Introduction
The incidence rate of metabolic-associated fatty
liver disease (MAFLD) has rapidly increased, and it has become a
major health concern, posting a threat to numerous lives (1-3).
The ectopic accumulation of liver fat aggravates the metabolic
burden, and when accompanied by multiple factors, such as lipotoxic
substances, the inflammatory response and oxidative stress, MAFLD
can progress from steatosis to cirrhosis, and in some cases, even
hepatocellular carcinoma. Exercise is often used as the main
treatment modality for MAFLD, which can effectively improve
intrahepatic lipid accumulation (4-6).
Previous research and a randomized controlled trials (RCT) on the
effects of aerobic exercise training doses on liver fat
accumulation have revealed that even a minimal amount of physical
activity (150-180 min of moderate-intensity exercise per week) can
reduce liver fat in the short-term (7,8).
However, the majority of patients who were included in the RCT went
through the diet and exercise program without success (8). As with all clinical trials, some
patients cannot tolerate or adhere to the prescribed exercise
program due to their physical condition or lifestyle habits. Thus,
exploring an effective alternative therapy to simulate exercise may
provide a feasible solution.
MicroRNAs (miRNAs/miRs) are evolutionarily
conserved, small (~21 nucleotides) non-coding RNAs (9). miRNA precursors (pre-miRNAs) are
stem-loop structures which are exported to the cytoplasm by the
karyopherin, exportin 5 (Exp5) transporter. In the cytoplasm,
pre-miRNAs are processed by the Dicer enzyme to produce mature
miRNAs of ~22 nucleotides in length (9). In a previous study by the authors,
miR-212 was identified to be closely related to lipid metabolism
(10). It has also been shown to
be involved in a variety of pathophysiological processes, including
cell proliferation (11),
angiogenesis (12), intestinal
permeability (13) and blood
glucose metabolism (14).
Furthermore, its dysregulation is strongly related to metabolic
diseases, such as hypertension, cardiac hypertrophy (15) and alcoholic liver disease
(13). Mature miR-212 consists
mainly of miR-212-3p and miR-212-5p, both of which are closely
related to lipid metabolism. Leucine deprivation suppresses
triglyceride (TG) accumulation in the liver, which usually causes
elevated levels of miR-212-5p (16). A high-fat diet (HFD) accelerates
liver lipid accumulation, which usually leads to elevated levels of
miR-212-3p (10). In addition,
exercise improves hepatic lipid accumulation, which decreases
miR-212-3p levels in patients with MAFLD. The study of miR-212 may
contribute to the development of potential exercise replacement
therapies for MAFLD in the future. The therapeutic potential of
miR-212 (an alternative treatment which can mimic the effects of
exercise), as well as the detailed functional mechanisms of MAFLD
warrant further investigation.
The present study explored three functional roles of
miR-212-3p in MAFLD; i.e., the therapeutic effects of targeting
miR-212-3p, its association with exercise and the mechanisms
underlying its functions. The data confirmed that the inhibition of
miR-212-3p exerted beneficial effects on MAFLD similar to those of
exercise by targeting fibroblast growth factor 21 (FGF21), rather
than chromodomain helicase DNA binding protein 1 (CHD1). In
addition, the regulatory effects of exercise on miR-212-3p were
investigated. Exercise played a protective role in MAFLD by
activating early growth response 1 (EGR1), which transcriptionally
suppressed miR-212-3p and increased FGF21 expression. The
overexpression of miR-212-3p abolished the regulatory effects of
EGR1 on FGF21, validating a regulatory association among EGR1,
miR-212-3p and FGF21. miR-212-3p was a key molecule in the effects
of exercise on MAFLD. These findings suggest a potential
therapeutic effect of targeting miR-212-3p on MAFLD.
Materials and methods
Animals experiments and grouping
A total of 30 male C57BL/6 mice (aged 8 weeks;
weighing 20-23 g; mean ± standard error, 21.7±0.9 g) were purchased
from the Shanghai SLAC Laboratory Animal Co., Ltd. The mice were
maintained on a 12-h light/dark cycle at 25°C, a humidity of 50-60%
and lighting with ad libitum access to food and water. The
animal study was approved by the Animal Ethics Committee of Tongji
University, Shanghai, China (approval no. TJHBLAC-2 019-024). All
procedures performed in experiments involving animals were in
accordance with the ethical standards of the institution or
practice at which the studies were conducted. The mice were
randomly divided into five groups as follows: i) The chow group
(n=6), in which mice were fed standard chow for 16 weeks, and
administered tail vein injections of adeno-associated virus (AAV)
serotype 8 gene vectors [1×1011 viral genomes in 100
µl saline; Hanheng Biotechnology (Shanghai) Co., Ltd.] in
the 4th week; ii) the HFD group (n=6), in which mice were fed a HFD
for 16 weeks, and received a tail vein injection of AAV serotype 8
gene vectors [1×1011 viral genomes in 100 µl
saline; Hanheng Biotechnology (Shanghai) Co., Ltd.] in the 4th
week; iii) the HFD and SP group (n=6), in which mice were fed a HFD
for 16 weeks, and received a tail vein injection of recombinant AAV
serotype 8 gene vectors carrying miR-212-3P sponge
[1×1011 viral genomes in 100 µl saline; Hanheng
Biotechnology (Shanghai) Co., Ltd.] in the 4th week; iv) the HE
group (n=6), in which mice were fed a HFD, subjected to daily
exercise on a running machine (10 m/min, 60 min/day, 16 weeks) and
received a tail vein injection of recombinant AAV serotype 8 gene
vectors [1×1011 viral genomes in 100 µl saline;
Hanheng Biotechnology (Shanghai) Co., Ltd.] in the 4th week; and v)
the HE and OE group (n=6), in which mice were fed a HFD, subjected
to daily exercise on a running machine (10 m/min, 60 min/day, 16
weeks) and received a tail vein injection of recombinant AAV
serotype 8 gene vectors carrying miR-212-3P [1×1011
viral genomes in 100 µl saline; Hanheng Biotechnology
(Shanghai) Co., Ltd.] in the 4th week. The standard chow diet (cat.
no. LAD3001M; 14.1% kcal protein, 75.9% carbohydrate and 10.0% fat;
3.6 kcal/g) and HFD (cat. no. TP23400; 14.1% kcal protein, 25.9%
carbohydrate and 60.0% fat; 5.0 kcal/g) were obtained from Trophic
Animal Feed High-tech Co. Ltd.
For the glucose tolerance test (GTT), the mice were
fasted overnight for 16 h with free access to water and injected
intraperitoneally (i.p.) with 2 g/kg glucose. For the insulin
tolerance test (ITT), the mice were fasted 4 h prior to being i.p.
injected with human insulin (0.75 U/kg; Eli Lilly and Company). The
fasting blood glucose levels were detected after the mice were
fasted for 6 h.
The criteria for humane endpoints in this experiment
were as follows: Markedly reduced food or water intake, waddling,
dyspnea, ruffled fur or self-mutilation, inability to stand and
unresponsiveness to external stimuli. Any animals reaching these
endpoints were to be euthanized with 1% pentobarbital sodium (150
mg/kg, i.p., P3761, MilliporeSigma). All animals were observed
daily for behavior and food or water intake to assess their health
status. No abnormal signs indicating the humane endpoint of the
experiment were observed in any of the mice during the
experiment.
At the end of the experiment (16 weeks), a total of
30 male C57BL/6 mice were euthanized by an overdose of inhalant
anesthetic (isoflurane; concentration: 5%; exposure time, 5 min),
followed by ensuring the mouse was unconscious by testing the
retraction of the foot pedal and the reflexes of the eyelids, and
finally exsanguination under anesthesia to ensure death. When the
mice exhibited no respiration, no heartbeat and no response to any
external stimuli, they were considered dead. Samples required for
testing were collected.
Microarray data and human samples
Microarray data (GSE65978) were obtained from the
Gene Expression Omnibus (GEO) database, as previously described
(10). Non-coding RNA expression
information was measured for liver tissue samples from 4 patients
with MAFLD and 4 patients with non-fatty liver at Tongji Hospital,
Tongji University. The National Institute of Diabetes and Digestive
and Kidney Diseases (NIDDK) Nonalcoholic Steatohepatitis Clinical
Research Network (NASH CRN) criteria (17) were used to assess liver histology.
Combined with the clinical history, the Alcohol Use Disorders
Identification Test (AUDIT) questionnaire (18) was used to exclude the interference
of a history of alcohol consumption. The present study was approved
by the Ethics Committee of Tongji Hospital (approval no.
K-KYSB-2020-139) and was performed in accordance with the
Declaration of Helsinki. Written informed consent was obtained from
the patients or their guardians.
Measurement of serum and liver
metabolites
Serum was separated at 5,000 × g for 15 min at room
temperature and stored at −80°C. Alanine transaminase (ALT),
aspartate transaminase (AST) and TG levels were measured using
routine clinical chemical assays (cat. nos. C009-2-1, C010-2-1 and
A110-1-1, respectively; Nanjing Jiancheng Bioengineering
Institute). The process was carried out according to the
manufacturer's protocols. Serum FGF21 and insulin levels were
measured using a mouse FGF21 ELISA kit and a mouse insulin ELISA
kit (JM-03104M1 and JM-02862M1, respectively; Jiangsu Jingmei
Biotechnology, Co., Ltd.), according to the manufacturer's
protocols.
Histopathological analysis and
immunohistochemistry
The liver tissue samples were fixed in 4%
paraformaldehyde for 24 h at temperature. For the preparation of
paraffin-embedded tissues, the tissues were routinely dehydrated at
room temperature using an increasing ethanol gradient followed by
xylene. The tissues were then embedded in paraffin. The
non-alcoholic fatty liver disease (NAFLD) activity score (NAS) was
calculated by determining steatosis, inflammation and ballooning.
