Introduction
The human skeleton is a dynamic system (1). Osteoclasts, osteoblasts and
osteocytes regulate bone mass, and the disruption of the balance of
activity between these cells can result in bone diseases, such as
osteoporosis and periodontal disease, which can severely endanger
human health (2,3). Controlling the differentiation and
functionality of these cells is thus crucial from a therapeutic
standpoint. An imbalance in the activity of osteoclasts and
osteoblasts caused by an increase in the quantity and function of
osteoclasts as a result of estrogen deprivation is the hallmark of
postmenopausal osteoporosis, a prevalent skeletal illness affecting
older women (4,5). The course of postmenopausal
osteoporosis can be attenuated by suppressing osteoclast formation
and activity, according to pertinent studies (6,7).
Osteoclasts are known for their bone resorbing
function and are a special type of terminally differentiated cell
that originates from the blood lineage monocyte-macrophage system
(8). Macrophage
colony-stimulating factor (M-CSF) and receptor activator of nuclear
factor κB (NF-κB) ligand (RANKL) control osteoclast formation
(9,10). Multiple intracellular signaling
events are activated by the interaction of RANKL with its receptor,
RANK, which influences the function and survival of osteoclasts.
These effectors stimulate the expression and activation of
transcription factors, such as nuclear factor of activated T cells,
cytoplasmic 1 (NFATc1), which is required for osteoclast precursor
differentiation (11,12). The inhibition of the intracellular
process that is triggered by RANKL is thus considered to be a
crucial therapeutic objective for the treatment of postmenopausal
osteoporosis (13).
Casticin (Cas) is a methoxy flavonoid compound
obtained from Vitex trifolia with medicinal properties
(14). Cas has been shown to
possess anti-inflammatory and antitumor activities (15,16). In addition, Cas has been found to
promote wound healing and to improve the symptoms of menopause in
ovariectomized rats (17,18). Previous studies have demonstrated
that osteoclast activity and osteolytic bone diseases are directly
associated to inflammation (19,20). The aim of the present study was to
ascertain whether Cas can affect the differentiation and function
of osteoclasts, and explore its probable mechanisms of action, as
its role in osteoclastogenesis has not yet been clarified. The
protective effects of Cas in a mouse model of estrogen deficiency
highlight its potential to prevent postmenopausal osteoporosis.
Materials and methods
Cell source and animal care
In this part of the experiment, 20 SPF grade C57BL/6
female mice aged 6-8 weeks and weighing ~20 g were obtained from
the Animal Experiment Center of Guangxi Medical University
(Nanning, China). The animal experiments were performed after
approval (approval no. 202110004) from the Animal Care and Welfare
Committee of Guangxi Medical University. The mice were euthanized
after being suffocated with carbon dioxide at a displacement rate
of 50%/min, the long bones of both lower limbs were removed, and
the bone marrow cavities were rinsed with α-MEM (Thermo Fisher
Scientific, Inc.) to collect cells. Mouse bone macrophage precursor
cells were collected by flushing the bone marrow volume of all long
bones and culturing for 4 days under complete medium [α-MEM, 1%
penicillin/streptomycin-mixed antibiotics and 10% fetal bovine
serum (FBS; Thermo Fisher Scientific, Inc.) conditions and 25 ng/ml
M-CSF (R&D Systems, Inc.)], followed by digestion of the cells
with trypsin for 5 min, centrifugation at 120 × g, at 25°C for 5
min, and suspension counting, for use in subsequent experiments.
The mice were maintained at the laboratory of pathogen-free animals
of Guangxi Medical University under conditions of constant
temperature at 25°C, 60% constant humidity, 12 h of alternating
light, unrestricted activity, and SPF-grade normal mouse chow
available ad libitum. The cell culture conditions were a
37°C constant temperature and a constant CO2
concentration of 5%.
Antibodies and reagents
Cas (purity, ≥98%; purchased from Chengdu Must
Biotechnology Co., Ltd.) was dissolved in dimethyl sulfoxide (DMSO)
and diluted with α-MEM (Thermo Fisher Scientific, Inc.) to the
desired experimental concentration. FBS was obtained from Thermo
Fisher Scientific, Inc. MedChemExpress provided the CCK-8 kit.
R&D Systems, Inc. provided recombinant mouse M-CSF and RANKL.
Penicillin/streptomycin-mixed antibiotics, trypsin and bovine serum
albumin (BSA) were from Gibco; Thermo Fisher Scientific, Inc.
Rhodamine-phalloidin, DAPI, PBS, 4% paraformaldehyde tissue
fixative, Triton X-100, β-estradiol and Alizarin Red S solution
were purchased from Beijing Solarbio Science & Technology Co.,
Ltd., and Beyotime Institute of Biotechnology provided the alkaline
phosphatase assay kit. Santa Cruz Biotechnology, Inc. provided the
p65 fluorescent antibodies (cat. no. sc-8008 Alexa
Fluor® 546; 1:100 dilution) used for p65 nuclear
translocation, and the primary antibodies, NFATc1 (cat. no. sc7294;
1:200 dilution) and cathepsin K (CTSK; cat. no. sc-48353; 1:200
dilution), used for western blot analysis. Abcam provided the c-Fos
antibody (cat. no. ab134122; 1:500 dilution). The majority of the
remaining primary antibodies were produced by Cell Signaling
Technology, Inc., including NF-κB p65 (product no. 8242; 1:1,000
dilution), phosphorylated (p)-p65 (product no. 3033; 1:1,000
dilution), NF-κB1 p105/p50 (product no. 13586; 1:1,000 dilution),
IκBα (product no. 4812, 1:1,000 dilution), ERK1/2 (product no.
4695; 1:1,000 dilution), p-ERK1/2 (product no. 4370; 1:2,000
dilution), p38 (product no. 8690; 1:1,000 dilution), p-p38 (product
no. 4511; 1:1,000 dilution), JNK (product no. 9252; 1:1,000
dilution), p-JNK (product no. 4668; 1:1,000 dilution), TAK1
(product no. 4505; 1:1,000 dilution), p-TAK1 (product no. 9339;
1:1,000 dilution), Akt (product no. 9272; 1:1,000 dilution), p-Akt
(product no. 4060; 1:2,000 dilution), GSK3β (product no. 5676;
1:1,000 dilution) and p-GSK3β (product no. 9327; 1:1,000 dilution).
T-75 aerated cell culture flasks, cell-grade sterilized 96-well
cell culture plates, cell-grade sterilized 6-well cell culture
plates, sterile centrifuge tubes, and bovine bone chips were
purchased from Eppendorf.
Tartrate-resistant acid phosphatase
(TRAP) staining and osteoclast differentiation
Bone marrow-derived macrophages (BMMs) were
inoculated in 96-well plates at a density of 6×103
cells/well and incubated overnight at 37°C and 5% CO2.
Following attachment, the plate was transferred to a biosafety
cabinet, and the required concentration of RANKL and Cas was
prepared with pre-warmed medium. Apart from the negative control
group, the positive control and drug intervention groups were
treated with 50 ng/ml RANKL and 0, 0.5, 1, 1.5 or 2 µM Cas.
At the aforementioned concentrations, once per day, the medium was
changed. The plate was gently rinsed with PBS and fixed for 1 h at
25°C with 4% paraformaldehyde following 5-7 days of cell culture.
The cells were then incubated at 50 µl/well for 15 min with
TRAP (Sigma-Aldrich; Merck KGaA) staining solution at room
temperature. The BMMs were stimulated with 50 ng/ml RANKL in the
presence of 2 µM Cas at three stages of osteoclast
development, and control wells were set up and stained as described
above. The 96-well plates were then photographed using Cytation 5
(BioTek Instruments, Inc.) and the number of osteoclasts were
counted using ImageJ software 1.51 (National Institutes of Health,
Bethesda, MD), followed by statistical analysis, specifying
positive staining, with a nuclei number of ≥3 as valid counts.
Cytotoxicity assay
The BMMs were cultured in 96-well plates at a
density of 6×103 cells/well, as described above. The
plates were incubated at 37°C and 5% CO2. The following
day, Cas (0, 0.5, 1, 1.5, 2 or 2.5 µM) was added, and after
96 h, 10 µl CCK-8 reagent was added to each well and
incubated for 2 h at 37°C and 5% CO2. A multimode
microplate reader (Berthold Technologies Gmbh & Co. KG) was
used to measure the absorbance values.
F-actin ring staining
The cell culture conditions were identical to those
used in the osteoclast differentiation assay. Following cell
maturation, the cells were fixed for 1 h at 25°C with 4%
paraformaldehyde, permeabilized for 5 min with a 0.1%
Trixon-100/PBS mixture, and incubated for 30 min with a 3% BSA/PBS
mixture at room temperature. The plate was then incubated for 1 h
at 25°C with rhodamine-phalloidin at a concentration of 1:200
diluted with 0.2% BSA solution. Subsequently, the wells were washed
with 0.2% BSA and stained for 5 min at 25°C with a 1:1,000 DAPI/PBS
mixture, excess liquid was discarded, and the cells were washed and
photographed with a Cytation 5 imaging multimode reader.
