
*P<0.05 vs. NC; #P<0.05 vs.
TGF-β1+pcDNA3.1; &P<0.05 vs. TGF-β1+si-NC. lncRNA
MALAT1, long non-coding RNA metastasis associated lung
adenocarcinoma transcript 1; miR, microRNA; NF-κB, nuclear
factor-κB; TLR4, toll-like receptor-4; siRNA, small interfering
RNA; NC, negative control; α-SMA, α-smooth muscle actin; COL1,
collagen 1; TIMP-1, tissue inhibitor of metalloprotease-1; TGF-β1,
transforming growth factor-β1. |
Viability (Fig.
5D) and proliferation (Fig.
5E) of LX2 cells were increased, and the apoptotic rate
decreased (Fig. 5F) following
TGF-β1 treatment (P<0.05). Transfection of pcDNA3.1-MALAT1 in
the presence of TGF-β1 increased viability and proliferation, and
decreased apoptosis, even further than TGF-β1 alone (all
P<0.05). In contrast, transfection with si-MALAT1 in the
presence of TGF-β1 decreased viability and proliferation, and
increased apoptosis compared with TGF-β1 alone (P<0.05). Bax and
cleaved caspase-3 expression were decreased, and Bcl-2 expression
was increased by transfection with pcDNA3.1-MALAT1 in the presence
of TGF-β1, when compared with TGF-β1 alone (P<0.05; Fig. 5G). In contrast, Bax and cleaved
caspase-3 levels were upregulated, and Bcl-2 was downregulated
following si-MALAT1 transfection in the presence of TGF-β1,
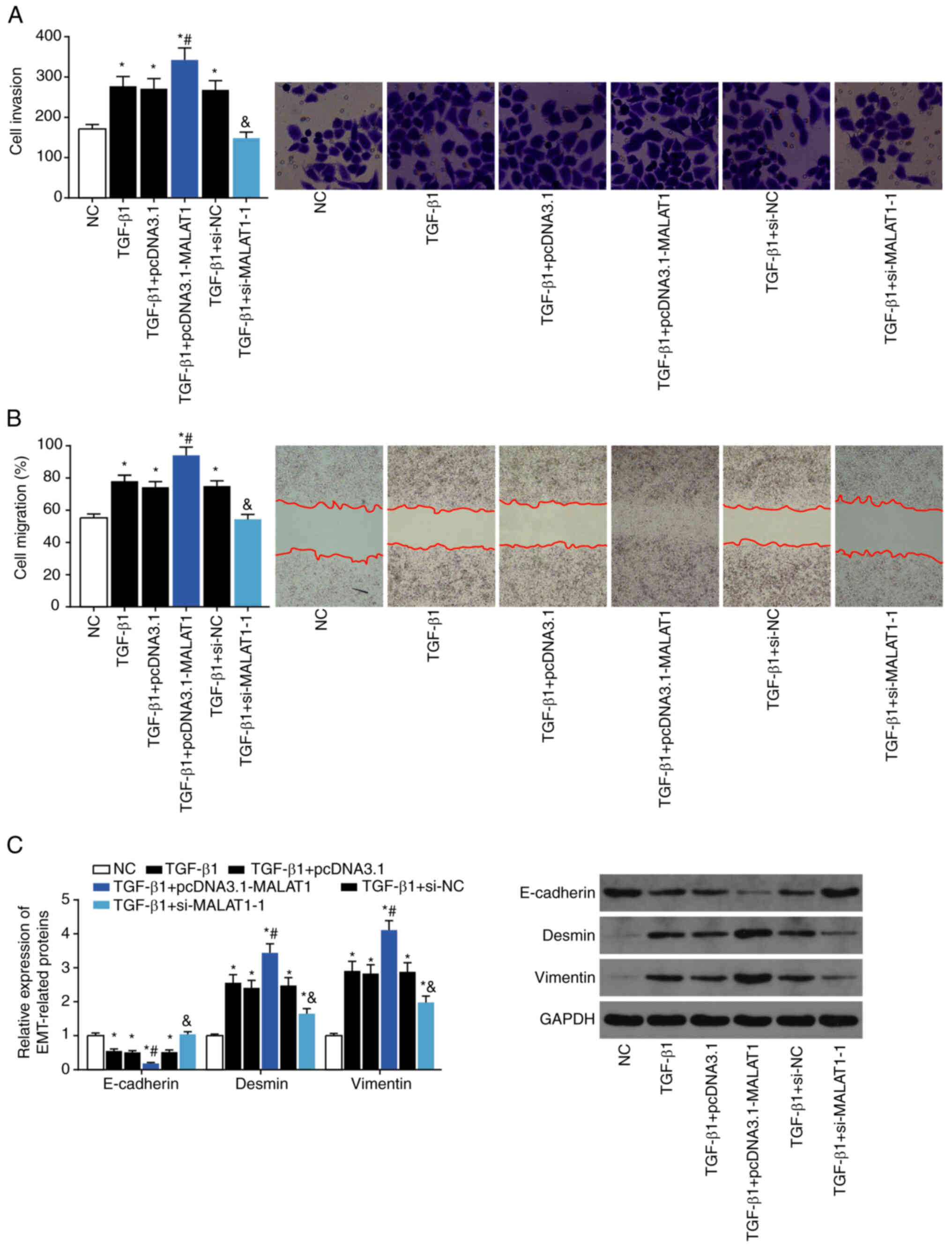
compared with TGF-β1 alone (P<0.05). Migration and adhesion of
TGF-β1-treated LX2 cells were increased by pcDNA3.1-MALAT1
transfection (P<0.05), and reduced by si-MALAT1 transfection
(P<0.05; Fig. 6A and B).