For the preparation of frozen tissues, tissues were routinely
dehydrated at room temperature using an increasing sucrose followed
by optimal cutting temperature compound embedding, and preservation
at −80°C. Liver sections (5-µm-thick) were respectively
stained with hematoxylin and eosin (H&E) dye (cat. no. G1120;
Beijing Solarbio Science & Technology Co., Ltd.) and Oil Red O
dye (cat. no. D027-1-1; Nanjing Jiancheng Bioengineering Institute)
to observe hepatic lipid droplets. For H&E staining, the tissue
sections were stained with hematoxylin solution for 10 min at room
temperature and washed in running water for 5 min, followed by the
addition of differentiation solution for 10 sec at room
temperature. The tissue sections were subsequently stained with
eosin solution for 10 sec at room temperature, dehydrated in
alcohol (75, 85, 95 and 100%; 2-3 sec each at room temperature),
and rinsed in 100% alcohol for 1 min at room temperature. Finally,
the tissue sections were cleared with xylene and sealed with
neutral balsam. For Oil Red O staining, frozen sections were dried
at room temperature for 15-20 min, followed by incubation in 100%
isopropanol for 5 min. Subsequently, 0.5% Oil Red O solution was
incubated for 8 min at 60°C, distilled water was cleaned for 5 min.
Hematoxylin was stained for 10 min and washed with distilled water
for 5 min. Finally, the sections were sealed with glycerol gelatin.
The 'analyze particles' function (size-pixel2 from 0.1
to infinity; circularity from 0.1 to 1) of Image J software
(Version: 1.52a; National Institutes of Health) was used for lipid
droplet quantification. The sections were scanned using a
Pannoramic Whole Slide Scanner (Pannoramic Desk; 3DHISTECH Ltd.)
and viewed using CaseViewer 2.2 (3DHISTECH Ltd.).
For immunohistochemical staining, the sections were
immersed in 0.01 M sodium citrate (100°C, 20 min) for antigen
retrieval. Endogenous peroxidase activity was quenched by
incubation with 3% H2O2 for 10 min at room
temperature. The sections were then incubated with 10% bovine serum
albumin for 1 h at room temperature, and then incubated with
anti-FGF21 antibody (1:200, cat. no. ab171941, Abcam) at 4°C
overnight. A goat anti-rabbit IgG H&L (HRP) secondary antibody
(1:1,000, cat. no. ab97051, Abcam) specific to the primary antibody
was selected and allowed to react at room temperature for 60 min.
DAB (MilliporeSigma) color reaction solution was used for slice
color development. The sections were scanned using a Pannoramic
Whole Slide Scanner (Pannoramic Desk; 3DHISTECH Ltd.) and viewed
using CaseViewer 2.2 (3DHISTECH Ltd.).
Cell transfection and Nile Red
staining
The liver cancer cell line (HepG2) was purchased
from The Cell Bank of Type Culture Collection of The Chinese
Academy of Sciences (serial no. SCSP-510). The liver cancer cell
line (HepG2) used in the study was authenticated by STR profiling.
The liver cancer cell line (HepG2) was treated with 1 mM long-chain
free fatty acid (FFA; oleate, palmitate at a molar ration of 2:1;
MilliporeSigma) in 1% bovine serum albumin for 24 h to induce
lipogenesis in vitro. The liver cancer cells (HepG2) were
plated in six-well plates in antibiotic-free medium for 24 h prior
to transfection and transfected at 70% confluency for 48 h using
Lipofectamine 2000® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. The
miR-212-3p mimics and inhibitor purchased from Guangzhou Ruibo Co.,
Ltd. The sequence of siRNA duplex targeting EGR1 was referenced
from the literature (19) and
synthesized by Sangon Biotech (Shanghai) Co., Ltd. EGR1
overexpression plasmid with the pcDNA3.1 vector were purchased from
Changsha Youzhe Co., Ltd. For the transfection experiments, the
HepG2 cells were transfected with miR-212-3p mimics/control (50
nM), inhibitors/control (100 nM), FGF21-siRNA/control (75 nM),
CHD1-siRNA/control (75 nM), EGR1-siRNA/control (75 nM) and EGR1
overexpression plasmids/blank vector (1.25 µg/ml) according
to the experimental purpose. The efficiency of transfection was
evaluated using reverse transcription-quantitative (RT-qPCR).
For Nile Red staining, the cells were washed three
times with phosphate-buffered saline (PBS) and then fixed with 3.7%
paraformaldehyde for 1 h at room temperature and washed thrice
again with PBS. To evaluate intracellular lipogenesis, lipid
droplets were stained with Nile Red dye (0.1 µmol/ml; CAS
no. 7385-67-3; Sigma-Aldrich; Merck KGaA) for 15 min in the dark at
room temperature, and cell nuclei were stained with DAPI (CAS no.
KGA215-10; Jiangsu Keygen Biotech Corp., Ltd) for 5 min in the dark
at room temperature. Finally, the cells were washed three times
with PBS to clean the dye solution. The results were analyzed using
a Leica fluorescence microscope (Leica DM6B; Leica Microsystems,
Inc.). The whole staining process was conducted at room temperature
avoiding direct light. Each experiment was performed in triplicate.
The siRNA and negative control sequences used in the experiments
are presented in Table SI.
Luciferase reporter assay
The 293T cell line was purchased from The Cell Bank
of Type Culture Collection of The Chinese Academy of Sciences
(Serial: SCSP-502). To test for promoter activity, the 293T cells
were seeded at 5×104 per well in 24-well plates and
co-transfected with pGL3-miR-promoter/pGL3-basic (containing the
−1,500 to +500 bp promoter sequence of miR-212) (1 µg/ml),
pRL-TK (1 µg/ml) (control for fluorescence) and
EGR1-siRNA/control (75 nM) or pcDNA3.1-EGR1/pcDNA3.1-basic (1
µg/ml) using Lipofectamine 2000® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to
manufacturer's instructions. At 48 h post-transfection, the
luciferase activities were determined using a Dual Luciferase
Reporter Assay kit (DL101, Nanjing Vazyme Biotech Co., Ltd), and
normalized to Renilla luciferase activity, respectively.
Each experiment was performed in triplicate.
Western blot analysis
The liver tissues and cells were lysed using RIPA
lysis buffer (Beyotime Institute of Biotechnology) containing 1%
phenylmethanesulfonyl fluoride. A total of 30 µg protein was
loaded onto 10 and 12.5% SDS-polyacrylamide gels, and transferred
onto PVDF membranes. The membranes were incubated overnight at 4°C
with primary antibodies (as indicated below) followed by an
incubation with the horseradish peroxidase-conjugated goat
anti-rabbit IgG secondary antibody (1:10,000; cat. no. A0208) and
goat anti-mouse IgG (1:10,000; cat. no. A0216) (both from Beyotime
Institute of Biotechnology) at room temperature for 1 h. The
protein bands were visualized using ECL Moon from Beyotime
Institute of Biotechnology. The blots were scanned using an Odyssey
Two-color Infrared Laser Imaging System (LI-COR Biosciences). The
primary antibodies used were as follows: Anti-EGR1 (1:1,000; cat.
no. 55117-1-AP, ProteinTech Group, Inc.), anti-CHD1 (1:1,000; cat.
no. 4351S, Cell Signaling Technology, Inc.), anti-FGF21 (1:1,000;
cat. no. ab171941, Abcam), anti-ERK1/2 (1:1,000; cat. no. ab184699,
Abcam), anti-phosphorylated (p-)ERK1/2 (1:1,000; cat. no. 4377s,
Cell Signaling Technology, Inc.), anti-fatty acid synthase (FASN;
1:5,000; cat. no. ab128870, Abcam), anti-carnitine
palmitoyltransferase (CPT)1α (1:1,000; cat. no. 15184-1-AP,
ProteinTech Group, Inc.) and anti-β-actin (1:5,000; cat. no.
MA1-140, Thermo Fisher Scientific, Inc.). The protein bands were
visualized using BeyoECL Plus (Beyotime Institute of
Biotechnology). QuantityOne v4.6.6 software (Bio Rad Laboratories,
Inc.) was used for the analysis and for the ratio of the gray value
of the target protein band to the gray.
RNA isolation and RT-qPCR
Total RNA was isolated from the cells and tissues
using TRIzol reagent (Sigma-Aldrich; Merck KGaA). For mRNA
detection, RNA was reverse transcribed into cDNA using PrimeScript™
RT Master Mix (cat. no. RR036B; Takara Bio, Inc.). The temperature
protocol was as follows: 37°C for 15 min, 85°C for 5 sec. qPCR was
carried out using TB Green® Premix Ex Taq™ (cat. no.
RR042B; Takara Bio, Inc.) with the ABI Q5 system. The full
thermocycling conditions for qPCR are as follows: Initial
denaturation at 95°C for 30 sec, followed by 40 cycles of 95°C for
5 sec and 60°C for 30 sec. For miRNA analysis, miRNA was reverse
transcribed using the miRNA 1st Strand cDNA Synthesis kit (cat. no.
MR101-01; Nanjing Vazyme Biotech Co., Ltd), and amplified with
miRNA universal SYBR qPCR Master Mix (cat. no. MQ101-02; Nanjing
Vazyme Biotech Co., Ltd). The temperature protocol for cDNA
synthesis were as follows: 25°C for 5 min, 50°C for 15 min, 85°C
for 5 min. The full thermocycling conditions for qPCR are as
follows: Initial denaturation at 95°C for 5 min, followed by 40
cycles of 95°C for 10 sec and 60°C for 30 sec. β-actin was used as
an internal control for each coding genes of interests. U6 was used
as an internal control for each non-coding genes of interests. The
primers used for RT-qPCR are presented in Table SI.