Osteoclast acidification assay
To assess the effects of Cas on osteoclast
acidification, acridine orange dye [3,6-bis(dimethylamino)acridine]
(MedChemExpress) was used as previously describd (21). The osteoclasts were cultured in
the presence or absence of 0, 1 or 2 µM Cas, and the culture
conditions were the same as those aforementioned. The cells were
incubated with a mixture of 0 µg/ml acridine orange and
α-MEM for 15 min at 37°C and 5% CO2, and the 96-well
plates washed with PBS were photographed with a Cytation 5 imaging
multimode reader, and images of the wells were obtained and
saved.
Bone resorption assay
Bovine bone slices were processed in advance;
1×105 BMMs were inoculated in six-well plates, and RANKL
(50 ng/ml) was added the following day. The formed osteoclasts were
inoculated in 96-well plates and cultured with the addition of Cas
(0, 1 or 2 µM). Following 1 h of fixation in 4%
paraformaldehyde at room temperature, the bone sections were
cleaned with a brush and imaged using a Regulus 8100 scanning
electron microscope (Hitachi Corporation). ImageJ software 1.51 was
used to analyze the areas of bone resorption.
Reverse transcription-quantitative PCR
(RT-qPCR)
For culture into osteoclasts, 1×105 BMMs
were added to six-well plates and subjected to Cas (0, 1 or 2
µM). Total RNA was extracted using TRIzol®
reagent 5 days later, and cDNA was produced using a Revert-Aid RT
kit (Thermo Fisher Scientific, Inc.). SYBR Green Master (Roche
Diagnostics) dye was used for RT-qPCR. A real-time fluorescence
quantitative PCR instrument was then used to determine the mRNA
expression. The reaction system involved heating to 95°C for
denaturation and holding at 4°C for 55 cycles (99°C, 15 sec; 60°C,
15 sec; 72°C, 40 sec). β-actin, a housekeeping gene, was
used to normalize the gene expression, and the 2−ΔΔCq
method was applied to analyze the data (22). The primer sequences used are
listed in Table I.
 | Table IPrimer sequences used in reverse
transcription-quantitative PCR. |
Table I
Primer sequences used in reverse
transcription-quantitative PCR.
| Gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
|
Atp6v0d2 |
GTGAGACCTTGGAAGACCTGAA |
GAGAAATGTGCTCAGGGGCT |
| Dcstamp |
TCTGCTGTATCGGCTCATCTC |
ACTCCTTGGGTTCCTTGCTT |
| Fos |
CCAGTCAAGAGCATCAGCAA |
AAGTAGTGCAGCCCGGAGTA |
| Nfatc1 |
GGTGCTGTCTGGCCATAACT |
GAAACGCTGGTACTGGCTTC |
| Ctsk |
AGGCGGCTATATGACCACTG |
TCTTCAGGGCTTTCTCGTTC |
| Mmp9 |
GAAGGCAAACCCTGTGTT |
AGAGTACTGCTTGCCCAGGA |
| β-actin |
TCTGCTGGAAGGTGGACAGT |
CCTCTATGCCAACACAGTGC |
Western blot analysis
To demonstrate the mechanisms through which Cas
inhibits signaling pathways, the BMMs were seeded in six-well
plates at a density of 5×105 cells per well. Before
being stimulated for 1 h with 2 µM Cas, the BMMs were
starved with α-MEM for 3 h, and RANKL (50 ng/ml) stimulation was
then applied for 0, 5, 10, 20, 30 and 60 min. The BMMs were
inoculated at 1×105 cells/well in six-well plates and
stimulated for 1, 3 and 5 days with Cas (2 µM) and RANKL (50
ng/ml), followed by lysis of the cells using RIPA buffer (Beijing
Kangwei Century Biotechnology Co., Ltd.) to extract the proteins
and protein concentrations were assessed using a BCA kit (Beyotime
Institute of Biotechnology). Subsequently, 10% gels were prepared
according to the PAGE Gel Fast Preparation kit instructions (cat.
no. PG112; Shanghai Epizyme Biomedical Technology Co., Ltd.), 40
µg of proteins were loaded per lane, which were separated
using the 10% SDS-PAGE, before transfer of the isolated proteins
onto nitrocellulose membranes and blocking with 5% skim milk powder
for 1 h at room temperature. The membranes were incubated with
primary antibodies diluted in 5% BSA-PBS for 18 h before being
placed in cold storage (4°C). The membranes were then incubated
with the corresponding secondary antibodies [goat anti-rabbit/mouse
IgG (H+L), DyLight 800 4×PEG, cat. nos. SA5-35571/SA5-35521,
respectively, Invitrogen; Thermo Fisher Scientific, Inc.; diluted
in 5% skim milk] the following day for 1 h at room temperature and
imaged using an Image Quant LAS-4000 system (GE Healthcare; Cytiva)
to observe the target bands. ImageJ software 1.51 was used for
densitometric analysis.
p65 nuclear translocation
The BMMs were seeded in confocal culture dishes at a
cell density of 1×105 and starved with α-MEM for 3 h
after 1 day, followed by Cas (2 µM) intervention for 1 h and
RANKL stimulation for 10 min. The cells were then fixed with 4%
paraformaldehyde for 10 min at 37°C and 5% CO2. This was
followed by washing with PBS, dialysis with 0.1% Triton X-100 for 5
min, and sealing with 3% BSA for 30 min. The nuclei were stained
with DAPI for 5 min at 25°C following incubation with p65
fluorescent antibodies diluted in 0.3% BSA-PBS according to the
manufacturer's instructions at room temperature, and the 96-well
plates were imaged using a Cytation 5 reader; images were obtained
and stored. ImageJ software 1.51 was used for analysis.
Mouse model of ovariectomy
Ovariectomy and micro-computed tomography (micro-CT)
were performed as previously described (23). A total of 30 female C57BL/6 mice,
weighing ~20 g and aged 9 weeks, were purchased from GemPharmatech
LLC., and were randomly assigned to one of five groups as follows:
The sham-operated group, ovariectomized (OVX) group, estrogen
[estradiol (E2)] group, and the Cas (2.5 mg/kg) and Cas
(5 mg/kg) treatment groups. The mice were maintained at the
laboratory of pathogen-free animals of Guangxi Medical University
under conditions of constant temperature at 25°C, 60% constant
humidity, 12 h of alternating light, unrestricted activity, and
SPF-grade normal mouse chow available ad libitum. Following
acclimatization, tribromoethanol (150 mg/kg) was injected
intraperitoneally for bilateral ovariectomy, and at 1 week after
surgery, mice in the treatment group were intraperitoneally
injected with 2.5 or 5 mg/kg Cas, mice in the E2 group
were injected with 100 ng/kg E2 and mice in the sham
group and OVX group were injected with the same volume of normal
saline as the treatment group, once every 2 days for 42 consecutive
days. After 6 weeks, the mice were asphyxiated by carbon dioxide.
The flow rate of CO2 used for euthanasia was 50%/min,
for a 10-liter volume chamber, using a flow rate of 5 liters per
minute. Their lower limbs and viscera were fixed in 4%
paraformaldehyde for 48 h at 25°C, and their tibiae were examined
using micro-CT (SCANO MEDICAL AG).
Micro-CT and histological analysis
The isolated whole left tibiae were imaged and
quantified using micro-CT. The analysis method was the same as in
that in a previous study (24).
The following were the scanning parameters: 50 kV source voltage;
500 A source current; 0.5 mm Al filter; 9 m pixel size; 180 degree
rotation step. The region of interest in the distal tibia was
selected and Mimics 19.0 (Materialise) was used to perform 3D
reconstruction. Using SkyScan CT software (version 1.15.22;
Bruker), the 3D images were used to calculate bone volume/tissue
volume (BV/TV), trabecular number (Tb.N), trabecular thickness
(Tb.Th), and trabecular separation (Tb.Sp). The sample was fixed in
4% paraformaldehyde at 25°C for 24 h and decalcified at a
temperature of 4°C for at least 2 weeks with 10% EDTA before being
cut into 5-µm-thick slices for histological staining and
analysis. Next, the sections were stained with hematoxylin and
eosin (H&E) for 5 min or TRAP staining for 30 min at 25°C. The
heart, liver and kidney of mice were also stained with H&E
staining after the same preprocessing. The images of sections were
obtained using uSCOPE MXII-20 Digital Microscope Slide Scanner
(Microscopes International) and analyzed using Bioquent Osteo
software 2019 (BIOQUENT).
Osteoblast proliferation and
differentiation
MC3T3-E1 cells (product code FH0384; Shanghai Fuheng
Biotechnology Co., Ltd.) were removed from -80°C for rewarming, and
cultured with a mixture of 10% FBS, 1% penicillin/streptomycin
antibiotics and DMEM for 48 h at 37°C. The cells were then digested
with EDTA-trypsin for 5 min when the culture flask was full, and
centrifuged at 120 × g for 5 min at room temperature. The cells
were then inoculated in a 48-well plate (5×104
cells/well) and Cas (0, 1 and 2 µM) was used to treat the
cultured cells. After 7 days, the alkaline phosphatase (ALP)
activity was determined using a BCIP/NBT alkaline phosphatase
chromogenic kit (cat. no. C3206; Beyotime Institute of
Biotechnology) for 15 min at room temperature. The cells were
stained using Alizarin Red S solution kit (cat. no. G1452, Beijing
Solarbio Science & Technology Co., Ltd.) for 15 min at 25°C to
observe the formation of bone nodules by a Cytation 5 reader, 21
days later.