Transfection of LX2 cells with pcDNA3.1-MALAT1 in the presence of
TGF-β1 increased Desmin and Vimentin expression, and decreased
E-cadherin expression, compared with TGF-β1 alone (all P<0.05).
In contrast, transfection with si-MALAT1 in the presence of TGF-β1
upregulated E-cadherin, and downregulated Desmin and Vimentin,
compared with TGF-β1 alone (P<0.05; Fig. 6C).
miR-181a-5p inhibits collagen production
and reduces viability, proliferation, migration and adhesion of
HSCs
Expression of TLR4, NF-κB, α-SMA, COL1 and TIMP-1
were significantly increased following transfection of miR-181a-5p
inhibitor in the presence of TGF-β1 (all P<0.05), whereas they
were markedly reduced by transfection of miR-181a-5p mimic,
compared to TGF-β1 alone (all P<0.05; Fig. 7A and B). In the presence of
TGF-β1, viability and proliferation of LX2 cells was increased by
transfection with miR-181a-5p inhibitor (both P<0.05), and
decreased by transfection with miR-181a-5p mimic (both P<0.05),
when compared with TGF-β1 alone (Fig. 7C and D). Conversely, the
apoptotic rate of LX2 cells was increased by transfection with
miR-181a-5p mimic and decreased by transfection with miR-181a-5p
inhibitor, compared with TGF-β1 alone (all P<0.05; Fig. 7E). In concordance with these