Bioinformatics analysis
For the determination of target genes, the
TargetScan (20-25) (http://www.targetscan.org/vert_80/) and miRwalk
(26) (http://mirwalk.umm.uni-heidelberg.de/) online website
software tools were used that query possible downstream gene sets
by inputing miR-212-3p in the search bar of the website.
Subsequently, enrichment analysis was carried out using Gene
Ontology (GO) to identify biological processes and pathways.
Specifically, terms with a P-value <0.01, a minimum count of 3,
and an enrichment factor >1.5 (the enrichment factor is the
ratio between the observed counts and the counts expected by
chance) were collected and grouped into clusters based on their
membership similarities. Finally, experimentally confirmed
literatures were examined as the final screening (10,27). For transcription factor
prediction, the UCSC online website (http://genome.ucsc.edu/) (28) was queried for promoter region
sequences of miR-212-3p. Subsequently, the sequence was input into
Jasper (https://jaspar.genereg.net/)
(29) and regRNA 2.0 (http://regrna2.mbc.nctu.edu.tw.) (30) online software to obtain predicted
transcription factors. MGI mammalian phenotype (http://www.informatics.jax.org/) was used for
functional screening (31).
Statistical analysis
GraphPad Prism software (Version: 7.00; GraphPad
Software, Inc.) and SPSS 20.0 (IBM Corp.) were used for statistical
analysis. The Metascape online website (http://metascape.org/gp/index.html#/main/step1)
was used for functional enrichment analysis (32). The homeostasis model assessment of
insulin resistance (HOMA-IR) was calculated as (fasting glucose
level x fasting insulin level)/22.4. An unpaired Student's t-test
was used for comparisons between two groups. One-way analysis of
variance (ANOVA) was used for comparisons of three or more groups,
and following ANOVA, Dunnett's test or Tukey's test were used as
post hoc tests according to the analytical purpose. A Mann-Whitney
U test was used to evaluate non-parametric data. All data are
expressed as the mean ± standard error of the mean (SEM), or median
(25-75th percentile). P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-212-3p, but not miR-212-5p is
involved in lipid metabolism in MAFLD
The authors have previously demonstrated that a high
expression of miR-212 promotes lipogenesis in vitro
(10). In the present study, it
was found that among miR-212, both miR-212-3p and miR-212-5p were
stably expressed in vivo, as determined using miRBase
database. The association between miR-212-3p and miR-212-5p in
lipid metabolism warrants clarification. Firstly, following the
chow diet, the HFD and HE models, the microarray data and RT-qPCR
revealed that miR-212-3p expression was elevated in the HFD model
and decreased with exercise intervention. However, miR-212-5p
expression was not consistent with miR-212-3p; this did not
increase with the HFD (Fig. 1A
and Table SII). Secondly,
miR-212-3p and miR-212-5p were detected in human samples from
patients with MAFLD. The expression of miR-212-3p, but not that of
miR-212-5p, was significantly elevated in liver tissues from
patients with MAFLD compared with normal livers (Fig. 1B). Finally, the expression of
miR-212-3p, but not that of miR-212-5p, was significantly
upregulated in the FFA-stimulated cell model (Fig. 1C).
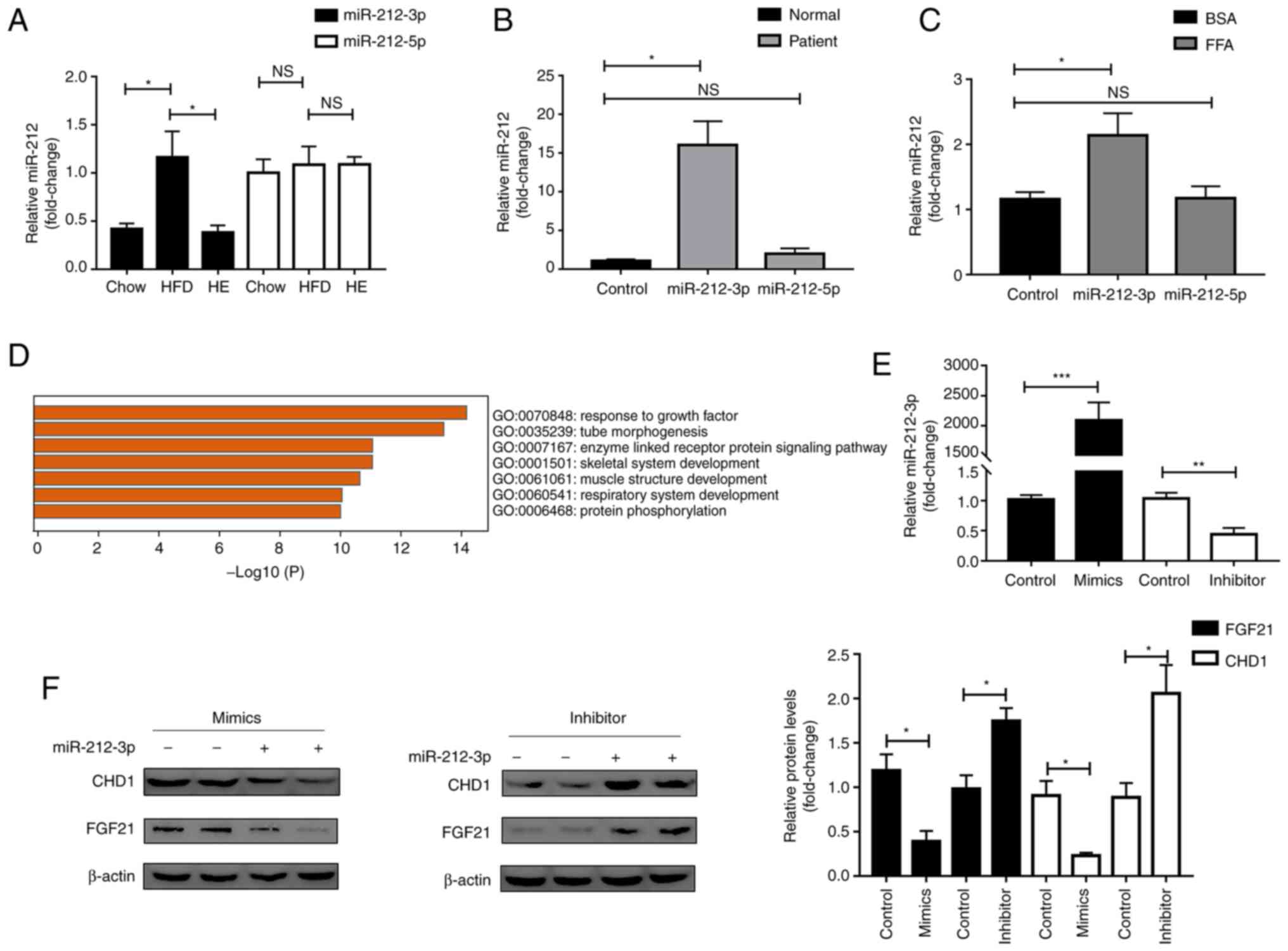 | Figure 1miR-212-3p, but not miR-212-5p is
involved in MAFLD. (A) miR-212-3p, but not miR-212-5p expression,
was elevated in HFD and decreased by exercise (n=6). (B)
miR-212-3p, but not miR-212-5p expression, was elevated in patients
with MAFLD (n=4). (C) miR-212-3p, but not miR-212-5p expression,
was elevated in the FFA-treated cell model (n=3). (D) Functional
enrichment analysis of the target gene. (E) miR-212-3p was
overexpressed/knocked down by miR-212-3p mimics/inhibitor,
respectively. (F) miR-212-3p negatively regulated FGF21/CHD1 at the