Statistical analysis
The aforementioned experiments were performed three
separate times, the results were input into Graphpad Prism 8.00
(GraphPad Software, Inc.) for analysis, and the findings are
expressed as the mean ± standard deviation (SD). The comparison of
multicomponent means was performed using unpaired Student's t-test
or one-way ANOVA, followed by Tukey's post hoc analysis. A P-value
<0.05 was considered to indicate a statistically significant
difference.
Results
Cas inhibits RANKL-induced osteoclast
differentiation
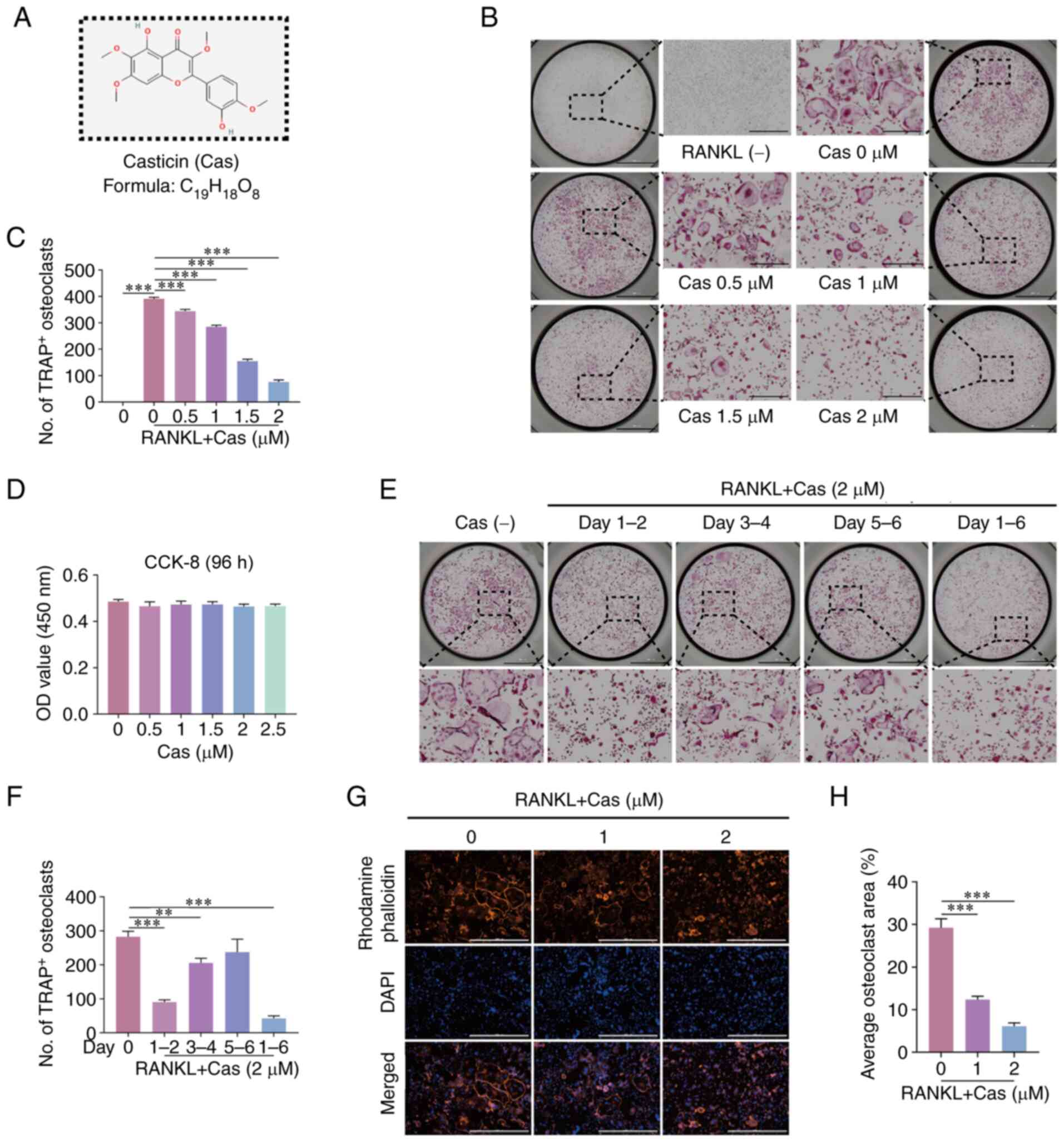
Cas (Fig. 1A) has
been shown to exhibit anti-inflammatory properties (25). The present study thus investigated
its role in osteoclast differentiation. In vitro experiments
with TRAP staining revealed that with the increasing Cas
concentration, the inhibition of RANKL-induced osteoclast
differentiation was observed in a concentration-dependent manner
(Fig. 1B and C). The viability of
BMMs was not affected at this concentration range (Fig. 1D), particularly at 2 µM,
where the inhibitory effect was particularly pronounced. In
addition, during the 6 days of incubation of the BMMs with RANKL,
which is the entire differentiation process of osteoclasts, Cas
continued to inhibit osteoclast differentiation and was most
effective in the early stages of differentiation (Fig. 1E and F). Mature osteoclasts can
form an actin ring, which is essential for osteoclasts to perform
their bone resorption functions (26). Herein, the F-actin belt formed by
osteoclast differentiation was stained with rhodamine-phalloidin
and it was found that the formation of the F-actin belt was
significantly hindered by Cas (Fig.
1G and H).
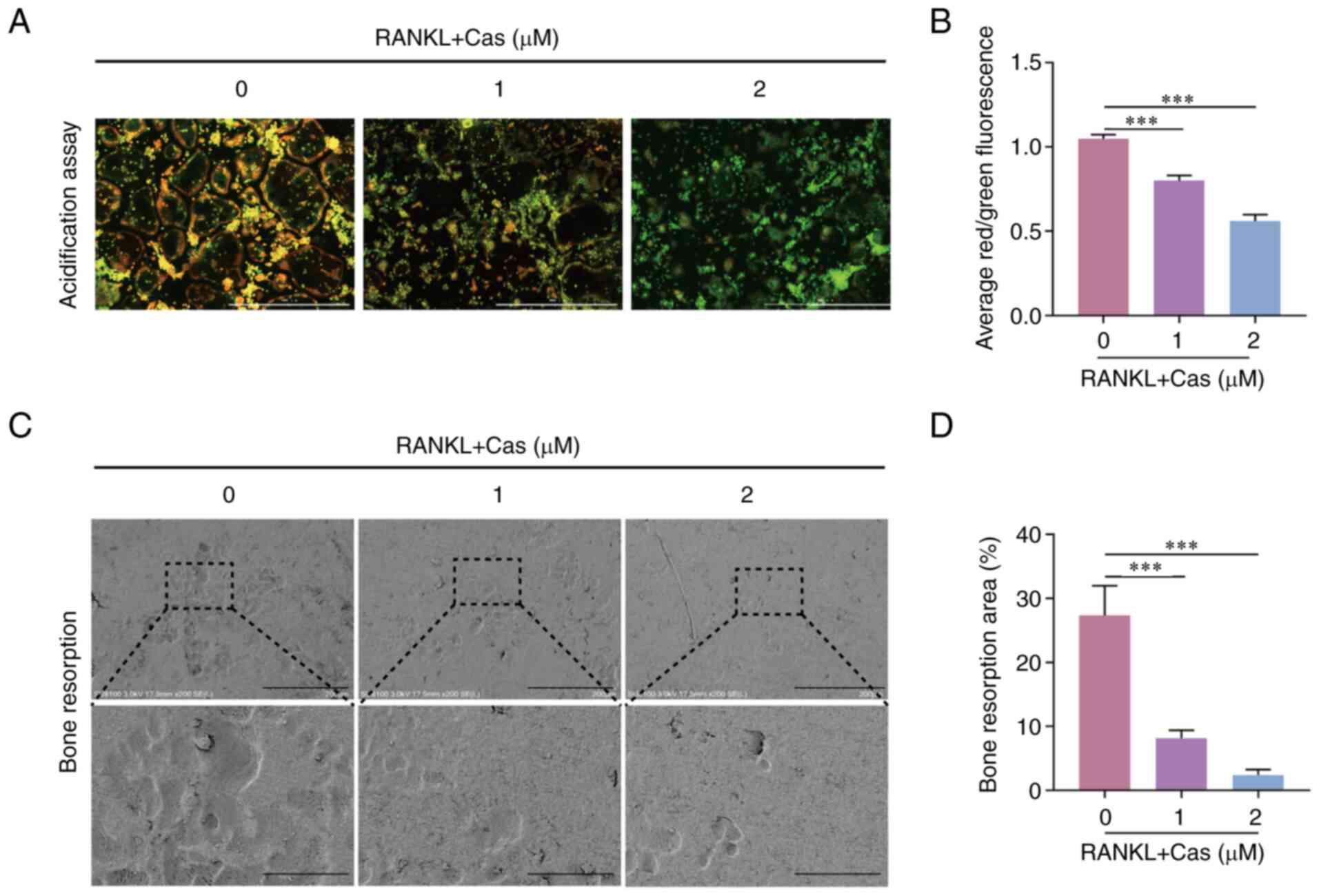
Cas inhibits the acidification and bone
resorption of osteoclasts
Osteoclast cytoplasmic protons are transported to
confined areas to form an acidic extracellular microenvironment for
bone resorption. The present study then determined the effects of
Cas on bone resorption function by growing RANKL-stimulated
osteoclasts in 96-well plates for 2 days, while intervening with
Cas and staining with acridine orange. The results revealed a
decrease in acid secretion from osteoclasts (Fig. 2A and B). In addition,
RANKL-stimulated osteoclasts were grown on bone chips for 2 days
with Cas concentrations of 1 and 2 µM, and under an electron
microscope, the area of resorption pits on the bone surface
appeared to be reduced to varying degrees (Fig. 2C and D). These results verified
that Cas inhibited the acid secretion and bone resorption function
of osteoclasts.