data, Bax and cleaved caspase-3 expression were reduced, and Bcl-2
expression was increased following miR-181a-5p inhibitor
transfection, and vice versa for miR-181a-5p mimic
transfection (all P<0.05; Fig.
7F).
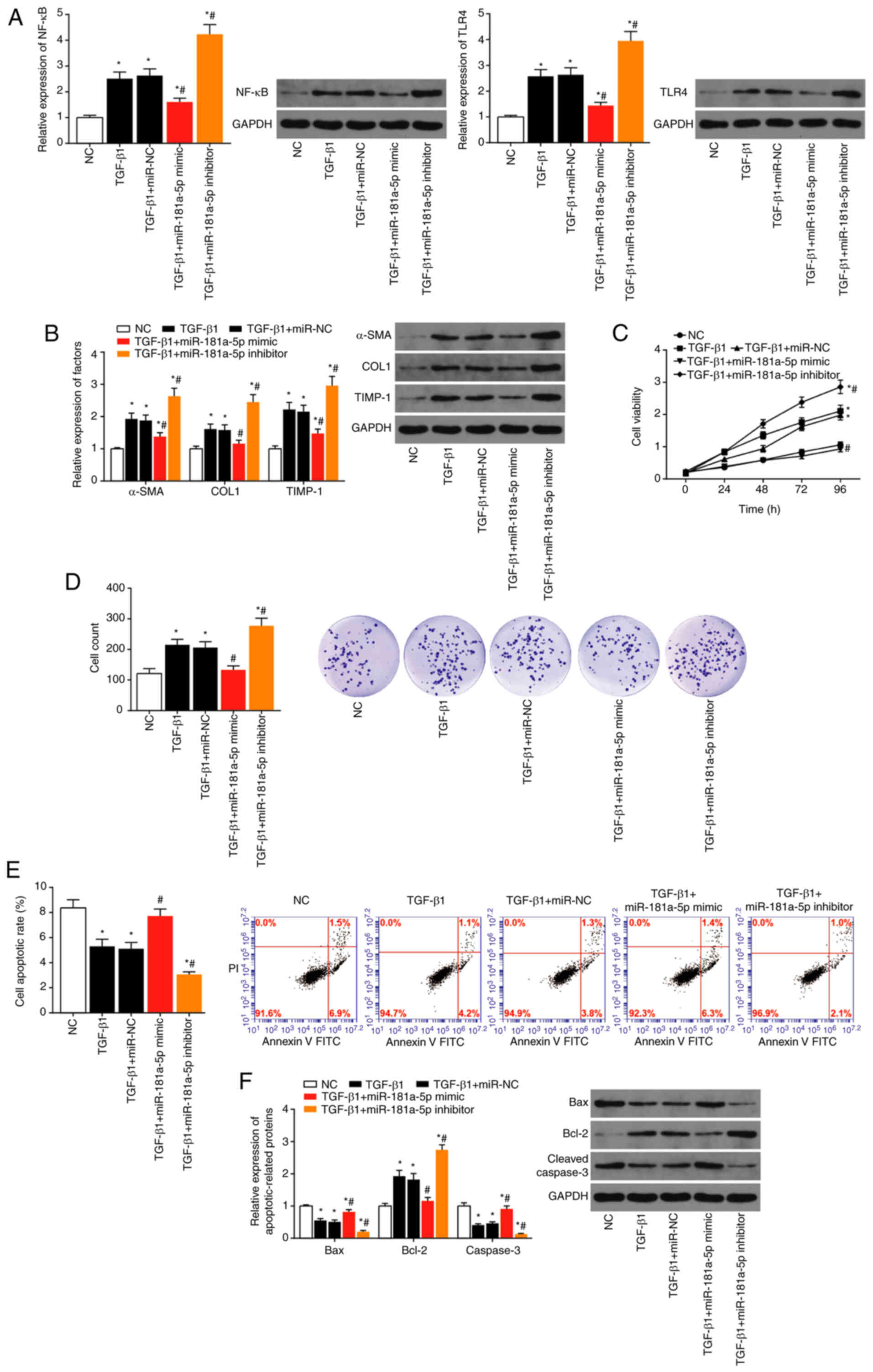 | Figure 7Effect of miR-181a-5p on the
viability and proliferation of LX2 cells. (A) Expression levels of
NF-κB and TLR4 were detected in LX2 cells treated with TGF-β1,
TGF-β1+miR-NC, TGF-β1+miR-181a-5p mimic and TGF-β1+miR-181a-5p
inhibitor. *P<0.05 vs. NC; #P<0.05 vs.
TGF-β1+miR-NC. (B) The expression levels of α-SMA, COL1 and TIMP-1
were evaluated in the LX2 cells treated with TGF-β1, TGF-β1+miR-NC,
TGF-β1+miR-181a-5p mimic and TGF-β1+miR-181a-5p inhibitor.
*P<0.05 vs. NC; #P<0.05 vs.
TGF-β1+miR-NC group. (C) Viability, (D) proliferation, (E)
apoptosis activity as well as (F) apoptin expression in LX2 cells
treated with TGF-β1, TGF-β1+miR-NC, TGF-β1+miR-181a-5p mimic and
TGF-β1+miR-181a-5p inhibitor were determined. *P<0.05
vs. NC; #P<0.05 vs. TGF-β1+miR-NC. miR, microRNA; NC,
negative control; NF-κB, nuclear factor-κB; TLR4, toll-like
receptor-4; α-SMA, α-smooth muscle actin; COL1, collagen 1; TIMP-1,
tissue inhibitor of metalloprotease-1; TGF-β1, transforming growth
factor-β1. |
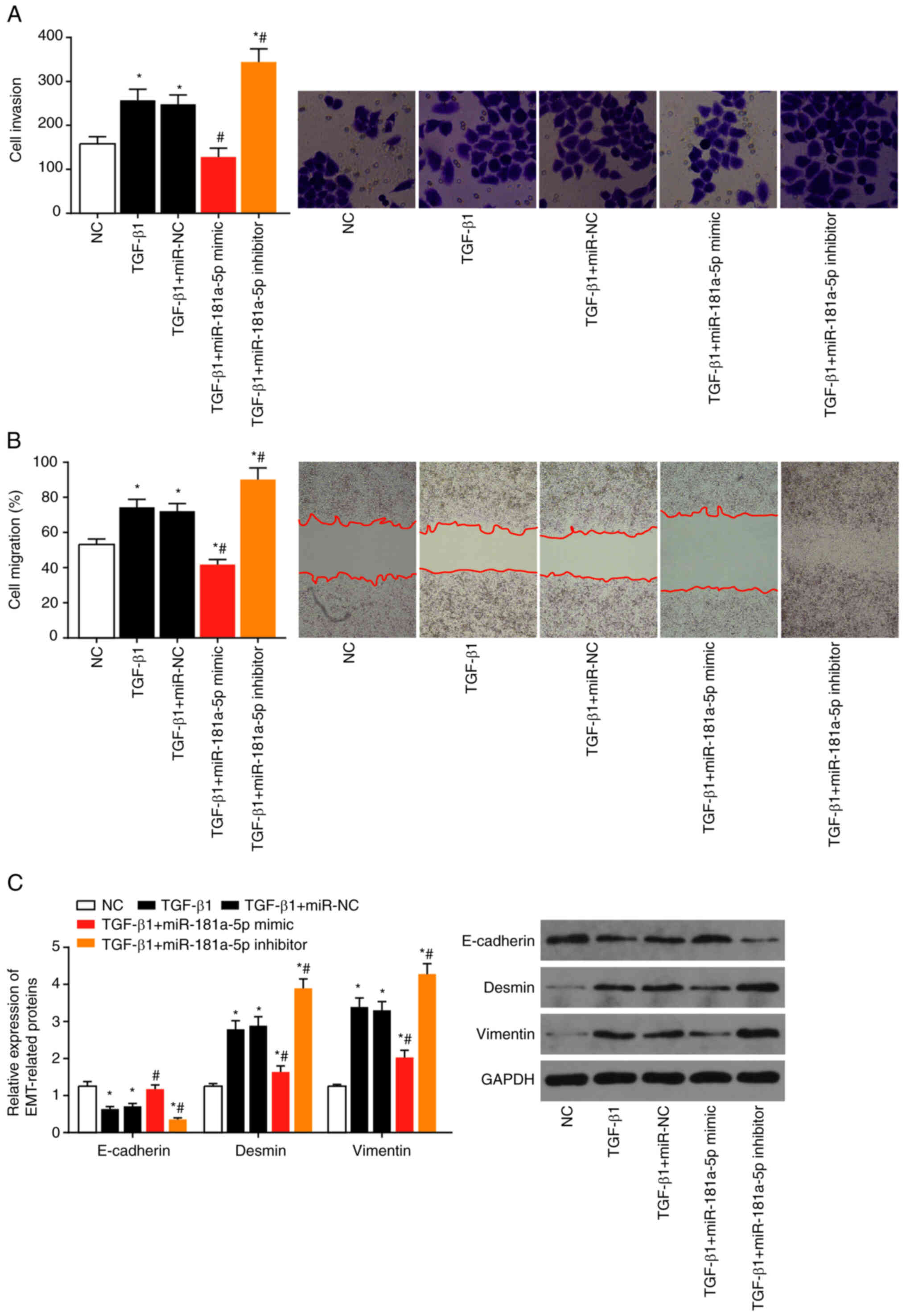