protein level under FFA treatment. The data are presented as the
mean ± SEM. *P<0.05, **P<0.01 and
***P<0.001. NS, not significant; MAFLD,
metabolic-associated fatty liver disease; HFD, high-fat diet; FFA,
free fatty acid; FGF21, fibroblast growth factor 21; CHD1,
chromodomain helicase DNA binding protein 1; HE, high-fat diet and
exercise. |
Subsequently, the target genes (CHD1 and FGF21) of
miR-212-3p involved in lipid metabolism were selected using
TargetScan (20-25), miRwalk (26) and experimental validation
(10,27), combined with functional enrichment
analysis (Fig. 1D), in which
FGF21 was one of the molecules previously identified and reported
by the authors (10). On this
basis, miR-212-3p mimics/inhibitor were separately transfected for
further validation (Fig. 1E). The
overexpression of miR-212-3p suppressed the protein levels, but not
the mRNA levels of CHD1 and FGF21. Conversely, the inhibition of
miR-212-3p significantly upregulated the protein levels, but not
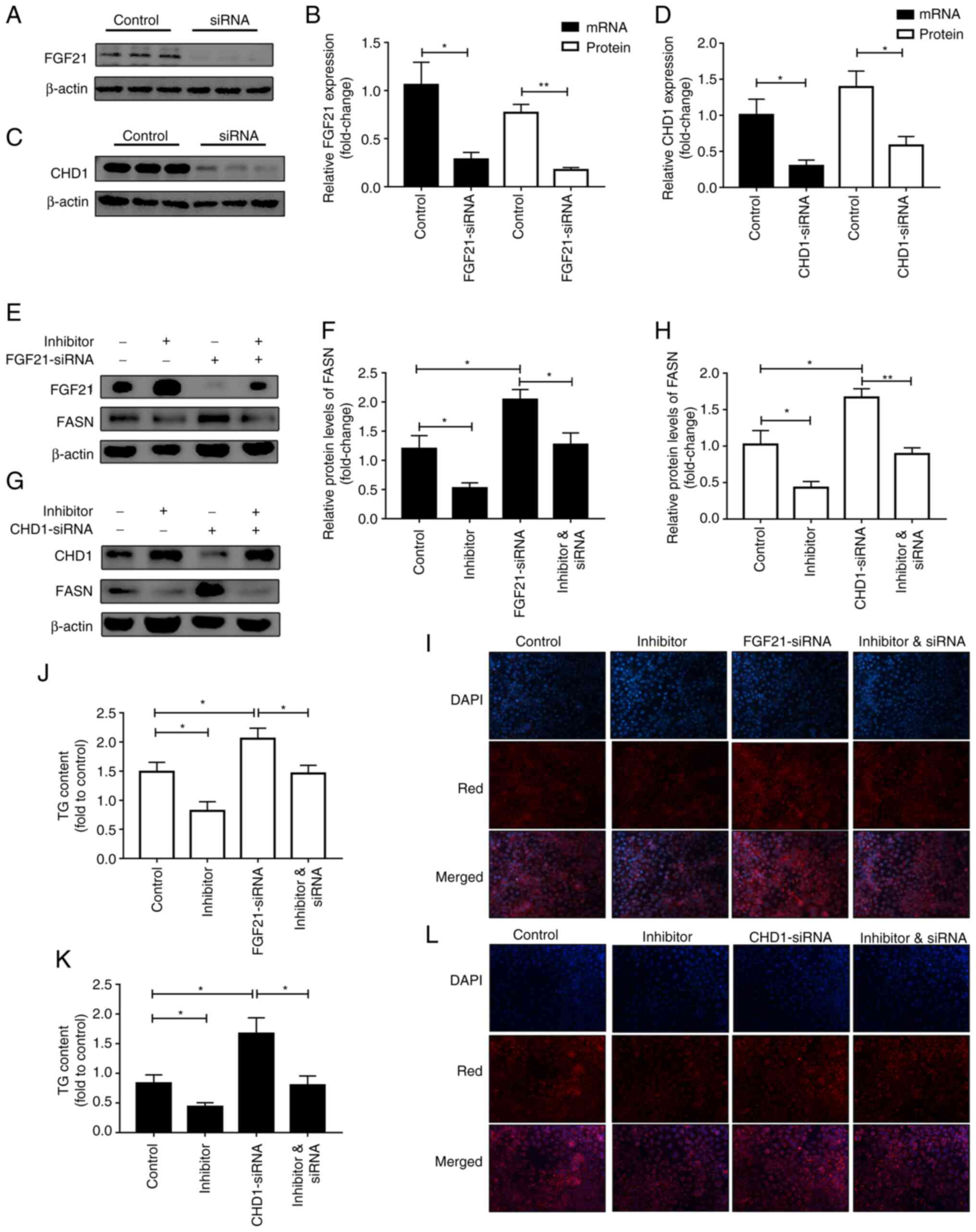
the mRNA levels of CHD1 and FGF21 (Figs. 1F and S1). Furthermore, a rescue experiment
was performed to further verify whether the lipogenic effect of
miR-212-3p was realized by regulating FGF21/CHD1. FGF21/CHD1 was
knocked down using siRNA (Fig.
2A-D). Subsequently, following co-transfection with miR-212-3p
inhibitor and FGF21-siRNA, the decrease in FASN expression induced
by transfection with miR-212-3p inhibitor alone was abolished by
FGF21-siRNA (Fig. 2E and F).
Moreover, Nile Red staining and TG content assay also revealed that
the silencing of FGF21 significantly attenuated the suppressive
effects of miR-212 inhibitor on lipogenesis (Fig. 2I and J). The same experiments were
also performed for miR-212-3p and CHD1 (Fig. 2G, H, K and L). The data indicated
that the effect of miR-212-3p on lipogenesis was achieved by
negatively regulating FGF21/CHD1. FGF-21/CHD1 is a target gene of
miR-212-3p in lipogenesis.
Inhibition of miR-212-3p expression
mimics exercise to improve lipid accumulation induced by a HFD
miR-212-3p was significantly elevated in MAFLD and
could be decreased through exercise. On this basis, a HFD and HE
model was constructed to explore the therapeutic potential of
targeting miR-212-3p. The workflow is illustrated in Fig. 3A. Exercise therapy was used as a
positive control. After 16 weeks on a HFD, the mice were obese. The
increased body weight and liver index were reduced by inhibiting
miR-212-3p as effectively as exercise therapy (Fig. 3B and C). The long-term feeding of
the HFD led to an elevation in the levels of ALT and AST, which
were reduced through the inhibition of miR-212-3p (Fig. 3D). The liver and serum TG contents
both exhibited similar downward trends following the inhibition of
miR-212-3p and exercise therapy (Fig.
3E and F). Intrahepatic fat accumulation was determined using
H&E and Oil Red O staining; the inhibition of miR-212-3p
exerted a similar ameliorative effect to that of exercise, as
assessed using the NAS score and lipid droplet quantification
(Fig. 3G-J). The GTT and ITT were
performed to examine glucose metabolism homeostasis. Similar to
exercise therapy, both the GTT and ITT revealed substantially
improved insulin sensitivity and glucose tolerance in the HFD and
SP group (Fig. 3K-M). The fasting
blood glucose and insulin levels of the mice in each group were
detected to construct a HOMA-IR. The results revealed that the
inhibition of miR-212-3p ameliorated the impairment of insulin
signaling in mice fed the HFD, similar to that of exercise therapy
(Fig. 3N-P). These results
indicated that the inhibition of miR-212-3p exerted ameliorative
effects on MAFLD similar to those of exercise.
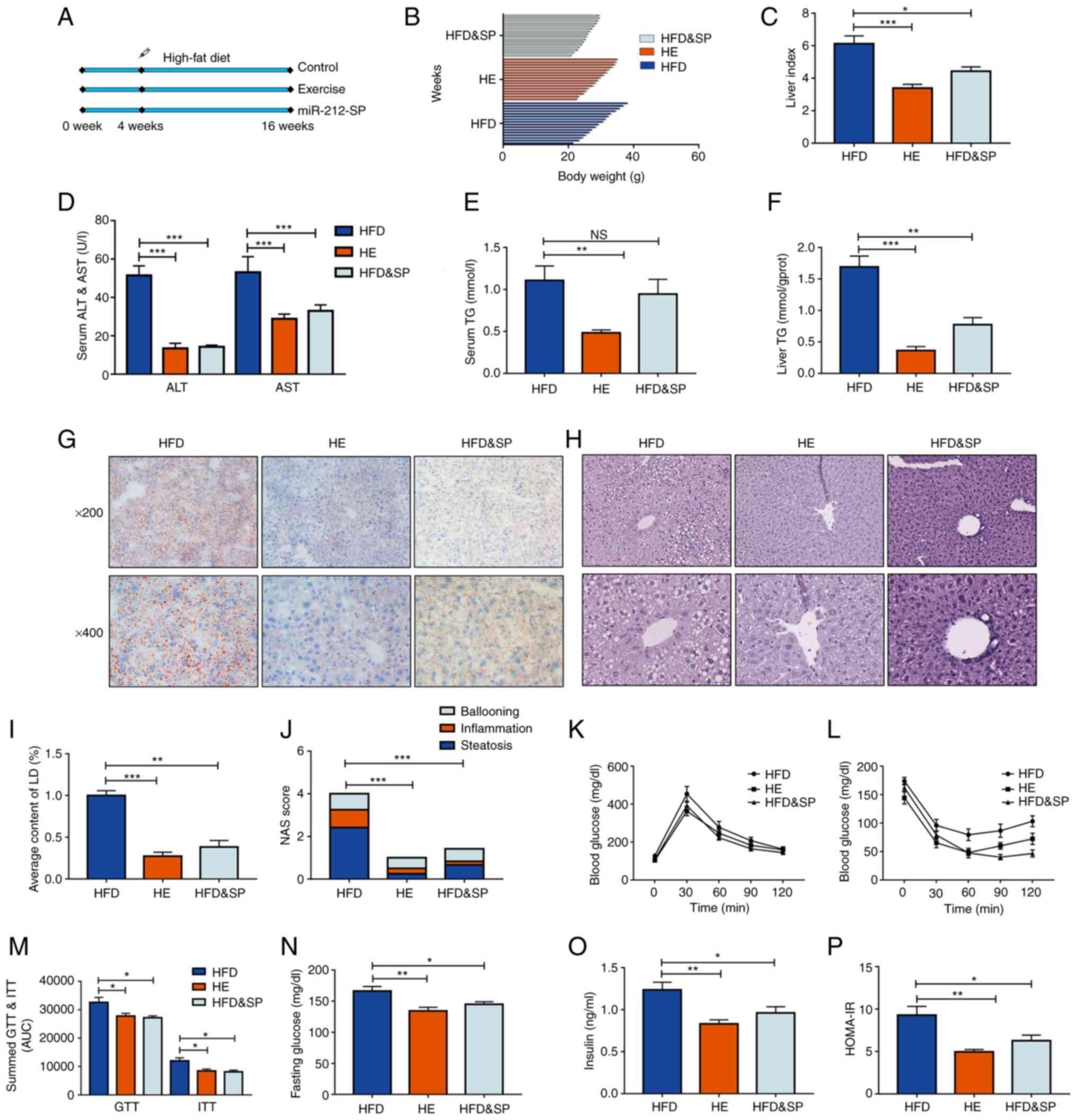 | Figure 3Inhibition of miR-212-3p expression
mimics the effects of exercise to improve lipid accumulation
induced by a HFD. (A) The experimental protocol for the animals.