Cas attenuates the expression of NFATc1,
related genes and downstream proteins
The osteoclast differentiation process involves the
expression of a series of specific genes that promote osteoclast
maturation and bone resorption. The present study used RT-qPCR to
detect osteoclast-specific gene expression under the influence of
various concentrations of Cas. The results demonstrated that Cas
inhibited the expression of osteoclast-specific genes, including
Nfatc1, Fos, ATPase H+ transporting V0 subunit D2
(Atp6v0d2), Ctsk, dendrocyte expressed seven transmembrane
protein (Dcstamp) and matrix metalloproteinase 9
(Mmp9) (Fig. 3A-F).
Subsequently, western blot analysis was used to examine the effects
of Cas on the expression of NFATc1 and its downstream proteins. Cas
treatment significantly reduced the protein expression levels of
NFATc1, c-Fos, CTSK and ATP6V0D2 (Fig. 3G-K). In summary, these findings
indicated that Cas inhibited NFATc1 and downstream gene expression
in vitro.
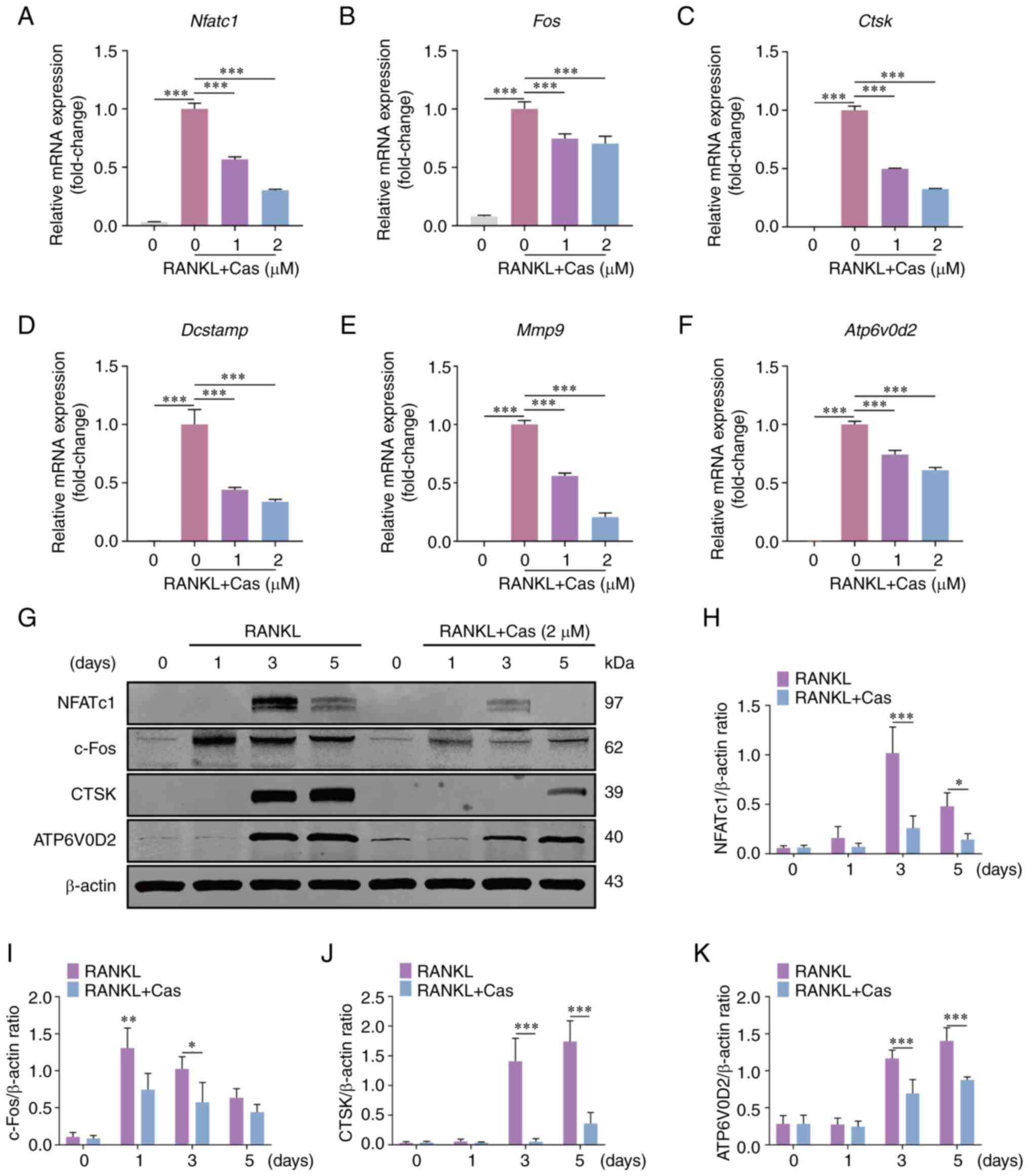 | Figure 3Cas inhibits the expression of
NFATc1, downstream related genes and proteins. (A-F) Cas inhibited
the expression of osteoclast-related genes Nfatc1,
Fos, Ctsk, Dcstamp, Mmp9 and
Atp6v0d2 stimulated by RANKL. (G) Typical western blot
images of NFATc1, c-Fos, CTSK and ATP6V0D2 protein expression in
osteoclasts stimulated by RANKL and 2 µM Cas for 0, 1, 3 and
5 days. (H-K) The relative ratio of the gray values of NFATc1,
c-Fos, CTSK and ATP6V0D2 to β-actin was quantified.