In the presence of TGF-β1, migration and adhesion of
LX2 cells was significantly increased by miR-181a-5p inhibitor
transfection, and decreased by miR-181a-5p mimic transfection (all
P<0.05) (Fig. 8A and B).
Expression of Desmin and Vimentin were decreased and expression of
E-cadherin was increased following miR-181a-5p inhibitor
transfection (all P<0.05), and vice versa for miR-181a-5p
mimic transfection, compared with TGF-β1 alone (P<0.05; Fig. 8C).
Discussion
Several studies have highlighted the importance of
TLR4/NF-κB signaling for the induction of liver fibrosis (20). Activation of NF-κB signaling
inhibits apoptosis of HSCs, and enables HSCs to maintain a
continuously-activated state (21). LPS-activated TLR4/NF-κB signaling
led to the release of inflammatory factors (such as IL-8) and
adhesion molecules (such as monocyte chemoattractant protein-1),
both of which are markers of liver fibrosis (22-24). Following binding to receptors on
the HSC surface, the TLR4/NF-κB axis participates in
phosphorylation of Smad, thereby triggering secretion of collagen,
whose deposition contributes to liver fibrosis progression
(9,10). Given the importance of TLR4/NF-κB
signaling in liver fibrosis, upstream molecules that modulate this
pathway are worthy of intensive exploration.