(B-D) The body weight, liver index, and serum ALT and AST levels
were increased in the HFD group, while they were decreased in the
HFD and SP, and HE group. (E and F) The liver TG content and serum
TG content exhibited a similar decrease following the inhibition of
miR-212-3p compared with exercise treatment. (G-J) Hematoxylin and
eosin and Oil Red O staining confirmed the ameliorative effect of
miR-212-3p inhibition on lipid accumulation. (K-M) Similar to
exercise treatment, the inhibition of miR-212-3p improved glucose
homeostasis through GTT and ITT. (N-P) Similar to exercise therapy,
the inhibition of miR-212-3p improved insulin resistance through
fasting blood glucose, serum insulin and the calculated HOMA-IR.
The data are presented as the mean ± SEM. *P<0.05,
**P<0.01 and ***P<0.001. NS, not
significant; HFD, high-fat diet; ALT, alanine transaminase; AST,
aspartate transaminase; SP, serotype (mice that received the tail
vein injection of recombinant adeno-associated virus serotype 8
gene vectors); TG, triglyceride; LD, lipid droplets; NAS, NAFLD
activity score; GTT, glucose tolerance test; ITT, insulin tolerance
test; HOMA-IR, homeostasis model assessment of insulin resistance;
HE, high-fat diet and exercise. |
miR-212-3p plays a key role in the
effects of exercise on MAFLD
The aforementioned results suggested that the
inhibition of miR-212-3p reduced lipid accumulation as effectively
as exercise therapy. Of note, exercise decreased miR-212-3p
expression in the HFD group. However, whether miR-212-3p plays a
key role in the effects of exercise on MAFLD remained to be
determined. To further explore this, hepatic miR-212-3p was
overexpressed in the HE group through AAV to explore its function.
The HFD group was used as a negative control to verify the effects
of exercise. The workflow is presented in Fig. 4A. Body weight, liver index, serum
ALT and AST all increased as a result of the high expression of
miR-212-3p in the HE and OE group, compared with the HE group
(Fig. 4B-D). With the
overexpression of miR-212-3p, hepatic TG and serum TG levels also
significantly increased even with the intervention of exercise
(Fig. 4E and F). Pathological
evidence collected through the consideration of the NAS score and
lipid droplet quantification further indicated that the
overexpression of miR-212-3p promoted hepatic fatty infiltration
similar to that of the HFD group (Fig. 4G-J). The GTT and ITT assays
indicated that the overexpression of miR-212-3p induced a
dysregulation of glucose homeostasis compared to that of the HE
group. The corresponding curve was significantly higher in the HE
and OE group (Fig. 4K-M).
Moreover, the HOMA-IR also revealed that the overexpression of
miR-212-3p abolished the ameliorative effects of long-term regular
exercise on insulin resistance (Fig.
4N-P). These results indicated that the overexpression of
miR-212-3p abolished the therapeutic effects of exercise on
MAFLD.
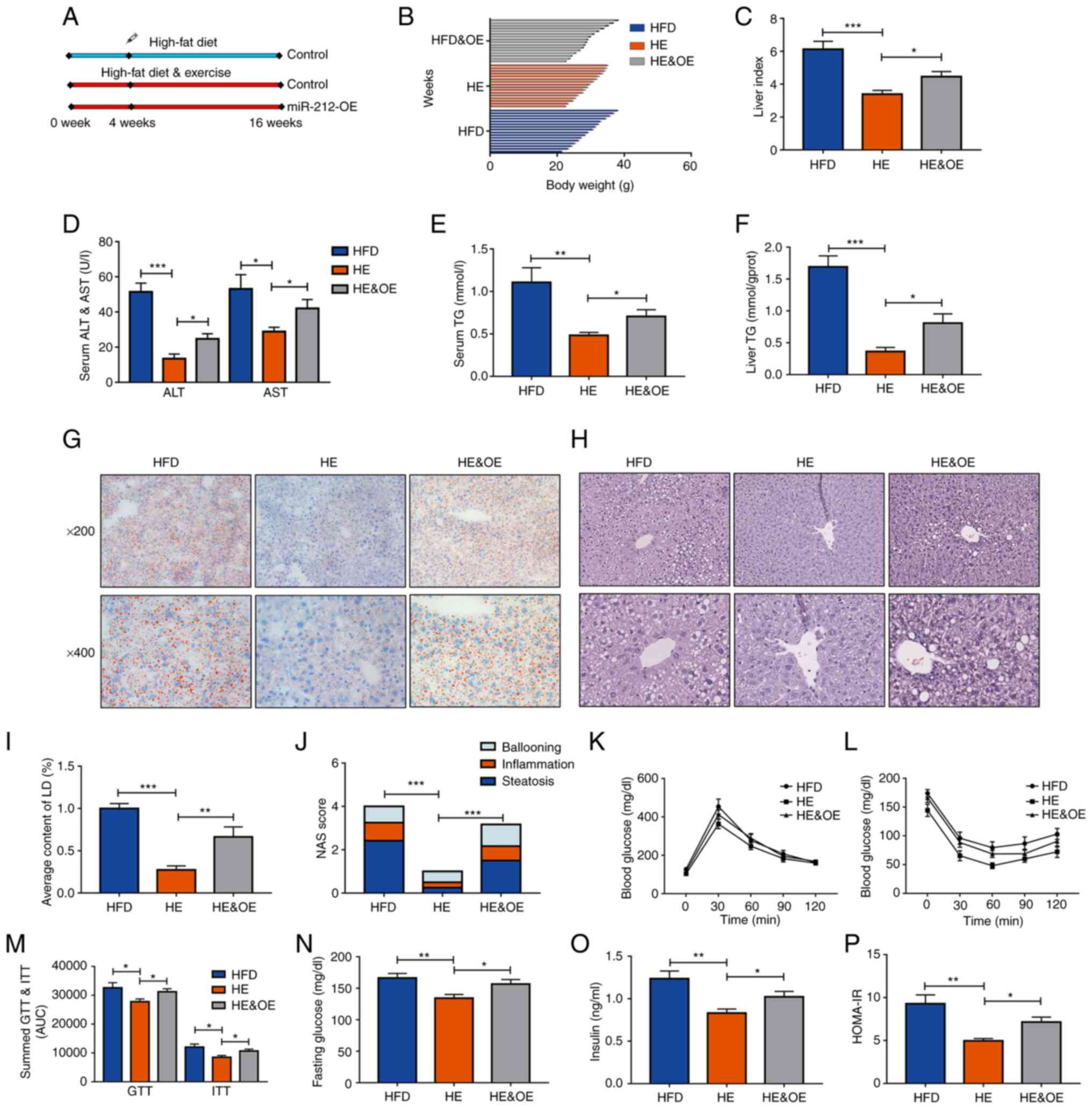 | Figure 4The overexpression of miR-212-3p
blocks the beneficial effects of exercise on MAFLD. (A) The
experimental procedures on animals. (B-D) The body weight, liver
index, and serum ALT and AST levels were decreased in the HE group,
while they were increased in the HE and OE, and HFD group. (E and
F) The liver and serum TG content exhibited similar increase
following the overexpression of miR-212-3p compared with the HFD
group. (G-J) Pathological evidence by NAS score and lipid droplet
quantification suggested that the overexpression of miR-212-3p
promoted hepatic fatty infiltration similar to the HFD group. (K-M)
The high expressed miR-212-3p resulted in dysregulation of glucose
homeostasis, as determined by GTT and ITT. (N-P) Overexpression of
miR-212-3p abolished the ameliorative effects of long-term regular
exercise in insulin resistance. The data are presented as the mean
± SEM. *P<0.05, **P<0.01 and
***P<0.001. HFD, high-fat diet; ALT, alanine
transaminase; AST, aspartate transaminase; OE, overexpression; TG,
triglyceride; LD, lipid droplets; NAS, NAFLD activity score; GTT,
glucose tolerance test; ITT, insulin tolerance test; HOMA-IR,
homeostasis model assessment of insulin resistance; HE, high-fat
diet and exercise. |
The function of miR-212-3p is associated
with regulating hepatic FGF21
The inhibition of miR-212-3p exerted beneficial
effects on MAFLD similar to those of exercise therapy. Conversely,
the overexpression of miR-212 abolished the ameliorative effects of
exercise on MAFLD. The present study then further explored the
mechanisms of miR-212-3p in these processes. Consistent with the
in vitro experiments, FGF21 was a target gene of miR-212-3p
in vivo (Fig. 5A and D).
However, CHD1 was also detected and it was not regulated by
miR-212-3p in both the HFD and HE models (Fig. 5A and F). These results suggested
that miR-212-3p played a role in the beneficial effects of exercise
on MAFLD by regulating FGF21 rather than CHD1 in vivo.
Subsequently, the activation state of the FGF21 pathway was
detected to further verify the association between miR-212-3p and
FGF21. Compared with the HFD group, the expression of p-ERK1/2 and
EGR1 significantly increased following the inhibition of miR-212-3p
(Fig. 5A, B, E, I and J).
Conversely, the expression of p-ERK1/2 markedly decreased following
the overexpression of miR-212-3p (Fig. 5A and B). In addition, the
overexpression of miR-212-3p enhanced lipogenic activity, as shown
by the upregulation of FASN mRNA and protein levels (Fig. 5C, G and J). The inhibition of
miR-212-3p enhanced lipolytic activity, as shown by the increase in
CPT1α mRNA and protein levels (Fig.
5C, H and J). Finally, ELISA, immunohistochemistry and
luciferase reporter assay were used to re-validate the association
between miR-212-3p and FGF21. The results confirmed that the
functions of miR-212-3p were associated with the negative
regulation of hepatic FGF21 (Figs. 5K
and L; and S2).