*P<0.05, **P<0.01 and
***P<0.001. All data are expressed as the mean ± SD.
Cas, casticin; NFATc1, nuclear factor of activated T cells,
cytoplasmic 1; Ctsk, cathepsin K; Dcstamp, dendrocyte
expressed seven transmembrane protein; Mmp9, matrix
metalloproteinase 9; Atp6v0d2, ATPase H+
transporting V0 subunit D2; RANKL, receptor activator of nuclear
factor κB ligand. |
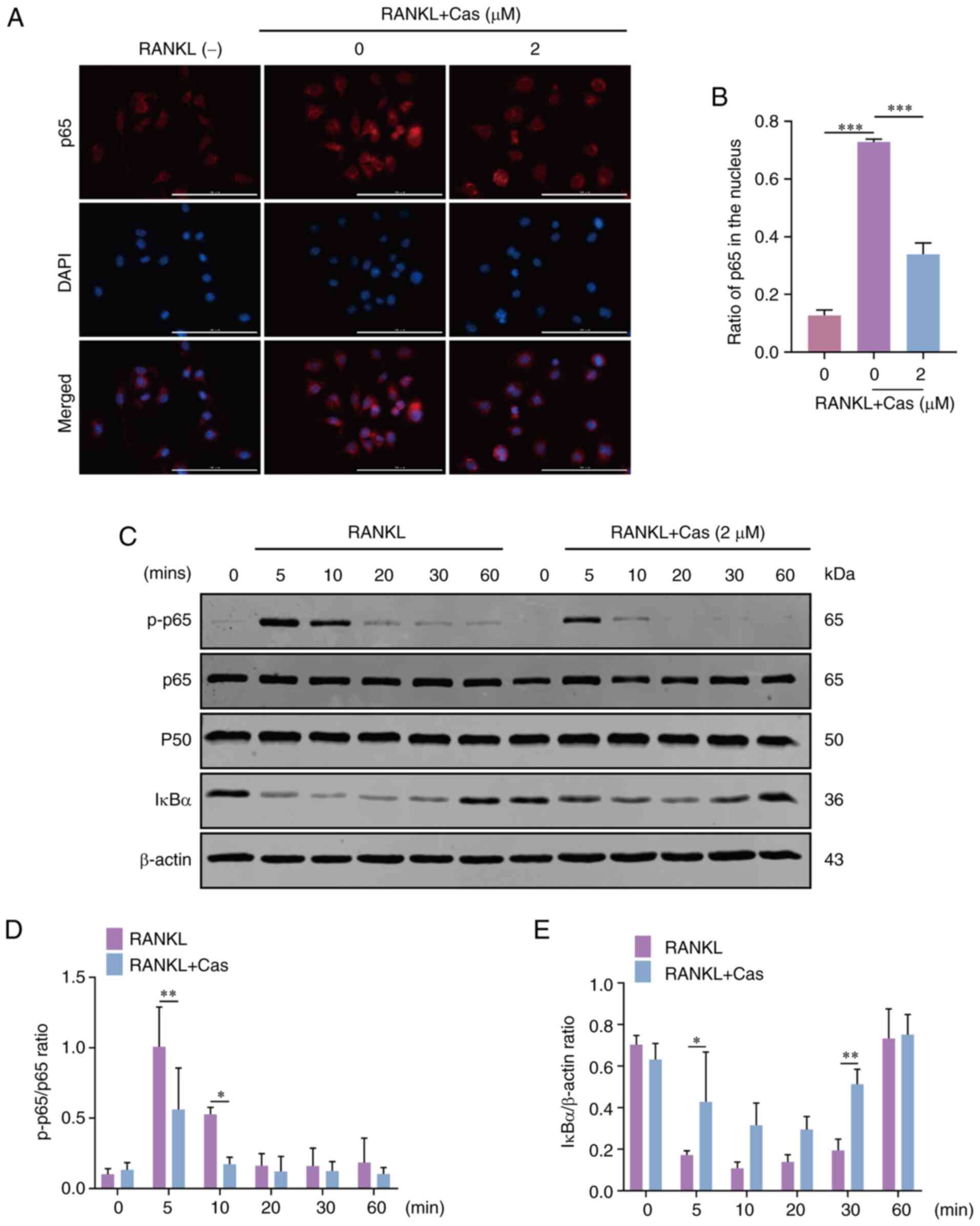
Cas inhibits the RANKL-induced activation
of the AKT/ERK and NF-κB pathways
The main signaling pathways activated during
osteoclastogenesis are considered to be NF-κB, MAPK and AKT
(27). In the present study, to
investigate the potential mechanisms affecting osteoclasts
following Cas treatment, western blot analysis was used to examine
the phosphorylation levels of signaling cascades for osteoclast
differentiation. When the BMMs were stimulated with RANKL, p65
phosphorylation was inhibited. Immunofluorescence was also used to
detect NF-κB p65 nuclear translocation (Fig. 4A and B). In addition, IκBα
degradation was reduced following exposure to 2 µM Cas
(Fig. 4C-E). The results revealed
that various Cas concentrations significantly altered the nuclear
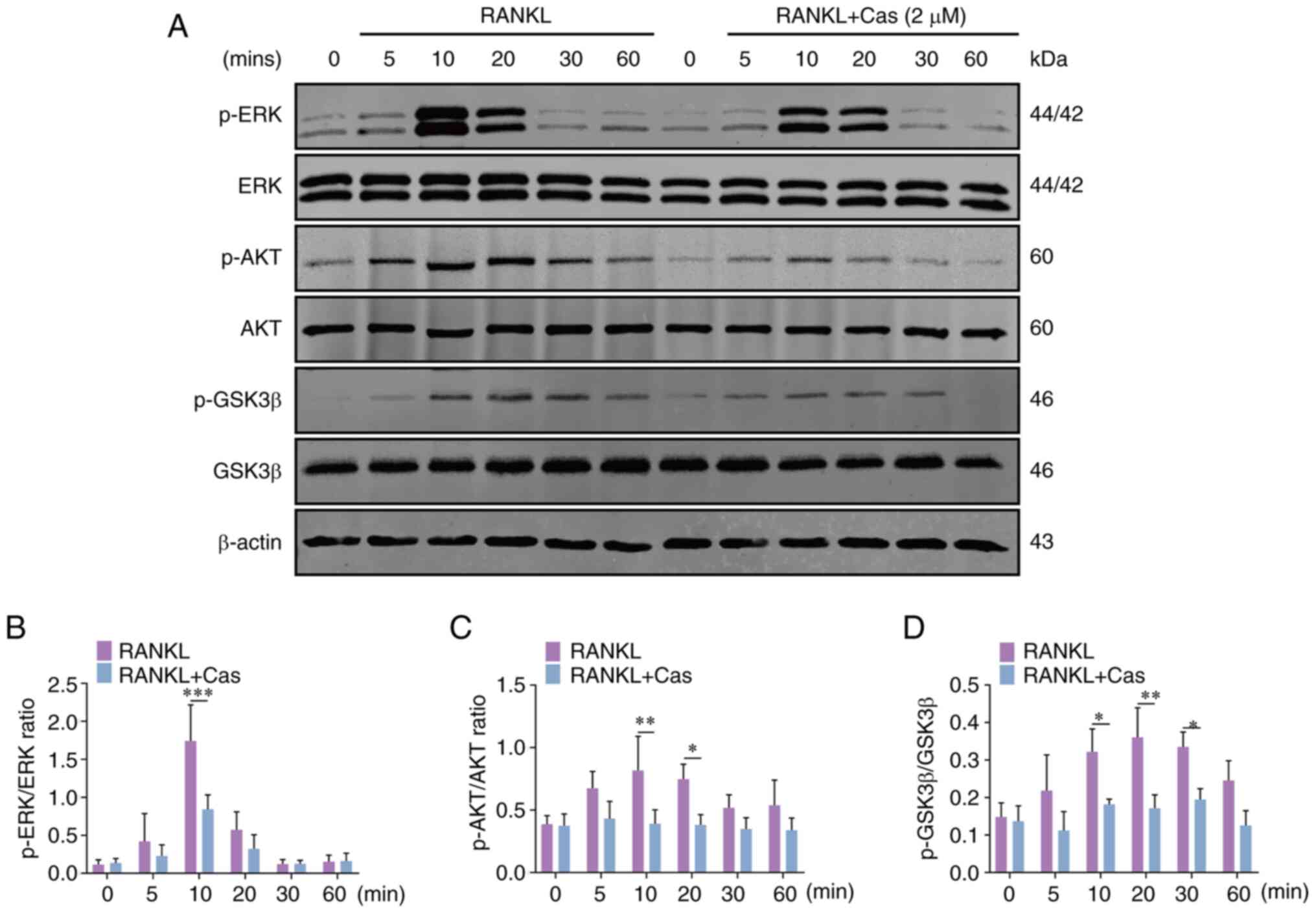
translocation of p65. Moreover, Cas significantly inhibited ERK
phosphorylation at 10 min (Fig. 5A
and B), while JNK and p38 phosphorylation were unaffected. TAK1
belongs to the MAPK kinase family, and herein, it was verified that
Cas had no significant effect on the phosphorylation of TAK1
upstream of the signaling pathway (Fig. S1). The AKT signaling pathway is
also involved in osteoclast differentiation and survival (28). Cas inhibited the phosphorylation
of AKT and GSK3β (Fig. 5A, C and
D), a downstream signal of AKT. These findings indicated that
Cas exerts an inhibitory effect on the activation of the AKT/ERK
and NF-κB signaling pathways.
Cas reduces ovariectomy-induced bone loss
in mice
To further investigate whether Cas can prevent
osteoporosis, a mouse model of osteoporosis was employed using mice
with devitalized ovaries. The mice were then injected 2.5 and 5
mg/kg Cas in the experimental group and with saline 100 ng/kg
E2 in the control group every 2 days for 42 days. No
mortality or major adverse effects were observed during the surgery
and the therapeutic intervention period. Cas had no significant
toxic effects on the heart, liver, or kidneys, according to H&E
staining (Fig. S2). Cas and
E2 prevented bone loss in mice compared with the control
group (Fig. 6A), and quantitative
analysis confirmed the trabecular parameters. BV/TV and Tb.N were
significantly increased, Tb.sp was reduced in the experimental and
E2 groups when compared to the vehicle group (Fig. 6B-E). The amount of bone trabeculae
was significantly higher in the Cas-treated animals, according to
H&E staining (Fig. 6F and G).
The groups treated with Cas had fewer osteoclasts, according to
TRAP staining (Fig. 6H and I).
These results further confirmed that Cas inhibits osteoclast
formation and function in vivo.