In the present study, lncRNA MALAT1, initially
identified in non-small cell lung cancer in 2003 (25), was found to promote TLR4 and
NF-κB expression in an HSC cell line, thus not only increasing
TGF-β1-induced proliferation, migration and adhesion, but also
exacerbating LPS-triggered release of inflammatory cytokines
(including IL-6, IL-1β and TNF-α) and collagen synthesis. It has
been reported that MALAT1 regulates the malignant activity and
inflammatory responses of several cell types. For example, knockout
of MALAT1 reduced proliferation and migration of esophageal
squamous cell carcinoma cells (26), and knock-down of MALAT1 led to
G2/M arrest by compromising the cytoplasmic transfer of
Heterogeneous nuclear ribonucleoproteins C (27), which all support the notion that
MALAT1 enhanced migration and proliferation of cells, just as was
observed in HSCs. MALAT1 has also been suggested to promote glucose
induced secretion of inflammatory cytokines, such as TNF-α and
IL-6, by activating SAA3 (28).
However, one study reported that MALAT1 negatively regulated NF-κB
and thereby inhibited the release of inflammatory cytokines from
LPS-activated macrophages (29),
which does not agree with the results of the present study. It is
possible that the influence of MALAT1 on inflammation varies
between cell types, and that MALAT1 may perform distinct functions
in different disorders.
In agreement with the competing endogenous RNA
hypothesis, it was further shown that miR-181a was sponged and
downregulated by MALAT1 in HSCs. It was previously reported that
TGF-β1, which facilitates liver fibrosis by activating HSCs and
driving synthesis of extracellular matrix proteins (such as type
I/III collagen) (30), promoted
shear maturation of pri-miR-181a through activation of Smad2/3 and
the Drosha miRNA processing complex (31). Moreover, TGF-β also induced high
levels of precursor and mature miR-181 in hepatic cells by
targeting and controlling TIMP3 expression (32). Here, it was proposed that TGF-β
may downregulate miR-181 expression in HSCs by increasing MALAT1
expression, and that the MALAT1/miR-181a axis may be involved in
TGF-β-mediated HSC activation. In agreement with the HSC data in
the present study, miR-181a was reported to reduce proliferation
and migration of cutaneous squamous cell carcinoma cells (33), oral squamous cell carcinoma cells
(34), non-small cell lung
cancer cells (35) and malignant
glioma cells (36). However,
miR-181a appeared to facilitate oncogenesis in breast cancer
(37) and liver cancer (38). To explain this seeming
contradiction, it is speculated that there may be additional
mechanisms yet to be discovered, such as bi-directional regulation
in breast cancer (39), and that
miR-181a could exert different effects in different diseases.
miR-181a was reported to regulate the development of embryonic
organs, the lymphatic system and blood cell lines (40), as well as the differentiation of
megakaryocytes and maturation of T lymphocytes (41). Consistent with an earlier study
that reported downregulation of miR-181a in a LPS-induced ALI rat
model (42), it was shown in the
present study that miR-181a repressed inflammation in HSCs by
deactivation of the TLR4/NF-κB axis. Together, the results of the
present study showed that miR-181a was pivotal for the contribution
of MALAT1 to the activation and inflammation of the LX2 cell
line.
In conclusion, inflammation and HSC activation were
of significance to the development of liver fibrosis (43,44). TGF-β1 is typically used to induce
HSC activation and proliferation (45). LPS, a constituent of the cell
wall of gram-negative bacteria, elicits a series of inflammation
responses, and is often applied to establish cell models of
inflammation in the study of liver fibrosis (46). This investigation used TGF-β1 to
trigger HSC activation and utilized LPS to construct models of
inflammation of HSC. In both cell models, a
MALAT1/miR181a/TLR4/NF-κB axis was studied, and an association of
MALAT1/miR181a/TLR4/NF-κB axis with HSC activation and inflammation
was identified. There are several potential shortcomings in the
experimental design used. Firstly, the liver fibrosis cohort were
all HBV-infected, so it is unclear whether the
MALAT1-miR-181-TLR4/NF-kB axis behaves similarly in cases of
non-HBV liver fibrosis. Secondly, only a single cell line was
included; the data would be more convincing if other cell lines or
primary cells were also studied. In future studies, these mentioned
limitations will be addressed and animal models of liver fibrosis
will be used to verify these findings in vitro, which will
provide support for the proposed model of the role of the
MALAT1/miR-181 axis in liver fibrosis.
Supplementary Data
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YW, QM, ZZ, LuZ and LiZ conceived and designed the
experiments. YW, QM and ZZ performed the experiments. ZZ analyzed
the data. LuZ and LiZ drafted the manuscript. YW and QM confirm the
authenticity of all the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Affiliated Hospital of Guizhou Medical University.