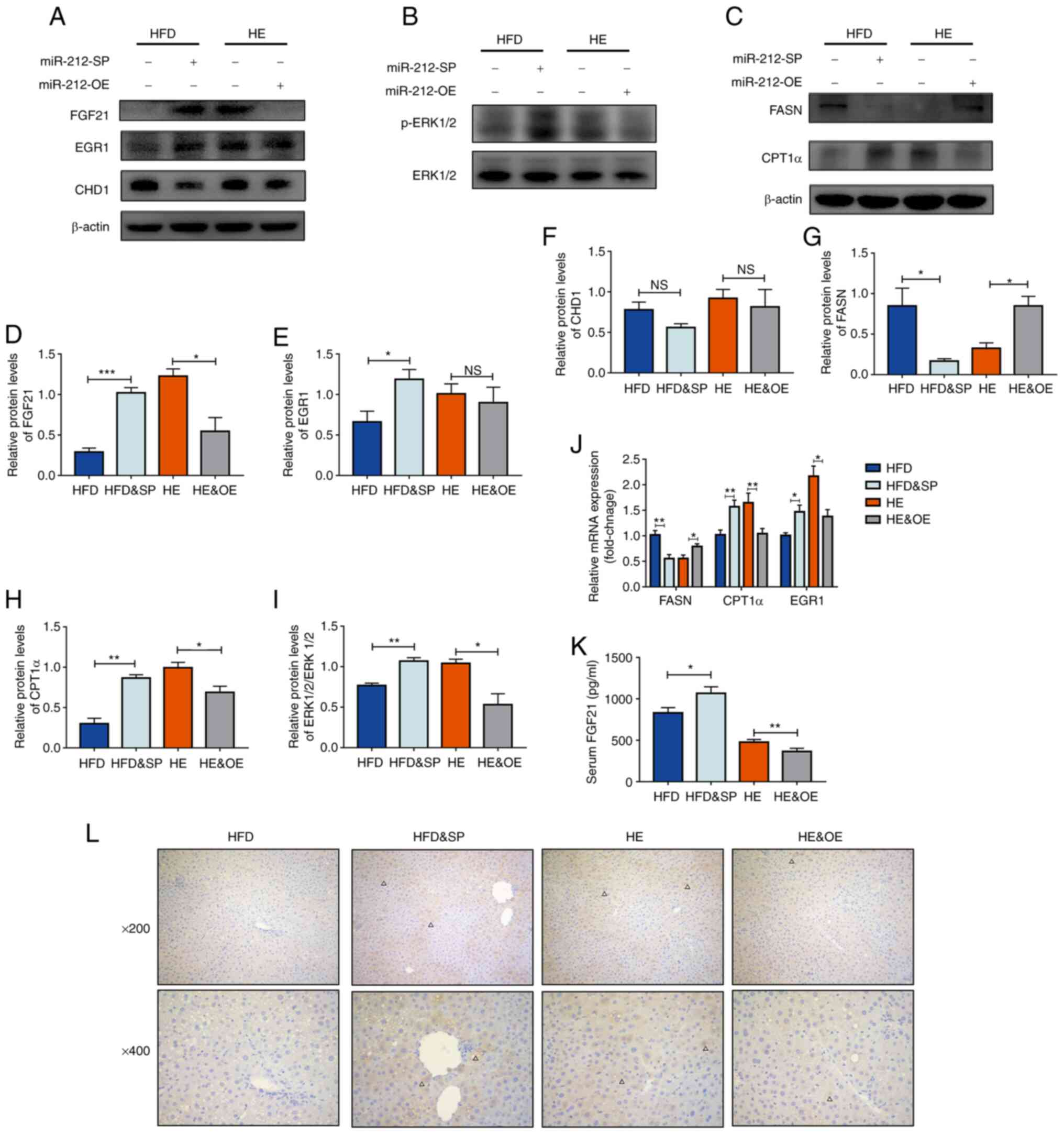 | Figure 5The function of miR-212-3p is
associated with the regulation of hepatic FGF21. (A and B)
miR-212-3p mimics the effects of exercise on MAFLD by regulating
the FGF21 pathway, but not CHD1. (C) The inhibition of miR-212-3p
suppresses lipogenesis, whereas the overexpression of miR-212-3p
promotes lipogenesis. (D-I) Protein quantification of FGF21, EGR1,
CHD1, p-ERK1/2, FASN and CPT1α. (J) Relative mRNA levels of FASN,
CPT1α and EGR1. (K) Serum ELISA levels of FGF21. (L)
Immunohistochemistry of FGF21. The positive areas are marked by
triangles. The data are presented as the mean ± SEM.
*P<0.05, **P<0.01 and
***P<0.001. NS, not significant; FGF21, fibroblast
growth factor 21; CHD1, chromodomain helicase DNA binding protein
1; EGR1, early growth response 1; FASN, fatty acid synthase; CPT1α,
carnitine palmitoyltransferase 1α; HFD, high-fat diet; HE, high-fat
diet and exercise; SP, serotype (mice that received the tail vein
injection of recombinant adeno-associated virus serotype 8 gene
vectors). |
Exercise improves lipid accumulation by
inhibiting miR-212-3p via activating EGR1
The inhibition of miR-212-3p mimicked the
therapeutic effects of exercise on MAFLD, while the overexpression
of miR-212-3p abolished the beneficial effects of exercise on
MAFLD. miR-212-3p appeared to play a key role in the beneficial
effects of exercise on MAFLD. To confirm the link between exercise
and miR-212-3p, a transcription factor motif analysis of the
miR-212-3p promoter region was performed. Sequences of genes in the
upstream region of the miR-212-encoding gene in human and mouse
genomes were accessed through the online genome browser UCSC
(28) (Fig. 6A). A total of 18 transcription
factors that could bind to the sequences in this region were also
predicted online using Jasper and regRNA 2.0 software (29,30). Four transcription factors were
found to have simultaneous upstream region binding sites for
miR-212 in both humans and mice (Table SIII). Subsequently, the MGI
mammalian phenotype was used to analyze the functions and pathways
of the aforementioned 18 transcription factors and found that EGR1
may be simultaneously involved in regulating body weight, abnormal
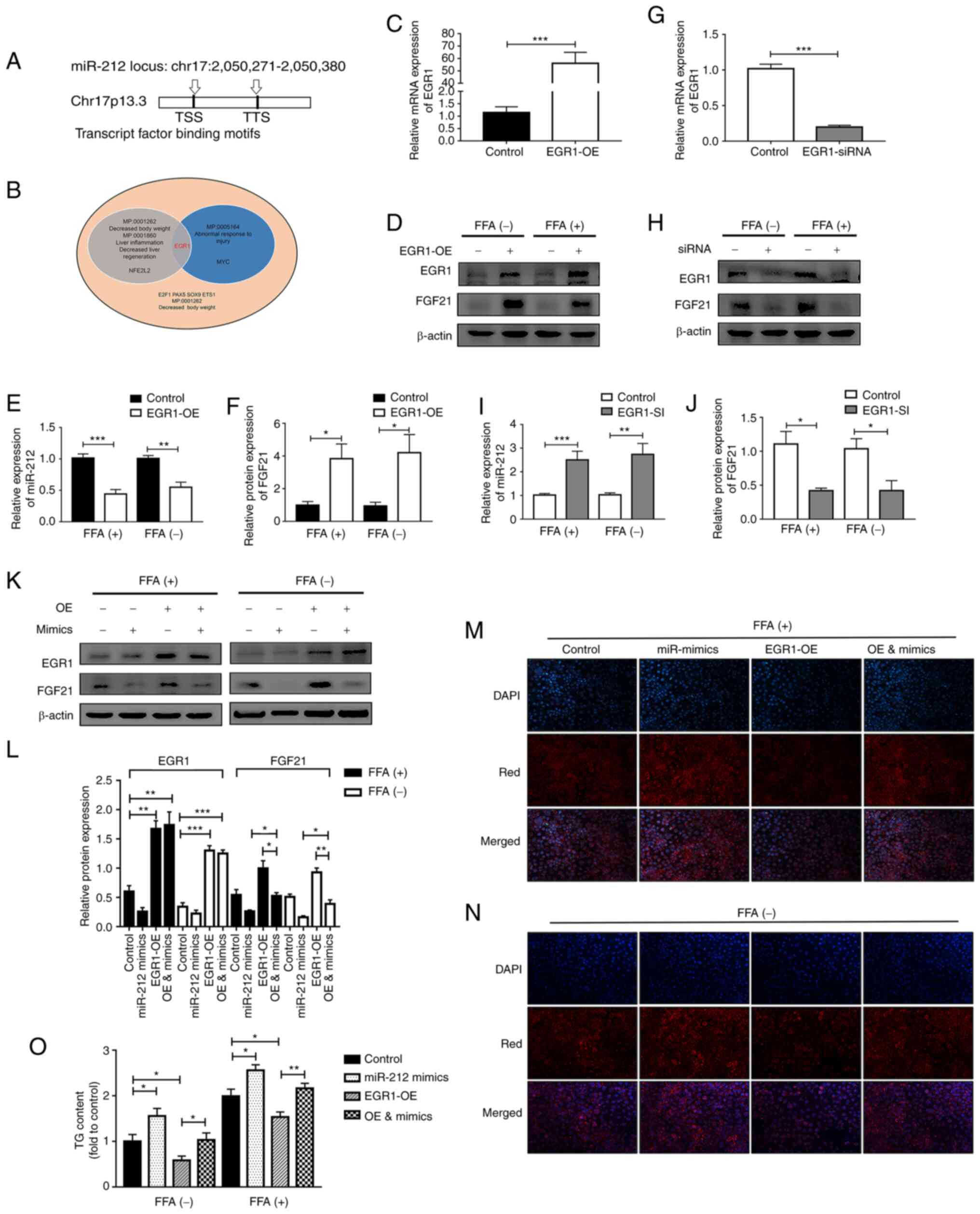
injury response and liver inflammation (Fig. 6B). Of note, EGR1 is an
exercise-related transcription factor; its expression can increase
11.4-fold following exercise (33). Notably, EGR1 was also found to be
a downstream target gene of FGF21.