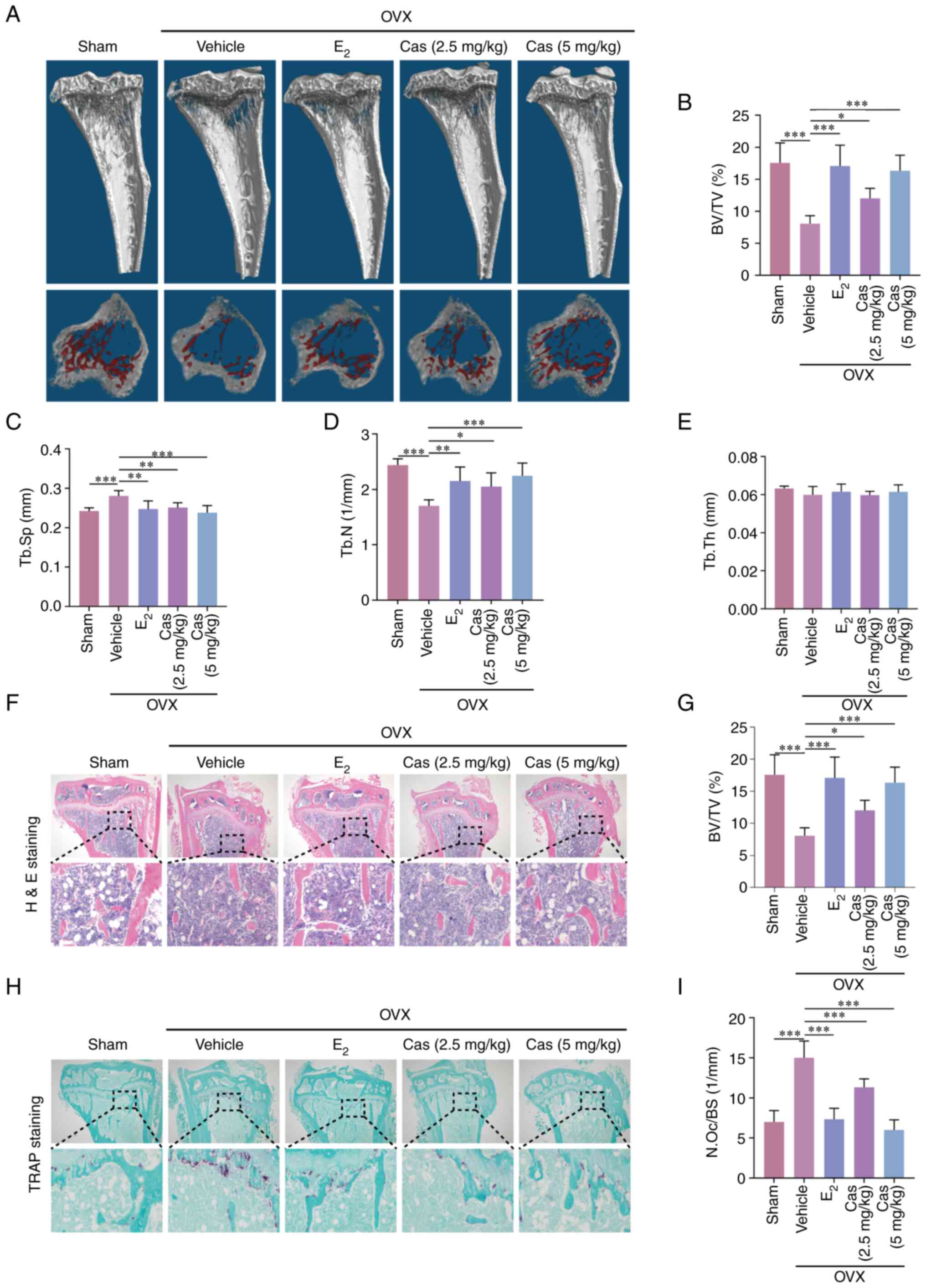 | Figure 6Casticin inhibits
ovariectomized-induced bone loss. (A) 3D representative images of
different groups of tibial plateau of mice. (B-E) Tibial cancellous
bone data of BV/TV, trabecular separation, trabecular number, and
trabecular thickness (n=6). (F) Representative H&E staining
results of tibial plateau sections. (G) Quantification of H&E
staining BV/TV in tibial plateau. (H) Representative TRAP staining
results of tibial plateau sections. (I) Quantification of the
number of osteoclasts in tibial plateau after TRAP staining.
*P<0.05, **P<0.01 and
***P<0.001. All data are expressed as the mean ± SD.
BV/TV, volume/tissue volume; H&E, hematoxylin and eosin; TRAP,
tartrate-resistant acid phosphatase; OVX, ovariectomized;
E2, estradiol; Cas, casticin; Tb.Sp, trabecular
separation; Tb.N, trabecular number; Tb.Th, trabecular
thickness. |
Cas has no significant effect on
osteoblasts in vitro
The present study also explored the effects of Cas
on osteoblasts. MC3T3-E1 cells were cultured in the presence of Cas
(1 and 2 µM). After 7 days of culture with osteoblast
induction solution, the cells were stained with alkaline
phosphatase and Alizarin Red S solutions after 21 days. Cas had no
effect on the MC3T3-E1 cells (Fig.
S3).
Discussion
The most common bone disease affecting women is
postmenopausal osteoporosis. Estrogen deficiency causes excessive
osteoclast activation and bone resorption, ultimately leading to a
decrease in bone mass (29).
Currently, there are a number of methods used to treat osteoporosis
(30), and bisphosphonates are
commonly used clinically, with considerable adverse effects, such
as causing bone discontinuity after fracture and inhibiting bone
formation (31). Therefore, the
development of a class of drugs that are effective in the treatment
of osteoporosis without severe side-effects is urgently required.
The benefits of natural plant-based active components for safety
over synthetic chemical medicines or estrogens are more pronounced
(32-34). Cas, as a natural compound with
multiple pharmacological activities, has proven to be effective in
several diseases and does not damage vital organs in mice (19,35,36). Bone mass is regulated by both
osteoclasts and osteoblasts (37). The effects of Cas on osteoclast
formation, fusion, and bone resorption capacity in vitro, as
well as the osteoblast lineage, were investigated in the present
study, while in vivo, the biological effects of Cas were
examined on ovariectomy-induced osteoporosis. Cas inhibited
osteoclast formation by suppressing the activation of the AKT/ERK
and NF-κB pathways. As a result, in vivo analyses revealed
that Cas inhibited the bone loss caused by a lack of estrogen.
Multiple cytokines, including RANKL and M-CSF,
stimulate osteoclast differentiation, both of which lead to the
activation of associated signaling cascades by binding to their
downstream receptors to promote osteoclast activation (38). RANKL triggers the activation of
downstream pathways through RANK signaling, followed by TRAF6 and
downstream molecules, including NF-κB, MAPK, AKT-GSK3β, etc., all
of which play critical roles in osteoclast development (39-41). During osteoclast differentiation,
the phosphorylation of p65 and the degradation of the IκBα via
proteasomes can result in the nuclear dimerization of p50 and
p65/RelA. The activation of ERK in the MAPK pathway is also
essential for osteoclast survival (42). Herein, Cas was found to hinder
osteoclast differentiation and activation by affecting the
phosphorylation of p65 and nuclear localization, the degradation of
IκBα, as well as the phosphorylation of ERK. In comparison, TAK1 is
a member of the MAPK kinase family (43), and Cas had no effect on the
phosphorylation of TAK1.