Written informed consent was obtained from all participants
included in the present study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Yin C, Evason KJ, Asahina K and Stainier
DY: Hepatic stellate cells in liver development, regeneration, and
cancer. J Clin Invest. 123:1902–1910. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Higashi T, Friedman SL and Hoshida Y:
Hepatic stellate cells as key target in liver fibrosis. Adv Drug
Deliv Rev. 121:27–42. 2017. View Article : Google Scholar
|
|
3
|
Dixon LJ, Barnes M, Tang H, Pritchard MT
and Nagy LE: Kupffer cells in the liver. Compr Physiol. 3:785–797.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Koyama Y and Brenner DA: Liver
inflammation and fibrosis. J Clin Invest. 127:55–64. 2017.
View Article : Google Scholar
|
|
5
|
Dhar D, Baglieri J, Kisseleva T and
Brenner DA: Mechanisms of liver fibrosis and its role in liver
cancer. Exp Biol Med (Maywood). 245:96–108. 2020. View Article : Google Scholar
|
|
6
|
Peng H, Wan LY, Liang JJ, Zhang YQ, Ai WB
and Wu JF: The roles of lncRNA in hepatic fibrosis. Cell Biosci.
8:632018. View Article : Google Scholar :
|
|
7
|
Zhang K, Han X, Zhang Z, Zheng L, Hu Z,
Yao Q, Cui H, Shu G, Si M, Li C, et al: The liver-enriched
lnc-LFAR1 promotes liver fibrosis by activating TGFβ and Notch
pathways. Nat Commun. 8:1442017. View Article : Google Scholar
|
|
8
|
Wang X, Cheng Z, Dai L, Jiang T, Jia L,
Jing X, An L, Wang H and Liu M: Knockdown of long noncoding RNA H19
represses the progress of pulmonary fibrosis through the
transforming growth factor β/smad3 pathway by regulating MicroRNA
140. Mol Cell Biol. 39:e00143–19. 2019. View Article : Google Scholar
|
|
9
|
Dooley S and ten Dijke P: TGF-β in
progression of liver disease. Cell Tissue Res. 347:245–256. 2012.
View Article : Google Scholar
|
|
10
|
Yang JW, Hien TT, Lim SC, Jun DW, Choi HS,
Yoon JH, Cho IJ and Kang KW: Pin1 induction in the fibrotic liver
and its roles in TGF-β1 expression and Smad2/3 phosphorylation. J
Hepatol. 60:1235–1241. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu Y, Liu X, Zhou Q, Huang C, Meng X, Xu F
and Li J: Silent information regulator 1 (SIRT1) ameliorates liver
fibrosis via promoting activated stellate cell apoptosis and
reversion. Toxicol Appl Pharmacol. 289:163–176. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jiang R, Zhou Y, Wang S, Pang N, Huang Y,
Ye M, Wan T, Qiu Y, Pei L, Jiang X, et al: Nicotinamide riboside
protects against liver fibrosis induced by CCl4 via
regulating the acetylation of smads signaling pathway. Life Sci.
225:20–28. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yu F, Lu Z, Cai J, Huang K, Chen B, Li G,
Dong P and Zheng J: MALAT1 functions as a competing endogenous RNA
to mediate Rac1 expression by sequestering miR-101b in liver
fibrosis. Cell Cycle. 14:3885–3896. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dai X, Chen C, Xue J, Xiao T, Mostofa G,
Wang D, Chen X, Xu H, Sun Q, Li J, et al: Exosomal MALAT1 derived
from hepatic cells is involved in the activation of hepatic
stellate cells via miRNA-26b in fibrosis induced by arsenite.
Toxicol Lett. 316:73–84. 2019. View Article : Google Scholar
|
|
15
|
Sun Y, Jiang T, Jia Y, Zou J, Wang X and
Gu W: LncRNA MALAT1/miR-181a-5p affects the proliferation and
adhesion of myeloma cells via regulation of Hippo-YAP signaling
pathway. Cell Cycle. 18:2509–2523. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiang K, Guo S, Zhang T, Yang Y, Zhao G,
Shaukat A, Wu H and Deng G: Downregulation of TLR4 by miR-181a
provides negative feedback regulation to lipopolysaccharide-induced
inflammation. Front Pharmacol. 9:1422018. View Article : Google Scholar :
|
|
17
|
Sahin H, Borkham-Kamphorst E, do O NT,
Berres ML, Kaldenbach M, Schmitz P, Weiskirchen R, Liedtke C,
Streetz KL, Maedler K, et al: Proapoptotic effects of the
chemokine, CXCL 10 are mediated by the noncognate receptor TLR4 in
hepatocytes. Hepatology. 57:797–805. 2013. View Article : Google Scholar
|
|
18
|
Zhu D, He X, Duan Y, Chen J, Wang J, Sun
X, Qian H, Feng J, Sun W, Xu F and Zhang L: Expression of
microRNA-454 in TGF-β1-stimulated hepatic stellate cells and in
mouse livers infected with Schistosoma japonicum. Parasit Vectors.