Gain and loss of function assays for EGR1 were
performed separately. In the case of EGR1 overexpression, the
expression of miR-212-3p was suppressed and that of FGF21
significantly increased, both with or without FFA treatment
(Fig. 6C-F). Conversely, the
knockdown of EGR1 significantly increased miR-212-3p expression,
whereas it decreased FGF21 expression (Fig. 6G-J). Subsequently, rescue
experiments were performed to clarify whether EGR1 regulates FGF21
via miR-212-3p. The corresponding increase in FGF21 expression when
EGR1 was overexpressed was abolished by co-transfection with
miR-212-3p mimics (Fig. 6K and
L). As detected using Nile Red staining and by examining the TG
content, EGR1 overexpression induced a notable decrease in the
lipid content, while this effect was blocked through the high
expression of miR-212-3p, regardless of FFA treatment (Fig. 6M-O).
Finally, luciferase reporter assays were conducted
to further illustrate the direct mechanisms involved in the
interaction between EGR1 and miR-212. 293T cells were transiently
transfected with modified pGL3 vectors containing the 2 kb promoter
segment of miR-212 (containing the -1,500 to +500 bp promoter
sequence of miR-212) (Fig. S3A).
Firefly luciferase expression was used to assess the activity of
the transcriptional promoter. The pRL-TK vector expressing
Renilla luciferase was co-transfected to control for
transfection efficiency. To further analyze the regulatory effects
of EGR1 on pri-miR-212 transcription, EGR1 protein levels were
altered in 293T cells using siRNA and an overexpression plasmid.
Compared to the control group, the overexpression of EGR1 decreased
the luciferase activity of the miR-212 promoter by >50%
(Fig. S3B). Conversely, the
knockdown of EGR1 increased the transcriptional activity (Fig. S3C). It was thus concluded that
the overexpression of miR-212-3p abolished the effects of exercise
on MAFLD by exogenously relieving the transcriptional repression of
EGR1.
Discussion
Exercise is known to be beneficial for MAFLD
treatment (34). During the
course of studying the effects of exercise in the treatment of
MAFLD, a previous study by the authors demonstrated that miR-212-3p
was a key factor in delivering the beneficial effects of exercise
on MAFLD (10). An HFD promotes
liver lipid accumulation, which causes elevated levels of
miR-212-3p. Moreover, exercise improves hepatic lipid accumulation,
which decreases miR-212-3p expression in patients with MAFLD.
Despite the strong association between miR-212-3p, exercise and
MAFLD, existing data on the gain of function, loss of function and
rescue experiments lack clarity on whether miR-212-3p is critical
for exercise to protect against MAFLD (10). Therefore, the present study
focused on the role of miR-212-3p in the progression of MAFLD.
Mature miRNAs are derived from the shearing of
precursors, whereby miR-212-3p and miR-212-5p are stably expressed.
Studies have reported that a leucine-deficient diet can lead to an
increase in the expression of miR-212-5p, which in turn decreases
lipogenesis by targeting FASN and stearoyl-CoA desaturase 1
(16). Therefore, the present
study first examined whether miR-212-5p also plays a role using a
mouse model of HFD-induced MAFLD. In the HFD and exercise mouse
model, miR-212-3p, but not miR-212-5p, was regulated by the
high-fat environment and exercise intervention. However, whether
the elevation of miR-212-3p expressions was responsible for the
HFD-induced intrahepatic lipid accumulation in MAFLD also needed to
be determined. Pathological and serological indicators revealed
that the inhibition of miR-212-3p effectively inhibited
intrahepatic lipid accumulation. Exercise inhibited the high
expression of miR-212-3p in MAFLD, while the exogenous high
expression of miR-212-3p effectively blocked the lipid-lowering
effects of exercise. Based on the aforementioned results, it can be
suggested that miR-212-3p is a key factor involved in the
effectiveness of exercise. After confirming that miR-212-3p was
critical for the protective effects of exercise against MAFLD, the
present study further examined the mechanisms through which the
effects of miR-212-3p were achieved. For this purpose, two
potential targets were systematically screened out through target
prediction, functional enrichment and literature curation.
Furthermore, it was confirmed that miR-212-3p exerted its effects
on lipogenesis through the regulation of CHD1 and FGF21 in
vitro. However, under a more complex environment in
vivo, only FGF21 was regulated by miR-212-3p. Therefore, these
results suggested that the utility of miR-212-3p in lipogenesis was
achieved through the regulation of FGF21 both in vitro and
in vivo.
To elucidate the association between exercise and
miR-212-3p, a transcription factor motif prediction analysis was
performed along with mammalian phenotyping to gain further insight
into the mechanisms (35). EGR1
is a member of the immediate early genes family and plays a
critical role in regulating cell growth and proliferation (36). It has been shown that EGR1 is
involved in the inhibitory effects of leptin on PPARγ, suggesting
that EGR1 may inhibit lipid accumulation (37). Of note, EGR1 responds to exercise
in the early stages, that is, exercise can directly and rapidly
alter the expression level of EGR1 (33). An increase of as much as 11.4-fold
in EGR1 expression can be observed following exercise. EGR1
expression is increased after an organism is stimulated with the
recombinant protein FGF21, and EGR1 has been found to be the
downstream target gene of the FGF21 signaling pathway (38,39). Taken together, these findings may
explain why the overexpression of miR-212-3p in the exercise model
did not cause an increase in EGR1 expression (Fig. 5A and E). The association between
EGR1, miR-212-3p and FGF21 is crucial for delivering the beneficial
effects of exercise on MAFLD. Through a luciferase reporter gene,
gain of function, loss of function and rescue experiments, the
present study successfully confirmed that EGR1 can increase FGF21
expression through the transcriptional inhibition of miR-212-3p.
FGF21 is a myokine which can improve the metabolic regulation of
systemic organs when produced by skeletal muscle in response to
exercise (15). A previous study
demonstrated that mice overexpressing FGF21 are were to resist
HFD-induced obesity (40).
Another study demonstrated that the injection of FGF21 recombinant
protein was able to attenuate obesity-induced hyper-TG levels and
liver damage (41). An increasing
number of studies have suggested that FGF21 is a potent anti-MAFLD
molecule (42,43). In the present study, in the
exercise model, it was observed that the overexpression of
miR-212-3p blocked the increase in the expression of FGF21 by EGR1,
and significantly increased lipid accumulation. At the same time,
the FGF21 pathway, which was activated through exercise, was also
blocked by miR-212-3p. Furthermore, when miR-212-3p expression was
inhibited, the regulation of FGF21 by EGR1 was restored. The data
also indicated that the FGF21 pathway was once again activated and
lipid accumulation was reduced.
Finally, the present study still has some
limitations. It was observed that targeting miR-212-3p
significantly improved lipid deposition and prevented NAFLD.
However, a greater number of patients need to be treated when they
already have NAFLD. In this case, it remains to be determined
whether miR-212-3p would be equally effective. In addition, the
regulatory association between EGR1, miR-212-3p and FGF21 was
supported by in vitro experiments; however, whether the
in vivo environment would still effective warrants to be
further exploration.
In conclusion, the inhibition of miR-212-3p exerts
effects on MAFLD similar to those of exercise. Gene therapy
targeting miR-212-3p may thus mimic the ameliorative effects of
exercise on MAFLD and may provide novel strategies for the
comprehensive treatment of patients with MAFLD, particularly those
who cannot adhere to or tolerate exercise programs.
Supplementary Data
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
BS and YZ were involved in data curation and
analyses. MZ was involved in the study supervision and data
validation. RL and WY were involved in the conceptualization of the
study, in the formal analysis and in funding acquisition. BS and YZ
confirm the authenticity of all the raw data. All authors have read
and approved the final manuscript.
Ethics approval and consent to
participate
For the use of patient samples, the present study
was approved by the Ethics Committee of Tongji Hospital (approval
no. K-KYSB-2020-139) and was performed in accordance with the
Declaration of Helsinki. Written informed consent was obtained from
the patients or their guardians. The animal experiments were
approved by the Animal Ethics Committee of Tongji University
(approval no. TJHBLAC-2019-024).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was supported by a grant from the National
Natural Science Foundation of China (grant no. 81873567).
References
|
1
|
Simon TG, Roelstraete B, Khalili H,
Hagström H and Ludvigsson JF: Mortality in biopsy-confirmed
nonalcoholic fatty liver disease: Results from a nationwide cohort.
Gut. 70:1375–1382. 2021. View Article : Google Scholar
|
|
2
|
Cotter TG and Rinella M: Nonalcoholic
fatty liver disease 2020: The state of the disease.
Gastroenterology. 158:1851–11864. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Younossi ZM: Patient-reported outcomes and
the economic effects of nonalcoholic fatty liver disease and
nonalcoholic steatohepatitis: The value proposition. Hepatology.
68:2405–2412. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rohilla S, Awasthi A, Kaur S and Puria R:
Evolutionary conservation of long non-coding RNAs in non-alcoholic
fatty liver disease. Life Sci. 264:1185602021. View Article : Google Scholar
|
|
5
|
El-Agroudy NN, Kurzbach A, Rodionov RN,
O'Sullivan J, Roden M, Birkenfeld AL and Pesta DH: Are lifestyle
therapies effective for NAFLD treatment? Trends Endocrinol Metab.