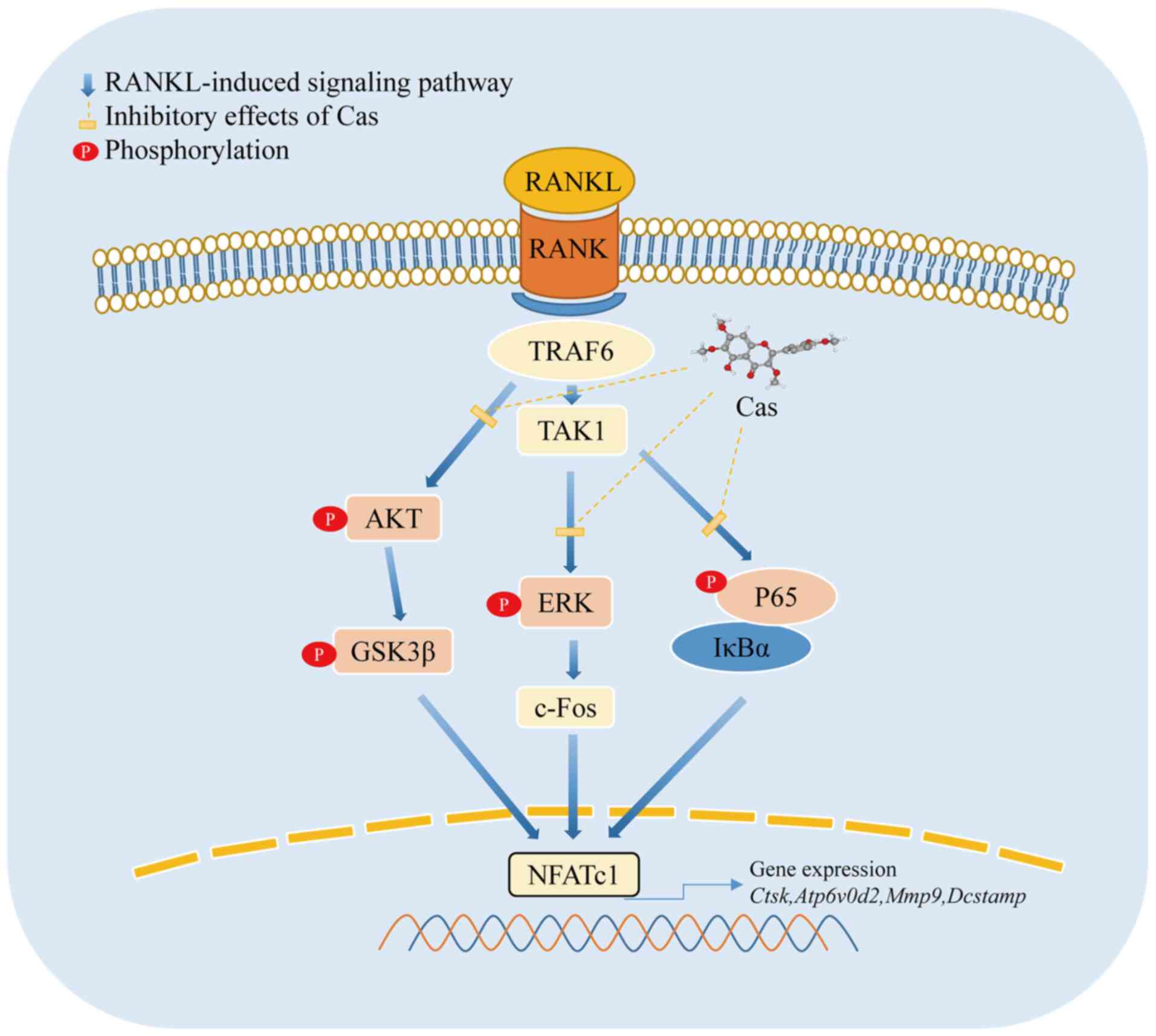
A crucial transcription factor for osteoclasts is
NFATc1 (44), of which a high
expression is regulated by NF-κB and c-Fos. The persistent
production of c-Fos is supported by the phosphorylation of ERK,
which in turn stimulates NFATc1 in osteoclast progenitors and
ultimately results in osteoclastogenesis (45). According to previous research, AKT
promoted the formation of the inactive form of GSK3β (p-GSK3β), as
well as the nuclear localization of NFATc1, whereas constitutively
active GSK3β overexpression inhibited osteoclast formation by
downregulating NFATc1 (46-48). The results of the present study
demonstrated that Cas inhibited the phosphorylation of AKT/GSK3β
and thus prevented the expression of related proteins and
transcription factors required for osteoclast maturation, as well
as the expression of the osteoclast-associated genes, Nfatc1,
Fos, Atp6v0d2, Ctsk, Dcstamp, and Mmp9 (Fig. 7), which are directly or indirectly
regulated by NFATc1 (49).
There are some limitations to the present study. The
study explored the mechanism by which Cas inhibits osteoclast
activation, which has been shown to be multi-pathway. Studies on
NF-κB, MAPK, AKT and other conventional pathways have been widely
carried out (27,50), and most studies use pathway
inhibitors as experimental controls (51-53), thus, in the future, the mechanism
of Cas inhibition of osteoclast activation using inhibitors of the
ERK/AKT/GSK3β pathway will be further investigated. The association
between the Cas-inhibited RANKL-induced ERK/AKT/GSK3β signaling
pathway and NFATc1 and its downstream proteins requires further
study. Notably, the molecular mechanism of Cas inhibition needs
further study in vivo. Cas has recently been reported to
inhibit reactive oxygen species (ROS) to reduce cartilage
degeneration associated with osteoarthritis (54), and whether there is a link with
ROS clearance in osteoclasts and bone cells needs to be further
investigated. Cas also plays a role in oncological disease, and its
role in tumor bone metastasis may represent an interest of future
investigation.
In conclusion, the results of the present study
demonstrated that Cas prevents ovariectomy-induced bone loss by
suppressing the effects of RANKL on osteoclast differentiation and
function via regulating the AKT/ERK and NF-κB signaling pathways.
Thus, Cas may have potential for use as a therapeutic agent for the
prevention of postmenopausal osteoporosis.
Supplementary Data
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ, QL and JX conceived and designed the study. FY
and YS drafted the manuscript and performed the experiments. JL and
KW performed the experiments. HL and JC acquired and analyzed the
data. JX revised the manuscript. All of the authors confirm the
authenticity of all the raw data. All authors read and approved the
manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
The animal experiments were performed after approval
(approval no. 202110004) from the Animal Care and Welfare Committee
of Guangxi Medical University (Nanning, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
Funding
The present study was funded by the National Natural Science
Foundation of China (grant no. 81960405), and the Guangxi Science,
Technology Base and Talent Special Project (grant no. GuikeAD
19254003).
References
|
1
|
Michalski MN and McCauley LK: Macrophages
and skeletal health. Pharmacol Ther. 174:43–54. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kular J, Tickner J, Chim SM and Xu J: An
overview of the regulation of bone remodelling at the cellular
level. Clin Biochem. 45:863–873. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sobh MM, Abdalbary M, Elnagar S, Nagy E,
Elshabrawy N, Abdelsalam M, Asadipooya K and El-Husseini A:
Secondary osteoporosis and metabolic bone diseases. J Clin Med.
11:23822022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Han Y, You X, Xing W, Zhang Z and Zou W:
Paracrine and endocrine actions of bone-the functions of secretory
proteins from osteoblasts, osteocytes, and osteoclasts. Bone Res.
6:162018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bae SJ, Shin MW, Son T, Lee HS, Chae JS,
Jeon S, Oh GT and Kim KW: Ninjurin1 positively regulates osteoclast
development by enhancing the survival of prefusion osteoclasts. Exp
Mol Med. 51:1–16. 2019.
|
|
7
|
Chen K, Qiu P, Yuan Y, Zheng L, He J, Wang
C, Guo Q, Kenny J, Liu Q, Zhao J, et al: Pseurotin A inhibits
osteoclastogenesis and prevents ovariectomized-induced bone loss by
suppressing reactive oxygen species. Theranostics. 9:1634–1650.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sun Y, Li J, Xie X, Gu F, Sui Z, Zhang K
and Yu T: Macrophage-osteoclast associations: Origin, polarization,
and subgroups. Front Immunol. 12:7780782021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tobeiha M, Moghadasian MH, Amin N and
Jafarnejad S: RANKL/RANK/OPG pathway: A mechanism involved in
exercise-induced bone remodeling. Biomed Res Int. 2020:69103122020.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Asagiri M and Takayanagi H: The molecular
understanding of osteoclast differentiation. Bone. 40:251–264.
2007. View Article : Google Scholar
|
|
11
|
Takayanagi H: The role of NFAT in
osteoclast formation. Ann N Y Acad Sci. 1116:227–237. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Grigoriadis AE, Wang ZQ, Cecchini MG,
Hofstetter W, Felix R, Fleisch HA and Wagner EF: c-Fos: A key
regulator of osteoclast-macrophage lineage determination and bone
remodeling. Science. 266:443–448. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bellavia D, Dimarco E, Costa V, Carina V,
De Luca A, Raimondi L, Fini M, Gentile C, Caradonna F and Giavaresi
G: Flavonoids in bone erosive diseases: Perspectives in
osteoporosis treatment. Trends Endocrinol Metab. 32:76–94. 2021.
View Article : Google Scholar
|
|
14
|
Mesaik MA, Murad S, Khan KM, Tareen RB,
Ahmed A and Choudhary MI: Isolation and immunomodulatory properties
of a flavonoid, casticin from Vitex agnus-castus. Phytother Res.
23:1516–1520. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chan EWC, Wong SK and Chan HT: Casticin
from Vitex species: A short review on its anticancer and
anti-inflammatory properties. J Integr Med. 16:147–152. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ramchandani S, Naz I, Lee JH, Khan MR and
Ahn KS: An overview of the potential antineoplastic effects of
casticin. Molecules. 25:12872020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun C, Yan H, Jiang K and Huang L:
Protective effect of casticin on experimental skin wound healing of
rats. J Surg Res. 274:145–152. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee JH, Lee S, Nguyen QN, Phung HM, Shin
MS, Kim JY, Choi H, Shim SH and Kang KS: Identification of the
active ingredient and beneficial effects of Vitex rotundifolia
fruits on menopausal symptoms in ovariectomized rats. Biomolecules.
11:10332021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li J, Qiu C, Xu P, Lu Y and Chen R:
Casticin improves respiratory dysfunction and attenuates oxidative
stress and inflammation via inhibition of NF-ĸB in a chronic
obstructive pulmonary disease model of chronic cigarette
Smoke-exposed rats. Drug Des Devel Ther. 14:5019–5027. 2020.