7:1482014. View Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
20
|
Lai L, Chen Y, Tian X, Li X, Zhang X, Lei
J, Bi Y, Fang B and Song X: Artesunate alleviates hepatic fibrosis
induced by multiple pathogenic factors and inflammation through the
inhibition of LPS/TLR4/NF-κB signaling pathway in rats. Eur J
Pharmacol. 765:234–241. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen Z, Amro EM, Becker F, Hölzer M, Rasa
SMM, Njeru SN, Han B, Di Sanzo S, Chen Y, Tang D, et al:
Cohesin-mediated NF-κB signaling limits hematopoietic stem cell
self-renewal in aging and inflammation. J Exp Med. 216:152–175.
2019. View Article : Google Scholar :
|
|
22
|
Kobayashi K, Yoshioka T, Miyauchi J,
Nakazawa A, Yamazaki S, Ono H, Tatsuno M, Iijima K, Takahashi C,
Okada Y, et al: Monocyte chemoattractant protein-1 (MCP-1) as a
potential therapeutic target and a noninvasive biomarker of liver
fibrosis associated with transient myeloproliferative disorder in
down syndrome. J Pediatr Hematol Oncol. 39:e285–e289. 2017.
View Article : Google Scholar
|
|
23
|
Zimmermann HW, Seidler S, Gassler N,
Nattermann J, Luedde T, Trautwein C and Tacke F: Interleukin-8 is
activated in patients with chronic liver diseases and associated
with hepatic macrophage accumulation in human liver fibrosis. PLoS
One. 6:e213812011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vespasiani-Gentilucci U, Carotti S,
Perrone G, Mazzarelli C, Galati G, Onetti-Muda A, Picardi A and
Morini S: Hepatic toll-like receptor 4 expression is associated
with portal inflammation and fibrosis in patients with NAFLD. Liver
Int. 35:569–581. 2015. View Article : Google Scholar
|
|
25
|
Ji P, Diederichs S, Wang W, Böing S,
Metzger R, Schneider PM, Tidow N, Brandt B, Buerger H, Bulk E, et
al: MALAT-1, a novel noncoding RNA, and thymosin beta4 predict
metastasis and survival in early-stage non-small cell lung cancer.
Oncogene. 22:8031–8041. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang X, Li M, Wang Z, Han S, Tang X, Ge Y,
Zhou L, Zhou C, Yuan Q and Yang M: Silencing of long noncoding RNA
MALAT1 by miR-101 and miR-217 inhibits proliferation, migration,
and invasion of esophageal squamous cell carcinoma cells. J Biol
Chem. 290:3925–3935. 2015. View Article : Google Scholar :
|
|
27
|
Yang F, Yi F, Han X, Du Q and Liang Z:
MALAT-1 interacts with hnRNP C in cell cycle regulation. FEBS Lett.
587:3175–3181. 2013. View Article : Google Scholar
|
|
28
|
Puthanveetil P, Chen S, Feng B, Gautam A
and Chakrabarti S: Long non-coding RNA MALAT1 regulates
hyperglycaemia induced inflammatory process in the endothelial
cells. J Cell Mol Med. 19:1418–1425. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhao G, Su Z, Song D, Mao Y and Mao X: The
long noncoding RNA MALAT1 regulates the lipopolysaccharide-induced
inflammatory response through its interaction with NF-κB. FEBS
Lett. 590:2884–2895. 2016. View Article : Google Scholar
|
|
30
|
Liu N, Feng J, Lu X, Yao Z, Liu Q, Lv Y,
Han Y, Deng J and Zhou Y: Isorhamnetin inhibits liver fibrosis by
reducing autophagy and inhibiting extracellular matrix formation
via the TGF-β1/smad3 and TGF-β1/p38 MAPK pathways. Mediators
Inflamm. 2019:61750912019. View Article : Google Scholar
|
|
31
|
Wang Y, Yu Y, Tsuyada A, Ren X, Wu X,
Stubblefield K, Rankin-Gee EK and Wang SE: Transforming growth
factor-β regulates the sphere-initiating stem cell-like feature in
breast cancer through miRNA-181 and ATM. Oncogene. 30:1470–1480.