30:701–709. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Takahashi H, Kotani K, Tanaka K, Egucih Y
and Anzai K: Therapeutic approaches to nonalcoholic fatty liver
disease: Exercise intervention and related mechanisms. Front
Endocrinol (Lausanne). 9:5882018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Keating SE, Hackett DA, Parker HM,
O'Connor HT, Gerofi JA, Sainsbury A, Baker MK, Chuter VH, Caterson
ID, George J and Johnson NA: Effect of aerobic exercise training
dose on liver fat and visceral adiposity. J Hepatol. 63:174–182.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Glass O, Filozof C, Noureddin M,
Berner-Hansen M, Schabel E, Omokaro SO, Schattenberg JM, Barradas
K, Miller V, Francque S, et al: Standardisation of diet and
exercise in clinical trials of NAFLD-NASH: Recommendations from the
liver forum. J Hepatol. 73:680–693. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fabian MR and Sonenberg N: The mechanics
of miRNA-mediated gene silencing: A look under the hood of miRISC.
Nat Struct Mol Biol. 19:586–593. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xiao J, Bei Y, Liu J, Dimitrova-Shumkovska
J, Kuang D, Zhou Q, Li J, Yang Y, Xiang Y, Wang F, et al: MiR-212
downregulation contributes to the protective effect of exercise
against non-alcoholic fatty liver via targeting FGF-21. J Cell Mol
Med. 20:204–216. 2016. View Article : Google Scholar
|
|
11
|
Liang X, Zeng J, Wang L, Fang M, Wang Q,
Zhao M, Xu X, Liu Z, Li W, Liu S, et al: Histone demethylase
retinoblastoma binding protein 2 is overexpressed in hepatocellular
carcinoma and negatively regulated by hsa-miR-212. PLoS One.
8:e697842013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kumarswamy R, Volkmann I, Beermann J, Napp
LC, Jabs O, Bhayadia R, Melk A, Ucar A, Chowdhury K, Lorenzen JM,
et al: Vascular importance of the miR-212/132 cluster. Eur Heart J.
35:3224–3231. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tang Y, Banan A, Forsyth CB, Fields JZ,
Lau CK, Zhang LJ and Keshavarzian A: Effect of alcohol on miR-212
expression in intestinal epithelial cells and its potential role in
alcoholic liver disease. Alcohol Clin Exp Res. 32:355–364. 2008.
View Article : Google Scholar
|
|
14
|
Mollet IG, Malm HA, Wendt A, Orho-Melander
M and Eliasson L: Integrator of stress responses calmodulin binding
transcription activator 1 (Camta1) regulates miR-212/miR-132
expression and insulin secretion. J Biol Chem. 291:18440–18452.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ucar A, Gupta SK, Fiedler J, Erikci E,
Kardasinski M, Batkai S, Dangwal S, Kumarswamy R, Bang C, Holzmann
A, et al: The miRNA-212/132 family regulates both cardiac
hypertrophy and cardiomyocyte autophagy. Nat Commun. 3:10782012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guo Y, Yu J, Wang C, Li K, Liu B, Du Y,
Xiao F, Chen S and Guo F: miR-212-5p suppresses lipid accumulation
by targeting FAS and SCD1. J Mol Endocrinol. 59:205–217. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kleiner DE, Brunt EM, Van Natta M, Behling
C, Contos MJ, Cummings OW, Ferrell LD, Liu YC, Torbenson MS,
Unalp-Arida A, et al: Design and validation of a histological
scoring system for nonalcoholic fatty liver disease. Hepatology.
41:1313–1321. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Saunders JB, Aasland OG, Babor TF, de la
Fuente JR and Grant M: Development of the alcohol use disorders
identification test (AUDIT): WHO collaborative project on early
detection of persons with harmful alcohol consumption-II.
Addiction. 88:791–804. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Park SE, Lee SW, Hossain MA, Kim MY, Kim
MN, Ahn EY, Park YC, Suh H, Kim GY, Choi YH and Kim ND: A
chenodeoxycholic derivative, HS-1200, induces apoptosis and cell
cycle modulation via Egr-1 gene expression control on human
hepatoma cells. Cancer Lett. 270:77–86. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
McGeary SE, Lin KS, Shi CY, Pham TM,
Bisaria N, Kelley GM and Bartel DP: The biochemical basis of
microRNA targeting efficacy. Science. 366:eaav17412019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
ELife. 4:e050052015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Garcia DM, Baek D, Shin C, Bell GW,
Grimson A and Bartel DP: Weak seed-pairing stability and high
target-site abundance decrease the proficiency of lsy-6 and other
microRNAs. Nat Struct Mol Biol. 18:1139–1146. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar :
|
|
24
|
Grimson A, Farh KK, Johnston WK,
Garrett-Engele P, Lim LP and Bartel DP: MicroRNA targeting
specificity in mammals: Determinants beyond seed pairing. Mol Cell.
27:91–105. 2007. View Article : Google Scholar
|
|
25
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sticht C, De La Torre C, Parveen A and
Gretz N: MiRWalk: An online resource for prediction of microRNA
binding sites. PLoS One. 13:e02062392018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Green CD, Huang Y, Dou X, Yang L, Liu Y
and Han JDJ: Impact of dietary interventions on noncoding RNA
networks and mRNAs encoding chromatin-related factors. Cell Rep.
18:2957–2968. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gonzalez JN, Zweig AS, Speir ML, Schmelter
D, Rosenbloom KR, Raney BJ, Powell CC, Nassar LR, Maulding ND, Lee
CM, et al: The UCSC genome browser database: 2021 update. Nucleic
Acids Res. 49:D1046–D1057. 2021. View Article : Google Scholar
|
|
29
|
Fornes O, Castro-Mondragon JA, Khan A, van
der Lee R, Zhang X, Richmond PA, Modi BP, Correard S, Gheorghe M,
Baranašić D, et al: JASPAR 2020: Update of the open-access database
of transcription factor binding profiles. Nucleic Acids Res.
48:D87–D92. 2020.
|
|
30
|
Chang TH, Huang HY, Hsu JB, Weng SL, Horng
JT and Huang HD: An enhanced computational platform for
investigating the roles of regulatory RNA and for identifying
functional RNA motifs. BMC Bioinformatics. 14(Suppl 2): S42013.
View Article : Google Scholar
|
|
31
|
Smith CL and Eppig JT: The mammalian
phenotype ontology: Enabling robust annotation and comparative
analysis. Wiley Interdiscip Rev Syst Biol Med. 1:390–399. 2009.
View Article : Google Scholar
|
|
32
|
Zhou Y, Zhou B, Pache L, Chang M,
Khodabakhshi AH, Tanaseichuk O, Benner C and Chanda SK: Metascape
provides a biologist-oriented resource for the analysis of
systems-level datasets. Nat Commun. 10:15232019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
McLean CS, Mielke C, Cordova JM, Langlais
PR, Bowen B, Miranda D, Coletta DK and Mandarino LJ: Gene and
microRNA expression responses to exercise; relationship with
insulin sensitivity. PLoS One. 10:e01270892015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Orci LA, Gariani K, Oldani G, Delaune V,
Morel P and Toso C: Exercise-based interventions for nonalcoholic
fatty liver disease: A meta-analysis and meta-regression. Clin
Gastroenterol Hepatol. 14:1398–1411. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hayamizu TF, Baldock RA and Ringwald M:
Mouse anatomy ontologies: Enhancements and tools for exploring and
integrating biomedical data. Mamm Genome. 26:422–430. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Han MS, Perry RJ, Camporez JP, Scherer PE,
Shulman GI, Gao G and Davis RJ: A feed-forward regulatory loop in
adipose tissue promotes signaling by the hepatokine FGF21. Genes
Dev. 35:133–146. 2021. View Article : Google Scholar :
|
|
37
|
Zhou Y, Jia X, Zhou M and Liu J: Egr-1 is
involved in the inhibitory effect of leptin on PPARgamma expression
in hepatic stellate cell in vitro. Life Sci. 84:544–551. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li J, Xu C, Liu Y, Li Y, Du S, Zhang R,
Sun Y, Zhang R, Wang Y, Xue H, et al: Fibroblast growth factor 21
inhibited ischemic arrhythmias via targeting miR-143/EGR1 axis.
Basic Res Cardiol. 115:92020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Adams AC, Yang C, Coskun T, Cheng CC,
Gimeno RE, Luo Y and Kharitonenkov A: The breadth of FGF21's
metabolic actions are governed by FGFR1 in adipose tissue. Mol
Metab. 2:31–37. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Samms RJ, Murphy M, Fowler MJ, Cooper S,
Emmerson P, Coskun T, Adams AC, Kharitonenkov A, Ebling FJP and
Tsintzas K: Dual effects of fibroblast growth factor 21 on hepatic
energy metabolism. J Endocrinol. 227:37–47. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang WF, Li SM, Ren GP, Zheng W, Lu YJ, Yu
YH, Xu WJ, Li TH, Zhou LH, Liu Y and Li DS: Recombinant murine
fibroblast growth factor 21 ameliorates obesity-related
inflammation in monosodium glutamate-induced obesity rats.
Endocrine. 49:119–129. 2015. View Article : Google Scholar
|
|
42
|
Maratos-Flier E: Fatty liver and FGF21
physiology. Exp Cell Res. 360:2–5. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Byun S, Seok S, Kim YC, Zhang Y, Yau P,
Iwamori N, Xu HE, Ma J, Kemper B and Kemper JK: Fasting-induced
FGF21 signaling activates hepatic autophagy and lipid degradation
via JMJD3 histone demethylase. Nat Commun. 11:8072020. View Article : Google Scholar : PubMed/NCBI
|