View Article : Google Scholar :
|
|
20
|
Xu J, Wu HF, Ang ES, Yip K, Woloszyn M,
Zheng MH and Tan RX: NF-kappaB modulators in osteolytic bone
diseases. Cytokine Growth Factor Rev. 20:7–17. 2009. View Article : Google Scholar
|
|
21
|
Qin A, Cheng TS, Lin Z, Pavlos NJ, Jiang
Q, Xu J, Dai KR and Zheng MH: Versatile roles of V-ATPases
accessory subunit Ac45 in osteoclast formation and function. PLoS
One. 6:e271552011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
23
|
Xiao L, Zhong M, Huang Y, Zhu J, Tang W,
Li D, Shi J, Lu A, Yang H, Geng D, et al: Puerarin alleviates
osteoporosis in the ovariectomy-induced mice by suppressing
osteoclastogenesis via inhibition of TRAF6/ROS-dependent MAPK/NF-κB
signaling pathways. Aging (Albany NY). 12:21706–21729. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sapra L, Shokeen N, Porwal K, Saini C,
Bhardwaj A, Mathew M, Mishra PK, Chattopadhyay N, Dar HY, Verma B
and Srivastava RK: Bifidobacterium longum ameliorates
ovariectomy-induced bone loss via enhancing anti-osteoclastogenic
and immunomodulatory potential of regulatory B cells (Bregs). Front
Immunol. 13:8757882022. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li X, Mei W, Huang Z, Zhang L, Zhang L, Xu
B, Shi X, Xiao Y, Ma Z, Liao T, et al: Casticin suppresses
monoiodoacetic acid-induced knee osteoarthritis through inhibiting
HIF-1α/NLRP3 inflammasome signaling. Int Immunopharmacol.
86:1067452020. View Article : Google Scholar
|
|
26
|
Teitelbaum SL: Bone resorption by
osteoclasts. Science. 289:1504–1508. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pereira M, Petretto E, Gordon S, Bassett
JHD, Williams GR and Behmoaras J: Common signalling pathways in
macrophage and osteoclast multinucleation. J Cell Sci.
131:jcs2162672018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Adamik J, Pulugulla SH, Zhang P, Sun Q,
Lontos K, Macar DA, Auron PE and Galson DL: EZH2 supports
osteoclast differentiation and bone resorption via epigenetic and
cytoplasmic targets. J Bone Miner Res. 35:181–195. 2020. View Article : Google Scholar
|
|
29
|
Khosla S, Oursler MJ and Monroe DG:
Estrogen and the skeleton. Trends Endocrinol Metab. 23:576–581.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Song S, Guo Y, Yang Y and Fu D: Advances
in pathogenesis and therapeutic strategies for osteoporosis.
Pharmacol Ther. 237:1081682022. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim HS, Jung HY, Kim MO, Joa KL, Kim YJ,
Kwon SY and Kim CH: Successful conservative treatment: Multiple
atypical fractures in osteoporotic patients after bisphosphate
medication: A unique case report. Medicine (Baltimore).
94:e4462015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang X, Liang J, Wang Z, Su Y, Zhan Y, Wu
Z, Li J, Li X, Chen R, Zhao J, et al: Sesamolin protects mice from
ovariectomized bone loss by inhibiting osteoclastogenesis and
RANKL-mediated NF-κB and MAPK signaling pathways. Front Pharmacol.
12:6646972021. View Article : Google Scholar
|
|
33
|
Xian Y, Su Y, Liang J, Long F, Feng X,
Xiao Y, Lian H, Xu J, Zhao J, Liu Q and Song F: Oroxylin A reduces
osteoclast formation and bone resorption via suppressing
RANKL-induced ROS and NFATc1 activation. Biochem Pharmacol.
193:1147612021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ding D, Yan J, Feng G, Zhou Y, Ma L and
Jin Q: Dihydroartemisinin attenuates osteoclast formation and bone
resorption via inhibiting the NF-κB, MAPK and NFATc1 signaling
pathways and alleviates osteoarthritis. Int J Mol Med. 49:42022.
View Article : Google Scholar
|
|
35
|
Kowalski M, Assa A, Patil K, Terrell C,
Holliday N and Pai SB: Casticin impacts key signaling pathways in
colorectal cancer cells leading to cell death with therapeutic
implications. Genes (Basel). 13:8152022. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fan L, Zhang Y, Zhou Q, Liu Y, Gong B, Lü
J, Zhu H, Zhu G, Xu Y and Huang G: Casticin inhibits breast cancer
cell migration and invasion by down-regulation of PI3K/Akt
signaling pathway. Biosci Rep. 38:BSR201807382018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang L, You X, Zhang L, Zhang C and Zou W:
Mechanical regulation of bone remodeling. Bone Res. 10:162022.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Takayanagi H: RANKL as the master
regulator of osteoclast differentiation. J Bone Miner Metab.
39:13–18. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sun Y, Li J, Xie X, Gu F, Sui Z, Zhang K
and Yu T: Recent advances in osteoclast biological behavior. Front
Cell Dev Biol. 9:7886802021. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Novack DV: Role of NF-κB in the skeleton.
Cell Res. 21:169–182. 2011. View Article : Google Scholar
|
|
41
|
Cao H, Zhu K, Qiu L, Li S, Niu H, Hao M,
Yang S, Zhao Z, Lai Y, Anderson JL, et al: Critical role of AKT
protein in myeloma-induced osteoclast formation and osteolysis. J
Biol Chem. 288:30399–30410. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Soysa NS, Alles N, Aoki K and Ohya K:
Osteoclast formation and differentiation: an overview. J Med Dent
Sci. 59:65–74. 2012.PubMed/NCBI
|
|
43
|
Jo YJ, Lee HI, Kim N, Hwang D, Lee J, Lee
GR, Hong SE, Lee H, Kwon M, Kim NY, et al: Cinchonine inhibits
osteoclast differentiation by regulating TAK1 and AKT, and promotes
osteogenesis. J Cell Physiol. 236:1854–1865. 2021. View Article : Google Scholar
|
|
44
|
Sitara D and Aliprantis AO:
Transcriptional regulation of bone and joint remodeling by NFAT.
Immunol Rev. 233:286–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xu W, Chen X, Wang Y, Fan B, Guo K, Yang
C, Yu S, Pang Y and Zhang S: Chitooligosaccharide inhibits
RANKL-induced osteoclastogenesis and ligation-induced periodontitis
by suppressing MAPK/c-fos/NFATC1 signaling. J Cell Physiol.
235:3022–3032. 2020. View Article : Google Scholar
|
|
46
|
Zhang Q, Hu S, He Y, Song Z, Shen Y, Zhao
Z and Zhang Q, Qin L and Zhang Q: Monotropein protects against
inflammatory bone loss and suppresses osteoclast formation and bone
resorption by inhibiting NFATc1 via NF-κB and Akt/GSK-3β pathway.
Nutrients. 14:39782022. View Article : Google Scholar
|
|
47
|
Fan X, Xiong H, Wei J, Gao X, Feng Y, Liu
X, Zhang G, He QY, Xu J and Liu L: Cytoplasmic hnRNPK interacts
with GSK3β and is essential for the osteoclast differentiation. Sci
Rep. 5:177322015. View Article : Google Scholar
|
|
48
|
Yang S, Song D, Wang Z, Su Y, Chen J, Xian
Y, Huang J, Li J, Xu J, Zhao J and Liu Q: AKT/GSK3β/NFATc1 and ROS
signal axes are involved in AZD1390-mediated inhibitory effects on
osteoclast and OVX-induced osteoporosis. Int Immunopharmacol.
113:1093702022. View Article : Google Scholar
|
|
49
|
Song I, Kim JH, Kim K, Jin HM, Youn BU and
Kim N: Regulatory mechanism of NFATc1 in RANKL-induced osteoclast
activation. FEBS Lett. 583:2435–2440. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Meng B, Wu D, Cheng Y, Huang P, Liu Y, Gan
L, Liu C and Cao Y: Interleukin-20 differentially regulates bone
mesenchymal stem cell activities in RANKL-induced
osteoclastogenesis through the OPG/RANKL/RANK axis and the NF-κB,
MAPK and AKT signalling pathways. Scand J Immunol. 91:e128742020.
View Article : Google Scholar
|
|
51
|
Jiang T, Gong Y, Zhang W, Qiu J, Zheng X,
Li Z, Yang G and Hong Z: PD0325901, an ERK inhibitor, attenuates
RANKL-induced osteoclast formation and mitigates cartilage
inflammation by inhibiting the NF-κB and MAPK pathways. Bioorg
Chem. 132:1063212023. View Article : Google Scholar
|
|
52
|
Lee ZH, Lee SE, Kim CW, Lee SH, Kim SW,
Kwack K, Walsh K and Kim HH: IL-1alpha stimulation of osteoclast
survival through the PI 3-kinase/Akt and ERK pathways. J Biochem.
131:161–166. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Chaisson ML, Branstetter DG, Derry JM,
Armstrong AP, Tometsko ME, Takeda K, Akira S and Dougall WC:
Osteoclast differentiation is impaired in the absence of inhibitor
of kappa B kinase alpha. J Biol Chem. 279:54841–54848. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chu J, Yan B, Zhang J, Peng L, Ao X, Zheng
Z, Jiang T and Zhang Z: Casticin attenuates osteoarthritis-related
cartilage degeneration by inhibiting the ROS-Mediated NF-κB
signaling pathway in vitro and in vivo. Inflammation. 43:810–820.
2020. View Article : Google Scholar : PubMed/NCBI
|