2011. View Article : Google Scholar
|
|
32
|
Wang B, Hsu SH, Majumder S, Kutay H, Huang
W, Jacob ST and Ghoshal K: TGFbeta-mediated upregulation of hepatic
miR-181b promotes hepatocarcinogenesis by targeting TIMP3.
Oncogene. 29:1787–1797. 2010. View Article : Google Scholar
|
|
33
|
Neu J, Dziunycz PJ, Dzung A, Lefort K,
Falke M, Denzler R, Freiberger SN, Iotzova-Weiss G, Kuzmanov A,
Levesque MP, et al: miR-181a decelerates proliferation in cutaneous
squamous cell carcinoma by targeting the proto-oncogene KRAS. PLoS
One. 12:e01850282017. View Article : Google Scholar
|
|
34
|
Li GH, Ma ZH and Wang X: Long non-coding
RNA CCAT1 is a prognostic biomarker for the progression of oral
squamous cell carcinoma via miR-181a-mediated Wnt/β-catenin
signaling pathway. Cell cycle. 18:2902–2913. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shi Q, Zhou Z, Ye N, Chen Q, Zheng X and
Fang M: MiR-181a inhibits non-small cell lung cancer cell
proliferation by targeting CDK1. Cancer Biomark. 20:539–546. 2017.
View Article : Google Scholar
|
|
36
|
Ma J, Yao Y, Wang P, Liu Y, Zhao L, Li Z,
Li Z and Xue Y: MiR-181a regulates blood-tumor barrier permeability
by targeting Krüppel-like factor 6. J Cereb Blood Flow Metab.
34:1826–1836. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Taylor MA, Sossey-Alaoui K, Thompson CL,
Danielpour D and Schiemann WP: TGF-β upregulates miR-181a
expression to promote breast cancer metastasis. J Clin Invest.
123:150–163. 2013. View Article : Google Scholar
|
|
38
|
Zhuang X, Chen Y, Wu Z, Xu Q, Chen M, Shao
M, Cao X, Zhou Y, Xie M, Shi Y, et al: Mitochondrial miR-181a-5p
promotes glucose metabolism reprogramming in liver cancer by
regulating the electron transport chain. Carcinogenesis.
41:972–983. 2020. View Article : Google Scholar
|
|
39
|
Yang C, Tabatabaei SN, Ruan X and Hardy P:
The dual regulatory role of MiR-181a in breast cancer. Cell Physiol
Biochem. 44:843–856. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Seoudi AM, Lashine YA and Abdelaziz AI:
MicroRNA-181a-a tale of discrepancies. Expert Rev Mol Med.
14:e52012. View Article : Google Scholar
|
|
41
|
Kim C, Jadhav RR, Gustafson CE, Smithey
MJ, Hirsch AJ, Uhrlaub JL, Hildebrand WH, Nikolich-Žugich J, Weyand
CM and Goronzy JJ: Defects in antiviral T cell responses inflicted
by aging-associated miR-181a deficiency. Cell Rep. 29:2202–2216.e5.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cai ZG, Zhang SM, Zhang Y, Zhou YY, Wu HB
and Xu XP: MicroRNAs are dynamically regulated and play an
important role in LPS-induced lung injury. Can J Physiol Pharmacol.
90:37–43. 2012. View Article : Google Scholar
|
|
43
|
Anthony B, Allen JT, Li YS and McManus DP:
Hepatic stellate cells and parasite-induced liver fibrosis. Parasit
Vectors. 3:602010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Parola M and Pinzani M: Liver fibrosis:
Pathophysiology, pathogenetic targets and clinical issues. Mol
Aspects Med. 65:37–55. 2019. View Article : Google Scholar
|
|
45
|
Mu M, Zuo S, Wu RM, Deng KS, Lu S, Zhu JJ,
Zou GL, Yang J, Cheng ML and Zhao XK: Ferulic acid attenuates liver
fibrosis and hepatic stellate cell activation via inhibition of
TGF-β/Smad signaling pathway. Drug Des Devel Ther. 12:4107–4115.
2018. View Article : Google Scholar
|
|
46
|
Hu N, Wang C, Dai X, Zhou M, Gong L, Yu L,
Peng C and Li Y: Phillygenin inhibits LPS-induced activation and
inflammation of LX2 cells by TLR4/MyD88/NF-κB signaling pathway. J
Ethnopharmacol. 248:1123612020. View Article : Google Scholar
|